Embed Size (px)
Citation preview

NATURE NEUROSCIENCE VOLUME 10 | NUMBER 1 | JANUARY 2007 9
N E W S A N D V I E W S
Tolerance to opiate reward: role of midbrain IRS2-Akt pathwayBrandon K Harvey, Bruce T Hope & Yavin Shaham
Addicts report that opiate drugs lose their rewarding effects over time, but the molecular mechanisms underlying this effect are unknown. A study now reports that tolerance to morphine reward in rats is due to downregulation of IRS2-Akt signaling in the ventral tegmental area (VTA), the cell body region of the mesolimbic dopamine reward system.
In 1952, Abraham Wikler1 reported that an addicted subject given free access to morphine increased his daily drug intake from about 50 mg per day to up to 1,400 mg per day over 100 days. This dramatic observation, and results from subsequent studies, suggest that the progression of opiate addiction is accompanied by tolerance (a decrease in behavioral or physiological responses to a drug over repeated administration) to the drug’s rewarding effects2. Animal models of addiction also provide some evidence of tolerance to opiate reward. Rats allowed to self- administer drugs (animals are typically trained to perform an operant response, usually a lever-press for drug injections) and given unlimited access to heroin increase their drug intake over time3,4. In addition, rats previ-ously exposed to high morphine doses become less sensitive to the drug’s rewarding effects, as measured in the conditioned place preference procedure5,6. Yet, although numerous studies have been done on cellular events underlying different behavioral effects of opiate drugs7,8, the cellular mechanisms of opiate reward tolerance have not been explored. Now, Russo et al.9 use an impressive array of behavioral, neuroanatomical and molecular techniques to demonstrate that the IRS2-Akt signaling pathway in the VTA has a critical role in mediating this tolerance to opiate reward.
One day after withdrawal from chronic morphine exposure, the size of VTA dopamine (but not non-dopamine) neurons
decreases by ~25% (ref. 10). This effect is reversed by brain-derived neurotrophic factor (BDNF), which signals through three main pathways: the protein insulin receptor substrate 2 (IRS2), phospholipase Cγ and extracellular signal regulated kinase (ERK) pathways. However, the cellular mechanisms underlying this morphine-induced reduction in VTA dopamine cell size, and the behavioral significance of this morphological change, were not clear.
To determine which cellular mechanism is involved, Russo et al.9 explored the role of the
IRS2-Akt pathway. When IRS2 is phosphorylated by receptor tyrosine kinases (RTKs), this leads to activation of protein kinase B or Akt, a kinase known to regulate several cellular processes such as apoptosis, cell survival, axonal growth and cell differentiation. Russo et al. were guided by their and others’ previous findings that chronic morphine exposure decreases VTA IRS2 protein levels and Akt activity but increases phospholipase Cγ and ERK activity. They first confirmed that VTA cell size and IRS2 protein levels decreased 1 day after withdrawal from chronic exposure to morphine pellets (75 mg per day for 5 d).
The authors are at the Intramural Research
Program, National Institute on Drug Abuse,
National Institutes of Health, Department of
Health and Human Services, 5500 Nathan
Shock Drive, Baltimore, Maryland 21224, USA.
e-mail: [email protected]
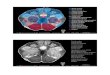
IntegrinsRTKs
IRS2
PIP2
PIP2PIP3
PI3K Akt
pAkt
PDK1 PTEN
D
B
C
A
GPCRs
Multiple targets:kinases (e.g., GSK3)
transcription factors (e.g., NK-ΚB)
Cell death or survival,cell growth, cell differentiation
Figure 1 Manipulation of the IRS2-Akt pathway in the VTA using HSV amplicon vector-mediated gene delivery. Ligand binding to the extracellular side of receptor tyrosine kinases (RTK) activates tyrosine kinases within the intracellular portions of these receptors. Activated RTKs autophosphorylate themselves as well as phosphorylating tyrosine residues on other membrane-bound substrates such as insulin receptor substrate 2 (IRS2). Phosphorylated IRS2 recruits phosphotidylinositol-3 kinase (PI3K) to the membrane, where it phosphorylates the intracellular membrane lipid phosphatidyl inositol (4,5)-bisphosphate (PIP2) to produce phosphatidyl inositol (3,4,5)-trisphosphate (PIP3). The presence of PIP3 in the membrane attracts inactive Akt (also known as protein kinase B) to the membrane, where membrane-bound phosphoinositide-dependent kinase-1 (PDK1) can phosphorylate and activate Akt (pAkt). Membrane-bound phosphatase and tensin homolog deleted in chromosome 10 (PTEN) dephosphorylates PIP3 to PIP2 and inactivates the lipid signaling pathway. Activated Akt can dissociate from the membrane and phosphorylate serine and threonine residues on at least 20 known substrates that control diverse cellular pathways. Other receptors, such as integrins and certain G protein–coupled receptors (GPCRs), can also activate PI3K, resulting in PIP3 production and Akt activation (dashed lines). Russo et al.9 used HSV vectors to disrupt (A, C; blue) or augment (B, D; blue) IRS2-Akt signaling in cells of the VTA. GSK3, glycogen synthase kinase-3β.
Kim
berly
Cae
sar
©20
07 N
atur
e P
ublis
hing
Gro
up
http
://w
ww
.nat
ure.
com
/nat
uren
euro
scie
nce

10 VOLUME 10 | NUMBER 1 | JANUARY 2007 NATURE NEUROSCIENCE
N E W S A N D V I E W S
These effects were accompanied by decreases in phosphorylation of IRS2 downstream targets, Akt and glycogen synthase kinase-3β (Fig. 1).
To demonstrate that downregulation of the IRS2-Akt pathway mediates morphine-induced decreases in the cell size of VTA neurons, Russo et al. used in vivo viral- mediated gene transfer11,12, a method where a functional gene of interest is incorporated into the genome of a virus that has been made non-replicating but retains the capacity to deliver its genome into cells. They used this method to express dominant- negative and wild-type IRS2 proteins (IRS2dn and IRS2wt) using a herpes simplex virus (HSV) vector. They injected these virus constructs into the VTA of drug-naive or morphine-treated rats and found that an IRS2dn virus, which decreased IRS2 function, reduced VTA cell size in morphine-naive rats, whereas the IRS2wt virus, which presumably restored morphine-downregulated IRS2 function, prevented morphine-induced decreases in cell size. These findings indicate that chronic morphine exposure decreases VTA cell size through an IRS2-dependent mechanism.
Russo et al. then assessed the roles of VTA cell size reduction and downregulation of the IRS2 pathway in morphine reward by using the conditioned place preference (CPP) procedure. During the conditioning (training) phase of this procedure, a distinct environment is paired with drug injections and effects, while another environment is paired with vehicle (control) injections; during the subsequent drug-free test phase, animals are allowed to choose between the two environments. Mice and rats show increased preference for the previously drug-paired environment over the non-drug environment, and this increase in preference is used to measure the drug’s rewarding effects. The authors found that the morphine-dependent rats were less sensitive to morphine reward during early withdrawal (1 d); during this time, they observed rewarding effects of morphine in these rats only at doses 20 times higher than those that would be rewarding in non-drug-dependent rats. Russo et al. also found that sensitivity to morphine reward was associated with changes in VTA cell size: when these cells returned to their normal size after 4 weeks of withdrawal, the previously morphine-dependent rats were no longer tolerant to morphine reward. Together with the previous molecular and anatomical findings, these results demonstrate that tolerance to morphine’s rewarding effects in drug- dependent rats is associated with reduced VTA cell size and downregulation of the IRS2-Akt pathway.
The authors then found that IRS2dn VTA injections, like chronic morphine administration, decreased morphine reward, whereas IRS2wt injections modestly enhanced
drug reward. These effects are likely to be mediated by the IRS2-Akt pathway: VTA injections of an Aktdn virus mimicked the effect of the IRS2dn virus. This newly identified molecular mechanism for opiate reward tolerance suggests that morphine downregulates the IRS2-Akt pathway in VTA cells, resulting in decreases in VTA dopamine cell size, which lead to tolerance to morphine reward.
However, the generality of these findings with regard to the self-administration of tolerance-inducing high doses of opiate drugs remains unclear. The CPP procedure that the authors used to assess the rewarding effect of morphine has some limitations: drug exposure is not under the rat’s control and the amount of drug exposure is generally very low (typically only two to four acute intraperitoneal or subcutaneous injections of low to moderate doses of drug). Thus, the relevance of CPP results to voluntary, compulsive intake of high doses of opiate drugs in humans1,2 is unknown. Russo et al. reported that, as with morphine pellets, 26 days of 4 h per day of heroin self- administration decreased VTA cell size, suggesting that the results from the CPP procedure generalize to opiate self- administration. However, because they did not report the rats’ daily heroin intake, it is unknown whether the reduction in VTA dopamine cell size was associated with tolerance to self-administered heroin. In a previous study in which rats were trained for 4 h per day to lever-press for heroin injections, Martin et al.13 found that daily drug intake remained unchanged for many weeks (that is, no drug tolerance developed). Based on the results of Martin et al., an important question for future research is whether opiate-induced decreases in VTA cell size also mediate tolerance to the rewarding effects of opiate drugs in the self-administration procedure in rats given unlimited drug access3,4.
The interpretation of data from addiction studies using viral-mediated gene transfer procedures requires some consideration. Over the last decade, this method has been used to study the role of drug-induced neuroadaptations in locomotor sensitization, reward and relapse11,14. Typically, in these studies, investigators target proteins that are upregulated in VTA and its projection areas by chronic exposure to addictive drugs. They inject protein-specific wild-type or dominant-negative viruses into the brain and assess the drugs’ behavioral effects. If injections of the wild-type virus enhance these behavioral effects, investigators conclude that the specific neuroadaptations promote drug-taking behavior. In contrast, if injections of the dominant- negative virus enhance drugs’ behavioral effects, investigators conclude that the specific neuroadaptations decrease
drug- taking behavior. However, a potential problem with this common interpretation is that investigators usually do not provide data comparing the expression levels of the target protein in rats repeatedly exposed to drugs with those of drug-naive rats injected with the viral constructs. In most cases, it is likely that the virus manipulation increases (or decreases) protein levels to a much greater degree than that induced by repeated drug exposure (typically about ~50% or less of normal levels)15. Thus, the relevance of findings from viral-mediated gene transfer studies, in which proteins levels increase (or decrease) to non-physiological values, to the understanding of the role of drug-induced neuroadaptations in drug reward and relapse is questionable.
Taking these caveats into consideration, a particularly strong feature of Russo et al.’s report is that the viral- mediated gene delivery of IRS2dn into the VTA caused both cellular and behavioral responses that were very similar to those induced by chronic morphine. This finding indicates that the viral-mediated gene transfer used here caused physiologically relevant changes, and thus allows the authors to conclude that the VTA IRS2-Akt pathway mediates tolerance to morphine reward in drug-dependent rats. This experimental procedure and findings should serve as a model for future studies in which the in vivo viral- mediated gene transfer method is used to assess the role of drug-induced neuroadaptations in addiction. Understanding the mechanisms behind drug tolerance may also allow us to develop effective therapeutic targets for reversing or delaying the development of opiate addiction.
1. Wikler, A. Psychiatr. Q. 26, 270–293 (1952).2. Jaffe, J.H. Drug addiction and drug abuse. in
Goodman & Gilman’s the Pharmacological Basis of Therapeutics (ed. Gilman, A.G., Rall, T.W., Nies, A.S. & Taylor, P.) 522–573 (Pergamon Press, New York, 1990).
3. Bozarth, M.A. & Wise, R.A. J. Am. Med. Assoc. 254, 81–83 (1985).
4. Ahmed, S.H., Walker, J.R. & Koob, G.F. Neuropsychopharmacology 22, 413–421 (2000).
5. Shippenberg, T.S., Emmett-Oglesby, M.W., Ayesta, F.J. & Herz, A. Psychopharmacology (Berl.) 96, 110–115 (1988).
6. Bechara, A. & van der Kooy, D. Behav. Neurosci. 106, 364–373 (1992).
7. Nestler, E.J. & Aghajanian, G.K. Science 278, 58–63 (1997).
8. De Vries, T.J. & Shippenberg, T.S. J. Neurosci. 22, 3321–3325 (2002).
9. Russo, S.J. et al. Nat. Neurosci. 10, 93–99 (2007).10. Sklair-Tavron, L. et al. Proc. Natl. Acad. Sci. USA 93,
11202–11207 (1996).11. Neve, R.L., Neve, K.A., Nestler, E.J. & Carlezon, W.A.,
Jr. Biotechniques 39, 381–391 (2005).12. Simonato, M., Manservigi, R., Marconi, P. & Glorioso, J.
Trends Neurosci. 23, 183–190 (2000).13. Martin, T.J., Walker, L.E., Sizemore, G.M., Smith, J.E.
& Dworkin, S.I. Drug Alcohol Depend. 41, 93–100 (1996).
14. Self, D.W. Biol. Psychiatry 57, 1479–1484 (2005).15. Lu, L., Grimm, J.W., Shaham, Y. & Hope, B.T.
J. Neurochem. 85, 1604–1613 (2003).
©20
07 N
atur
e P
ublis
hing
Gro
up
http
://w
ww
.nat
ure.
com
/nat
uren
euro
scie
nce