Embed Size (px)
Citation preview

8/13/2019 The xylem sap of maple [Acer platanoides) trees—sap obtained by a novel method shows changes with season a…
http://slidepdf.com/reader/full/the-xylem-sap-of-maple-acer-platanoides-treessap-obtained-by-a-novel-method 1/11
Journal o f xperimental Botany Vol. 47, No. 294, pp. 123-133, January 1996Journal ofExperimentalBotany
Th e xylem sap of maple [Acer platanoides) t rees—sap
obtained by a novel method shows changes with season
and height
Volker Schill1-4'5, Wolfram Hartun g 2 , Birgit Or then 3 and Manfred H. Weisenseel4
11nstitut f r Mechanische Verfahrenstechnik und Mechanik, Universita t Karlsruhe (TH), D-76128 Karlsruhe,
Germany2 Julius-von-Sachs-lnstitut fQr Biowisse nschaften der Universita t WOrzburg Lehrstuhl Botanik I,
Mittlerer Da llenbergweg 64, D-9708 2 WOrzburg Germany3 Institut f r Okologie der Pflanzen, Westfa lische Wilhelms-Universita t, D-48143 Munster, Germany
4 Botanisches Institut, Lehrstuhl I Universita t Karlsruhe (TH), D-76128 Karlsruhe, G ermany
Received 9 May 1995; Acc epted 18 September 1995
A b s t r a c t
Xylem sap of log pieces of maple trees was collected
by a novel pressure/decompression method developed
recently for the mechanical drying of timber. Seasonal
changes and spatial distributions of the osmotic poten-
t ial , the pH and the concentrations of major solutes
and of the plant stress-hormone abscisic acid (ABA)
were investigated.
Sucrose and quebrachitol were the main compon-ents of the xylem sap. The sucrose concentration
varied betwee n 10 mM and 25 mM during the winter
months and declined to a minimum during the period
of bud burst. Quebrachitol was found in concentrations
of up to 7 mM with a high variability throughout the
year. High concentrations of ABA were measured
during the summer season and in mid-winter. Rainfall
caused an increase of ABA in some samples.
The osmotic potential of the xylem sap increased
with the height of the collection site. The pH of the
sap decreased by approximately one unit between the
base of the trunk and the crown. The increase of
the osmotic potential was mainly due to sucrose,
quebrachitol and potassium. M alate contr ibuted to the
decrease of the pH. ABA of the xylem sap increased
with decreasing moisture content of the wood.
Key words: Pressure/decompression method, xylem sap,
abscisic acid, sucrose, quebrachitol, Acer platanoides.
In t r o d u c t io n
Recently, Schill e t al. (1992) developed a new method forthe drying of wood. In contrast to conventional dryingprocesses, the novel method drives water as a liquid outof the wood by establishing a pressure difference betweenthe centre of a wood piece and its surface. The liquidobtained is pure xylem sap (see Discussion). The methodis rapid and straightforward. Its application requires
neither wood samples of a special shape nor seals atcertain surfaces. The novel technique is also useful forresearch into xylem transport in ligneous plants becausexylem sap from roots, trunks, branches, and twigs ofthese plants can be obtained rapidly and easily in largequantities.
In this study the xylem sap of young Acer platanoidestrees was investigated. The seasonal changes in the con-centrations of specific carbohydrates were recorded. Inaddition, the plant stress-hormone abscisic acid (ABA)was measured because it plays a major role as a root-to-shoot stress signal that is transported in the xylem (Davies
and Zhang, 1991; Khalil and Grace, 1993). The ABAconcentration in the xylem sap was plotted ag ainst rainfallto see whether a low soil moisture status, or increasingdrought stress at the root, respectively, may act as astimulus for the loading of the xylem with ABA in trees.
The spatial distribution between the base of the trunkand the crown of the xylem sap osmotic potential (n),the pH-value and those solutes that contribute most to n
8 To whom correspondence should be addressed. Fax: +49 72 69 39 65.
Abbreviations: ABA, abscisic acid; wmc, wood moisture content; bb, bud burst; Is, leaf senescence; la, leaf abscission.
© Oxford Un iversity Press 1996

8/13/2019 The xylem sap of maple [Acer platanoides) trees—sap obtained by a novel method shows changes with season a…
http://slidepdf.com/reader/full/the-xylem-sap-of-maple-acer-platanoides-treessap-obtained-by-a-novel-method 2/11
12 4 Schill et al .
and pH was also investigated. Potassium (K+) in the
xylem sap was measured to find out whether living woodcells are damaged during the pressure treatment and,therefore, release K
+ into the apoplast. The ABA concen-
tration in the xylem sap and the wood moisture contentof individual stem segments were examined to obtaininformation about whether the local moisture status of
the tissue can influence the xylem sap ABA content.
Materials and methods
Preparation of wood samples
In 1992 and 1993 appro xim ately 50, 8-10-year- old trees ofNorway maple (Acer platcmoides) were felled. The trees grew inthe Hardtwald forest near Karlsruhe, Germany, and measured4- 6 m in height. Imm ediately after cu tting the trees between09.00 h an d 10.00 h, segm ents of 25 cm in length were tak enfrom the basal part of the logs, approximately 0.1-1.1 m above-ground , and trimmed to a length of 15 cm. The bark of thesegments was removed, and ap proxim ately 50% of the segmentswere cleaved so that they would fit into the sap collecting
con taine rs (F ig. 1). All samples were sto red in plastic foilduring the transport to the laboratory and subjected to xylemextraction w ithin 30 min after cutting. To measure the distribu-tion of solutes between the base of the trunk and the crown,two separate maple trees were cut into segments as shown inFig. 6. Furth er preparatio n of the samples was as describedabove.
air out
soundabsorber
bottle compressor
Fig. 1. Schematic drawing of the apparatus used for the collection ofxylem sap (not to scale). After releasing the pressure, xylem sap leavesthe wood samples and is collected at the bottom of the containersinside the pressure chamber. Pressure chamber dimensions are: Insidediameter =5 3 mm, length = 3 m; maximum permissible pressure =lOMPa.
Collection of xylem sap
The log samples were placed into the sap collecting containersof a pressure chamber (Fig. 1). A pressure of 4 MPa was thenapplied for 30 s (=p ress ure period) and afterwards releasedsuddenly to ambient pressure (=decompression period). In theperiod of ambient pressure, xylem sap starts to move withinthe wood along the xylem vessels and leaves at the cuttingfaces. After 4.5 min, the pressure/decompression treatmen t wasrepeated. Routinely, five successive cycles of pressure anddecompression were applied. The xylem sap that accumulatedduring the treatment in the containers was frozen and stored at— 18 °C for further analysis. The dry weight of the log sampleswas measured according to the German stand ard DI N 52183procedure. The wood moisture content (wmc) was calculatedfrom the weight of the freshly prepared samples (n' frMh) and theweight of the dried samples (w dr y):
wm c = ( w fTCtb - irdr y)/u'dr y x 100%.
Analysis of sugars and sugar alcohols in the xylem sap
A vo lume of 4 ml of xylem sap was mixed with 2 ml of an a-D-methyl-glucopyranos id solut ion (S m g m T 1 ) as internal stand-ard. The mixture was prepurified with a 10 x3 0 mm A 12O 3
column and forced thro ugh 0.45 ^m syringe filters. Analysis of
sugars was conducted with an HPLC apparatus using a4.6 x 250 mm colum n filled with Poly gosil-N H 2 5 fim 60 A(Macherey-Nagel, Dtlren, Germany), a type 6000A pump(Waters, Eschborn, Germany), a type R401 RI detector(Waters) and a type CR3A Chromatopac integrator (Shimadzu,Duisburg, Germany). The sugar content of the xylem sap wascalculated from separate measurements using solutions ofstandard sugars (Merck, Darmstadt, Germany, analytical grade)or quebrachitol (Aldrich, Steinheim, Germany, pure grade).The limit for quantitative detection was approximately 1.5 mMfor glucose and fructose and approximately 1 mM for sucroseand quebrachitol.
Analysis of quebrachitol and organic acids
The xylem sap was deionized with bonded silica columns
(Sepralyte SCX and NH 2 , ICT-Ass-chem, Bad Homburg,Germany). The neutral fraction was eluated with aqua dest. ,the acid fraction with 1 N HC1. Quebrachitol in the neutralfraction was measured as trimethylsilylether on glass columnsof 2 m x 2 mm packed either with 3 % SE 30 or 3 % Dexsil 300(Richter et al, 1990). Orga nic acids were analysed as trimethyl-silyl derivatives on a glass column packed with 3% OV 225(ICT-Ass-chem). The detector and injector (type 3600, Varian,Darm stadt, Germ any) temperatures were 250 °C and 220 °C,respectively. The temperature of the column oven was initially100 °C and program med to r ise at S X m i n
1 to 230 °C . T he
carrier gas was nitrog en with a flow rate of 25 ml m in1.
Measurement of pH and osmotic potential
The pH of the xylem sap was measured using a type pH196pH meter (WTW, Weilheim, Germany) with a type 403electrode (Ingold, Steinbach, Germany). The osmotic potentialwas measured cryoscopically employing an Osm oma t 030nanoliter osmometer (Gonotec, Berlin, Germany).
Analysis of ABA and potassium
Abscisic acid was measured immunologically using an ELISAwith monoclonal antibodies as described by Mertens et al(1983). Aliquots of xylem sap were mixed with TBS-buffer(TRIS-buftered sa line: 50 m o lm
3 T R I S , 1 m o l m ~
3 MgCl2 ,
150 mol m ~3 NaCl; pH 7.8) and subjected to the ELISA. For
8/13/2019 The xylem sap of maple [Acer platanoides) trees—sap obtained by a novel method shows changes with season a…
http://slidepdf.com/reader/full/the-xylem-sap-of-maple-acer-platanoides-treessap-obtained-by-a-novel-method 3/11
Xylem sap of maple 125
analysis of potassium in the xylem sap a type FMD 3 atomicabsorption spectrometer (Zeiss, Oberkochen. Germany) wasemployed.
Preparation of scanning electron micrographs
Cubes of approx ima te ly 7 x 7 x 7 mm 3 were cut out of severaldifferent samples of maple wood that had been subjected to thepressure/decompression treatment as described above. The
cubes were soaked with ethanol overnight. Subsequently theywere cut with a razor blade and air-dried for approximately24 h. After sputter ing with go ld to a layer of 180 nm , th esamples were examined under a type JSM 6400 scanningelectron microscope (Japan Electron Optics Laboratory Ltd. ,Tokyo, Japa n). Th e acceleration voltage was 25 or 28 kV,respectively.
Results
Seasonal changes of wood moisture content and xylem sap
components
The wood moisture content of 52 A. platanoides trees was
measured in the course of two successive years, and theresults are shown in Fig. 2. The highest values, i.e. wood
moisture contents of 85-100%, were found in late winter
(January and February) and in spring during the period
from dormancy to bud burst, which started in the middle
of March. Up to autumn, the wood moisture content
decreased gradually and reached very low values of about
55% in September and October. Afterwards the wood
moisture content increased again. This increase started
during leaf abscission early in November.
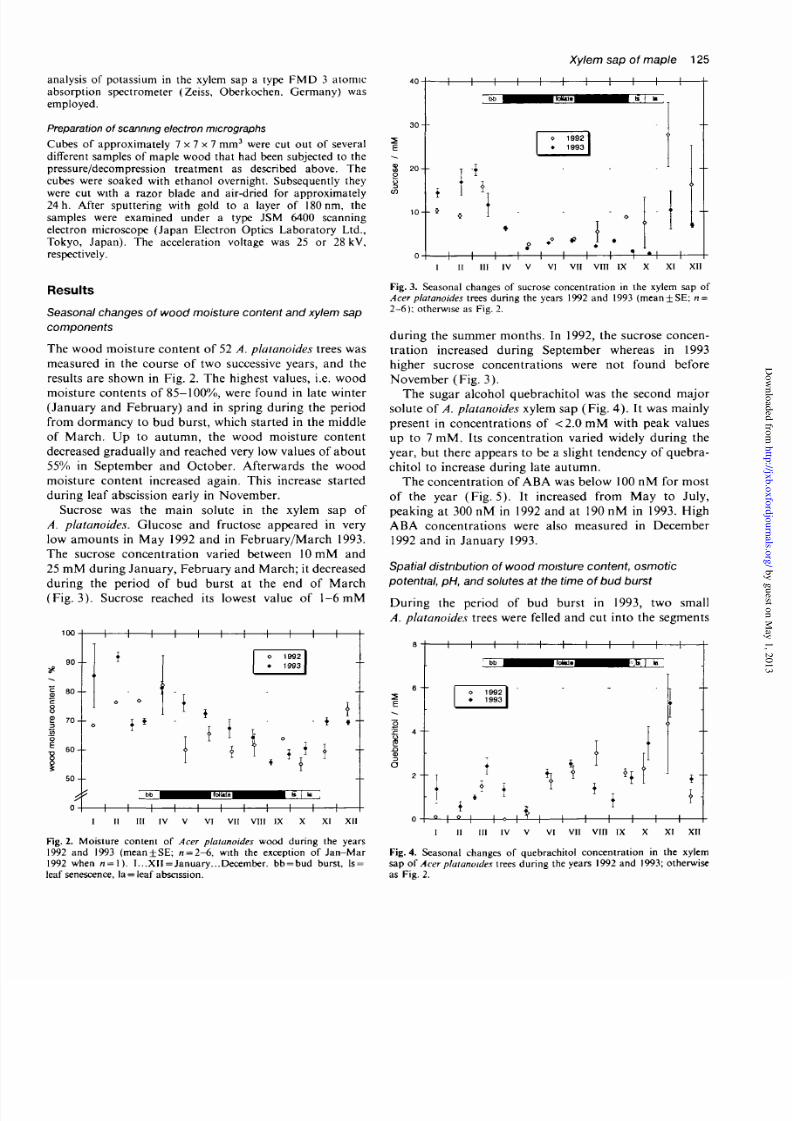
Sucrose was the main solute in the xylem sap of
A. platanoides. Glucose and fructose appeared in very
low amounts in May 1992 and in February/March 1993.
The sucrose conce ntration varied between 10 mM and25 mM during Janu ary, F ebruary and March; i t decreased
during the period of bud burst at the end of March
(Fig . 3) . Sucrose reached its lowest value of 1-6 mM
9 0 - -
g 8 0
S 7 0 - -
6 0 - -
5 0 - -
h
--
o
h
o
—1
o
f
1—
H -
bt 1
— 1 — 1 —
* i
I ^I
—1 1
-1 h-
i
-1 r-
— f-
0
•
Ii
— t -
1992
1993
o
—I
H 1
- i
H 1
I
1
III IV VI VII VTII IX XI XII
Fig. 2. Moisture content of Acer platanoides wood during the years1992 and 1993 (mean±SE; n = 2-6, with the exception of Ja n-M ar1992 when n = \). I . . .XII = January. . .D ecem ber. bb = bud burst , ls =leaf senescence, la = leaf absc ission .
XII
Fig. 3. Seasonal chan ges of sucrose concentration in the xylem sa p ofAcer platanoides trees during the years 1992 and 1993 (m ea n± SE ; n =2- 6) ; otherwise as Fig. 2.
during the summer months. In 1992, the sucrose concen-
tration increased during September whereas in 1993
higher sucrose concentrations were not found beforeNovember (Fig. 3) .
The sugar alcohol quebrachitol was the second major
solute of A. platanoides xylem sap (Fig. 4). It was mainly
present in concentrations of < 2 .0 m M with peak values
up to 7 mM . Its concen tration varied widely durin g the
year, but there appears to be a slight tendency of quebra-
chitol to increase during late autumn.
The concentration of ABA was below 100 nM for most
of the year (Fig. 5). It increased from May to July,
peak ing at 300 nM in 1992 and a t 190 nM in 19 93. High
ABA concentrations were also measured in December
1992 and in January 1993.
Spatial distribution of wood moisture content, osmo tic
potential, pH, and solutes at the time of bud burst
During the period of bud burst in 1993, two small
A. platanoides trees were felled and cut into the segments
I
I 1 1 1 1 1 1 1 1
I V vi VII vin ix XI XII
Fig. 4. Seasonal changes of quebrachitol conc entrat ion in the xylem
sap of Acer platanoides trees during the years 1992 and 1993; otherwise
as Fig. 2.

8/13/2019 The xylem sap of maple [Acer platanoides) trees—sap obtained by a novel method shows changes with season a…
http://slidepdf.com/reader/full/the-xylem-sap-of-maple-acer-platanoides-treessap-obtained-by-a-novel-method 4/11
126 Schill et al.
500-
400-
S 300-c
2< 200-
100-
0 -
1 1
fI
I o
1 1
1 h -
-
1 1 —
— 1 1 h -
f
—I 1 1 1-
—I
m
§
— i —
EEK]
o 1992• 1993
1
H
--
H r-II III IV V I V II V III IX X I XII
Fig. 5. Season al changes of abscisic acid (AB A) con centration in thexylem sap of Acer platanoides trees during the years 1992 and 1993;otherwise as Fig. 2.
shown in Fig. 6. Among the segments of tree A the woodmoisture content varied within a narrow range, i.e. from
61-75%. In tree B one branch showed a particularly highwood moisture content, i.e. more than 80%, whereas allother segments had wood moisture contents similar totree A.
The osmotic potential (77) of xylem sap of segmentsclose to the ground was 0.05 MPa (Fig. 7A). It increasedto approximately 0.2 MPa in the crown segments of the
trees. The pH-value of xylem sap, on the other hand,decreased with the sampling height by approximately1 pH unit (Fig. 7B). Xylem sap of the base segments wasneutral or slightly alkaline, whereas the pH of the crownsegments was 6.1-6.3. The solutes which contributedmost to 77 and pH were sucrose, quebrachitol, potassium,and malate (Fig. 8A -D) . All these substances increased
with height, except for potassium in tree B. The increaseof malate by a factor of 5 may be responsible for thedecrease of pH with height. Citrate was also measured inthe xylem sap (Fig. 8D). The citrate concentration, how-ever, remained constant at < 100 p M along the tree.
The concentration of ABA in the xylem sap at the timeof bud b urst was between 41 nM and 187 nM in tree Aand between 40 nM and 145 nM in tree B (Fig . 9).However, ABA was distributed neither homogeneouslynor polarly. There were areas of high ABA con centration,predominantly at a height of 1.3-1.9 m, next to areas oflow ABA concentration. The xylem sap ABA concentra-tion was low, i.e. 63 8 + 5.3 nM (mean + SE), in the
segments with a wood moisture content above 80%(cf. tree B*). The ABA concentration was higher, i.e.75 8 + 6.2 nM , in parts of the tree with a wood moisturecontent of 70-80%, and highest, i.e. 102.9±9.5nM, inthose segments with a wood moisture content of less than70%. In tree A, where the wood moisture content wasnowhere above 80%, the average ABA concentration was
10 0
not measured
Fig. 6 . Morphology of two A. platanoides trees and spatial distribution of wood moisture content at the time of bud burst. In the scale on the left,
black indicates 100% wood moistu re content and w hite 50% wood moisture content. N umb ers next to the trees state percentage local wood
moisture content .
b y g u e s t onM a y1 ,2 0 1 3
h t t p : / / j x b . oxf or d j o ur n a l s . or g /
D
o wnl o a d e d f r om

8/13/2019 The xylem sap of maple [Acer platanoides) trees—sap obtained by a novel method shows changes with season a…
http://slidepdf.com/reader/full/the-xylem-sap-of-maple-acer-platanoides-treessap-obtained-by-a-novel-method 5/11
Xylem sap of maple 127
m above ground
5 .1 -
4.1
3 . 1 - -
2.1
1 . 1 - - -
0.1
• o oSo
• °CO
C D O
• OCO
» o o
a o
o •
« °
B
o o] •CD Q •
o o
o o
I °jo oj o • i
0.1 0.2 0 3 6
n MPa
6 5p H
7 5
Fig. 7. Spatial distribution of (A) osmotic potentia l n and B) pH in two trees of Acer platanoides Solid circles: tree A, open circles: tree B.
95.2+ 12.2 nM. Precipitation had no clear effect on ABAin the xylem. When the ABA concentration of the xylemsap is plotted against rainfall within the previous 8 d,
only a slight tendency for ABA to increase with soilmoisture may become evident (Fig. 10).
Discussion
The first question to address is the purity of the sap, i.e.
whether living cells, for instance ray parenchyma cells,are damaged by the pressure/decompression treatmentand contaminate the collected xylem sap. The answer isno. Firstly, the low potassium concentration of 3-6 mMindicates that the collected xylem sap is free of cytosoliccontamination. Assuming that the concentration of K
+
in the protoplast is 100 mM and 5 m M in the apop last,and that the proportion of parenchyma cells in Acerwood is 16-18% (Wagenfllhr and Scheiber, 1985), thena mixture of the solutions of both compartments wouldyield a K
+ concentration of 20-23 mM which is not the
case. K+ was used already by Hartung et al. (1988) as a
marker for cytosolic contamination in apoplastic sap
of cotton leaves. In their experiments, pressures of
3.5-4.0 MPa were applied. This did not cause rupture of
living cells and release of K+ into the apoplast. Potassium
concentrations similar to those measured in the presentexperiments have been found in the xylem sap of beech,spruce and eucalypt without pressure/decompressiontreatment (Glavac et al 1989; Kazda and Weilgony,1988; Stark, 1992).
Secondly, other living plants, for instance Nitella sp.
and Elodea canadenis, with cell walls much thinner thanwood parenchyma cells, have been subjected to the
pressure/decompression process. Immediately afterdecompression, the plants were examined with a light
microscope. No damage of the cell walls or other struc-tures was observed. The cells showed lively cytoplasmicstreaming, indicating their vitality and intactness.
Thirdly, scanning electron micrographs of woodsamples after pressure/decompression treatment showthat no damage of the pit membranes between cells of
the wood parenchym a and the xylem vessels had occurred.On the other hand, perforations of the intervessel pits did
occur occasionally (Fig . 11). However, this kind of
damage does not cause contamination of xylem sap withcytosolic solutes.
Sucrose was the main solute of Acer platanoides xylemsap. Its concentration changed markedly with the seasons,rising to a long lasting maximum in autumn and winter.After bud burst and onset of leaf growth sucrose declinedto concentrations of 2-5 mM. Starch degradation duringthe winter dormancy was reported for A . pseudoplatanusby Essiamah and Eschrich (1985). They found that the
cells of the xylem parenchyma were completely filled withstarch at the end of October, but no starch was found inFebruary. This change may be involved in protectionagainst frost damage (Essiamah and Eschrich, 1985;
Cortes and Sinclair, 1985), but further experiments are
needed to verify this hypothesis.Starting in March with bud burst, a loading of the
xylem with sucrose was observed in A . platanoides in thepresent investigation. As the trees were devoid of an
appreciable leaf surface at the time, a transpiration streamas driving force is unlikely. Transpiration from the barkmay account for sap movement only to a minor extent.Therefore, the high sucrose content in the crown mustoriginate from neighbouring parenchyma cells rather thanfrom cells of the basal part of the stem or from the roots.But sucrose might also be transported, as the occurrenceof a high sucrose content in the stem during bud burst
QANSER LIBRARY
MILLERSVILLE UNrVERS TY
MILLERSVILLE.-PA 1755J

8/13/2019 The xylem sap of maple [Acer platanoides) trees—sap obtained by a novel method shows changes with season a…
http://slidepdf.com/reader/full/the-xylem-sap-of-maple-acer-platanoides-treessap-obtained-by-a-novel-method 6/11
12 8 Schill et al.
m above ground
5.H r
4 1 - -
10 20Sucrose / mM
3 6Qtiebrachlto) / mM
m above ground
5.1-
4 .1 -
1 1
2 1 -
1.1-
0 1 -
: c
— _ _ _
o
o
o o
o o oC D
o ooo
o oo •o o •
o •o o
oo0
•
o •• o
o• o
o
11
•
•
r --
.
1
t
7
t
7
'
V
17
7 7
r7
7
:7
W
7
7
7
r •
1
D
• o<
o oo •
o • o
•o
• 0
• o
— t
exo
•oco o
o oo «
1
1
1
•
•
o
T7
o
1— 1—
o
o
o
Citrate (A)Citrate (B)Malate (A)Malate (B)
\-
3 6Potassium / mM
9 0 400 800Citrate, Malate / |iM
1200
Fig. 8. S patial distribution of (A) sucrose , (B) que brach itol, (C) potassium , and (D) citrate and malate in two trees of Acer plalunoides during budburst in March 1993 Solid circles: tree A, open circles, tree B.
of A. pseudoplatanus indicates (Es siam ah, 1982). Hefound sucrose concentrations of approximately 50 mM inbleeding sap of stems at various heights above-ground.This suggests, in contrast to the results of our investi-gation, a loading of sucrose into the xylem sap in basalparts of the trunk or in the roots and its subsequentupward transport. Hydrolysis of sucrose apparently plays
no role in A. platanoides. Only very small am ounts ofglucose and fructose were found in February and March.
Quebrachitol, the second major carbohydrate inA. platanoides xylem sap , showed only m inor seasonalchanges with a small peak in N ovemb er. The physiologicalrole of quebrachitol is still mysterious. Quebrachitol issynthesized from m-inositol via epimerization of bornesi-tol and is found frequently in various tissues of membersof the genus Acer (Hegnauer, 1964; Plouvier, 1990;
Schilling et al., 1972). Perhaps it is only a companion ofsucrose without any special function. However, little isknown about the function of cyclitols and acyclic polyolsin higher plants. They may be cryoprotectants, organicosmolytes or carbon storage compounds (Richter andPopp, 1992). Furthermore, they may serve as mobilereserves, as storage material of reduction equivalents, and
they may be involved in the regulation of coenzymes(Loescher, 1987).
Using our novel technique, it was possible for the firsttime to measure the total spatial distribution of severalsolutes, pH and n in a tree. The measured rise of IT withthe sampling height coincided with an increase of sucrose,quebrachitol, K + , and m alate. Application of van't Hoff'sequation shows, however, that the sum of the measuredsolutes accounts only for n at the stem base. With
b y
g u e s t onM
a y1
,2
0 1
3
h t t p : / / j x b . oxf or d
j o ur n
a l s . or g
/
D o
wnl o a d e d f r om

8/13/2019 The xylem sap of maple [Acer platanoides) trees—sap obtained by a novel method shows changes with season a…
http://slidepdf.com/reader/full/the-xylem-sap-of-maple-acer-platanoides-treessap-obtained-by-a-novel-method 7/11

8/13/2019 The xylem sap of maple [Acer platanoides) trees—sap obtained by a novel method shows changes with season a…
http://slidepdf.com/reader/full/the-xylem-sap-of-maple-acer-platanoides-treessap-obtained-by-a-novel-method 8/11
130 Sd/i/7/etal.
• f
10 u ni
Fig. 11 . Scanning electron m icrographs of maple wood after five pressure/decompression treatments of 4.0 MPa each. ( A) Radial section: vessel-to-ray pits show entirely intact pit membranes (white arrowheads). (B) Tangential section: intervessel pits show some damaged pit membranes(black arrowheads), as well as intact pit membranes (white arrowheads).

8/13/2019 The xylem sap of maple [Acer platanoides) trees—sap obtained by a novel method shows changes with season a…
http://slidepdf.com/reader/full/the-xylem-sap-of-maple-acer-platanoides-treessap-obtained-by-a-novel-method 9/11
may be explained by two mechanisms. Firstly, ABA may
be transported at different rates at different sites from the
xylem vessels to the adjacent parenchyma cells, causing
regions of high and low ABA in the xylem. Secondly,
parenchyma cells at different locations within the stem
Xylem sap of maple 131
and branches may synthesize and release ABA to the
xylem at different rates causing regions of high and low
ABA in the xylem. The latter possibility is supported by
the ABA pattern of tree B. Here ABA is highest in the
branches with the lowest wood moisture content. Low
B
Fig. 12. Principle of the pressure/decomp ression meth od. (A ) Compressed air penetrates the pneum atic wood system (= intercellular spaces). Dueto the short diffusion paths (arro ws), air saturat ion of the xylem sap occurs within a few seconds. (B) Subsequent ra pid decom pression causesrelease of the air, forming small bubbles. (C) Air bubbles expand further and fuse to minimize the phase boundary area. (D) Air-water menisciform as a result of liquid expu lsion. ap = axial parench yma, ft = fibre-tracheid, ip = intervessel pitting, is = intercellular space, rp = ray parenc hym a,v = vessel, vrp = vessel-to-ray pittin g.

8/13/2019 The xylem sap of maple [Acer platanoides) trees—sap obtained by a novel method shows changes with season a…
http://slidepdf.com/reader/full/the-xylem-sap-of-maple-acer-platanoides-treessap-obtained-by-a-novel-method 10/11
132 Sc/7/7/etal.
wood moisture content is a likely signal for stimulatingABA synthesis and release.
In order to describe the xylem sap expulsion a qualitat-ive model concept was developed for hardwood (Fig. 12).It is based on the assumption that upon subjecting thewood to a high pressure atmosphere, compressed airpenetrates into the pneumatic wood system. In a simple
case the pneumatic wood system consists of the air-filledaxial and radial intercellular spaces, while all of the cellcavities are liquid-filled. The air components can, inaccordance with their partial pressures, become dissolvedthrough a large internal area (Fig . 12A). Due to the shortdiffusion path s (micron rang e), a comprehensive sa tura-tion of the xylem sap is possible in just a few seconds. Asubsequent rapid decompression releases the air whichforms small bubbles, displacing the xylem sap (Fig. 12B).The bubbles which are thus created expand further andunite to minimize the phase boundary area (Fig. 12C).Later on air-water menisci form as a result of thecontinuous sap displacement (Fig. 12D). However, some
of the dissolved air may diffuse back to the elements ofthe pneumatic wood system to escape unused. A moredetailed description of the physics of the sap displacementwill soon be given elsewhere.
Xylem sap of twigs and leaves is commonly obtainedby the pressure bomb method. The quantities of sapobtained are small, and the method fails to yield sap ofsturdy branches and trunks. Xylem sap of trunk wood isobtained, therefore, by the water displacement method,which likely causes dilution of the sap (Glavac et a/.,1989). Water-saturated butanol has also been used asa displacing agent in vacuum extraction experiments(Moreno and Garcia-Martinez, 1980). However, butanol
may destroy membranes of wood parenchyma cells andcause contamination of the apoplastic solution with sol-utes of the protoplast.
The novel pressure/decompression method employedwith maple trees provides an uncomplicated and rapidmeans of obtaining xylem sap in large quantities. Nosealing of the wood samples is needed, no special shapeis required. Compressed air, or an inert gas like nitrogen,can be used to displace the xylem sap. The risk of dilutionor contamination is practically zero. When the apparatusis mounted on a mobile stage, xylem sap can be obtainedat the site within a few minutes after cutting a tree.
Acknowledgements
Helpful comments of Professor W Stahl and ProfessorG Jurzitza are acknowledged. We thank D Honig for helpingus with the sugar analysis and B Dierich for analysis of ABA.J Farinha, H Lauer and C Wolff assisted with the preparationof the wood samples and the pressure/decompression experi-ments. M Kurz of the forest district Wildpark, Karlsruhe,authorized and supervised the cutting of maple trees. WH isgrateful to Deutsch e Forschungsgemeinschaft (S FB 251, TP
A3) for financial support and to Professor EW Weiler(Universitat Bochum) for a generous gift of immunochemicals.
References
Cortes PM, Sinclair TR. 1985. The role of osm otic potentia l inspring sap flow of mature sugar maple trees (Acer saccharum
Marsh) . Journal of Experimental Botany 36, 12-24.
Davies WJ, Zhang J. 1991. Ro ot signals and the regulation ofgrowth and development of plants in drying soil. Annual
Review of Plant Physiology and Plant Molecular Biology
42 , 55-76 .Essiamah SK. 1982. Fruhjahrsaktivitaten bei einheimischen
LaubbSumen. PhD thesis, Forstwirtschaftl iche Fakultat ,Georg-August-Universitat , GSttingen.
Essiamah SK, Eschrich W. 1985. Changes of starch content inthe storage tissues of deciduous trees during winter andspring. IAWA Bulletin n s. 6, 97-106.
Glavac V, Koenies H, Jochheim H, Ebben U. 1989. Mineralcomposition of the xylem sap from beech (Fagus sylvatica L.)and variations of its ion concentration along the axialgradient. Angewandte Botanik 63 , 471-86 .
Hartung W, Radin JW, Hendrix DL. 1988. Abscisic acid
movement into the apoplastic solution of water-stressedcotton leaves. Plant Physiology 86 , 908-13 .
Hegnauer R. 1964. Chemotaxonomie der Pflanzen, Vol. 3.,Birkhauser, Basel.
Jackson M B. 1991. Reg ulation of water relation ships in flooded
plants by ABA from leaves, roots and xylem sap. In: DaviesWJ, Jones HG, eds. Abscisic acid physiology and biochemistry.
Oxford: BIOS Scientific Publishers, 217-26.Kazda M, Weilgony P. 1988. Seasonal dynamics of major
cations in xylem sap and needles of Norway spruce (Piceaabies L. Karst.) in stands with different soil solution chemistry.
Plant and So ; /110 , 91-100 .Khalil AAM, Grace J. 1993. Does xylem sap ABA control the
stomatal behaviour of water-stressed sycamore (Acerpseudoplatanus L.) seedlings? Journal of Experimental Botany
4 4 ,1 1 2 7 -3 4 .Loescber WH. 1987. Physiology and m etabolism of sugar
alcohols in higher plants. Physiologia Plantarum 70, 553-7.Mertens RJ, Deus-Neumann B, Weiler EW. 1983. Monoclonal
antibodies for the detection and quantitation of the endogen-ous plant growth regulator, abscisic acid. FEBS Letters
160, 269-72 .Moreno J, Garcia-Martinez JL. 1980. Extraction of tracheal
sap from citrus and analysis of i ts nitrogenous compounds.Physiologia Plantarum 50 , 298-303 .
Pate JS. 1980. Transport and parti t ioning of nitrogenous
solutes. Annual Review of Plant Physiology 31 , 313-40 .Plouvier V. 1990. Alditols et cyclitols: repartition et taxinomie
chez les plantes superieures. Les recherches publiees depuis1962. Bulletin du Mus ee d Histoire naturelle, Pa ris, 4e serie
12, 209-23 .
Reinbothe H, Mothes K. 1962. Urea, ureides. and guanidines inplants. Annual Review of Plant Physiology 13, 129-50.
Richter A, Popp M. 1992. The physiological importance ofaccumulation of cyclitols in Viscum album L. New Phvtologist
121,431-8.Richter A, Thonke B, Popp M. 1990. lr>l-O-methyl-muco-
inositol in Viscum album and members of the Rhizophoraceae.Phytochemistry 29, 1785-6.
Schill V, Korger V, Stahl W. 1992. Mechanical drying of
wood—an alternative to seasoning drying? Proceedings of the3rd IUFRO conference on wood drying, Understanding the
b y
g u e s t onM
a y1
,2
01
3
h t t p : / / jx b . oxf or d
j o urn
al s . or g
/
D o wnl o
a d e dfr om

8/13/2019 The xylem sap of maple [Acer platanoides) trees—sap obtained by a novel method shows changes with season a…
http://slidepdf.com/reader/full/the-xylem-sap-of-maple-acer-platanoides-treessap-obtained-by-a-novel-method 11/11
wood drying process: a synthesis of th eory and practice ,Universitat fiir Bodenkultur, Vienna, Austria, August18-21.
Schilling N, Dittrich P, Kandler O. 1972. The form ation ofL-quebrachitol from D-bornesitol in leaves of Acer pseudo-platanus. Phytochemistry 11 , 1 4 0 1 - 4 .
Stark N. 1992. The effects of water and multi-nutrient stress onxylem sap chemistry, photosynthesis and transpiration of
seedlings of two Eucalypts. Trees
6, 7-12.
Xylem sap of maple 133
WagenfOhr R, Scheiber C. 1985. Holiatlas, 2nd edn. Leipzig:VEB Fachbuchverlag.
Wartinger A, Heilmeier H , Hartung W , Schulze E-D. 1990.Daily and seasonal courses of leaf conductance and abscisicacid in the xylem sap of almond trees [Prunus dulcis (Miller)D.A. Webb]. New Phytologist 116, 581-7 .
Zhang J, Davies WJ. 1987. ABA in roots and leaves of flooded
pea plants. Journal of Experimental Botany 38, 649-59.





























