Embed Size (px)
Citation preview

Function of Jasmonate in Response and Tolerance of Arabidopsisto Thrip Feeding
Hiroshi Abe1,*, Jun Ohnishi
2, Mari Narusaka
3, Shigemi Seo
4, Yoshihiro Narusaka
3, Shinya Tsuda
2and
Masatomo Kobayashi1
1 Department of Biological Systems, RIKEN BioResource Center, Tsukuba, 305-0074 Japan2 Department of Plant Pathology, National Agricultural Research Center, Tsukuba, 305-8666 Japan3 Research Institute for Biological Sciences, Okayama, 7549-1 Yoshikawa, Kibichuo-cho, Okayama 716-1241, Japan4 Department of Plant Physiology, National Institute of Agrobiological Sciences, Tsukuba, 305-8666 Japan
We analyzed the interaction between Arabidopsis and
western flower thrips (Frankliniella occidentalis), which are one
of the most serious insect pests of cultivated plants. We focused
on the function of the immunity-related plant hormones
jasmonate (JA), ethylene (ET) and salicylic acid (SA) in the
plant’s response to thrip feeding. Expression of the marker
genes for each hormone response was induced by thrip feeding in
wild-type (WT) plants. Further analyses in the hormone-related
mutants coi1-1 (JA insensitive), ein2-1 and ein3-1 (ET
insensitive) and eds16-1 (SA deficient) suggested the impor-
tance of these hormones in the plant response to feeding.
Comparative transcriptome analyses suggested a strong
relationship between thrip feeding and JA treatment, but not
ET or SA treatment. The JA content of WT plants was
significantly increased after thrip feeding. Moreover, coi1-1,
but not ein2-1, showed lower feeding tolerance against thrips
than the WT. Application of JA to WT plants before thrip
feeding enhanced the plants’ feeding tolerance. JA modulates
several defense responses in cooperation with ET, but applica-
tion of the ET precursor 1-aminocyclopropane-carboxylic acid
had a marked negative effect on feeding tolerance. Our results
indicate that JA plays an important role inArabidopsis in terms
of response to, and tolerance against, thrip feeding.
Keywords: Arabidopsis thaliana — Ethylene — Frankliniella
occidentalis — Insect feeding — Jasmonate — Western
flower thrips.
Abbreviations: ACC, 1-aminocyclopropane-carboxylic acid;AOC, allene oxide cyclase; AOS, allene oxide synthase; chiB,�-chitinase; coi1-1, coronatine insensitive 1-1; eds16-1, enhanceddisease susceptibility 16-1; ein2-1, ein3-1, ethylene-insensitive 2-1,3-1; ET, ethylene; JA, jasmonic acid; LOX2, lipoxygenase 2;OPDA, 12-oxo-phytodienoic acid; PDF1.2, plant defensin 1.2; SA,salicylic acid; VSP2, vegetative storage protein 2; WT, wild type.
Introduction
Insect feeding retards plant growth and decreases crop
productivity. Plants develop defense systems against insect
feeding by constitutive and inducible protective responses
(reviewed by Walling 2000, Kessler and Baldwin 2002,
Korth and Thompson 2006), many of which remain
unclear. Recently, plant responses to insect feeding have
been analyzed at the molecular, cellular and physiological
levels (reviewed by Kessler and Baldwin 2002, Korth and
Thompson 2006, Turlings and Ton 2006), although these
analyses of plant–herbivore interaction have focused on
limited types of insect species (mostly lepidopteran larvae
and aphids).
The western flower thrip (Frankliniella occidentalis
[Pergande]) is one of the most important insect herbivores
and is an exotic pest of greenhouse production in many
countries. Thrips use their mouthparts to pierce plant cells
and suck out the contents. This feeding mode differs from
those of the well-studied caterpillars or aphids (reviewed by
Walling 2000, Thompson and Goggin 2006), and thrips can
also act as virus vectors (reviewed by Parrella 1995).
Because the thigmokinetic behavior of western flower
thrips (i.e. their tendency to occupy narrow crevices
within or between plant parts) and the emergence of
insecticidal resistance, it is difficult to control this species
with insecticides (reviewed by Jensen 2000). Therefore,
elucidation of the molecular mechanism of the plant
response to the feeding of western flower thrips is important
for the development of new methods to prevent damage.
The plant hormones jasmonic acid (JA), ethylene (ET)
and salicylic acid (SA) mediate a variety of physiological
processes, including the basal plant defense responses
induced by various biotic stresses such as fungal, bacterial
and viral infections (reviewed by Pieterse et al. 2006). The
hormonal defense responses in plants are classified into two
major pathways: the SA pathway and the JA/ET pathway
(reviewed by Thomma et al. 2001). The SA-mediated
defense response provides resistance to biotrophic fungi
and bacteria such as Peranospora parasitica and
Pseudomonas syringae (Uknes et al. 1992). Activation of
the SA-mediated defense response is related to the
hypersensitive response, which triggers systemic acquired
resistance. In contrast, the JA/ET-mediated pathway
*Corresponding author: E-mail, [email protected]; Fax, þ81-29-836-9053.
Plant Cell Physiol. 49(1): 68–80 (2008)doi:10.1093/pcp/pcm168, available online at www.pcp.oxfordjournals.org� The Author 2007. Published by Oxford University Press on behalf of Japanese Society of Plant Physiologists.All rights reserved. For permissions, please email: [email protected]
68
Dow
nloaded from https://academ
ic.oup.com/pcp/article/49/1/68/1903854 by guest on 09 D
ecember 2021

appears to regulate resistance against necrotrophic fungi
such as Alternaria brassicicola and Botrytis cinerea
(Thomma et al. 1998).
In addition, there are at least two modes of interaction
between JA and ET, one synergistic and the other
antagonistic (reviewed by Pieterse et al. 2006). Expression
of �-chitinase (chiB) and plant defensin (PDF1.2), marker
genes of the JA/ET pathway, is induced by application of
either JA or ET, and a synergistic effect occurs when both
hormones are applied. However, expression of vegetative
storage protein 2 (VSP2) and lipoxygenase 2 (LOX2),
marker genes of the JA pathway, is induced by JA but
not by ET. The effect of JA on the expression of these genes
is antagonistically reduced by ET treatment (reviewed by
Lorenzo and Solano 2005). Campbell et al. (2002) reported
that expression of some defense-related genes might be
regulated through the coordination of multiple signaling
cascades. Thus, it is important to examine the involvement
of each hormone in each biotic stress response.
The plant hormones JA, ET and SA also appear to
have important functions in plant responses to insect
feeding (reviewed by Walling 2000, Korth and Thompson
2006). In Arabidopsis, Ellis et al. (2002) reported that the
constitutive JA signaling mutant cev1 shows higher
tolerance to aphids, whereas tolerance is decreased in the
JA-insensitive mutant coi1-1. The tomato spr2 mutant, a
mutant that shows loss of function of fatty acid desaturase
and is involved in JA biosynthesis, is compromised in
defense against attack by tobacco hornworm larvae
(Li et al. 2003). In Nicotiana attenuata, JA regulates the
accumulation of the potent defense metabolite nicotine, and
ET suppresses the JA-induced accumulation of nicotine.
Attack of the leaves of N. attenuata by its specialist
herbivore Manduca sexta elicits ET synthesis, and as a
result the nicotine content in N. attenuata is decreased or
unchanged (Kahl et al. 2000, Winz and Baldwin 2001). This
well-understood scheme demonstrates the relationship
between plant–insect interactions and the functions of
plant hormones.
Studies have also clarified the function of SA in plant
tolerance against insect attack. Application of the SA
analog benzothiadiazole to tomato reduces the population
growth of aphids (Cooper et al. 2004). In addition, cpr5 and
ssi2 mutants, which have an elevated SA level and a
constitutively active SA-dependent signal, respectively,
showed increased tolerance to aphid population growth
(Pegadaraju et al. 2005). In contrast, the reduced SA
accumulation mutant pad4 showed decreased tolerance to
aphid population growth (Pegadaraju et al. 2005). The
expression of various genes is induced by the feeding of
insects, such as aphids and caterpillars (Moran and
Thompson 2001, Moran et al. 2002, Hui et al. 2003,
Thompson and Goggin 2006); the expression of many of
these feeding-inducible genes is also induced by JA/ET or
SA treatment (Moran and Thompson 2001). However, the
molecular mechanism underlying the function of these plant
hormones in feeding response remains unclear.
In this study, we analyzed the function of JA, ET
and SA in the response of Arabidopsis to thrip feeding.
Expression analyses of marker genes suggested the involve-
ment of these hormones in the response to thrip feeding. The
expression of marker genes was reduced or canceled in the
hormonal mutants. In addition, we performed comparative
transcriptome analyses in Arabidopsis treated with thrips,
JA, ET or SA. JA contents were increased after thrip feeding,
and the results demonstrated the greater importance of JA
than ET or SA in the Arabidopsis response to thrip feeding.
In addition, the coi1-1 (JA-insensitive) mutant showed
decreased feeding tolerance against thrips; application of
JA enhanced its feeding tolerance against thrips, whereas ET
application had no such effect. We discuss the function of JA
and ET in the feeding tolerance of Arabidopsis.
Results
Expression of marker genes involved in hormone responses
We analyzed the JA-, ET- or SA-inducible expression
of several marker genes in Arabidopsis. Expression of VSP2
and LOX2, marker genes of the JA pathway, was induced
by thrip feedings. This induction was not observed in
control plants (Fig. 1A). Similarly, the expression of chiB
and PDF1.2, marker genes of the JA/ET pathway, and of
the SA-inducible genes PR1 and BGL2 was also induced by
thrip feeding (Fig. 1B, C). Because these results indicated
that JA, ET and SA production was affected by thrip
feeding, we used hormonal mutants (i.e. the JA-insensitive
mutant coi1-1, the ET-insensitive mutants ein2-1 and ein3-1,
and the SA-deficient mutant eds16-1) to analyze the
induction of expression of marker genes by thrip feeding
(Chao et al. 1997, Xie et al. 1998, Alonso et al. 1999,
Dewdney et al. 2000, Wildermuth et al. 2001). Expression of
VSP2 and chiB, marker genes of the JA and JA/ET
pathways, respectively, was induced less in the
JA-insensitive coi1-1 mutant than in the control plant
(Fig. 2A). Expression of chiB was also reduced in the
ET-insensitive ein2-1 and ein3-1 mutants (Fig. 2B), but
induction of expression of the VSP2 gene was not reduced
in these mutants (Fig. 2B). Expression of PR1 and BGL2 in
the SA-deficient mutant eds16-1was markedly reduced
(Fig. 2C). These results suggest the possible involvement
of JA, ET and SA signals in the feeding-inducible gene
expression of Arabidopsis by thrips.
Correlation between thrip feeding and hormone treatments
Microarrays are convenient tools that are used to
investigate the correlation in gene expression between insect
JA but not ET acts in Arabidopsis tolerance to thrips 69
Dow
nloaded from https://academ
ic.oup.com/pcp/article/49/1/68/1903854 by guest on 09 D
ecember 2021

5VSP2 geneA
4R
elat
ive
expr
essi
on
3
2
1
0
0h
5h
10h
24h
5h
10h
24h
−Feeding +Feeding
5LOX2 gene
4
Rel
ativ
e ex
pres
sion
3
2
1
0
0h
5h
10h
24h
5h
10h
24h
−Feeding +Feeding
5PDF1.2 geneB
4
Rel
ativ
e ex
pres
sion
3
2
1
0
0h
5h
10h
24h
5h
10h
24h
−Feeding +Feeding
5CHIB gene
4R
elat
ive
expr
essi
on
3
2
1
0
0h
5h
10h
24h
5h
10h
24h
−Feeding +Feeding
5BGL.2 geneC
4
Rel
ativ
e ex
pres
sion
3
2
1
0
0h
5h
10h
24h
5h
10h
24h
−Feeding +Feeding
5PR1 gene
4
Rel
ativ
e ex
pres
sion
3
2
1
0
0h
5h
10h
24h
5h
10h
24h
−Feeding +Feeding
Fig. 1 Expression of marker genes for JA, ET and SA signals is induced by thrip feeding in Arabidopsis. (A) VSP2 and LOX2 as markergenes for the JA pathway, (B) chiB and PDF1.2 as marker genes for the JA/ET pathway and (C) PR1 and BGL2 as marker genes for the SApathway. Five 3-week-old plants were grown in a single pot, and 25 adult female thrips were allowed to feed on plants. After 0, 5, 10 and24 h, total RNA was prepared from the plants with (þFeeding) or without (�Feeding) thrip feeding, and first-strand cDNA was synthesizedfor PCR analysis. Primer sequences used in this analysis are described in Materials and Methods. The expression level of each gene wasnormalized to the expression of CBP20 (control) and is shown as the relative value. Each value represents the average of 10 plants �SD ofthree replications.
70 JA but not ET acts in Arabidopsis tolerance to thrips
Dow
nloaded from https://academ
ic.oup.com/pcp/article/49/1/68/1903854 by guest on 09 D
ecember 2021

A5
VSP2 gene
4
3
Rel
ativ
e ex
pres
sion
2
1
0
WT
0h
5h
Fee
ding
24h
Fee
ding 0h
5h
Fee
ding
24h
Fee
ding
coi1-1
5CHIB gene
4
3
Rel
ativ
e ex
pres
sion
2
1
0
WT
0h
5h
Fee
ding
24h
Fee
ding 0h
5h
Fee
ding
24h
Fee
ding
coi1-1
C5PR1 gene
4
3
Rel
ativ
e ex
pres
sion
2
1
0
WT
0h
5h
Fee
ding
24h
Fee
ding 0h
5h
Fee
ding
24h
Fee
ding
eds16-1
5BGL2 gene
4
3
Rel
ativ
e ex
pres
sion
2
1
0
WT
0h
5h
Fee
ding
24h
Fee
ding 0h
5h
Fee
ding
24h
Fee
ding
eds16-1
B5
CHIB gene
4
3
Rel
ativ
e ex
pres
sion
2
1
0
WT
0h
5h
Fee
ding
5h
Fee
ding
24h
Fee
ding
24h
Fee
ding0hr
0hr
5h
Fee
ding
24h
Fee
ding
ein2-1 ein3-1 WT
0h
5h
Fee
ding
5h
Fee
ding
24h
Fee
ding
24h
Fee
ding0hr
0hr
5h
Fee
ding
24h
Fee
ding
ein2-1 ein3-1
5VSP2 gene
4
3
Rel
ativ
e ex
pres
sion
2
1
0
Fig. 2 Expression of marker genes for JA, ET and SA signals induced by thrip feeding is suppressed in coi1-1, ein2-1, ein3-1 and eds16-1mutants. (A) VSP2 and LOX2 in Columbia (WT) and the JA-insensitive mutant coi1-1, (B) chiB and PDF1.2 in WT and the ET-insensitivemutants ein2-1 and ein3-1, and (C) PR1 and BGL2 in WT and the SA-deficient mutant eds16-1. Analyses were the same as described in thelegend to Fig. 1.
JA but not ET acts in Arabidopsis tolerance to thrips 71
Dow
nloaded from https://academ
ic.oup.com/pcp/article/49/1/68/1903854 by guest on 09 D
ecember 2021

and hormone treatments. Narusaka et al. (2006) designed
an Arabidopsis microarray consisting of 1.2K full-length
cDNA clones representing the putative defense-related and
regulatory genes. These 1.2K genes were selected from
among the genes covered by the Arabidopsis 7K array
(RIKEN, Japan; Seki et al. 2002) and Arabidopsis oligo
microarray (Agilent Technologies, Santa Clara, CA, USA;
Narusaka et al. 2003a, Narusaka et al. 2003b, Narusaka
et al. 2004). Treatment of Arabidopsis Col-0 WT plants with
SA, JA and ET induced the expression of 282, 159 and 246
genes, respectively (Narusaka et al. 2006). We used the
1.2K cDNA microarray to compare the gene expression
patterns between thrip feeding and JA, ET or SA treatment.
Thrips fed on Arabidopsis plants for 2, 5, 10 or 24 h, and
plants were snap-frozen in liquid nitrogen and stored at –
808C until RNA extraction. The relationship between
experimental pairs was analyzed by means of Pearson’s
correlation coefficient with the software Excel 2004 for Mac
ver. 11.2.3 (Microsoft, Redmond, WA, USA). Several
authors have offered guidelines for the interpretation of
the correlation coefficient. Cohen (1998), for example, has
suggested the following interpretations for correlations:
0� r50.10 is regarded as an insubstantial; 0.10� r50.30 as
a small; 0.30� r50.50 as a medium; and 0.50� r51.00 as a
large correlation. The analyses revealed significant correla-
tions between thrip feeding and JA treatment but not ET or
SA treatment (Table 1A). The correlation coefficient was
large or medium between thrip feeding and JA treatment,
except thrip feeding for 2 h and JA treatment for 10 h. These
results indicate the importance of JA in the response to
thrip feeding. The correlation between thrip feeding and ET
or SA treatment was insubstantial. Most of these pairs,
except thrip feeding for 2 h and ET treatment for 2 h,
showed a correlation50.2 suggesting that they are not close
or show a negative correlation. We also detected the
correlation between JA and ET (Table 1B). The relationship
between JA and ET treatment, based on transcriptome
analyses, was reported previously (Schenk et al. 2000).
Therefore, we further analyzed the function of JA in the
response of Arabidopsis to thrip feeding to investigate
whether ET was also involved in the response.
Jasmonate functions as a regulator of plant response to thrip
feeding
To analyze the function of JA in the response to thrip
feeding, we measured the contents of JA in plants fed on by
thrips. The JA content of plants 24 h after thrip feeding was
11.8 times that before feeding; in plants that had not been
fed on, the JA level after 24 h was 1.4 times that before
feeding, i.e. virtually unchanged (Fig. 3A). Next, we
analyzed the expression of the genes allene oxide synthase
(AOS) and allene oxide cyclase 2 (AOC2), both of which
encode enzymes that catalyze JA biosynthesis (Delker et al.
2006; Fig. 3C), in plants with or without thrip feeding.
Induction of the expression of both genes was apparent 24
and 48 h after thrip feeding (Fig. 3D, E). However,
biosyntheses of ET was not increased in the plants 24 h
after thrip feeding (Fig. 3B). We detected elevated ET levels
in the eto1-1 mutant (an overproducer of ET) under control
conditions, suggesting that our experimental design was
valid (Fig. 3B).
Next, we used the JA-insensitive mutant coi1-1 to
analyze the function of JA in the tolerance of Arabidopsis to
thrip feeding. Arabidopsis rosette leaves were cut into discs
8mm in diameter. One adult female thrip was put on the
leaf disc and allowed to feed. After 1 or 2 d, we determine
the area of the feeding scar on each leaf disc (n¼ 10;
Fig. 4A, B). After 2 d of feeding, the area of feeding scars
was significantly larger than that after 1 d of feeding in both
WT and mutant plants, suggesting the validity of this
Table 1 Correlation coefficients between experimental pairs in the microarray analyses
JA 2 h JA 5h JA 10h JA 24 h ET 2h ET 5h ET 10 h ET 24 h SA 2h SA 5h SA10h SA 24 h
A Thrips 2 h 0.31� 0.44� 0.25� 0.31� 0.22� –0.04 0.02 0.06 0.07 0.02 0.09 0.24�
Thrips 5 h 0.38� 0.56� 0.51� 0.33� 0.01 –0.10 –0.07 0.01 –0.06 –0.06 0.00 0.15�
Thrips 10 h 0.30� 0.55� 0.55� 0.34� –0.07 –0.16� –0.11� –0.04 –0.09 –0.13� –0.09 0.07
Thrips 24 h 0.37� 0.60� 0.65� 0.42� –0.05 –0.11 –0.06 0.03 –0.10 –0.14� –0.09 0.10
B JA 2h – – – – 0.58� 0.30� 0.30� 0.50� 0.12� –0.04 –0.06 0.06
JA 5h – – – – 0.52� 0.41� 0.38� 0.54� 0.07 –0.03 0.06 0.13�
JA 10 h – – – – 0.44� 0.40� 0.44� 0.56� –0.03 –0.05 0.01 0.12�
JA 24 h – – – – 0.61� 0.43� 0.49� 0.76� –0.02 –0.17� –0.13� 0.18�
Total RNA was isolated 2, 5, 10, and 24 h after treatment with 5mM SA, 0.1mM JA or 1mM ET (Narusaka et al. 2006), or thrips feeding.Each experiment was replicated three times with 10–15 plants per sample. To obtain sufficient material, samples from replicateexperiments were pooled before RNA extraction. We conducted two or three independent microarray analyses with the same RNA.Asterisks indicate significant difference at P50.05.A complete data set is available at http://www.ncbi.nlm.nih.gov/geo/ with accession numbers GSM239486, GSM241219, GSM241220,GSM241221 and GSM86106.
72 JA but not ET acts in Arabidopsis tolerance to thrips
Dow
nloaded from https://academ
ic.oup.com/pcp/article/49/1/68/1903854 by guest on 09 D
ecember 2021

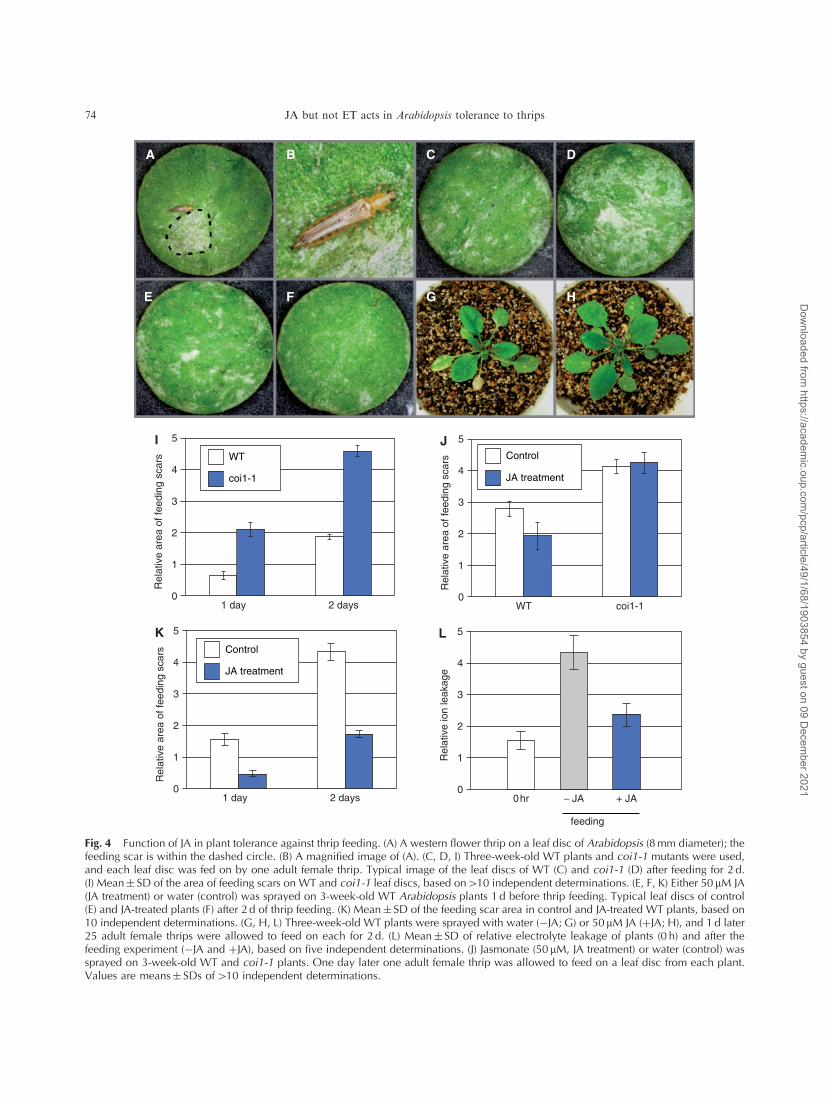
experimental system (Fig. 4I–K). The area of feeding scars
in coi1-1 mutants was significantly greater than that in WT
plants (Fig. 4C, D, I). To analyze the direct effect of JA on
thrip performance, we treated the coi1-1 mutant with 50 mMmethyl jasmonate 24 h before the beginning of feeding and
then determined the area of the feeding scar after 2 d of
thrip feeding. Application of JA to coi1-1 mutants did not
have a significant effect on thrip feeding (Fig. 4J). WT
plants were treated in the same manner; the feeding scar on
the JA-treated plants was significantly smaller than that on
untreated plants (Fig. 4E, F, K).
We next used whole Arabidopsis plants to analyze the
effect of JA treatment on feeding tolerance. JA treatment
enhanced feeding tolerance compared with that in untreated
plants (Fig. 4G, H). The effectiveness of JA application on
thrip feeding was evaluated by the electrolyte leakage test.
Electrolyte leakage was significantly lower in plants treated
with JA than in untreated plants (Fig. 4L). Together with
the results from the leaf disc assay, these results indicate
that JA is able to induce acquired resistance against thrip
feeding.
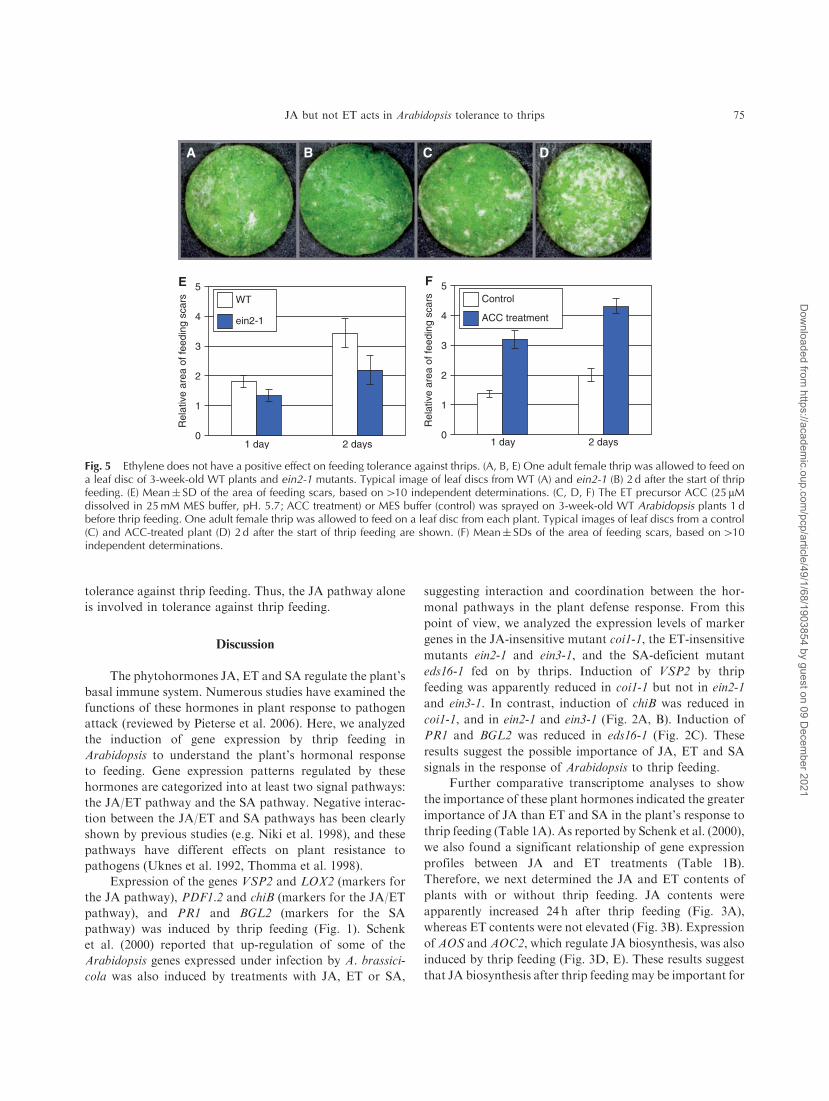
Ethylene does not enhance tolerance against thrip feeding
JA and ET have a synergistic role in some disease
responses via the JA/ET pathway (reviewed by Lorenzo and
Solano 2005). To understand the role of ET in tolerance
against thrip feeding, we analyzed the feeding rate in ein2-1
mutants. The ein2-1 mutants suffered a level of damage by
thrip feeding similar to that in WT plants (Fig. 5A, B, E).
Next, we analyzed the effect of ET application on thrip
feeding. Application of the ET precursor 1-aminocyclopro-
pane-carboxylic acid (ACC) 24 h before thrip feeding
increased the area of feeding scars compared with that in
untreated plants (Fig. 5C, D, F). These results indicate that
ET does not have any positive effect on Arabidopsis
400A
D
300
JA c
onte
nts
(ng/
gFW
)
200
100
0
5AOS gene
4
3
Rel
ativ
e ex
pres
sion
2
1
0
0h
0h 24h−Feeding
24h+Feeding
5B C
4
Rel
ativ
e co
nten
ts
2
3
1
024h
−Feeding
WT
24h−Feeding
24h+Feeding
eto1-1
Linoleic Acid (LA)
12-oxo-phytodieonic acid (OPDA)
LOX
AOS
AOC
OPR3
JA
24h
−Feeding
48h
48h
24h
E5
AOC2 gene
4
3
Rel
ativ
e ex
pres
sion
2
1
0
0h
24h
48h
48h
24h
+Feeding −Feeding +Feeding
Fig. 3 Effect of thrip feeding on the biosynthesis of JA and ET in Arabidopsis. (A, B) Endogenous levels of JA (JAþmethyl JA; A) and ET (B).Ten adult female thrips were allowed to feed on a 3-week-old plant in a closed container with air vents (þFeeding). The control plant waskept in a container without thrips (�Feeding). At the beginning of the experiment (0 h) and after 24 h from the start of feeding, 1 g of planttissue was harvested for the extraction of RNA. The results shown are means� SDs of three independent measurements. (C) Proposedmodel of the pathway of biosynthesis of JA in Arabidopsis. (D, E) Expression levels of the marker genes of JA biosynthesis, AOS (D) andAOC2 (E). Analyses were the same as described in the legend to Fig. 1.
JA but not ET acts in Arabidopsis tolerance to thrips 73
Dow
nloaded from https://academ
ic.oup.com/pcp/article/49/1/68/1903854 by guest on 09 D
ecember 2021
WT
coi1-1
5I
4
3
Rel
ativ
e ar
ea o
f fee
ding
sca
rs
2
1
01 day 2 days
Control
JA treatment
5J
4
3
Rel
ativ
e ar
ea o
f fee
ding
sca
rs
2
1
0WT coi1-1
Control
JA treatment
5K
4
3
Rel
ativ
e ar
ea o
f fee
ding
sca
rs
2
1
01 day 2 days
5L
4
3
Rel
ativ
e io
n le
akag
e
2
1
00hr
feeding
+ JA− JA
A B C D
E F G H
Fig. 4 Function of JA in plant tolerance against thrip feeding. (A) A western flower thrip on a leaf disc of Arabidopsis (8mm diameter); thefeeding scar is within the dashed circle. (B) A magnified image of (A). (C, D, I) Three-week-old WT plants and coi1-1 mutants were used,and each leaf disc was fed on by one adult female thrip. Typical image of the leaf discs of WT (C) and coi1-1 (D) after feeding for 2 d.(I) Mean� SD of the area of feeding scars on WT and coi1-1 leaf discs, based on410 independent determinations. (E, F, K) Either 50 mM JA(JA treatment) or water (control) was sprayed on 3-week-old WT Arabidopsis plants 1 d before thrip feeding. Typical leaf discs of control(E) and JA-treated plants (F) after 2 d of thrip feeding. (K) Mean� SD of the feeding scar area in control and JA-treated WT plants, based on10 independent determinations. (G, H, L) Three-week-old WT plants were sprayed with water (�JA; G) or 50 mM JA (þJA; H), and 1 d later25 adult female thrips were allowed to feed on each for 2 d. (L) Mean� SD of relative electrolyte leakage of plants (0 h) and after thefeeding experiment (�JA and þJA), based on five independent determinations. (J) Jasmonate (50 mM, JA treatment) or water (control) wassprayed on 3-week-old WT and coi1-1 plants. One day later one adult female thrip was allowed to feed on a leaf disc from each plant.Values are means� SDs of410 independent determinations.
74 JA but not ET acts in Arabidopsis tolerance to thrips
Dow
nloaded from https://academ
ic.oup.com/pcp/article/49/1/68/1903854 by guest on 09 D
ecember 2021
tolerance against thrip feeding. Thus, the JA pathway alone
is involved in tolerance against thrip feeding.
Discussion
The phytohormones JA, ET and SA regulate the plant’s
basal immune system. Numerous studies have examined the
functions of these hormones in plant response to pathogen
attack (reviewed by Pieterse et al. 2006). Here, we analyzed
the induction of gene expression by thrip feeding in
Arabidopsis to understand the plant’s hormonal response
to feeding. Gene expression patterns regulated by these
hormones are categorized into at least two signal pathways:
the JA/ET pathway and the SA pathway. Negative interac-
tion between the JA/ET and SA pathways has been clearly
shown by previous studies (e.g. Niki et al. 1998), and these
pathways have different effects on plant resistance to
pathogens (Uknes et al. 1992, Thomma et al. 1998).
Expression of the genes VSP2 and LOX2 (markers for
the JA pathway), PDF1.2 and chiB (markers for the JA/ET
pathway), and PR1 and BGL2 (markers for the SA
pathway) was induced by thrip feeding (Fig. 1). Schenk
et al. (2000) reported that up-regulation of some of the
Arabidopsis genes expressed under infection by A. brassici-
cola was also induced by treatments with JA, ET or SA,
suggesting interaction and coordination between the hor-
monal pathways in the plant defense response. From this
point of view, we analyzed the expression levels of marker
genes in the JA-insensitive mutant coi1-1, the ET-insensitive
mutants ein2-1 and ein3-1, and the SA-deficient mutant
eds16-1 fed on by thrips. Induction of VSP2 by thrip
feeding was apparently reduced in coi1-1 but not in ein2-1
and ein3-1. In contrast, induction of chiB was reduced in
coi1-1, and in ein2-1 and ein3-1 (Fig. 2A, B). Induction of
PR1 and BGL2 was reduced in eds16-1 (Fig. 2C). These
results suggest the possible importance of JA, ET and SA
signals in the response of Arabidopsis to thrip feeding.
Further comparative transcriptome analyses to show
the importance of these plant hormones indicated the greater
importance of JA than ET and SA in the plant’s response to
thrip feeding (Table 1A). As reported by Schenk et al. (2000),
we also found a significant relationship of gene expression
profiles between JA and ET treatments (Table 1B).
Therefore, we next determined the JA and ET contents of
plants with or without thrip feeding. JA contents were
apparently increased 24 h after thrip feeding (Fig. 3A),
whereas ET contents were not elevated (Fig. 3B). Expression
of AOS and AOC2, which regulate JA biosynthesis, was also
induced by thrip feeding (Fig. 3D, E). These results suggest
that JA biosynthesis after thrip feeding may be important for
A B C D
WT5E
4
3
2
Rel
ativ
e ar
ea o
f fee
ding
sca
rs
1
01 day 2 days
ein2-1
Control5F
4
3
2
Rel
ativ
e ar
ea o
f fee
ding
sca
rs
1
01 day 2 days
ACC treatment
Fig. 5 Ethylene does not have a positive effect on feeding tolerance against thrips. (A, B, E) One adult female thrip was allowed to feed ona leaf disc of 3-week-old WT plants and ein2-1 mutants. Typical image of leaf discs from WT (A) and ein2-1 (B) 2 d after the start of thripfeeding. (E) Mean� SD of the area of feeding scars, based on410 independent determinations. (C, D, F) The ET precursor ACC (25 mMdissolved in 25mM MES buffer, pH. 5.7; ACC treatment) or MES buffer (control) was sprayed on 3-week-old WT Arabidopsis plants 1 dbefore thrip feeding. One adult female thrip was allowed to feed on a leaf disc from each plant. Typical images of leaf discs from a control(C) and ACC-treated plant (D) 2 d after the start of thrip feeding are shown. (F) Mean� SDs of the area of feeding scars, based on410independent determinations.
JA but not ET acts in Arabidopsis tolerance to thrips 75
Dow
nloaded from https://academ
ic.oup.com/pcp/article/49/1/68/1903854 by guest on 09 D
ecember 2021

plant tolerance against thrips. The self-organizing maps
(SOMs) analyses to cluster the gene expression pattern found
how the correlation between thrip feeding and JA treatment
is different from the correlation between JA and ET treat-
ment in comparative transcriptome analyses. The correlation
between thrip feeding and JA treatment is based on the genes
which are induced by both thrip feeding and JA treatment,
but not by ET treatment. On the other hand, the correlation
between JA and ET treatment is based on the genes which are
induced by thrip feeding and repressed by JA and ET
treatment (data not shown).
JA and ET regulate the expression of PDF1.2 and chiB.
Expression of these genes was induced by thrip feeding, but
the induction was suppressed in the coi1-1, ein2-1 and ein3-1
mutants (Fig. 2A, B). Likewise, Penninckx et al. (1998)
reported that expression of PDF1.2 was not induced by JA
in ein2-1 nor by ET in coi1-1. Studies have demonstrated
that ERF1, a positive regulator of PDF1.2 and chiB gene
expression, acts as a key factor in the integration of both JA
and ET signals (reviewed by Lorenzo and Solano 2005). JA
and ET also induce the expression of ERF1. Consistent with
the expression of PDF1.2, the expression of ERF1 has not
been induced by JA in ein2-1, nor by ET in coi1-1. On the
basis of these findings, it has been suggested that the
simultaneous requirement for JA and ET signals for ERF1
expression explains the interaction of JA and ET signals
(Lorenzo and Solano 2005). Judging from these reports and
the absence of ET production after thrip feeding, expression
of PDF1.2 and chiB appears to be regulated by JA rather
than by the JA/ET pathway.
Although the SA-inducible genes PR1 and BGL2 were
induced by thrip feeding (Fig. 1B, C), correlation analyses
using transcriptome data did not indicate the importance of
SA in plant response to thrip feeding (Table 1A). Thus, the
biological significance of PR1 and BGL2 induction by thrip
feeding is unclear. Further investigations are needed to
clarify how Arabidopsis plants respond to thrip feeding
through the antagonistic plant hormones JA and SA.
Acquired resistance is one effective strategy for
protection against biotic or abiotic stresses (reviewed by
Thomashow 1999, Burke et al. 2000, Grant and Lamb
2006). We analyzed feeding tolerance in plants with or
without JA application to understand the function of JA in
acquired resistance against thrip feeding. Treatment of
Arabidopsis with JA enhanced tolerance to thrip feeding in
both leaf disc and whole-plant assays (Fig. 4J, K),
indicating that JA may induce acquired resistance against
thrip feeding in Arabidopsis. We also analyzed the feeding
tolerance in a JA-insensitive coi1-1 mutant. In contrast to
the results of the JA application assay, thrips fed on this
mutant more than on the WT plants (Fig. 4I).
Recent studies on plant–herbivore interactions indi-
cated the role of plant volatiles as olfactory signals for
insect behavior (Kessler and Baldwin 2001, Li et al. 2002,
Aharoni et al. 2003, Bruce et al. 2005). In this study we used
methyl jasmonate, a major plant volatile substance. To
eliminate the possibility that methyl jasmonate directly
changed the thrips’ behavior, we treated the coi1-1 mutant
with methyl jasmonate and found that this volatile had no
effect on thrip feeding on coi1-1 (Fig. 4L). Therefore, we
concluded that JA enhances resistance against thrip feeding
via JA-regulated defense responses.
Glucosinolates are secondary metabolites, the volatile
breakdown products of which have biological activity
against herbivores and pathogens (Bones et al. 1996,
Raybould and Moyes 2001, Barth and Jender 2006).
Sasaki-Sekimoto et al. (2005) reported that the biosynthesis
of glucosinolate is regulated by JA. Thus, glucosinolate may
be one of the metabolites that plays a role in JA-inducible
acquired resistance against thrip feeding.
In Arabidopsis, cev1, a constitutively active mutant for
JA signaling, showed higher tolerance to aphids than the
WT, whereas the JA-insensitive mutant coi1-1 showed
decreased tolerance against these insect pests (Ellis et al.
2002). Analyses of the interaction between the coi1-1
mutant and cabbage white butterfly larvae support the
importance of JA in insect feeding (Reymond et al. 2004).
Recently, the JAZ (jasmonate ZIM-domain) family of
repressors was identified as the negative regulator of JA
signaling (Chini et al. 2007, Thines et al. 2007, Yan et al.
2007). The JAZ repressor interacts with COI1 protein,
which leads the degradation of JAZ protein to induce
JA-responsive gene expression. Interestingly, JA-Ile but not
JA stimulates the interaction between JAZ repressor and
COI1. JA-Ile may be involved in the enhancement of
tolerance to thrip feeding promoted by JA application.
There is a long-term debate on a separate wounding
signal regulated by JA and the cyclopentenone precursor of
JA, 12-oxo-phytodienoic acid (OPDA). Taki et al. (2005)
identified a set of genes that specifically responded to
OPDA but not to JA, and they indicated a difference
between JA signaling and OPDA signaling. Interestingly, a
set of genes induced by thrip feeding includes these OPDA-
specific response genes (data not shown). Further analyses
using the opr3 mutant, which lack a major metabolizing
enzyme from OPDA to JA, would indicate the functional
importance and difference of JA and OPDA in plant
response and tolerance to thrip feeding.
Insect pests feed on plants through various modes.Most
studies have investigated lepidopteran larvae, which chew
plant tissues, and aphids, which suck phloem from plants.
Thrips, however, use their mouthparts to pierce plant cells
and suck out their contents (reviewed by Parrella 1995). Our
correlation analyses indicated the importance of JA in plant
response to thrip feeding, and similar results have
been reported in response to aphids and caterpillars.
76 JA but not ET acts in Arabidopsis tolerance to thrips
Dow
nloaded from https://academ
ic.oup.com/pcp/article/49/1/68/1903854 by guest on 09 D
ecember 2021

Reymond et al. (2004) reported that many genes induced by
Pieris rapae and Spodoptera littoralis feeding are controlled
by JA. To clarify the details of the various plant–herbivore
interactions, more research on the molecular differences in
plant responses to the different modes of insect feeding is
needed.
JA is involved in the wounding response of plants. In the
wounding pathway (JA pathway), ET has an antagonistic
effect (O’Donnell et al. 1996). However, JA acts in
cooperation with ET during the plant defense response to
fungal attack (JA/ET pathway). We analyzed the effect of
ET application on thrip feeding to understand the role of ET
in plant tolerance to thrip attack. When the ET precursor
ACC was applied to Arabidopsis, the area of thrip feeding
scars was greater than in the controls (Fig. 5F). In addition,
the ET-insensitive mutant ein2-1 was damaged by thrips to
an equal or lesser degree than were WT plants (Fig. 5E).
These results suggest that JA functions separately from ET
during the response to thrip feeding. Thus, the wounding
response may be involved in tolerance against the piercing
and sucking feeding mode of thrips.
Different gene expression in response to mechanical
wounding and insect feeding is reported (Reymond et al.
2000). It is known that gene expression is controlled by
insect oral factors as well as insect feeding (reviewed by
Korth 2003). In addition, Little et al. (2007) reported that
oviposition by Pieris brassicae triggers defense responses,
which are mostly independent of JA signaling. We should
analyze and understand the plant response challenged by
the complexity of insect attack including feeding, oral
factors and oviposition.
Our results demonstrated the importance of JA in the
plant response to, and tolerance against, thrip feeding. ET
did not have such an effect on thrip feeding, although its
action in the plant defense system is connected with that of
JA. In addition to feeding on plants, western flower thrips
also act as vectors of tospovirus (reviewed by Whitfield
et al. 2005). Thus, western flower thrips damage crops
through both feeding and associated disease. From this
point of view, apart from the JA pathway, there may be
other important pathway(s) that regulate the plant defense
response to feeding against viruliferous thrips. Further
research, including transcriptome and metabolome ana-
lyses, is required to better our understanding of plant–
herbivore interactions. Our results showed that the
Arabidopsis–thrips system is a good tool for clarifying the
detailed mechanisms of the plant defense response to this
important agricultural pest.
Materials and Methods
Plant materials and cultivation
Plants (A. thaliana ecotype Col-0) were grown in soil asdescribed previously (Weigel and Glazebrook 2002). Several
mutants related to plant hormone biosynthesis and signaling(coi1-1, ein2-1, ein3-1, eto1-1 and eds16–1) were used in this study.Arabidopsis seeds were sown on sterile soil in pots, moistened, andplaced in a cold room at 48C in the dark to synchronizegermination. The pots were then transferred to 228C with a long-day photoperiod (16 h light/8 h dark). Plants at the four-leafstage were transferred individually to pots and grown to therosette stage.
Identification of coi1-1 plants
Homozygous coi1-1 plants were selected by PCR analysisfollowed by BsmI digestion. Genomic DNA was extracted from asingle rosette leaf of each plant and purified with a DNeasy PlantKit (Qiagen, Valencia, CA, USA), and the PCR was performedwith ExTaq DNA polymerase (TAKARA, Kyoto, Japan).Nucleotide sequences of primers were as follows: forward primer,50-GGAAACAGGAGCCCGAGATC-30; reverse primer, 50-TGGATGTTTCTCGGAGCAGC-30. Amplified DNA was treated withBsmI, which digests DNA from the coi1-1mutant only. The reactionmixture was analyzed by agarose gel electrophoresis.
Thrip feeding
Laboratory colonies of F. occidentalis were maintained in aclosed environmental chamber, as described previously (Murai andLoomans 2001). For the assay, female adults 14–21 d afteremergence from the pupal stage were used. They were starved for2–3 h before feeding on test plants. Twenty-five adult females weregrouped and allowed to feed on each whole plant in an acrylcylinder chamber with air ventilation windows covered with a finemesh. This system confined the feeding adults to the chamber andprevented their escape.
Leaf disc assay
Leaf discs with an 8mm diameter were produced with abiopsy punch (Kay Industries, Oyana, Japan). Leaf discs werefloated on 1.5ml of distilled water in wells of a white 1.5ml sampletube stand (Assist, Tokyo, Japan). A single adult female that hadbeen starved for 2–3 h was placed on a single leaf disc. The hole ofthe sample tube was covered with ABI Prism Optical AdhesiveCover (Applied Biosystems, Foster City, CA, USA), and a few tinyholes for air vents during feeding were made by piercing theadhesive cover with a 27G fine injection needle. Thrips wereallowed to feed for 1 or 2 d at 228C. The area of thrip feeding scarson the surface of each leaf disc was measured by ImageJ software(Abramoff et al. 2004) on digitized images taken under a VHX-200digital microscope (Keyence, Osaka, Japan). The measured area ofleaf discs from each treatment was analyzed using JMP software,ver. 5.1 (SAS Institute, Inc., Cary, NC, USA).
Electrolyte leakage assay
Three-week-old Arabidopsis plants grown in soil were fed onby 25 adult female thrips for 1 or 2 d. An electrolyte leakage assaywas performed as described by Weigel et al. (2001).
Arabidopsis 1.2K cDNA microarray analysis
Total RNA was isolated with Trizol reagent (Invitrogen,Carlsbad, CA, USA). mRNA was prepared with an MACSmRNA Isolation Kit (Miltenyi Biotec, Bergisch Glabach,Germany). To ensure the reproducibility of the microarray results,we replicated the experiment twice with 10 Arabidopsis plants persample. To obtain sufficient material for the experiments, wepooled samples from duplicate experiments before RNA
JA but not ET acts in Arabidopsis tolerance to thrips 77
Dow
nloaded from https://academ
ic.oup.com/pcp/article/49/1/68/1903854 by guest on 09 D
ecember 2021

extraction. Preparation of fluorescent probes, microarray hybridi-zation, scanning and data analysis were as described previously(Seki et al. 2001). Image analyses and signal quantification wereperformed with ScanArray Express ver. 3.0 (PerkinElmer,Waltham, MA, USA). A lambda control template DNA fragment(TAKARA) was used as an external control to equalize thehybridization signals generated from different samples (Narusakaet al. 2006). The median of 82 signal values obtained from thelambda control template DNA fragment on the 1.2K microarraywas used as an external control. Each experiment was repeatedthree times.
Quantitative reverse–transcription PCR
Five 3-week-old Arabidopsis plants at the rosette stage grownin the same pot were fed on for 5, 10 or 24 h by 25 female adultthrips in the closed container with air vents. Experiments wererepeated twice. After feeding, the plants were frozen in liquidnitrogen for further analyses. Total RNA (2mg) isolated by usingTrizol reagent (Invitrogen) was treated with RNase-free DNase(TAKARA) to eliminate genomic DNA contamination. First-strand cDNA was synthesized with random oligo-hexamers withSuperscript III reverse transcriptase according to the manufac-turer’s instructions (Invitrogen). Quantitative real-time PCR wascarried out with SYBR Green PCR Master Mix (AppliedBiosystems) using the first-strand cDNA as a template on asequence detector system (ABI Prism 7900HT, AppliedBiosystems). Expression of CBP20 was used for normalization asa standard control gene. Nucleotide sequences of the gene-specificprimers were as follows: VSP2 (At5g24770; forward primer,50-GTTAGGGACCGGAGCATCAA-3; reverse primer, 50-AACGGTCACTGAGTATGATGGGT-30); LOX2 (At3g45140; forwardprimer, 50-TTGCTCGCCAGACACTTGC-30; reverse primer,50-GGGATCACCATAAACGGCC-30); chiB (At3g12500; forwardprimer, 50-ACGGAAGAGGACCAATGCAA-30; reverse primer,50-GTTGGCAACAAGGTCAGGGT-30);PDF1.2 (At5g44420; for-ward primer, 50-CCATCATCACCCTTATCTTCGC-30; reverseprimer, 50-TGTCCCACTTGGCTTCTCG-30); BGL2 (At3g57260;forward primer, 50-GCCGACAAGTGGGTTCAAGA-30; reverseprimer, 50-AACCCCCCAACTGAGGGTT-30); PR1 (At2g14610;forward primer, 50-GTTGCAGCCTATGCTCGGAG-30; reverseprimer, 50-CCGCTACCCCAGGCTAAGTT-30); CBP20 (At5g44200; forward primer, 50-CCTTGTGGCTTTTGTTTCGTC-30;reverse primer, 50-ACACGAATAGGCCGGTCATC-30); AOS(At5g42650; forward primer, 50-TCTTCTCTTCGCCACGTGC-30;reverse primer, 50-GGTTATGAACTTGATGACCCGC-3); AOC2(At3g25780; forward primer, 50-CGGCAAGAAACCAACAGAGC-30; reverse primer, 50-GACCTGCCGTGATTCCCAC-30).
Jasmonate quantification
Quantification of JA and its methyl ester was performed asdescribed previously (Seo et al. 1995), except that an HP6890 gaschromatograph fitted to a quadrupole mass spectrometer (Hewlett-Packard, Wilmington, DE, USA) was used for the analysis.Approximately 1 g of each Arabidopsis plant with or without thripfeeding was used for quantification. Analyses were performed onthree independent samples.
Ethylene quantification
ET was quantified with a gas chromatograph (GC-8A;Shimadzu, Kyoto, Japan) as described previously (Ohtsuboet al. 1999).
Accession numbers
The Arabidopsis Genome Initiative gene codes (AGI codes)and GenBank accession Nos., respectively, for the genes mentionedin this article are as follows: VSP2 (At5g24770, AB006778), LOX2(At3g45140, AYO62611), chiB (At3g12500, AY054628), PDF1.2(At5g44420, AY063779), BGL2 (At3g57260, AY099668), PR1(At2g14610, AY064023), CBP20 (At5g44200, AF140219), AOS(At5g42650, AY062828) and AOC2 (At3g25780, AJ308484).
Funding
Ministry of Education, Culture, Sports, Science and
Technology Grant-in-Aid for Scientific Research in area
‘Young Scientist (B)’ to H.A.; 2004 Industrial Technology
Research Grant Program of the New Energy and Industrial
Technology Development Organization of Japan to Y.N.
and H.A.
Acknowledgments
We thank I. Sasaki, S. Kawamura and F. Mori of RIKENBRC, and A. Kawashima, S. Nagai and Y. Matsumura of theNational Agricultural Research Center for their excellent technicalassistance. We also thank Dr. M. Watanabe of NIAS for his kindsupport and helpful discussions, and Dr. K. Yamaguchi-Shinozakiof JIRCAS and Dr. T. Shimoda of NARC for their helpfuldiscussions.
References
Abramoff, M.D., Magelhaes, P.J. and Ram, S.J. (2004) Image processingwith ImageJ. Biophotonics Int. 11: 36–42.
Aharoni, A., Giri, A.P., Deuerlein, S., Griepink, F., de Kogel, W.J.,Verstappen, F.W., Verhoeven, H.A., Jongsma, M.A., Schwab, W. andBouwmeester, H.J. (2003) Terpenoid metabolism in wild-type andtransgenic Arabidopsis plants. Plant Cell 15: 2866–2884.
Alonso, J.M., Hirayama, T., Roman, G., Nourizadeh, S. and Ecker, J.R.(1999) EIN2, a bifunctional transducer of ethylene and stress responses inArabidopsis. Science 284: 2148–2152.
Barth, C. and Jander, G. (2006) Arabidopsis myrosinases TGG1 and TGG2have redundant function in glucosinolate breakdown and insect defense.Plant J. 46: 549–562.
Bones, A.M., Tas, E. and Rossiter, J.T. (1996) The myrosinase–glucosinolate system, its organisation and biochemistry. Physiol. Plant97: 194–208.
Bruce, T.J., Wadhams, L.J. and Woodcock, C.M. (2005) Insect hostlocation: a volatile situation. Trends Plant Sci. 10: 269–274.
Burke, J.J., O’Mahony, P.J. and Oliver, M.J. (2000) Isolation of Arabidopsismutants lacking components of acquired thermotolerance. Plant Physiol.123: 575–588.
Campbell, M.A., Fitzgerald, H.A. and Ronald, P.C. (2002) Engineeringpathogen resistance in crop plants. Transgenic Res. 11: 599–613.
Chao, Q., Rothenberg, M., Solano, R., Roman, G., Terzaghi, W. andEcker, J.R. (1997) Activation of the ethylene gas response pathway inArabidopsis by the nuclear protein ETHYLENE-INSENSITIVE3 andrelated proteins. Cell 89: 1133–1144.
Chini, A., Fonseca, S., Fernandez, G., Adie, B., Chico, J.M., et al. (2007)The JAZ family of repressors is the missing link in jasmonate signaling.Nature 448: 666–671.
Cohen, J. (1998) Differences between correlation coefficients. In StatisticalPower Analysis for the Behavioral Sciences, 2nd edn. Edited by Cohen, J.pp. 109–143. Lawrence Erlbaum Associates, Florence.
78 JA but not ET acts in Arabidopsis tolerance to thrips
Dow
nloaded from https://academ
ic.oup.com/pcp/article/49/1/68/1903854 by guest on 09 D
ecember 2021

Cooper, W.C., Jia, L. and Goggin, F.L. (2004) Acquired and R-gene-mediated resistance against the potato aphid in tomato. J. Chem. Ecol.30: 2527–2542.
Delker, C., Stenzel, I., Hause, B., Miersch, O., Feussner, I. andWasternack, C. (2006) Jasmonate biosynthesis in Arabidopsis thaliana:enzymes, products, regulation. Plant Biol. (Stuttg.) 8: 297–306.
Dewdney, J., Reuber, T.L., Wildermuth, M.C., Devoto, A., Cui, J.,Stutius, L.M., Drummond, E.P. and Ausubel, F.M. (2000) Three uniquemutants of Arabidopsis identify eds loci required for limiting growth of abiotrophic fungal pathogen. Plant J. 24: 205–218.
Ellis, C., Karafyllidis, I. and Turner, J.G. (2002) Constitutive activation ofjasmonate signaling in an Arabidopsis mutant correlates with enhancedresistance to Erysiphe cichoracearum, Pseudomonas syringae, and Myzus
persicae. Mol. Plant-Microbe Interact. 15: 1025–1030.Grant, M. and Lamb, C. (2006) Systemic immunity. Curr. Opin. Plant Biol.
9: 414–420.Hui, D., Iqbal, J., Lehmann, K., Gase, K., Saluz, H.P. and Baldwin, I.T.
(2003) Molecular interactions between the specialist herbivore Manduca
sexta (Lepidoptera, Sphingidae) and its natural host Nicotiana attenuata.
V. Microarray analysis and further characterization of large-scalechanges in herbivore-induced mRNAs. Plant Physiol. 131: 1877–1893.
Jensen, S.E. (2000) Insecticide resistance in the western flower thrips,Frankliniella occidentalis. Integr. Pest. Manag. Rev. 5: 131–146.
Kahl, J., Siemens, D.H., Aerts, R.J., Gabler, R., Kuhnemann, F.,Preston, C.A. and Baldwin, I.T. (2000) Herbivore-induced ethylenesuppresses a direct defense but not a putative indirect defense against anadapted herbivore. Planta 210: 336–342.
Kessler, A. and Baldwin, I.T. (2001) Defensive function of herbivore-induced plant volatile emissions in nature. Science 291: 2141–2144.
Kessler, A. and Baldwin, I.T. (2002) Plant responses to insect herbivory: theemerging molecular analysis. Annu. Rev. Plant Biol. 53: 299–328.
Korth, K.L. (2003) Profiling the response of plants to herbivorous insects.Genome Biol. 4: 221.1–221.4.
Korth, K.L. and Thompson, G.A. (2006) Chemical signals in plants:jasmonates and the role of insect-derived elicitors responses toherbivores. In Multigenic and Induced Systemic Resistance in Plants.Edited by Tuzun, S. and Bent, E. pp. 259–278. Springer, New York.
Li, C., Liu, G., Xu, C., Lee, G.I., Bauer, P., Ling, H.Q., Ganal, M.W. andHowe, G.A. (2003) The tomato suppressor of prosystemin-mediated
responses2 gene encodes a fatty acid desaturase required for thebiosynthesis of jasmonic acid and the production of a systemic woundsignal for defense gene expression. Plant Cell 15: 1646–1661.
Li, X., Schuler, M.A. and Berenbaum, M.R. (2002) Jasmonate andsalicylate induce expression of herbivore cytochrome P450 genes.Nature 419: 712–715.
Little, D., Gouthier-Darimont, C., Bressow, F. and Reymond, P. (2007)Oviposition py pierid butterflies triggers defense responses inArabidopsis. Plant Physiol. 143: 784–800.
Lorenzo, O. and Solano, R. (2005) Molecular players regulating thejasmonate signalling network. Curr. Opin. Plant Biol. 8: 532–540.
Moran, P.J. and Thompson, G.A. (2001) Molecular responses to aphidfeeding in Arabidopsis in relation to plant defense pathways. Plant
Physiol. 125: 1074–1085.Moran, P.J., Cheng, Y., Cassell, J.L. and Thompson, G.A. (2002) Gene
expression profiling of Arabidopsis thaliana in compatible plant–aphidinteractions. Arch. Insect Biochem. Physiol. 51: 182–203.
Murai, T. and Loomans, A.J.M. (2001) Evaluation of an improved methodfor mass-rearing of thrips and a thrips parasitoid. Ent. Exp. Appl. 101:281–289.
Narusaka, M., Abe, H., Kobayashi, M., Kubo, Y., Kawai, K., Izawa, N.and Narusaka, Y. (2006) A model system to screen for candidate plantactivators using immmune-induction system in Arabidopsis. Plant
Biotech. 23: 321–327.Narusaka, Y., Narusaka, M., Park, P., Kubo, Y., Hirayama, T., et al.
(2004) RCH1, a locus in Arabidopsis that confers resistance to thehemibiotrophic fungal pathogen Colletotrichum higginsianum. Mol.Plant-Microbe Interact. 17: 749–762.
Narusaka, Y., Narusaka, M., Seki, M., Fujita, M., Ishida, J., et al. (2003a)Expression profiles of Arabidopsis phospholipase A IIA gene in responseto biotic and abiotic stresses. Plant Cell Physiol. 44: 1246–1252.
Narusaka, Y., Narusaka, M., Seki, M., Ishida, J., Nakashima, M., et al.(2003b) The cDNA microarray analysis using an Arabidopsis pad3mutant reveals the expression profiles and classification of genes inducedby Alternaria brassicicola attack. Plant Cell Physiol. 44: 377–387.
Niki, T., Mitsuhara, I., Seo, S., Ohtsubo, N. and Ohashi, Y. (1998)Antagonistic effect of salicylic acid and jasmonic acid on the expressionof pathogesesis-related (PR) protein genes in wounded mature tobaccoleaves. Plant Cell Physiol. 39: 500–507.
O’Donnell, P.J., Calvert, C., Atzorn, R., Wasternack, C., Leyser, H.M.O.and Bowles, D.J. (1996) Ethylene as a signal mediating the woundingresponse of tomato plants. Science 274: 1914–1917.
Ohtsubo, N., Mitsuhara, I., Koga, M., Seo, S. and Ohashi, Y. (1999)Ethylene promotes the necrotic lesion formation and basic PR geneexpression in TMV-infected tobacco. Plant Cell Physiol. 40: 808–817.
Parrella, M.P. (1995) IPM: approaches and prospects. In Thrips Biologyand Management. Edited by Parker, B.L., Skinner, M. and Lewis, T.pp. 357–364. Plenum, New York.
Pegadaraju, V., Knepper, C., Reese, J. and Shah, J. (2005) Premature leafsenescence modulated by the Arabidopsis PHYTOALEXINDEFICIENT4 gene is associated with defense against the phloem-feedinggreen peach aphid. Plant Physiol. 139: 1927–1934.
Penninckx, I.A., Thomma, B.P., Buchala, A., Metraux, J.P. andBroekaert, W.F. (1998) Concomitant activation of jasmonate andethylene response pathways is required for induction of a plant defensingene in Arabidopsis. Plant Cell 10: 2103–2113.
Pieterse, C.M.J., Schaller, A., Mauch-Mani, B. and Conrath, U. (2006)Signaling in plant resistance responses: divergence and cross-talk ofdefense pathways. In Multigenic and Induced Systemic Resistance inPlants. Edited by Tuzun, S. and Bent, E. pp. 166–196. Springer,New York.
Raybould, A.F. and Moyes, C.L. (2001) The ecological genetics of aliphaticglucosinolates. Heredity 87: 383–391.
Reymond, P., Bodenhausen, N., Van Poecke, R.M., Krishnamurthy, V.,Dicke, M. and Farmer, E.E. (2004) A conserved transcript pattern inresponse to a specialist and a generalist herbivore. Plant Cell 16:3132–3147.
Reymond, P., Weber, H., Damond, M. and Farmer, E.E. (2000)Differential gene expression in response to mechanical wounding andinsect feeding in Arabidopsis. Plant Cell 12: 707–719.
Sasaki-Sekimoto, Y., Taki, N., Obayashi, T., Aono, M., Matsumoto, F.,et al. (2005) Coordinated activation of metabolic pathways forantioxidants and defence compounds by jasmonates and their roles instress tolerance in Arabidopsis. Plant J. 44: 653–668.
Schenk, P.M., Kazan, K., Wilson, I., Anderson, J.P., Richmond, T.,Somerville, S.C. and Manners, J.M. (2000) Coordinated plant defenseresponses in Arabidopsis revealed by microarray analysis. Proc. NatlAcad. Sci. USA 97: 11655–11660.
Seki, M., Narusaka, M., Abe, H., Kasuga, M., Yamaguchi-Shinozaki, K.,Carninci, P., Hayashizaki, Y. and Shinozaki, K. (2001) Monitoring theexpression pattern of 1300 Arabidopsis genes under drought and coldstresses by using a full-length cDNA microarray. Plant Cell 13: 61–72.
Seki, M., Narusaka, M., Ishida, J., Nanjo, T., Fujita, M., et al. (2002)Monitoring the expression profiles of 7000 Arabidopsis genes underdrought, cold and high-salinity stresses using a full-length cDNAmicroarray. Plant J. 31: 279–292.
Seo, S., Okamoto, M., Seto, H., Ishizuka, K., Sano, H. and Ohashi, Y.(1995) Tobacco MAP kinase: a possible mediator in wound signaltransduction pathways. Science 270: 1988–1992.
Taki, N., Sekimoto, Y., Obayashi, T., Kikuta, A., Ainai, T., Kobayashi, Y.,Sakurai, N., Suzuki, H., Takamiya, K., Shibata, D. and Ohta, H. (2005)12-oxo-phytodienoic acid triggers expression of a distinct set of genesfrom jasmonates, and plays a role other than intermediate of jasmonatesbiosynthesis. Plant Physiol. 139: 1268–1283.
Thines, B., Katsir, L., Melotto, M., Niu, Y., Mandaokar, A., Liu, G.,Nomura, K., He, S.H., Howe, G.A. and Browse, J. (2007) JAZ repressorproteins are targets of the SCFCOI1 complex during jasmonate signaling.Nature 448: 661–665.
Thomashow, M.F. (1999) PLANT COLD ACCLIMATION: freezingtolerance genes and regulatory mechanisms. Annu. Rev. Plant Physiol.
Plant Mol. Biol. 50: 571–599.
JA but not ET acts in Arabidopsis tolerance to thrips 79
Dow
nloaded from https://academ
ic.oup.com/pcp/article/49/1/68/1903854 by guest on 09 D
ecember 2021

Thomma, B., Eggermont, K., Penninckx, I., Mauch-Mani, B.,Vogelsang, R., Cammue, B.P.A. and Broekaert, W.F. (1998) Separatejasmonate-dependent and salicylate-dependent defense-response path-ways in Arabidopsis are essential for resistance to distinct microbialpathogens. Proc. Natl Acad. Sci. USA 95: 15107–15111.
Thomma, B.P., Penninckx, I.A., Broekaert, W.F. and Cammue, B.P. (2001)The complexity of disease signaling in Arabidopsis. Curr. Opin. Immunol.13: 63–68.
Thompson, G.A. and Goggin, F.L. (2006) Transcriptomics and functionalgenomics of plant defence induction by phloem-feeding insects. J. Exp.Bot. 57: 755–766.
Turlings, T.C. and Ton, J. (2006) Exploiting scents of distress: the prospectof manipulating herbivore-induced plant odours to enhance the controlof agricultural pests. Curr. Opin. Plant Biol. 9: 421–427.
Uknes, S., Mauch-Mani, B., Moyer, M., Potter, S., Williams, S.,Dincher, S., Chandler, D., Slusarenko, A., Ward, E. and Ryals, J.(1992) Acquired resistance in Arabidopsis. Plant Cell 4: 645–656.
Walling, L.L. (2000) The myriad plant responses to herbivores. J. PlantGrowth Regul. 19: 195–216.
Weigel, D. and Glazebrook, J. (2002) How to grow Arabidopsis.In Arabidopsis: A Laboratory Manual. Edited by Weigel, D. andGlazebrook, J. pp. 1–18. Cold Spring Harbor Laboratory Press, ColdSpring Harbor, NY.
Weigel, R.R., Bauscher, C., Pfitzner, A.J. and Pfitzner, U.M. (2001)
NIMIN-1, NIMIN-2 and NIMIN-3, members of a novel family of
proteins from Arabidopsis that interact with NPR1/NIM1, a key
regulator of systemic acquired resistance in plants. Plant Mol. Biol. 46:
143–160.Whitfield, A.E., Ullman, D.E. and German, T.L. (2005) Tospovirus–thrips
interactions. Annu. Rev. Phytopathol. 43: 459–489.Wildermuth, M.C., Dewdney, J., Wu, G. and Ausubel, F.M. (2001)
Isochorismate synthase is required to synthesize salicylic acid for plant
defence. Nature 414: 562–565.Winz, R.A. and Baldwin, I.T. (2001) Molecular interactions between the
specialist herbivore Manduca sexta (Lepidoptera, Sphingidae) and its
natural host Nicotiana attenuata. IV. Insect-induced ethylene reduces
jasmonate-induced nicotine accumulation by regulating putrescine
N-methyltransferase transcripts. Plant Physiol. 125: 2189–2202.Xie, D.X., Feys, B.F., James, S., Nieto-Rostro, M. and Turner, J.G. (1998)
COI1: an Arabidopsis gene required for jasmonate-regulated defense and
fertility. Science 280: 1091–1094.Yan, Y., Stolz, S., Chetelat, A., Reymond, P., Pagni, M., Dubugnon, L. and
Farmera, E.E. (2007) A downstream mediator in the growth repression
limb of the jasmonate. Plant Cell 15: 2866–2884.
(Received November 9, 2007; Accepted November 19, 2007)
80 JA but not ET acts in Arabidopsis tolerance to thrips
Dow
nloaded from https://academ
ic.oup.com/pcp/article/49/1/68/1903854 by guest on 09 D
ecember 2021
![Physiology & Behavior - UZH7682ba26-1f51-45c5...ural feeding behavior of proboscis monkeys that cease feeding during the night [8], no food was provided after 18:00. The dry matter](https://img.dokumen.tips/doc/110x75/5f09f7957e708231d4295e5f/physiology-behavior-uzh-7682ba26-1f51-45c5-ural-feeding-behavior-of-proboscis.jpg)




























