Embed Size (px)
Citation preview

1
Facultad d
e C
iencia
s E
xperim
enta
les
ogía
UNIVERSIDAD DE JAÉN
Facultad de Ciencias Experimentales
Trabajo Fin de Grado
Trabajo Fin de Grado
Alumno: Serena Montoro, Mª Ángeles
Jaén, julio, 2015
Detección de hemoparásitos en
sangre de aves

2
Facultad d
e C
iencia
s E
xperim
enta
les
ogía
UNIVERSIDAD DE JAÉN
Facultad de Ciencias Experimentales
Trabajo Fin de Grado
Trabajo Fin de Grado
Alumno: Serena Montoro, Mª Ángeles
Jaén, julio, 2015
Detección de hemoparásitos en
sangre de aves

3
. ÍNDICE
1. Resumen ………………………………………………………………………… 7
1.1. Abstract ………………………………………………………………… 7
2. Introducción ……………………………………………………………………… 8
3. Objetivos …………..…………………………………………………………… 12
4. Materiales y métodos ………………………………………………………… 13
4.1. Situación geográfica de los puntos de muestreo ...…………….... 13
4.2. Toma de la muestra de sangre, realización y fijación de frotis
sanguíneos .………………………………………………………………………. 15
4.3. Tinción de Giemsa ...…………….………………………………… 21
4.4. Observaciones al microscopio y toma de fotografías de las muestras
positivas ………………………………………………………….………… 24
4.5. Procesamiento de imágenes y recuento de eritrocitos, glóbulos
blancos y gametocitos …………….……………………………………………… 25
4.6. Análisis estadístico …………………………………….………… 29
5. Resultados ……………………………………………………………............. 29
5.1. Observación de frotis sanguíneos ………………….....………… 29
5.1.1. Determinación del nivel de parasitemia ……………… 29
5.1.2. Recuento de campos analizados ................................... 31
5.1.3. Recuento de eritrocitos, glóbulos blancos,
macrogametocitos y microgametocitos…………………… 32
6. Discusión ……………………………………………………………………… 37
7. Conclusiones ………………………………………………………………… 40
8. Referencias bibliográficas ……………………………………………………… 41
9. Anexos ………………………………………………………………………….. 47
9.1. Anexo 1. Cuaderno de campo ………………………………………. 47
4
9.2. Anexo 2. Observación al microscopio ……………………………… 47
9.3. Anexo 3. Recuento de los datos de las fotografías positivas
…………………………………………………………………………………….. 48

5
ÍNDICE DE FIGURAS Y TABLAS
Figura 1A. Vector de haemoprotideo, mosquito picador Culicoides
nubeculosus…………………………………………………………………………… 9
Figura 1B. Vector de haemoprotideo, mosca chupadora de sangre
hipoboscídea Pseudolynchia canarienses……………………………………… 9
Figura 2A. Microgametocito de H. columbae …………………………………. 10
Figura 2B. Macrogametocito de H. columbae ……………………................... 10
Figura 3. Ciclo vital de hemoparásitos aviares ………………………………… 11
Figura 4. Mapa de España a escala 1:25 ………………………………………. 14
Figura 5. Fotografía en la que se desinfecta la pata del ave ......................... 16
Figura 6A. Vena metatarsal media de la pata de un ave ….…………………. 17
Figura 6B. Incisión en la pata del animal ………..……………………………... 17
Figura 6C. Salida de la sangre tras la incisión ………………………………… 17
Figura 7. Fotografía de cómo se lleva a cabo el paso 1 del frotis sanguíneo .. 18
Figuras 8A y 8B. Pasos dos (A) y tres (B, en caso excepcional) del frotis
sanguíneo ………………………………….…………………………………….... 18
Figura 9. Presión con un algodón en la pata del ave .………………………… 19
Figura 10. Identificación de la muestra en el portaobjetos …………………… 20
Figura 11. Fijación de la muestra en un tubo Falcon con metanol absoluto … 20
Figura 12. Fotografía con los portaobjetos en la caja de transporte ………… 21
Figuras 13A y 13B. Preparaciones durante su tinción (13A) y tras 45-60
minutos (13B) …………………………………..…………………………………. 23
Figura 14. Lavado de las preparaciones tras su tinción .…………………….. 23
Figura 15. Preparaciones secándose a temperatura ambiente …………….. 23

6
Figura 16. Recorrido en el frotis sanguíneo para la búsqueda e identificación
de gametocitos de Hemosporidios ……………………………………………… 24
Figura 17. Segundo paso una vez incorporada la imagen deseada al programa
ImageJ ..……………………………………………………………………………. 26
Figura 18. Se pueden ver las tres réplicas que se obtendrían de la imagen
tratada ...…………………………………………………………………………… 26
Figura 19. Resalto únicamente el núcleo del elemento ……………………… 27
Figura 20. Fotografía con los elementos ya resaltados ……………………… 27
Figuras 21A y 21B. Los últimos pasos a realizar del recuento de eritrocitos .. 28
Figura 22. Los elementos resaltados y enumerados …………………………. 28
Figura 23. Porcentaje de individuos parasitados y su nivel de parasitemia con
respecto a la población total ...…………………………………………………… 30
Tabla 1: Datos referidos a las muestras sanguíneas tomadas en los diferentes
palomares, fecha, número de individuos y especie ……………………………. 15
Tabla 2. Criterio a seguir para establecer el nivel de parasitemia …………… 31
Tabla 3. Número de campos analizados en el total de todas las muestras
dependiendo del palomar ...……………………………………………………… 32
Tabla 4. Número de eritrocitos, glóbulos blancos y gametocitos que hay en
todos los campos de cada muestra parasitada analizada …………………… 33
Tabla 5. Media, Varianza y Desviación estándar del número de eritrocitos,
glóbulos blancos y gametocitos en cada palomar .……………………………. 34
Tabla 6. Media, Varianza y Desviación estándar del número de eritrocitos,
glóbulos blancos y gametocitos en cada periodo de tiempo en Fuerte del rey..35
Tabla 7. Nivel de parasitemia de los individuos del palomar de Fuerte del Rey
que se les sacó sangre en la primera y segunda recogida de muestras …..... 36
Tabla 8. Criterio a seguir para el nivel de parasitemia ………………………… 37

7
1. RESUMEN.
Durante el año 2014 se llevó a cabo una investigación sobre el nivel de
parasitemia de Columba livia (Gmelin, 1789) capturadas en palomares en
condiciones de semilibertad. Para ello, se realizó el muestreo de 101 muestras
en dos poblaciones jiennenses, Mancha Real (15) y Fuerte del Rey (86), de las
cuales el 26,73% (n=27) han presentado hemoparásitos pertenecientes al
género Haemoproteus. Según su morfología se deben a la especie
Haemoproteus columbae (Kruse, 1890). La toma de muestras se llevó a cabo
en dos recogidas de periodo distinto. En la segunda población, además se
analizaron 4 muestras procedentes de Streptopelia risoria (Linnaeus, 1758)
resultando todas libres de parásitos. Otros objetivos son la frecuencia relativa
de aparición de macrogametocitos y microgametocitos de las especies de
hemosporidios en cada palomar, y el recuento de eritrocitos y glóbulos blancos
en relación al número de gametocitos. Cada población presentaba diferentes
niveles de parasitemia.
1.1. Abstract.
This investigation, concerning the level of parasitemia of Columba livia (Gmelin,
1789) captured in dovecotes in conditions semi-freedom, has been made during
2014. Ous sample included blood of 101 pigeons proceding from two different
places of Jaén [Mancha Real (15) and Fuerte del Rey (86)], of which 26.73% (n
= 27) had hemoparasites of gender Haemoproteus. According to their
morphology they could be assigned to the species Haemoproteus columbae
(Kruse, 1890). The sampling in Fuerte del Rey was done in two differents
moments. In the second population, also 4 samples of Streptopelia risoria
(Linnaeus, 1758) was analyzed, and it was free of parasites. Other objectives
are to study the relative frequency of occurrence of macrogametocytes and
microgametocytes of species of haemosporidia in each dovecotes, and the
erythrocyte and white blood cells count, in relation to the number of
gametocytes. Each one of studied population had different levels of
parasitemia.

8
2. INTRODUCCIÓN.
El parasitismo representa una de las formas de vida más exitosas sobre el
planeta, existen más de 31.000 especies de protozoarios reportadas, y se
calcula que falta por investigar y describir cerca del 80% (Matta et al., 2001).
Las palomas conviven con humanos y otras especies animales en la
naturaleza, y son criadas como fuente de comida, como un hobby, como
símbolo, y para fines experimentales (Harlin, 1994). La interacción de las
palomas con el hombre y otras aves domésticas y silvestres les hace un
portador potencial de parásitos zoonóticos (Adang et al., 2008). Varios
parásitos obstaculizarán de forma significativa el crecimiento de la paloma, el
desarrollo y la productividad que a veces puede conducir a la muerte,
especialmente en pichones jóvenes (Fatihu et al., 1991).Sin embargo, los
hemoparásitos están considerados por muchos autores a tener efectos
benignos o leves (Scaglione et al., 2015).
En la naturaleza, sólo una fracción de artrópodos llevan patógenos, y así los
huéspedes vertebrados están más a menudo expuestos a las picaduras de los
individuos no infectados. Sin embargo, incluso si no se transmiten agentes
patógenos, se alimentan de sangre que irritan y distraen a los hospedadores, a
menudo provocando conductas defensivas (tales como el aseo). Con cada
picadura, los artrópodos van a suministrar compuestos salivales que cambian
las condiciones fisiológicas locales en el lugar de la picadura, la creación de un
interfaz donde el sistema inmune del huésped puede interactuar con la suite de
compuestos en la saliva. Los compuestos salivales pueden mejorar la
adquisición de sangre del huésped por el bloqueo de hemostasia, causando
vasodilatación y reduciendo inflamación (Waite et al., 2014).
En la actualidad, la investigación de parásitos sanguíneos versus huésped
aviar, canaliza sus esfuerzos al análisis de la ecología comportamental y
evolutiva; efecto de estos parásitos sobre el huésped y su aplicación a la
conservación de especies en vida silvestre (Van Riper et al., 1994). Los
parásitos sanguíneos desempeñan un papel fundamental en la ecología y la
evolución de las aves, ya que son capaces de afectar a la aptitud de acogida y
la supervivencia (Fecchio, 2011). Dependiendo del tipo pueden hallarse en el

9
plasma, glóbulos rojos (eritrocitos) o glóbulos blancos (leucocitos). Los grupos
más frecuentes en el Mundo son: Haemoproteus; Leucocytozoon; microfilaria;
Plasmodium; Trypanosoma y en menores proporciones Atoxoplasma, Babesia,
Hepatozoon, Lankesterella y Toxoplasma (Matta et al., 2001).
Estudios hasta la fecha, han determinado que el parásito de la sangre más
común encontrado en palomas es Haemoproteus columbae y la tasa de
infección puede ser tan alta como 75% con límites de 6 y 86%. Se observan en
las células rojas de la sangre. Los gametocitos de H. columbae se desarrollan
en diminutas formas hasta llegar a formas alargadas, con un perfil de media
luna, que rodean parcialmente el núcleo de la célula huésped. El núcleo de la
célula huésped puede ser desplazado sin llegar necesariamente al borde de la
célula. H. columbae se encuentra ampliamente distribuida en todo el mundo, en
particular en climas cálidos y templados (Reza et al., 2011).
1A 1B
Figura 1A. Vector de haemoprotideo, mosquito picador Culicoides nubeculosus
(modificado por Gutsevich, 1973) (Valkiünas, 2004). Figura 1B. Vector de
haemoprotideo, mosca chupadora de sangre hipoboscídea Pseudolynchia
canarienses (Krylov, 1994) (Fotografía original).
10
Los principales vectores hemoparásitos de los pájaros son mosquitos que pican
(Diptera: Ceratopogonidae) (véase figura 1A) y las moscas hipoboscídeas
(Hippoboscidae) (véase figura 1B) (Valkiūnas, 2004). Tanto la mosca macho
como la hembra, se alimentan de sangre dos veces al día, bebiendo sangre en
20-80 combates por minuto; ambos sexos transmiten Haemoproteus. H.
columbae madura en una etapa infecciosa después de 10 días; estas etapas
migran a las glándulas salivales de la mosca y puede ser transmitida cuando la
mosca muerde a otro paloma. Por lo general, ambos sexos pasan la mayoría (~
70% -90%) de su tiempo en el cuerpo de la paloma. Las moscas macho
dejarán el ave para encontrar a otra, y las hembras lo que harán es dejar pupas
sobre las superficies, tales como el suelo de la jaula de un pájaro cautivo
(Waite et al., 2014).
2A 2B
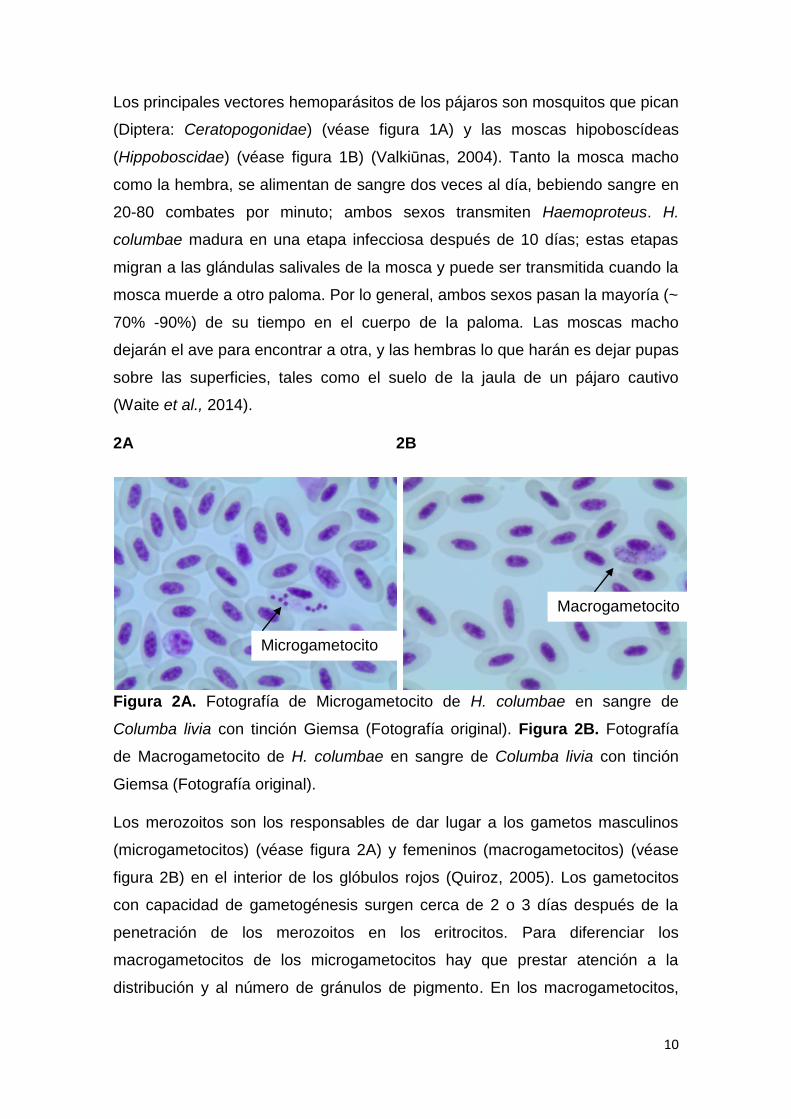
Figura 2A. Fotografía de Microgametocito de H. columbae en sangre de
Columba livia con tinción Giemsa (Fotografía original). Figura 2B. Fotografía
de Macrogametocito de H. columbae en sangre de Columba livia con tinción
Giemsa (Fotografía original).
Los merozoitos son los responsables de dar lugar a los gametos masculinos
(microgametocitos) (véase figura 2A) y femeninos (macrogametocitos) (véase
figura 2B) en el interior de los glóbulos rojos (Quiroz, 2005). Los gametocitos
con capacidad de gametogénesis surgen cerca de 2 o 3 días después de la
penetración de los merozoitos en los eritrocitos. Para diferenciar los
macrogametocitos de los microgametocitos hay que prestar atención a la
distribución y al número de gránulos de pigmento. En los macrogametocitos,
Macrogametocito
Microgametocito

11
los gránulos se encuentran de forma homogénea por todo el citoplasma y
núcleo. Los gránulos de pigmento de los microgametocitos tienden a agregarse
en grandes (más de 1 micrometro de diámetro) masas compactas sobre todo
en los polos y, como resultado, los gránulos son más grandes. El número de
gránulos en microgametocitos es aproximadamente la mitad que en
macrogametocitos (Valkiūnas, 2004).
Los ciclos de vida de hemosporidios (reino Chromalveolata, superfilo Alveolata,
filo Apicomplexa, clase Aconoidasida, orden Haemospororida, géneros más
importantes: Haemoproteus, Leucocytozoon y Plasmodium (Morrison, 2009))
son hemoparásitos obligados heteroxénicos y complicados (Valkiūnas, 2004).
Figura 3. Ciclo vital de hemoparásitos aviares (Matta et al., 2001).
En los haemoproteidos, el ciclo de vida comienza cuando el insecto
(hospedador definitivo) ingiere sangre que contiene macro y microgametocitos
(véase figura 3). En el intestino se induce una serie compleja de eventos,
donde los gametocitos maduran. Los microgametocitos producen de 8 a 12
células pequeñas en forma de hilo -microgametos-, que fecundan al
macrogameto. La fertilización origina zigotos móviles (ooquinetos), que
penetran las células epiteliales del intestino, originando un ooquiste. Cuando el
ooquiste madura, libera los esporozoitos (forma parasítica infectiva) al

12
hemocele. Estos migran y se acumulan en las glándulas salivares del insecto
que infecta al ave en la siguiente picadura (Atkinson, 1989).
En el ave (huésped intermedio), los esporozoitos pueden infectar el pulmón,
pasando a una forma asexual de reproducción denominada esquizonte. El
proceso final de maduración del esquizonte culmina con la liberación de cientos
de merozoitos (formas parasíticas infectantes). Estos pueden reinfectar los
tejidos sólidos iniciales, o migrar a la sangre y desarrollarse en gametocito
dentro de los glóbulos rojos. Los gametocitos se desarrollan hasta diferenciarse
morfológicamente en macrogametocitos y microgametocitos (Fallis y Desser,
1977). La infección se produce entre 9-12 meses. Recientemente se encontró
una relación entre los megaloesquizontes (esquizontes gigantes con más de
1.000 merozoitos) y reacciones inflamatorias en loros y pavos (Atkinson, 1988).
Cabe señalar que el modo de infección en aves (picadura de la mosca,
intramuscular, intravenosa, intraperitoneal y la inoculación de esporozoitos)
influye en la dinámica de la parasitemia (Valkiūnas, 2004).
3. OBJETIVOS.
El principal objetivo de este estudio es la búsqueda e identificación de
hemoparásitos en sangre de ave, en concreto del orden Columbiformes.
Otros objetivos llevados a cabo son:
- Establecer la frecuencia relativa de presencia de macrogametocitos y
microgametocitos, de las especies de Hemosporidios encontradas en
las muestras sanguíneas mediante la utilización de microscopio
óptico (40x y 100x).
- Realizar recuentos automáticos de eritrocitos, glóbulos blancos y
gametocitos mediante la toma de fotografías de las muestras
positivas con el uso del programa ImageJ.

13
- Calcular el índice de parasitemia presente en los individuos
parasitados por especies de Hemospororidos (géneros
Haemoproteus, Leucocytozoon y Plasmodium) en ambas
poblaciones estudiadas (Mancha Real y Fuerte del Rey).
- Comparación del nivel de eritrocitos, glóbulos blancos y nivel de
parasitemia entre las dos poblaciones (Mancha Real y Fuerte del
Rey).
- Comparación del nivel de eritrocitos, glóbulos blancos y nivel de
parasitemia en la población de Fuerte del Rey en distintos periodos
de tiempo.
4. MATERIALES Y MÉTODOS.
4.1. Situación geográfica de los puntos de muestreo.
La recogida de muestras sanguíneas se llevó a cabo durante el mes de agosto
del 2014. Posteriormente, una segunda recogida tuvo lugar a finales del mes
de octubre de ese mismo año. Dichas muestras pertenecían a aves de cuyos
palomares se encontraban en condiciones de semilibertad y en dos localidades
de Jaén, Mancha Real y Fuerte del Rey (véase figura 4).

14
Figura 4. Mapa de España a escala 1:25. Los puntos de muestreo son
designados por una letra indicada en el mapa, habiendo en la localidad de
Fuerte del Rey dos de ellos, en tiempos diferentes. (Google maps).
En cada municipio se tomaron un número determinado de muestras,
atendiendo al número de ejemplares disponibles. Una primera recogida fue en
Mancha Real y Fuerte del Rey. La segunda recogida se llevó a cabo en la
localidad de Fuerte del Rey sólo, y se hizo sobre algunos de los individuos que
ya se les había hecho en la primera recogida durante el mes de agosto (véase
tabla 1), en concreto a 35 individuos, además de a dos nuevos ejemplares.
15
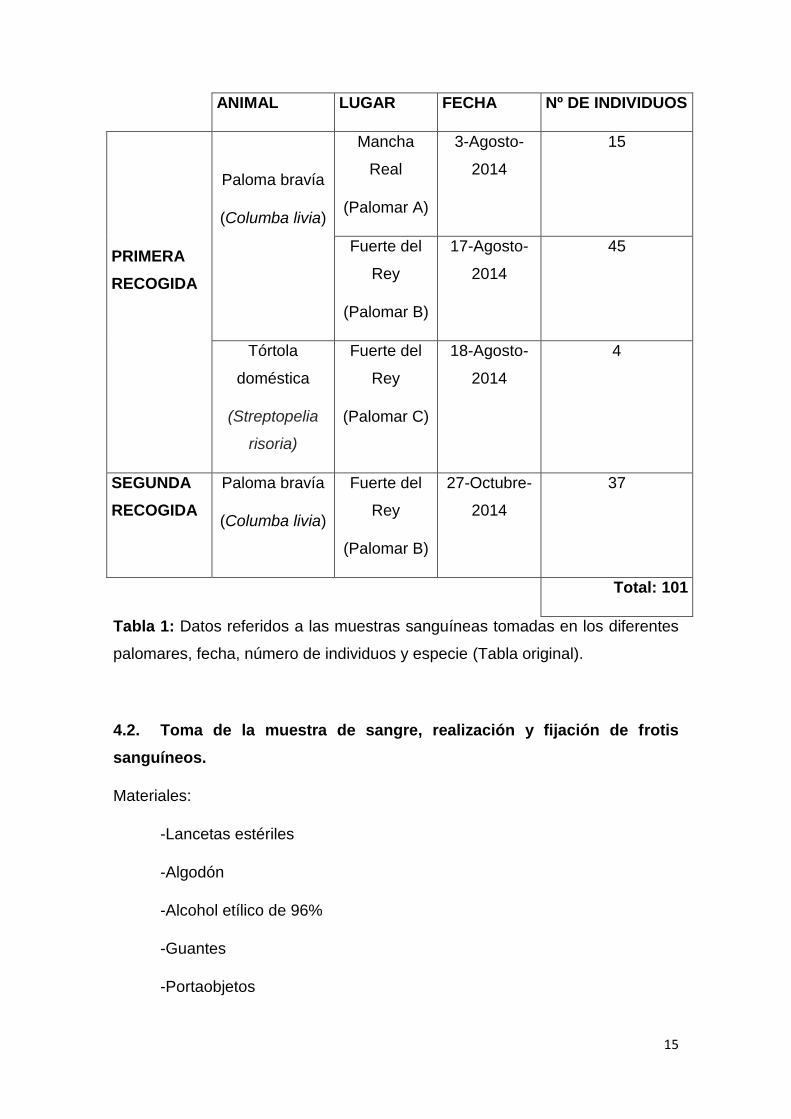
ANIMAL LUGAR FECHA Nº DE INDIVIDUOS
PRIMERA
RECOGIDA
Paloma bravía
(Columba livia)
Mancha
Real
(Palomar A)
3-Agosto-
2014
15
Fuerte del
Rey
(Palomar B)
17-Agosto-
2014
45
Tórtola
doméstica
(Streptopelia
risoria)
Fuerte del
Rey
(Palomar C)
18-Agosto-
2014
4
SEGUNDA
RECOGIDA
Paloma bravía
(Columba livia)
Fuerte del
Rey
(Palomar B)
27-Octubre-
2014
37
Total: 101
Tabla 1: Datos referidos a las muestras sanguíneas tomadas en los diferentes
palomares, fecha, número de individuos y especie (Tabla original).
4.2. Toma de la muestra de sangre, realización y fijación de frotis
sanguíneos.
Materiales:
-Lancetas estériles
-Algodón
-Alcohol etílico de 96%
-Guantes
-Portaobjetos

16
-Metanol
-Cajas para transportar las muestras
-Lápiz
-Cuaderno de campo
-Pinzas
-Tubos Falcon
-Rotulador permanente
Para la recogida de muestras sanguíneas, es necesaria la obtención de sangre
a partir de una pequeña incisión. Una vez conseguida, se realizan dos frotis
sanguíneos por cada individuo. Para ello, se sigue el siguiente protocolo:
-Desinfección con un algodón y alcohol etílico al 96% en la zona donde se
realizará la punción (véase figura 5).
Figura 5. Fotografía en la
que se desinfecta la pata del
ave en la que se actuará
para la obtención de sangre
(Fotografía original).
-Se llevará a cabo la incisión por medio de una lanceta estéril en la vena
metatarsal media, bien en la pata derecha o izquierda del ave (véase figura 6A,
6B y 6C).
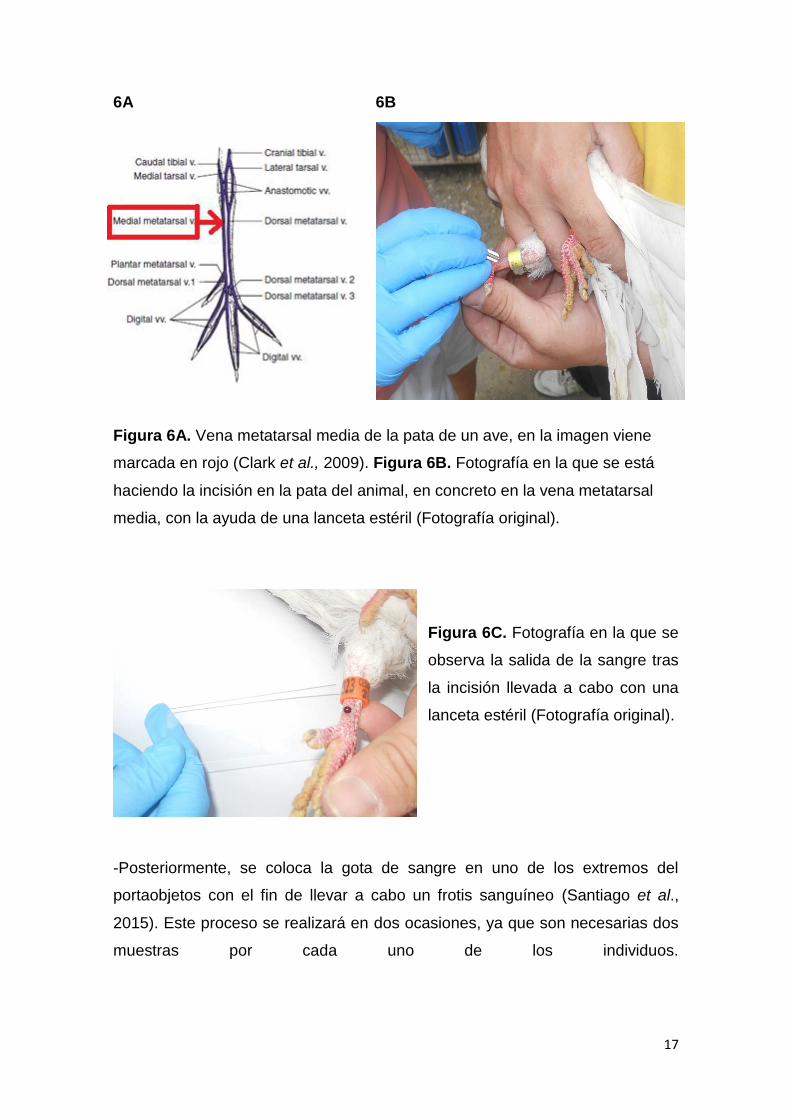
17
6A 6B
Figura 6A. Vena metatarsal media de la pata de un ave, en la imagen viene
marcada en rojo (Clark et al., 2009). Figura 6B. Fotografía en la que se está
haciendo la incisión en la pata del animal, en concreto en la vena metatarsal
media, con la ayuda de una lanceta estéril (Fotografía original).
Figura 6C. Fotografía en la que se
observa la salida de la sangre tras
la incisión llevada a cabo con una
lanceta estéril (Fotografía original).
-Posteriormente, se coloca la gota de sangre en uno de los extremos del
portaobjetos con el fin de llevar a cabo un frotis sanguíneo (Santiago et al.,
2015). Este proceso se realizará en dos ocasiones, ya que son necesarias dos
muestras por cada uno de los individuos.
18
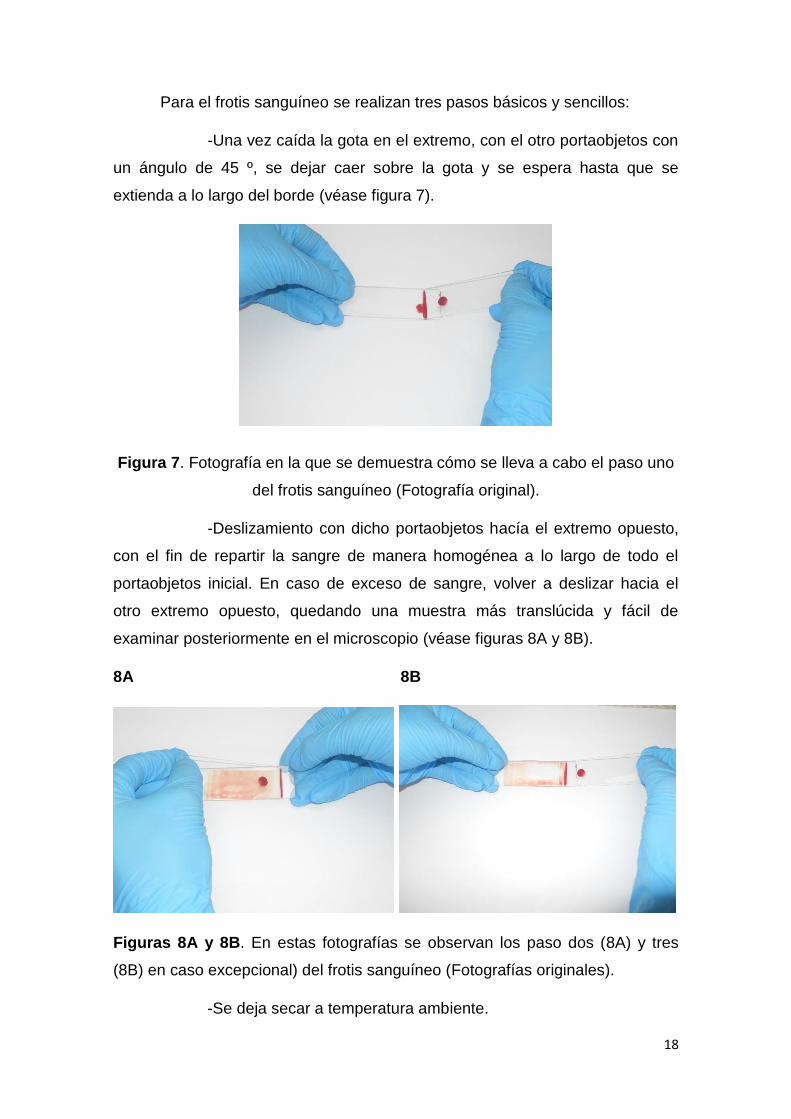
Para el frotis sanguíneo se realizan tres pasos básicos y sencillos:
-Una vez caída la gota en el extremo, con el otro portaobjetos con
un ángulo de 45 º, se dejar caer sobre la gota y se espera hasta que se
extienda a lo largo del borde (véase figura 7).
Figura 7. Fotografía en la que se demuestra cómo se lleva a cabo el paso uno
del frotis sanguíneo (Fotografía original).
-Deslizamiento con dicho portaobjetos hacía el extremo opuesto,
con el fin de repartir la sangre de manera homogénea a lo largo de todo el
portaobjetos inicial. En caso de exceso de sangre, volver a deslizar hacia el
otro extremo opuesto, quedando una muestra más translúcida y fácil de
examinar posteriormente en el microscopio (véase figuras 8A y 8B).
8A 8B
Figuras 8A y 8B. En estas fotografías se observan los paso dos (8A) y tres
(8B) en caso excepcional) del frotis sanguíneo (Fotografías originales).
-Se deja secar a temperatura ambiente.

19
-Una vez acabados los pasos anteriores, se vuelve a colocar un algodón
embebido de alcohol etílico y se le presiona en la zona de la incisión durante
unos segundos para cesar la salida de sangre (véase figura 9).
Figura 9. En esta fotografía se observa la presión con un algodón en la pata
del ave, en el lugar donde se tuvo la incisión (Fotografía original).
-Marcaje del portaobjetos con un lápiz poniéndole el número de identificación
de cada muestra (véase figura 10). Posteriormente, también se apunta en el
cuaderno de campo, dicho número de identificación, con su fecha y lugar
determinado (véase anexo 1).

20
Figura 10. Identificación de la
muestra en el portaobjetos
(Fotografía original).
-Fijar con metanol absoluto la muestra durante 15-20 segundos. Para ello se
requiere el uso de pinza y guantes (véase figura 11).
Figura 11. En esta fotografía se está
introduciendo la muestra, en un tubo
Falcon, para su fijación con metanol
absoluto (Fotografía original).
-Finalmente, colocar cada una de las muestras por orden de identificación, en
la caja especial para transportar los portaobjetos (véase figura 12).

21
Figura 12. Fotografía en la que se observan todas las muestras sanguíneas
colocadas ordenadamente, con su identificación determinada (Fotografía
original).
4.3. Tinción de Giemsa.
Recomendada para la detección e identificación de parásitos sanguíneos. En
mi caso, los frotis sanguíneos han sido teñidos con Giemsa al
2,5%.(Organización Mundial de la Salud, 1992).
1. Stock 100X Tampón Fosfato 0,67 M
Na2HPO4 59,24 g
Na2HPO4H2O 36,38 g
Agua desionizada 1000,00 ml
Autoclavar o esterilizar por filtrado (poro de 0,2 µm). El tampón estéril es
estable a temperatura ambiente durante 1 año.
2. Sol. trabajo tampón Fosfato 0,0067 M, pH 7,2
Stock tampón Fosfato (100x) 10,0 ml
Agua desionizada 990,0 ml

22
Comprobar el pH antes de su uso. Debe ser pH 7,2. Estable a
temperatura ambiente durante 1 mes.
3. Stock tinción de Giemsa (El colorante Giemsa está disponible
comercialmente. Teóricamente no caduca).
TÉCNICA DE TRABAJO
1. Solución de trabajo de colorante Giemsa (2,5%): haga solución fresca
para cada lote de extensiones.
Tampón de trabajo Giemsa: 195 ml
Sol. stock colorante Giemsa: 5 ml
Tinción
1. Preparar solución de trabajo de Giemsa en un vaso de tinción, de
acuerdo con las instrucciones anteriores. Debe considerarse que 200 ml
son suficientes para llenar un vaso Coplin horizontal, para recipientes de
otras dimensiones deben adaptarse los volúmenes mencionados para
no modificar las proporciones.
2. Coloque 200 ml de tampón de trabajo Giemsa en un segundo vaso
Coplin horizontal. Adapte el volumen a las dimensiones del recipiente
que esté utilizando.
3. Coloque los portaobjetos en la solución de trabajo Giemsa (2.5%)
durante 45-60 minutos (véase figuras 13A y 13B).

23
13A 13B
Figuras 13A y 13B. Fotografías en las que aparecen las preparaciones
durante su tinción (13A) y tras 45-60 minutos (13B) (Fotografías
originales).
4. Saque las preparaciones y lávelas sumergiéndolas 3-4 veces en el
tampón Giemsa. Las gotas gruesas deben mantenerse en el tampón
durante 5 minutos (véase figura 14).
Figura 14 Lavado de las preparaciones
tras su tinción (Fotografía original).
5. Dejar secar las preparaciones boca arriba a temperatura ambiente
(véase figura 15).
Figura 15. Fotografía con las preparaciones secándose a temperatura
ambiente (Fotografía original).

24
Nota: Para reducir los tiempos de tinción pueden utilizarse soluciones más
concentradas de Giemsa, de modo que podemos reducir el tiempo de tinción
desde los 45-60 minutos en la solución de tinción Giemsa al 2.5%, hasta 10
minuto en una solución Giemsa al 10%. Debe considerar que acortar los
tiempos de tinción mediante un incremento en la concentración de colorante
puede suponer una reducción en la predictibilidad de la calidad de los
resultados.
4.4. Observaciones al microscopio y toma de fotografías de las
muestras positivas.
Una vez teñidas las muestras mediante la tinción de Giemsa explicada
anteriormente, se procede al análisis en el microscopio óptico bajo aceite de
inmersión de todas las preparaciones, para posteriormente clasificarlas en
positivas (parasitadas) o negativas (no parasitadas), quedándonos así con las
positivas. Todo esto era anotado en la ficha de observación al microscopio
(véase anexo 2).
La metodología de observación en cada una de las preparaciones es básica, y
consiste en barridos en zig-zag por distintos campos al azar (véase figura 16).
El número de campos por cada preparación oscila en 20. En este proceso se
pretende además de la búsqueda de gametocitos de hemosporidios, su
identificación a nivel de especie.
Figura 16. Recorrido que se sigue
en cada uno de los campos de cada
frotis sanguíneo para la búsqueda e
identificación de gametocitos de
hemosporidios (Fotografía original).

25
Posteriormente, se lleva a cabo la toma de fotografías de las preparaciones
que son positivas con hemosporidios. Para ello, se requiere de un microscopio
óptico con cámara incorporada (Olympus) y utilizando el objetivo de 100x,
además del uso del aceite de inmersión y de la técnica de barrido explicada
anteriormente (véase figura 16). Por cada preparación se toma un número de
fotografías variable, entre 30-50 dependiendo de la calidad del frotis sanguíneo.
El hecho de la toma de dichas fotografías al azar implica que pueda ocurrir que
una muestra siendo positiva, a la hora de analizar sus campos no aparezca
ningún parásito. Esto se debe a un nivel de parasitemia muy bajo.
La técnica de procesado de imágenes se llevará a cabo mediante el software
ImageJ que se explicará en el siguiente apartado.
4.5. Procesamiento de imágenes y recuento de eritrocitos, glóbulos
blancos y gametocitos.
Tras la obtención de las fotografías parasitadas se procede al recuento de
eritrocitos tanto sanos como parasitados. Además del recuento de glóbulos
blancos y gametocitos, diferenciando entre macrogametocitos y
microgametocitos. Para llevar a cabo este recuento se ha requerido de la
utilización de un programa conocido como ImageJ (Abramoff et al., 2004). Todo
esto queda reflejado en una tabla tipo Excel (véase Anexo 3).
El tutorial de dicho programa a utilizar es el siguiente:
- Abrir el programa en un ordenador en el que fue instalado previamente.
- Para abrir la imagen objetivo, en el menú de ImageJ, se debe pinchar en
“File” “Open”, eligiendo la imagen que queremos analizar.
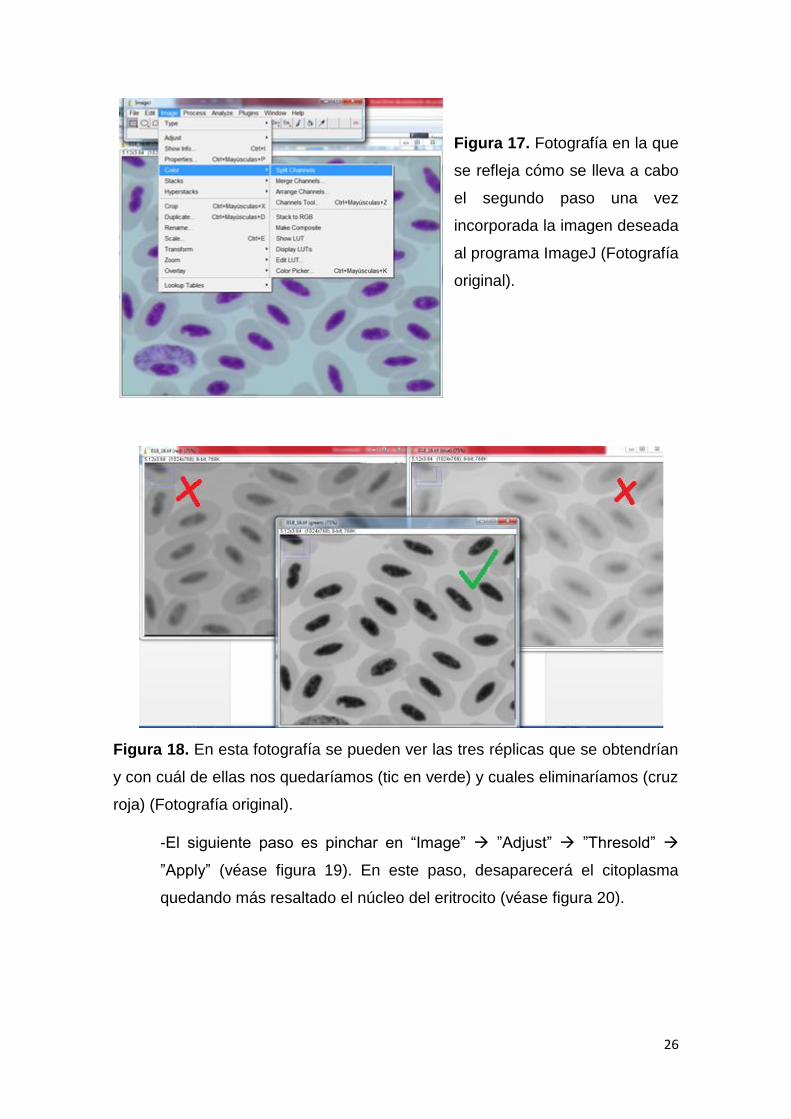
- Después de tener la imagen abierta con ImageJ, se pincha en el menú
en “Image” ”Color” ”Split Channels” (véase figura 17). Tras estos
clickeos, aparecerán tres réplicas en distintas tonalidades de grises. La
imagen que se escoge es en la que resalte más el núcleo de los
eritrocitos (véase figura 18).
26
Figura 17. Fotografía en la que
se refleja cómo se lleva a cabo
el segundo paso una vez
incorporada la imagen deseada
al programa ImageJ (Fotografía
original).
Figura 18. En esta fotografía se pueden ver las tres réplicas que se obtendrían
y con cuál de ellas nos quedaríamos (tic en verde) y cuales eliminaríamos (cruz
roja) (Fotografía original).
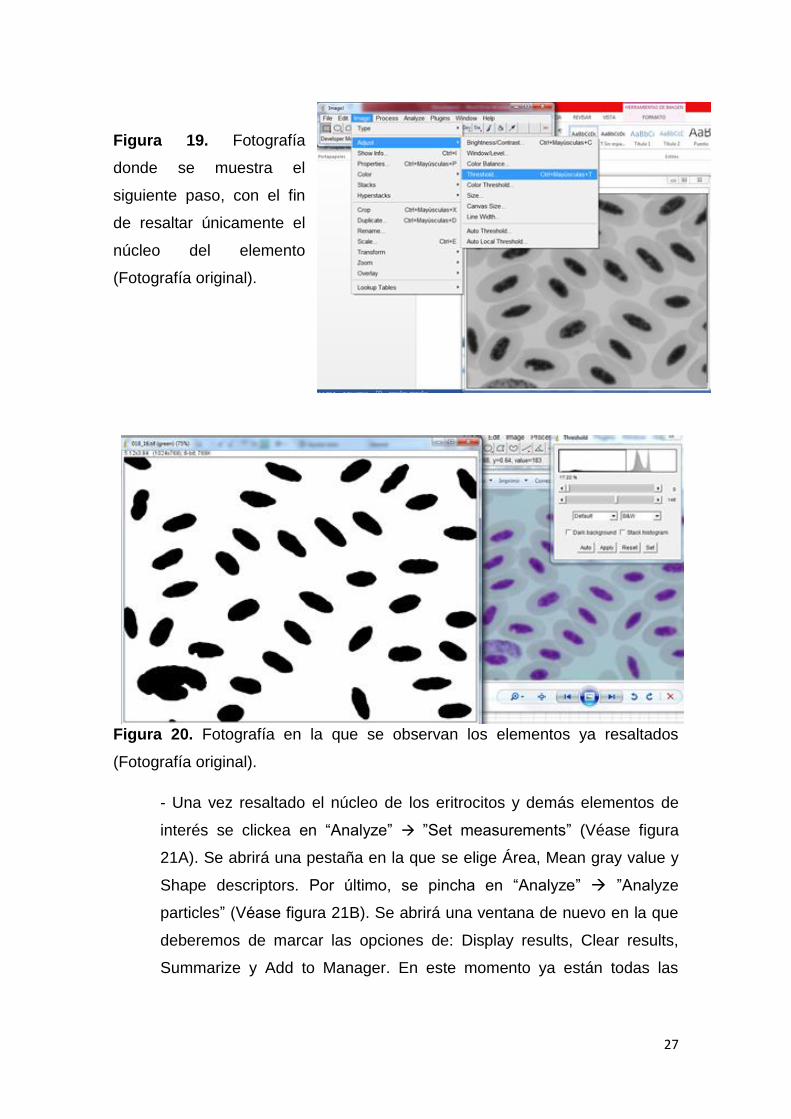
-El siguiente paso es pinchar en “Image” ”Adjust” ”Thresold”
”Apply” (véase figura 19). En este paso, desaparecerá el citoplasma
quedando más resaltado el núcleo del eritrocito (véase figura 20).
27
Figura 19. Fotografía
donde se muestra el
siguiente paso, con el fin
de resaltar únicamente el
núcleo del elemento
(Fotografía original).
Figura 20. Fotografía en la que se observan los elementos ya resaltados
(Fotografía original).
- Una vez resaltado el núcleo de los eritrocitos y demás elementos de
interés se clickea en “Analyze” ”Set measurements” (Véase figura
21A). Se abrirá una pestaña en la que se elige Área, Mean gray value y
Shape descriptors. Por último, se pincha en “Analyze” ”Analyze
particles” (Véase figura 21B). Se abrirá una ventana de nuevo en la que
deberemos de marcar las opciones de: Display results, Clear results,
Summarize y Add to Manager. En este momento ya están todas las

28
características necesarias seleccionadas para que se inicie el recuento
de los elementos que nos interesa.
21A 21B
Figuras 21A y 21B. En estas fotografías se observan los últimos pasos a
realizar indicados anteriormente (Fotografías originales).
- Al aplicar el último paso, se abrirá automáticamente una ventana con el
número de eritrocitos que corresponde a dicha imagen analizada (véase figura
22).
Figura 22. Fotografía en la que aparecen los elementos resaltados y
enumerados (Fotografía original).
-Todos estos pasos se repiten en cada una de las imágenes que
queremos llevar a cabo el recuento de elementos de nuestro interés.

29
4.6. Análisis estadístico.
Utilizando Excel se han calculado una serie de parámetros que nos permiten
comparar las dos poblaciones a estudiar, Mancha Real y Fuerte del Rey,
además de las dos subpoblaciones estudiadas en diferentes periodos de
tiempo en Fuerte del Rey. Para ello calculamos la media, la varianza y la
desviación estándar tanto de número de eritrocitos, como de glóbulos blancos y
gametocitos. Además del número de campos analizados en total y número de
parásitos por cada mil eritrocitos.
5. RESULTADOS.
5.1. Observación frotis sanguíneo.
5.1.1 Determinación del nivel de parasitemia.
De las 101 muestras de sangre en aves que fueron examinadas, sólo en 27 de
ellas se han observado hemoparásitos del género Haemoproteus. De esas 101
muestras, 97 pertenecen a paloma doméstica (Columba livia) y 4 provienen de
tórtola doméstica (Streptopelia risoria), resultando libres de hemoparásitos
estas últimas. Por lo que, todas las muestras parasitadas son de paloma
doméstica.
Tras el examen de todos esos frotis sanguíneos en el microscopio óptico, se
observa que el 26, 73 % del total (n=101) se encuentran parasitadas, en
concreto por Haemoproteus columbae. De los animales parasitados (n=27), el
37% presenta un nivel de parasitemia muy bajo, el 41% presenta un nivel de
parasitemia bajo, el 4% presenta un nivel de parasitemia medio, el 7% presenta
un nivel de parasitemia alto y por último, el 11% presenta un nivel de
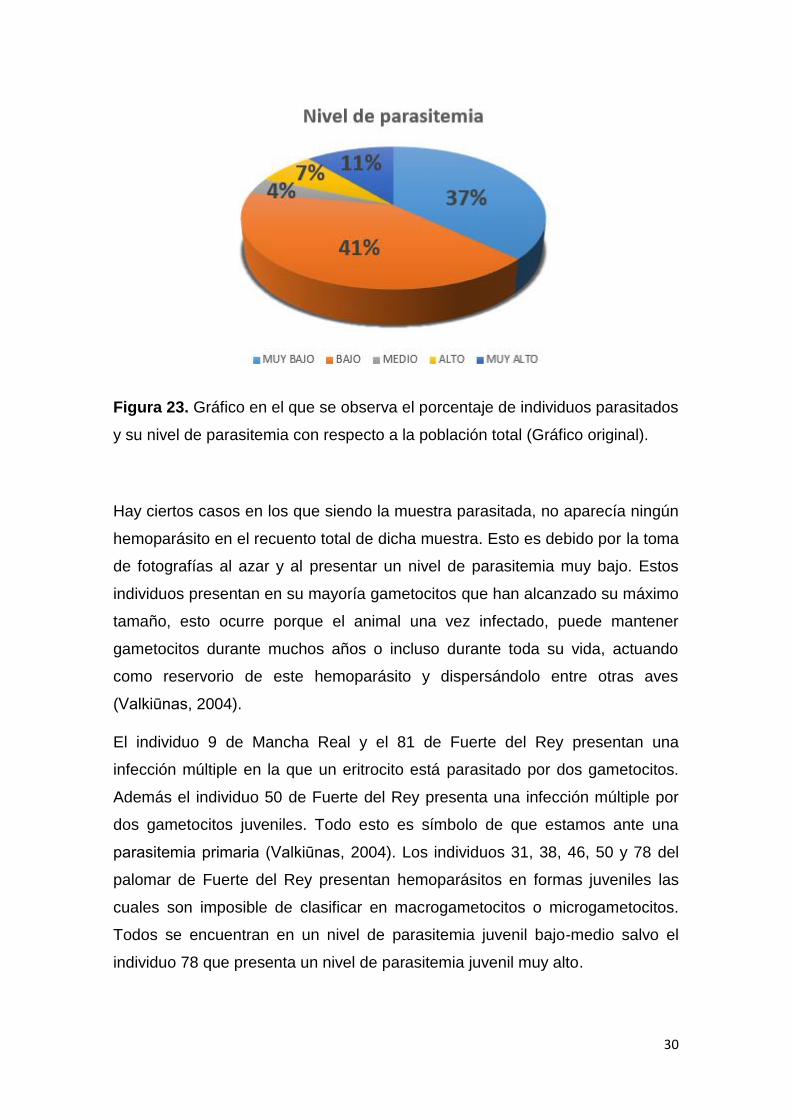
parasitemia muy alto (véase figura 23).
Si comparamos el nivel de parasitemia de cada población, se observa que en
Mancha Real el 53,33 % del total de sus muestras están parasitadas, mientras
que Fuerte del Rey presenta el 22.09% de muestras parasitadas. Claramente
podemos ver el grado de infección alto que muestra Mancha Real con respecto
a la otra población.
30
Figura 23. Gráfico en el que se observa el porcentaje de individuos parasitados
y su nivel de parasitemia con respecto a la población total (Gráfico original).
Hay ciertos casos en los que siendo la muestra parasitada, no aparecía ningún
hemoparásito en el recuento total de dicha muestra. Esto es debido por la toma
de fotografías al azar y al presentar un nivel de parasitemia muy bajo. Estos
individuos presentan en su mayoría gametocitos que han alcanzado su máximo
tamaño, esto ocurre porque el animal una vez infectado, puede mantener
gametocitos durante muchos años o incluso durante toda su vida, actuando
como reservorio de este hemoparásito y dispersándolo entre otras aves
(Valkiūnas, 2004).
El individuo 9 de Mancha Real y el 81 de Fuerte del Rey presentan una
infección múltiple en la que un eritrocito está parasitado por dos gametocitos.
Además el individuo 50 de Fuerte del Rey presenta una infección múltiple por
dos gametocitos juveniles. Todo esto es símbolo de que estamos ante una
parasitemia primaria (Valkiūnas, 2004). Los individuos 31, 38, 46, 50 y 78 del
palomar de Fuerte del Rey presentan hemoparásitos en formas juveniles las
cuales son imposible de clasificar en macrogametocitos o microgametocitos.
Todos se encuentran en un nivel de parasitemia juvenil bajo-medio salvo el
individuo 78 que presenta un nivel de parasitemia juvenil muy alto.

31
El criterio que se ha llevado a cabo para establecer el nivel de parasitemia es el
siguiente (véase tabla 2):
Nivel de
parasitemia
Rango de
parásitos
Parásitos totales
por 1000
eritrocitos
Porcentaje
nivel de
parasitismo
Muy bajo 0-5 10 37.04 %
Bajo 6-14 11 40.74%
Medio 15-20 1 3.70 %
Alto 21-26 2 7.41 %
Muy alto 26 o más 3 11.11 %
n=27
5.1.2. Recuento de campos analizados.
Se realizó un recuento de gametocitos (macrogametocitos y microgametocitos)
con un número variable de campos dependiendo de la calidad del frotis
sanguíneo, entre 30-50 por cada muestra. Además de un recuento de
eritrocitos y glóbulos blancos, con el objetivo de ver la relación entre ellos en
las dos poblaciones estudiadas de diferente lugar (Fuerte del Rey y Mancha
Real) y en la misma población en diferentes periodos de tiempo (Fuerte del
Rey).
El número de campos total analizados en cada población fue diferente,
dependiendo como se dijo anteriormente, de la calidad del frotis sanguíneo
(véase tabla 3).
Tabla 2. En ella se observa el criterio a seguir para establecer el nivel de
parasitemia (tabla original).

32
Situación geográfica Nº de campos analizados
Mancha Real 310
Fuerte del Rey (Mediados agosto) 340
Fuerte del Rey (Finales octubre) 335
n=985
Tabla 3. Número de campos analizados en el total de todas las muestras
dependiendo del palomar (tabla original).
5.1.3. Recuento de eritrocitos, glóbulos blancos y gametocitos.
Se lleva a cabo un recuento de eritrocitos totales por muestra teniendo en
cuenta que en cada una podemos encontrarnos entre 30-50 campos
analizados, dependiendo de la calidad del frotis sanguíneo. El recuento de
eritrocitos se ha llevado a cabo en todas aquellas muestras que estaban
parasitadas por hemosporidios. Al igual que con el recuento de eritrocitos, se
hace un recuento de glóbulos blancos, macrogametocitos y microgametocitos
(véase tabla 4).

33
Muestra
Palomar
Nº
Eritrocitos
total
Nº glóbulos
blancos
total
Macro-
gametos
Micro-
gametos
Relación
macro
/
micro
Nº
campos
1 A 3325 21 17 6 2.83 40
4 A 2799 19 7 0 - 35
7 A 1465 32 1 1 1 50
8 A 1825 12 1 1 1 35
9 A 1904 10 25 11 2.27 45
10 A 1624 16 14 16 0.875 35
12 A 1951 4 7 7 1 40
13 A 2049 14 4 8 0.5 30
21 B 1490 9 5 6 0.092 40
22 B 1540 4 0 1 0 40
27 B 2026 0 0 0 0 40
28 B 1380 1 5 1 5 40
31 B 1525 25 5 2 2.5 40
38 B 1616 4 8 2 4 30
46 B 1391 13 2 4 0.5 40
50 B 1509 1 33 5 6.6 30
51 B 1253 4 4 0 - 35
69 B 1269 8 10 8 1.25 40
72 B 1629 5 6 1 6 30
76 B 1139 5 0 0 0 40
78 B 2038 12 - - - 30
79 B 1516 9 8 3 2.67 30
81 B 1149 10 8 6 1.33 40
87 B 1668 7 2 3 0.67 30
90 B 1016 7 2 0 - 30
93 B 1275 20 2 2 1 30
94 B 1137 5 1 2 0.5 40
Tabla 4. Número de eritrocitos, glóbulos blancos, macrogametocitos y
microgametocitos que hay en todos los campos de cada muestra parasitada
analizada (tabla original).
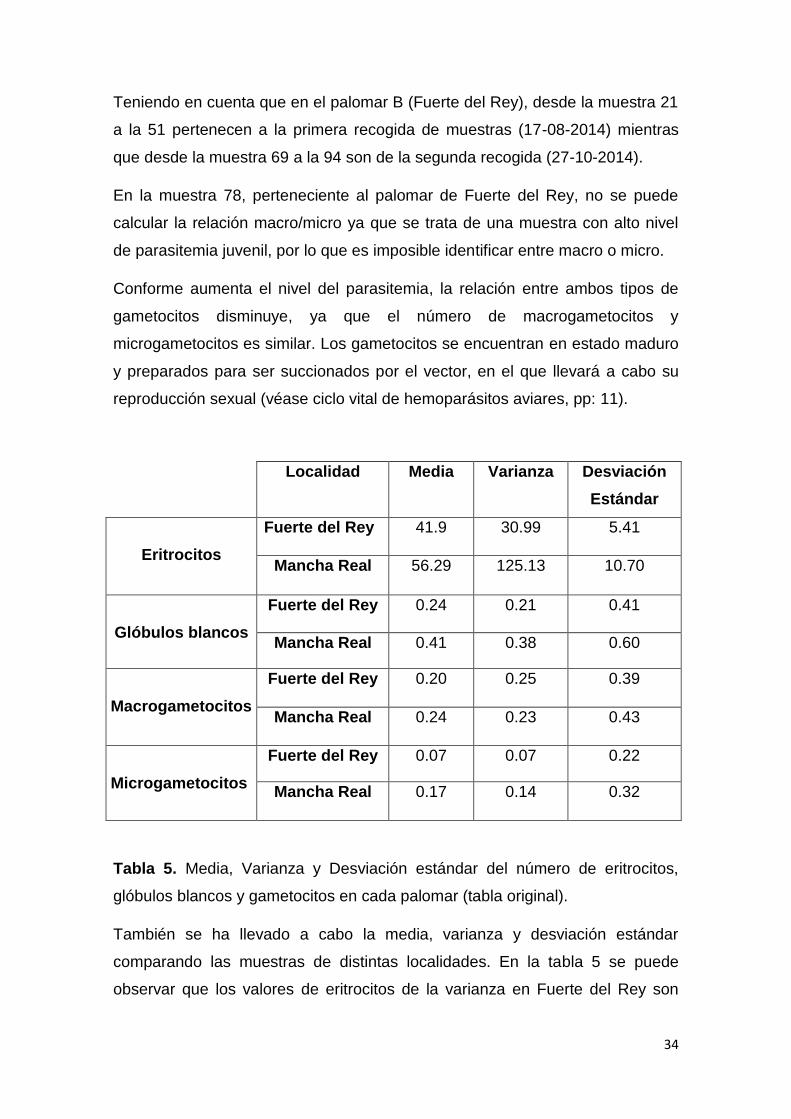
34
Teniendo en cuenta que en el palomar B (Fuerte del Rey), desde la muestra 21
a la 51 pertenecen a la primera recogida de muestras (17-08-2014) mientras
que desde la muestra 69 a la 94 son de la segunda recogida (27-10-2014).
En la muestra 78, perteneciente al palomar de Fuerte del Rey, no se puede
calcular la relación macro/micro ya que se trata de una muestra con alto nivel
de parasitemia juvenil, por lo que es imposible identificar entre macro o micro.
Conforme aumenta el nivel del parasitemia, la relación entre ambos tipos de
gametocitos disminuye, ya que el número de macrogametocitos y
microgametocitos es similar. Los gametocitos se encuentran en estado maduro
y preparados para ser succionados por el vector, en el que llevará a cabo su
reproducción sexual (véase ciclo vital de hemoparásitos aviares, pp: 11).
Tabla 5. Media, Varianza y Desviación estándar del número de eritrocitos,
glóbulos blancos y gametocitos en cada palomar (tabla original).
También se ha llevado a cabo la media, varianza y desviación estándar
comparando las muestras de distintas localidades. En la tabla 5 se puede
observar que los valores de eritrocitos de la varianza en Fuerte del Rey son
Localidad Media Varianza Desviación
Estándar
Eritrocitos
Fuerte del Rey 41.9 30.99 5.41
Mancha Real 56.29 125.13 10.70
Glóbulos blancos
Fuerte del Rey 0.24 0.21 0.41
Mancha Real 0.41 0.38 0.60
Macrogametocitos
Fuerte del Rey 0.20 0.25 0.39
Mancha Real 0.24 0.23 0.43
Microgametocitos
Fuerte del Rey 0.07 0.07 0.22
Mancha Real 0.17 0.14 0.32
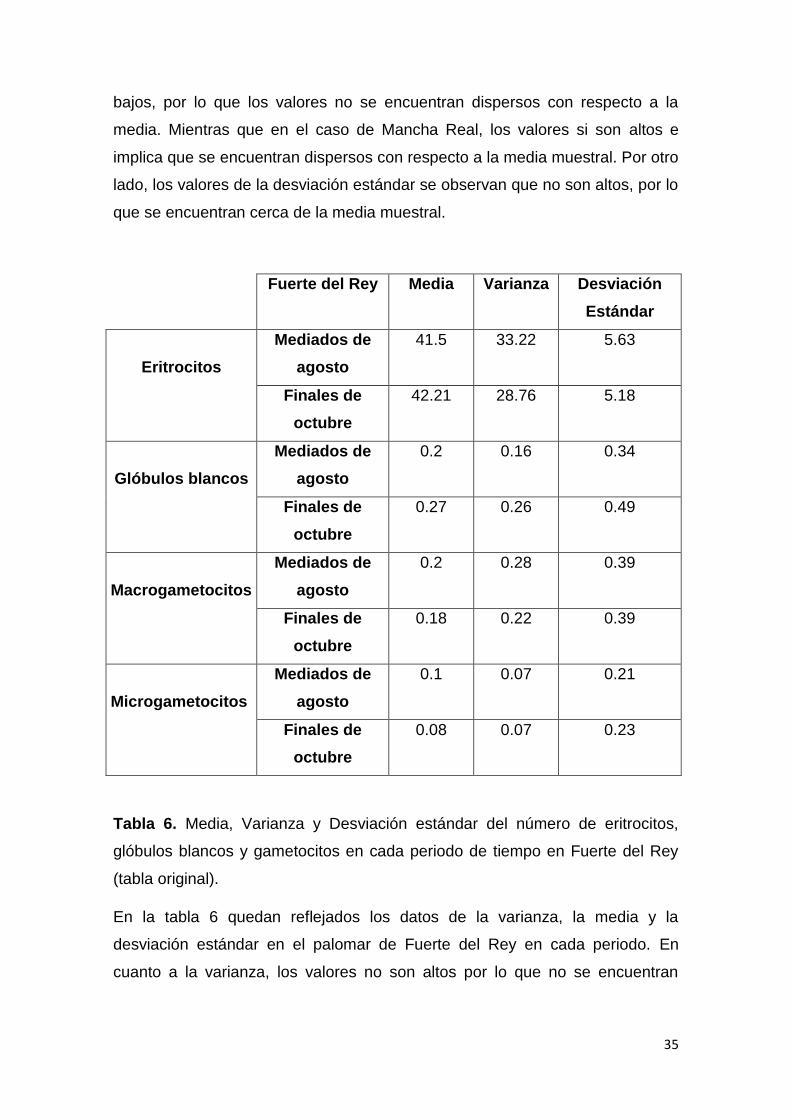
35
bajos, por lo que los valores no se encuentran dispersos con respecto a la
media. Mientras que en el caso de Mancha Real, los valores si son altos e
implica que se encuentran dispersos con respecto a la media muestral. Por otro
lado, los valores de la desviación estándar se observan que no son altos, por lo
que se encuentran cerca de la media muestral.
Tabla 6. Media, Varianza y Desviación estándar del número de eritrocitos,
glóbulos blancos y gametocitos en cada periodo de tiempo en Fuerte del Rey
(tabla original).
En la tabla 6 quedan reflejados los datos de la varianza, la media y la
desviación estándar en el palomar de Fuerte del Rey en cada periodo. En
cuanto a la varianza, los valores no son altos por lo que no se encuentran
Fuerte del Rey Media Varianza Desviación
Estándar
Eritrocitos
Mediados de
agosto
41.5 33.22 5.63
Finales de
octubre
42.21 28.76 5.18
Glóbulos blancos
Mediados de
agosto
0.2 0.16 0.34
Finales de
octubre
0.27 0.26 0.49
Macrogametocitos
Mediados de
agosto
0.2 0.28 0.39
Finales de
octubre
0.18 0.22 0.39
Microgametocitos
Mediados de
agosto
0.1 0.07 0.21
Finales de
octubre
0.08 0.07 0.23
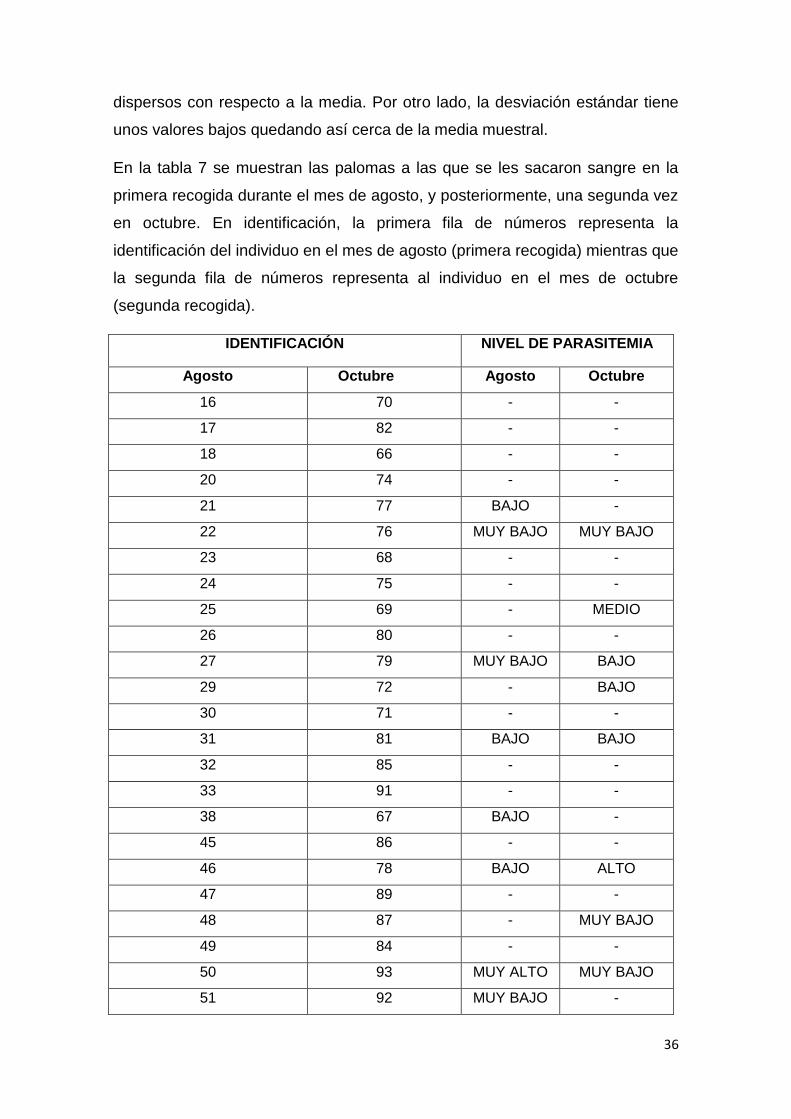
36
dispersos con respecto a la media. Por otro lado, la desviación estándar tiene
unos valores bajos quedando así cerca de la media muestral.
En la tabla 7 se muestran las palomas a las que se les sacaron sangre en la
primera recogida durante el mes de agosto, y posteriormente, una segunda vez
en octubre. En identificación, la primera fila de números representa la
identificación del individuo en el mes de agosto (primera recogida) mientras que
la segunda fila de números representa al individuo en el mes de octubre
(segunda recogida).
IDENTIFICACIÓN NIVEL DE PARASITEMIA
Agosto Octubre Agosto Octubre
16 70 - -
17 82 - -
18 66 - -
20 74 - -
21 77 BAJO -
22 76 MUY BAJO MUY BAJO
23 68 - -
24 75 - -
25 69 - MEDIO
26 80 - -
27 79 MUY BAJO BAJO
29 72 - BAJO
30 71 - -
31 81 BAJO BAJO
32 85 - -
33 91 - -
38 67 BAJO -
45 86 - -
46 78 BAJO ALTO
47 89 - -
48 87 - MUY BAJO
49 84 - -
50 93 MUY ALTO MUY BAJO
51 92 MUY BAJO -

37
54 73 - -
55 90 - MUY BAJO
56 88 - -
57 97 - -
58 100 - -
59 95 - -
60 101 - -
61 99 - -
62 94 - MUY BAJO
63 96 - -
Tabla 7. Nivel de parasitemia de los individuos del palomar de Fuerte del Rey
que se les sacó sangre en la primera y segunda recogida de muestras (Tabla
original)
De acuerdo al criterio que se siguió anteriormente, en el apartado 5.1.1 (Nivel
de parasitemia) (véase tabla 8) se designará el nivel de parasitemia que
presentan las dos subpoblaciones del municipio de Fuerte del Rey.
Tabla 8. Criterio a seguir para el nivel de
parasitemia (Tabla original).
6. DISCUSIÓN.
Tras el estudio de 101 frotis sanguíneos teñidos con Giemsa, se han detectado
hemoparásitos del género Haemoproteus, en concreto de Haemoproteus
columbae, y no se encontraron pertenecientes a otros géneros como son
Leucocytozoon y Plasmodium. Mediante el método anterior se ha logrado
detectar que el 26,73% del total de muestras (n=101) estaban parasitadas. El
parasito de sangre aviar más común según Desser y Bennett, (1993) es
Nivel de
parasitemia
Rango de
parásitos
Muy bajo 0-5
Bajo 6-14
Medio 15-20
Alto 21-26
Muy alto 26 o más

38
Haemoproteus. El motivo del porqué sólo apareció del género Haemoproteus y
no de Leucocytozoon ni Plasmodium es por la distribución geográfica de sus
vectores (Valkiūnas, 2004).
Para la búsqueda de detección de hemoparásitos, el método más eficaz
demostrado por estudios de Krone et al., (2008) es la observación de frotis
sanguíneos (Santiago et al., 2015), ya que las técnicas moleculares pueden
presentar problemas para detectar determinadas infecciones. Este estudio
podemos decir que se encuentra con valores normales de parasitemia dentro
de las poblaciones de paloma doméstica analizadas según algunos estudios
como el que llevó acabó Reza et al,. (2011), investigando en tres lugares
diferentes de Irán, (Babol, Iahijan and Firouz Koh) y encontraron que el 30 %
de palomas eran parasitadas habiendo un total de 120. Otro estudio iraní
realizado mediante un método molecular por Doosti et al., (2014) mostró que la
tasa de prevalencia de Haemoproteus columbae fue 23,18% (51/120). En
España, en Córdoba, Martínez-Moreno et al., (1989) realizaron un estudio de la
parasitofauna de 101 ejemplares, registrándose el 26,73% parasitadas por H.
columbae, como puede verse, presentando los mismos resultados del estudio
realizado en las dos poblaciones de Jaén.
Según en la estación climática en la que nos encontremos puede variar el nivel
de parasitemia. Un estudio de Klei et al., (1975) explica la razón del porqué no
existe esa diferencia en los valores de parasitemia en los diferentes periodos
de muestreo. Los picos estacionales de hemosporidios se observan
generalmente durante la primavera y el verano, y es debido a la recaída en
primavera asociado con cambios fisiológicos que ocurren durante la
reproducción o movimientos de las aves migratorias. La aparición estacional de
H. columbae difiere de otros hemosporidios en que la mayor prevalencia de
infección ocurren durante el otoño y el invierno y no existe una recaída en
primavera. La falta de recaída primavera en H. columbae puede explicarse por
la ausencia de los ciclos reproductivos estacionales, comportamiento no
migratorio de palomas y presencia del vector en todo el año. Existe una mayor
incidencia de H. columbae en las aves durante el otoño y el invierno. Esto
puede ser explicado por el aumento de los periodos de transmisión durante el

39
verano y el otoño. El porcentaje de aves jóvenes infectadas fueron más altos
durante verano y otoño, lo que indica que estas temporadas son periodos de
transmisión activa.
Las muestras sanguíneas analizadas tienen un número de macrogametocitos
mayor que el número de microgametocitos. Los estudios que investigan las
diferencias sexuales en la intensidad de parásitos en la sangre han
proporcionado variables resultados. Forbes et al., (1994) no encontraron
diferencias significativas por sexo en la prevalencia de Haemoproteus y
Plasmodium. Según Wiehn et al., (1998) los machos y las hembras pueden
diferir según los aspectos de su comportamiento que modifican la exposición a
vectores de insectos, resistencia a la infestación, y / o la capacidad de combatir
y eliminar las infecciones adquiridas tal vez como resultado de las interacciones
sexo-específicas con condiciones ambientales. Por el contrario, Deviche et al.
(2001a, b) no indicó diferencias sexuales en prevalencia de parásitos, sin
embargo, la intensidad de la infección disminuyó en los machos en verano.
Algunos han reportado que las hembras de insectos ectoparásitos son con
frecuencia más abundantes debido a su larga vida útil y capacidad de
dispersión en comparación con la de los machos (Hamilton1967; Clayton et
al.1992; Dick and Patterson, 2008).
La presencia de Haemoproteus columbae, en el ave depende principalmente
de la abundancia de moscas hipoboscídeas, ya que las palomas al ser
animales muy móviles están expuestas a las picaduras de sus vectores
(Nematollahi et al., 2012; Sol et al., 2000). Ellos están circulando
continuamente alrededor del mundo y cualquier variación de su prevalencia,
intensidad y el impacto de la salud, ya sea por sexo, estacionalidad o espacial
relacionada, podría depender de la susceptibilidad del huésped (Scaglione et
al., 2015). Numerosos factores endógenos y exógenos pueden tener una
influencia acumulativa en el estado de infección de ambos sexos de las
palomas por estos parásitos, tales como la hormonas del huésped y los
compuestos humorales, edad y nutricional condiciones, el comportamiento y
hábitos, así como la estación del año y las características ecológicas y físicas

40
de las regiones (Al-Barwari et al., 2012). Por lo general, las hembras son más
parasitados porque la reducción de la actividad de locomoción durante el
período de anidación es un factor que aumenta la probabilidad de su infección
con haemosporidians (Valkiūnas, 2004).
Un ave puede ser invadido por varios parásitos de la misma especie o de
diferentes taxones (Elahi et al., 2014). La interacción de dos o más
ectoparásitos en el mismo hospedador puede decirse que es una interacción
competitiva-interespecífica baja caracterizado por infestaciones simultáneas
que pueden no ser perjudicial para las dos especies (Adang et al., 2008).
No se han hallado datos del porque la población de Mancha Real tiene un
mayor porcentaje de parasitemia que la población de Fuerte del Rey. Sí que
podríamos decir que puede deberse por una serie de factores: como un río,
una balsa, un pantano, etc cerca del palomar, por el tratamiento que utilice el
cuidador con las palomas, en el caso de tenerlo, por la higiene de dicho
palomar o por el grado de libertad.
7. CONCLUSIONES.
-Los hemoparásitos encontrados en Columba livia pertenecen al género
Haemoproteus, en concreto a la especie H. columbae.
-De 101 muestras de sangre analizadas, el 26,73% (n=27) de palomas están
parasitadas, siendo valores normales de parasitemia según algunos estudios.
-En gran parte de las muestras estudiadas, el número de macrogametocitos es
mayor que el de microgametocitos. Aunque según estudios realizados por
varios autores se trata de un resultado variable.
-El muestreo en diferentes periodos de tiempo, en verano (agosto) y en otoño
(octubre), tiene valores similares de parasitemia. Esto es debido según Klei et
al., (1975) porque esas temporadas son los periodos de transmisión activa en
H. columbae, a diferencia de otros hemosporidios Los valores de prevalencia
registrados varían en función de la época del año en la que se realice el
muestreo (Gupta et al., 2011).

41
-La población de Mancha Real tiene un nivel de parasitemia mayor que la de
Fuerte del Rey, desconociéndose con seguridad el porqué.

42
8. REFERENCIAS BIBLIOGRÁFICAS.
1. Abramoff, M.D., Magalhaes, P.J. & Ram, S.J. 2004 .Image Processing with
ImageJ. Biophotonics International, 11 (7):36-42.
2. Adang, K.L., Oniye, S.J., Ezealor, A.U., Abdu, P.A. & Ajanusi, O.J. 2008.
Ectoparasites of Domestic Pigeon (Columba livia domestica, Linnaeus) in
Zaria, Nigeria. Research Journal Parasitology, 3(2):79–84.
3. Al-Barwari, S. & Saeed, I. 2012. The parasitic communities of the rock pigeon
Columba livia from Iraq: component and importance. Turkiye Parazitoloji
Dergisi, 36(4):232–239.
4. Atkinson, C.T. 1988. Epizootiology of Haemoproteus meleagridis (Protozoa:
Haemosporida) in Florida: potential vectors and prevalence in naturally
infected Culicoides (Diptera: Ceratopogonidae). Journal of Medical
Entomology, 25(1):39-44.
5. Atkinson, C.T. 1989. Ultrastructure of the ookinetes of Haemoproteus
meleagridis (Haemosporida: Haemoproteidae). Journal of Parasitology,
75(1): 135-141.
6. Clayton, D.H., Gregory, R.D. & Price, R.D. 1992. Comparative ecology of
Neotropical bird lice (Insecta: Phthiraptera). Journal Animal Ecology.
61(3):781–795.
7. Desser, S. S. & Bennett, G. F. (1993). The genera Leucocytozoon,
Haemoproteus and Hepatocystis. In parasitic protozoa. (2nd eds) Vol. IV.
Ed. Kreier J. Academic Press. New York, pp. 273-307.
8. Deviche, P., Greiner, E.C, & Manteca, H. (2001a). Interspecific variability of
prevalence in blood parasites of adult passerine birds during the breeding
season in Alaska. Journal of Wildlife Diseases, 37(1):28–35.

43
9. Deviche, P., Greiner, E.C. & Manteca, H. (2001b) Seasonal and age-related
changes in blood parasite prevalence in dark-eyed Juncos (Juncohy
emalis, Aves, Passeriformes). Journal of Experimental Zoology,
289(7):456–466.
10. Dick, C.W. & Patterson, B.D. 2008. An excess of males: skewed sex ratios
in bat flies (Diptera:Streblidae). Evolutionary Ecology, 22(6):757–769.
11. Doosti, A., Ahmadi, R., Mohammadalipour, Z & Zohoor, A .2014. Detection
of Haemoproteus columbae in iranian pigeons using PCR. International
Conference on Biological, Civil and Environmental Engineering (BCEE).
Dubai.
12. Elahi, R., Islam, A., Hossain, M.S., Mohiuddin, K., Mikolon, A., Paul, S.K., et
al., 2014. Prevalence and diversity of avian haematozoan parasites in
wetlands of Bangladesh. Journal of Parasitology Research, 2014:1-12.
13. Fallis, A. M. & Desser, S. S. 1977. On species of Leucocytozoon,
Haemoproteus, and Hepatocystis. In Parasitic Protozoa Vol III. Ed. Kreier
J. Academic Press. New York.
14. Fatihu, M.Y., Ogbogu, V.C., Njoku, C.V. & Sarror, D.I. 1991. Comparative
studies of gastrointestinal helminth of poultry in Zaria. Revue D’élevage et
de Médecine Vétérinaire Des Pays Tropicaux, 44(2):175–177.
15. Fecchio, A. 2011. Prevalência, diversidade e estructura da comunidade de
hemoparasitos (Haemoproteus e Plasmodium) em aves do Cerrado do
Brasil Central. Programa de Pós-graduação em Biología Animal.
Universidade de Brasília. http://repositorio.unb.br/handle/10482/8811
16. Forbes, M., Weatherhead, P.J. & Bennett, G.F. 1994. Blood parasites of
blue grouses: Variation in prevalence and patterns of inter specific
association. Oecologia, 97(4):520–525.

44
17. Gupta, D.K., Jahan, N. & Gupta, N. 2011. Distribution pattern of
apicomplexan parasites (Sporozoa: Haemosporida) in Columba livia,
Gmelin. Journal of Parasitic Diseases, 35(1):18-22.
18. Hamilton, W.D. 1967. Extraordinary sex ratios. Science, 156(3774):477–
488.
19. Harlin, R.W. 1994. Pigeons. The Veterinary Clinics of North America: Small
Animal Practice, 24(1):157–173.
20. Klei, T.R. & DeGiusti, D.L. 1975. Seasonal occurence of Haemoproteus
columbae Kruse and its vector Pseudolynchia canariensis Bequaert.
Journal of Widlife Diseases, 11(1):130-135.
21. Krone, O., Waldenström, J., Valkiūnas, G., Lessow, O., Müller, K., Iezhova,
T.A., Fickel, J. & Bensch, S. 2008. Haemosporidian blood parasites in
European birds of prey and owls. Journal Parasitol, 94(3):709-815.
22. Martínez-Moreno, FJ., Martínez-Moreno, A., Becerra-Martell, C. & Martínez-
Cruz, MS. 1989. Parasitofauna de la paloma (Columba livia) en la
Provincia de Córdoba (España). Revista Ibérica de Parasitología,
49(4):279-281.
23. Matta, N.E. & Rodriguez, O.A. 2001. Hemoparásitos aviares. Acta Biológica
Colombiana, 6(1):27-34.
24. Morrison, D.A. 2009. Evolution of the Apicomplexa: where are we now?
Trends Parasitol, 25(8):375-382.
25. Nematollahi, A., Ebrahimi, M., Ahmadi, A. & Himan, M. 2012. Prevalence of
Haemoproteus columbae and Trichomonas gallinae in pigeons (Columba

45
livia domestica) in Isfahan, Iran. Journal of Parasitic Diseases, 36(1):141-
142.
26. Organización Mundial de la Salud.1992.Métodos básicos de laboratorio en
parasitología médica. Parte 1. Ginebra, Pp 44
27. Quiroz, H. (2005). Parasitología y enfermedades parasitarias de animales
domésticos. México, D.F: Limusa, Grupo Noriega Editores.
28. Reza, M. & Taghi, M. 2011. Haemoproteus columbae in Columba livia
domestica of Three Areas in Iran in 2010. Global Veterinaria, 7(6):593-
595.
29. Santiago, D. & Carbó, P. 2015. Parásitos sanguíneos de malaria y géneros
relacionados (orden: haemosporida) en aves de México:
recomendaciones metodológicas para campo y laboratorio. Ornitología
Neotropical, 26(1):59–77.
30. Scaglione, F.S., Pregel, P., Cannizzo, F.T., Pérez-Rodríguez, A., Ferroglio,
E. & Bollo, E. 2015. Prevalence of new and known species of
haemoparasites in feral pigeons in northwest Italy. Malaria Journal, 14:99.
31. Sol, D., Jovani, R. & Torres, J. 2000. Geographical variation in blood
parasites in feral pigeons: the role of vectors. Ecography, 23(3):307-314.
32. Valkiūnas, G. 2004. Avian Malaria Parasites and other Haemosporidia.
Florida: CRC, Boca Ratón.
33. Van Riper, C., Atkinson, C. & Seed, T. 1994. Plasmodia of birds, in Parasitic
Protozoa, Vol 7, Kreier J.P. ed. New York and London. Academic Press.
34. Waite, J., Henry, A., Owen, J. & Clayton, D. 2014. An experimental test of
the effects of behavioral and immunological defenses against vectors: do

46
they interact to protect birds from blood parasites?. Parasites & Vectors,
7(1):104.
35. Wiehn, J. & Korpimaki, E. 1998. Resource levels reproduction and
resistance to haematozoan infections. The Royal Society of London,
265(1402):1197–1201.
36. Imagen Culicoides nubeculosus: Valkiūnas, G. 2004. Avian Malaria
Parasites and other Haemosporidia. Florida: CRC, Boca Ratón.
37. Imagen vena metatarsal: Clark, P., Boardman, W. & Raidal, S. 2009. Atlas
of clinical Avian Hematology. Singapore: Wiley-Blackwell.

47
9. ANEXO
9.1. Anexo 1
CUADERNO DE CAMPO
Localidad: Fecha: …/…/2014
NUMERO DE MUESTRA IDENTIFICACIÓN ESPECIE
9.2. Anexo 2
OBSERVACIÓN AL MICROSCOPIO
NUMERO DE MUESTRA PALOMAR POSITIVO/NEGATIVO

48
9.3. Anexo 3
RECUENTO DE LOS DATOS DE LAS FOTOGRAFÍAS POSITIVAS
NUMERO DE CAMPOS (1-50)
N º
M
U
E
S
T
R
A
ERITROCITOS
LEUCOCITOS
MACROGAMETOCITOS
MICROGAMETOCITOS
N º
M
U
E
S
T
R
A
ERITROCITOS
LEUCOCITOS
MACROGAMETOCITOS
MICROGAMETOCITOS