Embed Size (px)
Citation preview

Brain Research Reviews, 8 (1984) 143- 153 Elsevier
BRR 90015
143
The Superior Colliculus and Visual Neglect in Rat and Hamster. II. Possible Mechanisms
PAUL DEAN and PETER REDGRAVE
Department of Psychology, University of Sheffield, Sheffield SIOZTN (U.K.)
(Accepted July lOth, 1984)
Key words: superior colliculus - retinal ganglion cells - geniculostriate - rat - hamster
CONTENTS
1. Introduction .............................................................................................................................................
2. Unique retinal input to superior colliculus ........................................................................................................ 2.1. Size of collicular and geniculostriate projections .......................................................................................... 2.2. Nature of collicular and geniculostriate projections ......................................................................................
2.2.1. Visual field representation ............................................................................................................ 2.2.2. Retinal ganglion-cell properties in the two projections ..........................................................................
2.3. Unique retinal input to superior colliculus: conclusions .................................................................................
3. Superior colliculus affects geniculostriate function .............................................................................................. 3.1. Superior colliculus as output station for striate cortex .................................................................................... 3.2. Superior colliculus and ‘gating’ of striate function ........................................................................................
4. Mechanisms of neglect: conclusions ................................................................................................................
5. Summary .................................................................................................................................................
Acknowledgements ........................................................................................................................................
References ...................................................................................................................................................
143
144 144 145 146 146 148
148 148 149
150
151
151
151
1. INTRODUCTION
How can the visual neglect produced by collicular
damage in rats and hamsters be explained in terms of
the anatomy and physiology of their visual systems?
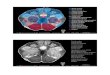
One widely-accepted proposal is illustrated diagra-
matically in Fig. 1 (cf. fig. 39 in ref. 62). This di- . . . . . . .-...
agram emlloclies two important assumpnons. rirsr, Fig. 1. Diagram of possible substrate of response-specific col- that the bulk of visual information passes from the
retina to the geniculostriate system, with only a sub-
set travelling to the superior colliculus. Secondly, the
superior colliculus is specifically responsible for visu-
ally triggered orienting movements, whereas striate
cortex (in conjunction with other areas of visual cor-
licular neglect, as discussed in section 1. The superior colliculus receives a subset of the information that travels to the geniculo- striate system, but is the only visual structure with effective connections to the systems that produce orienting movements. Hence removal of the superior colliculus will impair visually- elicited orienting. LGd, dorsal lateral geniculate nucleus of the thalamus. (Retinal projections to other structures, such as pre- tectal nuclei, omitted. See section 1).
Correspondence: P. Dean, Department of Psychology, University of Sheffield, Sheffield SlO 2TN, U.K.
0165-0173/84/$03.00 0 1984 Elsevier Science Publishers B.V.

144
tex) controls most other responses to visual stimula-
tion.
The plausibility of this model depends on collicular
neglect being a response-dependent deficit. Howev-
er, recent behavioural evidence indicates that the
neglect is not confined to orienting movements, but is
in some instances a failure to detect or attend to cer-
tain kinds of visual stimulusg. The model illustrated in
Fig. 1 therefore requires substantial revision to ac-
commodate this new evidence.
Possible revisions are additionally constrained by
the observation that in rats and hamsters lesions of
striate cortex do not produce striking visual neg-
lect20.37.~.61. This would suggest the following possi-
bilities.
(i) Contrary to the traditional view that all impor-
tant visual information passes to striate cortex, some
visual information travels only to the superior collicu-
lus.
(ii) In rats and hamsters an intact superior collicu-
lus is essential for striate cortex to function complete-
ly normally. Destruction of the superior colliculus
therefore affects both major visual pathways.
These are not exclusive alternatives, and indeed
the proposal that collicular neglect has more than one
behavioural component9 would indicate that both
may be correct. Thus, one of the components, name-
ly a deficit in the detection of certain kinds of visual
stimulus, favours suggestion (i): whereas another be-
havioural component, a disorder of attention in
which other kinds of visual stimulus are detected only
under particular circumstances, is more consistent
with suggestion (ii).
Anatomical and electrophysiological evidence rel-
evant to each of the suggestions is considered in sec-
tions 2 and 3. In these sections only occasional refer-
ence is made to retinal projections to areas other
than the superior colliculus or dorsal lateral genicu-
late nucleus (dLGN); for example to the ventral lat-
eral geniculate nucleus, the pretectal nuclei, and the
accessory optic nuclei. One reason for the omission is
that in rats and hamsters, as in other animals, visual
capability appears to be extremely limited after de-
struction of both the superior colliculus and the corti-
cal target of the dLGN (refs. in ref. 7, 8). Combined
with electrophysiological evidence (refs. in ref. S),
this observation suggests that although the functions
of these other structures are not fully understood,
they are unlikely to play any major role in the per-
formance of the sorts of visual task that are the sub-
ject of this review. One point of terminology - the
pathway from dorsal lateral geniculate nucleus of the
thalamus to cerebral cortex is referred to as the ge-
niculostriate pathway. The term is used because cur-
rent evidence suggests that in rat and hamster either
the dLGN projects solely to striate cortex, or any
projection to extrastriate cortex is very
slight g , 3 6 10,23,49,59,65,
2. UNIQUE RETINAL INPUT TO SUPERIOR COLLICU-
LUS
There are two different kinds of evidence to sug-
gest that in rats and hamsters the retinal projection to
the superior colliculus probably carries visual infor-
mation that does not reach the dLGN. One is evi-
dence relating to size of retinal projections, which in-
dicates that the projection to superior colliculus may
be larger than that to the dLGN (section 2.1). The
second kind of evidence deals more specifically with
the nature of the information carried by the two path-
ways (section 2.2).
2.1. Size of collicular and geniculostriate projections
Linden and Perry33 found that after injection of the
retrograde tracer horseradish peroxidase into the su-
perior colliculus of rats, up to about 47% of cells in
the corresponding area of the ganglion cell layer in
the contralateral retina were labelled. Because in
rats about 50% of the cells in this layer are displaced
amacrine cells which do not send an axon into the op-
tic nerveso, it appears that over 90% of ganglion cells
project to the superior colliculus from the regions of
retina investigated (which excluded the most tempo-
ral region, corresponding to the central part of the
field). The authors give reasons for thinking that a
similar proportion of retinal ganglion cells may pro-
ject to the superior colliculus in hamsters. Prelimi-
nary results suggested that after injections of horse-
radish peroxidase into rat dLGN, about 50% of true
retinal ganglion cells were labelled.
These findings are very much at odds with the as-
sumption that in all mammals the bulk of retinal out-
put travels to the dLGN, with a (small) subset of it di-
verging to the superior colliculus (cf. Fig. 1). Howev-

145
er, it has recently been shown that almost all retinal
ganglion cells project to the superior colliculus in the
rabbit? and there are a number of other findings in
hamster and in rat that are consistent with Linden
and Perry’s results rather than the traditional view.
(1) It has been observed that ‘the greater number
of fibres of the optic tract pass over the outer surface
of the lateral geniculate nucleus to reach the optic
thalamus and colliculi’ (ref. 29, p. 345). Also, there
are about 120,000 fibres in the rat’s optic nerve, but
only about 40,000 cells in the dorsal lateral geniculate
nucleus74, and the volume of the superficial layers of
the superior colliculus (i.e. those layers that receive
the bulk of the retinal input) is about twice that of the
dLGN3s.
(2) Some evidence suggests that the conduction ve-
locities of optic nerve fibres in rat and hamster have a
trimodal distribution. It has been established that the
superior colliculus receives input from all 3 groups, in
both rati7J9,67@ and hamsters6,57.
(3) Toga and Collins73 indirectly measured the dis-
tribution of radioactivity within the brains of albino
rats, following injection of radioactively labelled
amino acid into the eye. They found that the superior
colliculus received 64-76% of the radioactivity
found in the brain contralateral to the injection (in al-
bino rats the visual pathways are almost entirely
crossed). By comparison, the dorsal lateral genicu-
late nucleus received ll-17% (and the pretectal nu-
clei 6-14%). Toga and Collins also measured visu-
ally-produced changes in neural metabolism using
[i4C]2-deoxyglucose. Glucose utilization in response
to light flashes was greater in the superior colliculus
than in dLGN or visual cortex, an effect more pro-
nounced with higher frequencies of stimulation. The
authors interpret these results as suggesting that the
retina projects much more heavily to the superior
colliculus than to other structures, and that this dif-
ference is reflected in the metabolic changes that oc-
cur in response to certain kinds of visual stimulation.
(4) Sefton67 recorded from 34 single cells in rat
dLGN and found that all would respond to stimula-
tion of the superior colliculus. Moreover, the latency
of the response to collicular stimulation correlated
well with the latency of the response to stimulation of
the optic nerve. For this reason and others Sefton
concluded that the fibres innervating the dLGN all
bifurcated to innervate the superior colliculus.
(5) Olavarria and Torrealba44 found that the re-
ceptive fields of cells in lateral prestriate cortex (area
18a of Krieg”) were only slightly affected by ablation
of striate cortex (area 17), and suggested that a tecto-
cortical path was probably responsible for the spar-
ing.
All these results taken together do not completely
rule out the possibility that the dorsal lateral genicu-
late nucleus receives a small input not shared with the
superior colliculus (perhaps from temporal retina):
however, they are all consistent with the possibility
that 40-50% of retinal ganglion cells in the rat pro-
ject to the superior colliculus but not to the dorsal lat-
eral geniculate nucleus. Whether these cells project
to the superior colliculus alone is still unclear. Large
lesions of the superior colliculus in adult rats do not
cause retrograde degeneration of retinal ganglion
cellssiJ3, whereas section of the optic nerve doe+.
Either all retinotectal axons have collaterals, in
which case it is puzzling that the superior colliculus
receives about twice as much label from an injected
retina as all other structures put together, or retino-
tectal axons possess some unknown property that
renders them vulnerable to damage relatively near
the cell body, but not to damage near the axon termi-
nal. At present it is not known which of these alterna-
tives is correct. However, both behavioural and elec-
trophysiological evidence (refs. in e.g. ref. 7) suggest
that if retinotectal axons do bifurcate to reach struc-
tures such as the pretectal nuclei, the information
that reaches those nuclei cannot be used on its own to
solve tests of visual neglect (see section 1).
To summarize, there is some evidence to suggest
that in rats little if any visual information fails to
reach the superior colliculus, and moreover the ret-
inotectal pathway carries information that the genic-
ulostriate pathway does not. Whether the superior
colliculus receives projections that no other retinal
target receives is not known for certain.
2.2. Nature of collicular and geniculostriate projec- tions
The evidence above does not indicate what sort of
information the superior colliculus but not the dorsal
lateral geniculate nucleus receives from the retina.
The evidence considered now relates first to the rep-
resentation of different parts of the visual field in the

146
two structures (section 2.2.1), then secondly to the
properties of the retinal ganglion cells that make up
those representations (section 2.2.2).
2.2.1. Visual field representation
The experiments of Toga and Collins73 also pro-
vided information about visual field representation in
the retinotectal projection. Radioactive label after
eye-injection was not found uniformly across the su-
perficial layers of the superior colliculus: rather,
label was particularly dense in the medial portion of
the superior colliculus, where the upper part of the
visual field is representedi4@. A similar pattern was
revealed in the metabolic response to light flashes,
except that in this case the area of greatest response
extended further over the posterior part of the supe-
rior colliculus, where the far temporal part of the vis-
ual field is mapped.
These results raise the possibility that the informa-
tion received by the superior colliculus that is not
shared by the geniculostriate pathway concerns the
periphery of the visual field. Electrophysiological
studies of superior colliculus and striate cortex in rats
and hamsters provide some support for this idea.
There seems to be general agreement that in the su-
perior colliculus neither magnification factor nor re-
ceptive field size change greatly from central field to
periphery (refs. 5, 14, 68,72: though see ref. 12). In
contrast, there is a clear expansion of the central field
(corresponding to area centralis, which is nasal and
ventral to the projection of the optic disk in the visual
field) in dLGN and striate cortexiJ9Js,71. Thus, Fu-
kuda et al.18 found that about two-thirds of all genicu-
late units sampled had receptive-field centres within
25” of the centre of the area centralis, and Lennie and
Perry31 report a heavy preponderance of receptive
fields in the inferior nasal visual field (cf. ref. 55).
This difference in mapping could occur either be-
cause the central field is more fully represented in the
cortex, or the peripheral field more fully represented
in the superior colliculus, or some combination of
these. It appears that in the central parts of the visual
field, receptive fields of single units are similar in size
in both superior colliculus and striate cortex (about
5-15”: refs. 5, 12, 24, 71, 72, 76). However, in the
periphery of the field cortical cells may have larger
receptive fields than collicular cells, at least in ham-
sters71%72. Indeed in hamsters the extreme upper and
lateral portions of the visual field appear not to be
represented at all in striate cortex7i. This may also be
true for rats, in that recordings from dLGN have re-
vealed very few cells with receptive fields in the up-
per far peripheryss: however, the issue is still to be re-
solved, as other workers have claimed (ref. 39,
p. 230) that ‘The extent and contour of the visual
field represented in LGd is in close agreement with
that found in the superior colliculus’ (by Siminoff et
al.68).
Thus, to summarize, there is both anatomical and
electrophysiological evidence to suggest that the pe-
riphery of the visual field may be in some sense more
fully represented in the superior colliculus of rats and
hamsters, than in the geniculostriate system. Howev-
er, the precise nature of the difference is unclear. In
this context it would be useful to have a study in
which the peripheral field representation in striate
cortex (with superior colliculus removed) was specif-
ically compared with that in the superior colliculus
(with striate cortex removed).
2.2.2. Retinal ganglion-cell properties in the two pro-
jections
The evidence reviewed so far indicates that the su-
perior colliculus receives a heavier retinal projection
than the dorsal lateral geniculate nucleus, particular-
ly for peripheral regions of the visual field. In this
section we ask whether the retinal ganglion cells that
project to the superior colliculus but not to the dorsal
lateral geniculate nucleus have different morpholog-
ical or electrophysiological properties from other ret-
inal ganglion cells. Three studies suggest that they
are ceils with slowly conducting axons and small so-
mas.
(1) Schober and Gruschka65 measured the soma
sizes of retinal ganglion cells that were labelled with
horseradish peroxidase after injection of the enzyme
into either superior colliculus or dorsal lateral genic-
mate. A higher proportion of small retinal ganglion
cells (soma diameter less than about 9 pm) were la-
belled after injections into the superior colliculus
than into the dLGN.
(2) Suminoto et al.69 recorded the responses of sin-
gle cells in either the superior colliculus or the dorsal
lateral geniculate nucleus to stimulation of optic
nerve and optic chiasma. From the response latencies
to stimulation at these two sites, they were able to

147
calculate the conduction velocity of the fibres that produced the response in each cell. The superior col- liculus received a higher proportion than the dLGN nucleus of slowly conducting cells (less than about 4 m/s). The retinal projection to the dLGN could con- tain a lower proportion of slowly conducting cells with small somas than the projection to the supe- rior colliculus for one of two main reasons. Either (a) most cells project to dLGN, but only the small cells also project to the superior colliculus, or (b) most cells project to the superior colliculus, but only the larger cells also project to the dLGN. Linden and Perry’s32 data support the second alternative, as does, indirectly, a comparison of Sumitomo et al.‘s data with those obtained from measuring fibre diam- eters in rat optic nerve**. Ogden and Miller43 found that for rhesus monkey optic nerve, the external di- ameter of the fibre was related to conduction velocity by the equation
Velocity (m/s) = 3.2 x Diameter (inpm)
at least up to diameters of 5 pm. Applying this equa- tion to the data of Sumitomo et al.69 suggests that the distribution of fibre diameters that supply the superi- or colliculus (calculating from velocities, 50% of in- put fibres < 1.22pm) comes much closer to the optic nerve distribution obtained by Hughes (50% c 1.08 pm) than does the distribution of dLGN fibres (50% < 1.92 pm). The discrepancy between collicular and optic nerve distributions may be the result of com- pression of the optic nerve fibres during sectioning**.
(3) Direct support for the view that it is small cells that project to the superior colliculus but not to the dorsal lateral geniculate nucleus is provided by a re- cent study of Ni and DreherQ, who were able to visu- alize retinal ganglion cell morphology after injections of horseradish peroxidase into either target area. On morphological grounds they divided retinal ganglion cells into 5 classes, 4 of which projected to both the dorsal lateral geniculate nucleus and the superior col- liculus. In the remaining class (IIIa) however, many cells were labelled after injection of horseradish per- oxidase into superior colliculus, but few after injec- tion into dLGN. Class IIIA cells had small somas (mean 11.6 pm) and fine axons, as well as fine long primary dendrites with very few branches.
What sort of information is conveyed by these
slowly conducting retinal ganglion cells that project to the superior colliculus but not to the dorsal lateral geniculate nucleus? Unfortunately, there appear to have been few studies of the electrophysiological properties of retinal ganglion cells in the rat or ham- ster, and none relating those properties to conduc- tion velocity. An indirect method is to examine the receptive field properties of cells in the superior colli- culus that receive slowly conducting retinal affer- entsi6Js. The majority of these cells (87%) have been classified as either ON-OFF (responded phasically to both onset and offset of light spot), ON phasic (re- sponded phasically to light onset alone) and move- ment sensitive (responded only to moving light spots). They typically preferred slow movement (C 30 deg/s) , and small stimuli (C 10’) by virtue of a sup- pressive surround which by itself did not give re- sponses. ON-OFF and movement sensitive cells comprise just over 50% of all cell types recorded in the visual layers of the rat superior colliculus com- pared with about 20% in the dorsal lateral geniculate nucleusiQ0. The proportion of ON phasic cells is about the same in the two structures (about 32%); however, in dLGN but not superior colliculus the ma- jority of these cells have concentric off sur- rounds*iJ4Ji, and respond to fast movementls.
Somewhat similar differences between the prop- erties of cells in striate cortex and superior colliculus of hamsters have been described by Tiao and Blake- morerl,? (a) collicular cells frequently preferred small objects, by virtue of their strong suppressive surrounds. In contrast, cortical cells usually respond- ed best to long moving contours; (b) most collicular neurones preferred slower stimulus velocities than did cortical neurones, although there were a few col- licular cells that probably preferred faster movement than that needed by cortical cells; and (c) there was an overall preference for light objects on a dark ground in the cortex, and for dark on light in the colli- culus.
These indirect observations therefore suggest that in rats and hamsters the small, slowly-conducting cells that project to the superior colliculus but not to dLGN give phasic responses to the appearance, dis- appearance and slow movement of small stimuli, and their receptive fields lack concentric surrounds. These properties are consistent with what is known about retinotectal and retinogeniculate projections

148
in cats, the animal that has been most extensively in-
vestigated (e.g. refs. 25,75). In cats, retinal ganglion
cells have been classified into 3 groups (for review
see e.g. ref. 30) on the basis of a number of prop-
erties. Put very briefly: (1) Y cells, with large cell
bodies and fast conducting axons, project both to
dLGN and superior colliculus; (2) X cells, with medi-
um-sized cell bodies and moderately fast conducting
axons, project to the dLGN but hardly at all to the su-
perior colliculus. There is currently disagreement
about the existence of a separate X class in rat~18.21,31,42.48.50,77~ and (3) W cells, with small cell
bodies and slowly’conducting axons, more than half
of which project to the superior colliculus but not to
the dLGN. Electrophysiologically, W-cells appear to
be a rather heterogeneous group, but some of them
respond phasically to the appearance, disappearance
and slow movement of small stimuli, and their recep-
tive fields lack concentric surrounds.
It therefore appears that the retinal ganglion cells
in rat that project to superior colliculus but not dLGN
correspond to a subclass of W-cell in cat (possibly, it
has been suggested, to subclass W2;17). The electro-
physiological properties and dendritic morphology of
W-cells have led to speculations that they function as
primitive ‘feature-detectors’ for local change: that is,
they perform a highly non-linear transformation
upon their inputs to generate information concerning
the existence, whereabouts and crude characteristics
of localized luminance changes relative to back-
ground (e.g. ref. 27,34,53,54).
2.3. Unique retinal input to superior colliculus: con-
clusions
The evidence reviewed here indicates that, con-
trary to traditional assumptions, in rats and probably
hamsters the superior colliculus receives a heavier
retinal projection than the dorsal lateral geniculate
nucleus. In fact it appears that in at least some areas
of the retina very nearly all retinal ganglion cells pro-
ject to the superior colliculus. In contrast, the genicu-
lostriate projection does not include one class of W-
like retinal ganglion cell, and appears to have an im-
poverished representation of the upper and temporal
periphery. It is not at present clear whether this im-
poverished representation arises because the class of
retinal ganglion cell that projects only to the superior
colliculus is relatively more abundant in the periph-
ery, or whether in the periphery additional classes of
ganglion fail to project to the dLGN. Nor is it known
whether there are any retinal ganglion cells, in any
part of the retina, that project to dLGN but not supe-
rior colliculus.
These uncertainties, combined with lack of de-
tailed electrophysiological data, make it impossible
to characterize precisely the kind of information that
the superior colliculus receives but the dLGN does
not. The best guess at present seems to be that the su-
perior colliculus may get unique information about
small, transient stimuli (local ‘events’?) throughout
the visual field, and perhaps about a wider range of
stimuli in the upper and temporal periphery.
3. SUPERIOR COLLICULUS AFFECTS GENICULO-
STRIATE FUNCTION
An intact superior colliculus might be necessary
for the geniculostriate system to function normally. If
so, collicular damage would disturb both major visual
pathways, and affect the processing of visual infor-
mation that reached the dLGN (but not necessarily
the superior colliculus itself). As a potential substrate
for collicular neglect this is therefore complementary
to the one discussed in the previous section.
There are two obvious arrangements whereby ge-
niculostriate function could be partly dependent on
the superior colliculus: (1) the superior colliculus is it-
self an important output station for striate cortex
(Section 3.1); and (2) the superior colliculus controls
some of the information that reaches striate cortex,
or leaves it for other areas of the brain (section 3.2).
3.1. Superior colliculus as output station for striate
cortex
Anatomical evidence indicates that in rat and ham-
ster, as in other animals, there are numerous path-
ways by which the geniculiostriate system could use
the superior colliculus as an output station. For ex-
ample, there are direct projections from both striate
cortex and extrastriate visual areas to the superior
colliculus (e.g. refs. 32, 44), as well as indirect con-
nections via the basal ganglia (e.g. ref. 56).
Direct attempts to evaluate the importance of
these pathways in collicular neglect have compared
the effects of lesions restricted to the superficial lay-

149
ers of the superior colliculus, with lesions that also in-
vade intermediate and deep layers. The rationale for
this procedure is that most cortical afferents arrive at
the intermediate and deep layers, whereas retinal in-
put is restricted to the superficial layers. For exam-
ple, Mort et al.4 and Finlay et al.11, found that
orienting to sunflower seeds by hamsters was com-
pletely abolished only if the deep layers of the super-
ior colliculus were destroyed or undercut. If damage
were restricted to the superficial layers, there was
near total loss of orienting to upper and peripheral
visual fields, but some preserved orienting in lower
and anterior fields. The suggestion was made that fi-
bres from visual cortex to the deep layers of the supe-
rior colliculus might be responsible for the spared
function seen after lesions restricted to the superficial
layers. Related observations that neglect is observed
only if deep collicular layers are damaged have been
made in ratss’, tree shrews4 and gerbil+.
The suggestion that neglect in central areas of the
field requires destruction of corticotectal fibres is
consistent with what is known about the organization
of the retinotectal and geniculostriate pathways (sec-
tion 2), and with the finding of Toga and Collins’s
that electrical stimulation of striate cortex in rat in-
creased metabolic rate much more in that region of
the superior colliculus dealing with the central part of
the visual field than in the peripheral representation.
Taken with Toga and Collins’ other findings (sec-
tion 2.2.1), this result suggests that the superior colli-
culus receives visual information from peripheral
field primarily from the retina, whereas information
from central areas arrives from both retina and visual
cortex.
However, evidence concerning the effects of su-
perficial-layer lesions is at present insufficient to per-
mit any definite inferences about the role of cortico-
tectal projections in collicular neglect.
(a) In some experiments4JrJ6 the lesions of the su-
perficial layers that failed to produce total neglect
were made in neonatal animals. It is clear that in
hamsters and rats damage to the superior colliculus at
this age gives rise to abnormal retinal projections to
surviving structures (e.g. refs. 26, 52, 62, 64). The
role played by these projections in alleviating neglect
is not known.
(b) In cases where superficial lesions have been
made in adult animals, there are other problems of
interpretation. In Mort et al.‘s experimenta, the su-
perior colliculus was damaged by undercuts that were
horizontal in the mediolateral direction. Such cuts
spare deep layers if they are made too superficially.
But because in hamsters the surface of the superior
colliculus slopes steeply downwards from medial to
lateral, an inadequate cut is likely also to spare lateral
portions of the superficial layers. These lateral por-
tions correspond to the inferior part of the visual
field, so that Mort et al.% incomplete neglect may re-
flect at least in part incomplete damage to the super-
ficial layers. There also appears to have been some
sparing in the experiment of Midgley and Teess’: in
the example illustrated (ref. 37, fig. 8) lateral, medi-
al and caudal parts of the superficial layers were not
destroyed. Moreover, although the authors say (p.
326) that lesions restricted to the more superficial
laminae of the superior colliculus did not impair
orienting, their data do not seem entirely to support
this conclusion, in that the extent to which licking was
suppressed by a novel stimulus was similar in animals
with superficial and large lesions in all but two of the
15 examples given.
Thus, it is not clear from current data whether total
destruction of the superficial layers in adult rats and
hamsters does produce a less complete visual neglect
than lesions which in addition invade the deeper lay-
ers. Moreover, even if it were found that superficial
lesions did produce milder effects, there would still
remain problems of interpretation, because although
the bulk of the retinotectal projection terminates in
the superficial layers, there is a detectable retinal in-
put to the deep layers of the superior colliculus in
ratls. The behavioural role of direct or indirect corti-
cal projections to deep layers of superior colliculus
has yet to be properly evaluated. Consequently, it is
at present impossible to say to what extent collicular
neglect arises because the superior colliculus is an
output station for the geniculostriate system.
3.2. Superior colliculus and ‘gating’ of striate func- tion
Damage to the superior colliculus might interfere
with geniculostriate function because it turned off a
tonic or a phasic signal from the superior colliculus
necessary for certain kinds of information to pass to
or from striate cortex - that is, the superior collicu-
lus might act as a ‘gate’ on striate signals.

150
There are a number of collicular projections that
might plausibly carry such a signal. Examples are the
projections to: (i) the dorsal lateral geniculate nucle-
11~34~47.49 (it is also possible that the functions of the
dorsal lateral geniculate nucleus are altered by dam-
age to the superior colliculus, because at least some
retinogeniculate fibres are collaterals of retinotectal
fibres); (ii) extrastriate visual cortex via the lateral
posterior nucleus of the thalamus (e.g. refs. 2, 41,
49, 70); (iii) mesencephalic reticular formationis346;
and (iv) midline thalamic nuclei*.
Unfortunately, little is known concerning the
properties of these pathways. Olavarria and Torreal-
baa found that the receptive fields of cells in lateral
prestriate cortex (area 18a) were relatively slightly
affected by destruction of striate cortex, and they
suggested that the pathway from superior colliculus
to prestriate cortex, via the lateral posterior nucleus,
might be responsible for the spared function. Behav-
ioural evidence is consistent with this suggestion7JJ2.
However, to what extent this pathway is necessary
for normal prestriate function is unknown. There is
some evidences that the superior colliculus is in-
volved in the activation of the cortical EEG to brief
light flashes, but whether this mechanism involves
any gating of striate output is unclear. Finally, Midg-
ley and Tee@ have found that the some of the behav-
ioural symptoms of collicular destruction can be abol-
ished by administration of amphetamine, but the re-
lationship of this finding to putative gating systems
has yet to be established (see also ref. 9).
In summary, the evidence at present is insufficient
either to establish or to exclude gating of striate func-
tion as an important mechanism of collicular neglect
in rats and hamsters.
4. MECHANISMS OF NEGLECT: CONCLUSIONS
It was argued in section 1 that what is perhaps the
most widely-accepted mechanism underlying collic-
ular neglect (illustrated in Fig. 1) required revision in
the light of the findings: (a) the neglect is stimulus-
dependent and not confined to orienting responses;
and (b) it is not found after destruction of striate cor-
tex. Two possible revisions were proposed.
The first was that some visual information reaches
the superior colliculus but not the dLGN. The evi-
dence reviewed in section 2 indicates that this is al-
+?
->?
Fig. 2. Revision of Fig. 1 to illustrate multiple substrates for collicular neglect in rats and hamsters (section 4). (i) The supe- rior colliculus receives a massive retinal projection, which con- veys to it visual information not received by the geniculostriate system. Thus, removal of the superior colliculus will affect de- tection of certain kinds of visual stimulus. (ii) The superior col- liculus may be necessary for the geniculostriate system to func- tion properly, for example by controlling striate output in ex- trastriate cortex, using the projection from superior colliculus to visual cortex via the thalamus. Removal of the superior colli- culus may therefore affect processing of stimuli under partic- ular conditions of cortical activation, producing an ‘attentional’ deficit. (iii) The precise connections of superior colliculus and visual cortex with response systems are unclear. It is thus possi- ble that removal of the superior colliculus produces a response- specific impairment, although not necessarily of the orienting response.
most certainly the case (Fig. 2). One implication of
this conclusion is that after removal of the superior
colliculus there will be some visual information that
the animal is no longer able to process, i.e. there will
be some kinds of visual stimulation that the animal
can no longer detect. Unfortunately, current evi-
dence permits only a rough description of the infor-
mation that reaches the superior colliculus alone: it
appears to be related to localized, transient stimuli,
and to the periphery of the visual field. This is inter-
esting in the light of the behavioural evidence con-
cerning collicular neglects, which suggests that after
collicular damage rats and hamsters have difficulty
detecting small moving stimuli, an impairment that
may be more pronounced in the periphery. Thus, it is
possible that one behavioural component of collicular
neglect, namely an apparent failure to detect certain
classes of visual stimulus, arises because only the su-
perior colliculus (in rat and hamster) receives retinal
information concerning those kinds of stimulus. This
suggestion lends itself readily to experimental test,
with both electrophysiological and psychophysical
techniques.
The second revision proposed in section 1 was that
the superior colliculus might be necessary for normal

1.51
functioning of the geniculostriate system (Fig. 2).
Anatomical techniques have demonstrated numer-
ous pathways that could mediate such a dependence
of geniculostriate function upon the superior collicu- lus, but at present detailed information about the kind
of information that these pathways carry, and their
role in collicular neglect, is not available. One indi-
rect reason for suspecting their importance is the be-
havioural evidence9 that indicates collicular neglect
might have an attentional component. It appears that
collicular animals respond normally to certain stimuli
(possibly large stimuli in the central regions of the
field) only in particular circumstances (e.g. when the
animal is very alert). If this is the case, then informa-
tion about such stimuli must reach visual areas be-
sides the superior colliculus, but clearly those areas
are unable to function completely normally when the
colliculus is damaged. It is thus plausible to suppose
that dependence of the geniculostriate system upon
the superior colliculus underlies the attentional com-
ponent of collicular neglect, while recognizing the ex-
tent of the experimental work required to specify the
nature of that dependence in adequate detail.
Finally, Fig. 2 differs from Fig. 1 in leaving un-
specified the connections of superior colliculus and of
visual cortex with response mechanisms. It is argued
in the next paper that the superior colliculus in rats
and hamsters probably has widespread connections
with a variety of response mechanisms in addition to
those concerned with orienting movements. Howev-
er, it is unclear whether visual cortex also has effec-
tive (even if undirect) connections with the same
mechanisms. Thus, Fig. 2 leaves open the possibili-
ties that removal of the superior colliculus could pro-
duce a response-specific impairment; and that the re-
sponse concerned need not be orienting movements
(cf. Midgley and Tees finding concerning the effects
of collicular lesions on rearings’).
neglect, whereas damage to striate cortex does not,
require explanation in terms of the anatomy and
physiology of visual pathways in these animals. One
proposal is that neglect is produced because the supe-
rior colliculus is the only visual structure directly con-
cerned with the production of orienting movements.
However, recent behavioural evidence indicates that
this mechanism on its own is insufficient: collicular
neglect is not confined to orienting movements, but is
in part an inability to register particular kinds of visu-
al stimulus. Two additional mechanisms are consid-
ered.
(1) The superior colliculus receives visual informa-
tion that the geniculostriate pathway does not. A va-
riety of anatomical and electrophysiological evidence
suggests that this is the case. Although the exact na-
ture of the information is unclear, there is some sug-
gestion that the superior colliculus has a stronger rep-
resentation of the peripheral field than the geniculo-
striate pathway, and may be more concerned with
small transient stimuli throughout the visual field.
(2) An intact superior colliculus is essential for nor-
mal functioning of the geniculostriate system. Ana-
tomical evidence indicates that there are pathways
whereby: (a) visual cortex could use the superior col-
liculus as an output station; and (b) the superior colli-
culus could control signals entering or leaving the
geniculostriate system, although the nature of the in-
formation carried by the pathways is not yet under-
stood.
The precise contribution of these two additional
mechanisms to collicular neglect remains to be deter-
mined. However, it may be conjectured that the first
would underly the deficit in stimulus detection that
appears to be one component of collicular neglect,
whereas the second mechanism might underly an at-
tentional component.
ACKNOWLEDGEMENTS
5. SUMMARY
The observations that removal of the superior col-
liculus in rats and hamsters produces a striking visual
REFERENCES
1 Adams, A. D. and Forrester, J. M., The projections of the rat’s visual field on the cerebral cortex, Quart. J. exp. Phys- iol., 53 (1968) 327-336.
This work was supported by S.E.R.C. Grants
B/24707 and C/46963.
2 Antonetty, C. M., A Study of the Connections of the Superi- or Colliculus, Doctoral Thesis, University of London, 1973.
3 Bigl, V. and Schober, W., Cholinergic transmission in sub- cortical and cortical visual centers of rats: no evidence for the involvement of primary optic system, Exp. Bruin Res.,

152
27 (1977) 211-219. 4 Casagrande, V. A. and Diamond, I. T., Ablation study of
the superior colliculus in the tree shrew (Tupaia glis), 1. camp. Neural., 156 (1974) 207-238.
5 Chalupa, L. M. and Rhoades, R. W., Responses of visual, somatosensory, and auditory neurones in the golden ham- ster’s superior colliculus, J. Physiol. (Lend.), 270 (1977) 595-626.
6 Coleman, J. and Clerici, W. J., Extrastriate projections from thalamus to posterior occipito-temporal cortex in rat, Brain Res, 194 (1980) 205-209.
7 Dean, P., Grating detection and visual acuity after lesions of striate cortex in hooded rats, Exp. Brain Res., 43 (1978) 145-153.
8 Dean, P., Visual pathways and acuity in hooded rats, Be- hav. Brain Res, 3 (1981) 239-271.
9 Dean, P. and Redgrave, P., The superior colliculus and vis- ual neglect in rats and hamsters. I. Behavioural evidence, Brain Res. Rev., 8 (1984) 000-000.
10 Dursteler, M. R., Blakemore, C. and Garey, L. J., Projec- tions to the visual cortex in the golden hamster, J. camp Neural., 183 (1979) 185-204.
11 Finlay, B. L., Marder, K. and Cordon, D., Acquisition of visuomotor behavior after neonatal tectal lesions in the hamster: the role of visual experience, J. camp. physiol. F’sychol., 94 (1980) 506-518.
12 Finlay, B. L., Schneps, S. E., Wilson, K. G. and Schneider, G. E., Topography of visual and somatosensory projections to the superior colliculus of the golden hamster, Brain Res., 142 (1978) 223-235.
13 Foreman, N., Spatial and Attentional Functions of the Mid- brain Visual System, Doctoral Thesis, University of Not- tingham, 1980.
14 Forrester, J. M. and Lal, S. L., The projection of the rat’s visual field upon the superior colhculus, J. Physiol. (Lond.), 189 (1967) 25P-26P.
15 Frankfurter, A., Beckstead, R. M. and Harting, J. K., Ret- inal projections to the deep layers of the superior colliculus in the rat and cat, Anat. Rec., 202 (1982) 60A.
16 Fukuda. Y. and Iwama, K., Visual receptive-field prop-
17
18
19
20
21
erties of single cells in the rat superior colliculus, Jap. J. Physiol., 28 (1978) 385-400. Fukuda, Y., Sumitomo, I. and Sugitani, M., Mode of reti- nal projections to the three visual centers of the rat (dorsal and ventral lateral geniculate nuclei and superior collicu- lus). In Y. Katsuki, R. Norgren and M. Sato (Eds.), Brain Mechanisms of Sensation, John Wiley, New York, 1981, pp. 91-104. Fukuda, Y., Sumitomo, I., Sugitani, M. and Iwama, K., Receptive-field properties of cells in the dorsal part of the albino rat’s lateral geniculate nucleus, Jap. J. Physiol., 29 (1979) 283-307. Fukuda, Y., Suzuki, D. A. and Iwama, K., Characteristics of optic nerve innervation in the rat superior colliculus as revealed by field potential analysis, Jap. J. Physiol., 28 (1978) 347-365. Goodale, M. A. and Murison, R. C. C.. The effects of le- sions of the superior colliculus on locomotor orientation and the orienting reflex in the rat, Brain Res., 88 (1975) 243-261. Hale, P. T., Sefton, A. J. and Dreher, B., A correlation of receptive field properties with conduction velocity of cells in the rat’s retino-geniculo-cortical pathway. Exp. Brain Res., 35 (1979) 425-442.
22
23
24
25
26
27
28
29
Hughes, A., The pigmented-rat optic nerve: fibre count and fibre diameter spectrum, J. camp. Neural., 176 (1977) 263-268. Hughes, H. C., Anatomical and neurobehavioral investiga- tion concerning the thalamo-cortical organization of the rat’s visual system, J. camp. Neurol., 175 (1977) 311-336. Humphrey, N. K., Responses to visual stimuli of units in the superior colliculus of rats and monkeys, Exp. Neural., 20 (1968) 312-340. Ilhng, R.-B. and Wassle, H., The retinal projection to the thalamus in the cat: a quantitative investigation and a com- parison with the retinotectal pathway, J. camp. Neural.,
202 (1981) 265-285. Ingle, D. J., (1982) Organization of visuomotor behaviors in vertebrates. In D. J. Ingle, M. A. Goodale and R. J. W. Mansfield (Eds.), Analysis of Visual Behavior, M.I.T. Press, Cambridge, MA, 1982, pp. 67-109. Koch, C., Poggio, T. and Torre, V., Retinal ganglion cells. A functional interpretation of dendritic morphology, Phil. Trans. B, 298 (1982) 227-264. Krieg, W. J. S., Connections of the cerebral cortex. I. The albino rat. A. Topography of the cortical areas, J. camp. Neurol., 84 (1946) 221-275. Lashley, K. S., The mechanism of vision. VII. The projec- tion of the retina upon the primary optic centers in the rat, J. camp. Neurol., 59 (1934) 341-371.
30 Lennie, P., Parallel visual pathways: a review, Vision Res., 20 (1980) 561-594.
31 Lennie, P. and Perry, V. H., Spatial contrast sensitivity of cells in the lateral geniculate nucleus of the rat, J. Physiol. (Lond.), 315 (1981) 69-79.
32 Lent, R., The organization of subcortical projections of the hamster’s visual cortex, 1. camp. Neural., 206 (1982) 227-242.
33 Linden, P. and Perry, V. H., Massive retinotectal projec- tion in rats, Brain Res., 272 (1983) 145-149
34 MacKay-Sim, A., Sefton, A. J. and Martin, P. R., Subcor- tical projections to lateral geniculate and thalamic reticular nuclei in the hooded rat, J. camp. Neural., 213 (1983) 24-35.
35 Marr, D. C., Vision, W. H. Freeman, San Francisco, CA,
36
37
38
39
1982. Masterton, B., Skeen, L. C. and RoBards, M. J., Origins of anthropoid intelligence. II. Pulvinar-extrastriate system and visual reversal learning, Brain Behav. Evol., 10 (1974) 322-353. Midgley, G. C. and Tees, R. C.. Orienting behaviour by rats with visual cortical and subcortical lesions, Exp. Brain Res., 41(1981) 316-328. Midgley, G. C. and Tees, R. C., Reinstatement of orienting behavior in rats with superior colliculus lesions by n-am- phetamine, Sot. Neurosci. Abstr., 8 (1982) 894. Montero, V. M., Brugge. J. F. and Beitel, R. E., Relation of the visual beld to the lateral gemculate body of the albino rat. J. Neurophvsiol.. 31 (1968) 221-236.
40 Mort, E., Cairns, S., He&h, H. and Finlay, B., The role of the superior colliculus in visually guided locomotion and visual orienting in the hamster, Physiol. Psychol., 8 (1980) 20-28.
41 Nagel, S. M. and Butter. C. M., Tectal and cortical affer- ents to nucleus lateralis posterior in the rat, Sot. Neurosci. Abstr., 5 (1979) 799.
42 Ni, S. Y. and Dreher, B., Morphology of rat retinal gangli- on cells projecting to the thalamus and midbrain, Proc. aus-

43
44
45
46
47
tral. Physiol. Pharmacol. Sot., 12 (1981) 97P. Ogden, T. E. and Miller, R. F., Studies of the optic nerve of the Rhesus monkey: nerve fibre spectrum and physiological propetties, Vision Rex, 6 (1966) 485-506. Olavarria, J. and Torrealba, F., The effect of acute lesions of the striate cortex on the retinotopic organization of the lateral peristriate cortex in the rat, Brain Res., 151 (1978) 386-391. Olavarria, J. and Van Sluyters, C., The projection from striate and extrastriate cortical areas to superior colliculus in the rat, Brain Res., 242 (1982) 332-336. Otani, K., Kimura, M. and Yamada, J., The tectofugal pro- jections in the rat and cat, Proc. 10th int. Congr. Anat., 1975, p. 183. Pasquier, D. A. and Villar, M. J., Subcortical projections to the lateral geniculate body in the rat, Exp. Brain Res., 48 (1982) 409-419.
48 Perry, V. H., The ganglion cell layer of the retina of the rat: a Golgi study, Proc. roy. Sot., 204, B (1979) 363-375.
49 Perry, V. H., A tectocortical visual pathway in the rat, Neu- rosci., 5 (1980) 915-927.
50 Perry, V. H., Evidence for an amacrine cell system in the ganglion cell layer of the rat retina, Neurosci., 6 (1981) 931-934.
51 Perry, V. H. and Cowey, A., Changes in the retino-fugal pathways following cortical and tectal lesions in neonatal and adult rats, Exp. Brain Res., 35 (1979) 97-108.
52 Perry, V. H. and Cowey, A., A sensitive period for gangli- on cell degeneration and the formation of aberrant retino- fugal connections following tectal lesions in rats, Neurosci- ence, 7 (1982) 583-594.
53 Pinter, R. B., Product term non-linear lateral inhibition en- hances visual selectivity for small objects or edges, J. theor. Biol., 100 (1983) 525-531.
54 Pinter, R. B. and Harris, L. R., Temporal and spatial re- sponse characteristics of the cat superior colliculus, Brain Res., 207 (1981) 73-94.
55 Reese, B. E. and Jeffery, G., Crossed and uncrossed visual topography in dorsal lateral geniculate nucleus of the pig- mented rat, J. Neurophysiol., 49 (1983) 877-885.
56 Rhoades, R. W. and Chalupa, L. M., Conduction velocity distribution of the retinocollicular pathway in the golden hamster, Brain Res., 159 (1978) 396-401.
57 Rhoades, R. W. and Chalupa, L. M., Conduction velocity distribution of the retinal input to the hamster’s superior colliculus and a correlation with receptive field characteris- tics, J. camp. Neural., 184 (1979) 243-264.
58 Rhoades, R. W., Kuo, D. C., Polcer, J. D., Fish, S. E. and Voneida, T. J., Indirect visual cortical input to the deep lay- ers of the hamster’s superior colliculus via the basal ganglia, J. camp. Neural., 208 (1982) 239-254.
59 Ribak, C. E. and Peters, A., An autoradiographic study of the projections from the lateral geniculate body of the rat, Brain Res., 92 (1975) 341-368.
60 Schneider, G. E., Contrasting visuomotor functions of tec- turn and cortex in the golden hamster, Psychol. Forsch., 31
153
(1967) 52-62. 61 Schneider, G. E., Two visual systems, Science, 163 (1969)
895-902. 62 Schneider, G. E., Mechanisms of functional recovery fol-
lowing lesions of visual cortex or superior colliculus in neo- nate and adult hamsters, Brain Behav. Evol., 3 (1970) 295-323.
63 Schneider, G. E., Two visuomotor systems in the Syrian hamster. In D. J. Ingle and J. M. Sprague (Eds.), Sensori- motor Functions of Midbrain Tectum, Neurosci. Res. Progr. Bull., 13 (1975) 255-257.
64 Schneider, G. E., Is it really better to have your brain lesion early? A revision of the ‘Kennard Principle’, Neuropsycho- logia, 17 (1979) 557-583.
65 Schober, W. and Gruschka, H., Zur Projektion der einzel- nen Ganglienzellklassen der Retina der Albinoratte. Eine Studie mit Meerrettich-Peroxidase, Z. Mikrosk.-anat. Forsch. Leipzig, 92 (1978) 283-297.
66 Schober, W. and Winkelmann, E., Die geniculo-kortikale Projecktion bei Albinoratten, J. Hirnforsch., 18 (1977) l-20.
67 Sefton, A. J., The innervation of the lateral geniculate nu- cleus and anterior colliculus in the rat, Vision Res., 8 (1968) 867-881.
68 Siminoff, R., Schwassmann, H. 0. and Kruger, L., An electrophysiological study of the visual projection to the su- perior colliculus of the rat, J. camp. Neural., 127 (1966) 435-444.
69 Sumitomo, I., Ide, K., Iwama, K. and Arikuni, T., Conduc- tion velocity of optic nerve fibers innervating lateral genic- ulate body and superior colliculus in the rat, Exp. Neural., 25 (1969) 378-392.
70 Takahashi, T., Nuclear organization of the rat’s lateral thalamus, Sot. Neurosci. A&r., 8 (1982) 643.
71 Tiao, Y.-C. and Blakemore, C., Functional organization in the visual cortex of the golden hamster, J. camp. Neurol., 168 (1976) 459-482.
72 Tiao, Y.-C. and Blakemore, C., Functional organization in the superior colliculus of the golden hamster, J. camp. Neu- rol., 168 (1976) 483-504.
73 Toga, A. W. and Collins, R. C., Metabolic response of op- tic centers to visual stimuli in the albino rat: anatomical and physiological considerations, J. camp. Neural., 199 (1981) 443-464.
74 Vaney, D. I., Peichl, L., Wassle, H. and Illing, R.-B., Al- most all ganglion cells in the rabbit retina project to the su- perior colliculus, Brain Res, 212 (1981) 447-453.
75 Wassle, H. and Illing, R.-B., The retinal projection to the superior colliculus in the cat: a quantitative study with HRP, J. camp. Neural., 190 (1980) 333-356.
76 Wiesenfeld, Z. and Kornel, E. E., Receptive fields of sin- gle cells in the visual cortex of the hooded rat, Brain Res., 94 (1975) 401-412.
77 Winkelmann, E. and Brauer, K., Some aspects of neuronal organization in rat’s visual structures, Vision Res., 21 (1981) 1681-1683.