Embed Size (px)
Citation preview

See discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/332589621
The skeleton and the mineralized tissues of the living coelacanths
Article · March 2019
CITATIONS
4READS
226
3 authors:
Some of the authors of this publication are also working on these related projects:
Revisão do Ramo Gondwânico da família Mawsoniidae (Sarcopterygii: Actinistia: Coelacanthiformes) View project
Morpho-functionnal study of the paired fins of extant and extincts coelacanths - Considerations on the terrestrialisation process of vertebrates View project
François J. Meunier
116 PUBLICATIONS 2,475 CITATIONS
SEE PROFILE
Camila Cupello
Rio de Janeiro State University
15 PUBLICATIONS 49 CITATIONS
SEE PROFILE
Gaël Clément
Muséum National d'Histoire Naturelle
103 PUBLICATIONS 912 CITATIONS
SEE PROFILE
All content following this page was uploaded by Camila Cupello on 23 April 2019.
The user has requested enhancement of the downloaded file.

type according to the definition of MEUNIER et al. (2013). This organization is less complex than the three other plicidentine types (polyplocodont, eusthenodont and dendrodont), which are described in most of extinct and extant Sarcopterygii (SCHULTZE, 1969, 1970).The bony plates of the lung
The lung of Latimeria is reduced to a short oesophagus diverticulum (CUPELLO et al., 2015) surrounded by scattered small and thin ovoid plates (Fig. 9A, B). These plates cover the surface of the vestigial lung, and are composed of a fibrous matrix with fibrocytes: it is a cellular bony tissue (CUPELLO et al., 2017). Each plate is enclosed in a membranous bag. These plates look fragile in their middle plane since a central weakness zone opened on the microtome knife, creating an artefactual lumen (Fig. 9C) (CUPELLO et al., 2017). The fibres are collagenous and organized in superposed layers. The cells are true osteocytes (Fig. 9E) with a more or less star-shaped outline, and send cytoplasmic extensions in the thickness of the plate (Fig. 9D, E). Rare lining cells (osteoblasts) are
present on the surface of the plates, and conjunctive fibres (SHARPEY’s fibres) penetrate within the plate (Fig. 9C). The extracellular matrix shows various staining intensities with concentric and parallel (Fig. 9D) or globular shaped lines. These lines are considered to be Liesegang lines that characterize an active spheritic mineralizing process. The lung plates of Latimeria are thus true bony plates, and are homologous with the large bony plates known in the abdominal cavity of fossil actinistians (BRITO et al., 2010; CUPELLO et al., 2017). The calcified walls that surrounded the lung of the Cretaceous coelacanth Axelrodichthys are made of large osseous plates of various thickness (BRITO et al., 2010), as those of other fossil coelacanths (CLEMENT, 1999, 2005). The bony tissue of these plates is a vascularized cellular bone with more or less large vascular cavities and some internal remodelling (BRITO et al., 2010). Moreover the mineralization of the lung ossified plates in Axelrodichthys is spheritic as in Latimeria’s plates (CUPELLO et al., 2017). So there is a continuity of the histological structure of the lung plates in the coelacanths during their
evolution, but with an important reduction of the bony plates in Latimeria linked to the vestigial state of its lung (CUPELLO et al., 2015, 2019).
CONCLUSION
This overview of eighty years (1938–2018) of histological work on Latimeria skeletal tissues allows some anatomical and evolutionary considerations. It can be enlightened a drastic reduction of endochondral ossification during the long evolutionary history of coelacanths. The persistence of large volume of cartilage in the endoskeleton at adult stage can be compared to the “little bone and considerable cartilage” that characterize the skeleton of a number of demersal notothenioid telostean fishes (EASTMAN et al., 2014).
A processus of spheritic mineralization has been recently highlighted in various skeletal elements of Latimeria, by the presence of globular dentine in teeth, in odontodes of the tegumentary skeleton (scales, fin rays), in scales at the interface between the external layer and the basal plate, as well as in lung bony plates. The spheritic mineralization (i.e., a radiating arrangement of hydroxyapatite crystals and of the organic matrix) in vertebrate skeletal tissues is considered as a precursor of inotropic mineralization (specific interactions between collagen fibrils and the mineral phase) that possibly represents a derived evolutionary stage of calcification mechanisms (ØRVIG, 1951, 1968; FRANCILLON-VIEILLOT et al., 1990; ZYLBERBERG et al., 1992). The simultaneous presence of both mineralization processes (spheritic and inotropic) in the mineralized tissues of Latimeria and in other osteichthyans such as teleosteans (ZYLBERBERG and MEUNIER, 2008) is however questionable. The mineralized spherules present at the limit between the external layer and the basal plate of Latimeria scales are considered to be the product of an inotropic mineralization (MEUNIER and ZYLBERBERG, 1999). It thus appears that the state of the mineralized spherules observed in the various skeletal tissues of Latimeria, including the lung bony plates, must be studied with adapted ultrastructural techniques in order to test their true origin: spheritic or inotropic mineralization?
ACKNOWLEDGEMENTS
This contribution is linked to an international roundtable on coelacanths evolution held at the Kitakyushu Museum and at the Aquamarine Fukushima in August 2017. We would like to thank the anonymous reviewers for their constructive comments and valuable suggestions. C.C. was partially financed by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brasil (CAPES) – Finance Code 001 (Programa Nacional de Pós Doutorado-PNPD).
INTRODUCTION
The first study of fossil coelacanths was done by AGASSIZ (1833–44) and was followed by a lot of papers with the discovery of new fossils (see reviews of JANVIER, 1996 and FOREY, 1998). Fossil coelacanths constitute an important monophyletic group composed of more than forty genera, eighty species, and belong to the Sarcopterygii (ANDREWS, 1973; JANVIER, 1996; FOREY, 1998; UYENO and YABUMOTO, 2007; and many others). For a century coelacanths were considered extinct since the end of the Cretaceous time (Fig. 1).
With the discovery of a living coelacanth in 1938 (SMITH, 1939) the scientific community was soon highly interested to compare the incomplete fossil sarcopterygian fishes dataset to the recent anatomical and biological organization of Latimeria chalumnae. Many authors (see THOMSON, 1969; MILLOT and ANTHONY, 1958; MILLOT et al., 1978 FOREY, 1984, 1998; JANVIER, 1996) focused their work on Latimeria as a key-taxon to test or propose various hypotheses on the origin and evolutionary history of the early tetrapods and sarcopterygii in general, and to infer their possible biological characteristics. The fossil sarcopterygian fishes are known by their fossilized mineralized tissues, essentially their bones, scales and teeth. Together with the extant lungfishes, especially Neoceratodus, Latimeria offered a unique access to the skeleton as a whole: with both mineralized and unmineralized skeletal tissues. Here, we aim to present an overview of the last eighty years (1938–2018) of histological studies of Latimeria’s skeleton.
Since the discovery of the Raja Laut (“king of the sea” ), off Sulawesi Island in Indonesia (ERDMANN et al., 1998, 1999), the genus Latimeria comprises two species: L. chalumnae SMITH 1939 and L. menadoensis POUYAUD et al., 1999. However our knowledge of the anatomy and biology of the living coelacanths is so far essentially based on L. chalumnae.
THE LATIMERIA SKELETON
The first anatomical observations on the extant coelacanth were done by SMITH (1940). The anatomy of the Latimeria skeleton has been later described in detail by MILLOT and ANTHONY (1958). The living coelacanth shows a general organization similar to that of the other actinistian fishes (THOMSON, 1969; JANVIER, 1996; FOREY, 1998). The skeleton can be divided in various parts (Fig. 2): skull, axial skeleton, paired and unpaired fins, tegumentary skeleton (scales). The skull, axial skeleton and fins are constituted of endoskeletal elements that are overlain by exoskeletal elements. In the extant Latimeria the endoskeleton of the skull shows an important regression of the bones, especial ly in the neurocranium, which are replaced by cartilaginous tissues, as in its Mesozoic fossil actinistian relatives (ROMER, 1937, 1942; MILLOT and ANTHONY, 1958; BJERRING, 1973; FOREY, 1998). In the same way, the endoskeleton of paired and unpaired fins is essentially constituted of four axial cartilaginous elements (Fig. 2; MILLOT and ANTHONY, 1958), with pre and post-axial
elements and eventually minute superficial endochondral ossification (CASTANET et al., 1975). The axial skeleton is composed of the notochord which is coated by an unmineralized fibrillary sheath and is totally deprived of well developed vertebrae due to the lack of ossified centra, although some discrete neural and haemal arches develop in the anterior part of the notochord (MILLOT and ANTHONY, 1958). The anterior neural and haemal spines are relatively short but they progressively increase in length posteriorly. Importantly neural and haemal spines are composed of perichondral bone surrounding a cartilaginous core (Fig. 2).
The scales belong to the exoskeleton. In both extant coelacanth species they are of elasmoid type, composed of an upper external layer also called “external ornamented layer”, and of a lower thicker layer, called the basal plate, which is stratified and almost totally unmineralized (Fig. 3). The upper external layer is ornamented with radial crests. In the posterior area of the scale these radial reliefs are overlain by numerous denticulations, the odontodes, which can be superposed (Fig. 3). The anterior area of the external layer, which is covered by the anterior scales, shows crests with irregularities that have been interpreted as growth marks and tentatively used for ageing coelacanths (HUREAU and OZOUF, 1977; MILLER, 1979).
The tooth plates of the buccal cavity support series of teeth of various sizes (Fig. 4). They range in three morphotypes: the fangs (7–10 mm in height), middle-sized teeth (3–4 mm in height) and rounded tubercles (MILLOT and ANTHONY, 1958). The teeth of the two first categories are conical and sharp whereas those of the third category have an obtuse tip (Fig. 5). Fangs of Latimeria are inserted into a socket (MILLOT and ANTHONY, 1958; HOBDELL and MILER, 1969; CASTANET et al., 1975) and they have a smooth external surface (MILLOT et al., 1978; MEUNIER et al., 2015).
A particularity of the exoskeleton of Latimeria is the abundance of odontodes at the surface of various skull bones
Bull. Kitakyushu Mus. Nat. Hist. Hum. Hist., Ser. A, 17: 37–48, March 31, 2019
The skeleton and the mineralized tissues of the living coelacanths
ABSTRACT − The present overview of the histological studies on Latimeria mineralized tissues, since the discovery of the first living coelacanth in 1938, allows some anatomical and evolutionary considerations. It enlightens: i) a drastic reduction of cartilage to bone transformation processes during evolution; ii) the persistence of large volume of cartilage in the endoskeleton at adult stage; iii) the bony nature of the plates that surround the lung diverticle; iv) the presence of a developed process of spheritic mineralization in various skeletal organs: in teeth, in odontodes of the tegumentary skeleton (scales, fin rays), in scales at the interface between the external layer and the basal plate, as well as in lung bony plates.
François J. MEUNIER1, Camila CUPELLO2 and Gaël CLÉMENT3
1UMR 7208 (MNHN - Sorbonne Universités - Univ. Caen Normandie - Univ. Antilles – CNRS - IRD), BOREA, Département Adaptations du Vivant, Muséum national d'Histoire naturelle, C.P. 026, 43 rue Cuvier,
75231 Paris cedex 05, France. E-mail: [email protected] de Zoologia, Universidade do Estado do Rio de Janeiro, R. São Francisco Xavier,
524-Maracanã, Rio de Janeiro 20550–900, Brazil. E-mail: [email protected] 7207 (MNHN–Sorbonne Université–CNRS), Centre de Recherche sur la Paléobiodiversité et les
Paléoenvironnements, Département Origines & Evolution, Muséum national d’Histoire naturelle, CP38, 57 rue Cuvier, 75231 Paris cedex 05, France. E-mail: [email protected]
(Received August 12, 2018; accepted October 22, 2018)
(MILLOT and ANTHONY, 1958; BERNHAUSER, 1961), on the fin rays (CASTANET et al., 1975) and on the posterior (free) area of scales (Fig. 3) (MILLOT and ANTHONY, 1958; CASTANET et al., 1975; HADIATY and RACHNATIKA, 2003; MEUNIER et al., 2008).
Among the various components of the whole skeleton, and apart the SMITH’s pioneer work (SMITH, 1940), numerous studies have been carried out on the histological organization of scales (ROUX, 1942; SMITH et al., 1972; CASTANET et al., 1975; GIRAUD et al., 1978a; MILLER, 1979; SMITH, 1979; MEUNIER, 1980; MEUNIER and ZYLBERBERG, 1999; HADIATY and RACHMATIKA, 2003; MEUNIER et al., 2008) and teeth (MILLER and HOBDELL, 1968; GRADY, 1970; HOBDELL and MILLER, 1969; CASTANET et al., 1975; SHELLIS and POOLE, 1978;
SMITH, 1978; SASAGAWA et al., 1984; MEUNIER et al., 2015). Contrary to the superficial skeleton, there are clearly less histological studies of the skeletal bony elements (FRANCILLON et al., 1975). Meanwhile, in addition to the usual skeletal elements, specific bony tissues have been described in fossil (BRITO et al., 2010) and extant coelacanths (CUPELLO et al., 2015, 2017), such as the mineralized plates surrounding the lung. These bony plates in actinistians do not belong to the skeleton sensu stricto. They are specific bony specializations in relation to the lung (see below), as is the “rocker bone” in relation to the gas bladder of some ophidiiform teleosts (PARMENTIER et al., 2008). Despite the name, the rocker bone is not true bony tissue (PARMENTIER et al., 2008), contrary to
the bony plates of fossil and extant coelacanths (BRITO et al., 2010; CUPELLO et al., 2017).
ADULT LATIMERIA SKELETAL TISSUES
Bony tissuesCortical areas of the dermal skeleton are constituted of
primary periosteal bony t issue. I ts mineralization is heterogeneous and generally marked by series of growth lines, probably due to alternative physiological cycles linked to seasonal variations (Fig. 6). The cortical primary bone shows vascular canals and numerous SHARPEY’s fibres (Fig. 6B). The centre of these bones is frequently made of a spongiosa with vascular cavities surrounded by secondary bony deposits that result from remodelling processes (Fig. 6). This secondary bone is separated from the primary one by reversal cementing lines that are hypermineralized.
Bone remodelling in adult specimens can be more or less developed according to a bone and within a given bone. Primary bone is destroyed by osteoclasts and the deposition process of the secondary bone results from the activity of osteoblasts that have replaced the osteoclasts. The secondary bone does generally not replaced the whole primary bone areas, which can still be recognizable by the presence of
SHARPEY’s fibres and/or growth marks (Fig. 6).The dermal fin rays that sustain paired and unpaired fins
in Latimeria are true lepidotrichia (CASTANET et al., 1975). They are made of two parallel opposite gutter-shaped hemirays, each of which being a series of hemisegments articulated by a collagenous ligament. Each hemisegment is made of a mineralized collagenous fibrillary matrix with embedded osteocytes. The hemisegments are sometimes fused at the basal part of the rays due to centrifugal deposition of bony laminae around the primary articulation, and eventually to the mineralization of the ligament (see figs. 19, 20 in CASTANET et al., 1975). Odontodes observed on the external convex surface of the rays are similar to those present on the dermal bones of the skull and scales. The distal extremity of the lepidotrichia is overlapped by actinotrichia (GÉRAUDIE and MEUNIER, 1980) that are long tapered rods of elastoidin , a fibrous protein of collagenous nature (FAURÉ-FREMIET, 1936; GARRAULT, 1936). The presence of lepidotrichia with distal actinotrichia in Latimeria, as in the fins of Actinopterygii, is
regarded as a plesiomorphic character for Osteichthyes (GÉRAUDIE and MEUNIER, 1980, 1984).
The upper layer of the Latimeria scale is relatively thin and it is constituted of bony tissue with embedded osteocytes, whereas the basal plate is much thicker and composed of numerous strata made of thick collagenous fibres (Fig. 7). Between two collagenous layers there are star-shaped cells (SMITH et al., 1972, pl. VI; CASTANET et al., 1975, fig. 12), the elasmocytes, which cytoplasmic processes insert between the collagenous fibres. These fibres are set in a very specific network. In each layer, the fibres are parallel to each other and the direction of the fibres change from a layer to another one. This regular organization results in a spatial arrangement termed “twisted plywood” (GIRAUD et al., 1978a,b). The rotation of fibres direction from one layer to the next has a mean angle of 27° (GIRAUD et al., 1978a). This organization is found in each scale of L. chalumnae. In L. menadoensis, the basal plate of the scales is also a twisted plywood but its rotation angle seems to be slightly less regular (MEUNIER et al.,
2008). The basal plate in both species is unmineralized (Fig. 7) excepted at the contact between the superficial layer and the basal plate, where spheritic mineralized granules are seen in the very first layers of the basal plate (MEUNIER, 1980; MEUNIER and ZYLBERBERG, 1999; MEUNIER et al., 2008). There is no pore-canal system so Latimeria’s scales are not cosmoid, contrary to most of palaeozoic sarcopterygian fishes. Due to the presence of an unmineralized stratified basal plate, the scales of Latimeria are defined as elasmoid-like scales. However the plywood-like organization is considered to be homologous with the bony basal plate of cosmoid scales of extinct sarcopterygian fishes (MEUNIER, 1980; SIRE and HUYSSEUNE, 2003; MONDEJAR, 2018; SCHULTZE, 2018).Cartilages
The cartilaginous tissues in Latimeria are characterized by long chondrocytes (Fig. 8) contrary to those of teleostean fishes that are relatively spherical in shape (MEUNIER, 1979; ZYLBERBERG and MEUNIER, 2008). Cartilage tissues can show an endochondral ossification process. This phenomenon has been studied respectively on the urohyal (PEGUETA, 1968), the MECKEL’s cartilage and the proximal piece of the pectoral girdle (FRANCILLON et al., 1975). The endochondral ossification is relatively limited when it occurs and the volume of enchondral bone remains reduced to thin bony layers (Fig. 8). The cartilage is destroyed by chondroclasts in areas where endochondral ossification occurs. Serial chondrocytes, known at the origin of the calcified cartilage, seem to be lacking in the endochondral ossification process in Latimeria (FRANCILLON et al., 1975). This constitutes a true difference with teleostean endochondral ossifications (MEUNIER, 1979; ZYLBERBERG and MEUNIER, 2008). Tooth tissues
The three tooth morphotypes described in Latimeria are fangs (7–10 mm in height), middle-sized teeth (3–4 mm in height) and rounded tubercles (MILLOT and ANTHONY, 1958). These teeth and tubercles are constituted of a cone of orthodentine set around a large pulp cavity and overlain by an external hypermineralized layer considered as true enamel (GRADY, 1970; CASTANET et al., 1975; SHELLIS and POOLE, 1978; SMITH, 1978; SASAGAWA et al., 1984). Various studies have revealed the presence of globular dentine (Fig. 5) at the periphery of the first third of the tooth (MILLER and HOBDELL, 1968; HOBDELL and MILLER, 1969; SHELLIS and POOLE, 1978; SASAGAWA et al., 1984). This typical histological organization characterizes the various teeth of the bucco-pharyngeal cavity as well as the various odontodes of the dermal bony plates, the fin rays and the scales (CASTANET et al., 1975).
In the fang the inner wall of the pulp cavity displays a series of plies (Fig. 5; MEUNIER et al., 2015). These plies start at the base of the tooth and reach at least the first third to the half of the tooth towards its tip (Fig. 5). Such plies are also present but less developed in the small caniniform teeth (MEUNIER et al., 2015). These dentine plies in the pulp cavity characterizes a plicidentine organization of orthodentine type. These primary plies correspond to a simplexodont plicidentine
REFERENCES
AGASSIZ, L. 1833–44. Recherches sur les Poissons fossiles. Imprimerie Petitpierre, Neuchâtel. [5 volumes and atlas.]
ANDREWS, S. M. 1973. Interrelationships of crossopterygians. In: GREENWOOD, P. H., MILES, R. S. and PATTERSON, C. (eds.), Interrelationships of Fishes, Acad. Press, London, pp. 137–177.
BERNHAUSER, A. 1961. Zur Knochen- und Zah, histology von Latimeria chalumnae Smith und einiger Fossilformen. Sber. öst. Akad. Wiss., 170: 119–137.
BJERRING, H. C. 1973. Relationships of coelacanthiforms. In: GREENWOOD P. H., MILES R. S. and PATTERSON C. (eds.), Interrelationships of Fishes, Acad. Press, London, pp. 179–205.
BRITO, P. M., MEUNIER, F. J., CLÉMENT, G. and GEFFARD-KURIYAMA, D. 2010. The histological structure of the calcified lung of the fossil coelacanth Axelrodichthys araripensis (Actinistia: Mawsoniidae). Palaeontology, 53: 1281–1290.
CASANE, D. and LAURENTI, P. 2013. Why coelacanths are not ‘living fossils’. Bioessays, 35: 332–338. DOI 10.1002/bies.201200145.
CASTANET, J., MEUNIER, F., BERGOT, C. and FRANÇOIS, Y. 1975. Données pré l iminaires sur les s t ructures histologiques du squelette de Latimeria chalumnae. I - Dents, écailles, rayons de nageoires. In: Coll. Inter. CNRS, "Problèmes actuels de Paléontologie. Evolution des Vertébrés", Paris, 4–9 Juin 1973, 159–168, 4 Pls.
CLEMENT, G. 1999. The actinistian (Sarcopterygii) Piveteauia madagascarensis LEHMAN from the Lower Triassic of Northeastern Madagascar: a redescription on the basis of new material. Journal of Vertebrate Paleontology, 19: 234–242.
CLEMENT, G. 2005. A new coelacanth (Actinist ia, Sarcopterygii) from the Jurassic of France, and the question of the closest relative fossil to Latimeria. Journal of Vertebrate Paleontology, 25: 481–491.
CUPELLO, C., BRITO, P. M., MEUNIER, F. J., HERBIN, M., JANVIER, P., DUTEL, H. & CLÉMENT, G. 2015. Allometric growth in the extant coelacanth lung during ontogenetic development. Nature Communications, 6: 8222. DOI: 10.1038/ncomms9222.
CUPELLO, C., CLEMÉNT, G., MEUNIER, F. J., HERBIN, M., YABUMOTO, Y. and BRITO, P. M. 2019. The long-time adaptation of coelacanths to moderate deep water: reviewing the evidences. Bulletin of Kitakyushu Museum of Natural History and Human History Series A (Natural History), 17: 29–35.
CUPELLO, C., MEUNIER, F. J., HERBIN, M., JANVIER, P., CLÉMENT, G. and BRITO, P. M. 2017. The homology and funct ion of the lung pla tes in extant and foss i l coelacanths. Scientific Reports , 7 : 9244. DOI: 10.1038/s41598-017-09327-6.
EASTMAN, J. T., WITMER, L. M., RIDGELY, R. C. and KUHN, K.
L. 2014. Divergence in skeleta l mass and bone morphology in antarctic notothenioid fishes. Journal of Morphology, 275: 841–861. DOI: 10.1002/jmor.20258
ERDMANN, M. V., CALDWELL, R. L. and MOOSA, M. K. 1998. Indonesian “king of the sea” discovered. Nature, 395: 335.
ERDMANN, M. V., CALDWELL, R. L., JEWETT, S. L. and TJAKRAWIDJAJA, A. 1999. The second recorded living coelacanth from north Sulawesi. Environmental Biology of Fishes, 34: 445–451.
FAURÉ-FREMIET, M. 1936. La structure des fibres d’éastoidine. Archives d'Anatomie Microscopique, 32: 249–269.
FOREY, P. L.- 1984. The coelacanth as a living fossil. In: ELREDGE N. and STANLEY S.M. (eds.), Living fossils. Springer Verlag, Berlin, pp. 166–169.
FOREY, P. L.- 1998. History of the Coelacanth Fishes. Chapman and Hall, London, 419 pp.
FRANCILLON, H., MEUNIER, F., NGO TUAN PHONG, D. and RICQLÈS, A. (DE). 1975. Données préliminaires sur les structures histologiques du squelette de Latimeria chalumnae. I I- Tissus osseux et cartilages. In: Coll. Inter. CNRS, "Problèmes actuels de Paléontologie. Evolution des Vertébrés", Paris, 4–9 Juin 1973, Centre National de la Recherche Scientifique, pp. 169–174, 2 pls.
FRANCILLON-VIEILLOT H., BUFFRENIL V. DE, CASTANET J., GÉRAUDIE J., MEUNIER FJ., SIRE J-Y., ZYLBERBERG L., a n d R I C Q L È S A . D E 1 9 9 0 . M i c r o s t r u c t u r e a n d mineralization of vertebrate skeletal tissues. In: CARTER, J. G. (ed.), Skeletal Biomineralization: Patterns, Processes and Evolutionary Trends. Vol. I , Van Nostrand, New York, pp. 471–530
GARRAULT, H., 1936. Développement des fibres d’élastoidine (actinotrichia) chez les salmonides. Archives d'Anatomie Microscopique, 130: 105–137.
GÉRAUDIE, J. and MEUNIER, F. J. 1980. Elastoidin actinotrichia in coelacanth fins: a comparative study with teleosts. Tissue and Cell, 12: 637–645.
GÉRAUDIE, J. and MEUNIER, F. J. 1984. Structure and comparative morphology of camptotrichia of lungfish fins. Tissue and Cell, 16: 217–236.
GIRAUD, M. M., CASTANET, J., MEUNIER, F. J. and BOULIGAND, Y. 1978a. The fibrous structure of coelacanth scales: a twisted "plywood". Tissue and Cell, 10: 671–686.
GIRAUD, M. M., CASTANET, J., MEUNIER, F. J. and BOULIGAND, Y. 1978b. Organisation spatiale de l'isopédine des écailles du coelacanthe (Latimeria chalumnae SMITH). Comptes Rendus de l’Académie des Sciences, 287: 487–489.
GRADY, J . E. 1970. Tooth development in Latimeria chalumnae (SMITH). Journal of Morphology, 132: 377–388.
HADIATY, R. K. and RACHMATIKA, I. 2003. Morphological study of the scales of Latimeria menadoensis POUYAUD et al., 1999. Treubia (A Journal on Zoology of the Indo-Australian Archipelago), 33: 1–11.
HOBDELL, M. H. and MILLER, W. A. 1969. Radiographic
anatomy of the teeth and tooth supporting tissues of Latimeria chalumnae. Archives of Oral Biology, 14: 855–858.
HUREAU J.-Cl. and OZOUF, C. 1977. Détermination de l’âge et croissance du cœlacanthe Latimeria chalumnae SMITH, 1939 (Poisson, Crossoptérygien, Coelacanthidé). Cybium, 2: 129–137.
JANVIER, P. 1996. Early Vertebrates. Clarendon Press, Oxford, 393 pp.
MEUNIER, F. J. 1979. Etude histologique et microradiographique du cartilage hémal de la vertèbre de la carpe, Cyprinus carpio L. (Pisces, Teleostei, Cyprinidae). Acta Zoologica, 60: 19–31.
MEUNIER, F. J. 1980. Les relations isopédine-tissu osseux dans le post-temporal et les écailles de la ligne latérale de Latimeria chalumnae (SMITH). Zoologica Scripta, 9: 307–317.
MEUNIER, F. J. and ZYLBERBERG, L. 1999. The structure of the external layer and of the odontodes of scales in Latimeria chalumnae (Sarcopterygii, Actinistia, Coelacanthidae) revisited using scanning and transmission electron microscopy. In: SÉRET, B. and SIRE, J.-Y. (eds.), Proc. 5th Indo-Pacific Fish Conf., Nouméa, 1997, Soc. Fr. Ichtyol., Paris, pp. 109–116.
MEUNIER, F. J., ERDMANN, M. V., FERMON, Y. and CALDWELL, R. L. 2008. Can the comparative study of the morphology and histology of the scales of Latimeria menadoensis and L. chalumnae (Sarcopterygii, Actinistia, Coelacanthidae) bring new insight on the taxonomy and the biogeography of recent coelacanthids? Geological Society, London, Special Publications, 295: 351–360.
MEUNIER, F. J., BRITO, P. M. and LEAL, M.-E. 2013. Quelques données morphologiques et histologiques sur la structure de la plaque dentaire linguale d’Arapaima gigas (Osteoglossidae; Teleostei). Cybium, 37: 263–271
MEUNIER, F. J., MONDEJAR-FERNANDEZ, J., GOUSSARD, F., CLÉMENT, G. and HERBIN, M. 2015. Presence of plicidentine in the oral teeth of Latimeria chalumnae (Sarcopterygii; Actinistia; Coelacanthidae). Journal of Structural Biology, 190: 31–37.
MILLER, W. A. 1979. Observations on the structure of mineralized tissue of the coelacanth, including scales and their associate odontodes. Occasional Papers of the Californian Academy of Sciences, 134: 68–78.
MILLER, W. A. and HOBDELL, M. H. 1968. Preliminary report on the histology of the dental and paradental tissues of Latimeria chalumnae (SMITH) with a note on tooth replacement. Archives of Oral Biology, 13: 1289–1291.
MILLOT, J. and ANTHONY, J. 1958. Anatomie de Latimeria chalumnae. T. 1. Squelette, muscles et formations de soutien. CNRS Edr., Paris. 122 pp., 80 pls.
MILLOT, J., ANTHONY, J. and ROBINEAU, D. 1978. Anatomie de Latimeria chalumnae. T. 3. Appareil digestif, téguments, écailles. CNRS Edr., Paris. pp. 7–28, 135–139.
MONDEJAR-FERNANDEZ, J. 2018. On cosmine: its origins, b io logy and impl ica t ions for sarcopterygian interrelationships. Cybium, 42: 41–65.
ØRVIG T. 1951. Histologic studies of Placoderms and fossil Elasmobranchs. 1: The endoskeleton, with remarks on the hard tissues of lower vertebrates in general. Arkiv för Zoologi, 2: 321–454, 8 pl.
ØRVIG T. 1968. The dermal skeleton; general considerations. In: ØRVIG T. (ed.), Current problems of lower vertebrate phylogeny. Proceedings of the fourth Nobel Symposium, Stockholm, 1967, J. Wiley, New-York. pp. 373–397.
PARMENTIER, E., COMPÈRE, P., CASADEWALL, M., FONTENELLE, N., CLOOTS, R. and HENRIST, C. 2008. The rocker bone: a new kind of mineralized tissue? Cell and Tissue Research, 334: 67–79.
PEGUETA, V. P. 1968. Enchondral ossification in Latimeria chalumnae SMITH. Dopovidi Akademii Nauk Ukrainia SSR., 7: 653–656 (in Russian; English abstr.).
POUYAUD, L., WIRJOATMODIO, S., RACHMATIKA, L., TJAKRAWIDJAJA, A., HADIATY, R. K. and HADIE, W. 1999. Une nouvelle espèce de coelacanthe, preuves génétiques et morphologiques. Comptes Rendus de l’Académie des Sciences, 322: 261–267.
ROMER, A.S. 1937. The braincase of the carboniferous crossopterygian Megalichthys nitidus. Bulletin of the Museum of Comparative Zoology, 824: 1–73.
ROMER, A.S. 1942. Cartilage as an embryonic adaptation. American Naturalist, 76: 394–404.
ROUX, G. H. 1942. The microscopic anatomy of the Latimeria scale. South African Journal of Medical Sciences, 7: 1–18.
SASAGAWA, I., ISHIYAMA, M. and KODERA, H. 1984. Fine structure of the pharyngeal teeth in the coelacanth fish (Latimeria chalumnae). In: FEARNHEAD, R. W. and SUGA, S. (eds.), Tooth enamel IV, Elsevier Science Publishers, Amsterdam, pp. 462–466.
SCHULTZE, H.-P. 1969. Die Faltenzähne der Rhipidistiden Crossopterygier, der Tetrapoden und der Actinopterygi-er-Gattung Lepisosteus nebst einer Beschreibung der Zahnstruktur von Onychodus (Struniiformer Crossoptery-gier). Palaeontologica Italica, New Series, 35(65): 63–137.
SCHULTZE, H.-P. 1970. Folded teeth and the monophyletic origin of Tetrapods. American Museum Novitates, 2408: 1–10.
SCHULTZE, H.-P. 2018. Hard tissues in fish evolution: history and current issues. Cybium, 42(1): 29–39.
SHELLIS, R. P. and POOLE, D. F. G. 1978. The structure of the dental hard tissues of the coelacanthid fish Latimeria chalumnae SMITH. Archives of Oral Biology, 23: 1105–1113.
SIRE, J.-Y. and HUYSSEUNE, A. 2003. Formation of dermal skeletal and dental tissues in fish: a comparative and evolutionary approach. Biological Review, 78: 219–249.
SMITH, J. L. B. 1939. A living fish of the Mesozoic type. Nature, 143: 455–456.
SMITH, J. L. B. 1940. A living coelacanth fish from South Africa. Transactions of the Royal Society of South Africa, 28: 1–106, 44 Pls.
SMITH, M. M. 1978. Enamel in the oral teeth of Latimeria chalumnae (Pisces: Actinistia): a scanning electron microscopic study. Journal of Zoology, 185: 355–369.
SMITH, M. M. 1979. Scanning electron microscopy of odontodes in the scales of coelacanth embryo, Latimeria chalumnae SMITH. Archives of Oral Biology, 24: 179–183.
SMITH, M. M., HOBDELL, M. H. and MILLER, W. A. 1972. The structure of the scales of Latimeria chalumnae. Journal of Zoology, London, 167: 501–509.
THOMSON, K. S. 1969. The biology of the lobe-finned fishes.
Biological Reviews, 44: 91–154. UYENO, T. and YABUMOTO, Y. 2007. Origin of extant
coelacanths. The Coelacanth, Fathom the Mystery 2007. Aquamarine Fukushima, pp. 24–26.
ZYLBERBERG, L. and MEUNIER, F. J. 2008. New data on the structure and the chondrocyte populations of the haemal cartilage of abdominal vertebrae in the adult carp Cyprinus carpio (Teleostei, Ostariophysii, Cyprinidae). Cybium, 32: 225–239.
ZYLBERBERG, L., GÉRAUDIE, J., MEUNIER, F. J., & SIRE J. Y., 1992. Biomineralization in the integumental skeleton of the living lower Vertebrates. In: HALL, B. K. (ed), Bone, Volume 4, CRC Press, Boca Raton, pp. 171–224.

type according to the definition of MEUNIER et al. (2013). This organization is less complex than the three other plicidentine types (polyplocodont, eusthenodont and dendrodont), which are described in most of extinct and extant Sarcopterygii (SCHULTZE, 1969, 1970).The bony plates of the lung
The lung of Latimeria is reduced to a short oesophagus diverticulum (CUPELLO et al., 2015) surrounded by scattered small and thin ovoid plates (Fig. 9A, B). These plates cover the surface of the vestigial lung, and are composed of a fibrous matrix with fibrocytes: it is a cellular bony tissue (CUPELLO et al., 2017). Each plate is enclosed in a membranous bag. These plates look fragile in their middle plane since a central weakness zone opened on the microtome knife, creating an artefactual lumen (Fig. 9C) (CUPELLO et al., 2017). The fibres are collagenous and organized in superposed layers. The cells are true osteocytes (Fig. 9E) with a more or less star-shaped outline, and send cytoplasmic extensions in the thickness of the plate (Fig. 9D, E). Rare lining cells (osteoblasts) are
present on the surface of the plates, and conjunctive fibres (SHARPEY’s fibres) penetrate within the plate (Fig. 9C). The extracellular matrix shows various staining intensities with concentric and parallel (Fig. 9D) or globular shaped lines. These lines are considered to be Liesegang lines that characterize an active spheritic mineralizing process. The lung plates of Latimeria are thus true bony plates, and are homologous with the large bony plates known in the abdominal cavity of fossil actinistians (BRITO et al., 2010; CUPELLO et al., 2017). The calcified walls that surrounded the lung of the Cretaceous coelacanth Axelrodichthys are made of large osseous plates of various thickness (BRITO et al., 2010), as those of other fossil coelacanths (CLEMENT, 1999, 2005). The bony tissue of these plates is a vascularized cellular bone with more or less large vascular cavities and some internal remodelling (BRITO et al., 2010). Moreover the mineralization of the lung ossified plates in Axelrodichthys is spheritic as in Latimeria’s plates (CUPELLO et al., 2017). So there is a continuity of the histological structure of the lung plates in the coelacanths during their
evolution, but with an important reduction of the bony plates in Latimeria linked to the vestigial state of its lung (CUPELLO et al., 2015, 2019).
CONCLUSION
This overview of eighty years (1938–2018) of histological work on Latimeria skeletal tissues allows some anatomical and evolutionary considerations. It can be enlightened a drastic reduction of endochondral ossification during the long evolutionary history of coelacanths. The persistence of large volume of cartilage in the endoskeleton at adult stage can be compared to the “little bone and considerable cartilage” that characterize the skeleton of a number of demersal notothenioid telostean fishes (EASTMAN et al., 2014).
A processus of spheritic mineralization has been recently highlighted in various skeletal elements of Latimeria, by the presence of globular dentine in teeth, in odontodes of the tegumentary skeleton (scales, fin rays), in scales at the interface between the external layer and the basal plate, as well as in lung bony plates. The spheritic mineralization (i.e., a radiating arrangement of hydroxyapatite crystals and of the organic matrix) in vertebrate skeletal tissues is considered as a precursor of inotropic mineralization (specific interactions between collagen fibrils and the mineral phase) that possibly represents a derived evolutionary stage of calcification mechanisms (ØRVIG, 1951, 1968; FRANCILLON-VIEILLOT et al., 1990; ZYLBERBERG et al., 1992). The simultaneous presence of both mineralization processes (spheritic and inotropic) in the mineralized tissues of Latimeria and in other osteichthyans such as teleosteans (ZYLBERBERG and MEUNIER, 2008) is however questionable. The mineralized spherules present at the limit between the external layer and the basal plate of Latimeria scales are considered to be the product of an inotropic mineralization (MEUNIER and ZYLBERBERG, 1999). It thus appears that the state of the mineralized spherules observed in the various skeletal tissues of Latimeria, including the lung bony plates, must be studied with adapted ultrastructural techniques in order to test their true origin: spheritic or inotropic mineralization?
ACKNOWLEDGEMENTS
This contribution is linked to an international roundtable on coelacanths evolution held at the Kitakyushu Museum and at the Aquamarine Fukushima in August 2017. We would like to thank the anonymous reviewers for their constructive comments and valuable suggestions. C.C. was partially financed by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brasil (CAPES) – Finance Code 001 (Programa Nacional de Pós Doutorado-PNPD).
38 The skeleton and the mineralized tissues of the living coelacanths 39
INTRODUCTION
The first study of fossil coelacanths was done by AGASSIZ (1833–44) and was followed by a lot of papers with the discovery of new fossils (see reviews of JANVIER, 1996 and FOREY, 1998). Fossil coelacanths constitute an important monophyletic group composed of more than forty genera, eighty species, and belong to the Sarcopterygii (ANDREWS, 1973; JANVIER, 1996; FOREY, 1998; UYENO and YABUMOTO, 2007; and many others). For a century coelacanths were considered extinct since the end of the Cretaceous time (Fig. 1).
With the discovery of a living coelacanth in 1938 (SMITH, 1939) the scientific community was soon highly interested to compare the incomplete fossil sarcopterygian fishes dataset to the recent anatomical and biological organization of Latimeria chalumnae. Many authors (see THOMSON, 1969; MILLOT and ANTHONY, 1958; MILLOT et al., 1978 FOREY, 1984, 1998; JANVIER, 1996) focused their work on Latimeria as a key-taxon to test or propose various hypotheses on the origin and evolutionary history of the early tetrapods and sarcopterygii in general, and to infer their possible biological characteristics. The fossil sarcopterygian fishes are known by their fossilized mineralized tissues, essentially their bones, scales and teeth. Together with the extant lungfishes, especially Neoceratodus, Latimeria offered a unique access to the skeleton as a whole: with both mineralized and unmineralized skeletal tissues. Here, we aim to present an overview of the last eighty years (1938–2018) of histological studies of Latimeria’s skeleton.
Since the discovery of the Raja Laut (“king of the sea” ), off Sulawesi Island in Indonesia (ERDMANN et al., 1998, 1999), the genus Latimeria comprises two species: L. chalumnae SMITH 1939 and L. menadoensis POUYAUD et al., 1999. However our knowledge of the anatomy and biology of the living coelacanths is so far essentially based on L. chalumnae.
THE LATIMERIA SKELETON
The first anatomical observations on the extant coelacanth were done by SMITH (1940). The anatomy of the Latimeria skeleton has been later described in detail by MILLOT and ANTHONY (1958). The living coelacanth shows a general organization similar to that of the other actinistian fishes (THOMSON, 1969; JANVIER, 1996; FOREY, 1998). The skeleton can be divided in various parts (Fig. 2): skull, axial skeleton, paired and unpaired fins, tegumentary skeleton (scales). The skull, axial skeleton and fins are constituted of endoskeletal elements that are overlain by exoskeletal elements. In the extant Latimeria the endoskeleton of the skull shows an important regression of the bones, especial ly in the neurocranium, which are replaced by cartilaginous tissues, as in its Mesozoic fossil actinistian relatives (ROMER, 1937, 1942; MILLOT and ANTHONY, 1958; BJERRING, 1973; FOREY, 1998). In the same way, the endoskeleton of paired and unpaired fins is essentially constituted of four axial cartilaginous elements (Fig. 2; MILLOT and ANTHONY, 1958), with pre and post-axial
elements and eventually minute superficial endochondral ossification (CASTANET et al., 1975). The axial skeleton is composed of the notochord which is coated by an unmineralized fibrillary sheath and is totally deprived of well developed vertebrae due to the lack of ossified centra, although some discrete neural and haemal arches develop in the anterior part of the notochord (MILLOT and ANTHONY, 1958). The anterior neural and haemal spines are relatively short but they progressively increase in length posteriorly. Importantly neural and haemal spines are composed of perichondral bone surrounding a cartilaginous core (Fig. 2).
The scales belong to the exoskeleton. In both extant coelacanth species they are of elasmoid type, composed of an upper external layer also called “external ornamented layer”, and of a lower thicker layer, called the basal plate, which is stratified and almost totally unmineralized (Fig. 3). The upper external layer is ornamented with radial crests. In the posterior area of the scale these radial reliefs are overlain by numerous denticulations, the odontodes, which can be superposed (Fig. 3). The anterior area of the external layer, which is covered by the anterior scales, shows crests with irregularities that have been interpreted as growth marks and tentatively used for ageing coelacanths (HUREAU and OZOUF, 1977; MILLER, 1979).
The tooth plates of the buccal cavity support series of teeth of various sizes (Fig. 4). They range in three morphotypes: the fangs (7–10 mm in height), middle-sized teeth (3–4 mm in height) and rounded tubercles (MILLOT and ANTHONY, 1958). The teeth of the two first categories are conical and sharp whereas those of the third category have an obtuse tip (Fig. 5). Fangs of Latimeria are inserted into a socket (MILLOT and ANTHONY, 1958; HOBDELL and MILER, 1969; CASTANET et al., 1975) and they have a smooth external surface (MILLOT et al., 1978; MEUNIER et al., 2015).
A particularity of the exoskeleton of Latimeria is the abundance of odontodes at the surface of various skull bones
Fig. 3. Latimeria chalumnae and L. menadoensis. Details of the posterior field of scales (SEM). A, L. chalumnae. General view of the odontodes. The posterior margin of the scale is on the upper left. (scale = 500 µm). B, L. menadoensis. General view of the odontodes. The posterior margin of the scale is on the upper right. (scale = 500 µm). C, L. menadoensis. Closed-up view of the odontodes positioned on the radial crests of the external layer. (scale = 100 µm). D, L. chalumnae. Closed-up view of the odontodes positioned on the radial crests of the external layer, and two superimposed odontodes are seen on the right (*). (scale = 100 µm). E, L. chalumnae. Closed-up view of two superimposed odontodes with the apical part of the lower one being eroded (arrow). (scale = 100 µm). (After MEUNIER et al., 2008).
Fig. 4. Latimeria chalumnae. Buccal roof showing the different tooth types: fangs (black arrow-heads), small caniniform teeth (white arrow-heads) and patches of round shaped teeth (black arrows), especially on the parasphenoid tooth plate (t.par) and the medial side of the pterygoid (t.pt). The front side is on the top of the figure. (After MILLOT and ANTHONY, 1958). (scale = 10 mm).
(MILLOT and ANTHONY, 1958; BERNHAUSER, 1961), on the fin rays (CASTANET et al., 1975) and on the posterior (free) area of scales (Fig. 3) (MILLOT and ANTHONY, 1958; CASTANET et al., 1975; HADIATY and RACHNATIKA, 2003; MEUNIER et al., 2008).
Among the various components of the whole skeleton, and apart the SMITH’s pioneer work (SMITH, 1940), numerous studies have been carried out on the histological organization of scales (ROUX, 1942; SMITH et al., 1972; CASTANET et al., 1975; GIRAUD et al., 1978a; MILLER, 1979; SMITH, 1979; MEUNIER, 1980; MEUNIER and ZYLBERBERG, 1999; HADIATY and RACHMATIKA, 2003; MEUNIER et al., 2008) and teeth (MILLER and HOBDELL, 1968; GRADY, 1970; HOBDELL and MILLER, 1969; CASTANET et al., 1975; SHELLIS and POOLE, 1978;
SMITH, 1978; SASAGAWA et al., 1984; MEUNIER et al., 2015). Contrary to the superficial skeleton, there are clearly less histological studies of the skeletal bony elements (FRANCILLON et al., 1975). Meanwhile, in addition to the usual skeletal elements, specific bony tissues have been described in fossil (BRITO et al., 2010) and extant coelacanths (CUPELLO et al., 2015, 2017), such as the mineralized plates surrounding the lung. These bony plates in actinistians do not belong to the skeleton sensu stricto. They are specific bony specializations in relation to the lung (see below), as is the “rocker bone” in relation to the gas bladder of some ophidiiform teleosts (PARMENTIER et al., 2008). Despite the name, the rocker bone is not true bony tissue (PARMENTIER et al., 2008), contrary to
the bony plates of fossil and extant coelacanths (BRITO et al., 2010; CUPELLO et al., 2017).
ADULT LATIMERIA SKELETAL TISSUES
Bony tissuesCortical areas of the dermal skeleton are constituted of
primary periosteal bony t issue. I ts mineralization is heterogeneous and generally marked by series of growth lines, probably due to alternative physiological cycles linked to seasonal variations (Fig. 6). The cortical primary bone shows vascular canals and numerous SHARPEY’s fibres (Fig. 6B). The centre of these bones is frequently made of a spongiosa with vascular cavities surrounded by secondary bony deposits that result from remodelling processes (Fig. 6). This secondary bone is separated from the primary one by reversal cementing lines that are hypermineralized.
Bone remodelling in adult specimens can be more or less developed according to a bone and within a given bone. Primary bone is destroyed by osteoclasts and the deposition process of the secondary bone results from the activity of osteoblasts that have replaced the osteoclasts. The secondary bone does generally not replaced the whole primary bone areas, which can still be recognizable by the presence of
SHARPEY’s fibres and/or growth marks (Fig. 6).The dermal fin rays that sustain paired and unpaired fins
in Latimeria are true lepidotrichia (CASTANET et al., 1975). They are made of two parallel opposite gutter-shaped hemirays, each of which being a series of hemisegments articulated by a collagenous ligament. Each hemisegment is made of a mineralized collagenous fibrillary matrix with embedded osteocytes. The hemisegments are sometimes fused at the basal part of the rays due to centrifugal deposition of bony laminae around the primary articulation, and eventually to the mineralization of the ligament (see figs. 19, 20 in CASTANET et al., 1975). Odontodes observed on the external convex surface of the rays are similar to those present on the dermal bones of the skull and scales. The distal extremity of the lepidotrichia is overlapped by actinotrichia (GÉRAUDIE and MEUNIER, 1980) that are long tapered rods of elastoidin , a fibrous protein of collagenous nature (FAURÉ-FREMIET, 1936; GARRAULT, 1936). The presence of lepidotrichia with distal actinotrichia in Latimeria, as in the fins of Actinopterygii, is
regarded as a plesiomorphic character for Osteichthyes (GÉRAUDIE and MEUNIER, 1980, 1984).
The upper layer of the Latimeria scale is relatively thin and it is constituted of bony tissue with embedded osteocytes, whereas the basal plate is much thicker and composed of numerous strata made of thick collagenous fibres (Fig. 7). Between two collagenous layers there are star-shaped cells (SMITH et al., 1972, pl. VI; CASTANET et al., 1975, fig. 12), the elasmocytes, which cytoplasmic processes insert between the collagenous fibres. These fibres are set in a very specific network. In each layer, the fibres are parallel to each other and the direction of the fibres change from a layer to another one. This regular organization results in a spatial arrangement termed “twisted plywood” (GIRAUD et al., 1978a,b). The rotation of fibres direction from one layer to the next has a mean angle of 27° (GIRAUD et al., 1978a). This organization is found in each scale of L. chalumnae. In L. menadoensis, the basal plate of the scales is also a twisted plywood but its rotation angle seems to be slightly less regular (MEUNIER et al.,
2008). The basal plate in both species is unmineralized (Fig. 7) excepted at the contact between the superficial layer and the basal plate, where spheritic mineralized granules are seen in the very first layers of the basal plate (MEUNIER, 1980; MEUNIER and ZYLBERBERG, 1999; MEUNIER et al., 2008). There is no pore-canal system so Latimeria’s scales are not cosmoid, contrary to most of palaeozoic sarcopterygian fishes. Due to the presence of an unmineralized stratified basal plate, the scales of Latimeria are defined as elasmoid-like scales. However the plywood-like organization is considered to be homologous with the bony basal plate of cosmoid scales of extinct sarcopterygian fishes (MEUNIER, 1980; SIRE and HUYSSEUNE, 2003; MONDEJAR, 2018; SCHULTZE, 2018).Cartilages
The cartilaginous tissues in Latimeria are characterized by long chondrocytes (Fig. 8) contrary to those of teleostean fishes that are relatively spherical in shape (MEUNIER, 1979; ZYLBERBERG and MEUNIER, 2008). Cartilage tissues can show an endochondral ossification process. This phenomenon has been studied respectively on the urohyal (PEGUETA, 1968), the MECKEL’s cartilage and the proximal piece of the pectoral girdle (FRANCILLON et al., 1975). The endochondral ossification is relatively limited when it occurs and the volume of enchondral bone remains reduced to thin bony layers (Fig. 8). The cartilage is destroyed by chondroclasts in areas where endochondral ossification occurs. Serial chondrocytes, known at the origin of the calcified cartilage, seem to be lacking in the endochondral ossification process in Latimeria (FRANCILLON et al., 1975). This constitutes a true difference with teleostean endochondral ossifications (MEUNIER, 1979; ZYLBERBERG and MEUNIER, 2008). Tooth tissues
The three tooth morphotypes described in Latimeria are fangs (7–10 mm in height), middle-sized teeth (3–4 mm in height) and rounded tubercles (MILLOT and ANTHONY, 1958). These teeth and tubercles are constituted of a cone of orthodentine set around a large pulp cavity and overlain by an external hypermineralized layer considered as true enamel (GRADY, 1970; CASTANET et al., 1975; SHELLIS and POOLE, 1978; SMITH, 1978; SASAGAWA et al., 1984). Various studies have revealed the presence of globular dentine (Fig. 5) at the periphery of the first third of the tooth (MILLER and HOBDELL, 1968; HOBDELL and MILLER, 1969; SHELLIS and POOLE, 1978; SASAGAWA et al., 1984). This typical histological organization characterizes the various teeth of the bucco-pharyngeal cavity as well as the various odontodes of the dermal bony plates, the fin rays and the scales (CASTANET et al., 1975).
In the fang the inner wall of the pulp cavity displays a series of plies (Fig. 5; MEUNIER et al., 2015). These plies start at the base of the tooth and reach at least the first third to the half of the tooth towards its tip (Fig. 5). Such plies are also present but less developed in the small caniniform teeth (MEUNIER et al., 2015). These dentine plies in the pulp cavity characterizes a plicidentine organization of orthodentine type. These primary plies correspond to a simplexodont plicidentine
REFERENCES
AGASSIZ, L. 1833–44. Recherches sur les Poissons fossiles. Imprimerie Petitpierre, Neuchâtel. [5 volumes and atlas.]
ANDREWS, S. M. 1973. Interrelationships of crossopterygians. In: GREENWOOD, P. H., MILES, R. S. and PATTERSON, C. (eds.), Interrelationships of Fishes, Acad. Press, London, pp. 137–177.
BERNHAUSER, A. 1961. Zur Knochen- und Zah, histology von Latimeria chalumnae Smith und einiger Fossilformen. Sber. öst. Akad. Wiss., 170: 119–137.
BJERRING, H. C. 1973. Relationships of coelacanthiforms. In: GREENWOOD P. H., MILES R. S. and PATTERSON C. (eds.), Interrelationships of Fishes, Acad. Press, London, pp. 179–205.
BRITO, P. M., MEUNIER, F. J., CLÉMENT, G. and GEFFARD-KURIYAMA, D. 2010. The histological structure of the calcified lung of the fossil coelacanth Axelrodichthys araripensis (Actinistia: Mawsoniidae). Palaeontology, 53: 1281–1290.
CASANE, D. and LAURENTI, P. 2013. Why coelacanths are not ‘living fossils’. Bioessays, 35: 332–338. DOI 10.1002/bies.201200145.
CASTANET, J., MEUNIER, F., BERGOT, C. and FRANÇOIS, Y. 1975. Données pré l iminaires sur les s t ructures histologiques du squelette de Latimeria chalumnae. I - Dents, écailles, rayons de nageoires. In: Coll. Inter. CNRS, "Problèmes actuels de Paléontologie. Evolution des Vertébrés", Paris, 4–9 Juin 1973, 159–168, 4 Pls.
CLEMENT, G. 1999. The actinistian (Sarcopterygii) Piveteauia madagascarensis LEHMAN from the Lower Triassic of Northeastern Madagascar: a redescription on the basis of new material. Journal of Vertebrate Paleontology, 19: 234–242.
CLEMENT, G. 2005. A new coelacanth (Actinist ia, Sarcopterygii) from the Jurassic of France, and the question of the closest relative fossil to Latimeria. Journal of Vertebrate Paleontology, 25: 481–491.
CUPELLO, C., BRITO, P. M., MEUNIER, F. J., HERBIN, M., JANVIER, P., DUTEL, H. & CLÉMENT, G. 2015. Allometric growth in the extant coelacanth lung during ontogenetic development. Nature Communications, 6: 8222. DOI: 10.1038/ncomms9222.
CUPELLO, C., CLEMÉNT, G., MEUNIER, F. J., HERBIN, M., YABUMOTO, Y. and BRITO, P. M. 2019. The long-time adaptation of coelacanths to moderate deep water: reviewing the evidences. Bulletin of Kitakyushu Museum of Natural History and Human History Series A (Natural History), 17: 29–35.
CUPELLO, C., MEUNIER, F. J., HERBIN, M., JANVIER, P., CLÉMENT, G. and BRITO, P. M. 2017. The homology and funct ion of the lung pla tes in extant and foss i l coelacanths. Scientific Reports , 7 : 9244. DOI: 10.1038/s41598-017-09327-6.
EASTMAN, J. T., WITMER, L. M., RIDGELY, R. C. and KUHN, K.
L. 2014. Divergence in skeleta l mass and bone morphology in antarctic notothenioid fishes. Journal of Morphology, 275: 841–861. DOI: 10.1002/jmor.20258
ERDMANN, M. V., CALDWELL, R. L. and MOOSA, M. K. 1998. Indonesian “king of the sea” discovered. Nature, 395: 335.
ERDMANN, M. V., CALDWELL, R. L., JEWETT, S. L. and TJAKRAWIDJAJA, A. 1999. The second recorded living coelacanth from north Sulawesi. Environmental Biology of Fishes, 34: 445–451.
FAURÉ-FREMIET, M. 1936. La structure des fibres d’éastoidine. Archives d'Anatomie Microscopique, 32: 249–269.
FOREY, P. L.- 1984. The coelacanth as a living fossil. In: ELREDGE N. and STANLEY S.M. (eds.), Living fossils. Springer Verlag, Berlin, pp. 166–169.
FOREY, P. L.- 1998. History of the Coelacanth Fishes. Chapman and Hall, London, 419 pp.
FRANCILLON, H., MEUNIER, F., NGO TUAN PHONG, D. and RICQLÈS, A. (DE). 1975. Données préliminaires sur les structures histologiques du squelette de Latimeria chalumnae. I I- Tissus osseux et cartilages. In: Coll. Inter. CNRS, "Problèmes actuels de Paléontologie. Evolution des Vertébrés", Paris, 4–9 Juin 1973, Centre National de la Recherche Scientifique, pp. 169–174, 2 pls.
FRANCILLON-VIEILLOT H., BUFFRENIL V. DE, CASTANET J., GÉRAUDIE J., MEUNIER FJ., SIRE J-Y., ZYLBERBERG L., a n d R I C Q L È S A . D E 1 9 9 0 . M i c r o s t r u c t u r e a n d mineralization of vertebrate skeletal tissues. In: CARTER, J. G. (ed.), Skeletal Biomineralization: Patterns, Processes and Evolutionary Trends. Vol. I , Van Nostrand, New York, pp. 471–530
GARRAULT, H., 1936. Développement des fibres d’élastoidine (actinotrichia) chez les salmonides. Archives d'Anatomie Microscopique, 130: 105–137.
GÉRAUDIE, J. and MEUNIER, F. J. 1980. Elastoidin actinotrichia in coelacanth fins: a comparative study with teleosts. Tissue and Cell, 12: 637–645.
GÉRAUDIE, J. and MEUNIER, F. J. 1984. Structure and comparative morphology of camptotrichia of lungfish fins. Tissue and Cell, 16: 217–236.
GIRAUD, M. M., CASTANET, J., MEUNIER, F. J. and BOULIGAND, Y. 1978a. The fibrous structure of coelacanth scales: a twisted "plywood". Tissue and Cell, 10: 671–686.
GIRAUD, M. M., CASTANET, J., MEUNIER, F. J. and BOULIGAND, Y. 1978b. Organisation spatiale de l'isopédine des écailles du coelacanthe (Latimeria chalumnae SMITH). Comptes Rendus de l’Académie des Sciences, 287: 487–489.
GRADY, J . E. 1970. Tooth development in Latimeria chalumnae (SMITH). Journal of Morphology, 132: 377–388.
HADIATY, R. K. and RACHMATIKA, I. 2003. Morphological study of the scales of Latimeria menadoensis POUYAUD et al., 1999. Treubia (A Journal on Zoology of the Indo-Australian Archipelago), 33: 1–11.
HOBDELL, M. H. and MILLER, W. A. 1969. Radiographic
anatomy of the teeth and tooth supporting tissues of Latimeria chalumnae. Archives of Oral Biology, 14: 855–858.
HUREAU J.-Cl. and OZOUF, C. 1977. Détermination de l’âge et croissance du cœlacanthe Latimeria chalumnae SMITH, 1939 (Poisson, Crossoptérygien, Coelacanthidé). Cybium, 2: 129–137.
JANVIER, P. 1996. Early Vertebrates. Clarendon Press, Oxford, 393 pp.
MEUNIER, F. J. 1979. Etude histologique et microradiographique du cartilage hémal de la vertèbre de la carpe, Cyprinus carpio L. (Pisces, Teleostei, Cyprinidae). Acta Zoologica, 60: 19–31.
MEUNIER, F. J. 1980. Les relations isopédine-tissu osseux dans le post-temporal et les écailles de la ligne latérale de Latimeria chalumnae (SMITH). Zoologica Scripta, 9: 307–317.
MEUNIER, F. J. and ZYLBERBERG, L. 1999. The structure of the external layer and of the odontodes of scales in Latimeria chalumnae (Sarcopterygii, Actinistia, Coelacanthidae) revisited using scanning and transmission electron microscopy. In: SÉRET, B. and SIRE, J.-Y. (eds.), Proc. 5th Indo-Pacific Fish Conf., Nouméa, 1997, Soc. Fr. Ichtyol., Paris, pp. 109–116.
MEUNIER, F. J., ERDMANN, M. V., FERMON, Y. and CALDWELL, R. L. 2008. Can the comparative study of the morphology and histology of the scales of Latimeria menadoensis and L. chalumnae (Sarcopterygii, Actinistia, Coelacanthidae) bring new insight on the taxonomy and the biogeography of recent coelacanthids? Geological Society, London, Special Publications, 295: 351–360.
MEUNIER, F. J., BRITO, P. M. and LEAL, M.-E. 2013. Quelques données morphologiques et histologiques sur la structure de la plaque dentaire linguale d’Arapaima gigas (Osteoglossidae; Teleostei). Cybium, 37: 263–271
MEUNIER, F. J., MONDEJAR-FERNANDEZ, J., GOUSSARD, F., CLÉMENT, G. and HERBIN, M. 2015. Presence of plicidentine in the oral teeth of Latimeria chalumnae (Sarcopterygii; Actinistia; Coelacanthidae). Journal of Structural Biology, 190: 31–37.
MILLER, W. A. 1979. Observations on the structure of mineralized tissue of the coelacanth, including scales and their associate odontodes. Occasional Papers of the Californian Academy of Sciences, 134: 68–78.
MILLER, W. A. and HOBDELL, M. H. 1968. Preliminary report on the histology of the dental and paradental tissues of Latimeria chalumnae (SMITH) with a note on tooth replacement. Archives of Oral Biology, 13: 1289–1291.
MILLOT, J. and ANTHONY, J. 1958. Anatomie de Latimeria chalumnae. T. 1. Squelette, muscles et formations de soutien. CNRS Edr., Paris. 122 pp., 80 pls.
MILLOT, J., ANTHONY, J. and ROBINEAU, D. 1978. Anatomie de Latimeria chalumnae. T. 3. Appareil digestif, téguments, écailles. CNRS Edr., Paris. pp. 7–28, 135–139.
MONDEJAR-FERNANDEZ, J. 2018. On cosmine: its origins, b io logy and impl ica t ions for sarcopterygian interrelationships. Cybium, 42: 41–65.
ØRVIG T. 1951. Histologic studies of Placoderms and fossil Elasmobranchs. 1: The endoskeleton, with remarks on the hard tissues of lower vertebrates in general. Arkiv för Zoologi, 2: 321–454, 8 pl.
ØRVIG T. 1968. The dermal skeleton; general considerations. In: ØRVIG T. (ed.), Current problems of lower vertebrate phylogeny. Proceedings of the fourth Nobel Symposium, Stockholm, 1967, J. Wiley, New-York. pp. 373–397.
PARMENTIER, E., COMPÈRE, P., CASADEWALL, M., FONTENELLE, N., CLOOTS, R. and HENRIST, C. 2008. The rocker bone: a new kind of mineralized tissue? Cell and Tissue Research, 334: 67–79.
PEGUETA, V. P. 1968. Enchondral ossification in Latimeria chalumnae SMITH. Dopovidi Akademii Nauk Ukrainia SSR., 7: 653–656 (in Russian; English abstr.).
POUYAUD, L., WIRJOATMODIO, S., RACHMATIKA, L., TJAKRAWIDJAJA, A., HADIATY, R. K. and HADIE, W. 1999. Une nouvelle espèce de coelacanthe, preuves génétiques et morphologiques. Comptes Rendus de l’Académie des Sciences, 322: 261–267.
ROMER, A.S. 1937. The braincase of the carboniferous crossopterygian Megalichthys nitidus. Bulletin of the Museum of Comparative Zoology, 824: 1–73.
ROMER, A.S. 1942. Cartilage as an embryonic adaptation. American Naturalist, 76: 394–404.
ROUX, G. H. 1942. The microscopic anatomy of the Latimeria scale. South African Journal of Medical Sciences, 7: 1–18.
SASAGAWA, I., ISHIYAMA, M. and KODERA, H. 1984. Fine structure of the pharyngeal teeth in the coelacanth fish (Latimeria chalumnae). In: FEARNHEAD, R. W. and SUGA, S. (eds.), Tooth enamel IV, Elsevier Science Publishers, Amsterdam, pp. 462–466.
SCHULTZE, H.-P. 1969. Die Faltenzähne der Rhipidistiden Crossopterygier, der Tetrapoden und der Actinopterygi-er-Gattung Lepisosteus nebst einer Beschreibung der Zahnstruktur von Onychodus (Struniiformer Crossoptery-gier). Palaeontologica Italica, New Series, 35(65): 63–137.
SCHULTZE, H.-P. 1970. Folded teeth and the monophyletic origin of Tetrapods. American Museum Novitates, 2408: 1–10.
SCHULTZE, H.-P. 2018. Hard tissues in fish evolution: history and current issues. Cybium, 42(1): 29–39.
SHELLIS, R. P. and POOLE, D. F. G. 1978. The structure of the dental hard tissues of the coelacanthid fish Latimeria chalumnae SMITH. Archives of Oral Biology, 23: 1105–1113.
SIRE, J.-Y. and HUYSSEUNE, A. 2003. Formation of dermal skeletal and dental tissues in fish: a comparative and evolutionary approach. Biological Review, 78: 219–249.
SMITH, J. L. B. 1939. A living fish of the Mesozoic type. Nature, 143: 455–456.
SMITH, J. L. B. 1940. A living coelacanth fish from South Africa. Transactions of the Royal Society of South Africa, 28: 1–106, 44 Pls.
SMITH, M. M. 1978. Enamel in the oral teeth of Latimeria chalumnae (Pisces: Actinistia): a scanning electron microscopic study. Journal of Zoology, 185: 355–369.
SMITH, M. M. 1979. Scanning electron microscopy of odontodes in the scales of coelacanth embryo, Latimeria chalumnae SMITH. Archives of Oral Biology, 24: 179–183.
SMITH, M. M., HOBDELL, M. H. and MILLER, W. A. 1972. The structure of the scales of Latimeria chalumnae. Journal of Zoology, London, 167: 501–509.
THOMSON, K. S. 1969. The biology of the lobe-finned fishes.
Biological Reviews, 44: 91–154. UYENO, T. and YABUMOTO, Y. 2007. Origin of extant
coelacanths. The Coelacanth, Fathom the Mystery 2007. Aquamarine Fukushima, pp. 24–26.
ZYLBERBERG, L. and MEUNIER, F. J. 2008. New data on the structure and the chondrocyte populations of the haemal cartilage of abdominal vertebrae in the adult carp Cyprinus carpio (Teleostei, Ostariophysii, Cyprinidae). Cybium, 32: 225–239.
ZYLBERBERG, L., GÉRAUDIE, J., MEUNIER, F. J., & SIRE J. Y., 1992. Biomineralization in the integumental skeleton of the living lower Vertebrates. In: HALL, B. K. (ed), Bone, Volume 4, CRC Press, Boca Raton, pp. 171–224.
François J. MEUNIER, Camila CUPELLO and Gaël CLÉMENT
Fig. 1. Schematic phylogeny of nine fossil genera of Actinistia and the extant Latimeria. (modified from CASANE and LAURENTI, 2013, fig. 1).
Fig. 2. Latimeria chalumnae. Skeleton (bones, cartilages and fin rays) of entire fish based on MILLOT and ANTHONY (1958) and after FOREY (1998). Note i) the monobasal insertion of pectoral, pelvic, anal and second dorsal fins on their respective girdle; ii) the presence of minute neural and haemal arches (respectively arrow-heads and arrows). (Scale = 100 mm).

type according to the definition of MEUNIER et al. (2013). This organization is less complex than the three other plicidentine types (polyplocodont, eusthenodont and dendrodont), which are described in most of extinct and extant Sarcopterygii (SCHULTZE, 1969, 1970).The bony plates of the lung
The lung of Latimeria is reduced to a short oesophagus diverticulum (CUPELLO et al., 2015) surrounded by scattered small and thin ovoid plates (Fig. 9A, B). These plates cover the surface of the vestigial lung, and are composed of a fibrous matrix with fibrocytes: it is a cellular bony tissue (CUPELLO et al., 2017). Each plate is enclosed in a membranous bag. These plates look fragile in their middle plane since a central weakness zone opened on the microtome knife, creating an artefactual lumen (Fig. 9C) (CUPELLO et al., 2017). The fibres are collagenous and organized in superposed layers. The cells are true osteocytes (Fig. 9E) with a more or less star-shaped outline, and send cytoplasmic extensions in the thickness of the plate (Fig. 9D, E). Rare lining cells (osteoblasts) are
present on the surface of the plates, and conjunctive fibres (SHARPEY’s fibres) penetrate within the plate (Fig. 9C). The extracellular matrix shows various staining intensities with concentric and parallel (Fig. 9D) or globular shaped lines. These lines are considered to be Liesegang lines that characterize an active spheritic mineralizing process. The lung plates of Latimeria are thus true bony plates, and are homologous with the large bony plates known in the abdominal cavity of fossil actinistians (BRITO et al., 2010; CUPELLO et al., 2017). The calcified walls that surrounded the lung of the Cretaceous coelacanth Axelrodichthys are made of large osseous plates of various thickness (BRITO et al., 2010), as those of other fossil coelacanths (CLEMENT, 1999, 2005). The bony tissue of these plates is a vascularized cellular bone with more or less large vascular cavities and some internal remodelling (BRITO et al., 2010). Moreover the mineralization of the lung ossified plates in Axelrodichthys is spheritic as in Latimeria’s plates (CUPELLO et al., 2017). So there is a continuity of the histological structure of the lung plates in the coelacanths during their
evolution, but with an important reduction of the bony plates in Latimeria linked to the vestigial state of its lung (CUPELLO et al., 2015, 2019).
CONCLUSION
This overview of eighty years (1938–2018) of histological work on Latimeria skeletal tissues allows some anatomical and evolutionary considerations. It can be enlightened a drastic reduction of endochondral ossification during the long evolutionary history of coelacanths. The persistence of large volume of cartilage in the endoskeleton at adult stage can be compared to the “little bone and considerable cartilage” that characterize the skeleton of a number of demersal notothenioid telostean fishes (EASTMAN et al., 2014).
A processus of spheritic mineralization has been recently highlighted in various skeletal elements of Latimeria, by the presence of globular dentine in teeth, in odontodes of the tegumentary skeleton (scales, fin rays), in scales at the interface between the external layer and the basal plate, as well as in lung bony plates. The spheritic mineralization (i.e., a radiating arrangement of hydroxyapatite crystals and of the organic matrix) in vertebrate skeletal tissues is considered as a precursor of inotropic mineralization (specific interactions between collagen fibrils and the mineral phase) that possibly represents a derived evolutionary stage of calcification mechanisms (ØRVIG, 1951, 1968; FRANCILLON-VIEILLOT et al., 1990; ZYLBERBERG et al., 1992). The simultaneous presence of both mineralization processes (spheritic and inotropic) in the mineralized tissues of Latimeria and in other osteichthyans such as teleosteans (ZYLBERBERG and MEUNIER, 2008) is however questionable. The mineralized spherules present at the limit between the external layer and the basal plate of Latimeria scales are considered to be the product of an inotropic mineralization (MEUNIER and ZYLBERBERG, 1999). It thus appears that the state of the mineralized spherules observed in the various skeletal tissues of Latimeria, including the lung bony plates, must be studied with adapted ultrastructural techniques in order to test their true origin: spheritic or inotropic mineralization?
ACKNOWLEDGEMENTS
This contribution is linked to an international roundtable on coelacanths evolution held at the Kitakyushu Museum and at the Aquamarine Fukushima in August 2017. We would like to thank the anonymous reviewers for their constructive comments and valuable suggestions. C.C. was partially financed by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brasil (CAPES) – Finance Code 001 (Programa Nacional de Pós Doutorado-PNPD).
38 The skeleton and the mineralized tissues of the living coelacanths 39
INTRODUCTION
The first study of fossil coelacanths was done by AGASSIZ (1833–44) and was followed by a lot of papers with the discovery of new fossils (see reviews of JANVIER, 1996 and FOREY, 1998). Fossil coelacanths constitute an important monophyletic group composed of more than forty genera, eighty species, and belong to the Sarcopterygii (ANDREWS, 1973; JANVIER, 1996; FOREY, 1998; UYENO and YABUMOTO, 2007; and many others). For a century coelacanths were considered extinct since the end of the Cretaceous time (Fig. 1).
With the discovery of a living coelacanth in 1938 (SMITH, 1939) the scientific community was soon highly interested to compare the incomplete fossil sarcopterygian fishes dataset to the recent anatomical and biological organization of Latimeria chalumnae. Many authors (see THOMSON, 1969; MILLOT and ANTHONY, 1958; MILLOT et al., 1978 FOREY, 1984, 1998; JANVIER, 1996) focused their work on Latimeria as a key-taxon to test or propose various hypotheses on the origin and evolutionary history of the early tetrapods and sarcopterygii in general, and to infer their possible biological characteristics. The fossil sarcopterygian fishes are known by their fossilized mineralized tissues, essentially their bones, scales and teeth. Together with the extant lungfishes, especially Neoceratodus, Latimeria offered a unique access to the skeleton as a whole: with both mineralized and unmineralized skeletal tissues. Here, we aim to present an overview of the last eighty years (1938–2018) of histological studies of Latimeria’s skeleton.
Since the discovery of the Raja Laut (“king of the sea” ), off Sulawesi Island in Indonesia (ERDMANN et al., 1998, 1999), the genus Latimeria comprises two species: L. chalumnae SMITH 1939 and L. menadoensis POUYAUD et al., 1999. However our knowledge of the anatomy and biology of the living coelacanths is so far essentially based on L. chalumnae.
THE LATIMERIA SKELETON
The first anatomical observations on the extant coelacanth were done by SMITH (1940). The anatomy of the Latimeria skeleton has been later described in detail by MILLOT and ANTHONY (1958). The living coelacanth shows a general organization similar to that of the other actinistian fishes (THOMSON, 1969; JANVIER, 1996; FOREY, 1998). The skeleton can be divided in various parts (Fig. 2): skull, axial skeleton, paired and unpaired fins, tegumentary skeleton (scales). The skull, axial skeleton and fins are constituted of endoskeletal elements that are overlain by exoskeletal elements. In the extant Latimeria the endoskeleton of the skull shows an important regression of the bones, especial ly in the neurocranium, which are replaced by cartilaginous tissues, as in its Mesozoic fossil actinistian relatives (ROMER, 1937, 1942; MILLOT and ANTHONY, 1958; BJERRING, 1973; FOREY, 1998). In the same way, the endoskeleton of paired and unpaired fins is essentially constituted of four axial cartilaginous elements (Fig. 2; MILLOT and ANTHONY, 1958), with pre and post-axial
elements and eventually minute superficial endochondral ossification (CASTANET et al., 1975). The axial skeleton is composed of the notochord which is coated by an unmineralized fibrillary sheath and is totally deprived of well developed vertebrae due to the lack of ossified centra, although some discrete neural and haemal arches develop in the anterior part of the notochord (MILLOT and ANTHONY, 1958). The anterior neural and haemal spines are relatively short but they progressively increase in length posteriorly. Importantly neural and haemal spines are composed of perichondral bone surrounding a cartilaginous core (Fig. 2).
The scales belong to the exoskeleton. In both extant coelacanth species they are of elasmoid type, composed of an upper external layer also called “external ornamented layer”, and of a lower thicker layer, called the basal plate, which is stratified and almost totally unmineralized (Fig. 3). The upper external layer is ornamented with radial crests. In the posterior area of the scale these radial reliefs are overlain by numerous denticulations, the odontodes, which can be superposed (Fig. 3). The anterior area of the external layer, which is covered by the anterior scales, shows crests with irregularities that have been interpreted as growth marks and tentatively used for ageing coelacanths (HUREAU and OZOUF, 1977; MILLER, 1979).
The tooth plates of the buccal cavity support series of teeth of various sizes (Fig. 4). They range in three morphotypes: the fangs (7–10 mm in height), middle-sized teeth (3–4 mm in height) and rounded tubercles (MILLOT and ANTHONY, 1958). The teeth of the two first categories are conical and sharp whereas those of the third category have an obtuse tip (Fig. 5). Fangs of Latimeria are inserted into a socket (MILLOT and ANTHONY, 1958; HOBDELL and MILER, 1969; CASTANET et al., 1975) and they have a smooth external surface (MILLOT et al., 1978; MEUNIER et al., 2015).
A particularity of the exoskeleton of Latimeria is the abundance of odontodes at the surface of various skull bones
Fig. 3. Latimeria chalumnae and L. menadoensis. Details of the posterior field of scales (SEM). A, L. chalumnae. General view of the odontodes. The posterior margin of the scale is on the upper left. (scale = 500 µm). B, L. menadoensis. General view of the odontodes. The posterior margin of the scale is on the upper right. (scale = 500 µm). C, L. menadoensis. Closed-up view of the odontodes positioned on the radial crests of the external layer. (scale = 100 µm). D, L. chalumnae. Closed-up view of the odontodes positioned on the radial crests of the external layer, and two superimposed odontodes are seen on the right (*). (scale = 100 µm). E, L. chalumnae. Closed-up view of two superimposed odontodes with the apical part of the lower one being eroded (arrow). (scale = 100 µm). (After MEUNIER et al., 2008).
Fig. 4. Latimeria chalumnae. Buccal roof showing the different tooth types: fangs (black arrow-heads), small caniniform teeth (white arrow-heads) and patches of round shaped teeth (black arrows), especially on the parasphenoid tooth plate (t.par) and the medial side of the pterygoid (t.pt). The front side is on the top of the figure. (After MILLOT and ANTHONY, 1958). (scale = 10 mm).
(MILLOT and ANTHONY, 1958; BERNHAUSER, 1961), on the fin rays (CASTANET et al., 1975) and on the posterior (free) area of scales (Fig. 3) (MILLOT and ANTHONY, 1958; CASTANET et al., 1975; HADIATY and RACHNATIKA, 2003; MEUNIER et al., 2008).
Among the various components of the whole skeleton, and apart the SMITH’s pioneer work (SMITH, 1940), numerous studies have been carried out on the histological organization of scales (ROUX, 1942; SMITH et al., 1972; CASTANET et al., 1975; GIRAUD et al., 1978a; MILLER, 1979; SMITH, 1979; MEUNIER, 1980; MEUNIER and ZYLBERBERG, 1999; HADIATY and RACHMATIKA, 2003; MEUNIER et al., 2008) and teeth (MILLER and HOBDELL, 1968; GRADY, 1970; HOBDELL and MILLER, 1969; CASTANET et al., 1975; SHELLIS and POOLE, 1978;
SMITH, 1978; SASAGAWA et al., 1984; MEUNIER et al., 2015). Contrary to the superficial skeleton, there are clearly less histological studies of the skeletal bony elements (FRANCILLON et al., 1975). Meanwhile, in addition to the usual skeletal elements, specific bony tissues have been described in fossil (BRITO et al., 2010) and extant coelacanths (CUPELLO et al., 2015, 2017), such as the mineralized plates surrounding the lung. These bony plates in actinistians do not belong to the skeleton sensu stricto. They are specific bony specializations in relation to the lung (see below), as is the “rocker bone” in relation to the gas bladder of some ophidiiform teleosts (PARMENTIER et al., 2008). Despite the name, the rocker bone is not true bony tissue (PARMENTIER et al., 2008), contrary to
the bony plates of fossil and extant coelacanths (BRITO et al., 2010; CUPELLO et al., 2017).
ADULT LATIMERIA SKELETAL TISSUES
Bony tissuesCortical areas of the dermal skeleton are constituted of
primary periosteal bony t issue. I ts mineralization is heterogeneous and generally marked by series of growth lines, probably due to alternative physiological cycles linked to seasonal variations (Fig. 6). The cortical primary bone shows vascular canals and numerous SHARPEY’s fibres (Fig. 6B). The centre of these bones is frequently made of a spongiosa with vascular cavities surrounded by secondary bony deposits that result from remodelling processes (Fig. 6). This secondary bone is separated from the primary one by reversal cementing lines that are hypermineralized.
Bone remodelling in adult specimens can be more or less developed according to a bone and within a given bone. Primary bone is destroyed by osteoclasts and the deposition process of the secondary bone results from the activity of osteoblasts that have replaced the osteoclasts. The secondary bone does generally not replaced the whole primary bone areas, which can still be recognizable by the presence of
SHARPEY’s fibres and/or growth marks (Fig. 6).The dermal fin rays that sustain paired and unpaired fins
in Latimeria are true lepidotrichia (CASTANET et al., 1975). They are made of two parallel opposite gutter-shaped hemirays, each of which being a series of hemisegments articulated by a collagenous ligament. Each hemisegment is made of a mineralized collagenous fibrillary matrix with embedded osteocytes. The hemisegments are sometimes fused at the basal part of the rays due to centrifugal deposition of bony laminae around the primary articulation, and eventually to the mineralization of the ligament (see figs. 19, 20 in CASTANET et al., 1975). Odontodes observed on the external convex surface of the rays are similar to those present on the dermal bones of the skull and scales. The distal extremity of the lepidotrichia is overlapped by actinotrichia (GÉRAUDIE and MEUNIER, 1980) that are long tapered rods of elastoidin , a fibrous protein of collagenous nature (FAURÉ-FREMIET, 1936; GARRAULT, 1936). The presence of lepidotrichia with distal actinotrichia in Latimeria, as in the fins of Actinopterygii, is
regarded as a plesiomorphic character for Osteichthyes (GÉRAUDIE and MEUNIER, 1980, 1984).
The upper layer of the Latimeria scale is relatively thin and it is constituted of bony tissue with embedded osteocytes, whereas the basal plate is much thicker and composed of numerous strata made of thick collagenous fibres (Fig. 7). Between two collagenous layers there are star-shaped cells (SMITH et al., 1972, pl. VI; CASTANET et al., 1975, fig. 12), the elasmocytes, which cytoplasmic processes insert between the collagenous fibres. These fibres are set in a very specific network. In each layer, the fibres are parallel to each other and the direction of the fibres change from a layer to another one. This regular organization results in a spatial arrangement termed “twisted plywood” (GIRAUD et al., 1978a,b). The rotation of fibres direction from one layer to the next has a mean angle of 27° (GIRAUD et al., 1978a). This organization is found in each scale of L. chalumnae. In L. menadoensis, the basal plate of the scales is also a twisted plywood but its rotation angle seems to be slightly less regular (MEUNIER et al.,
2008). The basal plate in both species is unmineralized (Fig. 7) excepted at the contact between the superficial layer and the basal plate, where spheritic mineralized granules are seen in the very first layers of the basal plate (MEUNIER, 1980; MEUNIER and ZYLBERBERG, 1999; MEUNIER et al., 2008). There is no pore-canal system so Latimeria’s scales are not cosmoid, contrary to most of palaeozoic sarcopterygian fishes. Due to the presence of an unmineralized stratified basal plate, the scales of Latimeria are defined as elasmoid-like scales. However the plywood-like organization is considered to be homologous with the bony basal plate of cosmoid scales of extinct sarcopterygian fishes (MEUNIER, 1980; SIRE and HUYSSEUNE, 2003; MONDEJAR, 2018; SCHULTZE, 2018).Cartilages
The cartilaginous tissues in Latimeria are characterized by long chondrocytes (Fig. 8) contrary to those of teleostean fishes that are relatively spherical in shape (MEUNIER, 1979; ZYLBERBERG and MEUNIER, 2008). Cartilage tissues can show an endochondral ossification process. This phenomenon has been studied respectively on the urohyal (PEGUETA, 1968), the MECKEL’s cartilage and the proximal piece of the pectoral girdle (FRANCILLON et al., 1975). The endochondral ossification is relatively limited when it occurs and the volume of enchondral bone remains reduced to thin bony layers (Fig. 8). The cartilage is destroyed by chondroclasts in areas where endochondral ossification occurs. Serial chondrocytes, known at the origin of the calcified cartilage, seem to be lacking in the endochondral ossification process in Latimeria (FRANCILLON et al., 1975). This constitutes a true difference with teleostean endochondral ossifications (MEUNIER, 1979; ZYLBERBERG and MEUNIER, 2008). Tooth tissues
The three tooth morphotypes described in Latimeria are fangs (7–10 mm in height), middle-sized teeth (3–4 mm in height) and rounded tubercles (MILLOT and ANTHONY, 1958). These teeth and tubercles are constituted of a cone of orthodentine set around a large pulp cavity and overlain by an external hypermineralized layer considered as true enamel (GRADY, 1970; CASTANET et al., 1975; SHELLIS and POOLE, 1978; SMITH, 1978; SASAGAWA et al., 1984). Various studies have revealed the presence of globular dentine (Fig. 5) at the periphery of the first third of the tooth (MILLER and HOBDELL, 1968; HOBDELL and MILLER, 1969; SHELLIS and POOLE, 1978; SASAGAWA et al., 1984). This typical histological organization characterizes the various teeth of the bucco-pharyngeal cavity as well as the various odontodes of the dermal bony plates, the fin rays and the scales (CASTANET et al., 1975).
In the fang the inner wall of the pulp cavity displays a series of plies (Fig. 5; MEUNIER et al., 2015). These plies start at the base of the tooth and reach at least the first third to the half of the tooth towards its tip (Fig. 5). Such plies are also present but less developed in the small caniniform teeth (MEUNIER et al., 2015). These dentine plies in the pulp cavity characterizes a plicidentine organization of orthodentine type. These primary plies correspond to a simplexodont plicidentine
REFERENCES
AGASSIZ, L. 1833–44. Recherches sur les Poissons fossiles. Imprimerie Petitpierre, Neuchâtel. [5 volumes and atlas.]
ANDREWS, S. M. 1973. Interrelationships of crossopterygians. In: GREENWOOD, P. H., MILES, R. S. and PATTERSON, C. (eds.), Interrelationships of Fishes, Acad. Press, London, pp. 137–177.
BERNHAUSER, A. 1961. Zur Knochen- und Zah, histology von Latimeria chalumnae Smith und einiger Fossilformen. Sber. öst. Akad. Wiss., 170: 119–137.
BJERRING, H. C. 1973. Relationships of coelacanthiforms. In: GREENWOOD P. H., MILES R. S. and PATTERSON C. (eds.), Interrelationships of Fishes, Acad. Press, London, pp. 179–205.
BRITO, P. M., MEUNIER, F. J., CLÉMENT, G. and GEFFARD-KURIYAMA, D. 2010. The histological structure of the calcified lung of the fossil coelacanth Axelrodichthys araripensis (Actinistia: Mawsoniidae). Palaeontology, 53: 1281–1290.
CASANE, D. and LAURENTI, P. 2013. Why coelacanths are not ‘living fossils’. Bioessays, 35: 332–338. DOI 10.1002/bies.201200145.
CASTANET, J., MEUNIER, F., BERGOT, C. and FRANÇOIS, Y. 1975. Données pré l iminaires sur les s t ructures histologiques du squelette de Latimeria chalumnae. I - Dents, écailles, rayons de nageoires. In: Coll. Inter. CNRS, "Problèmes actuels de Paléontologie. Evolution des Vertébrés", Paris, 4–9 Juin 1973, 159–168, 4 Pls.
CLEMENT, G. 1999. The actinistian (Sarcopterygii) Piveteauia madagascarensis LEHMAN from the Lower Triassic of Northeastern Madagascar: a redescription on the basis of new material. Journal of Vertebrate Paleontology, 19: 234–242.
CLEMENT, G. 2005. A new coelacanth (Actinist ia, Sarcopterygii) from the Jurassic of France, and the question of the closest relative fossil to Latimeria. Journal of Vertebrate Paleontology, 25: 481–491.
CUPELLO, C., BRITO, P. M., MEUNIER, F. J., HERBIN, M., JANVIER, P., DUTEL, H. & CLÉMENT, G. 2015. Allometric growth in the extant coelacanth lung during ontogenetic development. Nature Communications, 6: 8222. DOI: 10.1038/ncomms9222.
CUPELLO, C., CLEMÉNT, G., MEUNIER, F. J., HERBIN, M., YABUMOTO, Y. and BRITO, P. M. 2019. The long-time adaptation of coelacanths to moderate deep water: reviewing the evidences. Bulletin of Kitakyushu Museum of Natural History and Human History Series A (Natural History), 17: 29–35.
CUPELLO, C., MEUNIER, F. J., HERBIN, M., JANVIER, P., CLÉMENT, G. and BRITO, P. M. 2017. The homology and funct ion of the lung pla tes in extant and foss i l coelacanths. Scientific Reports , 7 : 9244. DOI: 10.1038/s41598-017-09327-6.
EASTMAN, J. T., WITMER, L. M., RIDGELY, R. C. and KUHN, K.
L. 2014. Divergence in skeleta l mass and bone morphology in antarctic notothenioid fishes. Journal of Morphology, 275: 841–861. DOI: 10.1002/jmor.20258
ERDMANN, M. V., CALDWELL, R. L. and MOOSA, M. K. 1998. Indonesian “king of the sea” discovered. Nature, 395: 335.
ERDMANN, M. V., CALDWELL, R. L., JEWETT, S. L. and TJAKRAWIDJAJA, A. 1999. The second recorded living coelacanth from north Sulawesi. Environmental Biology of Fishes, 34: 445–451.
FAURÉ-FREMIET, M. 1936. La structure des fibres d’éastoidine. Archives d'Anatomie Microscopique, 32: 249–269.
FOREY, P. L.- 1984. The coelacanth as a living fossil. In: ELREDGE N. and STANLEY S.M. (eds.), Living fossils. Springer Verlag, Berlin, pp. 166–169.
FOREY, P. L.- 1998. History of the Coelacanth Fishes. Chapman and Hall, London, 419 pp.
FRANCILLON, H., MEUNIER, F., NGO TUAN PHONG, D. and RICQLÈS, A. (DE). 1975. Données préliminaires sur les structures histologiques du squelette de Latimeria chalumnae. I I- Tissus osseux et cartilages. In: Coll. Inter. CNRS, "Problèmes actuels de Paléontologie. Evolution des Vertébrés", Paris, 4–9 Juin 1973, Centre National de la Recherche Scientifique, pp. 169–174, 2 pls.
FRANCILLON-VIEILLOT H., BUFFRENIL V. DE, CASTANET J., GÉRAUDIE J., MEUNIER FJ., SIRE J-Y., ZYLBERBERG L., a n d R I C Q L È S A . D E 1 9 9 0 . M i c r o s t r u c t u r e a n d mineralization of vertebrate skeletal tissues. In: CARTER, J. G. (ed.), Skeletal Biomineralization: Patterns, Processes and Evolutionary Trends. Vol. I , Van Nostrand, New York, pp. 471–530
GARRAULT, H., 1936. Développement des fibres d’élastoidine (actinotrichia) chez les salmonides. Archives d'Anatomie Microscopique, 130: 105–137.
GÉRAUDIE, J. and MEUNIER, F. J. 1980. Elastoidin actinotrichia in coelacanth fins: a comparative study with teleosts. Tissue and Cell, 12: 637–645.
GÉRAUDIE, J. and MEUNIER, F. J. 1984. Structure and comparative morphology of camptotrichia of lungfish fins. Tissue and Cell, 16: 217–236.
GIRAUD, M. M., CASTANET, J., MEUNIER, F. J. and BOULIGAND, Y. 1978a. The fibrous structure of coelacanth scales: a twisted "plywood". Tissue and Cell, 10: 671–686.
GIRAUD, M. M., CASTANET, J., MEUNIER, F. J. and BOULIGAND, Y. 1978b. Organisation spatiale de l'isopédine des écailles du coelacanthe (Latimeria chalumnae SMITH). Comptes Rendus de l’Académie des Sciences, 287: 487–489.
GRADY, J . E. 1970. Tooth development in Latimeria chalumnae (SMITH). Journal of Morphology, 132: 377–388.
HADIATY, R. K. and RACHMATIKA, I. 2003. Morphological study of the scales of Latimeria menadoensis POUYAUD et al., 1999. Treubia (A Journal on Zoology of the Indo-Australian Archipelago), 33: 1–11.
HOBDELL, M. H. and MILLER, W. A. 1969. Radiographic
anatomy of the teeth and tooth supporting tissues of Latimeria chalumnae. Archives of Oral Biology, 14: 855–858.
HUREAU J.-Cl. and OZOUF, C. 1977. Détermination de l’âge et croissance du cœlacanthe Latimeria chalumnae SMITH, 1939 (Poisson, Crossoptérygien, Coelacanthidé). Cybium, 2: 129–137.
JANVIER, P. 1996. Early Vertebrates. Clarendon Press, Oxford, 393 pp.
MEUNIER, F. J. 1979. Etude histologique et microradiographique du cartilage hémal de la vertèbre de la carpe, Cyprinus carpio L. (Pisces, Teleostei, Cyprinidae). Acta Zoologica, 60: 19–31.
MEUNIER, F. J. 1980. Les relations isopédine-tissu osseux dans le post-temporal et les écailles de la ligne latérale de Latimeria chalumnae (SMITH). Zoologica Scripta, 9: 307–317.
MEUNIER, F. J. and ZYLBERBERG, L. 1999. The structure of the external layer and of the odontodes of scales in Latimeria chalumnae (Sarcopterygii, Actinistia, Coelacanthidae) revisited using scanning and transmission electron microscopy. In: SÉRET, B. and SIRE, J.-Y. (eds.), Proc. 5th Indo-Pacific Fish Conf., Nouméa, 1997, Soc. Fr. Ichtyol., Paris, pp. 109–116.
MEUNIER, F. J., ERDMANN, M. V., FERMON, Y. and CALDWELL, R. L. 2008. Can the comparative study of the morphology and histology of the scales of Latimeria menadoensis and L. chalumnae (Sarcopterygii, Actinistia, Coelacanthidae) bring new insight on the taxonomy and the biogeography of recent coelacanthids? Geological Society, London, Special Publications, 295: 351–360.
MEUNIER, F. J., BRITO, P. M. and LEAL, M.-E. 2013. Quelques données morphologiques et histologiques sur la structure de la plaque dentaire linguale d’Arapaima gigas (Osteoglossidae; Teleostei). Cybium, 37: 263–271
MEUNIER, F. J., MONDEJAR-FERNANDEZ, J., GOUSSARD, F., CLÉMENT, G. and HERBIN, M. 2015. Presence of plicidentine in the oral teeth of Latimeria chalumnae (Sarcopterygii; Actinistia; Coelacanthidae). Journal of Structural Biology, 190: 31–37.
MILLER, W. A. 1979. Observations on the structure of mineralized tissue of the coelacanth, including scales and their associate odontodes. Occasional Papers of the Californian Academy of Sciences, 134: 68–78.
MILLER, W. A. and HOBDELL, M. H. 1968. Preliminary report on the histology of the dental and paradental tissues of Latimeria chalumnae (SMITH) with a note on tooth replacement. Archives of Oral Biology, 13: 1289–1291.
MILLOT, J. and ANTHONY, J. 1958. Anatomie de Latimeria chalumnae. T. 1. Squelette, muscles et formations de soutien. CNRS Edr., Paris. 122 pp., 80 pls.
MILLOT, J., ANTHONY, J. and ROBINEAU, D. 1978. Anatomie de Latimeria chalumnae. T. 3. Appareil digestif, téguments, écailles. CNRS Edr., Paris. pp. 7–28, 135–139.
MONDEJAR-FERNANDEZ, J. 2018. On cosmine: its origins, b io logy and impl ica t ions for sarcopterygian interrelationships. Cybium, 42: 41–65.
ØRVIG T. 1951. Histologic studies of Placoderms and fossil Elasmobranchs. 1: The endoskeleton, with remarks on the hard tissues of lower vertebrates in general. Arkiv för Zoologi, 2: 321–454, 8 pl.
ØRVIG T. 1968. The dermal skeleton; general considerations. In: ØRVIG T. (ed.), Current problems of lower vertebrate phylogeny. Proceedings of the fourth Nobel Symposium, Stockholm, 1967, J. Wiley, New-York. pp. 373–397.
PARMENTIER, E., COMPÈRE, P., CASADEWALL, M., FONTENELLE, N., CLOOTS, R. and HENRIST, C. 2008. The rocker bone: a new kind of mineralized tissue? Cell and Tissue Research, 334: 67–79.
PEGUETA, V. P. 1968. Enchondral ossification in Latimeria chalumnae SMITH. Dopovidi Akademii Nauk Ukrainia SSR., 7: 653–656 (in Russian; English abstr.).
POUYAUD, L., WIRJOATMODIO, S., RACHMATIKA, L., TJAKRAWIDJAJA, A., HADIATY, R. K. and HADIE, W. 1999. Une nouvelle espèce de coelacanthe, preuves génétiques et morphologiques. Comptes Rendus de l’Académie des Sciences, 322: 261–267.
ROMER, A.S. 1937. The braincase of the carboniferous crossopterygian Megalichthys nitidus. Bulletin of the Museum of Comparative Zoology, 824: 1–73.
ROMER, A.S. 1942. Cartilage as an embryonic adaptation. American Naturalist, 76: 394–404.
ROUX, G. H. 1942. The microscopic anatomy of the Latimeria scale. South African Journal of Medical Sciences, 7: 1–18.
SASAGAWA, I., ISHIYAMA, M. and KODERA, H. 1984. Fine structure of the pharyngeal teeth in the coelacanth fish (Latimeria chalumnae). In: FEARNHEAD, R. W. and SUGA, S. (eds.), Tooth enamel IV, Elsevier Science Publishers, Amsterdam, pp. 462–466.
SCHULTZE, H.-P. 1969. Die Faltenzähne der Rhipidistiden Crossopterygier, der Tetrapoden und der Actinopterygi-er-Gattung Lepisosteus nebst einer Beschreibung der Zahnstruktur von Onychodus (Struniiformer Crossoptery-gier). Palaeontologica Italica, New Series, 35(65): 63–137.
SCHULTZE, H.-P. 1970. Folded teeth and the monophyletic origin of Tetrapods. American Museum Novitates, 2408: 1–10.
SCHULTZE, H.-P. 2018. Hard tissues in fish evolution: history and current issues. Cybium, 42(1): 29–39.
SHELLIS, R. P. and POOLE, D. F. G. 1978. The structure of the dental hard tissues of the coelacanthid fish Latimeria chalumnae SMITH. Archives of Oral Biology, 23: 1105–1113.
SIRE, J.-Y. and HUYSSEUNE, A. 2003. Formation of dermal skeletal and dental tissues in fish: a comparative and evolutionary approach. Biological Review, 78: 219–249.
SMITH, J. L. B. 1939. A living fish of the Mesozoic type. Nature, 143: 455–456.
SMITH, J. L. B. 1940. A living coelacanth fish from South Africa. Transactions of the Royal Society of South Africa, 28: 1–106, 44 Pls.
SMITH, M. M. 1978. Enamel in the oral teeth of Latimeria chalumnae (Pisces: Actinistia): a scanning electron microscopic study. Journal of Zoology, 185: 355–369.
SMITH, M. M. 1979. Scanning electron microscopy of odontodes in the scales of coelacanth embryo, Latimeria chalumnae SMITH. Archives of Oral Biology, 24: 179–183.
SMITH, M. M., HOBDELL, M. H. and MILLER, W. A. 1972. The structure of the scales of Latimeria chalumnae. Journal of Zoology, London, 167: 501–509.
THOMSON, K. S. 1969. The biology of the lobe-finned fishes.
Biological Reviews, 44: 91–154. UYENO, T. and YABUMOTO, Y. 2007. Origin of extant
coelacanths. The Coelacanth, Fathom the Mystery 2007. Aquamarine Fukushima, pp. 24–26.
ZYLBERBERG, L. and MEUNIER, F. J. 2008. New data on the structure and the chondrocyte populations of the haemal cartilage of abdominal vertebrae in the adult carp Cyprinus carpio (Teleostei, Ostariophysii, Cyprinidae). Cybium, 32: 225–239.
ZYLBERBERG, L., GÉRAUDIE, J., MEUNIER, F. J., & SIRE J. Y., 1992. Biomineralization in the integumental skeleton of the living lower Vertebrates. In: HALL, B. K. (ed), Bone, Volume 4, CRC Press, Boca Raton, pp. 171–224.
François J. MEUNIER, Camila CUPELLO and Gaël CLÉMENT
Fig. 1. Schematic phylogeny of nine fossil genera of Actinistia and the extant Latimeria. (modified from CASANE and LAURENTI, 2013, fig. 1).
Fig. 2. Latimeria chalumnae. Skeleton (bones, cartilages and fin rays) of entire fish based on MILLOT and ANTHONY (1958) and after FOREY (1998). Note i) the monobasal insertion of pectoral, pelvic, anal and second dorsal fins on their respective girdle; ii) the presence of minute neural and haemal arches (respectively arrow-heads and arrows). (Scale = 100 mm).

type according to the definition of MEUNIER et al. (2013). This organization is less complex than the three other plicidentine types (polyplocodont, eusthenodont and dendrodont), which are described in most of extinct and extant Sarcopterygii (SCHULTZE, 1969, 1970).The bony plates of the lung
The lung of Latimeria is reduced to a short oesophagus diverticulum (CUPELLO et al., 2015) surrounded by scattered small and thin ovoid plates (Fig. 9A, B). These plates cover the surface of the vestigial lung, and are composed of a fibrous matrix with fibrocytes: it is a cellular bony tissue (CUPELLO et al., 2017). Each plate is enclosed in a membranous bag. These plates look fragile in their middle plane since a central weakness zone opened on the microtome knife, creating an artefactual lumen (Fig. 9C) (CUPELLO et al., 2017). The fibres are collagenous and organized in superposed layers. The cells are true osteocytes (Fig. 9E) with a more or less star-shaped outline, and send cytoplasmic extensions in the thickness of the plate (Fig. 9D, E). Rare lining cells (osteoblasts) are
present on the surface of the plates, and conjunctive fibres (SHARPEY’s fibres) penetrate within the plate (Fig. 9C). The extracellular matrix shows various staining intensities with concentric and parallel (Fig. 9D) or globular shaped lines. These lines are considered to be Liesegang lines that characterize an active spheritic mineralizing process. The lung plates of Latimeria are thus true bony plates, and are homologous with the large bony plates known in the abdominal cavity of fossil actinistians (BRITO et al., 2010; CUPELLO et al., 2017). The calcified walls that surrounded the lung of the Cretaceous coelacanth Axelrodichthys are made of large osseous plates of various thickness (BRITO et al., 2010), as those of other fossil coelacanths (CLEMENT, 1999, 2005). The bony tissue of these plates is a vascularized cellular bone with more or less large vascular cavities and some internal remodelling (BRITO et al., 2010). Moreover the mineralization of the lung ossified plates in Axelrodichthys is spheritic as in Latimeria’s plates (CUPELLO et al., 2017). So there is a continuity of the histological structure of the lung plates in the coelacanths during their
evolution, but with an important reduction of the bony plates in Latimeria linked to the vestigial state of its lung (CUPELLO et al., 2015, 2019).
CONCLUSION
This overview of eighty years (1938–2018) of histological work on Latimeria skeletal tissues allows some anatomical and evolutionary considerations. It can be enlightened a drastic reduction of endochondral ossification during the long evolutionary history of coelacanths. The persistence of large volume of cartilage in the endoskeleton at adult stage can be compared to the “little bone and considerable cartilage” that characterize the skeleton of a number of demersal notothenioid telostean fishes (EASTMAN et al., 2014).
A processus of spheritic mineralization has been recently highlighted in various skeletal elements of Latimeria, by the presence of globular dentine in teeth, in odontodes of the tegumentary skeleton (scales, fin rays), in scales at the interface between the external layer and the basal plate, as well as in lung bony plates. The spheritic mineralization (i.e., a radiating arrangement of hydroxyapatite crystals and of the organic matrix) in vertebrate skeletal tissues is considered as a precursor of inotropic mineralization (specific interactions between collagen fibrils and the mineral phase) that possibly represents a derived evolutionary stage of calcification mechanisms (ØRVIG, 1951, 1968; FRANCILLON-VIEILLOT et al., 1990; ZYLBERBERG et al., 1992). The simultaneous presence of both mineralization processes (spheritic and inotropic) in the mineralized tissues of Latimeria and in other osteichthyans such as teleosteans (ZYLBERBERG and MEUNIER, 2008) is however questionable. The mineralized spherules present at the limit between the external layer and the basal plate of Latimeria scales are considered to be the product of an inotropic mineralization (MEUNIER and ZYLBERBERG, 1999). It thus appears that the state of the mineralized spherules observed in the various skeletal tissues of Latimeria, including the lung bony plates, must be studied with adapted ultrastructural techniques in order to test their true origin: spheritic or inotropic mineralization?
ACKNOWLEDGEMENTS
This contribution is linked to an international roundtable on coelacanths evolution held at the Kitakyushu Museum and at the Aquamarine Fukushima in August 2017. We would like to thank the anonymous reviewers for their constructive comments and valuable suggestions. C.C. was partially financed by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brasil (CAPES) – Finance Code 001 (Programa Nacional de Pós Doutorado-PNPD).
INTRODUCTION
The first study of fossil coelacanths was done by AGASSIZ (1833–44) and was followed by a lot of papers with the discovery of new fossils (see reviews of JANVIER, 1996 and FOREY, 1998). Fossil coelacanths constitute an important monophyletic group composed of more than forty genera, eighty species, and belong to the Sarcopterygii (ANDREWS, 1973; JANVIER, 1996; FOREY, 1998; UYENO and YABUMOTO, 2007; and many others). For a century coelacanths were considered extinct since the end of the Cretaceous time (Fig. 1).
With the discovery of a living coelacanth in 1938 (SMITH, 1939) the scientific community was soon highly interested to compare the incomplete fossil sarcopterygian fishes dataset to the recent anatomical and biological organization of Latimeria chalumnae. Many authors (see THOMSON, 1969; MILLOT and ANTHONY, 1958; MILLOT et al., 1978 FOREY, 1984, 1998; JANVIER, 1996) focused their work on Latimeria as a key-taxon to test or propose various hypotheses on the origin and evolutionary history of the early tetrapods and sarcopterygii in general, and to infer their possible biological characteristics. The fossil sarcopterygian fishes are known by their fossilized mineralized tissues, essentially their bones, scales and teeth. Together with the extant lungfishes, especially Neoceratodus, Latimeria offered a unique access to the skeleton as a whole: with both mineralized and unmineralized skeletal tissues. Here, we aim to present an overview of the last eighty years (1938–2018) of histological studies of Latimeria’s skeleton.
Since the discovery of the Raja Laut (“king of the sea” ), off Sulawesi Island in Indonesia (ERDMANN et al., 1998, 1999), the genus Latimeria comprises two species: L. chalumnae SMITH 1939 and L. menadoensis POUYAUD et al., 1999. However our knowledge of the anatomy and biology of the living coelacanths is so far essentially based on L. chalumnae.
THE LATIMERIA SKELETON
The first anatomical observations on the extant coelacanth were done by SMITH (1940). The anatomy of the Latimeria skeleton has been later described in detail by MILLOT and ANTHONY (1958). The living coelacanth shows a general organization similar to that of the other actinistian fishes (THOMSON, 1969; JANVIER, 1996; FOREY, 1998). The skeleton can be divided in various parts (Fig. 2): skull, axial skeleton, paired and unpaired fins, tegumentary skeleton (scales). The skull, axial skeleton and fins are constituted of endoskeletal elements that are overlain by exoskeletal elements. In the extant Latimeria the endoskeleton of the skull shows an important regression of the bones, especial ly in the neurocranium, which are replaced by cartilaginous tissues, as in its Mesozoic fossil actinistian relatives (ROMER, 1937, 1942; MILLOT and ANTHONY, 1958; BJERRING, 1973; FOREY, 1998). In the same way, the endoskeleton of paired and unpaired fins is essentially constituted of four axial cartilaginous elements (Fig. 2; MILLOT and ANTHONY, 1958), with pre and post-axial
elements and eventually minute superficial endochondral ossification (CASTANET et al., 1975). The axial skeleton is composed of the notochord which is coated by an unmineralized fibrillary sheath and is totally deprived of well developed vertebrae due to the lack of ossified centra, although some discrete neural and haemal arches develop in the anterior part of the notochord (MILLOT and ANTHONY, 1958). The anterior neural and haemal spines are relatively short but they progressively increase in length posteriorly. Importantly neural and haemal spines are composed of perichondral bone surrounding a cartilaginous core (Fig. 2).
The scales belong to the exoskeleton. In both extant coelacanth species they are of elasmoid type, composed of an upper external layer also called “external ornamented layer”, and of a lower thicker layer, called the basal plate, which is stratified and almost totally unmineralized (Fig. 3). The upper external layer is ornamented with radial crests. In the posterior area of the scale these radial reliefs are overlain by numerous denticulations, the odontodes, which can be superposed (Fig. 3). The anterior area of the external layer, which is covered by the anterior scales, shows crests with irregularities that have been interpreted as growth marks and tentatively used for ageing coelacanths (HUREAU and OZOUF, 1977; MILLER, 1979).
The tooth plates of the buccal cavity support series of teeth of various sizes (Fig. 4). They range in three morphotypes: the fangs (7–10 mm in height), middle-sized teeth (3–4 mm in height) and rounded tubercles (MILLOT and ANTHONY, 1958). The teeth of the two first categories are conical and sharp whereas those of the third category have an obtuse tip (Fig. 5). Fangs of Latimeria are inserted into a socket (MILLOT and ANTHONY, 1958; HOBDELL and MILER, 1969; CASTANET et al., 1975) and they have a smooth external surface (MILLOT et al., 1978; MEUNIER et al., 2015).
A particularity of the exoskeleton of Latimeria is the abundance of odontodes at the surface of various skull bones
40 41
Fig. 5. Latimeria chalumnae. Coronoid and coronoid tooth structure. A, Tomographic imaging of a coronoid element. Three-dimensional reconstruction of the dental plate showing a fang, one mid-sized caniniform tooth (*) and numerous small round-shaped teeth (arrow-heads). The virtual section shows i) the plies of the dentine in the lower half of the pulp cavity (pc); ii) the bone of attachment (ba) between the tooth and the coronoid bony tissue. On the right of the figure, the coronoid shows a network of vascular cavities and canals (vc). (de = dentine; en = enamel) (scale = 1 mm). (After MEUNIER et al., 2015, Fig. 9-10). B, Microradiography of a cross section at the base of a fang showing several plies of the dentine wall (white arrow-heads). (bo = bone; pc = pulp cavity; de = dentine). (scale = 1 mm). (from CASTANET et al., 1975). C, Scanning Electron Microscopy of the spheritic dentine. The white and black asterisks point respectively to the external surface of the tooth and to the wall of the pulp cavity. (scale = 25 µm).
(MILLOT and ANTHONY, 1958; BERNHAUSER, 1961), on the fin rays (CASTANET et al., 1975) and on the posterior (free) area of scales (Fig. 3) (MILLOT and ANTHONY, 1958; CASTANET et al., 1975; HADIATY and RACHNATIKA, 2003; MEUNIER et al., 2008).
Among the various components of the whole skeleton, and apart the SMITH’s pioneer work (SMITH, 1940), numerous studies have been carried out on the histological organization of scales (ROUX, 1942; SMITH et al., 1972; CASTANET et al., 1975; GIRAUD et al., 1978a; MILLER, 1979; SMITH, 1979; MEUNIER, 1980; MEUNIER and ZYLBERBERG, 1999; HADIATY and RACHMATIKA, 2003; MEUNIER et al., 2008) and teeth (MILLER and HOBDELL, 1968; GRADY, 1970; HOBDELL and MILLER, 1969; CASTANET et al., 1975; SHELLIS and POOLE, 1978;
SMITH, 1978; SASAGAWA et al., 1984; MEUNIER et al., 2015). Contrary to the superficial skeleton, there are clearly less histological studies of the skeletal bony elements (FRANCILLON et al., 1975). Meanwhile, in addition to the usual skeletal elements, specific bony tissues have been described in fossil (BRITO et al., 2010) and extant coelacanths (CUPELLO et al., 2015, 2017), such as the mineralized plates surrounding the lung. These bony plates in actinistians do not belong to the skeleton sensu stricto. They are specific bony specializations in relation to the lung (see below), as is the “rocker bone” in relation to the gas bladder of some ophidiiform teleosts (PARMENTIER et al., 2008). Despite the name, the rocker bone is not true bony tissue (PARMENTIER et al., 2008), contrary to
the bony plates of fossil and extant coelacanths (BRITO et al., 2010; CUPELLO et al., 2017).
ADULT LATIMERIA SKELETAL TISSUES
Bony tissuesCortical areas of the dermal skeleton are constituted of
primary periosteal bony t issue. I ts mineralization is heterogeneous and generally marked by series of growth lines, probably due to alternative physiological cycles linked to seasonal variations (Fig. 6). The cortical primary bone shows vascular canals and numerous SHARPEY’s fibres (Fig. 6B). The centre of these bones is frequently made of a spongiosa with vascular cavities surrounded by secondary bony deposits that result from remodelling processes (Fig. 6). This secondary bone is separated from the primary one by reversal cementing lines that are hypermineralized.
Bone remodelling in adult specimens can be more or less developed according to a bone and within a given bone. Primary bone is destroyed by osteoclasts and the deposition process of the secondary bone results from the activity of osteoblasts that have replaced the osteoclasts. The secondary bone does generally not replaced the whole primary bone areas, which can still be recognizable by the presence of
SHARPEY’s fibres and/or growth marks (Fig. 6).The dermal fin rays that sustain paired and unpaired fins
in Latimeria are true lepidotrichia (CASTANET et al., 1975). They are made of two parallel opposite gutter-shaped hemirays, each of which being a series of hemisegments articulated by a collagenous ligament. Each hemisegment is made of a mineralized collagenous fibrillary matrix with embedded osteocytes. The hemisegments are sometimes fused at the basal part of the rays due to centrifugal deposition of bony laminae around the primary articulation, and eventually to the mineralization of the ligament (see figs. 19, 20 in CASTANET et al., 1975). Odontodes observed on the external convex surface of the rays are similar to those present on the dermal bones of the skull and scales. The distal extremity of the lepidotrichia is overlapped by actinotrichia (GÉRAUDIE and MEUNIER, 1980) that are long tapered rods of elastoidin , a fibrous protein of collagenous nature (FAURÉ-FREMIET, 1936; GARRAULT, 1936). The presence of lepidotrichia with distal actinotrichia in Latimeria, as in the fins of Actinopterygii, is
regarded as a plesiomorphic character for Osteichthyes (GÉRAUDIE and MEUNIER, 1980, 1984).
The upper layer of the Latimeria scale is relatively thin and it is constituted of bony tissue with embedded osteocytes, whereas the basal plate is much thicker and composed of numerous strata made of thick collagenous fibres (Fig. 7). Between two collagenous layers there are star-shaped cells (SMITH et al., 1972, pl. VI; CASTANET et al., 1975, fig. 12), the elasmocytes, which cytoplasmic processes insert between the collagenous fibres. These fibres are set in a very specific network. In each layer, the fibres are parallel to each other and the direction of the fibres change from a layer to another one. This regular organization results in a spatial arrangement termed “twisted plywood” (GIRAUD et al., 1978a,b). The rotation of fibres direction from one layer to the next has a mean angle of 27° (GIRAUD et al., 1978a). This organization is found in each scale of L. chalumnae. In L. menadoensis, the basal plate of the scales is also a twisted plywood but its rotation angle seems to be slightly less regular (MEUNIER et al.,
2008). The basal plate in both species is unmineralized (Fig. 7) excepted at the contact between the superficial layer and the basal plate, where spheritic mineralized granules are seen in the very first layers of the basal plate (MEUNIER, 1980; MEUNIER and ZYLBERBERG, 1999; MEUNIER et al., 2008). There is no pore-canal system so Latimeria’s scales are not cosmoid, contrary to most of palaeozoic sarcopterygian fishes. Due to the presence of an unmineralized stratified basal plate, the scales of Latimeria are defined as elasmoid-like scales. However the plywood-like organization is considered to be homologous with the bony basal plate of cosmoid scales of extinct sarcopterygian fishes (MEUNIER, 1980; SIRE and HUYSSEUNE, 2003; MONDEJAR, 2018; SCHULTZE, 2018).Cartilages
The cartilaginous tissues in Latimeria are characterized by long chondrocytes (Fig. 8) contrary to those of teleostean fishes that are relatively spherical in shape (MEUNIER, 1979; ZYLBERBERG and MEUNIER, 2008). Cartilage tissues can show an endochondral ossification process. This phenomenon has been studied respectively on the urohyal (PEGUETA, 1968), the MECKEL’s cartilage and the proximal piece of the pectoral girdle (FRANCILLON et al., 1975). The endochondral ossification is relatively limited when it occurs and the volume of enchondral bone remains reduced to thin bony layers (Fig. 8). The cartilage is destroyed by chondroclasts in areas where endochondral ossification occurs. Serial chondrocytes, known at the origin of the calcified cartilage, seem to be lacking in the endochondral ossification process in Latimeria (FRANCILLON et al., 1975). This constitutes a true difference with teleostean endochondral ossifications (MEUNIER, 1979; ZYLBERBERG and MEUNIER, 2008). Tooth tissues
The three tooth morphotypes described in Latimeria are fangs (7–10 mm in height), middle-sized teeth (3–4 mm in height) and rounded tubercles (MILLOT and ANTHONY, 1958). These teeth and tubercles are constituted of a cone of orthodentine set around a large pulp cavity and overlain by an external hypermineralized layer considered as true enamel (GRADY, 1970; CASTANET et al., 1975; SHELLIS and POOLE, 1978; SMITH, 1978; SASAGAWA et al., 1984). Various studies have revealed the presence of globular dentine (Fig. 5) at the periphery of the first third of the tooth (MILLER and HOBDELL, 1968; HOBDELL and MILLER, 1969; SHELLIS and POOLE, 1978; SASAGAWA et al., 1984). This typical histological organization characterizes the various teeth of the bucco-pharyngeal cavity as well as the various odontodes of the dermal bony plates, the fin rays and the scales (CASTANET et al., 1975).
In the fang the inner wall of the pulp cavity displays a series of plies (Fig. 5; MEUNIER et al., 2015). These plies start at the base of the tooth and reach at least the first third to the half of the tooth towards its tip (Fig. 5). Such plies are also present but less developed in the small caniniform teeth (MEUNIER et al., 2015). These dentine plies in the pulp cavity characterizes a plicidentine organization of orthodentine type. These primary plies correspond to a simplexodont plicidentine
REFERENCES
AGASSIZ, L. 1833–44. Recherches sur les Poissons fossiles. Imprimerie Petitpierre, Neuchâtel. [5 volumes and atlas.]
ANDREWS, S. M. 1973. Interrelationships of crossopterygians. In: GREENWOOD, P. H., MILES, R. S. and PATTERSON, C. (eds.), Interrelationships of Fishes, Acad. Press, London, pp. 137–177.
BERNHAUSER, A. 1961. Zur Knochen- und Zah, histology von Latimeria chalumnae Smith und einiger Fossilformen. Sber. öst. Akad. Wiss., 170: 119–137.
BJERRING, H. C. 1973. Relationships of coelacanthiforms. In: GREENWOOD P. H., MILES R. S. and PATTERSON C. (eds.), Interrelationships of Fishes, Acad. Press, London, pp. 179–205.
BRITO, P. M., MEUNIER, F. J., CLÉMENT, G. and GEFFARD-KURIYAMA, D. 2010. The histological structure of the calcified lung of the fossil coelacanth Axelrodichthys araripensis (Actinistia: Mawsoniidae). Palaeontology, 53: 1281–1290.
CASANE, D. and LAURENTI, P. 2013. Why coelacanths are not ‘living fossils’. Bioessays, 35: 332–338. DOI 10.1002/bies.201200145.
CASTANET, J., MEUNIER, F., BERGOT, C. and FRANÇOIS, Y. 1975. Données pré l iminaires sur les s t ructures histologiques du squelette de Latimeria chalumnae. I - Dents, écailles, rayons de nageoires. In: Coll. Inter. CNRS, "Problèmes actuels de Paléontologie. Evolution des Vertébrés", Paris, 4–9 Juin 1973, 159–168, 4 Pls.
CLEMENT, G. 1999. The actinistian (Sarcopterygii) Piveteauia madagascarensis LEHMAN from the Lower Triassic of Northeastern Madagascar: a redescription on the basis of new material. Journal of Vertebrate Paleontology, 19: 234–242.
CLEMENT, G. 2005. A new coelacanth (Actinist ia, Sarcopterygii) from the Jurassic of France, and the question of the closest relative fossil to Latimeria. Journal of Vertebrate Paleontology, 25: 481–491.
CUPELLO, C., BRITO, P. M., MEUNIER, F. J., HERBIN, M., JANVIER, P., DUTEL, H. & CLÉMENT, G. 2015. Allometric growth in the extant coelacanth lung during ontogenetic development. Nature Communications, 6: 8222. DOI: 10.1038/ncomms9222.
CUPELLO, C., CLEMÉNT, G., MEUNIER, F. J., HERBIN, M., YABUMOTO, Y. and BRITO, P. M. 2019. The long-time adaptation of coelacanths to moderate deep water: reviewing the evidences. Bulletin of Kitakyushu Museum of Natural History and Human History Series A (Natural History), 17: 29–35.
CUPELLO, C., MEUNIER, F. J., HERBIN, M., JANVIER, P., CLÉMENT, G. and BRITO, P. M. 2017. The homology and funct ion of the lung pla tes in extant and foss i l coelacanths. Scientific Reports , 7 : 9244. DOI: 10.1038/s41598-017-09327-6.
EASTMAN, J. T., WITMER, L. M., RIDGELY, R. C. and KUHN, K.
L. 2014. Divergence in skeleta l mass and bone morphology in antarctic notothenioid fishes. Journal of Morphology, 275: 841–861. DOI: 10.1002/jmor.20258
ERDMANN, M. V., CALDWELL, R. L. and MOOSA, M. K. 1998. Indonesian “king of the sea” discovered. Nature, 395: 335.
ERDMANN, M. V., CALDWELL, R. L., JEWETT, S. L. and TJAKRAWIDJAJA, A. 1999. The second recorded living coelacanth from north Sulawesi. Environmental Biology of Fishes, 34: 445–451.
FAURÉ-FREMIET, M. 1936. La structure des fibres d’éastoidine. Archives d'Anatomie Microscopique, 32: 249–269.
FOREY, P. L.- 1984. The coelacanth as a living fossil. In: ELREDGE N. and STANLEY S.M. (eds.), Living fossils. Springer Verlag, Berlin, pp. 166–169.
FOREY, P. L.- 1998. History of the Coelacanth Fishes. Chapman and Hall, London, 419 pp.
FRANCILLON, H., MEUNIER, F., NGO TUAN PHONG, D. and RICQLÈS, A. (DE). 1975. Données préliminaires sur les structures histologiques du squelette de Latimeria chalumnae. I I- Tissus osseux et cartilages. In: Coll. Inter. CNRS, "Problèmes actuels de Paléontologie. Evolution des Vertébrés", Paris, 4–9 Juin 1973, Centre National de la Recherche Scientifique, pp. 169–174, 2 pls.
FRANCILLON-VIEILLOT H., BUFFRENIL V. DE, CASTANET J., GÉRAUDIE J., MEUNIER FJ., SIRE J-Y., ZYLBERBERG L., a n d R I C Q L È S A . D E 1 9 9 0 . M i c r o s t r u c t u r e a n d mineralization of vertebrate skeletal tissues. In: CARTER, J. G. (ed.), Skeletal Biomineralization: Patterns, Processes and Evolutionary Trends. Vol. I , Van Nostrand, New York, pp. 471–530
GARRAULT, H., 1936. Développement des fibres d’élastoidine (actinotrichia) chez les salmonides. Archives d'Anatomie Microscopique, 130: 105–137.
GÉRAUDIE, J. and MEUNIER, F. J. 1980. Elastoidin actinotrichia in coelacanth fins: a comparative study with teleosts. Tissue and Cell, 12: 637–645.
GÉRAUDIE, J. and MEUNIER, F. J. 1984. Structure and comparative morphology of camptotrichia of lungfish fins. Tissue and Cell, 16: 217–236.
GIRAUD, M. M., CASTANET, J., MEUNIER, F. J. and BOULIGAND, Y. 1978a. The fibrous structure of coelacanth scales: a twisted "plywood". Tissue and Cell, 10: 671–686.
GIRAUD, M. M., CASTANET, J., MEUNIER, F. J. and BOULIGAND, Y. 1978b. Organisation spatiale de l'isopédine des écailles du coelacanthe (Latimeria chalumnae SMITH). Comptes Rendus de l’Académie des Sciences, 287: 487–489.
GRADY, J . E. 1970. Tooth development in Latimeria chalumnae (SMITH). Journal of Morphology, 132: 377–388.
HADIATY, R. K. and RACHMATIKA, I. 2003. Morphological study of the scales of Latimeria menadoensis POUYAUD et al., 1999. Treubia (A Journal on Zoology of the Indo-Australian Archipelago), 33: 1–11.
HOBDELL, M. H. and MILLER, W. A. 1969. Radiographic
anatomy of the teeth and tooth supporting tissues of Latimeria chalumnae. Archives of Oral Biology, 14: 855–858.
HUREAU J.-Cl. and OZOUF, C. 1977. Détermination de l’âge et croissance du cœlacanthe Latimeria chalumnae SMITH, 1939 (Poisson, Crossoptérygien, Coelacanthidé). Cybium, 2: 129–137.
JANVIER, P. 1996. Early Vertebrates. Clarendon Press, Oxford, 393 pp.
MEUNIER, F. J. 1979. Etude histologique et microradiographique du cartilage hémal de la vertèbre de la carpe, Cyprinus carpio L. (Pisces, Teleostei, Cyprinidae). Acta Zoologica, 60: 19–31.
MEUNIER, F. J. 1980. Les relations isopédine-tissu osseux dans le post-temporal et les écailles de la ligne latérale de Latimeria chalumnae (SMITH). Zoologica Scripta, 9: 307–317.
MEUNIER, F. J. and ZYLBERBERG, L. 1999. The structure of the external layer and of the odontodes of scales in Latimeria chalumnae (Sarcopterygii, Actinistia, Coelacanthidae) revisited using scanning and transmission electron microscopy. In: SÉRET, B. and SIRE, J.-Y. (eds.), Proc. 5th Indo-Pacific Fish Conf., Nouméa, 1997, Soc. Fr. Ichtyol., Paris, pp. 109–116.
MEUNIER, F. J., ERDMANN, M. V., FERMON, Y. and CALDWELL, R. L. 2008. Can the comparative study of the morphology and histology of the scales of Latimeria menadoensis and L. chalumnae (Sarcopterygii, Actinistia, Coelacanthidae) bring new insight on the taxonomy and the biogeography of recent coelacanthids? Geological Society, London, Special Publications, 295: 351–360.
MEUNIER, F. J., BRITO, P. M. and LEAL, M.-E. 2013. Quelques données morphologiques et histologiques sur la structure de la plaque dentaire linguale d’Arapaima gigas (Osteoglossidae; Teleostei). Cybium, 37: 263–271
MEUNIER, F. J., MONDEJAR-FERNANDEZ, J., GOUSSARD, F., CLÉMENT, G. and HERBIN, M. 2015. Presence of plicidentine in the oral teeth of Latimeria chalumnae (Sarcopterygii; Actinistia; Coelacanthidae). Journal of Structural Biology, 190: 31–37.
MILLER, W. A. 1979. Observations on the structure of mineralized tissue of the coelacanth, including scales and their associate odontodes. Occasional Papers of the Californian Academy of Sciences, 134: 68–78.
MILLER, W. A. and HOBDELL, M. H. 1968. Preliminary report on the histology of the dental and paradental tissues of Latimeria chalumnae (SMITH) with a note on tooth replacement. Archives of Oral Biology, 13: 1289–1291.
MILLOT, J. and ANTHONY, J. 1958. Anatomie de Latimeria chalumnae. T. 1. Squelette, muscles et formations de soutien. CNRS Edr., Paris. 122 pp., 80 pls.
MILLOT, J., ANTHONY, J. and ROBINEAU, D. 1978. Anatomie de Latimeria chalumnae. T. 3. Appareil digestif, téguments, écailles. CNRS Edr., Paris. pp. 7–28, 135–139.
MONDEJAR-FERNANDEZ, J. 2018. On cosmine: its origins, b io logy and impl ica t ions for sarcopterygian interrelationships. Cybium, 42: 41–65.
ØRVIG T. 1951. Histologic studies of Placoderms and fossil Elasmobranchs. 1: The endoskeleton, with remarks on the hard tissues of lower vertebrates in general. Arkiv för Zoologi, 2: 321–454, 8 pl.
ØRVIG T. 1968. The dermal skeleton; general considerations. In: ØRVIG T. (ed.), Current problems of lower vertebrate phylogeny. Proceedings of the fourth Nobel Symposium, Stockholm, 1967, J. Wiley, New-York. pp. 373–397.
PARMENTIER, E., COMPÈRE, P., CASADEWALL, M., FONTENELLE, N., CLOOTS, R. and HENRIST, C. 2008. The rocker bone: a new kind of mineralized tissue? Cell and Tissue Research, 334: 67–79.
PEGUETA, V. P. 1968. Enchondral ossification in Latimeria chalumnae SMITH. Dopovidi Akademii Nauk Ukrainia SSR., 7: 653–656 (in Russian; English abstr.).
POUYAUD, L., WIRJOATMODIO, S., RACHMATIKA, L., TJAKRAWIDJAJA, A., HADIATY, R. K. and HADIE, W. 1999. Une nouvelle espèce de coelacanthe, preuves génétiques et morphologiques. Comptes Rendus de l’Académie des Sciences, 322: 261–267.
ROMER, A.S. 1937. The braincase of the carboniferous crossopterygian Megalichthys nitidus. Bulletin of the Museum of Comparative Zoology, 824: 1–73.
ROMER, A.S. 1942. Cartilage as an embryonic adaptation. American Naturalist, 76: 394–404.
ROUX, G. H. 1942. The microscopic anatomy of the Latimeria scale. South African Journal of Medical Sciences, 7: 1–18.
SASAGAWA, I., ISHIYAMA, M. and KODERA, H. 1984. Fine structure of the pharyngeal teeth in the coelacanth fish (Latimeria chalumnae). In: FEARNHEAD, R. W. and SUGA, S. (eds.), Tooth enamel IV, Elsevier Science Publishers, Amsterdam, pp. 462–466.
SCHULTZE, H.-P. 1969. Die Faltenzähne der Rhipidistiden Crossopterygier, der Tetrapoden und der Actinopterygi-er-Gattung Lepisosteus nebst einer Beschreibung der Zahnstruktur von Onychodus (Struniiformer Crossoptery-gier). Palaeontologica Italica, New Series, 35(65): 63–137.
SCHULTZE, H.-P. 1970. Folded teeth and the monophyletic origin of Tetrapods. American Museum Novitates, 2408: 1–10.
SCHULTZE, H.-P. 2018. Hard tissues in fish evolution: history and current issues. Cybium, 42(1): 29–39.
SHELLIS, R. P. and POOLE, D. F. G. 1978. The structure of the dental hard tissues of the coelacanthid fish Latimeria chalumnae SMITH. Archives of Oral Biology, 23: 1105–1113.
SIRE, J.-Y. and HUYSSEUNE, A. 2003. Formation of dermal skeletal and dental tissues in fish: a comparative and evolutionary approach. Biological Review, 78: 219–249.
SMITH, J. L. B. 1939. A living fish of the Mesozoic type. Nature, 143: 455–456.
SMITH, J. L. B. 1940. A living coelacanth fish from South Africa. Transactions of the Royal Society of South Africa, 28: 1–106, 44 Pls.
SMITH, M. M. 1978. Enamel in the oral teeth of Latimeria chalumnae (Pisces: Actinistia): a scanning electron microscopic study. Journal of Zoology, 185: 355–369.
SMITH, M. M. 1979. Scanning electron microscopy of odontodes in the scales of coelacanth embryo, Latimeria chalumnae SMITH. Archives of Oral Biology, 24: 179–183.
SMITH, M. M., HOBDELL, M. H. and MILLER, W. A. 1972. The structure of the scales of Latimeria chalumnae. Journal of Zoology, London, 167: 501–509.
THOMSON, K. S. 1969. The biology of the lobe-finned fishes.
Biological Reviews, 44: 91–154. UYENO, T. and YABUMOTO, Y. 2007. Origin of extant
coelacanths. The Coelacanth, Fathom the Mystery 2007. Aquamarine Fukushima, pp. 24–26.
ZYLBERBERG, L. and MEUNIER, F. J. 2008. New data on the structure and the chondrocyte populations of the haemal cartilage of abdominal vertebrae in the adult carp Cyprinus carpio (Teleostei, Ostariophysii, Cyprinidae). Cybium, 32: 225–239.
ZYLBERBERG, L., GÉRAUDIE, J., MEUNIER, F. J., & SIRE J. Y., 1992. Biomineralization in the integumental skeleton of the living lower Vertebrates. In: HALL, B. K. (ed), Bone, Volume 4, CRC Press, Boca Raton, pp. 171–224.
The skeleton and the mineralized tissues of the living coelacanthsFrançois J. MEUNIER, Camila CUPELLO and Gaël CLÉMENT
Fig. 6. Latimeria chalumnae. Bone histology. A, Thin section in a coronoid bone (Basic fuchsin staining) showing primary bone (pb) with osteocytes, secondary bone (sb), a cementing line (arrow-heads) and a vascular cavity (vc). (scale = 50 µm). Inset: detail of osteocytes (arrow-heads) and their osteocytic canalicles. (scale = 25 µm). B, Thin section in a coronoid bone (Hemalun-PIC staining) showing cellular primary bone (pb) and its periostic membrane (white arrows). On the right, numerous collagenous fibres (cf) enter the bony matrix where they form SHARPEY’s fibres. The asterisks points to secondary vascular cavities limited by cementing lines (white arrow-heads); a primary vascular canal (vc) is seen. (scale = 100 µm). C, X-ray ground section in the cortical region of the angular showing primary bone with regular alternative of heterogeneous mineralized bony tissue. The black arrow-heads point to primary vascular canals, the white arrows to cement ing l ines , and the white arrow-heads to erosive bays. The asterisks point to two confluent secondary vascular cavities). (scale = 200 µm). D, Ground section of the angular, in a deeper region than C. Several bulks of secondary bone (sb) are limited by cementing lines (black arrow-heads). Primary bone (pb) persists in different areas. The white asterisks point to secondary vascular cavities. (scale = 200 µm). (C, D: After FRANCILLON et al., 1975).

type according to the definition of MEUNIER et al. (2013). This organization is less complex than the three other plicidentine types (polyplocodont, eusthenodont and dendrodont), which are described in most of extinct and extant Sarcopterygii (SCHULTZE, 1969, 1970).The bony plates of the lung
The lung of Latimeria is reduced to a short oesophagus diverticulum (CUPELLO et al., 2015) surrounded by scattered small and thin ovoid plates (Fig. 9A, B). These plates cover the surface of the vestigial lung, and are composed of a fibrous matrix with fibrocytes: it is a cellular bony tissue (CUPELLO et al., 2017). Each plate is enclosed in a membranous bag. These plates look fragile in their middle plane since a central weakness zone opened on the microtome knife, creating an artefactual lumen (Fig. 9C) (CUPELLO et al., 2017). The fibres are collagenous and organized in superposed layers. The cells are true osteocytes (Fig. 9E) with a more or less star-shaped outline, and send cytoplasmic extensions in the thickness of the plate (Fig. 9D, E). Rare lining cells (osteoblasts) are
present on the surface of the plates, and conjunctive fibres (SHARPEY’s fibres) penetrate within the plate (Fig. 9C). The extracellular matrix shows various staining intensities with concentric and parallel (Fig. 9D) or globular shaped lines. These lines are considered to be Liesegang lines that characterize an active spheritic mineralizing process. The lung plates of Latimeria are thus true bony plates, and are homologous with the large bony plates known in the abdominal cavity of fossil actinistians (BRITO et al., 2010; CUPELLO et al., 2017). The calcified walls that surrounded the lung of the Cretaceous coelacanth Axelrodichthys are made of large osseous plates of various thickness (BRITO et al., 2010), as those of other fossil coelacanths (CLEMENT, 1999, 2005). The bony tissue of these plates is a vascularized cellular bone with more or less large vascular cavities and some internal remodelling (BRITO et al., 2010). Moreover the mineralization of the lung ossified plates in Axelrodichthys is spheritic as in Latimeria’s plates (CUPELLO et al., 2017). So there is a continuity of the histological structure of the lung plates in the coelacanths during their
evolution, but with an important reduction of the bony plates in Latimeria linked to the vestigial state of its lung (CUPELLO et al., 2015, 2019).
CONCLUSION
This overview of eighty years (1938–2018) of histological work on Latimeria skeletal tissues allows some anatomical and evolutionary considerations. It can be enlightened a drastic reduction of endochondral ossification during the long evolutionary history of coelacanths. The persistence of large volume of cartilage in the endoskeleton at adult stage can be compared to the “little bone and considerable cartilage” that characterize the skeleton of a number of demersal notothenioid telostean fishes (EASTMAN et al., 2014).
A processus of spheritic mineralization has been recently highlighted in various skeletal elements of Latimeria, by the presence of globular dentine in teeth, in odontodes of the tegumentary skeleton (scales, fin rays), in scales at the interface between the external layer and the basal plate, as well as in lung bony plates. The spheritic mineralization (i.e., a radiating arrangement of hydroxyapatite crystals and of the organic matrix) in vertebrate skeletal tissues is considered as a precursor of inotropic mineralization (specific interactions between collagen fibrils and the mineral phase) that possibly represents a derived evolutionary stage of calcification mechanisms (ØRVIG, 1951, 1968; FRANCILLON-VIEILLOT et al., 1990; ZYLBERBERG et al., 1992). The simultaneous presence of both mineralization processes (spheritic and inotropic) in the mineralized tissues of Latimeria and in other osteichthyans such as teleosteans (ZYLBERBERG and MEUNIER, 2008) is however questionable. The mineralized spherules present at the limit between the external layer and the basal plate of Latimeria scales are considered to be the product of an inotropic mineralization (MEUNIER and ZYLBERBERG, 1999). It thus appears that the state of the mineralized spherules observed in the various skeletal tissues of Latimeria, including the lung bony plates, must be studied with adapted ultrastructural techniques in order to test their true origin: spheritic or inotropic mineralization?
ACKNOWLEDGEMENTS
This contribution is linked to an international roundtable on coelacanths evolution held at the Kitakyushu Museum and at the Aquamarine Fukushima in August 2017. We would like to thank the anonymous reviewers for their constructive comments and valuable suggestions. C.C. was partially financed by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brasil (CAPES) – Finance Code 001 (Programa Nacional de Pós Doutorado-PNPD).
INTRODUCTION
The first study of fossil coelacanths was done by AGASSIZ (1833–44) and was followed by a lot of papers with the discovery of new fossils (see reviews of JANVIER, 1996 and FOREY, 1998). Fossil coelacanths constitute an important monophyletic group composed of more than forty genera, eighty species, and belong to the Sarcopterygii (ANDREWS, 1973; JANVIER, 1996; FOREY, 1998; UYENO and YABUMOTO, 2007; and many others). For a century coelacanths were considered extinct since the end of the Cretaceous time (Fig. 1).
With the discovery of a living coelacanth in 1938 (SMITH, 1939) the scientific community was soon highly interested to compare the incomplete fossil sarcopterygian fishes dataset to the recent anatomical and biological organization of Latimeria chalumnae. Many authors (see THOMSON, 1969; MILLOT and ANTHONY, 1958; MILLOT et al., 1978 FOREY, 1984, 1998; JANVIER, 1996) focused their work on Latimeria as a key-taxon to test or propose various hypotheses on the origin and evolutionary history of the early tetrapods and sarcopterygii in general, and to infer their possible biological characteristics. The fossil sarcopterygian fishes are known by their fossilized mineralized tissues, essentially their bones, scales and teeth. Together with the extant lungfishes, especially Neoceratodus, Latimeria offered a unique access to the skeleton as a whole: with both mineralized and unmineralized skeletal tissues. Here, we aim to present an overview of the last eighty years (1938–2018) of histological studies of Latimeria’s skeleton.
Since the discovery of the Raja Laut (“king of the sea” ), off Sulawesi Island in Indonesia (ERDMANN et al., 1998, 1999), the genus Latimeria comprises two species: L. chalumnae SMITH 1939 and L. menadoensis POUYAUD et al., 1999. However our knowledge of the anatomy and biology of the living coelacanths is so far essentially based on L. chalumnae.
THE LATIMERIA SKELETON
The first anatomical observations on the extant coelacanth were done by SMITH (1940). The anatomy of the Latimeria skeleton has been later described in detail by MILLOT and ANTHONY (1958). The living coelacanth shows a general organization similar to that of the other actinistian fishes (THOMSON, 1969; JANVIER, 1996; FOREY, 1998). The skeleton can be divided in various parts (Fig. 2): skull, axial skeleton, paired and unpaired fins, tegumentary skeleton (scales). The skull, axial skeleton and fins are constituted of endoskeletal elements that are overlain by exoskeletal elements. In the extant Latimeria the endoskeleton of the skull shows an important regression of the bones, especial ly in the neurocranium, which are replaced by cartilaginous tissues, as in its Mesozoic fossil actinistian relatives (ROMER, 1937, 1942; MILLOT and ANTHONY, 1958; BJERRING, 1973; FOREY, 1998). In the same way, the endoskeleton of paired and unpaired fins is essentially constituted of four axial cartilaginous elements (Fig. 2; MILLOT and ANTHONY, 1958), with pre and post-axial
elements and eventually minute superficial endochondral ossification (CASTANET et al., 1975). The axial skeleton is composed of the notochord which is coated by an unmineralized fibrillary sheath and is totally deprived of well developed vertebrae due to the lack of ossified centra, although some discrete neural and haemal arches develop in the anterior part of the notochord (MILLOT and ANTHONY, 1958). The anterior neural and haemal spines are relatively short but they progressively increase in length posteriorly. Importantly neural and haemal spines are composed of perichondral bone surrounding a cartilaginous core (Fig. 2).
The scales belong to the exoskeleton. In both extant coelacanth species they are of elasmoid type, composed of an upper external layer also called “external ornamented layer”, and of a lower thicker layer, called the basal plate, which is stratified and almost totally unmineralized (Fig. 3). The upper external layer is ornamented with radial crests. In the posterior area of the scale these radial reliefs are overlain by numerous denticulations, the odontodes, which can be superposed (Fig. 3). The anterior area of the external layer, which is covered by the anterior scales, shows crests with irregularities that have been interpreted as growth marks and tentatively used for ageing coelacanths (HUREAU and OZOUF, 1977; MILLER, 1979).
The tooth plates of the buccal cavity support series of teeth of various sizes (Fig. 4). They range in three morphotypes: the fangs (7–10 mm in height), middle-sized teeth (3–4 mm in height) and rounded tubercles (MILLOT and ANTHONY, 1958). The teeth of the two first categories are conical and sharp whereas those of the third category have an obtuse tip (Fig. 5). Fangs of Latimeria are inserted into a socket (MILLOT and ANTHONY, 1958; HOBDELL and MILER, 1969; CASTANET et al., 1975) and they have a smooth external surface (MILLOT et al., 1978; MEUNIER et al., 2015).
A particularity of the exoskeleton of Latimeria is the abundance of odontodes at the surface of various skull bones
40 41
Fig. 5. Latimeria chalumnae. Coronoid and coronoid tooth structure. A, Tomographic imaging of a coronoid element. Three-dimensional reconstruction of the dental plate showing a fang, one mid-sized caniniform tooth (*) and numerous small round-shaped teeth (arrow-heads). The virtual section shows i) the plies of the dentine in the lower half of the pulp cavity (pc); ii) the bone of attachment (ba) between the tooth and the coronoid bony tissue. On the right of the figure, the coronoid shows a network of vascular cavities and canals (vc). (de = dentine; en = enamel) (scale = 1 mm). (After MEUNIER et al., 2015, Fig. 9-10). B, Microradiography of a cross section at the base of a fang showing several plies of the dentine wall (white arrow-heads). (bo = bone; pc = pulp cavity; de = dentine). (scale = 1 mm). (from CASTANET et al., 1975). C, Scanning Electron Microscopy of the spheritic dentine. The white and black asterisks point respectively to the external surface of the tooth and to the wall of the pulp cavity. (scale = 25 µm).
(MILLOT and ANTHONY, 1958; BERNHAUSER, 1961), on the fin rays (CASTANET et al., 1975) and on the posterior (free) area of scales (Fig. 3) (MILLOT and ANTHONY, 1958; CASTANET et al., 1975; HADIATY and RACHNATIKA, 2003; MEUNIER et al., 2008).
Among the various components of the whole skeleton, and apart the SMITH’s pioneer work (SMITH, 1940), numerous studies have been carried out on the histological organization of scales (ROUX, 1942; SMITH et al., 1972; CASTANET et al., 1975; GIRAUD et al., 1978a; MILLER, 1979; SMITH, 1979; MEUNIER, 1980; MEUNIER and ZYLBERBERG, 1999; HADIATY and RACHMATIKA, 2003; MEUNIER et al., 2008) and teeth (MILLER and HOBDELL, 1968; GRADY, 1970; HOBDELL and MILLER, 1969; CASTANET et al., 1975; SHELLIS and POOLE, 1978;
SMITH, 1978; SASAGAWA et al., 1984; MEUNIER et al., 2015). Contrary to the superficial skeleton, there are clearly less histological studies of the skeletal bony elements (FRANCILLON et al., 1975). Meanwhile, in addition to the usual skeletal elements, specific bony tissues have been described in fossil (BRITO et al., 2010) and extant coelacanths (CUPELLO et al., 2015, 2017), such as the mineralized plates surrounding the lung. These bony plates in actinistians do not belong to the skeleton sensu stricto. They are specific bony specializations in relation to the lung (see below), as is the “rocker bone” in relation to the gas bladder of some ophidiiform teleosts (PARMENTIER et al., 2008). Despite the name, the rocker bone is not true bony tissue (PARMENTIER et al., 2008), contrary to
the bony plates of fossil and extant coelacanths (BRITO et al., 2010; CUPELLO et al., 2017).
ADULT LATIMERIA SKELETAL TISSUES
Bony tissuesCortical areas of the dermal skeleton are constituted of
primary periosteal bony t issue. I ts mineralization is heterogeneous and generally marked by series of growth lines, probably due to alternative physiological cycles linked to seasonal variations (Fig. 6). The cortical primary bone shows vascular canals and numerous SHARPEY’s fibres (Fig. 6B). The centre of these bones is frequently made of a spongiosa with vascular cavities surrounded by secondary bony deposits that result from remodelling processes (Fig. 6). This secondary bone is separated from the primary one by reversal cementing lines that are hypermineralized.
Bone remodelling in adult specimens can be more or less developed according to a bone and within a given bone. Primary bone is destroyed by osteoclasts and the deposition process of the secondary bone results from the activity of osteoblasts that have replaced the osteoclasts. The secondary bone does generally not replaced the whole primary bone areas, which can still be recognizable by the presence of
SHARPEY’s fibres and/or growth marks (Fig. 6).The dermal fin rays that sustain paired and unpaired fins
in Latimeria are true lepidotrichia (CASTANET et al., 1975). They are made of two parallel opposite gutter-shaped hemirays, each of which being a series of hemisegments articulated by a collagenous ligament. Each hemisegment is made of a mineralized collagenous fibrillary matrix with embedded osteocytes. The hemisegments are sometimes fused at the basal part of the rays due to centrifugal deposition of bony laminae around the primary articulation, and eventually to the mineralization of the ligament (see figs. 19, 20 in CASTANET et al., 1975). Odontodes observed on the external convex surface of the rays are similar to those present on the dermal bones of the skull and scales. The distal extremity of the lepidotrichia is overlapped by actinotrichia (GÉRAUDIE and MEUNIER, 1980) that are long tapered rods of elastoidin , a fibrous protein of collagenous nature (FAURÉ-FREMIET, 1936; GARRAULT, 1936). The presence of lepidotrichia with distal actinotrichia in Latimeria, as in the fins of Actinopterygii, is
regarded as a plesiomorphic character for Osteichthyes (GÉRAUDIE and MEUNIER, 1980, 1984).
The upper layer of the Latimeria scale is relatively thin and it is constituted of bony tissue with embedded osteocytes, whereas the basal plate is much thicker and composed of numerous strata made of thick collagenous fibres (Fig. 7). Between two collagenous layers there are star-shaped cells (SMITH et al., 1972, pl. VI; CASTANET et al., 1975, fig. 12), the elasmocytes, which cytoplasmic processes insert between the collagenous fibres. These fibres are set in a very specific network. In each layer, the fibres are parallel to each other and the direction of the fibres change from a layer to another one. This regular organization results in a spatial arrangement termed “twisted plywood” (GIRAUD et al., 1978a,b). The rotation of fibres direction from one layer to the next has a mean angle of 27° (GIRAUD et al., 1978a). This organization is found in each scale of L. chalumnae. In L. menadoensis, the basal plate of the scales is also a twisted plywood but its rotation angle seems to be slightly less regular (MEUNIER et al.,
2008). The basal plate in both species is unmineralized (Fig. 7) excepted at the contact between the superficial layer and the basal plate, where spheritic mineralized granules are seen in the very first layers of the basal plate (MEUNIER, 1980; MEUNIER and ZYLBERBERG, 1999; MEUNIER et al., 2008). There is no pore-canal system so Latimeria’s scales are not cosmoid, contrary to most of palaeozoic sarcopterygian fishes. Due to the presence of an unmineralized stratified basal plate, the scales of Latimeria are defined as elasmoid-like scales. However the plywood-like organization is considered to be homologous with the bony basal plate of cosmoid scales of extinct sarcopterygian fishes (MEUNIER, 1980; SIRE and HUYSSEUNE, 2003; MONDEJAR, 2018; SCHULTZE, 2018).Cartilages
The cartilaginous tissues in Latimeria are characterized by long chondrocytes (Fig. 8) contrary to those of teleostean fishes that are relatively spherical in shape (MEUNIER, 1979; ZYLBERBERG and MEUNIER, 2008). Cartilage tissues can show an endochondral ossification process. This phenomenon has been studied respectively on the urohyal (PEGUETA, 1968), the MECKEL’s cartilage and the proximal piece of the pectoral girdle (FRANCILLON et al., 1975). The endochondral ossification is relatively limited when it occurs and the volume of enchondral bone remains reduced to thin bony layers (Fig. 8). The cartilage is destroyed by chondroclasts in areas where endochondral ossification occurs. Serial chondrocytes, known at the origin of the calcified cartilage, seem to be lacking in the endochondral ossification process in Latimeria (FRANCILLON et al., 1975). This constitutes a true difference with teleostean endochondral ossifications (MEUNIER, 1979; ZYLBERBERG and MEUNIER, 2008). Tooth tissues
The three tooth morphotypes described in Latimeria are fangs (7–10 mm in height), middle-sized teeth (3–4 mm in height) and rounded tubercles (MILLOT and ANTHONY, 1958). These teeth and tubercles are constituted of a cone of orthodentine set around a large pulp cavity and overlain by an external hypermineralized layer considered as true enamel (GRADY, 1970; CASTANET et al., 1975; SHELLIS and POOLE, 1978; SMITH, 1978; SASAGAWA et al., 1984). Various studies have revealed the presence of globular dentine (Fig. 5) at the periphery of the first third of the tooth (MILLER and HOBDELL, 1968; HOBDELL and MILLER, 1969; SHELLIS and POOLE, 1978; SASAGAWA et al., 1984). This typical histological organization characterizes the various teeth of the bucco-pharyngeal cavity as well as the various odontodes of the dermal bony plates, the fin rays and the scales (CASTANET et al., 1975).
In the fang the inner wall of the pulp cavity displays a series of plies (Fig. 5; MEUNIER et al., 2015). These plies start at the base of the tooth and reach at least the first third to the half of the tooth towards its tip (Fig. 5). Such plies are also present but less developed in the small caniniform teeth (MEUNIER et al., 2015). These dentine plies in the pulp cavity characterizes a plicidentine organization of orthodentine type. These primary plies correspond to a simplexodont plicidentine
REFERENCES
AGASSIZ, L. 1833–44. Recherches sur les Poissons fossiles. Imprimerie Petitpierre, Neuchâtel. [5 volumes and atlas.]
ANDREWS, S. M. 1973. Interrelationships of crossopterygians. In: GREENWOOD, P. H., MILES, R. S. and PATTERSON, C. (eds.), Interrelationships of Fishes, Acad. Press, London, pp. 137–177.
BERNHAUSER, A. 1961. Zur Knochen- und Zah, histology von Latimeria chalumnae Smith und einiger Fossilformen. Sber. öst. Akad. Wiss., 170: 119–137.
BJERRING, H. C. 1973. Relationships of coelacanthiforms. In: GREENWOOD P. H., MILES R. S. and PATTERSON C. (eds.), Interrelationships of Fishes, Acad. Press, London, pp. 179–205.
BRITO, P. M., MEUNIER, F. J., CLÉMENT, G. and GEFFARD-KURIYAMA, D. 2010. The histological structure of the calcified lung of the fossil coelacanth Axelrodichthys araripensis (Actinistia: Mawsoniidae). Palaeontology, 53: 1281–1290.
CASANE, D. and LAURENTI, P. 2013. Why coelacanths are not ‘living fossils’. Bioessays, 35: 332–338. DOI 10.1002/bies.201200145.
CASTANET, J., MEUNIER, F., BERGOT, C. and FRANÇOIS, Y. 1975. Données pré l iminaires sur les s t ructures histologiques du squelette de Latimeria chalumnae. I - Dents, écailles, rayons de nageoires. In: Coll. Inter. CNRS, "Problèmes actuels de Paléontologie. Evolution des Vertébrés", Paris, 4–9 Juin 1973, 159–168, 4 Pls.
CLEMENT, G. 1999. The actinistian (Sarcopterygii) Piveteauia madagascarensis LEHMAN from the Lower Triassic of Northeastern Madagascar: a redescription on the basis of new material. Journal of Vertebrate Paleontology, 19: 234–242.
CLEMENT, G. 2005. A new coelacanth (Actinist ia, Sarcopterygii) from the Jurassic of France, and the question of the closest relative fossil to Latimeria. Journal of Vertebrate Paleontology, 25: 481–491.
CUPELLO, C., BRITO, P. M., MEUNIER, F. J., HERBIN, M., JANVIER, P., DUTEL, H. & CLÉMENT, G. 2015. Allometric growth in the extant coelacanth lung during ontogenetic development. Nature Communications, 6: 8222. DOI: 10.1038/ncomms9222.
CUPELLO, C., CLEMÉNT, G., MEUNIER, F. J., HERBIN, M., YABUMOTO, Y. and BRITO, P. M. 2019. The long-time adaptation of coelacanths to moderate deep water: reviewing the evidences. Bulletin of Kitakyushu Museum of Natural History and Human History Series A (Natural History), 17: 29–35.
CUPELLO, C., MEUNIER, F. J., HERBIN, M., JANVIER, P., CLÉMENT, G. and BRITO, P. M. 2017. The homology and funct ion of the lung pla tes in extant and foss i l coelacanths. Scientific Reports , 7 : 9244. DOI: 10.1038/s41598-017-09327-6.
EASTMAN, J. T., WITMER, L. M., RIDGELY, R. C. and KUHN, K.
L. 2014. Divergence in skeleta l mass and bone morphology in antarctic notothenioid fishes. Journal of Morphology, 275: 841–861. DOI: 10.1002/jmor.20258
ERDMANN, M. V., CALDWELL, R. L. and MOOSA, M. K. 1998. Indonesian “king of the sea” discovered. Nature, 395: 335.
ERDMANN, M. V., CALDWELL, R. L., JEWETT, S. L. and TJAKRAWIDJAJA, A. 1999. The second recorded living coelacanth from north Sulawesi. Environmental Biology of Fishes, 34: 445–451.
FAURÉ-FREMIET, M. 1936. La structure des fibres d’éastoidine. Archives d'Anatomie Microscopique, 32: 249–269.
FOREY, P. L.- 1984. The coelacanth as a living fossil. In: ELREDGE N. and STANLEY S.M. (eds.), Living fossils. Springer Verlag, Berlin, pp. 166–169.
FOREY, P. L.- 1998. History of the Coelacanth Fishes. Chapman and Hall, London, 419 pp.
FRANCILLON, H., MEUNIER, F., NGO TUAN PHONG, D. and RICQLÈS, A. (DE). 1975. Données préliminaires sur les structures histologiques du squelette de Latimeria chalumnae. I I- Tissus osseux et cartilages. In: Coll. Inter. CNRS, "Problèmes actuels de Paléontologie. Evolution des Vertébrés", Paris, 4–9 Juin 1973, Centre National de la Recherche Scientifique, pp. 169–174, 2 pls.
FRANCILLON-VIEILLOT H., BUFFRENIL V. DE, CASTANET J., GÉRAUDIE J., MEUNIER FJ., SIRE J-Y., ZYLBERBERG L., a n d R I C Q L È S A . D E 1 9 9 0 . M i c r o s t r u c t u r e a n d mineralization of vertebrate skeletal tissues. In: CARTER, J. G. (ed.), Skeletal Biomineralization: Patterns, Processes and Evolutionary Trends. Vol. I , Van Nostrand, New York, pp. 471–530
GARRAULT, H., 1936. Développement des fibres d’élastoidine (actinotrichia) chez les salmonides. Archives d'Anatomie Microscopique, 130: 105–137.
GÉRAUDIE, J. and MEUNIER, F. J. 1980. Elastoidin actinotrichia in coelacanth fins: a comparative study with teleosts. Tissue and Cell, 12: 637–645.
GÉRAUDIE, J. and MEUNIER, F. J. 1984. Structure and comparative morphology of camptotrichia of lungfish fins. Tissue and Cell, 16: 217–236.
GIRAUD, M. M., CASTANET, J., MEUNIER, F. J. and BOULIGAND, Y. 1978a. The fibrous structure of coelacanth scales: a twisted "plywood". Tissue and Cell, 10: 671–686.
GIRAUD, M. M., CASTANET, J., MEUNIER, F. J. and BOULIGAND, Y. 1978b. Organisation spatiale de l'isopédine des écailles du coelacanthe (Latimeria chalumnae SMITH). Comptes Rendus de l’Académie des Sciences, 287: 487–489.
GRADY, J . E. 1970. Tooth development in Latimeria chalumnae (SMITH). Journal of Morphology, 132: 377–388.
HADIATY, R. K. and RACHMATIKA, I. 2003. Morphological study of the scales of Latimeria menadoensis POUYAUD et al., 1999. Treubia (A Journal on Zoology of the Indo-Australian Archipelago), 33: 1–11.
HOBDELL, M. H. and MILLER, W. A. 1969. Radiographic
anatomy of the teeth and tooth supporting tissues of Latimeria chalumnae. Archives of Oral Biology, 14: 855–858.
HUREAU J.-Cl. and OZOUF, C. 1977. Détermination de l’âge et croissance du cœlacanthe Latimeria chalumnae SMITH, 1939 (Poisson, Crossoptérygien, Coelacanthidé). Cybium, 2: 129–137.
JANVIER, P. 1996. Early Vertebrates. Clarendon Press, Oxford, 393 pp.
MEUNIER, F. J. 1979. Etude histologique et microradiographique du cartilage hémal de la vertèbre de la carpe, Cyprinus carpio L. (Pisces, Teleostei, Cyprinidae). Acta Zoologica, 60: 19–31.
MEUNIER, F. J. 1980. Les relations isopédine-tissu osseux dans le post-temporal et les écailles de la ligne latérale de Latimeria chalumnae (SMITH). Zoologica Scripta, 9: 307–317.
MEUNIER, F. J. and ZYLBERBERG, L. 1999. The structure of the external layer and of the odontodes of scales in Latimeria chalumnae (Sarcopterygii, Actinistia, Coelacanthidae) revisited using scanning and transmission electron microscopy. In: SÉRET, B. and SIRE, J.-Y. (eds.), Proc. 5th Indo-Pacific Fish Conf., Nouméa, 1997, Soc. Fr. Ichtyol., Paris, pp. 109–116.
MEUNIER, F. J., ERDMANN, M. V., FERMON, Y. and CALDWELL, R. L. 2008. Can the comparative study of the morphology and histology of the scales of Latimeria menadoensis and L. chalumnae (Sarcopterygii, Actinistia, Coelacanthidae) bring new insight on the taxonomy and the biogeography of recent coelacanthids? Geological Society, London, Special Publications, 295: 351–360.
MEUNIER, F. J., BRITO, P. M. and LEAL, M.-E. 2013. Quelques données morphologiques et histologiques sur la structure de la plaque dentaire linguale d’Arapaima gigas (Osteoglossidae; Teleostei). Cybium, 37: 263–271
MEUNIER, F. J., MONDEJAR-FERNANDEZ, J., GOUSSARD, F., CLÉMENT, G. and HERBIN, M. 2015. Presence of plicidentine in the oral teeth of Latimeria chalumnae (Sarcopterygii; Actinistia; Coelacanthidae). Journal of Structural Biology, 190: 31–37.
MILLER, W. A. 1979. Observations on the structure of mineralized tissue of the coelacanth, including scales and their associate odontodes. Occasional Papers of the Californian Academy of Sciences, 134: 68–78.
MILLER, W. A. and HOBDELL, M. H. 1968. Preliminary report on the histology of the dental and paradental tissues of Latimeria chalumnae (SMITH) with a note on tooth replacement. Archives of Oral Biology, 13: 1289–1291.
MILLOT, J. and ANTHONY, J. 1958. Anatomie de Latimeria chalumnae. T. 1. Squelette, muscles et formations de soutien. CNRS Edr., Paris. 122 pp., 80 pls.
MILLOT, J., ANTHONY, J. and ROBINEAU, D. 1978. Anatomie de Latimeria chalumnae. T. 3. Appareil digestif, téguments, écailles. CNRS Edr., Paris. pp. 7–28, 135–139.
MONDEJAR-FERNANDEZ, J. 2018. On cosmine: its origins, b io logy and impl ica t ions for sarcopterygian interrelationships. Cybium, 42: 41–65.
ØRVIG T. 1951. Histologic studies of Placoderms and fossil Elasmobranchs. 1: The endoskeleton, with remarks on the hard tissues of lower vertebrates in general. Arkiv för Zoologi, 2: 321–454, 8 pl.
ØRVIG T. 1968. The dermal skeleton; general considerations. In: ØRVIG T. (ed.), Current problems of lower vertebrate phylogeny. Proceedings of the fourth Nobel Symposium, Stockholm, 1967, J. Wiley, New-York. pp. 373–397.
PARMENTIER, E., COMPÈRE, P., CASADEWALL, M., FONTENELLE, N., CLOOTS, R. and HENRIST, C. 2008. The rocker bone: a new kind of mineralized tissue? Cell and Tissue Research, 334: 67–79.
PEGUETA, V. P. 1968. Enchondral ossification in Latimeria chalumnae SMITH. Dopovidi Akademii Nauk Ukrainia SSR., 7: 653–656 (in Russian; English abstr.).
POUYAUD, L., WIRJOATMODIO, S., RACHMATIKA, L., TJAKRAWIDJAJA, A., HADIATY, R. K. and HADIE, W. 1999. Une nouvelle espèce de coelacanthe, preuves génétiques et morphologiques. Comptes Rendus de l’Académie des Sciences, 322: 261–267.
ROMER, A.S. 1937. The braincase of the carboniferous crossopterygian Megalichthys nitidus. Bulletin of the Museum of Comparative Zoology, 824: 1–73.
ROMER, A.S. 1942. Cartilage as an embryonic adaptation. American Naturalist, 76: 394–404.
ROUX, G. H. 1942. The microscopic anatomy of the Latimeria scale. South African Journal of Medical Sciences, 7: 1–18.
SASAGAWA, I., ISHIYAMA, M. and KODERA, H. 1984. Fine structure of the pharyngeal teeth in the coelacanth fish (Latimeria chalumnae). In: FEARNHEAD, R. W. and SUGA, S. (eds.), Tooth enamel IV, Elsevier Science Publishers, Amsterdam, pp. 462–466.
SCHULTZE, H.-P. 1969. Die Faltenzähne der Rhipidistiden Crossopterygier, der Tetrapoden und der Actinopterygi-er-Gattung Lepisosteus nebst einer Beschreibung der Zahnstruktur von Onychodus (Struniiformer Crossoptery-gier). Palaeontologica Italica, New Series, 35(65): 63–137.
SCHULTZE, H.-P. 1970. Folded teeth and the monophyletic origin of Tetrapods. American Museum Novitates, 2408: 1–10.
SCHULTZE, H.-P. 2018. Hard tissues in fish evolution: history and current issues. Cybium, 42(1): 29–39.
SHELLIS, R. P. and POOLE, D. F. G. 1978. The structure of the dental hard tissues of the coelacanthid fish Latimeria chalumnae SMITH. Archives of Oral Biology, 23: 1105–1113.
SIRE, J.-Y. and HUYSSEUNE, A. 2003. Formation of dermal skeletal and dental tissues in fish: a comparative and evolutionary approach. Biological Review, 78: 219–249.
SMITH, J. L. B. 1939. A living fish of the Mesozoic type. Nature, 143: 455–456.
SMITH, J. L. B. 1940. A living coelacanth fish from South Africa. Transactions of the Royal Society of South Africa, 28: 1–106, 44 Pls.
SMITH, M. M. 1978. Enamel in the oral teeth of Latimeria chalumnae (Pisces: Actinistia): a scanning electron microscopic study. Journal of Zoology, 185: 355–369.
SMITH, M. M. 1979. Scanning electron microscopy of odontodes in the scales of coelacanth embryo, Latimeria chalumnae SMITH. Archives of Oral Biology, 24: 179–183.
SMITH, M. M., HOBDELL, M. H. and MILLER, W. A. 1972. The structure of the scales of Latimeria chalumnae. Journal of Zoology, London, 167: 501–509.
THOMSON, K. S. 1969. The biology of the lobe-finned fishes.
Biological Reviews, 44: 91–154. UYENO, T. and YABUMOTO, Y. 2007. Origin of extant
coelacanths. The Coelacanth, Fathom the Mystery 2007. Aquamarine Fukushima, pp. 24–26.
ZYLBERBERG, L. and MEUNIER, F. J. 2008. New data on the structure and the chondrocyte populations of the haemal cartilage of abdominal vertebrae in the adult carp Cyprinus carpio (Teleostei, Ostariophysii, Cyprinidae). Cybium, 32: 225–239.
ZYLBERBERG, L., GÉRAUDIE, J., MEUNIER, F. J., & SIRE J. Y., 1992. Biomineralization in the integumental skeleton of the living lower Vertebrates. In: HALL, B. K. (ed), Bone, Volume 4, CRC Press, Boca Raton, pp. 171–224.
The skeleton and the mineralized tissues of the living coelacanthsFrançois J. MEUNIER, Camila CUPELLO and Gaël CLÉMENT
Fig. 6. Latimeria chalumnae. Bone histology. A, Thin section in a coronoid bone (Basic fuchsin staining) showing primary bone (pb) with osteocytes, secondary bone (sb), a cementing line (arrow-heads) and a vascular cavity (vc). (scale = 50 µm). Inset: detail of osteocytes (arrow-heads) and their osteocytic canalicles. (scale = 25 µm). B, Thin section in a coronoid bone (Hemalun-PIC staining) showing cellular primary bone (pb) and its periostic membrane (white arrows). On the right, numerous collagenous fibres (cf) enter the bony matrix where they form SHARPEY’s fibres. The asterisks points to secondary vascular cavities limited by cementing lines (white arrow-heads); a primary vascular canal (vc) is seen. (scale = 100 µm). C, X-ray ground section in the cortical region of the angular showing primary bone with regular alternative of heterogeneous mineralized bony tissue. The black arrow-heads point to primary vascular canals, the white arrows to cement ing l ines , and the white arrow-heads to erosive bays. The asterisks point to two confluent secondary vascular cavities). (scale = 200 µm). D, Ground section of the angular, in a deeper region than C. Several bulks of secondary bone (sb) are limited by cementing lines (black arrow-heads). Primary bone (pb) persists in different areas. The white asterisks point to secondary vascular cavities. (scale = 200 µm). (C, D: After FRANCILLON et al., 1975).

type according to the definition of MEUNIER et al. (2013). This organization is less complex than the three other plicidentine types (polyplocodont, eusthenodont and dendrodont), which are described in most of extinct and extant Sarcopterygii (SCHULTZE, 1969, 1970).The bony plates of the lung
The lung of Latimeria is reduced to a short oesophagus diverticulum (CUPELLO et al., 2015) surrounded by scattered small and thin ovoid plates (Fig. 9A, B). These plates cover the surface of the vestigial lung, and are composed of a fibrous matrix with fibrocytes: it is a cellular bony tissue (CUPELLO et al., 2017). Each plate is enclosed in a membranous bag. These plates look fragile in their middle plane since a central weakness zone opened on the microtome knife, creating an artefactual lumen (Fig. 9C) (CUPELLO et al., 2017). The fibres are collagenous and organized in superposed layers. The cells are true osteocytes (Fig. 9E) with a more or less star-shaped outline, and send cytoplasmic extensions in the thickness of the plate (Fig. 9D, E). Rare lining cells (osteoblasts) are
present on the surface of the plates, and conjunctive fibres (SHARPEY’s fibres) penetrate within the plate (Fig. 9C). The extracellular matrix shows various staining intensities with concentric and parallel (Fig. 9D) or globular shaped lines. These lines are considered to be Liesegang lines that characterize an active spheritic mineralizing process. The lung plates of Latimeria are thus true bony plates, and are homologous with the large bony plates known in the abdominal cavity of fossil actinistians (BRITO et al., 2010; CUPELLO et al., 2017). The calcified walls that surrounded the lung of the Cretaceous coelacanth Axelrodichthys are made of large osseous plates of various thickness (BRITO et al., 2010), as those of other fossil coelacanths (CLEMENT, 1999, 2005). The bony tissue of these plates is a vascularized cellular bone with more or less large vascular cavities and some internal remodelling (BRITO et al., 2010). Moreover the mineralization of the lung ossified plates in Axelrodichthys is spheritic as in Latimeria’s plates (CUPELLO et al., 2017). So there is a continuity of the histological structure of the lung plates in the coelacanths during their
evolution, but with an important reduction of the bony plates in Latimeria linked to the vestigial state of its lung (CUPELLO et al., 2015, 2019).
CONCLUSION
This overview of eighty years (1938–2018) of histological work on Latimeria skeletal tissues allows some anatomical and evolutionary considerations. It can be enlightened a drastic reduction of endochondral ossification during the long evolutionary history of coelacanths. The persistence of large volume of cartilage in the endoskeleton at adult stage can be compared to the “little bone and considerable cartilage” that characterize the skeleton of a number of demersal notothenioid telostean fishes (EASTMAN et al., 2014).
A processus of spheritic mineralization has been recently highlighted in various skeletal elements of Latimeria, by the presence of globular dentine in teeth, in odontodes of the tegumentary skeleton (scales, fin rays), in scales at the interface between the external layer and the basal plate, as well as in lung bony plates. The spheritic mineralization (i.e., a radiating arrangement of hydroxyapatite crystals and of the organic matrix) in vertebrate skeletal tissues is considered as a precursor of inotropic mineralization (specific interactions between collagen fibrils and the mineral phase) that possibly represents a derived evolutionary stage of calcification mechanisms (ØRVIG, 1951, 1968; FRANCILLON-VIEILLOT et al., 1990; ZYLBERBERG et al., 1992). The simultaneous presence of both mineralization processes (spheritic and inotropic) in the mineralized tissues of Latimeria and in other osteichthyans such as teleosteans (ZYLBERBERG and MEUNIER, 2008) is however questionable. The mineralized spherules present at the limit between the external layer and the basal plate of Latimeria scales are considered to be the product of an inotropic mineralization (MEUNIER and ZYLBERBERG, 1999). It thus appears that the state of the mineralized spherules observed in the various skeletal tissues of Latimeria, including the lung bony plates, must be studied with adapted ultrastructural techniques in order to test their true origin: spheritic or inotropic mineralization?
ACKNOWLEDGEMENTS
This contribution is linked to an international roundtable on coelacanths evolution held at the Kitakyushu Museum and at the Aquamarine Fukushima in August 2017. We would like to thank the anonymous reviewers for their constructive comments and valuable suggestions. C.C. was partially financed by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brasil (CAPES) – Finance Code 001 (Programa Nacional de Pós Doutorado-PNPD).
INTRODUCTION
The first study of fossil coelacanths was done by AGASSIZ (1833–44) and was followed by a lot of papers with the discovery of new fossils (see reviews of JANVIER, 1996 and FOREY, 1998). Fossil coelacanths constitute an important monophyletic group composed of more than forty genera, eighty species, and belong to the Sarcopterygii (ANDREWS, 1973; JANVIER, 1996; FOREY, 1998; UYENO and YABUMOTO, 2007; and many others). For a century coelacanths were considered extinct since the end of the Cretaceous time (Fig. 1).
With the discovery of a living coelacanth in 1938 (SMITH, 1939) the scientific community was soon highly interested to compare the incomplete fossil sarcopterygian fishes dataset to the recent anatomical and biological organization of Latimeria chalumnae. Many authors (see THOMSON, 1969; MILLOT and ANTHONY, 1958; MILLOT et al., 1978 FOREY, 1984, 1998; JANVIER, 1996) focused their work on Latimeria as a key-taxon to test or propose various hypotheses on the origin and evolutionary history of the early tetrapods and sarcopterygii in general, and to infer their possible biological characteristics. The fossil sarcopterygian fishes are known by their fossilized mineralized tissues, essentially their bones, scales and teeth. Together with the extant lungfishes, especially Neoceratodus, Latimeria offered a unique access to the skeleton as a whole: with both mineralized and unmineralized skeletal tissues. Here, we aim to present an overview of the last eighty years (1938–2018) of histological studies of Latimeria’s skeleton.
Since the discovery of the Raja Laut (“king of the sea” ), off Sulawesi Island in Indonesia (ERDMANN et al., 1998, 1999), the genus Latimeria comprises two species: L. chalumnae SMITH 1939 and L. menadoensis POUYAUD et al., 1999. However our knowledge of the anatomy and biology of the living coelacanths is so far essentially based on L. chalumnae.
THE LATIMERIA SKELETON
The first anatomical observations on the extant coelacanth were done by SMITH (1940). The anatomy of the Latimeria skeleton has been later described in detail by MILLOT and ANTHONY (1958). The living coelacanth shows a general organization similar to that of the other actinistian fishes (THOMSON, 1969; JANVIER, 1996; FOREY, 1998). The skeleton can be divided in various parts (Fig. 2): skull, axial skeleton, paired and unpaired fins, tegumentary skeleton (scales). The skull, axial skeleton and fins are constituted of endoskeletal elements that are overlain by exoskeletal elements. In the extant Latimeria the endoskeleton of the skull shows an important regression of the bones, especial ly in the neurocranium, which are replaced by cartilaginous tissues, as in its Mesozoic fossil actinistian relatives (ROMER, 1937, 1942; MILLOT and ANTHONY, 1958; BJERRING, 1973; FOREY, 1998). In the same way, the endoskeleton of paired and unpaired fins is essentially constituted of four axial cartilaginous elements (Fig. 2; MILLOT and ANTHONY, 1958), with pre and post-axial
elements and eventually minute superficial endochondral ossification (CASTANET et al., 1975). The axial skeleton is composed of the notochord which is coated by an unmineralized fibrillary sheath and is totally deprived of well developed vertebrae due to the lack of ossified centra, although some discrete neural and haemal arches develop in the anterior part of the notochord (MILLOT and ANTHONY, 1958). The anterior neural and haemal spines are relatively short but they progressively increase in length posteriorly. Importantly neural and haemal spines are composed of perichondral bone surrounding a cartilaginous core (Fig. 2).
The scales belong to the exoskeleton. In both extant coelacanth species they are of elasmoid type, composed of an upper external layer also called “external ornamented layer”, and of a lower thicker layer, called the basal plate, which is stratified and almost totally unmineralized (Fig. 3). The upper external layer is ornamented with radial crests. In the posterior area of the scale these radial reliefs are overlain by numerous denticulations, the odontodes, which can be superposed (Fig. 3). The anterior area of the external layer, which is covered by the anterior scales, shows crests with irregularities that have been interpreted as growth marks and tentatively used for ageing coelacanths (HUREAU and OZOUF, 1977; MILLER, 1979).
The tooth plates of the buccal cavity support series of teeth of various sizes (Fig. 4). They range in three morphotypes: the fangs (7–10 mm in height), middle-sized teeth (3–4 mm in height) and rounded tubercles (MILLOT and ANTHONY, 1958). The teeth of the two first categories are conical and sharp whereas those of the third category have an obtuse tip (Fig. 5). Fangs of Latimeria are inserted into a socket (MILLOT and ANTHONY, 1958; HOBDELL and MILER, 1969; CASTANET et al., 1975) and they have a smooth external surface (MILLOT et al., 1978; MEUNIER et al., 2015).
A particularity of the exoskeleton of Latimeria is the abundance of odontodes at the surface of various skull bones
42 43
(MILLOT and ANTHONY, 1958; BERNHAUSER, 1961), on the fin rays (CASTANET et al., 1975) and on the posterior (free) area of scales (Fig. 3) (MILLOT and ANTHONY, 1958; CASTANET et al., 1975; HADIATY and RACHNATIKA, 2003; MEUNIER et al., 2008).
Among the various components of the whole skeleton, and apart the SMITH’s pioneer work (SMITH, 1940), numerous studies have been carried out on the histological organization of scales (ROUX, 1942; SMITH et al., 1972; CASTANET et al., 1975; GIRAUD et al., 1978a; MILLER, 1979; SMITH, 1979; MEUNIER, 1980; MEUNIER and ZYLBERBERG, 1999; HADIATY and RACHMATIKA, 2003; MEUNIER et al., 2008) and teeth (MILLER and HOBDELL, 1968; GRADY, 1970; HOBDELL and MILLER, 1969; CASTANET et al., 1975; SHELLIS and POOLE, 1978;
SMITH, 1978; SASAGAWA et al., 1984; MEUNIER et al., 2015). Contrary to the superficial skeleton, there are clearly less histological studies of the skeletal bony elements (FRANCILLON et al., 1975). Meanwhile, in addition to the usual skeletal elements, specific bony tissues have been described in fossil (BRITO et al., 2010) and extant coelacanths (CUPELLO et al., 2015, 2017), such as the mineralized plates surrounding the lung. These bony plates in actinistians do not belong to the skeleton sensu stricto. They are specific bony specializations in relation to the lung (see below), as is the “rocker bone” in relation to the gas bladder of some ophidiiform teleosts (PARMENTIER et al., 2008). Despite the name, the rocker bone is not true bony tissue (PARMENTIER et al., 2008), contrary to
the bony plates of fossil and extant coelacanths (BRITO et al., 2010; CUPELLO et al., 2017).
ADULT LATIMERIA SKELETAL TISSUES
Bony tissuesCortical areas of the dermal skeleton are constituted of
primary periosteal bony t issue. I ts mineralization is heterogeneous and generally marked by series of growth lines, probably due to alternative physiological cycles linked to seasonal variations (Fig. 6). The cortical primary bone shows vascular canals and numerous SHARPEY’s fibres (Fig. 6B). The centre of these bones is frequently made of a spongiosa with vascular cavities surrounded by secondary bony deposits that result from remodelling processes (Fig. 6). This secondary bone is separated from the primary one by reversal cementing lines that are hypermineralized.
Bone remodelling in adult specimens can be more or less developed according to a bone and within a given bone. Primary bone is destroyed by osteoclasts and the deposition process of the secondary bone results from the activity of osteoblasts that have replaced the osteoclasts. The secondary bone does generally not replaced the whole primary bone areas, which can still be recognizable by the presence of
SHARPEY’s fibres and/or growth marks (Fig. 6).The dermal fin rays that sustain paired and unpaired fins
in Latimeria are true lepidotrichia (CASTANET et al., 1975). They are made of two parallel opposite gutter-shaped hemirays, each of which being a series of hemisegments articulated by a collagenous ligament. Each hemisegment is made of a mineralized collagenous fibrillary matrix with embedded osteocytes. The hemisegments are sometimes fused at the basal part of the rays due to centrifugal deposition of bony laminae around the primary articulation, and eventually to the mineralization of the ligament (see figs. 19, 20 in CASTANET et al., 1975). Odontodes observed on the external convex surface of the rays are similar to those present on the dermal bones of the skull and scales. The distal extremity of the lepidotrichia is overlapped by actinotrichia (GÉRAUDIE and MEUNIER, 1980) that are long tapered rods of elastoidin , a fibrous protein of collagenous nature (FAURÉ-FREMIET, 1936; GARRAULT, 1936). The presence of lepidotrichia with distal actinotrichia in Latimeria, as in the fins of Actinopterygii, is
regarded as a plesiomorphic character for Osteichthyes (GÉRAUDIE and MEUNIER, 1980, 1984).
The upper layer of the Latimeria scale is relatively thin and it is constituted of bony tissue with embedded osteocytes, whereas the basal plate is much thicker and composed of numerous strata made of thick collagenous fibres (Fig. 7). Between two collagenous layers there are star-shaped cells (SMITH et al., 1972, pl. VI; CASTANET et al., 1975, fig. 12), the elasmocytes, which cytoplasmic processes insert between the collagenous fibres. These fibres are set in a very specific network. In each layer, the fibres are parallel to each other and the direction of the fibres change from a layer to another one. This regular organization results in a spatial arrangement termed “twisted plywood” (GIRAUD et al., 1978a,b). The rotation of fibres direction from one layer to the next has a mean angle of 27° (GIRAUD et al., 1978a). This organization is found in each scale of L. chalumnae. In L. menadoensis, the basal plate of the scales is also a twisted plywood but its rotation angle seems to be slightly less regular (MEUNIER et al.,
2008). The basal plate in both species is unmineralized (Fig. 7) excepted at the contact between the superficial layer and the basal plate, where spheritic mineralized granules are seen in the very first layers of the basal plate (MEUNIER, 1980; MEUNIER and ZYLBERBERG, 1999; MEUNIER et al., 2008). There is no pore-canal system so Latimeria’s scales are not cosmoid, contrary to most of palaeozoic sarcopterygian fishes. Due to the presence of an unmineralized stratified basal plate, the scales of Latimeria are defined as elasmoid-like scales. However the plywood-like organization is considered to be homologous with the bony basal plate of cosmoid scales of extinct sarcopterygian fishes (MEUNIER, 1980; SIRE and HUYSSEUNE, 2003; MONDEJAR, 2018; SCHULTZE, 2018).Cartilages
The cartilaginous tissues in Latimeria are characterized by long chondrocytes (Fig. 8) contrary to those of teleostean fishes that are relatively spherical in shape (MEUNIER, 1979; ZYLBERBERG and MEUNIER, 2008). Cartilage tissues can show an endochondral ossification process. This phenomenon has been studied respectively on the urohyal (PEGUETA, 1968), the MECKEL’s cartilage and the proximal piece of the pectoral girdle (FRANCILLON et al., 1975). The endochondral ossification is relatively limited when it occurs and the volume of enchondral bone remains reduced to thin bony layers (Fig. 8). The cartilage is destroyed by chondroclasts in areas where endochondral ossification occurs. Serial chondrocytes, known at the origin of the calcified cartilage, seem to be lacking in the endochondral ossification process in Latimeria (FRANCILLON et al., 1975). This constitutes a true difference with teleostean endochondral ossifications (MEUNIER, 1979; ZYLBERBERG and MEUNIER, 2008). Tooth tissues
The three tooth morphotypes described in Latimeria are fangs (7–10 mm in height), middle-sized teeth (3–4 mm in height) and rounded tubercles (MILLOT and ANTHONY, 1958). These teeth and tubercles are constituted of a cone of orthodentine set around a large pulp cavity and overlain by an external hypermineralized layer considered as true enamel (GRADY, 1970; CASTANET et al., 1975; SHELLIS and POOLE, 1978; SMITH, 1978; SASAGAWA et al., 1984). Various studies have revealed the presence of globular dentine (Fig. 5) at the periphery of the first third of the tooth (MILLER and HOBDELL, 1968; HOBDELL and MILLER, 1969; SHELLIS and POOLE, 1978; SASAGAWA et al., 1984). This typical histological organization characterizes the various teeth of the bucco-pharyngeal cavity as well as the various odontodes of the dermal bony plates, the fin rays and the scales (CASTANET et al., 1975).
In the fang the inner wall of the pulp cavity displays a series of plies (Fig. 5; MEUNIER et al., 2015). These plies start at the base of the tooth and reach at least the first third to the half of the tooth towards its tip (Fig. 5). Such plies are also present but less developed in the small caniniform teeth (MEUNIER et al., 2015). These dentine plies in the pulp cavity characterizes a plicidentine organization of orthodentine type. These primary plies correspond to a simplexodont plicidentine
REFERENCES
AGASSIZ, L. 1833–44. Recherches sur les Poissons fossiles. Imprimerie Petitpierre, Neuchâtel. [5 volumes and atlas.]
ANDREWS, S. M. 1973. Interrelationships of crossopterygians. In: GREENWOOD, P. H., MILES, R. S. and PATTERSON, C. (eds.), Interrelationships of Fishes, Acad. Press, London, pp. 137–177.
BERNHAUSER, A. 1961. Zur Knochen- und Zah, histology von Latimeria chalumnae Smith und einiger Fossilformen. Sber. öst. Akad. Wiss., 170: 119–137.
BJERRING, H. C. 1973. Relationships of coelacanthiforms. In: GREENWOOD P. H., MILES R. S. and PATTERSON C. (eds.), Interrelationships of Fishes, Acad. Press, London, pp. 179–205.
BRITO, P. M., MEUNIER, F. J., CLÉMENT, G. and GEFFARD-KURIYAMA, D. 2010. The histological structure of the calcified lung of the fossil coelacanth Axelrodichthys araripensis (Actinistia: Mawsoniidae). Palaeontology, 53: 1281–1290.
CASANE, D. and LAURENTI, P. 2013. Why coelacanths are not ‘living fossils’. Bioessays, 35: 332–338. DOI 10.1002/bies.201200145.
CASTANET, J., MEUNIER, F., BERGOT, C. and FRANÇOIS, Y. 1975. Données pré l iminaires sur les s t ructures histologiques du squelette de Latimeria chalumnae. I - Dents, écailles, rayons de nageoires. In: Coll. Inter. CNRS, "Problèmes actuels de Paléontologie. Evolution des Vertébrés", Paris, 4–9 Juin 1973, 159–168, 4 Pls.
CLEMENT, G. 1999. The actinistian (Sarcopterygii) Piveteauia madagascarensis LEHMAN from the Lower Triassic of Northeastern Madagascar: a redescription on the basis of new material. Journal of Vertebrate Paleontology, 19: 234–242.
CLEMENT, G. 2005. A new coelacanth (Actinist ia, Sarcopterygii) from the Jurassic of France, and the question of the closest relative fossil to Latimeria. Journal of Vertebrate Paleontology, 25: 481–491.
CUPELLO, C., BRITO, P. M., MEUNIER, F. J., HERBIN, M., JANVIER, P., DUTEL, H. & CLÉMENT, G. 2015. Allometric growth in the extant coelacanth lung during ontogenetic development. Nature Communications, 6: 8222. DOI: 10.1038/ncomms9222.
CUPELLO, C., CLEMÉNT, G., MEUNIER, F. J., HERBIN, M., YABUMOTO, Y. and BRITO, P. M. 2019. The long-time adaptation of coelacanths to moderate deep water: reviewing the evidences. Bulletin of Kitakyushu Museum of Natural History and Human History Series A (Natural History), 17: 29–35.
CUPELLO, C., MEUNIER, F. J., HERBIN, M., JANVIER, P., CLÉMENT, G. and BRITO, P. M. 2017. The homology and funct ion of the lung pla tes in extant and foss i l coelacanths. Scientific Reports , 7 : 9244. DOI: 10.1038/s41598-017-09327-6.
EASTMAN, J. T., WITMER, L. M., RIDGELY, R. C. and KUHN, K.
L. 2014. Divergence in skeleta l mass and bone morphology in antarctic notothenioid fishes. Journal of Morphology, 275: 841–861. DOI: 10.1002/jmor.20258
ERDMANN, M. V., CALDWELL, R. L. and MOOSA, M. K. 1998. Indonesian “king of the sea” discovered. Nature, 395: 335.
ERDMANN, M. V., CALDWELL, R. L., JEWETT, S. L. and TJAKRAWIDJAJA, A. 1999. The second recorded living coelacanth from north Sulawesi. Environmental Biology of Fishes, 34: 445–451.
FAURÉ-FREMIET, M. 1936. La structure des fibres d’éastoidine. Archives d'Anatomie Microscopique, 32: 249–269.
FOREY, P. L.- 1984. The coelacanth as a living fossil. In: ELREDGE N. and STANLEY S.M. (eds.), Living fossils. Springer Verlag, Berlin, pp. 166–169.
FOREY, P. L.- 1998. History of the Coelacanth Fishes. Chapman and Hall, London, 419 pp.
FRANCILLON, H., MEUNIER, F., NGO TUAN PHONG, D. and RICQLÈS, A. (DE). 1975. Données préliminaires sur les structures histologiques du squelette de Latimeria chalumnae. I I- Tissus osseux et cartilages. In: Coll. Inter. CNRS, "Problèmes actuels de Paléontologie. Evolution des Vertébrés", Paris, 4–9 Juin 1973, Centre National de la Recherche Scientifique, pp. 169–174, 2 pls.
FRANCILLON-VIEILLOT H., BUFFRENIL V. DE, CASTANET J., GÉRAUDIE J., MEUNIER FJ., SIRE J-Y., ZYLBERBERG L., a n d R I C Q L È S A . D E 1 9 9 0 . M i c r o s t r u c t u r e a n d mineralization of vertebrate skeletal tissues. In: CARTER, J. G. (ed.), Skeletal Biomineralization: Patterns, Processes and Evolutionary Trends. Vol. I , Van Nostrand, New York, pp. 471–530
GARRAULT, H., 1936. Développement des fibres d’élastoidine (actinotrichia) chez les salmonides. Archives d'Anatomie Microscopique, 130: 105–137.
GÉRAUDIE, J. and MEUNIER, F. J. 1980. Elastoidin actinotrichia in coelacanth fins: a comparative study with teleosts. Tissue and Cell, 12: 637–645.
GÉRAUDIE, J. and MEUNIER, F. J. 1984. Structure and comparative morphology of camptotrichia of lungfish fins. Tissue and Cell, 16: 217–236.
GIRAUD, M. M., CASTANET, J., MEUNIER, F. J. and BOULIGAND, Y. 1978a. The fibrous structure of coelacanth scales: a twisted "plywood". Tissue and Cell, 10: 671–686.
GIRAUD, M. M., CASTANET, J., MEUNIER, F. J. and BOULIGAND, Y. 1978b. Organisation spatiale de l'isopédine des écailles du coelacanthe (Latimeria chalumnae SMITH). Comptes Rendus de l’Académie des Sciences, 287: 487–489.
GRADY, J . E. 1970. Tooth development in Latimeria chalumnae (SMITH). Journal of Morphology, 132: 377–388.
HADIATY, R. K. and RACHMATIKA, I. 2003. Morphological study of the scales of Latimeria menadoensis POUYAUD et al., 1999. Treubia (A Journal on Zoology of the Indo-Australian Archipelago), 33: 1–11.
HOBDELL, M. H. and MILLER, W. A. 1969. Radiographic
anatomy of the teeth and tooth supporting tissues of Latimeria chalumnae. Archives of Oral Biology, 14: 855–858.
HUREAU J.-Cl. and OZOUF, C. 1977. Détermination de l’âge et croissance du cœlacanthe Latimeria chalumnae SMITH, 1939 (Poisson, Crossoptérygien, Coelacanthidé). Cybium, 2: 129–137.
JANVIER, P. 1996. Early Vertebrates. Clarendon Press, Oxford, 393 pp.
MEUNIER, F. J. 1979. Etude histologique et microradiographique du cartilage hémal de la vertèbre de la carpe, Cyprinus carpio L. (Pisces, Teleostei, Cyprinidae). Acta Zoologica, 60: 19–31.
MEUNIER, F. J. 1980. Les relations isopédine-tissu osseux dans le post-temporal et les écailles de la ligne latérale de Latimeria chalumnae (SMITH). Zoologica Scripta, 9: 307–317.
MEUNIER, F. J. and ZYLBERBERG, L. 1999. The structure of the external layer and of the odontodes of scales in Latimeria chalumnae (Sarcopterygii, Actinistia, Coelacanthidae) revisited using scanning and transmission electron microscopy. In: SÉRET, B. and SIRE, J.-Y. (eds.), Proc. 5th Indo-Pacific Fish Conf., Nouméa, 1997, Soc. Fr. Ichtyol., Paris, pp. 109–116.
MEUNIER, F. J., ERDMANN, M. V., FERMON, Y. and CALDWELL, R. L. 2008. Can the comparative study of the morphology and histology of the scales of Latimeria menadoensis and L. chalumnae (Sarcopterygii, Actinistia, Coelacanthidae) bring new insight on the taxonomy and the biogeography of recent coelacanthids? Geological Society, London, Special Publications, 295: 351–360.
MEUNIER, F. J., BRITO, P. M. and LEAL, M.-E. 2013. Quelques données morphologiques et histologiques sur la structure de la plaque dentaire linguale d’Arapaima gigas (Osteoglossidae; Teleostei). Cybium, 37: 263–271
MEUNIER, F. J., MONDEJAR-FERNANDEZ, J., GOUSSARD, F., CLÉMENT, G. and HERBIN, M. 2015. Presence of plicidentine in the oral teeth of Latimeria chalumnae (Sarcopterygii; Actinistia; Coelacanthidae). Journal of Structural Biology, 190: 31–37.
MILLER, W. A. 1979. Observations on the structure of mineralized tissue of the coelacanth, including scales and their associate odontodes. Occasional Papers of the Californian Academy of Sciences, 134: 68–78.
MILLER, W. A. and HOBDELL, M. H. 1968. Preliminary report on the histology of the dental and paradental tissues of Latimeria chalumnae (SMITH) with a note on tooth replacement. Archives of Oral Biology, 13: 1289–1291.
MILLOT, J. and ANTHONY, J. 1958. Anatomie de Latimeria chalumnae. T. 1. Squelette, muscles et formations de soutien. CNRS Edr., Paris. 122 pp., 80 pls.
MILLOT, J., ANTHONY, J. and ROBINEAU, D. 1978. Anatomie de Latimeria chalumnae. T. 3. Appareil digestif, téguments, écailles. CNRS Edr., Paris. pp. 7–28, 135–139.
MONDEJAR-FERNANDEZ, J. 2018. On cosmine: its origins, b io logy and impl ica t ions for sarcopterygian interrelationships. Cybium, 42: 41–65.
ØRVIG T. 1951. Histologic studies of Placoderms and fossil Elasmobranchs. 1: The endoskeleton, with remarks on the hard tissues of lower vertebrates in general. Arkiv för Zoologi, 2: 321–454, 8 pl.
ØRVIG T. 1968. The dermal skeleton; general considerations. In: ØRVIG T. (ed.), Current problems of lower vertebrate phylogeny. Proceedings of the fourth Nobel Symposium, Stockholm, 1967, J. Wiley, New-York. pp. 373–397.
PARMENTIER, E., COMPÈRE, P., CASADEWALL, M., FONTENELLE, N., CLOOTS, R. and HENRIST, C. 2008. The rocker bone: a new kind of mineralized tissue? Cell and Tissue Research, 334: 67–79.
PEGUETA, V. P. 1968. Enchondral ossification in Latimeria chalumnae SMITH. Dopovidi Akademii Nauk Ukrainia SSR., 7: 653–656 (in Russian; English abstr.).
POUYAUD, L., WIRJOATMODIO, S., RACHMATIKA, L., TJAKRAWIDJAJA, A., HADIATY, R. K. and HADIE, W. 1999. Une nouvelle espèce de coelacanthe, preuves génétiques et morphologiques. Comptes Rendus de l’Académie des Sciences, 322: 261–267.
ROMER, A.S. 1937. The braincase of the carboniferous crossopterygian Megalichthys nitidus. Bulletin of the Museum of Comparative Zoology, 824: 1–73.
ROMER, A.S. 1942. Cartilage as an embryonic adaptation. American Naturalist, 76: 394–404.
ROUX, G. H. 1942. The microscopic anatomy of the Latimeria scale. South African Journal of Medical Sciences, 7: 1–18.
SASAGAWA, I., ISHIYAMA, M. and KODERA, H. 1984. Fine structure of the pharyngeal teeth in the coelacanth fish (Latimeria chalumnae). In: FEARNHEAD, R. W. and SUGA, S. (eds.), Tooth enamel IV, Elsevier Science Publishers, Amsterdam, pp. 462–466.
SCHULTZE, H.-P. 1969. Die Faltenzähne der Rhipidistiden Crossopterygier, der Tetrapoden und der Actinopterygi-er-Gattung Lepisosteus nebst einer Beschreibung der Zahnstruktur von Onychodus (Struniiformer Crossoptery-gier). Palaeontologica Italica, New Series, 35(65): 63–137.
SCHULTZE, H.-P. 1970. Folded teeth and the monophyletic origin of Tetrapods. American Museum Novitates, 2408: 1–10.
SCHULTZE, H.-P. 2018. Hard tissues in fish evolution: history and current issues. Cybium, 42(1): 29–39.
SHELLIS, R. P. and POOLE, D. F. G. 1978. The structure of the dental hard tissues of the coelacanthid fish Latimeria chalumnae SMITH. Archives of Oral Biology, 23: 1105–1113.
SIRE, J.-Y. and HUYSSEUNE, A. 2003. Formation of dermal skeletal and dental tissues in fish: a comparative and evolutionary approach. Biological Review, 78: 219–249.
SMITH, J. L. B. 1939. A living fish of the Mesozoic type. Nature, 143: 455–456.
SMITH, J. L. B. 1940. A living coelacanth fish from South Africa. Transactions of the Royal Society of South Africa, 28: 1–106, 44 Pls.
SMITH, M. M. 1978. Enamel in the oral teeth of Latimeria chalumnae (Pisces: Actinistia): a scanning electron microscopic study. Journal of Zoology, 185: 355–369.
SMITH, M. M. 1979. Scanning electron microscopy of odontodes in the scales of coelacanth embryo, Latimeria chalumnae SMITH. Archives of Oral Biology, 24: 179–183.
SMITH, M. M., HOBDELL, M. H. and MILLER, W. A. 1972. The structure of the scales of Latimeria chalumnae. Journal of Zoology, London, 167: 501–509.
THOMSON, K. S. 1969. The biology of the lobe-finned fishes.
Biological Reviews, 44: 91–154. UYENO, T. and YABUMOTO, Y. 2007. Origin of extant
coelacanths. The Coelacanth, Fathom the Mystery 2007. Aquamarine Fukushima, pp. 24–26.
ZYLBERBERG, L. and MEUNIER, F. J. 2008. New data on the structure and the chondrocyte populations of the haemal cartilage of abdominal vertebrae in the adult carp Cyprinus carpio (Teleostei, Ostariophysii, Cyprinidae). Cybium, 32: 225–239.
ZYLBERBERG, L., GÉRAUDIE, J., MEUNIER, F. J., & SIRE J. Y., 1992. Biomineralization in the integumental skeleton of the living lower Vertebrates. In: HALL, B. K. (ed), Bone, Volume 4, CRC Press, Boca Raton, pp. 171–224.
The skeleton and the mineralized tissues of the living coelacanthsFrançois J. MEUNIER, Camila CUPELLO and Gaël CLÉMENT
Fig. 7. Latimeria menadoensis and L. chalumnae. Ground sections of scales. A, L. menadoensis. Longitudinal section; X-ray. The superficial bony layer (bo) and the odontodes (arrow-heads) are mineralized whereas the basal plate (BP) totally lacks mineralization (asterisks). Vascular canals are present within the superficial bony layer. The arrow on the upper right indicates the direction of the head. B–C, L. chalumnae. Transversal sections; respectively polarized light and X-ray. B, The basal plate is made of stratified layers of collagenous fibres (arrow). The arrow-heads point to the superficial odontodes. C, The superficial bony layer and the odontodes (arrow-heads) are mineralized whereas the basal plate (BP) totally lacks mineralization (asterisks). D, L. chalumnae. Longitudinal section; X-ray. The superficial bony layer (bo) and the odontodes (arrow-head) are mineralized whereas the basal plate (BP) totally lacks mineralization (asterisks). (af = anterior field; pf = posterior field). E, L. menadoensis. Broken frozen scale showing the different fibres directions in the successive layers of the basal plate. The rotation of the fibres directions from one layer to another characterizes a plywood-like organization. The arrow points to the crests of the superficial layer. (A, B, C: scale = 1 mm; D: scale = 2 mm; E: scale = 200 µm). (B, C, D after CASTANET et al., 1975; A, E after MEUNIER et al., 2008).
Fig. 8. Latimeria chalumnae. Cartilage histology (after FRANCILLON et al., 1975). A, MECKEL’s cartilage (thin histological section; Hemalun-PIC staining). An erosive vascular cavity (black asterisk) is separated from the medullary cavity (mc) by a layer of enchondral bone (eb). Elongate chondrocytes (arrow-heads) are seen in the cartilage. (scale = 50 µm). B, Scapulocoracoid (ground section 3 mm thick; x-ray). The cartilage (ca) is invaded by resorption canals (white asterisks), their wall is covered by thin bony layers, here clearly visible in white colour. The white arrow-heads point to the erosion front of the cartilage. (scale = 1 mm). C, Scapulocoracoid (thin histological section; Stoelzner staining). In the cartilage (ca), the vascular cavities (vc), resulting from a chondroclastic activity, are bordered by thin bony lamellae here visible in black colour (arrow-heads). (scale = 250 µm).

type according to the definition of MEUNIER et al. (2013). This organization is less complex than the three other plicidentine types (polyplocodont, eusthenodont and dendrodont), which are described in most of extinct and extant Sarcopterygii (SCHULTZE, 1969, 1970).The bony plates of the lung
The lung of Latimeria is reduced to a short oesophagus diverticulum (CUPELLO et al., 2015) surrounded by scattered small and thin ovoid plates (Fig. 9A, B). These plates cover the surface of the vestigial lung, and are composed of a fibrous matrix with fibrocytes: it is a cellular bony tissue (CUPELLO et al., 2017). Each plate is enclosed in a membranous bag. These plates look fragile in their middle plane since a central weakness zone opened on the microtome knife, creating an artefactual lumen (Fig. 9C) (CUPELLO et al., 2017). The fibres are collagenous and organized in superposed layers. The cells are true osteocytes (Fig. 9E) with a more or less star-shaped outline, and send cytoplasmic extensions in the thickness of the plate (Fig. 9D, E). Rare lining cells (osteoblasts) are
present on the surface of the plates, and conjunctive fibres (SHARPEY’s fibres) penetrate within the plate (Fig. 9C). The extracellular matrix shows various staining intensities with concentric and parallel (Fig. 9D) or globular shaped lines. These lines are considered to be Liesegang lines that characterize an active spheritic mineralizing process. The lung plates of Latimeria are thus true bony plates, and are homologous with the large bony plates known in the abdominal cavity of fossil actinistians (BRITO et al., 2010; CUPELLO et al., 2017). The calcified walls that surrounded the lung of the Cretaceous coelacanth Axelrodichthys are made of large osseous plates of various thickness (BRITO et al., 2010), as those of other fossil coelacanths (CLEMENT, 1999, 2005). The bony tissue of these plates is a vascularized cellular bone with more or less large vascular cavities and some internal remodelling (BRITO et al., 2010). Moreover the mineralization of the lung ossified plates in Axelrodichthys is spheritic as in Latimeria’s plates (CUPELLO et al., 2017). So there is a continuity of the histological structure of the lung plates in the coelacanths during their
evolution, but with an important reduction of the bony plates in Latimeria linked to the vestigial state of its lung (CUPELLO et al., 2015, 2019).
CONCLUSION
This overview of eighty years (1938–2018) of histological work on Latimeria skeletal tissues allows some anatomical and evolutionary considerations. It can be enlightened a drastic reduction of endochondral ossification during the long evolutionary history of coelacanths. The persistence of large volume of cartilage in the endoskeleton at adult stage can be compared to the “little bone and considerable cartilage” that characterize the skeleton of a number of demersal notothenioid telostean fishes (EASTMAN et al., 2014).
A processus of spheritic mineralization has been recently highlighted in various skeletal elements of Latimeria, by the presence of globular dentine in teeth, in odontodes of the tegumentary skeleton (scales, fin rays), in scales at the interface between the external layer and the basal plate, as well as in lung bony plates. The spheritic mineralization (i.e., a radiating arrangement of hydroxyapatite crystals and of the organic matrix) in vertebrate skeletal tissues is considered as a precursor of inotropic mineralization (specific interactions between collagen fibrils and the mineral phase) that possibly represents a derived evolutionary stage of calcification mechanisms (ØRVIG, 1951, 1968; FRANCILLON-VIEILLOT et al., 1990; ZYLBERBERG et al., 1992). The simultaneous presence of both mineralization processes (spheritic and inotropic) in the mineralized tissues of Latimeria and in other osteichthyans such as teleosteans (ZYLBERBERG and MEUNIER, 2008) is however questionable. The mineralized spherules present at the limit between the external layer and the basal plate of Latimeria scales are considered to be the product of an inotropic mineralization (MEUNIER and ZYLBERBERG, 1999). It thus appears that the state of the mineralized spherules observed in the various skeletal tissues of Latimeria, including the lung bony plates, must be studied with adapted ultrastructural techniques in order to test their true origin: spheritic or inotropic mineralization?
ACKNOWLEDGEMENTS
This contribution is linked to an international roundtable on coelacanths evolution held at the Kitakyushu Museum and at the Aquamarine Fukushima in August 2017. We would like to thank the anonymous reviewers for their constructive comments and valuable suggestions. C.C. was partially financed by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brasil (CAPES) – Finance Code 001 (Programa Nacional de Pós Doutorado-PNPD).
INTRODUCTION
The first study of fossil coelacanths was done by AGASSIZ (1833–44) and was followed by a lot of papers with the discovery of new fossils (see reviews of JANVIER, 1996 and FOREY, 1998). Fossil coelacanths constitute an important monophyletic group composed of more than forty genera, eighty species, and belong to the Sarcopterygii (ANDREWS, 1973; JANVIER, 1996; FOREY, 1998; UYENO and YABUMOTO, 2007; and many others). For a century coelacanths were considered extinct since the end of the Cretaceous time (Fig. 1).
With the discovery of a living coelacanth in 1938 (SMITH, 1939) the scientific community was soon highly interested to compare the incomplete fossil sarcopterygian fishes dataset to the recent anatomical and biological organization of Latimeria chalumnae. Many authors (see THOMSON, 1969; MILLOT and ANTHONY, 1958; MILLOT et al., 1978 FOREY, 1984, 1998; JANVIER, 1996) focused their work on Latimeria as a key-taxon to test or propose various hypotheses on the origin and evolutionary history of the early tetrapods and sarcopterygii in general, and to infer their possible biological characteristics. The fossil sarcopterygian fishes are known by their fossilized mineralized tissues, essentially their bones, scales and teeth. Together with the extant lungfishes, especially Neoceratodus, Latimeria offered a unique access to the skeleton as a whole: with both mineralized and unmineralized skeletal tissues. Here, we aim to present an overview of the last eighty years (1938–2018) of histological studies of Latimeria’s skeleton.
Since the discovery of the Raja Laut (“king of the sea” ), off Sulawesi Island in Indonesia (ERDMANN et al., 1998, 1999), the genus Latimeria comprises two species: L. chalumnae SMITH 1939 and L. menadoensis POUYAUD et al., 1999. However our knowledge of the anatomy and biology of the living coelacanths is so far essentially based on L. chalumnae.
THE LATIMERIA SKELETON
The first anatomical observations on the extant coelacanth were done by SMITH (1940). The anatomy of the Latimeria skeleton has been later described in detail by MILLOT and ANTHONY (1958). The living coelacanth shows a general organization similar to that of the other actinistian fishes (THOMSON, 1969; JANVIER, 1996; FOREY, 1998). The skeleton can be divided in various parts (Fig. 2): skull, axial skeleton, paired and unpaired fins, tegumentary skeleton (scales). The skull, axial skeleton and fins are constituted of endoskeletal elements that are overlain by exoskeletal elements. In the extant Latimeria the endoskeleton of the skull shows an important regression of the bones, especial ly in the neurocranium, which are replaced by cartilaginous tissues, as in its Mesozoic fossil actinistian relatives (ROMER, 1937, 1942; MILLOT and ANTHONY, 1958; BJERRING, 1973; FOREY, 1998). In the same way, the endoskeleton of paired and unpaired fins is essentially constituted of four axial cartilaginous elements (Fig. 2; MILLOT and ANTHONY, 1958), with pre and post-axial
elements and eventually minute superficial endochondral ossification (CASTANET et al., 1975). The axial skeleton is composed of the notochord which is coated by an unmineralized fibrillary sheath and is totally deprived of well developed vertebrae due to the lack of ossified centra, although some discrete neural and haemal arches develop in the anterior part of the notochord (MILLOT and ANTHONY, 1958). The anterior neural and haemal spines are relatively short but they progressively increase in length posteriorly. Importantly neural and haemal spines are composed of perichondral bone surrounding a cartilaginous core (Fig. 2).
The scales belong to the exoskeleton. In both extant coelacanth species they are of elasmoid type, composed of an upper external layer also called “external ornamented layer”, and of a lower thicker layer, called the basal plate, which is stratified and almost totally unmineralized (Fig. 3). The upper external layer is ornamented with radial crests. In the posterior area of the scale these radial reliefs are overlain by numerous denticulations, the odontodes, which can be superposed (Fig. 3). The anterior area of the external layer, which is covered by the anterior scales, shows crests with irregularities that have been interpreted as growth marks and tentatively used for ageing coelacanths (HUREAU and OZOUF, 1977; MILLER, 1979).
The tooth plates of the buccal cavity support series of teeth of various sizes (Fig. 4). They range in three morphotypes: the fangs (7–10 mm in height), middle-sized teeth (3–4 mm in height) and rounded tubercles (MILLOT and ANTHONY, 1958). The teeth of the two first categories are conical and sharp whereas those of the third category have an obtuse tip (Fig. 5). Fangs of Latimeria are inserted into a socket (MILLOT and ANTHONY, 1958; HOBDELL and MILER, 1969; CASTANET et al., 1975) and they have a smooth external surface (MILLOT et al., 1978; MEUNIER et al., 2015).
A particularity of the exoskeleton of Latimeria is the abundance of odontodes at the surface of various skull bones
42 43
(MILLOT and ANTHONY, 1958; BERNHAUSER, 1961), on the fin rays (CASTANET et al., 1975) and on the posterior (free) area of scales (Fig. 3) (MILLOT and ANTHONY, 1958; CASTANET et al., 1975; HADIATY and RACHNATIKA, 2003; MEUNIER et al., 2008).
Among the various components of the whole skeleton, and apart the SMITH’s pioneer work (SMITH, 1940), numerous studies have been carried out on the histological organization of scales (ROUX, 1942; SMITH et al., 1972; CASTANET et al., 1975; GIRAUD et al., 1978a; MILLER, 1979; SMITH, 1979; MEUNIER, 1980; MEUNIER and ZYLBERBERG, 1999; HADIATY and RACHMATIKA, 2003; MEUNIER et al., 2008) and teeth (MILLER and HOBDELL, 1968; GRADY, 1970; HOBDELL and MILLER, 1969; CASTANET et al., 1975; SHELLIS and POOLE, 1978;
SMITH, 1978; SASAGAWA et al., 1984; MEUNIER et al., 2015). Contrary to the superficial skeleton, there are clearly less histological studies of the skeletal bony elements (FRANCILLON et al., 1975). Meanwhile, in addition to the usual skeletal elements, specific bony tissues have been described in fossil (BRITO et al., 2010) and extant coelacanths (CUPELLO et al., 2015, 2017), such as the mineralized plates surrounding the lung. These bony plates in actinistians do not belong to the skeleton sensu stricto. They are specific bony specializations in relation to the lung (see below), as is the “rocker bone” in relation to the gas bladder of some ophidiiform teleosts (PARMENTIER et al., 2008). Despite the name, the rocker bone is not true bony tissue (PARMENTIER et al., 2008), contrary to
the bony plates of fossil and extant coelacanths (BRITO et al., 2010; CUPELLO et al., 2017).
ADULT LATIMERIA SKELETAL TISSUES
Bony tissuesCortical areas of the dermal skeleton are constituted of
primary periosteal bony t issue. I ts mineralization is heterogeneous and generally marked by series of growth lines, probably due to alternative physiological cycles linked to seasonal variations (Fig. 6). The cortical primary bone shows vascular canals and numerous SHARPEY’s fibres (Fig. 6B). The centre of these bones is frequently made of a spongiosa with vascular cavities surrounded by secondary bony deposits that result from remodelling processes (Fig. 6). This secondary bone is separated from the primary one by reversal cementing lines that are hypermineralized.
Bone remodelling in adult specimens can be more or less developed according to a bone and within a given bone. Primary bone is destroyed by osteoclasts and the deposition process of the secondary bone results from the activity of osteoblasts that have replaced the osteoclasts. The secondary bone does generally not replaced the whole primary bone areas, which can still be recognizable by the presence of
SHARPEY’s fibres and/or growth marks (Fig. 6).The dermal fin rays that sustain paired and unpaired fins
in Latimeria are true lepidotrichia (CASTANET et al., 1975). They are made of two parallel opposite gutter-shaped hemirays, each of which being a series of hemisegments articulated by a collagenous ligament. Each hemisegment is made of a mineralized collagenous fibrillary matrix with embedded osteocytes. The hemisegments are sometimes fused at the basal part of the rays due to centrifugal deposition of bony laminae around the primary articulation, and eventually to the mineralization of the ligament (see figs. 19, 20 in CASTANET et al., 1975). Odontodes observed on the external convex surface of the rays are similar to those present on the dermal bones of the skull and scales. The distal extremity of the lepidotrichia is overlapped by actinotrichia (GÉRAUDIE and MEUNIER, 1980) that are long tapered rods of elastoidin , a fibrous protein of collagenous nature (FAURÉ-FREMIET, 1936; GARRAULT, 1936). The presence of lepidotrichia with distal actinotrichia in Latimeria, as in the fins of Actinopterygii, is
regarded as a plesiomorphic character for Osteichthyes (GÉRAUDIE and MEUNIER, 1980, 1984).
The upper layer of the Latimeria scale is relatively thin and it is constituted of bony tissue with embedded osteocytes, whereas the basal plate is much thicker and composed of numerous strata made of thick collagenous fibres (Fig. 7). Between two collagenous layers there are star-shaped cells (SMITH et al., 1972, pl. VI; CASTANET et al., 1975, fig. 12), the elasmocytes, which cytoplasmic processes insert between the collagenous fibres. These fibres are set in a very specific network. In each layer, the fibres are parallel to each other and the direction of the fibres change from a layer to another one. This regular organization results in a spatial arrangement termed “twisted plywood” (GIRAUD et al., 1978a,b). The rotation of fibres direction from one layer to the next has a mean angle of 27° (GIRAUD et al., 1978a). This organization is found in each scale of L. chalumnae. In L. menadoensis, the basal plate of the scales is also a twisted plywood but its rotation angle seems to be slightly less regular (MEUNIER et al.,
2008). The basal plate in both species is unmineralized (Fig. 7) excepted at the contact between the superficial layer and the basal plate, where spheritic mineralized granules are seen in the very first layers of the basal plate (MEUNIER, 1980; MEUNIER and ZYLBERBERG, 1999; MEUNIER et al., 2008). There is no pore-canal system so Latimeria’s scales are not cosmoid, contrary to most of palaeozoic sarcopterygian fishes. Due to the presence of an unmineralized stratified basal plate, the scales of Latimeria are defined as elasmoid-like scales. However the plywood-like organization is considered to be homologous with the bony basal plate of cosmoid scales of extinct sarcopterygian fishes (MEUNIER, 1980; SIRE and HUYSSEUNE, 2003; MONDEJAR, 2018; SCHULTZE, 2018).Cartilages
The cartilaginous tissues in Latimeria are characterized by long chondrocytes (Fig. 8) contrary to those of teleostean fishes that are relatively spherical in shape (MEUNIER, 1979; ZYLBERBERG and MEUNIER, 2008). Cartilage tissues can show an endochondral ossification process. This phenomenon has been studied respectively on the urohyal (PEGUETA, 1968), the MECKEL’s cartilage and the proximal piece of the pectoral girdle (FRANCILLON et al., 1975). The endochondral ossification is relatively limited when it occurs and the volume of enchondral bone remains reduced to thin bony layers (Fig. 8). The cartilage is destroyed by chondroclasts in areas where endochondral ossification occurs. Serial chondrocytes, known at the origin of the calcified cartilage, seem to be lacking in the endochondral ossification process in Latimeria (FRANCILLON et al., 1975). This constitutes a true difference with teleostean endochondral ossifications (MEUNIER, 1979; ZYLBERBERG and MEUNIER, 2008). Tooth tissues
The three tooth morphotypes described in Latimeria are fangs (7–10 mm in height), middle-sized teeth (3–4 mm in height) and rounded tubercles (MILLOT and ANTHONY, 1958). These teeth and tubercles are constituted of a cone of orthodentine set around a large pulp cavity and overlain by an external hypermineralized layer considered as true enamel (GRADY, 1970; CASTANET et al., 1975; SHELLIS and POOLE, 1978; SMITH, 1978; SASAGAWA et al., 1984). Various studies have revealed the presence of globular dentine (Fig. 5) at the periphery of the first third of the tooth (MILLER and HOBDELL, 1968; HOBDELL and MILLER, 1969; SHELLIS and POOLE, 1978; SASAGAWA et al., 1984). This typical histological organization characterizes the various teeth of the bucco-pharyngeal cavity as well as the various odontodes of the dermal bony plates, the fin rays and the scales (CASTANET et al., 1975).
In the fang the inner wall of the pulp cavity displays a series of plies (Fig. 5; MEUNIER et al., 2015). These plies start at the base of the tooth and reach at least the first third to the half of the tooth towards its tip (Fig. 5). Such plies are also present but less developed in the small caniniform teeth (MEUNIER et al., 2015). These dentine plies in the pulp cavity characterizes a plicidentine organization of orthodentine type. These primary plies correspond to a simplexodont plicidentine
REFERENCES
AGASSIZ, L. 1833–44. Recherches sur les Poissons fossiles. Imprimerie Petitpierre, Neuchâtel. [5 volumes and atlas.]
ANDREWS, S. M. 1973. Interrelationships of crossopterygians. In: GREENWOOD, P. H., MILES, R. S. and PATTERSON, C. (eds.), Interrelationships of Fishes, Acad. Press, London, pp. 137–177.
BERNHAUSER, A. 1961. Zur Knochen- und Zah, histology von Latimeria chalumnae Smith und einiger Fossilformen. Sber. öst. Akad. Wiss., 170: 119–137.
BJERRING, H. C. 1973. Relationships of coelacanthiforms. In: GREENWOOD P. H., MILES R. S. and PATTERSON C. (eds.), Interrelationships of Fishes, Acad. Press, London, pp. 179–205.
BRITO, P. M., MEUNIER, F. J., CLÉMENT, G. and GEFFARD-KURIYAMA, D. 2010. The histological structure of the calcified lung of the fossil coelacanth Axelrodichthys araripensis (Actinistia: Mawsoniidae). Palaeontology, 53: 1281–1290.
CASANE, D. and LAURENTI, P. 2013. Why coelacanths are not ‘living fossils’. Bioessays, 35: 332–338. DOI 10.1002/bies.201200145.
CASTANET, J., MEUNIER, F., BERGOT, C. and FRANÇOIS, Y. 1975. Données pré l iminaires sur les s t ructures histologiques du squelette de Latimeria chalumnae. I - Dents, écailles, rayons de nageoires. In: Coll. Inter. CNRS, "Problèmes actuels de Paléontologie. Evolution des Vertébrés", Paris, 4–9 Juin 1973, 159–168, 4 Pls.
CLEMENT, G. 1999. The actinistian (Sarcopterygii) Piveteauia madagascarensis LEHMAN from the Lower Triassic of Northeastern Madagascar: a redescription on the basis of new material. Journal of Vertebrate Paleontology, 19: 234–242.
CLEMENT, G. 2005. A new coelacanth (Actinist ia, Sarcopterygii) from the Jurassic of France, and the question of the closest relative fossil to Latimeria. Journal of Vertebrate Paleontology, 25: 481–491.
CUPELLO, C., BRITO, P. M., MEUNIER, F. J., HERBIN, M., JANVIER, P., DUTEL, H. & CLÉMENT, G. 2015. Allometric growth in the extant coelacanth lung during ontogenetic development. Nature Communications, 6: 8222. DOI: 10.1038/ncomms9222.
CUPELLO, C., CLEMÉNT, G., MEUNIER, F. J., HERBIN, M., YABUMOTO, Y. and BRITO, P. M. 2019. The long-time adaptation of coelacanths to moderate deep water: reviewing the evidences. Bulletin of Kitakyushu Museum of Natural History and Human History Series A (Natural History), 17: 29–35.
CUPELLO, C., MEUNIER, F. J., HERBIN, M., JANVIER, P., CLÉMENT, G. and BRITO, P. M. 2017. The homology and funct ion of the lung pla tes in extant and foss i l coelacanths. Scientific Reports , 7 : 9244. DOI: 10.1038/s41598-017-09327-6.
EASTMAN, J. T., WITMER, L. M., RIDGELY, R. C. and KUHN, K.
L. 2014. Divergence in skeleta l mass and bone morphology in antarctic notothenioid fishes. Journal of Morphology, 275: 841–861. DOI: 10.1002/jmor.20258
ERDMANN, M. V., CALDWELL, R. L. and MOOSA, M. K. 1998. Indonesian “king of the sea” discovered. Nature, 395: 335.
ERDMANN, M. V., CALDWELL, R. L., JEWETT, S. L. and TJAKRAWIDJAJA, A. 1999. The second recorded living coelacanth from north Sulawesi. Environmental Biology of Fishes, 34: 445–451.
FAURÉ-FREMIET, M. 1936. La structure des fibres d’éastoidine. Archives d'Anatomie Microscopique, 32: 249–269.
FOREY, P. L.- 1984. The coelacanth as a living fossil. In: ELREDGE N. and STANLEY S.M. (eds.), Living fossils. Springer Verlag, Berlin, pp. 166–169.
FOREY, P. L.- 1998. History of the Coelacanth Fishes. Chapman and Hall, London, 419 pp.
FRANCILLON, H., MEUNIER, F., NGO TUAN PHONG, D. and RICQLÈS, A. (DE). 1975. Données préliminaires sur les structures histologiques du squelette de Latimeria chalumnae. I I- Tissus osseux et cartilages. In: Coll. Inter. CNRS, "Problèmes actuels de Paléontologie. Evolution des Vertébrés", Paris, 4–9 Juin 1973, Centre National de la Recherche Scientifique, pp. 169–174, 2 pls.
FRANCILLON-VIEILLOT H., BUFFRENIL V. DE, CASTANET J., GÉRAUDIE J., MEUNIER FJ., SIRE J-Y., ZYLBERBERG L., a n d R I C Q L È S A . D E 1 9 9 0 . M i c r o s t r u c t u r e a n d mineralization of vertebrate skeletal tissues. In: CARTER, J. G. (ed.), Skeletal Biomineralization: Patterns, Processes and Evolutionary Trends. Vol. I , Van Nostrand, New York, pp. 471–530
GARRAULT, H., 1936. Développement des fibres d’élastoidine (actinotrichia) chez les salmonides. Archives d'Anatomie Microscopique, 130: 105–137.
GÉRAUDIE, J. and MEUNIER, F. J. 1980. Elastoidin actinotrichia in coelacanth fins: a comparative study with teleosts. Tissue and Cell, 12: 637–645.
GÉRAUDIE, J. and MEUNIER, F. J. 1984. Structure and comparative morphology of camptotrichia of lungfish fins. Tissue and Cell, 16: 217–236.
GIRAUD, M. M., CASTANET, J., MEUNIER, F. J. and BOULIGAND, Y. 1978a. The fibrous structure of coelacanth scales: a twisted "plywood". Tissue and Cell, 10: 671–686.
GIRAUD, M. M., CASTANET, J., MEUNIER, F. J. and BOULIGAND, Y. 1978b. Organisation spatiale de l'isopédine des écailles du coelacanthe (Latimeria chalumnae SMITH). Comptes Rendus de l’Académie des Sciences, 287: 487–489.
GRADY, J . E. 1970. Tooth development in Latimeria chalumnae (SMITH). Journal of Morphology, 132: 377–388.
HADIATY, R. K. and RACHMATIKA, I. 2003. Morphological study of the scales of Latimeria menadoensis POUYAUD et al., 1999. Treubia (A Journal on Zoology of the Indo-Australian Archipelago), 33: 1–11.
HOBDELL, M. H. and MILLER, W. A. 1969. Radiographic
anatomy of the teeth and tooth supporting tissues of Latimeria chalumnae. Archives of Oral Biology, 14: 855–858.
HUREAU J.-Cl. and OZOUF, C. 1977. Détermination de l’âge et croissance du cœlacanthe Latimeria chalumnae SMITH, 1939 (Poisson, Crossoptérygien, Coelacanthidé). Cybium, 2: 129–137.
JANVIER, P. 1996. Early Vertebrates. Clarendon Press, Oxford, 393 pp.
MEUNIER, F. J. 1979. Etude histologique et microradiographique du cartilage hémal de la vertèbre de la carpe, Cyprinus carpio L. (Pisces, Teleostei, Cyprinidae). Acta Zoologica, 60: 19–31.
MEUNIER, F. J. 1980. Les relations isopédine-tissu osseux dans le post-temporal et les écailles de la ligne latérale de Latimeria chalumnae (SMITH). Zoologica Scripta, 9: 307–317.
MEUNIER, F. J. and ZYLBERBERG, L. 1999. The structure of the external layer and of the odontodes of scales in Latimeria chalumnae (Sarcopterygii, Actinistia, Coelacanthidae) revisited using scanning and transmission electron microscopy. In: SÉRET, B. and SIRE, J.-Y. (eds.), Proc. 5th Indo-Pacific Fish Conf., Nouméa, 1997, Soc. Fr. Ichtyol., Paris, pp. 109–116.
MEUNIER, F. J., ERDMANN, M. V., FERMON, Y. and CALDWELL, R. L. 2008. Can the comparative study of the morphology and histology of the scales of Latimeria menadoensis and L. chalumnae (Sarcopterygii, Actinistia, Coelacanthidae) bring new insight on the taxonomy and the biogeography of recent coelacanthids? Geological Society, London, Special Publications, 295: 351–360.
MEUNIER, F. J., BRITO, P. M. and LEAL, M.-E. 2013. Quelques données morphologiques et histologiques sur la structure de la plaque dentaire linguale d’Arapaima gigas (Osteoglossidae; Teleostei). Cybium, 37: 263–271
MEUNIER, F. J., MONDEJAR-FERNANDEZ, J., GOUSSARD, F., CLÉMENT, G. and HERBIN, M. 2015. Presence of plicidentine in the oral teeth of Latimeria chalumnae (Sarcopterygii; Actinistia; Coelacanthidae). Journal of Structural Biology, 190: 31–37.
MILLER, W. A. 1979. Observations on the structure of mineralized tissue of the coelacanth, including scales and their associate odontodes. Occasional Papers of the Californian Academy of Sciences, 134: 68–78.
MILLER, W. A. and HOBDELL, M. H. 1968. Preliminary report on the histology of the dental and paradental tissues of Latimeria chalumnae (SMITH) with a note on tooth replacement. Archives of Oral Biology, 13: 1289–1291.
MILLOT, J. and ANTHONY, J. 1958. Anatomie de Latimeria chalumnae. T. 1. Squelette, muscles et formations de soutien. CNRS Edr., Paris. 122 pp., 80 pls.
MILLOT, J., ANTHONY, J. and ROBINEAU, D. 1978. Anatomie de Latimeria chalumnae. T. 3. Appareil digestif, téguments, écailles. CNRS Edr., Paris. pp. 7–28, 135–139.
MONDEJAR-FERNANDEZ, J. 2018. On cosmine: its origins, b io logy and impl ica t ions for sarcopterygian interrelationships. Cybium, 42: 41–65.
ØRVIG T. 1951. Histologic studies of Placoderms and fossil Elasmobranchs. 1: The endoskeleton, with remarks on the hard tissues of lower vertebrates in general. Arkiv för Zoologi, 2: 321–454, 8 pl.
ØRVIG T. 1968. The dermal skeleton; general considerations. In: ØRVIG T. (ed.), Current problems of lower vertebrate phylogeny. Proceedings of the fourth Nobel Symposium, Stockholm, 1967, J. Wiley, New-York. pp. 373–397.
PARMENTIER, E., COMPÈRE, P., CASADEWALL, M., FONTENELLE, N., CLOOTS, R. and HENRIST, C. 2008. The rocker bone: a new kind of mineralized tissue? Cell and Tissue Research, 334: 67–79.
PEGUETA, V. P. 1968. Enchondral ossification in Latimeria chalumnae SMITH. Dopovidi Akademii Nauk Ukrainia SSR., 7: 653–656 (in Russian; English abstr.).
POUYAUD, L., WIRJOATMODIO, S., RACHMATIKA, L., TJAKRAWIDJAJA, A., HADIATY, R. K. and HADIE, W. 1999. Une nouvelle espèce de coelacanthe, preuves génétiques et morphologiques. Comptes Rendus de l’Académie des Sciences, 322: 261–267.
ROMER, A.S. 1937. The braincase of the carboniferous crossopterygian Megalichthys nitidus. Bulletin of the Museum of Comparative Zoology, 824: 1–73.
ROMER, A.S. 1942. Cartilage as an embryonic adaptation. American Naturalist, 76: 394–404.
ROUX, G. H. 1942. The microscopic anatomy of the Latimeria scale. South African Journal of Medical Sciences, 7: 1–18.
SASAGAWA, I., ISHIYAMA, M. and KODERA, H. 1984. Fine structure of the pharyngeal teeth in the coelacanth fish (Latimeria chalumnae). In: FEARNHEAD, R. W. and SUGA, S. (eds.), Tooth enamel IV, Elsevier Science Publishers, Amsterdam, pp. 462–466.
SCHULTZE, H.-P. 1969. Die Faltenzähne der Rhipidistiden Crossopterygier, der Tetrapoden und der Actinopterygi-er-Gattung Lepisosteus nebst einer Beschreibung der Zahnstruktur von Onychodus (Struniiformer Crossoptery-gier). Palaeontologica Italica, New Series, 35(65): 63–137.
SCHULTZE, H.-P. 1970. Folded teeth and the monophyletic origin of Tetrapods. American Museum Novitates, 2408: 1–10.
SCHULTZE, H.-P. 2018. Hard tissues in fish evolution: history and current issues. Cybium, 42(1): 29–39.
SHELLIS, R. P. and POOLE, D. F. G. 1978. The structure of the dental hard tissues of the coelacanthid fish Latimeria chalumnae SMITH. Archives of Oral Biology, 23: 1105–1113.
SIRE, J.-Y. and HUYSSEUNE, A. 2003. Formation of dermal skeletal and dental tissues in fish: a comparative and evolutionary approach. Biological Review, 78: 219–249.
SMITH, J. L. B. 1939. A living fish of the Mesozoic type. Nature, 143: 455–456.
SMITH, J. L. B. 1940. A living coelacanth fish from South Africa. Transactions of the Royal Society of South Africa, 28: 1–106, 44 Pls.
SMITH, M. M. 1978. Enamel in the oral teeth of Latimeria chalumnae (Pisces: Actinistia): a scanning electron microscopic study. Journal of Zoology, 185: 355–369.
SMITH, M. M. 1979. Scanning electron microscopy of odontodes in the scales of coelacanth embryo, Latimeria chalumnae SMITH. Archives of Oral Biology, 24: 179–183.
SMITH, M. M., HOBDELL, M. H. and MILLER, W. A. 1972. The structure of the scales of Latimeria chalumnae. Journal of Zoology, London, 167: 501–509.
THOMSON, K. S. 1969. The biology of the lobe-finned fishes.
Biological Reviews, 44: 91–154. UYENO, T. and YABUMOTO, Y. 2007. Origin of extant
coelacanths. The Coelacanth, Fathom the Mystery 2007. Aquamarine Fukushima, pp. 24–26.
ZYLBERBERG, L. and MEUNIER, F. J. 2008. New data on the structure and the chondrocyte populations of the haemal cartilage of abdominal vertebrae in the adult carp Cyprinus carpio (Teleostei, Ostariophysii, Cyprinidae). Cybium, 32: 225–239.
ZYLBERBERG, L., GÉRAUDIE, J., MEUNIER, F. J., & SIRE J. Y., 1992. Biomineralization in the integumental skeleton of the living lower Vertebrates. In: HALL, B. K. (ed), Bone, Volume 4, CRC Press, Boca Raton, pp. 171–224.
The skeleton and the mineralized tissues of the living coelacanthsFrançois J. MEUNIER, Camila CUPELLO and Gaël CLÉMENT
Fig. 7. Latimeria menadoensis and L. chalumnae. Ground sections of scales. A, L. menadoensis. Longitudinal section; X-ray. The superficial bony layer (bo) and the odontodes (arrow-heads) are mineralized whereas the basal plate (BP) totally lacks mineralization (asterisks). Vascular canals are present within the superficial bony layer. The arrow on the upper right indicates the direction of the head. B–C, L. chalumnae. Transversal sections; respectively polarized light and X-ray. B, The basal plate is made of stratified layers of collagenous fibres (arrow). The arrow-heads point to the superficial odontodes. C, The superficial bony layer and the odontodes (arrow-heads) are mineralized whereas the basal plate (BP) totally lacks mineralization (asterisks). D, L. chalumnae. Longitudinal section; X-ray. The superficial bony layer (bo) and the odontodes (arrow-head) are mineralized whereas the basal plate (BP) totally lacks mineralization (asterisks). (af = anterior field; pf = posterior field). E, L. menadoensis. Broken frozen scale showing the different fibres directions in the successive layers of the basal plate. The rotation of the fibres directions from one layer to another characterizes a plywood-like organization. The arrow points to the crests of the superficial layer. (A, B, C: scale = 1 mm; D: scale = 2 mm; E: scale = 200 µm). (B, C, D after CASTANET et al., 1975; A, E after MEUNIER et al., 2008).
Fig. 8. Latimeria chalumnae. Cartilage histology (after FRANCILLON et al., 1975). A, MECKEL’s cartilage (thin histological section; Hemalun-PIC staining). An erosive vascular cavity (black asterisk) is separated from the medullary cavity (mc) by a layer of enchondral bone (eb). Elongate chondrocytes (arrow-heads) are seen in the cartilage. (scale = 50 µm). B, Scapulocoracoid (ground section 3 mm thick; x-ray). The cartilage (ca) is invaded by resorption canals (white asterisks), their wall is covered by thin bony layers, here clearly visible in white colour. The white arrow-heads point to the erosion front of the cartilage. (scale = 1 mm). C, Scapulocoracoid (thin histological section; Stoelzner staining). In the cartilage (ca), the vascular cavities (vc), resulting from a chondroclastic activity, are bordered by thin bony lamellae here visible in black colour (arrow-heads). (scale = 250 µm).

44 45
type according to the definition of MEUNIER et al. (2013). This organization is less complex than the three other plicidentine types (polyplocodont, eusthenodont and dendrodont), which are described in most of extinct and extant Sarcopterygii (SCHULTZE, 1969, 1970).The bony plates of the lung
The lung of Latimeria is reduced to a short oesophagus diverticulum (CUPELLO et al., 2015) surrounded by scattered small and thin ovoid plates (Fig. 9A, B). These plates cover the surface of the vestigial lung, and are composed of a fibrous matrix with fibrocytes: it is a cellular bony tissue (CUPELLO et al., 2017). Each plate is enclosed in a membranous bag. These plates look fragile in their middle plane since a central weakness zone opened on the microtome knife, creating an artefactual lumen (Fig. 9C) (CUPELLO et al., 2017). The fibres are collagenous and organized in superposed layers. The cells are true osteocytes (Fig. 9E) with a more or less star-shaped outline, and send cytoplasmic extensions in the thickness of the plate (Fig. 9D, E). Rare lining cells (osteoblasts) are
present on the surface of the plates, and conjunctive fibres (SHARPEY’s fibres) penetrate within the plate (Fig. 9C). The extracellular matrix shows various staining intensities with concentric and parallel (Fig. 9D) or globular shaped lines. These lines are considered to be Liesegang lines that characterize an active spheritic mineralizing process. The lung plates of Latimeria are thus true bony plates, and are homologous with the large bony plates known in the abdominal cavity of fossil actinistians (BRITO et al., 2010; CUPELLO et al., 2017). The calcified walls that surrounded the lung of the Cretaceous coelacanth Axelrodichthys are made of large osseous plates of various thickness (BRITO et al., 2010), as those of other fossil coelacanths (CLEMENT, 1999, 2005). The bony tissue of these plates is a vascularized cellular bone with more or less large vascular cavities and some internal remodelling (BRITO et al., 2010). Moreover the mineralization of the lung ossified plates in Axelrodichthys is spheritic as in Latimeria’s plates (CUPELLO et al., 2017). So there is a continuity of the histological structure of the lung plates in the coelacanths during their
evolution, but with an important reduction of the bony plates in Latimeria linked to the vestigial state of its lung (CUPELLO et al., 2015, 2019).
CONCLUSION
This overview of eighty years (1938–2018) of histological work on Latimeria skeletal tissues allows some anatomical and evolutionary considerations. It can be enlightened a drastic reduction of endochondral ossification during the long evolutionary history of coelacanths. The persistence of large volume of cartilage in the endoskeleton at adult stage can be compared to the “little bone and considerable cartilage” that characterize the skeleton of a number of demersal notothenioid telostean fishes (EASTMAN et al., 2014).
A processus of spheritic mineralization has been recently highlighted in various skeletal elements of Latimeria, by the presence of globular dentine in teeth, in odontodes of the tegumentary skeleton (scales, fin rays), in scales at the interface between the external layer and the basal plate, as well as in lung bony plates. The spheritic mineralization (i.e., a radiating arrangement of hydroxyapatite crystals and of the organic matrix) in vertebrate skeletal tissues is considered as a precursor of inotropic mineralization (specific interactions between collagen fibrils and the mineral phase) that possibly represents a derived evolutionary stage of calcification mechanisms (ØRVIG, 1951, 1968; FRANCILLON-VIEILLOT et al., 1990; ZYLBERBERG et al., 1992). The simultaneous presence of both mineralization processes (spheritic and inotropic) in the mineralized tissues of Latimeria and in other osteichthyans such as teleosteans (ZYLBERBERG and MEUNIER, 2008) is however questionable. The mineralized spherules present at the limit between the external layer and the basal plate of Latimeria scales are considered to be the product of an inotropic mineralization (MEUNIER and ZYLBERBERG, 1999). It thus appears that the state of the mineralized spherules observed in the various skeletal tissues of Latimeria, including the lung bony plates, must be studied with adapted ultrastructural techniques in order to test their true origin: spheritic or inotropic mineralization?
ACKNOWLEDGEMENTS
This contribution is linked to an international roundtable on coelacanths evolution held at the Kitakyushu Museum and at the Aquamarine Fukushima in August 2017. We would like to thank the anonymous reviewers for their constructive comments and valuable suggestions. C.C. was partially financed by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brasil (CAPES) – Finance Code 001 (Programa Nacional de Pós Doutorado-PNPD).
INTRODUCTION
The first study of fossil coelacanths was done by AGASSIZ (1833–44) and was followed by a lot of papers with the discovery of new fossils (see reviews of JANVIER, 1996 and FOREY, 1998). Fossil coelacanths constitute an important monophyletic group composed of more than forty genera, eighty species, and belong to the Sarcopterygii (ANDREWS, 1973; JANVIER, 1996; FOREY, 1998; UYENO and YABUMOTO, 2007; and many others). For a century coelacanths were considered extinct since the end of the Cretaceous time (Fig. 1).
With the discovery of a living coelacanth in 1938 (SMITH, 1939) the scientific community was soon highly interested to compare the incomplete fossil sarcopterygian fishes dataset to the recent anatomical and biological organization of Latimeria chalumnae. Many authors (see THOMSON, 1969; MILLOT and ANTHONY, 1958; MILLOT et al., 1978 FOREY, 1984, 1998; JANVIER, 1996) focused their work on Latimeria as a key-taxon to test or propose various hypotheses on the origin and evolutionary history of the early tetrapods and sarcopterygii in general, and to infer their possible biological characteristics. The fossil sarcopterygian fishes are known by their fossilized mineralized tissues, essentially their bones, scales and teeth. Together with the extant lungfishes, especially Neoceratodus, Latimeria offered a unique access to the skeleton as a whole: with both mineralized and unmineralized skeletal tissues. Here, we aim to present an overview of the last eighty years (1938–2018) of histological studies of Latimeria’s skeleton.
Since the discovery of the Raja Laut (“king of the sea” ), off Sulawesi Island in Indonesia (ERDMANN et al., 1998, 1999), the genus Latimeria comprises two species: L. chalumnae SMITH 1939 and L. menadoensis POUYAUD et al., 1999. However our knowledge of the anatomy and biology of the living coelacanths is so far essentially based on L. chalumnae.
THE LATIMERIA SKELETON
The first anatomical observations on the extant coelacanth were done by SMITH (1940). The anatomy of the Latimeria skeleton has been later described in detail by MILLOT and ANTHONY (1958). The living coelacanth shows a general organization similar to that of the other actinistian fishes (THOMSON, 1969; JANVIER, 1996; FOREY, 1998). The skeleton can be divided in various parts (Fig. 2): skull, axial skeleton, paired and unpaired fins, tegumentary skeleton (scales). The skull, axial skeleton and fins are constituted of endoskeletal elements that are overlain by exoskeletal elements. In the extant Latimeria the endoskeleton of the skull shows an important regression of the bones, especial ly in the neurocranium, which are replaced by cartilaginous tissues, as in its Mesozoic fossil actinistian relatives (ROMER, 1937, 1942; MILLOT and ANTHONY, 1958; BJERRING, 1973; FOREY, 1998). In the same way, the endoskeleton of paired and unpaired fins is essentially constituted of four axial cartilaginous elements (Fig. 2; MILLOT and ANTHONY, 1958), with pre and post-axial
elements and eventually minute superficial endochondral ossification (CASTANET et al., 1975). The axial skeleton is composed of the notochord which is coated by an unmineralized fibrillary sheath and is totally deprived of well developed vertebrae due to the lack of ossified centra, although some discrete neural and haemal arches develop in the anterior part of the notochord (MILLOT and ANTHONY, 1958). The anterior neural and haemal spines are relatively short but they progressively increase in length posteriorly. Importantly neural and haemal spines are composed of perichondral bone surrounding a cartilaginous core (Fig. 2).
The scales belong to the exoskeleton. In both extant coelacanth species they are of elasmoid type, composed of an upper external layer also called “external ornamented layer”, and of a lower thicker layer, called the basal plate, which is stratified and almost totally unmineralized (Fig. 3). The upper external layer is ornamented with radial crests. In the posterior area of the scale these radial reliefs are overlain by numerous denticulations, the odontodes, which can be superposed (Fig. 3). The anterior area of the external layer, which is covered by the anterior scales, shows crests with irregularities that have been interpreted as growth marks and tentatively used for ageing coelacanths (HUREAU and OZOUF, 1977; MILLER, 1979).
The tooth plates of the buccal cavity support series of teeth of various sizes (Fig. 4). They range in three morphotypes: the fangs (7–10 mm in height), middle-sized teeth (3–4 mm in height) and rounded tubercles (MILLOT and ANTHONY, 1958). The teeth of the two first categories are conical and sharp whereas those of the third category have an obtuse tip (Fig. 5). Fangs of Latimeria are inserted into a socket (MILLOT and ANTHONY, 1958; HOBDELL and MILER, 1969; CASTANET et al., 1975) and they have a smooth external surface (MILLOT et al., 1978; MEUNIER et al., 2015).
A particularity of the exoskeleton of Latimeria is the abundance of odontodes at the surface of various skull bones
(MILLOT and ANTHONY, 1958; BERNHAUSER, 1961), on the fin rays (CASTANET et al., 1975) and on the posterior (free) area of scales (Fig. 3) (MILLOT and ANTHONY, 1958; CASTANET et al., 1975; HADIATY and RACHNATIKA, 2003; MEUNIER et al., 2008).
Among the various components of the whole skeleton, and apart the SMITH’s pioneer work (SMITH, 1940), numerous studies have been carried out on the histological organization of scales (ROUX, 1942; SMITH et al., 1972; CASTANET et al., 1975; GIRAUD et al., 1978a; MILLER, 1979; SMITH, 1979; MEUNIER, 1980; MEUNIER and ZYLBERBERG, 1999; HADIATY and RACHMATIKA, 2003; MEUNIER et al., 2008) and teeth (MILLER and HOBDELL, 1968; GRADY, 1970; HOBDELL and MILLER, 1969; CASTANET et al., 1975; SHELLIS and POOLE, 1978;
SMITH, 1978; SASAGAWA et al., 1984; MEUNIER et al., 2015). Contrary to the superficial skeleton, there are clearly less histological studies of the skeletal bony elements (FRANCILLON et al., 1975). Meanwhile, in addition to the usual skeletal elements, specific bony tissues have been described in fossil (BRITO et al., 2010) and extant coelacanths (CUPELLO et al., 2015, 2017), such as the mineralized plates surrounding the lung. These bony plates in actinistians do not belong to the skeleton sensu stricto. They are specific bony specializations in relation to the lung (see below), as is the “rocker bone” in relation to the gas bladder of some ophidiiform teleosts (PARMENTIER et al., 2008). Despite the name, the rocker bone is not true bony tissue (PARMENTIER et al., 2008), contrary to
the bony plates of fossil and extant coelacanths (BRITO et al., 2010; CUPELLO et al., 2017).
ADULT LATIMERIA SKELETAL TISSUES
Bony tissuesCortical areas of the dermal skeleton are constituted of
primary periosteal bony t issue. I ts mineralization is heterogeneous and generally marked by series of growth lines, probably due to alternative physiological cycles linked to seasonal variations (Fig. 6). The cortical primary bone shows vascular canals and numerous SHARPEY’s fibres (Fig. 6B). The centre of these bones is frequently made of a spongiosa with vascular cavities surrounded by secondary bony deposits that result from remodelling processes (Fig. 6). This secondary bone is separated from the primary one by reversal cementing lines that are hypermineralized.
Bone remodelling in adult specimens can be more or less developed according to a bone and within a given bone. Primary bone is destroyed by osteoclasts and the deposition process of the secondary bone results from the activity of osteoblasts that have replaced the osteoclasts. The secondary bone does generally not replaced the whole primary bone areas, which can still be recognizable by the presence of
SHARPEY’s fibres and/or growth marks (Fig. 6).The dermal fin rays that sustain paired and unpaired fins
in Latimeria are true lepidotrichia (CASTANET et al., 1975). They are made of two parallel opposite gutter-shaped hemirays, each of which being a series of hemisegments articulated by a collagenous ligament. Each hemisegment is made of a mineralized collagenous fibrillary matrix with embedded osteocytes. The hemisegments are sometimes fused at the basal part of the rays due to centrifugal deposition of bony laminae around the primary articulation, and eventually to the mineralization of the ligament (see figs. 19, 20 in CASTANET et al., 1975). Odontodes observed on the external convex surface of the rays are similar to those present on the dermal bones of the skull and scales. The distal extremity of the lepidotrichia is overlapped by actinotrichia (GÉRAUDIE and MEUNIER, 1980) that are long tapered rods of elastoidin , a fibrous protein of collagenous nature (FAURÉ-FREMIET, 1936; GARRAULT, 1936). The presence of lepidotrichia with distal actinotrichia in Latimeria, as in the fins of Actinopterygii, is
regarded as a plesiomorphic character for Osteichthyes (GÉRAUDIE and MEUNIER, 1980, 1984).
The upper layer of the Latimeria scale is relatively thin and it is constituted of bony tissue with embedded osteocytes, whereas the basal plate is much thicker and composed of numerous strata made of thick collagenous fibres (Fig. 7). Between two collagenous layers there are star-shaped cells (SMITH et al., 1972, pl. VI; CASTANET et al., 1975, fig. 12), the elasmocytes, which cytoplasmic processes insert between the collagenous fibres. These fibres are set in a very specific network. In each layer, the fibres are parallel to each other and the direction of the fibres change from a layer to another one. This regular organization results in a spatial arrangement termed “twisted plywood” (GIRAUD et al., 1978a,b). The rotation of fibres direction from one layer to the next has a mean angle of 27° (GIRAUD et al., 1978a). This organization is found in each scale of L. chalumnae. In L. menadoensis, the basal plate of the scales is also a twisted plywood but its rotation angle seems to be slightly less regular (MEUNIER et al.,
2008). The basal plate in both species is unmineralized (Fig. 7) excepted at the contact between the superficial layer and the basal plate, where spheritic mineralized granules are seen in the very first layers of the basal plate (MEUNIER, 1980; MEUNIER and ZYLBERBERG, 1999; MEUNIER et al., 2008). There is no pore-canal system so Latimeria’s scales are not cosmoid, contrary to most of palaeozoic sarcopterygian fishes. Due to the presence of an unmineralized stratified basal plate, the scales of Latimeria are defined as elasmoid-like scales. However the plywood-like organization is considered to be homologous with the bony basal plate of cosmoid scales of extinct sarcopterygian fishes (MEUNIER, 1980; SIRE and HUYSSEUNE, 2003; MONDEJAR, 2018; SCHULTZE, 2018).Cartilages
The cartilaginous tissues in Latimeria are characterized by long chondrocytes (Fig. 8) contrary to those of teleostean fishes that are relatively spherical in shape (MEUNIER, 1979; ZYLBERBERG and MEUNIER, 2008). Cartilage tissues can show an endochondral ossification process. This phenomenon has been studied respectively on the urohyal (PEGUETA, 1968), the MECKEL’s cartilage and the proximal piece of the pectoral girdle (FRANCILLON et al., 1975). The endochondral ossification is relatively limited when it occurs and the volume of enchondral bone remains reduced to thin bony layers (Fig. 8). The cartilage is destroyed by chondroclasts in areas where endochondral ossification occurs. Serial chondrocytes, known at the origin of the calcified cartilage, seem to be lacking in the endochondral ossification process in Latimeria (FRANCILLON et al., 1975). This constitutes a true difference with teleostean endochondral ossifications (MEUNIER, 1979; ZYLBERBERG and MEUNIER, 2008). Tooth tissues
The three tooth morphotypes described in Latimeria are fangs (7–10 mm in height), middle-sized teeth (3–4 mm in height) and rounded tubercles (MILLOT and ANTHONY, 1958). These teeth and tubercles are constituted of a cone of orthodentine set around a large pulp cavity and overlain by an external hypermineralized layer considered as true enamel (GRADY, 1970; CASTANET et al., 1975; SHELLIS and POOLE, 1978; SMITH, 1978; SASAGAWA et al., 1984). Various studies have revealed the presence of globular dentine (Fig. 5) at the periphery of the first third of the tooth (MILLER and HOBDELL, 1968; HOBDELL and MILLER, 1969; SHELLIS and POOLE, 1978; SASAGAWA et al., 1984). This typical histological organization characterizes the various teeth of the bucco-pharyngeal cavity as well as the various odontodes of the dermal bony plates, the fin rays and the scales (CASTANET et al., 1975).
In the fang the inner wall of the pulp cavity displays a series of plies (Fig. 5; MEUNIER et al., 2015). These plies start at the base of the tooth and reach at least the first third to the half of the tooth towards its tip (Fig. 5). Such plies are also present but less developed in the small caniniform teeth (MEUNIER et al., 2015). These dentine plies in the pulp cavity characterizes a plicidentine organization of orthodentine type. These primary plies correspond to a simplexodont plicidentine
REFERENCES
AGASSIZ, L. 1833–44. Recherches sur les Poissons fossiles. Imprimerie Petitpierre, Neuchâtel. [5 volumes and atlas.]
ANDREWS, S. M. 1973. Interrelationships of crossopterygians. In: GREENWOOD, P. H., MILES, R. S. and PATTERSON, C. (eds.), Interrelationships of Fishes, Acad. Press, London, pp. 137–177.
BERNHAUSER, A. 1961. Zur Knochen- und Zah, histology von Latimeria chalumnae Smith und einiger Fossilformen. Sber. öst. Akad. Wiss., 170: 119–137.
BJERRING, H. C. 1973. Relationships of coelacanthiforms. In: GREENWOOD P. H., MILES R. S. and PATTERSON C. (eds.), Interrelationships of Fishes, Acad. Press, London, pp. 179–205.
BRITO, P. M., MEUNIER, F. J., CLÉMENT, G. and GEFFARD-KURIYAMA, D. 2010. The histological structure of the calcified lung of the fossil coelacanth Axelrodichthys araripensis (Actinistia: Mawsoniidae). Palaeontology, 53: 1281–1290.
CASANE, D. and LAURENTI, P. 2013. Why coelacanths are not ‘living fossils’. Bioessays, 35: 332–338. DOI 10.1002/bies.201200145.
CASTANET, J., MEUNIER, F., BERGOT, C. and FRANÇOIS, Y. 1975. Données pré l iminaires sur les s t ructures histologiques du squelette de Latimeria chalumnae. I - Dents, écailles, rayons de nageoires. In: Coll. Inter. CNRS, "Problèmes actuels de Paléontologie. Evolution des Vertébrés", Paris, 4–9 Juin 1973, 159–168, 4 Pls.
CLEMENT, G. 1999. The actinistian (Sarcopterygii) Piveteauia madagascarensis LEHMAN from the Lower Triassic of Northeastern Madagascar: a redescription on the basis of new material. Journal of Vertebrate Paleontology, 19: 234–242.
CLEMENT, G. 2005. A new coelacanth (Actinist ia, Sarcopterygii) from the Jurassic of France, and the question of the closest relative fossil to Latimeria. Journal of Vertebrate Paleontology, 25: 481–491.
CUPELLO, C., BRITO, P. M., MEUNIER, F. J., HERBIN, M., JANVIER, P., DUTEL, H. & CLÉMENT, G. 2015. Allometric growth in the extant coelacanth lung during ontogenetic development. Nature Communications, 6: 8222. DOI: 10.1038/ncomms9222.
CUPELLO, C., CLEMÉNT, G., MEUNIER, F. J., HERBIN, M., YABUMOTO, Y. and BRITO, P. M. 2019. The long-time adaptation of coelacanths to moderate deep water: reviewing the evidences. Bulletin of Kitakyushu Museum of Natural History and Human History Series A (Natural History), 17: 29–35.
CUPELLO, C., MEUNIER, F. J., HERBIN, M., JANVIER, P., CLÉMENT, G. and BRITO, P. M. 2017. The homology and funct ion of the lung pla tes in extant and foss i l coelacanths. Scientific Reports , 7 : 9244. DOI: 10.1038/s41598-017-09327-6.
EASTMAN, J. T., WITMER, L. M., RIDGELY, R. C. and KUHN, K.
L. 2014. Divergence in skeleta l mass and bone morphology in antarctic notothenioid fishes. Journal of Morphology, 275: 841–861. DOI: 10.1002/jmor.20258
ERDMANN, M. V., CALDWELL, R. L. and MOOSA, M. K. 1998. Indonesian “king of the sea” discovered. Nature, 395: 335.
ERDMANN, M. V., CALDWELL, R. L., JEWETT, S. L. and TJAKRAWIDJAJA, A. 1999. The second recorded living coelacanth from north Sulawesi. Environmental Biology of Fishes, 34: 445–451.
FAURÉ-FREMIET, M. 1936. La structure des fibres d’éastoidine. Archives d'Anatomie Microscopique, 32: 249–269.
FOREY, P. L.- 1984. The coelacanth as a living fossil. In: ELREDGE N. and STANLEY S.M. (eds.), Living fossils. Springer Verlag, Berlin, pp. 166–169.
FOREY, P. L.- 1998. History of the Coelacanth Fishes. Chapman and Hall, London, 419 pp.
FRANCILLON, H., MEUNIER, F., NGO TUAN PHONG, D. and RICQLÈS, A. (DE). 1975. Données préliminaires sur les structures histologiques du squelette de Latimeria chalumnae. I I- Tissus osseux et cartilages. In: Coll. Inter. CNRS, "Problèmes actuels de Paléontologie. Evolution des Vertébrés", Paris, 4–9 Juin 1973, Centre National de la Recherche Scientifique, pp. 169–174, 2 pls.
FRANCILLON-VIEILLOT H., BUFFRENIL V. DE, CASTANET J., GÉRAUDIE J., MEUNIER FJ., SIRE J-Y., ZYLBERBERG L., a n d R I C Q L È S A . D E 1 9 9 0 . M i c r o s t r u c t u r e a n d mineralization of vertebrate skeletal tissues. In: CARTER, J. G. (ed.), Skeletal Biomineralization: Patterns, Processes and Evolutionary Trends. Vol. I , Van Nostrand, New York, pp. 471–530
GARRAULT, H., 1936. Développement des fibres d’élastoidine (actinotrichia) chez les salmonides. Archives d'Anatomie Microscopique, 130: 105–137.
GÉRAUDIE, J. and MEUNIER, F. J. 1980. Elastoidin actinotrichia in coelacanth fins: a comparative study with teleosts. Tissue and Cell, 12: 637–645.
GÉRAUDIE, J. and MEUNIER, F. J. 1984. Structure and comparative morphology of camptotrichia of lungfish fins. Tissue and Cell, 16: 217–236.
GIRAUD, M. M., CASTANET, J., MEUNIER, F. J. and BOULIGAND, Y. 1978a. The fibrous structure of coelacanth scales: a twisted "plywood". Tissue and Cell, 10: 671–686.
GIRAUD, M. M., CASTANET, J., MEUNIER, F. J. and BOULIGAND, Y. 1978b. Organisation spatiale de l'isopédine des écailles du coelacanthe (Latimeria chalumnae SMITH). Comptes Rendus de l’Académie des Sciences, 287: 487–489.
GRADY, J . E. 1970. Tooth development in Latimeria chalumnae (SMITH). Journal of Morphology, 132: 377–388.
HADIATY, R. K. and RACHMATIKA, I. 2003. Morphological study of the scales of Latimeria menadoensis POUYAUD et al., 1999. Treubia (A Journal on Zoology of the Indo-Australian Archipelago), 33: 1–11.
HOBDELL, M. H. and MILLER, W. A. 1969. Radiographic
anatomy of the teeth and tooth supporting tissues of Latimeria chalumnae. Archives of Oral Biology, 14: 855–858.
HUREAU J.-Cl. and OZOUF, C. 1977. Détermination de l’âge et croissance du cœlacanthe Latimeria chalumnae SMITH, 1939 (Poisson, Crossoptérygien, Coelacanthidé). Cybium, 2: 129–137.
JANVIER, P. 1996. Early Vertebrates. Clarendon Press, Oxford, 393 pp.
MEUNIER, F. J. 1979. Etude histologique et microradiographique du cartilage hémal de la vertèbre de la carpe, Cyprinus carpio L. (Pisces, Teleostei, Cyprinidae). Acta Zoologica, 60: 19–31.
MEUNIER, F. J. 1980. Les relations isopédine-tissu osseux dans le post-temporal et les écailles de la ligne latérale de Latimeria chalumnae (SMITH). Zoologica Scripta, 9: 307–317.
MEUNIER, F. J. and ZYLBERBERG, L. 1999. The structure of the external layer and of the odontodes of scales in Latimeria chalumnae (Sarcopterygii, Actinistia, Coelacanthidae) revisited using scanning and transmission electron microscopy. In: SÉRET, B. and SIRE, J.-Y. (eds.), Proc. 5th Indo-Pacific Fish Conf., Nouméa, 1997, Soc. Fr. Ichtyol., Paris, pp. 109–116.
MEUNIER, F. J., ERDMANN, M. V., FERMON, Y. and CALDWELL, R. L. 2008. Can the comparative study of the morphology and histology of the scales of Latimeria menadoensis and L. chalumnae (Sarcopterygii, Actinistia, Coelacanthidae) bring new insight on the taxonomy and the biogeography of recent coelacanthids? Geological Society, London, Special Publications, 295: 351–360.
MEUNIER, F. J., BRITO, P. M. and LEAL, M.-E. 2013. Quelques données morphologiques et histologiques sur la structure de la plaque dentaire linguale d’Arapaima gigas (Osteoglossidae; Teleostei). Cybium, 37: 263–271
MEUNIER, F. J., MONDEJAR-FERNANDEZ, J., GOUSSARD, F., CLÉMENT, G. and HERBIN, M. 2015. Presence of plicidentine in the oral teeth of Latimeria chalumnae (Sarcopterygii; Actinistia; Coelacanthidae). Journal of Structural Biology, 190: 31–37.
MILLER, W. A. 1979. Observations on the structure of mineralized tissue of the coelacanth, including scales and their associate odontodes. Occasional Papers of the Californian Academy of Sciences, 134: 68–78.
MILLER, W. A. and HOBDELL, M. H. 1968. Preliminary report on the histology of the dental and paradental tissues of Latimeria chalumnae (SMITH) with a note on tooth replacement. Archives of Oral Biology, 13: 1289–1291.
MILLOT, J. and ANTHONY, J. 1958. Anatomie de Latimeria chalumnae. T. 1. Squelette, muscles et formations de soutien. CNRS Edr., Paris. 122 pp., 80 pls.
MILLOT, J., ANTHONY, J. and ROBINEAU, D. 1978. Anatomie de Latimeria chalumnae. T. 3. Appareil digestif, téguments, écailles. CNRS Edr., Paris. pp. 7–28, 135–139.
MONDEJAR-FERNANDEZ, J. 2018. On cosmine: its origins, b io logy and impl ica t ions for sarcopterygian interrelationships. Cybium, 42: 41–65.
ØRVIG T. 1951. Histologic studies of Placoderms and fossil Elasmobranchs. 1: The endoskeleton, with remarks on the hard tissues of lower vertebrates in general. Arkiv för Zoologi, 2: 321–454, 8 pl.
ØRVIG T. 1968. The dermal skeleton; general considerations. In: ØRVIG T. (ed.), Current problems of lower vertebrate phylogeny. Proceedings of the fourth Nobel Symposium, Stockholm, 1967, J. Wiley, New-York. pp. 373–397.
PARMENTIER, E., COMPÈRE, P., CASADEWALL, M., FONTENELLE, N., CLOOTS, R. and HENRIST, C. 2008. The rocker bone: a new kind of mineralized tissue? Cell and Tissue Research, 334: 67–79.
PEGUETA, V. P. 1968. Enchondral ossification in Latimeria chalumnae SMITH. Dopovidi Akademii Nauk Ukrainia SSR., 7: 653–656 (in Russian; English abstr.).
POUYAUD, L., WIRJOATMODIO, S., RACHMATIKA, L., TJAKRAWIDJAJA, A., HADIATY, R. K. and HADIE, W. 1999. Une nouvelle espèce de coelacanthe, preuves génétiques et morphologiques. Comptes Rendus de l’Académie des Sciences, 322: 261–267.
ROMER, A.S. 1937. The braincase of the carboniferous crossopterygian Megalichthys nitidus. Bulletin of the Museum of Comparative Zoology, 824: 1–73.
ROMER, A.S. 1942. Cartilage as an embryonic adaptation. American Naturalist, 76: 394–404.
ROUX, G. H. 1942. The microscopic anatomy of the Latimeria scale. South African Journal of Medical Sciences, 7: 1–18.
SASAGAWA, I., ISHIYAMA, M. and KODERA, H. 1984. Fine structure of the pharyngeal teeth in the coelacanth fish (Latimeria chalumnae). In: FEARNHEAD, R. W. and SUGA, S. (eds.), Tooth enamel IV, Elsevier Science Publishers, Amsterdam, pp. 462–466.
SCHULTZE, H.-P. 1969. Die Faltenzähne der Rhipidistiden Crossopterygier, der Tetrapoden und der Actinopterygi-er-Gattung Lepisosteus nebst einer Beschreibung der Zahnstruktur von Onychodus (Struniiformer Crossoptery-gier). Palaeontologica Italica, New Series, 35(65): 63–137.
SCHULTZE, H.-P. 1970. Folded teeth and the monophyletic origin of Tetrapods. American Museum Novitates, 2408: 1–10.
SCHULTZE, H.-P. 2018. Hard tissues in fish evolution: history and current issues. Cybium, 42(1): 29–39.
SHELLIS, R. P. and POOLE, D. F. G. 1978. The structure of the dental hard tissues of the coelacanthid fish Latimeria chalumnae SMITH. Archives of Oral Biology, 23: 1105–1113.
SIRE, J.-Y. and HUYSSEUNE, A. 2003. Formation of dermal skeletal and dental tissues in fish: a comparative and evolutionary approach. Biological Review, 78: 219–249.
SMITH, J. L. B. 1939. A living fish of the Mesozoic type. Nature, 143: 455–456.
SMITH, J. L. B. 1940. A living coelacanth fish from South Africa. Transactions of the Royal Society of South Africa, 28: 1–106, 44 Pls.
SMITH, M. M. 1978. Enamel in the oral teeth of Latimeria chalumnae (Pisces: Actinistia): a scanning electron microscopic study. Journal of Zoology, 185: 355–369.
SMITH, M. M. 1979. Scanning electron microscopy of odontodes in the scales of coelacanth embryo, Latimeria chalumnae SMITH. Archives of Oral Biology, 24: 179–183.
SMITH, M. M., HOBDELL, M. H. and MILLER, W. A. 1972. The structure of the scales of Latimeria chalumnae. Journal of Zoology, London, 167: 501–509.
THOMSON, K. S. 1969. The biology of the lobe-finned fishes.
Biological Reviews, 44: 91–154. UYENO, T. and YABUMOTO, Y. 2007. Origin of extant
coelacanths. The Coelacanth, Fathom the Mystery 2007. Aquamarine Fukushima, pp. 24–26.
ZYLBERBERG, L. and MEUNIER, F. J. 2008. New data on the structure and the chondrocyte populations of the haemal cartilage of abdominal vertebrae in the adult carp Cyprinus carpio (Teleostei, Ostariophysii, Cyprinidae). Cybium, 32: 225–239.
ZYLBERBERG, L., GÉRAUDIE, J., MEUNIER, F. J., & SIRE J. Y., 1992. Biomineralization in the integumental skeleton of the living lower Vertebrates. In: HALL, B. K. (ed), Bone, Volume 4, CRC Press, Boca Raton, pp. 171–224.
The skeleton and the mineralized tissues of the living coelacanthsFrançois J. MEUNIER, Camila CUPELLO and Gaël CLÉMENT
Fig. 9. Latimeria chalumnae. Lung plates. A, Isolated lung plate of an adult specimen (CCC24). (scale = 500 µm). B, Cross section (azocarmin staining) in the anterior part of the vestigial lung showing its lumen (lu), two thin lung diverticles (arrows) and four surrounding plates (1–4) (oe = ventral wall of the oesophagous). (scale = 1 mm). C, Detail of the lung plate n°3 of B, showing the central weakness zone (*) and SHARPEY’s fibres (s) that enter the plate. (scale = 50 µm). D, Detail of a section (hematoxylin-Eosin staining) in a lung plate showing an osteocyte lacuna (*) and the Liesegang lines (arrow-heads) that characterize a spheritic mineralization. (scale = 20 µm). E, Detail (hematoxylin-Eosin staining) of two osteocyte lacunae (*) with their osteocytic processes (arrow-heads) that penetrate the extracellular collagenous matrix. (scale = 20 µm). (B after MILLOT and ANTHONY, 1978; C, D after CUPELLO et al., 2017).
Fig. 10. Latimeria chalumnae. Various odontodes. Ground sections in A) the posterior field (= free ornamented area) of a scale and B, C) a dermal bone. A, Odontode lying on the external layer. Spheritic mineralized granules are seen (arrow-heads) at the frontier of the external layer and the first layers of the basal plate. B, Odontode lying at the surface of a dermal bone. C, Dermal bone overlaid by the dermis (white asterisks). Two odontodes are visible: one at the surface of the bone with its tip eroded (arrow-heads), and an older one (*) covered by more recent bony layers. (scale for A, B, C = 100 µm). (bp = basal plate; bo = bone; de = dentine; pc = pulp cavity; vc = vascular cavity).

44 45
type according to the definition of MEUNIER et al. (2013). This organization is less complex than the three other plicidentine types (polyplocodont, eusthenodont and dendrodont), which are described in most of extinct and extant Sarcopterygii (SCHULTZE, 1969, 1970).The bony plates of the lung
The lung of Latimeria is reduced to a short oesophagus diverticulum (CUPELLO et al., 2015) surrounded by scattered small and thin ovoid plates (Fig. 9A, B). These plates cover the surface of the vestigial lung, and are composed of a fibrous matrix with fibrocytes: it is a cellular bony tissue (CUPELLO et al., 2017). Each plate is enclosed in a membranous bag. These plates look fragile in their middle plane since a central weakness zone opened on the microtome knife, creating an artefactual lumen (Fig. 9C) (CUPELLO et al., 2017). The fibres are collagenous and organized in superposed layers. The cells are true osteocytes (Fig. 9E) with a more or less star-shaped outline, and send cytoplasmic extensions in the thickness of the plate (Fig. 9D, E). Rare lining cells (osteoblasts) are
present on the surface of the plates, and conjunctive fibres (SHARPEY’s fibres) penetrate within the plate (Fig. 9C). The extracellular matrix shows various staining intensities with concentric and parallel (Fig. 9D) or globular shaped lines. These lines are considered to be Liesegang lines that characterize an active spheritic mineralizing process. The lung plates of Latimeria are thus true bony plates, and are homologous with the large bony plates known in the abdominal cavity of fossil actinistians (BRITO et al., 2010; CUPELLO et al., 2017). The calcified walls that surrounded the lung of the Cretaceous coelacanth Axelrodichthys are made of large osseous plates of various thickness (BRITO et al., 2010), as those of other fossil coelacanths (CLEMENT, 1999, 2005). The bony tissue of these plates is a vascularized cellular bone with more or less large vascular cavities and some internal remodelling (BRITO et al., 2010). Moreover the mineralization of the lung ossified plates in Axelrodichthys is spheritic as in Latimeria’s plates (CUPELLO et al., 2017). So there is a continuity of the histological structure of the lung plates in the coelacanths during their
evolution, but with an important reduction of the bony plates in Latimeria linked to the vestigial state of its lung (CUPELLO et al., 2015, 2019).
CONCLUSION
This overview of eighty years (1938–2018) of histological work on Latimeria skeletal tissues allows some anatomical and evolutionary considerations. It can be enlightened a drastic reduction of endochondral ossification during the long evolutionary history of coelacanths. The persistence of large volume of cartilage in the endoskeleton at adult stage can be compared to the “little bone and considerable cartilage” that characterize the skeleton of a number of demersal notothenioid telostean fishes (EASTMAN et al., 2014).
A processus of spheritic mineralization has been recently highlighted in various skeletal elements of Latimeria, by the presence of globular dentine in teeth, in odontodes of the tegumentary skeleton (scales, fin rays), in scales at the interface between the external layer and the basal plate, as well as in lung bony plates. The spheritic mineralization (i.e., a radiating arrangement of hydroxyapatite crystals and of the organic matrix) in vertebrate skeletal tissues is considered as a precursor of inotropic mineralization (specific interactions between collagen fibrils and the mineral phase) that possibly represents a derived evolutionary stage of calcification mechanisms (ØRVIG, 1951, 1968; FRANCILLON-VIEILLOT et al., 1990; ZYLBERBERG et al., 1992). The simultaneous presence of both mineralization processes (spheritic and inotropic) in the mineralized tissues of Latimeria and in other osteichthyans such as teleosteans (ZYLBERBERG and MEUNIER, 2008) is however questionable. The mineralized spherules present at the limit between the external layer and the basal plate of Latimeria scales are considered to be the product of an inotropic mineralization (MEUNIER and ZYLBERBERG, 1999). It thus appears that the state of the mineralized spherules observed in the various skeletal tissues of Latimeria, including the lung bony plates, must be studied with adapted ultrastructural techniques in order to test their true origin: spheritic or inotropic mineralization?
ACKNOWLEDGEMENTS
This contribution is linked to an international roundtable on coelacanths evolution held at the Kitakyushu Museum and at the Aquamarine Fukushima in August 2017. We would like to thank the anonymous reviewers for their constructive comments and valuable suggestions. C.C. was partially financed by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brasil (CAPES) – Finance Code 001 (Programa Nacional de Pós Doutorado-PNPD).
INTRODUCTION
The first study of fossil coelacanths was done by AGASSIZ (1833–44) and was followed by a lot of papers with the discovery of new fossils (see reviews of JANVIER, 1996 and FOREY, 1998). Fossil coelacanths constitute an important monophyletic group composed of more than forty genera, eighty species, and belong to the Sarcopterygii (ANDREWS, 1973; JANVIER, 1996; FOREY, 1998; UYENO and YABUMOTO, 2007; and many others). For a century coelacanths were considered extinct since the end of the Cretaceous time (Fig. 1).
With the discovery of a living coelacanth in 1938 (SMITH, 1939) the scientific community was soon highly interested to compare the incomplete fossil sarcopterygian fishes dataset to the recent anatomical and biological organization of Latimeria chalumnae. Many authors (see THOMSON, 1969; MILLOT and ANTHONY, 1958; MILLOT et al., 1978 FOREY, 1984, 1998; JANVIER, 1996) focused their work on Latimeria as a key-taxon to test or propose various hypotheses on the origin and evolutionary history of the early tetrapods and sarcopterygii in general, and to infer their possible biological characteristics. The fossil sarcopterygian fishes are known by their fossilized mineralized tissues, essentially their bones, scales and teeth. Together with the extant lungfishes, especially Neoceratodus, Latimeria offered a unique access to the skeleton as a whole: with both mineralized and unmineralized skeletal tissues. Here, we aim to present an overview of the last eighty years (1938–2018) of histological studies of Latimeria’s skeleton.
Since the discovery of the Raja Laut (“king of the sea” ), off Sulawesi Island in Indonesia (ERDMANN et al., 1998, 1999), the genus Latimeria comprises two species: L. chalumnae SMITH 1939 and L. menadoensis POUYAUD et al., 1999. However our knowledge of the anatomy and biology of the living coelacanths is so far essentially based on L. chalumnae.
THE LATIMERIA SKELETON
The first anatomical observations on the extant coelacanth were done by SMITH (1940). The anatomy of the Latimeria skeleton has been later described in detail by MILLOT and ANTHONY (1958). The living coelacanth shows a general organization similar to that of the other actinistian fishes (THOMSON, 1969; JANVIER, 1996; FOREY, 1998). The skeleton can be divided in various parts (Fig. 2): skull, axial skeleton, paired and unpaired fins, tegumentary skeleton (scales). The skull, axial skeleton and fins are constituted of endoskeletal elements that are overlain by exoskeletal elements. In the extant Latimeria the endoskeleton of the skull shows an important regression of the bones, especial ly in the neurocranium, which are replaced by cartilaginous tissues, as in its Mesozoic fossil actinistian relatives (ROMER, 1937, 1942; MILLOT and ANTHONY, 1958; BJERRING, 1973; FOREY, 1998). In the same way, the endoskeleton of paired and unpaired fins is essentially constituted of four axial cartilaginous elements (Fig. 2; MILLOT and ANTHONY, 1958), with pre and post-axial
elements and eventually minute superficial endochondral ossification (CASTANET et al., 1975). The axial skeleton is composed of the notochord which is coated by an unmineralized fibrillary sheath and is totally deprived of well developed vertebrae due to the lack of ossified centra, although some discrete neural and haemal arches develop in the anterior part of the notochord (MILLOT and ANTHONY, 1958). The anterior neural and haemal spines are relatively short but they progressively increase in length posteriorly. Importantly neural and haemal spines are composed of perichondral bone surrounding a cartilaginous core (Fig. 2).
The scales belong to the exoskeleton. In both extant coelacanth species they are of elasmoid type, composed of an upper external layer also called “external ornamented layer”, and of a lower thicker layer, called the basal plate, which is stratified and almost totally unmineralized (Fig. 3). The upper external layer is ornamented with radial crests. In the posterior area of the scale these radial reliefs are overlain by numerous denticulations, the odontodes, which can be superposed (Fig. 3). The anterior area of the external layer, which is covered by the anterior scales, shows crests with irregularities that have been interpreted as growth marks and tentatively used for ageing coelacanths (HUREAU and OZOUF, 1977; MILLER, 1979).
The tooth plates of the buccal cavity support series of teeth of various sizes (Fig. 4). They range in three morphotypes: the fangs (7–10 mm in height), middle-sized teeth (3–4 mm in height) and rounded tubercles (MILLOT and ANTHONY, 1958). The teeth of the two first categories are conical and sharp whereas those of the third category have an obtuse tip (Fig. 5). Fangs of Latimeria are inserted into a socket (MILLOT and ANTHONY, 1958; HOBDELL and MILER, 1969; CASTANET et al., 1975) and they have a smooth external surface (MILLOT et al., 1978; MEUNIER et al., 2015).
A particularity of the exoskeleton of Latimeria is the abundance of odontodes at the surface of various skull bones
(MILLOT and ANTHONY, 1958; BERNHAUSER, 1961), on the fin rays (CASTANET et al., 1975) and on the posterior (free) area of scales (Fig. 3) (MILLOT and ANTHONY, 1958; CASTANET et al., 1975; HADIATY and RACHNATIKA, 2003; MEUNIER et al., 2008).
Among the various components of the whole skeleton, and apart the SMITH’s pioneer work (SMITH, 1940), numerous studies have been carried out on the histological organization of scales (ROUX, 1942; SMITH et al., 1972; CASTANET et al., 1975; GIRAUD et al., 1978a; MILLER, 1979; SMITH, 1979; MEUNIER, 1980; MEUNIER and ZYLBERBERG, 1999; HADIATY and RACHMATIKA, 2003; MEUNIER et al., 2008) and teeth (MILLER and HOBDELL, 1968; GRADY, 1970; HOBDELL and MILLER, 1969; CASTANET et al., 1975; SHELLIS and POOLE, 1978;
SMITH, 1978; SASAGAWA et al., 1984; MEUNIER et al., 2015). Contrary to the superficial skeleton, there are clearly less histological studies of the skeletal bony elements (FRANCILLON et al., 1975). Meanwhile, in addition to the usual skeletal elements, specific bony tissues have been described in fossil (BRITO et al., 2010) and extant coelacanths (CUPELLO et al., 2015, 2017), such as the mineralized plates surrounding the lung. These bony plates in actinistians do not belong to the skeleton sensu stricto. They are specific bony specializations in relation to the lung (see below), as is the “rocker bone” in relation to the gas bladder of some ophidiiform teleosts (PARMENTIER et al., 2008). Despite the name, the rocker bone is not true bony tissue (PARMENTIER et al., 2008), contrary to
the bony plates of fossil and extant coelacanths (BRITO et al., 2010; CUPELLO et al., 2017).
ADULT LATIMERIA SKELETAL TISSUES
Bony tissuesCortical areas of the dermal skeleton are constituted of
primary periosteal bony t issue. I ts mineralization is heterogeneous and generally marked by series of growth lines, probably due to alternative physiological cycles linked to seasonal variations (Fig. 6). The cortical primary bone shows vascular canals and numerous SHARPEY’s fibres (Fig. 6B). The centre of these bones is frequently made of a spongiosa with vascular cavities surrounded by secondary bony deposits that result from remodelling processes (Fig. 6). This secondary bone is separated from the primary one by reversal cementing lines that are hypermineralized.
Bone remodelling in adult specimens can be more or less developed according to a bone and within a given bone. Primary bone is destroyed by osteoclasts and the deposition process of the secondary bone results from the activity of osteoblasts that have replaced the osteoclasts. The secondary bone does generally not replaced the whole primary bone areas, which can still be recognizable by the presence of
SHARPEY’s fibres and/or growth marks (Fig. 6).The dermal fin rays that sustain paired and unpaired fins
in Latimeria are true lepidotrichia (CASTANET et al., 1975). They are made of two parallel opposite gutter-shaped hemirays, each of which being a series of hemisegments articulated by a collagenous ligament. Each hemisegment is made of a mineralized collagenous fibrillary matrix with embedded osteocytes. The hemisegments are sometimes fused at the basal part of the rays due to centrifugal deposition of bony laminae around the primary articulation, and eventually to the mineralization of the ligament (see figs. 19, 20 in CASTANET et al., 1975). Odontodes observed on the external convex surface of the rays are similar to those present on the dermal bones of the skull and scales. The distal extremity of the lepidotrichia is overlapped by actinotrichia (GÉRAUDIE and MEUNIER, 1980) that are long tapered rods of elastoidin , a fibrous protein of collagenous nature (FAURÉ-FREMIET, 1936; GARRAULT, 1936). The presence of lepidotrichia with distal actinotrichia in Latimeria, as in the fins of Actinopterygii, is
regarded as a plesiomorphic character for Osteichthyes (GÉRAUDIE and MEUNIER, 1980, 1984).
The upper layer of the Latimeria scale is relatively thin and it is constituted of bony tissue with embedded osteocytes, whereas the basal plate is much thicker and composed of numerous strata made of thick collagenous fibres (Fig. 7). Between two collagenous layers there are star-shaped cells (SMITH et al., 1972, pl. VI; CASTANET et al., 1975, fig. 12), the elasmocytes, which cytoplasmic processes insert between the collagenous fibres. These fibres are set in a very specific network. In each layer, the fibres are parallel to each other and the direction of the fibres change from a layer to another one. This regular organization results in a spatial arrangement termed “twisted plywood” (GIRAUD et al., 1978a,b). The rotation of fibres direction from one layer to the next has a mean angle of 27° (GIRAUD et al., 1978a). This organization is found in each scale of L. chalumnae. In L. menadoensis, the basal plate of the scales is also a twisted plywood but its rotation angle seems to be slightly less regular (MEUNIER et al.,
2008). The basal plate in both species is unmineralized (Fig. 7) excepted at the contact between the superficial layer and the basal plate, where spheritic mineralized granules are seen in the very first layers of the basal plate (MEUNIER, 1980; MEUNIER and ZYLBERBERG, 1999; MEUNIER et al., 2008). There is no pore-canal system so Latimeria’s scales are not cosmoid, contrary to most of palaeozoic sarcopterygian fishes. Due to the presence of an unmineralized stratified basal plate, the scales of Latimeria are defined as elasmoid-like scales. However the plywood-like organization is considered to be homologous with the bony basal plate of cosmoid scales of extinct sarcopterygian fishes (MEUNIER, 1980; SIRE and HUYSSEUNE, 2003; MONDEJAR, 2018; SCHULTZE, 2018).Cartilages
The cartilaginous tissues in Latimeria are characterized by long chondrocytes (Fig. 8) contrary to those of teleostean fishes that are relatively spherical in shape (MEUNIER, 1979; ZYLBERBERG and MEUNIER, 2008). Cartilage tissues can show an endochondral ossification process. This phenomenon has been studied respectively on the urohyal (PEGUETA, 1968), the MECKEL’s cartilage and the proximal piece of the pectoral girdle (FRANCILLON et al., 1975). The endochondral ossification is relatively limited when it occurs and the volume of enchondral bone remains reduced to thin bony layers (Fig. 8). The cartilage is destroyed by chondroclasts in areas where endochondral ossification occurs. Serial chondrocytes, known at the origin of the calcified cartilage, seem to be lacking in the endochondral ossification process in Latimeria (FRANCILLON et al., 1975). This constitutes a true difference with teleostean endochondral ossifications (MEUNIER, 1979; ZYLBERBERG and MEUNIER, 2008). Tooth tissues
The three tooth morphotypes described in Latimeria are fangs (7–10 mm in height), middle-sized teeth (3–4 mm in height) and rounded tubercles (MILLOT and ANTHONY, 1958). These teeth and tubercles are constituted of a cone of orthodentine set around a large pulp cavity and overlain by an external hypermineralized layer considered as true enamel (GRADY, 1970; CASTANET et al., 1975; SHELLIS and POOLE, 1978; SMITH, 1978; SASAGAWA et al., 1984). Various studies have revealed the presence of globular dentine (Fig. 5) at the periphery of the first third of the tooth (MILLER and HOBDELL, 1968; HOBDELL and MILLER, 1969; SHELLIS and POOLE, 1978; SASAGAWA et al., 1984). This typical histological organization characterizes the various teeth of the bucco-pharyngeal cavity as well as the various odontodes of the dermal bony plates, the fin rays and the scales (CASTANET et al., 1975).
In the fang the inner wall of the pulp cavity displays a series of plies (Fig. 5; MEUNIER et al., 2015). These plies start at the base of the tooth and reach at least the first third to the half of the tooth towards its tip (Fig. 5). Such plies are also present but less developed in the small caniniform teeth (MEUNIER et al., 2015). These dentine plies in the pulp cavity characterizes a plicidentine organization of orthodentine type. These primary plies correspond to a simplexodont plicidentine
REFERENCES
AGASSIZ, L. 1833–44. Recherches sur les Poissons fossiles. Imprimerie Petitpierre, Neuchâtel. [5 volumes and atlas.]
ANDREWS, S. M. 1973. Interrelationships of crossopterygians. In: GREENWOOD, P. H., MILES, R. S. and PATTERSON, C. (eds.), Interrelationships of Fishes, Acad. Press, London, pp. 137–177.
BERNHAUSER, A. 1961. Zur Knochen- und Zah, histology von Latimeria chalumnae Smith und einiger Fossilformen. Sber. öst. Akad. Wiss., 170: 119–137.
BJERRING, H. C. 1973. Relationships of coelacanthiforms. In: GREENWOOD P. H., MILES R. S. and PATTERSON C. (eds.), Interrelationships of Fishes, Acad. Press, London, pp. 179–205.
BRITO, P. M., MEUNIER, F. J., CLÉMENT, G. and GEFFARD-KURIYAMA, D. 2010. The histological structure of the calcified lung of the fossil coelacanth Axelrodichthys araripensis (Actinistia: Mawsoniidae). Palaeontology, 53: 1281–1290.
CASANE, D. and LAURENTI, P. 2013. Why coelacanths are not ‘living fossils’. Bioessays, 35: 332–338. DOI 10.1002/bies.201200145.
CASTANET, J., MEUNIER, F., BERGOT, C. and FRANÇOIS, Y. 1975. Données pré l iminaires sur les s t ructures histologiques du squelette de Latimeria chalumnae. I - Dents, écailles, rayons de nageoires. In: Coll. Inter. CNRS, "Problèmes actuels de Paléontologie. Evolution des Vertébrés", Paris, 4–9 Juin 1973, 159–168, 4 Pls.
CLEMENT, G. 1999. The actinistian (Sarcopterygii) Piveteauia madagascarensis LEHMAN from the Lower Triassic of Northeastern Madagascar: a redescription on the basis of new material. Journal of Vertebrate Paleontology, 19: 234–242.
CLEMENT, G. 2005. A new coelacanth (Actinist ia, Sarcopterygii) from the Jurassic of France, and the question of the closest relative fossil to Latimeria. Journal of Vertebrate Paleontology, 25: 481–491.
CUPELLO, C., BRITO, P. M., MEUNIER, F. J., HERBIN, M., JANVIER, P., DUTEL, H. & CLÉMENT, G. 2015. Allometric growth in the extant coelacanth lung during ontogenetic development. Nature Communications, 6: 8222. DOI: 10.1038/ncomms9222.
CUPELLO, C., CLEMÉNT, G., MEUNIER, F. J., HERBIN, M., YABUMOTO, Y. and BRITO, P. M. 2019. The long-time adaptation of coelacanths to moderate deep water: reviewing the evidences. Bulletin of Kitakyushu Museum of Natural History and Human History Series A (Natural History), 17: 29–35.
CUPELLO, C., MEUNIER, F. J., HERBIN, M., JANVIER, P., CLÉMENT, G. and BRITO, P. M. 2017. The homology and funct ion of the lung pla tes in extant and foss i l coelacanths. Scientific Reports , 7 : 9244. DOI: 10.1038/s41598-017-09327-6.
EASTMAN, J. T., WITMER, L. M., RIDGELY, R. C. and KUHN, K.
L. 2014. Divergence in skeleta l mass and bone morphology in antarctic notothenioid fishes. Journal of Morphology, 275: 841–861. DOI: 10.1002/jmor.20258
ERDMANN, M. V., CALDWELL, R. L. and MOOSA, M. K. 1998. Indonesian “king of the sea” discovered. Nature, 395: 335.
ERDMANN, M. V., CALDWELL, R. L., JEWETT, S. L. and TJAKRAWIDJAJA, A. 1999. The second recorded living coelacanth from north Sulawesi. Environmental Biology of Fishes, 34: 445–451.
FAURÉ-FREMIET, M. 1936. La structure des fibres d’éastoidine. Archives d'Anatomie Microscopique, 32: 249–269.
FOREY, P. L.- 1984. The coelacanth as a living fossil. In: ELREDGE N. and STANLEY S.M. (eds.), Living fossils. Springer Verlag, Berlin, pp. 166–169.
FOREY, P. L.- 1998. History of the Coelacanth Fishes. Chapman and Hall, London, 419 pp.
FRANCILLON, H., MEUNIER, F., NGO TUAN PHONG, D. and RICQLÈS, A. (DE). 1975. Données préliminaires sur les structures histologiques du squelette de Latimeria chalumnae. I I- Tissus osseux et cartilages. In: Coll. Inter. CNRS, "Problèmes actuels de Paléontologie. Evolution des Vertébrés", Paris, 4–9 Juin 1973, Centre National de la Recherche Scientifique, pp. 169–174, 2 pls.
FRANCILLON-VIEILLOT H., BUFFRENIL V. DE, CASTANET J., GÉRAUDIE J., MEUNIER FJ., SIRE J-Y., ZYLBERBERG L., a n d R I C Q L È S A . D E 1 9 9 0 . M i c r o s t r u c t u r e a n d mineralization of vertebrate skeletal tissues. In: CARTER, J. G. (ed.), Skeletal Biomineralization: Patterns, Processes and Evolutionary Trends. Vol. I , Van Nostrand, New York, pp. 471–530
GARRAULT, H., 1936. Développement des fibres d’élastoidine (actinotrichia) chez les salmonides. Archives d'Anatomie Microscopique, 130: 105–137.
GÉRAUDIE, J. and MEUNIER, F. J. 1980. Elastoidin actinotrichia in coelacanth fins: a comparative study with teleosts. Tissue and Cell, 12: 637–645.
GÉRAUDIE, J. and MEUNIER, F. J. 1984. Structure and comparative morphology of camptotrichia of lungfish fins. Tissue and Cell, 16: 217–236.
GIRAUD, M. M., CASTANET, J., MEUNIER, F. J. and BOULIGAND, Y. 1978a. The fibrous structure of coelacanth scales: a twisted "plywood". Tissue and Cell, 10: 671–686.
GIRAUD, M. M., CASTANET, J., MEUNIER, F. J. and BOULIGAND, Y. 1978b. Organisation spatiale de l'isopédine des écailles du coelacanthe (Latimeria chalumnae SMITH). Comptes Rendus de l’Académie des Sciences, 287: 487–489.
GRADY, J . E. 1970. Tooth development in Latimeria chalumnae (SMITH). Journal of Morphology, 132: 377–388.
HADIATY, R. K. and RACHMATIKA, I. 2003. Morphological study of the scales of Latimeria menadoensis POUYAUD et al., 1999. Treubia (A Journal on Zoology of the Indo-Australian Archipelago), 33: 1–11.
HOBDELL, M. H. and MILLER, W. A. 1969. Radiographic
anatomy of the teeth and tooth supporting tissues of Latimeria chalumnae. Archives of Oral Biology, 14: 855–858.
HUREAU J.-Cl. and OZOUF, C. 1977. Détermination de l’âge et croissance du cœlacanthe Latimeria chalumnae SMITH, 1939 (Poisson, Crossoptérygien, Coelacanthidé). Cybium, 2: 129–137.
JANVIER, P. 1996. Early Vertebrates. Clarendon Press, Oxford, 393 pp.
MEUNIER, F. J. 1979. Etude histologique et microradiographique du cartilage hémal de la vertèbre de la carpe, Cyprinus carpio L. (Pisces, Teleostei, Cyprinidae). Acta Zoologica, 60: 19–31.
MEUNIER, F. J. 1980. Les relations isopédine-tissu osseux dans le post-temporal et les écailles de la ligne latérale de Latimeria chalumnae (SMITH). Zoologica Scripta, 9: 307–317.
MEUNIER, F. J. and ZYLBERBERG, L. 1999. The structure of the external layer and of the odontodes of scales in Latimeria chalumnae (Sarcopterygii, Actinistia, Coelacanthidae) revisited using scanning and transmission electron microscopy. In: SÉRET, B. and SIRE, J.-Y. (eds.), Proc. 5th Indo-Pacific Fish Conf., Nouméa, 1997, Soc. Fr. Ichtyol., Paris, pp. 109–116.
MEUNIER, F. J., ERDMANN, M. V., FERMON, Y. and CALDWELL, R. L. 2008. Can the comparative study of the morphology and histology of the scales of Latimeria menadoensis and L. chalumnae (Sarcopterygii, Actinistia, Coelacanthidae) bring new insight on the taxonomy and the biogeography of recent coelacanthids? Geological Society, London, Special Publications, 295: 351–360.
MEUNIER, F. J., BRITO, P. M. and LEAL, M.-E. 2013. Quelques données morphologiques et histologiques sur la structure de la plaque dentaire linguale d’Arapaima gigas (Osteoglossidae; Teleostei). Cybium, 37: 263–271
MEUNIER, F. J., MONDEJAR-FERNANDEZ, J., GOUSSARD, F., CLÉMENT, G. and HERBIN, M. 2015. Presence of plicidentine in the oral teeth of Latimeria chalumnae (Sarcopterygii; Actinistia; Coelacanthidae). Journal of Structural Biology, 190: 31–37.
MILLER, W. A. 1979. Observations on the structure of mineralized tissue of the coelacanth, including scales and their associate odontodes. Occasional Papers of the Californian Academy of Sciences, 134: 68–78.
MILLER, W. A. and HOBDELL, M. H. 1968. Preliminary report on the histology of the dental and paradental tissues of Latimeria chalumnae (SMITH) with a note on tooth replacement. Archives of Oral Biology, 13: 1289–1291.
MILLOT, J. and ANTHONY, J. 1958. Anatomie de Latimeria chalumnae. T. 1. Squelette, muscles et formations de soutien. CNRS Edr., Paris. 122 pp., 80 pls.
MILLOT, J., ANTHONY, J. and ROBINEAU, D. 1978. Anatomie de Latimeria chalumnae. T. 3. Appareil digestif, téguments, écailles. CNRS Edr., Paris. pp. 7–28, 135–139.
MONDEJAR-FERNANDEZ, J. 2018. On cosmine: its origins, b io logy and impl ica t ions for sarcopterygian interrelationships. Cybium, 42: 41–65.
ØRVIG T. 1951. Histologic studies of Placoderms and fossil Elasmobranchs. 1: The endoskeleton, with remarks on the hard tissues of lower vertebrates in general. Arkiv för Zoologi, 2: 321–454, 8 pl.
ØRVIG T. 1968. The dermal skeleton; general considerations. In: ØRVIG T. (ed.), Current problems of lower vertebrate phylogeny. Proceedings of the fourth Nobel Symposium, Stockholm, 1967, J. Wiley, New-York. pp. 373–397.
PARMENTIER, E., COMPÈRE, P., CASADEWALL, M., FONTENELLE, N., CLOOTS, R. and HENRIST, C. 2008. The rocker bone: a new kind of mineralized tissue? Cell and Tissue Research, 334: 67–79.
PEGUETA, V. P. 1968. Enchondral ossification in Latimeria chalumnae SMITH. Dopovidi Akademii Nauk Ukrainia SSR., 7: 653–656 (in Russian; English abstr.).
POUYAUD, L., WIRJOATMODIO, S., RACHMATIKA, L., TJAKRAWIDJAJA, A., HADIATY, R. K. and HADIE, W. 1999. Une nouvelle espèce de coelacanthe, preuves génétiques et morphologiques. Comptes Rendus de l’Académie des Sciences, 322: 261–267.
ROMER, A.S. 1937. The braincase of the carboniferous crossopterygian Megalichthys nitidus. Bulletin of the Museum of Comparative Zoology, 824: 1–73.
ROMER, A.S. 1942. Cartilage as an embryonic adaptation. American Naturalist, 76: 394–404.
ROUX, G. H. 1942. The microscopic anatomy of the Latimeria scale. South African Journal of Medical Sciences, 7: 1–18.
SASAGAWA, I., ISHIYAMA, M. and KODERA, H. 1984. Fine structure of the pharyngeal teeth in the coelacanth fish (Latimeria chalumnae). In: FEARNHEAD, R. W. and SUGA, S. (eds.), Tooth enamel IV, Elsevier Science Publishers, Amsterdam, pp. 462–466.
SCHULTZE, H.-P. 1969. Die Faltenzähne der Rhipidistiden Crossopterygier, der Tetrapoden und der Actinopterygi-er-Gattung Lepisosteus nebst einer Beschreibung der Zahnstruktur von Onychodus (Struniiformer Crossoptery-gier). Palaeontologica Italica, New Series, 35(65): 63–137.
SCHULTZE, H.-P. 1970. Folded teeth and the monophyletic origin of Tetrapods. American Museum Novitates, 2408: 1–10.
SCHULTZE, H.-P. 2018. Hard tissues in fish evolution: history and current issues. Cybium, 42(1): 29–39.
SHELLIS, R. P. and POOLE, D. F. G. 1978. The structure of the dental hard tissues of the coelacanthid fish Latimeria chalumnae SMITH. Archives of Oral Biology, 23: 1105–1113.
SIRE, J.-Y. and HUYSSEUNE, A. 2003. Formation of dermal skeletal and dental tissues in fish: a comparative and evolutionary approach. Biological Review, 78: 219–249.
SMITH, J. L. B. 1939. A living fish of the Mesozoic type. Nature, 143: 455–456.
SMITH, J. L. B. 1940. A living coelacanth fish from South Africa. Transactions of the Royal Society of South Africa, 28: 1–106, 44 Pls.
SMITH, M. M. 1978. Enamel in the oral teeth of Latimeria chalumnae (Pisces: Actinistia): a scanning electron microscopic study. Journal of Zoology, 185: 355–369.
SMITH, M. M. 1979. Scanning electron microscopy of odontodes in the scales of coelacanth embryo, Latimeria chalumnae SMITH. Archives of Oral Biology, 24: 179–183.
SMITH, M. M., HOBDELL, M. H. and MILLER, W. A. 1972. The structure of the scales of Latimeria chalumnae. Journal of Zoology, London, 167: 501–509.
THOMSON, K. S. 1969. The biology of the lobe-finned fishes.
Biological Reviews, 44: 91–154. UYENO, T. and YABUMOTO, Y. 2007. Origin of extant
coelacanths. The Coelacanth, Fathom the Mystery 2007. Aquamarine Fukushima, pp. 24–26.
ZYLBERBERG, L. and MEUNIER, F. J. 2008. New data on the structure and the chondrocyte populations of the haemal cartilage of abdominal vertebrae in the adult carp Cyprinus carpio (Teleostei, Ostariophysii, Cyprinidae). Cybium, 32: 225–239.
ZYLBERBERG, L., GÉRAUDIE, J., MEUNIER, F. J., & SIRE J. Y., 1992. Biomineralization in the integumental skeleton of the living lower Vertebrates. In: HALL, B. K. (ed), Bone, Volume 4, CRC Press, Boca Raton, pp. 171–224.
The skeleton and the mineralized tissues of the living coelacanthsFrançois J. MEUNIER, Camila CUPELLO and Gaël CLÉMENT
Fig. 9. Latimeria chalumnae. Lung plates. A, Isolated lung plate of an adult specimen (CCC24). (scale = 500 µm). B, Cross section (azocarmin staining) in the anterior part of the vestigial lung showing its lumen (lu), two thin lung diverticles (arrows) and four surrounding plates (1–4) (oe = ventral wall of the oesophagous). (scale = 1 mm). C, Detail of the lung plate n°3 of B, showing the central weakness zone (*) and SHARPEY’s fibres (s) that enter the plate. (scale = 50 µm). D, Detail of a section (hematoxylin-Eosin staining) in a lung plate showing an osteocyte lacuna (*) and the Liesegang lines (arrow-heads) that characterize a spheritic mineralization. (scale = 20 µm). E, Detail (hematoxylin-Eosin staining) of two osteocyte lacunae (*) with their osteocytic processes (arrow-heads) that penetrate the extracellular collagenous matrix. (scale = 20 µm). (B after MILLOT and ANTHONY, 1978; C, D after CUPELLO et al., 2017).
Fig. 10. Latimeria chalumnae. Various odontodes. Ground sections in A) the posterior field (= free ornamented area) of a scale and B, C) a dermal bone. A, Odontode lying on the external layer. Spheritic mineralized granules are seen (arrow-heads) at the frontier of the external layer and the first layers of the basal plate. B, Odontode lying at the surface of a dermal bone. C, Dermal bone overlaid by the dermis (white asterisks). Two odontodes are visible: one at the surface of the bone with its tip eroded (arrow-heads), and an older one (*) covered by more recent bony layers. (scale for A, B, C = 100 µm). (bp = basal plate; bo = bone; de = dentine; pc = pulp cavity; vc = vascular cavity).

type according to the definition of MEUNIER et al. (2013). This organization is less complex than the three other plicidentine types (polyplocodont, eusthenodont and dendrodont), which are described in most of extinct and extant Sarcopterygii (SCHULTZE, 1969, 1970).The bony plates of the lung
The lung of Latimeria is reduced to a short oesophagus diverticulum (CUPELLO et al., 2015) surrounded by scattered small and thin ovoid plates (Fig. 9A, B). These plates cover the surface of the vestigial lung, and are composed of a fibrous matrix with fibrocytes: it is a cellular bony tissue (CUPELLO et al., 2017). Each plate is enclosed in a membranous bag. These plates look fragile in their middle plane since a central weakness zone opened on the microtome knife, creating an artefactual lumen (Fig. 9C) (CUPELLO et al., 2017). The fibres are collagenous and organized in superposed layers. The cells are true osteocytes (Fig. 9E) with a more or less star-shaped outline, and send cytoplasmic extensions in the thickness of the plate (Fig. 9D, E). Rare lining cells (osteoblasts) are
present on the surface of the plates, and conjunctive fibres (SHARPEY’s fibres) penetrate within the plate (Fig. 9C). The extracellular matrix shows various staining intensities with concentric and parallel (Fig. 9D) or globular shaped lines. These lines are considered to be Liesegang lines that characterize an active spheritic mineralizing process. The lung plates of Latimeria are thus true bony plates, and are homologous with the large bony plates known in the abdominal cavity of fossil actinistians (BRITO et al., 2010; CUPELLO et al., 2017). The calcified walls that surrounded the lung of the Cretaceous coelacanth Axelrodichthys are made of large osseous plates of various thickness (BRITO et al., 2010), as those of other fossil coelacanths (CLEMENT, 1999, 2005). The bony tissue of these plates is a vascularized cellular bone with more or less large vascular cavities and some internal remodelling (BRITO et al., 2010). Moreover the mineralization of the lung ossified plates in Axelrodichthys is spheritic as in Latimeria’s plates (CUPELLO et al., 2017). So there is a continuity of the histological structure of the lung plates in the coelacanths during their
evolution, but with an important reduction of the bony plates in Latimeria linked to the vestigial state of its lung (CUPELLO et al., 2015, 2019).
CONCLUSION
This overview of eighty years (1938–2018) of histological work on Latimeria skeletal tissues allows some anatomical and evolutionary considerations. It can be enlightened a drastic reduction of endochondral ossification during the long evolutionary history of coelacanths. The persistence of large volume of cartilage in the endoskeleton at adult stage can be compared to the “little bone and considerable cartilage” that characterize the skeleton of a number of demersal notothenioid telostean fishes (EASTMAN et al., 2014).
A processus of spheritic mineralization has been recently highlighted in various skeletal elements of Latimeria, by the presence of globular dentine in teeth, in odontodes of the tegumentary skeleton (scales, fin rays), in scales at the interface between the external layer and the basal plate, as well as in lung bony plates. The spheritic mineralization (i.e., a radiating arrangement of hydroxyapatite crystals and of the organic matrix) in vertebrate skeletal tissues is considered as a precursor of inotropic mineralization (specific interactions between collagen fibrils and the mineral phase) that possibly represents a derived evolutionary stage of calcification mechanisms (ØRVIG, 1951, 1968; FRANCILLON-VIEILLOT et al., 1990; ZYLBERBERG et al., 1992). The simultaneous presence of both mineralization processes (spheritic and inotropic) in the mineralized tissues of Latimeria and in other osteichthyans such as teleosteans (ZYLBERBERG and MEUNIER, 2008) is however questionable. The mineralized spherules present at the limit between the external layer and the basal plate of Latimeria scales are considered to be the product of an inotropic mineralization (MEUNIER and ZYLBERBERG, 1999). It thus appears that the state of the mineralized spherules observed in the various skeletal tissues of Latimeria, including the lung bony plates, must be studied with adapted ultrastructural techniques in order to test their true origin: spheritic or inotropic mineralization?
ACKNOWLEDGEMENTS
This contribution is linked to an international roundtable on coelacanths evolution held at the Kitakyushu Museum and at the Aquamarine Fukushima in August 2017. We would like to thank the anonymous reviewers for their constructive comments and valuable suggestions. C.C. was partially financed by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brasil (CAPES) – Finance Code 001 (Programa Nacional de Pós Doutorado-PNPD).
INTRODUCTION
The first study of fossil coelacanths was done by AGASSIZ (1833–44) and was followed by a lot of papers with the discovery of new fossils (see reviews of JANVIER, 1996 and FOREY, 1998). Fossil coelacanths constitute an important monophyletic group composed of more than forty genera, eighty species, and belong to the Sarcopterygii (ANDREWS, 1973; JANVIER, 1996; FOREY, 1998; UYENO and YABUMOTO, 2007; and many others). For a century coelacanths were considered extinct since the end of the Cretaceous time (Fig. 1).
With the discovery of a living coelacanth in 1938 (SMITH, 1939) the scientific community was soon highly interested to compare the incomplete fossil sarcopterygian fishes dataset to the recent anatomical and biological organization of Latimeria chalumnae. Many authors (see THOMSON, 1969; MILLOT and ANTHONY, 1958; MILLOT et al., 1978 FOREY, 1984, 1998; JANVIER, 1996) focused their work on Latimeria as a key-taxon to test or propose various hypotheses on the origin and evolutionary history of the early tetrapods and sarcopterygii in general, and to infer their possible biological characteristics. The fossil sarcopterygian fishes are known by their fossilized mineralized tissues, essentially their bones, scales and teeth. Together with the extant lungfishes, especially Neoceratodus, Latimeria offered a unique access to the skeleton as a whole: with both mineralized and unmineralized skeletal tissues. Here, we aim to present an overview of the last eighty years (1938–2018) of histological studies of Latimeria’s skeleton.
Since the discovery of the Raja Laut (“king of the sea” ), off Sulawesi Island in Indonesia (ERDMANN et al., 1998, 1999), the genus Latimeria comprises two species: L. chalumnae SMITH 1939 and L. menadoensis POUYAUD et al., 1999. However our knowledge of the anatomy and biology of the living coelacanths is so far essentially based on L. chalumnae.
THE LATIMERIA SKELETON
The first anatomical observations on the extant coelacanth were done by SMITH (1940). The anatomy of the Latimeria skeleton has been later described in detail by MILLOT and ANTHONY (1958). The living coelacanth shows a general organization similar to that of the other actinistian fishes (THOMSON, 1969; JANVIER, 1996; FOREY, 1998). The skeleton can be divided in various parts (Fig. 2): skull, axial skeleton, paired and unpaired fins, tegumentary skeleton (scales). The skull, axial skeleton and fins are constituted of endoskeletal elements that are overlain by exoskeletal elements. In the extant Latimeria the endoskeleton of the skull shows an important regression of the bones, especial ly in the neurocranium, which are replaced by cartilaginous tissues, as in its Mesozoic fossil actinistian relatives (ROMER, 1937, 1942; MILLOT and ANTHONY, 1958; BJERRING, 1973; FOREY, 1998). In the same way, the endoskeleton of paired and unpaired fins is essentially constituted of four axial cartilaginous elements (Fig. 2; MILLOT and ANTHONY, 1958), with pre and post-axial
elements and eventually minute superficial endochondral ossification (CASTANET et al., 1975). The axial skeleton is composed of the notochord which is coated by an unmineralized fibrillary sheath and is totally deprived of well developed vertebrae due to the lack of ossified centra, although some discrete neural and haemal arches develop in the anterior part of the notochord (MILLOT and ANTHONY, 1958). The anterior neural and haemal spines are relatively short but they progressively increase in length posteriorly. Importantly neural and haemal spines are composed of perichondral bone surrounding a cartilaginous core (Fig. 2).
The scales belong to the exoskeleton. In both extant coelacanth species they are of elasmoid type, composed of an upper external layer also called “external ornamented layer”, and of a lower thicker layer, called the basal plate, which is stratified and almost totally unmineralized (Fig. 3). The upper external layer is ornamented with radial crests. In the posterior area of the scale these radial reliefs are overlain by numerous denticulations, the odontodes, which can be superposed (Fig. 3). The anterior area of the external layer, which is covered by the anterior scales, shows crests with irregularities that have been interpreted as growth marks and tentatively used for ageing coelacanths (HUREAU and OZOUF, 1977; MILLER, 1979).
The tooth plates of the buccal cavity support series of teeth of various sizes (Fig. 4). They range in three morphotypes: the fangs (7–10 mm in height), middle-sized teeth (3–4 mm in height) and rounded tubercles (MILLOT and ANTHONY, 1958). The teeth of the two first categories are conical and sharp whereas those of the third category have an obtuse tip (Fig. 5). Fangs of Latimeria are inserted into a socket (MILLOT and ANTHONY, 1958; HOBDELL and MILER, 1969; CASTANET et al., 1975) and they have a smooth external surface (MILLOT et al., 1978; MEUNIER et al., 2015).
A particularity of the exoskeleton of Latimeria is the abundance of odontodes at the surface of various skull bones
(MILLOT and ANTHONY, 1958; BERNHAUSER, 1961), on the fin rays (CASTANET et al., 1975) and on the posterior (free) area of scales (Fig. 3) (MILLOT and ANTHONY, 1958; CASTANET et al., 1975; HADIATY and RACHNATIKA, 2003; MEUNIER et al., 2008).
Among the various components of the whole skeleton, and apart the SMITH’s pioneer work (SMITH, 1940), numerous studies have been carried out on the histological organization of scales (ROUX, 1942; SMITH et al., 1972; CASTANET et al., 1975; GIRAUD et al., 1978a; MILLER, 1979; SMITH, 1979; MEUNIER, 1980; MEUNIER and ZYLBERBERG, 1999; HADIATY and RACHMATIKA, 2003; MEUNIER et al., 2008) and teeth (MILLER and HOBDELL, 1968; GRADY, 1970; HOBDELL and MILLER, 1969; CASTANET et al., 1975; SHELLIS and POOLE, 1978;
SMITH, 1978; SASAGAWA et al., 1984; MEUNIER et al., 2015). Contrary to the superficial skeleton, there are clearly less histological studies of the skeletal bony elements (FRANCILLON et al., 1975). Meanwhile, in addition to the usual skeletal elements, specific bony tissues have been described in fossil (BRITO et al., 2010) and extant coelacanths (CUPELLO et al., 2015, 2017), such as the mineralized plates surrounding the lung. These bony plates in actinistians do not belong to the skeleton sensu stricto. They are specific bony specializations in relation to the lung (see below), as is the “rocker bone” in relation to the gas bladder of some ophidiiform teleosts (PARMENTIER et al., 2008). Despite the name, the rocker bone is not true bony tissue (PARMENTIER et al., 2008), contrary to
the bony plates of fossil and extant coelacanths (BRITO et al., 2010; CUPELLO et al., 2017).
ADULT LATIMERIA SKELETAL TISSUES
Bony tissuesCortical areas of the dermal skeleton are constituted of
primary periosteal bony t issue. I ts mineralization is heterogeneous and generally marked by series of growth lines, probably due to alternative physiological cycles linked to seasonal variations (Fig. 6). The cortical primary bone shows vascular canals and numerous SHARPEY’s fibres (Fig. 6B). The centre of these bones is frequently made of a spongiosa with vascular cavities surrounded by secondary bony deposits that result from remodelling processes (Fig. 6). This secondary bone is separated from the primary one by reversal cementing lines that are hypermineralized.
Bone remodelling in adult specimens can be more or less developed according to a bone and within a given bone. Primary bone is destroyed by osteoclasts and the deposition process of the secondary bone results from the activity of osteoblasts that have replaced the osteoclasts. The secondary bone does generally not replaced the whole primary bone areas, which can still be recognizable by the presence of
SHARPEY’s fibres and/or growth marks (Fig. 6).The dermal fin rays that sustain paired and unpaired fins
in Latimeria are true lepidotrichia (CASTANET et al., 1975). They are made of two parallel opposite gutter-shaped hemirays, each of which being a series of hemisegments articulated by a collagenous ligament. Each hemisegment is made of a mineralized collagenous fibrillary matrix with embedded osteocytes. The hemisegments are sometimes fused at the basal part of the rays due to centrifugal deposition of bony laminae around the primary articulation, and eventually to the mineralization of the ligament (see figs. 19, 20 in CASTANET et al., 1975). Odontodes observed on the external convex surface of the rays are similar to those present on the dermal bones of the skull and scales. The distal extremity of the lepidotrichia is overlapped by actinotrichia (GÉRAUDIE and MEUNIER, 1980) that are long tapered rods of elastoidin , a fibrous protein of collagenous nature (FAURÉ-FREMIET, 1936; GARRAULT, 1936). The presence of lepidotrichia with distal actinotrichia in Latimeria, as in the fins of Actinopterygii, is
regarded as a plesiomorphic character for Osteichthyes (GÉRAUDIE and MEUNIER, 1980, 1984).
The upper layer of the Latimeria scale is relatively thin and it is constituted of bony tissue with embedded osteocytes, whereas the basal plate is much thicker and composed of numerous strata made of thick collagenous fibres (Fig. 7). Between two collagenous layers there are star-shaped cells (SMITH et al., 1972, pl. VI; CASTANET et al., 1975, fig. 12), the elasmocytes, which cytoplasmic processes insert between the collagenous fibres. These fibres are set in a very specific network. In each layer, the fibres are parallel to each other and the direction of the fibres change from a layer to another one. This regular organization results in a spatial arrangement termed “twisted plywood” (GIRAUD et al., 1978a,b). The rotation of fibres direction from one layer to the next has a mean angle of 27° (GIRAUD et al., 1978a). This organization is found in each scale of L. chalumnae. In L. menadoensis, the basal plate of the scales is also a twisted plywood but its rotation angle seems to be slightly less regular (MEUNIER et al.,
2008). The basal plate in both species is unmineralized (Fig. 7) excepted at the contact between the superficial layer and the basal plate, where spheritic mineralized granules are seen in the very first layers of the basal plate (MEUNIER, 1980; MEUNIER and ZYLBERBERG, 1999; MEUNIER et al., 2008). There is no pore-canal system so Latimeria’s scales are not cosmoid, contrary to most of palaeozoic sarcopterygian fishes. Due to the presence of an unmineralized stratified basal plate, the scales of Latimeria are defined as elasmoid-like scales. However the plywood-like organization is considered to be homologous with the bony basal plate of cosmoid scales of extinct sarcopterygian fishes (MEUNIER, 1980; SIRE and HUYSSEUNE, 2003; MONDEJAR, 2018; SCHULTZE, 2018).Cartilages
The cartilaginous tissues in Latimeria are characterized by long chondrocytes (Fig. 8) contrary to those of teleostean fishes that are relatively spherical in shape (MEUNIER, 1979; ZYLBERBERG and MEUNIER, 2008). Cartilage tissues can show an endochondral ossification process. This phenomenon has been studied respectively on the urohyal (PEGUETA, 1968), the MECKEL’s cartilage and the proximal piece of the pectoral girdle (FRANCILLON et al., 1975). The endochondral ossification is relatively limited when it occurs and the volume of enchondral bone remains reduced to thin bony layers (Fig. 8). The cartilage is destroyed by chondroclasts in areas where endochondral ossification occurs. Serial chondrocytes, known at the origin of the calcified cartilage, seem to be lacking in the endochondral ossification process in Latimeria (FRANCILLON et al., 1975). This constitutes a true difference with teleostean endochondral ossifications (MEUNIER, 1979; ZYLBERBERG and MEUNIER, 2008). Tooth tissues
The three tooth morphotypes described in Latimeria are fangs (7–10 mm in height), middle-sized teeth (3–4 mm in height) and rounded tubercles (MILLOT and ANTHONY, 1958). These teeth and tubercles are constituted of a cone of orthodentine set around a large pulp cavity and overlain by an external hypermineralized layer considered as true enamel (GRADY, 1970; CASTANET et al., 1975; SHELLIS and POOLE, 1978; SMITH, 1978; SASAGAWA et al., 1984). Various studies have revealed the presence of globular dentine (Fig. 5) at the periphery of the first third of the tooth (MILLER and HOBDELL, 1968; HOBDELL and MILLER, 1969; SHELLIS and POOLE, 1978; SASAGAWA et al., 1984). This typical histological organization characterizes the various teeth of the bucco-pharyngeal cavity as well as the various odontodes of the dermal bony plates, the fin rays and the scales (CASTANET et al., 1975).
In the fang the inner wall of the pulp cavity displays a series of plies (Fig. 5; MEUNIER et al., 2015). These plies start at the base of the tooth and reach at least the first third to the half of the tooth towards its tip (Fig. 5). Such plies are also present but less developed in the small caniniform teeth (MEUNIER et al., 2015). These dentine plies in the pulp cavity characterizes a plicidentine organization of orthodentine type. These primary plies correspond to a simplexodont plicidentine
46 47
REFERENCES
AGASSIZ, L. 1833–44. Recherches sur les Poissons fossiles. Imprimerie Petitpierre, Neuchâtel. [5 volumes and atlas.]
ANDREWS, S. M. 1973. Interrelationships of crossopterygians. In: GREENWOOD, P. H., MILES, R. S. and PATTERSON, C. (eds.), Interrelationships of Fishes, Acad. Press, London, pp. 137–177.
BERNHAUSER, A. 1961. Zur Knochen- und Zah, histology von Latimeria chalumnae Smith und einiger Fossilformen. Sber. öst. Akad. Wiss., 170: 119–137.
BJERRING, H. C. 1973. Relationships of coelacanthiforms. In: GREENWOOD P. H., MILES R. S. and PATTERSON C. (eds.), Interrelationships of Fishes, Acad. Press, London, pp. 179–205.
BRITO, P. M., MEUNIER, F. J., CLÉMENT, G. and GEFFARD-KURIYAMA, D. 2010. The histological structure of the calcified lung of the fossil coelacanth Axelrodichthys araripensis (Actinistia: Mawsoniidae). Palaeontology, 53: 1281–1290.
CASANE, D. and LAURENTI, P. 2013. Why coelacanths are not ‘living fossils’. Bioessays, 35: 332–338. DOI 10.1002/bies.201200145.
CASTANET, J., MEUNIER, F., BERGOT, C. and FRANÇOIS, Y. 1975. Données pré l iminaires sur les s t ructures histologiques du squelette de Latimeria chalumnae. I - Dents, écailles, rayons de nageoires. In: Coll. Inter. CNRS, "Problèmes actuels de Paléontologie. Evolution des Vertébrés", Paris, 4–9 Juin 1973, 159–168, 4 Pls.
CLEMENT, G. 1999. The actinistian (Sarcopterygii) Piveteauia madagascarensis LEHMAN from the Lower Triassic of Northeastern Madagascar: a redescription on the basis of new material. Journal of Vertebrate Paleontology, 19: 234–242.
CLEMENT, G. 2005. A new coelacanth (Actinist ia, Sarcopterygii) from the Jurassic of France, and the question of the closest relative fossil to Latimeria. Journal of Vertebrate Paleontology, 25: 481–491.
CUPELLO, C., BRITO, P. M., MEUNIER, F. J., HERBIN, M., JANVIER, P., DUTEL, H. & CLÉMENT, G. 2015. Allometric growth in the extant coelacanth lung during ontogenetic development. Nature Communications, 6: 8222. DOI: 10.1038/ncomms9222.
CUPELLO, C., CLEMÉNT, G., MEUNIER, F. J., HERBIN, M., YABUMOTO, Y. and BRITO, P. M. 2019. The long-time adaptation of coelacanths to moderate deep water: reviewing the evidences. Bulletin of Kitakyushu Museum of Natural History and Human History Series A (Natural History), 17: 29–35.
CUPELLO, C., MEUNIER, F. J., HERBIN, M., JANVIER, P., CLÉMENT, G. and BRITO, P. M. 2017. The homology and funct ion of the lung pla tes in extant and foss i l coelacanths. Scientific Reports , 7 : 9244. DOI: 10.1038/s41598-017-09327-6.
EASTMAN, J. T., WITMER, L. M., RIDGELY, R. C. and KUHN, K.
L. 2014. Divergence in skeleta l mass and bone morphology in antarctic notothenioid fishes. Journal of Morphology, 275: 841–861. DOI: 10.1002/jmor.20258
ERDMANN, M. V., CALDWELL, R. L. and MOOSA, M. K. 1998. Indonesian “king of the sea” discovered. Nature, 395: 335.
ERDMANN, M. V., CALDWELL, R. L., JEWETT, S. L. and TJAKRAWIDJAJA, A. 1999. The second recorded living coelacanth from north Sulawesi. Environmental Biology of Fishes, 34: 445–451.
FAURÉ-FREMIET, M. 1936. La structure des fibres d’éastoidine. Archives d'Anatomie Microscopique, 32: 249–269.
FOREY, P. L.- 1984. The coelacanth as a living fossil. In: ELREDGE N. and STANLEY S.M. (eds.), Living fossils. Springer Verlag, Berlin, pp. 166–169.
FOREY, P. L.- 1998. History of the Coelacanth Fishes. Chapman and Hall, London, 419 pp.
FRANCILLON, H., MEUNIER, F., NGO TUAN PHONG, D. and RICQLÈS, A. (DE). 1975. Données préliminaires sur les structures histologiques du squelette de Latimeria chalumnae. I I- Tissus osseux et cartilages. In: Coll. Inter. CNRS, "Problèmes actuels de Paléontologie. Evolution des Vertébrés", Paris, 4–9 Juin 1973, Centre National de la Recherche Scientifique, pp. 169–174, 2 pls.
FRANCILLON-VIEILLOT H., BUFFRENIL V. DE, CASTANET J., GÉRAUDIE J., MEUNIER FJ., SIRE J-Y., ZYLBERBERG L., a n d R I C Q L È S A . D E 1 9 9 0 . M i c r o s t r u c t u r e a n d mineralization of vertebrate skeletal tissues. In: CARTER, J. G. (ed.), Skeletal Biomineralization: Patterns, Processes and Evolutionary Trends. Vol. I , Van Nostrand, New York, pp. 471–530
GARRAULT, H., 1936. Développement des fibres d’élastoidine (actinotrichia) chez les salmonides. Archives d'Anatomie Microscopique, 130: 105–137.
GÉRAUDIE, J. and MEUNIER, F. J. 1980. Elastoidin actinotrichia in coelacanth fins: a comparative study with teleosts. Tissue and Cell, 12: 637–645.
GÉRAUDIE, J. and MEUNIER, F. J. 1984. Structure and comparative morphology of camptotrichia of lungfish fins. Tissue and Cell, 16: 217–236.
GIRAUD, M. M., CASTANET, J., MEUNIER, F. J. and BOULIGAND, Y. 1978a. The fibrous structure of coelacanth scales: a twisted "plywood". Tissue and Cell, 10: 671–686.
GIRAUD, M. M., CASTANET, J., MEUNIER, F. J. and BOULIGAND, Y. 1978b. Organisation spatiale de l'isopédine des écailles du coelacanthe (Latimeria chalumnae SMITH). Comptes Rendus de l’Académie des Sciences, 287: 487–489.
GRADY, J . E. 1970. Tooth development in Latimeria chalumnae (SMITH). Journal of Morphology, 132: 377–388.
HADIATY, R. K. and RACHMATIKA, I. 2003. Morphological study of the scales of Latimeria menadoensis POUYAUD et al., 1999. Treubia (A Journal on Zoology of the Indo-Australian Archipelago), 33: 1–11.
HOBDELL, M. H. and MILLER, W. A. 1969. Radiographic
anatomy of the teeth and tooth supporting tissues of Latimeria chalumnae. Archives of Oral Biology, 14: 855–858.
HUREAU J.-Cl. and OZOUF, C. 1977. Détermination de l’âge et croissance du cœlacanthe Latimeria chalumnae SMITH, 1939 (Poisson, Crossoptérygien, Coelacanthidé). Cybium, 2: 129–137.
JANVIER, P. 1996. Early Vertebrates. Clarendon Press, Oxford, 393 pp.
MEUNIER, F. J. 1979. Etude histologique et microradiographique du cartilage hémal de la vertèbre de la carpe, Cyprinus carpio L. (Pisces, Teleostei, Cyprinidae). Acta Zoologica, 60: 19–31.
MEUNIER, F. J. 1980. Les relations isopédine-tissu osseux dans le post-temporal et les écailles de la ligne latérale de Latimeria chalumnae (SMITH). Zoologica Scripta, 9: 307–317.
MEUNIER, F. J. and ZYLBERBERG, L. 1999. The structure of the external layer and of the odontodes of scales in Latimeria chalumnae (Sarcopterygii, Actinistia, Coelacanthidae) revisited using scanning and transmission electron microscopy. In: SÉRET, B. and SIRE, J.-Y. (eds.), Proc. 5th Indo-Pacific Fish Conf., Nouméa, 1997, Soc. Fr. Ichtyol., Paris, pp. 109–116.
MEUNIER, F. J., ERDMANN, M. V., FERMON, Y. and CALDWELL, R. L. 2008. Can the comparative study of the morphology and histology of the scales of Latimeria menadoensis and L. chalumnae (Sarcopterygii, Actinistia, Coelacanthidae) bring new insight on the taxonomy and the biogeography of recent coelacanthids? Geological Society, London, Special Publications, 295: 351–360.
MEUNIER, F. J., BRITO, P. M. and LEAL, M.-E. 2013. Quelques données morphologiques et histologiques sur la structure de la plaque dentaire linguale d’Arapaima gigas (Osteoglossidae; Teleostei). Cybium, 37: 263–271
MEUNIER, F. J., MONDEJAR-FERNANDEZ, J., GOUSSARD, F., CLÉMENT, G. and HERBIN, M. 2015. Presence of plicidentine in the oral teeth of Latimeria chalumnae (Sarcopterygii; Actinistia; Coelacanthidae). Journal of Structural Biology, 190: 31–37.
MILLER, W. A. 1979. Observations on the structure of mineralized tissue of the coelacanth, including scales and their associate odontodes. Occasional Papers of the Californian Academy of Sciences, 134: 68–78.
MILLER, W. A. and HOBDELL, M. H. 1968. Preliminary report on the histology of the dental and paradental tissues of Latimeria chalumnae (SMITH) with a note on tooth replacement. Archives of Oral Biology, 13: 1289–1291.
MILLOT, J. and ANTHONY, J. 1958. Anatomie de Latimeria chalumnae. T. 1. Squelette, muscles et formations de soutien. CNRS Edr., Paris. 122 pp., 80 pls.
MILLOT, J., ANTHONY, J. and ROBINEAU, D. 1978. Anatomie de Latimeria chalumnae. T. 3. Appareil digestif, téguments, écailles. CNRS Edr., Paris. pp. 7–28, 135–139.
MONDEJAR-FERNANDEZ, J. 2018. On cosmine: its origins, b io logy and impl ica t ions for sarcopterygian interrelationships. Cybium, 42: 41–65.
ØRVIG T. 1951. Histologic studies of Placoderms and fossil Elasmobranchs. 1: The endoskeleton, with remarks on the hard tissues of lower vertebrates in general. Arkiv för Zoologi, 2: 321–454, 8 pl.
ØRVIG T. 1968. The dermal skeleton; general considerations. In: ØRVIG T. (ed.), Current problems of lower vertebrate phylogeny. Proceedings of the fourth Nobel Symposium, Stockholm, 1967, J. Wiley, New-York. pp. 373–397.
PARMENTIER, E., COMPÈRE, P., CASADEWALL, M., FONTENELLE, N., CLOOTS, R. and HENRIST, C. 2008. The rocker bone: a new kind of mineralized tissue? Cell and Tissue Research, 334: 67–79.
PEGUETA, V. P. 1968. Enchondral ossification in Latimeria chalumnae SMITH. Dopovidi Akademii Nauk Ukrainia SSR., 7: 653–656 (in Russian; English abstr.).
POUYAUD, L., WIRJOATMODIO, S., RACHMATIKA, L., TJAKRAWIDJAJA, A., HADIATY, R. K. and HADIE, W. 1999. Une nouvelle espèce de coelacanthe, preuves génétiques et morphologiques. Comptes Rendus de l’Académie des Sciences, 322: 261–267.
ROMER, A.S. 1937. The braincase of the carboniferous crossopterygian Megalichthys nitidus. Bulletin of the Museum of Comparative Zoology, 824: 1–73.
ROMER, A.S. 1942. Cartilage as an embryonic adaptation. American Naturalist, 76: 394–404.
ROUX, G. H. 1942. The microscopic anatomy of the Latimeria scale. South African Journal of Medical Sciences, 7: 1–18.
SASAGAWA, I., ISHIYAMA, M. and KODERA, H. 1984. Fine structure of the pharyngeal teeth in the coelacanth fish (Latimeria chalumnae). In: FEARNHEAD, R. W. and SUGA, S. (eds.), Tooth enamel IV, Elsevier Science Publishers, Amsterdam, pp. 462–466.
SCHULTZE, H.-P. 1969. Die Faltenzähne der Rhipidistiden Crossopterygier, der Tetrapoden und der Actinopterygi-er-Gattung Lepisosteus nebst einer Beschreibung der Zahnstruktur von Onychodus (Struniiformer Crossoptery-gier). Palaeontologica Italica, New Series, 35(65): 63–137.
SCHULTZE, H.-P. 1970. Folded teeth and the monophyletic origin of Tetrapods. American Museum Novitates, 2408: 1–10.
SCHULTZE, H.-P. 2018. Hard tissues in fish evolution: history and current issues. Cybium, 42(1): 29–39.
SHELLIS, R. P. and POOLE, D. F. G. 1978. The structure of the dental hard tissues of the coelacanthid fish Latimeria chalumnae SMITH. Archives of Oral Biology, 23: 1105–1113.
SIRE, J.-Y. and HUYSSEUNE, A. 2003. Formation of dermal skeletal and dental tissues in fish: a comparative and evolutionary approach. Biological Review, 78: 219–249.
SMITH, J. L. B. 1939. A living fish of the Mesozoic type. Nature, 143: 455–456.
SMITH, J. L. B. 1940. A living coelacanth fish from South Africa. Transactions of the Royal Society of South Africa, 28: 1–106, 44 Pls.
SMITH, M. M. 1978. Enamel in the oral teeth of Latimeria chalumnae (Pisces: Actinistia): a scanning electron microscopic study. Journal of Zoology, 185: 355–369.
SMITH, M. M. 1979. Scanning electron microscopy of odontodes in the scales of coelacanth embryo, Latimeria chalumnae SMITH. Archives of Oral Biology, 24: 179–183.
SMITH, M. M., HOBDELL, M. H. and MILLER, W. A. 1972. The structure of the scales of Latimeria chalumnae. Journal of Zoology, London, 167: 501–509.
THOMSON, K. S. 1969. The biology of the lobe-finned fishes.
Biological Reviews, 44: 91–154. UYENO, T. and YABUMOTO, Y. 2007. Origin of extant
coelacanths. The Coelacanth, Fathom the Mystery 2007. Aquamarine Fukushima, pp. 24–26.
ZYLBERBERG, L. and MEUNIER, F. J. 2008. New data on the structure and the chondrocyte populations of the haemal cartilage of abdominal vertebrae in the adult carp Cyprinus carpio (Teleostei, Ostariophysii, Cyprinidae). Cybium, 32: 225–239.
ZYLBERBERG, L., GÉRAUDIE, J., MEUNIER, F. J., & SIRE J. Y., 1992. Biomineralization in the integumental skeleton of the living lower Vertebrates. In: HALL, B. K. (ed), Bone, Volume 4, CRC Press, Boca Raton, pp. 171–224.
François J. MEUNIER, Camila CUPELLO and Gaël CLÉMENT The skeleton and the mineralized tissues of the living coelacanths

type according to the definition of MEUNIER et al. (2013). This organization is less complex than the three other plicidentine types (polyplocodont, eusthenodont and dendrodont), which are described in most of extinct and extant Sarcopterygii (SCHULTZE, 1969, 1970).The bony plates of the lung
The lung of Latimeria is reduced to a short oesophagus diverticulum (CUPELLO et al., 2015) surrounded by scattered small and thin ovoid plates (Fig. 9A, B). These plates cover the surface of the vestigial lung, and are composed of a fibrous matrix with fibrocytes: it is a cellular bony tissue (CUPELLO et al., 2017). Each plate is enclosed in a membranous bag. These plates look fragile in their middle plane since a central weakness zone opened on the microtome knife, creating an artefactual lumen (Fig. 9C) (CUPELLO et al., 2017). The fibres are collagenous and organized in superposed layers. The cells are true osteocytes (Fig. 9E) with a more or less star-shaped outline, and send cytoplasmic extensions in the thickness of the plate (Fig. 9D, E). Rare lining cells (osteoblasts) are
present on the surface of the plates, and conjunctive fibres (SHARPEY’s fibres) penetrate within the plate (Fig. 9C). The extracellular matrix shows various staining intensities with concentric and parallel (Fig. 9D) or globular shaped lines. These lines are considered to be Liesegang lines that characterize an active spheritic mineralizing process. The lung plates of Latimeria are thus true bony plates, and are homologous with the large bony plates known in the abdominal cavity of fossil actinistians (BRITO et al., 2010; CUPELLO et al., 2017). The calcified walls that surrounded the lung of the Cretaceous coelacanth Axelrodichthys are made of large osseous plates of various thickness (BRITO et al., 2010), as those of other fossil coelacanths (CLEMENT, 1999, 2005). The bony tissue of these plates is a vascularized cellular bone with more or less large vascular cavities and some internal remodelling (BRITO et al., 2010). Moreover the mineralization of the lung ossified plates in Axelrodichthys is spheritic as in Latimeria’s plates (CUPELLO et al., 2017). So there is a continuity of the histological structure of the lung plates in the coelacanths during their
evolution, but with an important reduction of the bony plates in Latimeria linked to the vestigial state of its lung (CUPELLO et al., 2015, 2019).
CONCLUSION
This overview of eighty years (1938–2018) of histological work on Latimeria skeletal tissues allows some anatomical and evolutionary considerations. It can be enlightened a drastic reduction of endochondral ossification during the long evolutionary history of coelacanths. The persistence of large volume of cartilage in the endoskeleton at adult stage can be compared to the “little bone and considerable cartilage” that characterize the skeleton of a number of demersal notothenioid telostean fishes (EASTMAN et al., 2014).
A processus of spheritic mineralization has been recently highlighted in various skeletal elements of Latimeria, by the presence of globular dentine in teeth, in odontodes of the tegumentary skeleton (scales, fin rays), in scales at the interface between the external layer and the basal plate, as well as in lung bony plates. The spheritic mineralization (i.e., a radiating arrangement of hydroxyapatite crystals and of the organic matrix) in vertebrate skeletal tissues is considered as a precursor of inotropic mineralization (specific interactions between collagen fibrils and the mineral phase) that possibly represents a derived evolutionary stage of calcification mechanisms (ØRVIG, 1951, 1968; FRANCILLON-VIEILLOT et al., 1990; ZYLBERBERG et al., 1992). The simultaneous presence of both mineralization processes (spheritic and inotropic) in the mineralized tissues of Latimeria and in other osteichthyans such as teleosteans (ZYLBERBERG and MEUNIER, 2008) is however questionable. The mineralized spherules present at the limit between the external layer and the basal plate of Latimeria scales are considered to be the product of an inotropic mineralization (MEUNIER and ZYLBERBERG, 1999). It thus appears that the state of the mineralized spherules observed in the various skeletal tissues of Latimeria, including the lung bony plates, must be studied with adapted ultrastructural techniques in order to test their true origin: spheritic or inotropic mineralization?
ACKNOWLEDGEMENTS
This contribution is linked to an international roundtable on coelacanths evolution held at the Kitakyushu Museum and at the Aquamarine Fukushima in August 2017. We would like to thank the anonymous reviewers for their constructive comments and valuable suggestions. C.C. was partially financed by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brasil (CAPES) – Finance Code 001 (Programa Nacional de Pós Doutorado-PNPD).
INTRODUCTION
The first study of fossil coelacanths was done by AGASSIZ (1833–44) and was followed by a lot of papers with the discovery of new fossils (see reviews of JANVIER, 1996 and FOREY, 1998). Fossil coelacanths constitute an important monophyletic group composed of more than forty genera, eighty species, and belong to the Sarcopterygii (ANDREWS, 1973; JANVIER, 1996; FOREY, 1998; UYENO and YABUMOTO, 2007; and many others). For a century coelacanths were considered extinct since the end of the Cretaceous time (Fig. 1).
With the discovery of a living coelacanth in 1938 (SMITH, 1939) the scientific community was soon highly interested to compare the incomplete fossil sarcopterygian fishes dataset to the recent anatomical and biological organization of Latimeria chalumnae. Many authors (see THOMSON, 1969; MILLOT and ANTHONY, 1958; MILLOT et al., 1978 FOREY, 1984, 1998; JANVIER, 1996) focused their work on Latimeria as a key-taxon to test or propose various hypotheses on the origin and evolutionary history of the early tetrapods and sarcopterygii in general, and to infer their possible biological characteristics. The fossil sarcopterygian fishes are known by their fossilized mineralized tissues, essentially their bones, scales and teeth. Together with the extant lungfishes, especially Neoceratodus, Latimeria offered a unique access to the skeleton as a whole: with both mineralized and unmineralized skeletal tissues. Here, we aim to present an overview of the last eighty years (1938–2018) of histological studies of Latimeria’s skeleton.
Since the discovery of the Raja Laut (“king of the sea” ), off Sulawesi Island in Indonesia (ERDMANN et al., 1998, 1999), the genus Latimeria comprises two species: L. chalumnae SMITH 1939 and L. menadoensis POUYAUD et al., 1999. However our knowledge of the anatomy and biology of the living coelacanths is so far essentially based on L. chalumnae.
THE LATIMERIA SKELETON
The first anatomical observations on the extant coelacanth were done by SMITH (1940). The anatomy of the Latimeria skeleton has been later described in detail by MILLOT and ANTHONY (1958). The living coelacanth shows a general organization similar to that of the other actinistian fishes (THOMSON, 1969; JANVIER, 1996; FOREY, 1998). The skeleton can be divided in various parts (Fig. 2): skull, axial skeleton, paired and unpaired fins, tegumentary skeleton (scales). The skull, axial skeleton and fins are constituted of endoskeletal elements that are overlain by exoskeletal elements. In the extant Latimeria the endoskeleton of the skull shows an important regression of the bones, especial ly in the neurocranium, which are replaced by cartilaginous tissues, as in its Mesozoic fossil actinistian relatives (ROMER, 1937, 1942; MILLOT and ANTHONY, 1958; BJERRING, 1973; FOREY, 1998). In the same way, the endoskeleton of paired and unpaired fins is essentially constituted of four axial cartilaginous elements (Fig. 2; MILLOT and ANTHONY, 1958), with pre and post-axial
elements and eventually minute superficial endochondral ossification (CASTANET et al., 1975). The axial skeleton is composed of the notochord which is coated by an unmineralized fibrillary sheath and is totally deprived of well developed vertebrae due to the lack of ossified centra, although some discrete neural and haemal arches develop in the anterior part of the notochord (MILLOT and ANTHONY, 1958). The anterior neural and haemal spines are relatively short but they progressively increase in length posteriorly. Importantly neural and haemal spines are composed of perichondral bone surrounding a cartilaginous core (Fig. 2).
The scales belong to the exoskeleton. In both extant coelacanth species they are of elasmoid type, composed of an upper external layer also called “external ornamented layer”, and of a lower thicker layer, called the basal plate, which is stratified and almost totally unmineralized (Fig. 3). The upper external layer is ornamented with radial crests. In the posterior area of the scale these radial reliefs are overlain by numerous denticulations, the odontodes, which can be superposed (Fig. 3). The anterior area of the external layer, which is covered by the anterior scales, shows crests with irregularities that have been interpreted as growth marks and tentatively used for ageing coelacanths (HUREAU and OZOUF, 1977; MILLER, 1979).
The tooth plates of the buccal cavity support series of teeth of various sizes (Fig. 4). They range in three morphotypes: the fangs (7–10 mm in height), middle-sized teeth (3–4 mm in height) and rounded tubercles (MILLOT and ANTHONY, 1958). The teeth of the two first categories are conical and sharp whereas those of the third category have an obtuse tip (Fig. 5). Fangs of Latimeria are inserted into a socket (MILLOT and ANTHONY, 1958; HOBDELL and MILER, 1969; CASTANET et al., 1975) and they have a smooth external surface (MILLOT et al., 1978; MEUNIER et al., 2015).
A particularity of the exoskeleton of Latimeria is the abundance of odontodes at the surface of various skull bones
(MILLOT and ANTHONY, 1958; BERNHAUSER, 1961), on the fin rays (CASTANET et al., 1975) and on the posterior (free) area of scales (Fig. 3) (MILLOT and ANTHONY, 1958; CASTANET et al., 1975; HADIATY and RACHNATIKA, 2003; MEUNIER et al., 2008).
Among the various components of the whole skeleton, and apart the SMITH’s pioneer work (SMITH, 1940), numerous studies have been carried out on the histological organization of scales (ROUX, 1942; SMITH et al., 1972; CASTANET et al., 1975; GIRAUD et al., 1978a; MILLER, 1979; SMITH, 1979; MEUNIER, 1980; MEUNIER and ZYLBERBERG, 1999; HADIATY and RACHMATIKA, 2003; MEUNIER et al., 2008) and teeth (MILLER and HOBDELL, 1968; GRADY, 1970; HOBDELL and MILLER, 1969; CASTANET et al., 1975; SHELLIS and POOLE, 1978;
SMITH, 1978; SASAGAWA et al., 1984; MEUNIER et al., 2015). Contrary to the superficial skeleton, there are clearly less histological studies of the skeletal bony elements (FRANCILLON et al., 1975). Meanwhile, in addition to the usual skeletal elements, specific bony tissues have been described in fossil (BRITO et al., 2010) and extant coelacanths (CUPELLO et al., 2015, 2017), such as the mineralized plates surrounding the lung. These bony plates in actinistians do not belong to the skeleton sensu stricto. They are specific bony specializations in relation to the lung (see below), as is the “rocker bone” in relation to the gas bladder of some ophidiiform teleosts (PARMENTIER et al., 2008). Despite the name, the rocker bone is not true bony tissue (PARMENTIER et al., 2008), contrary to
the bony plates of fossil and extant coelacanths (BRITO et al., 2010; CUPELLO et al., 2017).
ADULT LATIMERIA SKELETAL TISSUES
Bony tissuesCortical areas of the dermal skeleton are constituted of
primary periosteal bony t issue. I ts mineralization is heterogeneous and generally marked by series of growth lines, probably due to alternative physiological cycles linked to seasonal variations (Fig. 6). The cortical primary bone shows vascular canals and numerous SHARPEY’s fibres (Fig. 6B). The centre of these bones is frequently made of a spongiosa with vascular cavities surrounded by secondary bony deposits that result from remodelling processes (Fig. 6). This secondary bone is separated from the primary one by reversal cementing lines that are hypermineralized.
Bone remodelling in adult specimens can be more or less developed according to a bone and within a given bone. Primary bone is destroyed by osteoclasts and the deposition process of the secondary bone results from the activity of osteoblasts that have replaced the osteoclasts. The secondary bone does generally not replaced the whole primary bone areas, which can still be recognizable by the presence of
SHARPEY’s fibres and/or growth marks (Fig. 6).The dermal fin rays that sustain paired and unpaired fins
in Latimeria are true lepidotrichia (CASTANET et al., 1975). They are made of two parallel opposite gutter-shaped hemirays, each of which being a series of hemisegments articulated by a collagenous ligament. Each hemisegment is made of a mineralized collagenous fibrillary matrix with embedded osteocytes. The hemisegments are sometimes fused at the basal part of the rays due to centrifugal deposition of bony laminae around the primary articulation, and eventually to the mineralization of the ligament (see figs. 19, 20 in CASTANET et al., 1975). Odontodes observed on the external convex surface of the rays are similar to those present on the dermal bones of the skull and scales. The distal extremity of the lepidotrichia is overlapped by actinotrichia (GÉRAUDIE and MEUNIER, 1980) that are long tapered rods of elastoidin , a fibrous protein of collagenous nature (FAURÉ-FREMIET, 1936; GARRAULT, 1936). The presence of lepidotrichia with distal actinotrichia in Latimeria, as in the fins of Actinopterygii, is
regarded as a plesiomorphic character for Osteichthyes (GÉRAUDIE and MEUNIER, 1980, 1984).
The upper layer of the Latimeria scale is relatively thin and it is constituted of bony tissue with embedded osteocytes, whereas the basal plate is much thicker and composed of numerous strata made of thick collagenous fibres (Fig. 7). Between two collagenous layers there are star-shaped cells (SMITH et al., 1972, pl. VI; CASTANET et al., 1975, fig. 12), the elasmocytes, which cytoplasmic processes insert between the collagenous fibres. These fibres are set in a very specific network. In each layer, the fibres are parallel to each other and the direction of the fibres change from a layer to another one. This regular organization results in a spatial arrangement termed “twisted plywood” (GIRAUD et al., 1978a,b). The rotation of fibres direction from one layer to the next has a mean angle of 27° (GIRAUD et al., 1978a). This organization is found in each scale of L. chalumnae. In L. menadoensis, the basal plate of the scales is also a twisted plywood but its rotation angle seems to be slightly less regular (MEUNIER et al.,
2008). The basal plate in both species is unmineralized (Fig. 7) excepted at the contact between the superficial layer and the basal plate, where spheritic mineralized granules are seen in the very first layers of the basal plate (MEUNIER, 1980; MEUNIER and ZYLBERBERG, 1999; MEUNIER et al., 2008). There is no pore-canal system so Latimeria’s scales are not cosmoid, contrary to most of palaeozoic sarcopterygian fishes. Due to the presence of an unmineralized stratified basal plate, the scales of Latimeria are defined as elasmoid-like scales. However the plywood-like organization is considered to be homologous with the bony basal plate of cosmoid scales of extinct sarcopterygian fishes (MEUNIER, 1980; SIRE and HUYSSEUNE, 2003; MONDEJAR, 2018; SCHULTZE, 2018).Cartilages
The cartilaginous tissues in Latimeria are characterized by long chondrocytes (Fig. 8) contrary to those of teleostean fishes that are relatively spherical in shape (MEUNIER, 1979; ZYLBERBERG and MEUNIER, 2008). Cartilage tissues can show an endochondral ossification process. This phenomenon has been studied respectively on the urohyal (PEGUETA, 1968), the MECKEL’s cartilage and the proximal piece of the pectoral girdle (FRANCILLON et al., 1975). The endochondral ossification is relatively limited when it occurs and the volume of enchondral bone remains reduced to thin bony layers (Fig. 8). The cartilage is destroyed by chondroclasts in areas where endochondral ossification occurs. Serial chondrocytes, known at the origin of the calcified cartilage, seem to be lacking in the endochondral ossification process in Latimeria (FRANCILLON et al., 1975). This constitutes a true difference with teleostean endochondral ossifications (MEUNIER, 1979; ZYLBERBERG and MEUNIER, 2008). Tooth tissues
The three tooth morphotypes described in Latimeria are fangs (7–10 mm in height), middle-sized teeth (3–4 mm in height) and rounded tubercles (MILLOT and ANTHONY, 1958). These teeth and tubercles are constituted of a cone of orthodentine set around a large pulp cavity and overlain by an external hypermineralized layer considered as true enamel (GRADY, 1970; CASTANET et al., 1975; SHELLIS and POOLE, 1978; SMITH, 1978; SASAGAWA et al., 1984). Various studies have revealed the presence of globular dentine (Fig. 5) at the periphery of the first third of the tooth (MILLER and HOBDELL, 1968; HOBDELL and MILLER, 1969; SHELLIS and POOLE, 1978; SASAGAWA et al., 1984). This typical histological organization characterizes the various teeth of the bucco-pharyngeal cavity as well as the various odontodes of the dermal bony plates, the fin rays and the scales (CASTANET et al., 1975).
In the fang the inner wall of the pulp cavity displays a series of plies (Fig. 5; MEUNIER et al., 2015). These plies start at the base of the tooth and reach at least the first third to the half of the tooth towards its tip (Fig. 5). Such plies are also present but less developed in the small caniniform teeth (MEUNIER et al., 2015). These dentine plies in the pulp cavity characterizes a plicidentine organization of orthodentine type. These primary plies correspond to a simplexodont plicidentine
46 47
REFERENCES
AGASSIZ, L. 1833–44. Recherches sur les Poissons fossiles. Imprimerie Petitpierre, Neuchâtel. [5 volumes and atlas.]
ANDREWS, S. M. 1973. Interrelationships of crossopterygians. In: GREENWOOD, P. H., MILES, R. S. and PATTERSON, C. (eds.), Interrelationships of Fishes, Acad. Press, London, pp. 137–177.
BERNHAUSER, A. 1961. Zur Knochen- und Zah, histology von Latimeria chalumnae Smith und einiger Fossilformen. Sber. öst. Akad. Wiss., 170: 119–137.
BJERRING, H. C. 1973. Relationships of coelacanthiforms. In: GREENWOOD P. H., MILES R. S. and PATTERSON C. (eds.), Interrelationships of Fishes, Acad. Press, London, pp. 179–205.
BRITO, P. M., MEUNIER, F. J., CLÉMENT, G. and GEFFARD-KURIYAMA, D. 2010. The histological structure of the calcified lung of the fossil coelacanth Axelrodichthys araripensis (Actinistia: Mawsoniidae). Palaeontology, 53: 1281–1290.
CASANE, D. and LAURENTI, P. 2013. Why coelacanths are not ‘living fossils’. Bioessays, 35: 332–338. DOI 10.1002/bies.201200145.
CASTANET, J., MEUNIER, F., BERGOT, C. and FRANÇOIS, Y. 1975. Données pré l iminaires sur les s t ructures histologiques du squelette de Latimeria chalumnae. I - Dents, écailles, rayons de nageoires. In: Coll. Inter. CNRS, "Problèmes actuels de Paléontologie. Evolution des Vertébrés", Paris, 4–9 Juin 1973, 159–168, 4 Pls.
CLEMENT, G. 1999. The actinistian (Sarcopterygii) Piveteauia madagascarensis LEHMAN from the Lower Triassic of Northeastern Madagascar: a redescription on the basis of new material. Journal of Vertebrate Paleontology, 19: 234–242.
CLEMENT, G. 2005. A new coelacanth (Actinist ia, Sarcopterygii) from the Jurassic of France, and the question of the closest relative fossil to Latimeria. Journal of Vertebrate Paleontology, 25: 481–491.
CUPELLO, C., BRITO, P. M., MEUNIER, F. J., HERBIN, M., JANVIER, P., DUTEL, H. & CLÉMENT, G. 2015. Allometric growth in the extant coelacanth lung during ontogenetic development. Nature Communications, 6: 8222. DOI: 10.1038/ncomms9222.
CUPELLO, C., CLEMÉNT, G., MEUNIER, F. J., HERBIN, M., YABUMOTO, Y. and BRITO, P. M. 2019. The long-time adaptation of coelacanths to moderate deep water: reviewing the evidences. Bulletin of Kitakyushu Museum of Natural History and Human History Series A (Natural History), 17: 29–35.
CUPELLO, C., MEUNIER, F. J., HERBIN, M., JANVIER, P., CLÉMENT, G. and BRITO, P. M. 2017. The homology and funct ion of the lung pla tes in extant and foss i l coelacanths. Scientific Reports , 7 : 9244. DOI: 10.1038/s41598-017-09327-6.
EASTMAN, J. T., WITMER, L. M., RIDGELY, R. C. and KUHN, K.
L. 2014. Divergence in skeleta l mass and bone morphology in antarctic notothenioid fishes. Journal of Morphology, 275: 841–861. DOI: 10.1002/jmor.20258
ERDMANN, M. V., CALDWELL, R. L. and MOOSA, M. K. 1998. Indonesian “king of the sea” discovered. Nature, 395: 335.
ERDMANN, M. V., CALDWELL, R. L., JEWETT, S. L. and TJAKRAWIDJAJA, A. 1999. The second recorded living coelacanth from north Sulawesi. Environmental Biology of Fishes, 34: 445–451.
FAURÉ-FREMIET, M. 1936. La structure des fibres d’éastoidine. Archives d'Anatomie Microscopique, 32: 249–269.
FOREY, P. L.- 1984. The coelacanth as a living fossil. In: ELREDGE N. and STANLEY S.M. (eds.), Living fossils. Springer Verlag, Berlin, pp. 166–169.
FOREY, P. L.- 1998. History of the Coelacanth Fishes. Chapman and Hall, London, 419 pp.
FRANCILLON, H., MEUNIER, F., NGO TUAN PHONG, D. and RICQLÈS, A. (DE). 1975. Données préliminaires sur les structures histologiques du squelette de Latimeria chalumnae. I I- Tissus osseux et cartilages. In: Coll. Inter. CNRS, "Problèmes actuels de Paléontologie. Evolution des Vertébrés", Paris, 4–9 Juin 1973, Centre National de la Recherche Scientifique, pp. 169–174, 2 pls.
FRANCILLON-VIEILLOT H., BUFFRENIL V. DE, CASTANET J., GÉRAUDIE J., MEUNIER FJ., SIRE J-Y., ZYLBERBERG L., a n d R I C Q L È S A . D E 1 9 9 0 . M i c r o s t r u c t u r e a n d mineralization of vertebrate skeletal tissues. In: CARTER, J. G. (ed.), Skeletal Biomineralization: Patterns, Processes and Evolutionary Trends. Vol. I , Van Nostrand, New York, pp. 471–530
GARRAULT, H., 1936. Développement des fibres d’élastoidine (actinotrichia) chez les salmonides. Archives d'Anatomie Microscopique, 130: 105–137.
GÉRAUDIE, J. and MEUNIER, F. J. 1980. Elastoidin actinotrichia in coelacanth fins: a comparative study with teleosts. Tissue and Cell, 12: 637–645.
GÉRAUDIE, J. and MEUNIER, F. J. 1984. Structure and comparative morphology of camptotrichia of lungfish fins. Tissue and Cell, 16: 217–236.
GIRAUD, M. M., CASTANET, J., MEUNIER, F. J. and BOULIGAND, Y. 1978a. The fibrous structure of coelacanth scales: a twisted "plywood". Tissue and Cell, 10: 671–686.
GIRAUD, M. M., CASTANET, J., MEUNIER, F. J. and BOULIGAND, Y. 1978b. Organisation spatiale de l'isopédine des écailles du coelacanthe (Latimeria chalumnae SMITH). Comptes Rendus de l’Académie des Sciences, 287: 487–489.
GRADY, J . E. 1970. Tooth development in Latimeria chalumnae (SMITH). Journal of Morphology, 132: 377–388.
HADIATY, R. K. and RACHMATIKA, I. 2003. Morphological study of the scales of Latimeria menadoensis POUYAUD et al., 1999. Treubia (A Journal on Zoology of the Indo-Australian Archipelago), 33: 1–11.
HOBDELL, M. H. and MILLER, W. A. 1969. Radiographic
anatomy of the teeth and tooth supporting tissues of Latimeria chalumnae. Archives of Oral Biology, 14: 855–858.
HUREAU J.-Cl. and OZOUF, C. 1977. Détermination de l’âge et croissance du cœlacanthe Latimeria chalumnae SMITH, 1939 (Poisson, Crossoptérygien, Coelacanthidé). Cybium, 2: 129–137.
JANVIER, P. 1996. Early Vertebrates. Clarendon Press, Oxford, 393 pp.
MEUNIER, F. J. 1979. Etude histologique et microradiographique du cartilage hémal de la vertèbre de la carpe, Cyprinus carpio L. (Pisces, Teleostei, Cyprinidae). Acta Zoologica, 60: 19–31.
MEUNIER, F. J. 1980. Les relations isopédine-tissu osseux dans le post-temporal et les écailles de la ligne latérale de Latimeria chalumnae (SMITH). Zoologica Scripta, 9: 307–317.
MEUNIER, F. J. and ZYLBERBERG, L. 1999. The structure of the external layer and of the odontodes of scales in Latimeria chalumnae (Sarcopterygii, Actinistia, Coelacanthidae) revisited using scanning and transmission electron microscopy. In: SÉRET, B. and SIRE, J.-Y. (eds.), Proc. 5th Indo-Pacific Fish Conf., Nouméa, 1997, Soc. Fr. Ichtyol., Paris, pp. 109–116.
MEUNIER, F. J., ERDMANN, M. V., FERMON, Y. and CALDWELL, R. L. 2008. Can the comparative study of the morphology and histology of the scales of Latimeria menadoensis and L. chalumnae (Sarcopterygii, Actinistia, Coelacanthidae) bring new insight on the taxonomy and the biogeography of recent coelacanthids? Geological Society, London, Special Publications, 295: 351–360.
MEUNIER, F. J., BRITO, P. M. and LEAL, M.-E. 2013. Quelques données morphologiques et histologiques sur la structure de la plaque dentaire linguale d’Arapaima gigas (Osteoglossidae; Teleostei). Cybium, 37: 263–271
MEUNIER, F. J., MONDEJAR-FERNANDEZ, J., GOUSSARD, F., CLÉMENT, G. and HERBIN, M. 2015. Presence of plicidentine in the oral teeth of Latimeria chalumnae (Sarcopterygii; Actinistia; Coelacanthidae). Journal of Structural Biology, 190: 31–37.
MILLER, W. A. 1979. Observations on the structure of mineralized tissue of the coelacanth, including scales and their associate odontodes. Occasional Papers of the Californian Academy of Sciences, 134: 68–78.
MILLER, W. A. and HOBDELL, M. H. 1968. Preliminary report on the histology of the dental and paradental tissues of Latimeria chalumnae (SMITH) with a note on tooth replacement. Archives of Oral Biology, 13: 1289–1291.
MILLOT, J. and ANTHONY, J. 1958. Anatomie de Latimeria chalumnae. T. 1. Squelette, muscles et formations de soutien. CNRS Edr., Paris. 122 pp., 80 pls.
MILLOT, J., ANTHONY, J. and ROBINEAU, D. 1978. Anatomie de Latimeria chalumnae. T. 3. Appareil digestif, téguments, écailles. CNRS Edr., Paris. pp. 7–28, 135–139.
MONDEJAR-FERNANDEZ, J. 2018. On cosmine: its origins, b io logy and impl ica t ions for sarcopterygian interrelationships. Cybium, 42: 41–65.
ØRVIG T. 1951. Histologic studies of Placoderms and fossil Elasmobranchs. 1: The endoskeleton, with remarks on the hard tissues of lower vertebrates in general. Arkiv för Zoologi, 2: 321–454, 8 pl.
ØRVIG T. 1968. The dermal skeleton; general considerations. In: ØRVIG T. (ed.), Current problems of lower vertebrate phylogeny. Proceedings of the fourth Nobel Symposium, Stockholm, 1967, J. Wiley, New-York. pp. 373–397.
PARMENTIER, E., COMPÈRE, P., CASADEWALL, M., FONTENELLE, N., CLOOTS, R. and HENRIST, C. 2008. The rocker bone: a new kind of mineralized tissue? Cell and Tissue Research, 334: 67–79.
PEGUETA, V. P. 1968. Enchondral ossification in Latimeria chalumnae SMITH. Dopovidi Akademii Nauk Ukrainia SSR., 7: 653–656 (in Russian; English abstr.).
POUYAUD, L., WIRJOATMODIO, S., RACHMATIKA, L., TJAKRAWIDJAJA, A., HADIATY, R. K. and HADIE, W. 1999. Une nouvelle espèce de coelacanthe, preuves génétiques et morphologiques. Comptes Rendus de l’Académie des Sciences, 322: 261–267.
ROMER, A.S. 1937. The braincase of the carboniferous crossopterygian Megalichthys nitidus. Bulletin of the Museum of Comparative Zoology, 824: 1–73.
ROMER, A.S. 1942. Cartilage as an embryonic adaptation. American Naturalist, 76: 394–404.
ROUX, G. H. 1942. The microscopic anatomy of the Latimeria scale. South African Journal of Medical Sciences, 7: 1–18.
SASAGAWA, I., ISHIYAMA, M. and KODERA, H. 1984. Fine structure of the pharyngeal teeth in the coelacanth fish (Latimeria chalumnae). In: FEARNHEAD, R. W. and SUGA, S. (eds.), Tooth enamel IV, Elsevier Science Publishers, Amsterdam, pp. 462–466.
SCHULTZE, H.-P. 1969. Die Faltenzähne der Rhipidistiden Crossopterygier, der Tetrapoden und der Actinopterygi-er-Gattung Lepisosteus nebst einer Beschreibung der Zahnstruktur von Onychodus (Struniiformer Crossoptery-gier). Palaeontologica Italica, New Series, 35(65): 63–137.
SCHULTZE, H.-P. 1970. Folded teeth and the monophyletic origin of Tetrapods. American Museum Novitates, 2408: 1–10.
SCHULTZE, H.-P. 2018. Hard tissues in fish evolution: history and current issues. Cybium, 42(1): 29–39.
SHELLIS, R. P. and POOLE, D. F. G. 1978. The structure of the dental hard tissues of the coelacanthid fish Latimeria chalumnae SMITH. Archives of Oral Biology, 23: 1105–1113.
SIRE, J.-Y. and HUYSSEUNE, A. 2003. Formation of dermal skeletal and dental tissues in fish: a comparative and evolutionary approach. Biological Review, 78: 219–249.
SMITH, J. L. B. 1939. A living fish of the Mesozoic type. Nature, 143: 455–456.
SMITH, J. L. B. 1940. A living coelacanth fish from South Africa. Transactions of the Royal Society of South Africa, 28: 1–106, 44 Pls.
SMITH, M. M. 1978. Enamel in the oral teeth of Latimeria chalumnae (Pisces: Actinistia): a scanning electron microscopic study. Journal of Zoology, 185: 355–369.
SMITH, M. M. 1979. Scanning electron microscopy of odontodes in the scales of coelacanth embryo, Latimeria chalumnae SMITH. Archives of Oral Biology, 24: 179–183.
SMITH, M. M., HOBDELL, M. H. and MILLER, W. A. 1972. The structure of the scales of Latimeria chalumnae. Journal of Zoology, London, 167: 501–509.
THOMSON, K. S. 1969. The biology of the lobe-finned fishes.
Biological Reviews, 44: 91–154. UYENO, T. and YABUMOTO, Y. 2007. Origin of extant
coelacanths. The Coelacanth, Fathom the Mystery 2007. Aquamarine Fukushima, pp. 24–26.
ZYLBERBERG, L. and MEUNIER, F. J. 2008. New data on the structure and the chondrocyte populations of the haemal cartilage of abdominal vertebrae in the adult carp Cyprinus carpio (Teleostei, Ostariophysii, Cyprinidae). Cybium, 32: 225–239.
ZYLBERBERG, L., GÉRAUDIE, J., MEUNIER, F. J., & SIRE J. Y., 1992. Biomineralization in the integumental skeleton of the living lower Vertebrates. In: HALL, B. K. (ed), Bone, Volume 4, CRC Press, Boca Raton, pp. 171–224.
François J. MEUNIER, Camila CUPELLO and Gaël CLÉMENT The skeleton and the mineralized tissues of the living coelacanths

Allometric growth in the extant coelacanth lung during ontogenetic development. Nature Communications, 6: 8222. DOI: 10.1038/ncomms9222
DE VOS, L. and OYUGI, D. 2002. First capture of a coelacanth, Latimeria chalumnae SMITH, 1939 (Pisces, Latimeriidae), off Kenya. South African Journal of Science, 98: 345–347.
ERDMANN, M. V., CALDWELL, R. L. and MOOSA, M. K. 1998. Indonesian ‘king of the sea’ discovered. Nature, 395: 335.
FRICKE, H., REINICKE, O., HOFER, H. and NACHTIGALL, W. 1987. Locomotion of the coelacanth Latimeria chalumnae in its natural environment. Nature, 329: 331–333.
FRICKE, H., HISSMANN, K., SCHAUER, J., REINICKE, O., KASANG, L. and PLANTE, R. 1991. Habitat and population size of coelacanth Latimeria chalumnae at Grand Comoro. Environmental Biology of Fishes, 32: 287–300.
FRICKE, H., HISSMANN, K., SCHAUER, J., ERDMANN, M., MOOSA, M. K. and PLANTE, R. 2000. Biogeography of the Indonesian coelacanth. Nature, 403: 38.
HEEMSTRA, P. C., FREEMAN, A. L., WONG, H. Y., HENSLEY, D. A. and RABESANDRATANA, H. D. 1996. First authentic
capture of a coelacanth off Madagascar. South African Journal of Science 92: 150–151.
POUYAUD, L., WIRJOATMODJO, S., RACHMATIKA, I., TJAKRAWIDJAJA, A., HADIATY, R. and HADIE, W. 1999. A new species of coelacanth. Genetic and morphologic proof. C. R. Academy of Science, 322: 261–267.
SMITH, J. L. B. 1939. The living coelacanth fish from South Africa. Nature, 143: 748–750.
SMITH, J. L. B. 1953. The second coelacanth. Nature, 171: 99–101.
SMITH, C. L., RAND, C. S., SCHAEFFER, B. and ATZ, J. W. 1975. Latimeria, the living coelacanth, is ovoviviparous. Science, 190: 1105–1106.
VENTER, P., TIMM, P., GUNN, G., LE ROUX, E., SERFONTEIN, E., SMITH, P., SMITH, E., BENSCH, M., HARDING, D. and HEEMSTRA, P. 2000. Discovery of a viable population of coelacanths (Latimeria chalumnae SMITH, 1939) at Sodwana Bay, South Africa. South African Journal of Science, 96: 567–568.
INTRODUCTION
The first living coelacanth, Latimeria chalumnae, was discovered in South Africa in the Indian Ocean in 1938 (SMITH, 1939). The first observation of the living coelacanth habitats using submersibles was reported in Comoros (FRICKE et al., 1987). Latimeria chalumnae hide in submarine caves along steep slopes between 100 m and 200 m depth during daytime and come out and drift near the ocean floor for hunting prey at night (FRICKE et al., 1991). However, since the record of juvenile or small (below 80 cm long) individuals is virtually absent, the growth, development and breeding biology of L. chalumnae remain poorly known. Our knowledge
of the reproduction biology of the coelacanths is almost restricted to an ovoviviparity strategy since some caught large females carried unfertilized eggs or embryos in their oviduct (SMITH et al., 1975; CUPELLO et al., 2015).
The Indonesian coelacanth, L. menadoensis, was first discovered in Manado, Sulawesi Island, Indonesia, in 1997. The first specimen was captured in 1998 (ERDMANN et al., 1998). According to a genetic study, the specimen differed from L. chalumnae and a new species was erected (POUYAUD et al., 1999). In 1999 two individuals were observed by submersible in a location 360 km away from Manado (FRICKE et al., 2000). Today 8 individuals were officially caught in the Indonesian seas. However, the details of their habitat have not
been described. The aim of this study is to clear their habitat and distribution.
METHODS
Two remotely operated vehicles (ROVs) (Kowa; VEGA300) were used for the surveys. The first ROV was replaced with the second one in 2007. Our ROVs are able to dive up to 300 m depth. These had two vertical, two horizontal and two right-left propellers and these were controlled from a boat on surface through a 400 m long tether cable. The underwater operations were visualized on a screen with information including directions of ROVs, depth, water temperature, date and time. All data were directly recorded on video-tapes on the boat. Since the 2007 survey, water temperature and depth were independently recorded with more accuracy by external measuring memories (Alec Electronics, later named JFE Alec; MDS-MKV/T, MDS-MKV/D). In addition two laser beam irradiators, that provide line lasers of 20-cm distance, were attached to the second ROV and have been used to register the size of encountered animals. The surveys were conducted off northern parts of Sulawesi Island and along the Biak Island located in northwestern New Guinea Island, Indonesia. The areas and time periods of each survey are shown in Table 1 and Fig. 1.
RESULTS
The field survey of the Indonesian coelacanth, Latimeria menadoensis, was conducted 14 times from 2005 to 2015 with 1173 underwater operations around Sulawesi and Biak islands in Indonesia (Fig. 1, Table 1). No coelacanth was encountered during the first surveys in 2005 although 452 underwater operations were conducted around Manado-tua Island and other close Indonesian islands from the 17th to 30th of April (Table 1, R1). The second field survey was held along the northern coast of Sulawesi from the 6th to 19th May in 2006, but once again despite of 107 underwater operations no coelacanth was recorded.
The first individual of Indonesian coelacanth (Table 2, ID 1) was recorded during the third field survey, in the morning of the 30th May 2006 (Table 1, R3). The coelacanth was observed in a cave of 165 m depth (Table 2, Boul-1) for 10 minutes (Table 2, E1). The adult individual was in stationary position in a cave (Buol-1) slowly moving its pectoral, pelvic, second dorsal and anal fins. The day after, another individual (ID 2) and an unidentified one (UN1) were found in the same cave (Buol-1). The unidentified one immediately hid deeper in the cave, where the ROV could not access, and only its lateral side of the body was recorded. The individual (ID 2) was observed for one minute before it hid inside the cave (Table 2: E2). When a new individual could be identified by the white
spots pattern on its body, an ID number was allocated. The individual which could not be distinguished was allocated an UN (unknown) + number.
In the end of the morning of the same day, a third individual (Table 2: E3, ID 3) was found in another cave (Buol-2), larger and located 20 m deeper (Buol-2) than the first cave (Buol-1). After 29 minutes of recording, this individual hid inside the cave. Although the water temperature increased of more than five degrees, from 14.8 to 20.4 ºC, it stayed at the same place, not moving away. Despite it disappeared from the camera frame we kept setting the ROV there in front of the cave until the next morning, however it did not appear again.
On the 4th June in 2006, three individuals, including ID 3 which was observed on the 31th May at Buol-2 and two new individuals, were observed in a vertical crack which was several hundred meters far from Buol-2 (Buol-3) for 2 hours and 14 minutes (Fig. 2C; Table 2: E4, ID 3, 4, 5). During this long period of time all individuals stayed near the rock wall always keeping their head against the water flow. The water
Bull. Kitakyushu Mus. Nat. Hist. Hum. Hist., Ser. A, 17: 49–56, March 31, 2019
Field surveys on the Indonesian coelacanth, Latimeria menadoensis using remotely operated vehicles from 2005 to 2015
ABSTRACT − Habitats of the Indonesian coelacanth, Latimeria menadoensis, were investigated by Remotely Operated Vehicles (ROVs) surveys in the northern coast of Sulawesi Island and southern coast of Biak Island by collaboration of Aquamarine Fukushima (Japan), and Indonesian Institute of Sciences and Sam Ratulangi University (Indonesia) from 2005 to 2015. The Remotely Operated Vehicles operations were conducted 1173 times and coelacanths were encountered 30 times. A total of 30 different individuals were observed at a depth range from 115.6 m to 218.9 m deep. The water temperature was between 12.4 to 21.5 ºC. Most of the individuals were found alone, however, schools of two, three and six individuals were also observed. The Indonesian coelacanth, L. menadoensis, was observed at a similar depth of the African coelacanth, Latimeria chalumnae, and the temperature range also seems to be very similar. However, Latimeria menadoensis was sometimes observed besides big rocks or the steep wall. It seems to be less sensitive to daylight than L. chalumnae. Here we report also, for the first time in the world, a juvenile coelacanth was observed in a small crack at 165 to 171 m depth during these surveys.
KEY WORDS: Indonesian coelacanth, underwater survey, Latimeria menadoensis, Sulawesi, Biak, ROV survey.
Masamitsu IWATA1, Yoshitaka YABUMOTO2, Toshiro SARUWATARI3,4, Shinya YAMAUCHI1, Kenichi FUJII1, Rintaro ISHII1, Toshiaki MORI1, Frensly D. HUKOM5, DIRHAMSYAH5, Teguh PERISTIWADY6, Augy SYAHAILATUA7,
Kawilarang W. A. MASENGI8, Ixchel F. MANDAGI8, Fransisco PANGALILA8 and Yoshitaka ABE1
1Aquamarine Fukushima, Marine Science Museum, 50 Tatsumi-cho, Onahama, Iwaki, Fukushima, 971-8101, Japan2Kitakyushu Museum of Natural History and Human History, 2-4-1 Higashida, Yahata Higashi-ku, Kitakyushu, Fukuoka,
805-0071, Japan3Atmosphere and Ocean Research Institute, The University of Tokyo, 5-1-5 Kashimanohara, Kashiwa-shi, Chiba, 277-8564, Japan
4Seikei Education and Research Center for Sustainable Development, Seikei Gakuen, 3-3-1 Kichijoji-Kitamachi, Musashino-shi, Tokyo, 180-8633, Japan
5Research Center for Oceanography, Indonesian Institute of Science, Jl. Pasir Putih I, Ancol Timur, Jakarta, 14430, Indonesia6Technical Implementation Unit Marine Biota Conservation Bitung, Indonesian Institute of Sciences, Jl. Colombo, Kec, Maesa,
Bitung Tengah, Maesa, Kota Bitung, Sulawesi Utara, 95511, Indonesia7Research Center for Deep Sea, Indonesian Institute of Science, Jl. Y. Syaranamual, Guru-Guru, Poka, Ambon, 97233, Indonesia
8Faculty of Fisheries and Marine Science, Sam Ratulangi University, Jl. Kampus UNSRAT Bahu, Manado, Sulawesi Utara, 95115, Indonesia
(Received August 24, 2018; accepted November 9, 2018)
temperature changed up and down by five degrees during the observation due to water flow reversals. When the water flow was reversed some times, all individuals always changed their directions against the water flow.
During the field survey carried on December of the same year, in the same Buol area (Table 1: R4) the sixth individual (Table 2: E5, ID 6) was recognized at 14:34 on the 12th December alongside a large rock. This individual stayed in the beginning of the observation and then moved ahead to deeper slowly. Eventually it reached 12 m deeper during one hour and 42 minutes.
On the 14th December, the individual ID 3 previously recorded at Buol-2 in a cave and at Buol-3 in a crack, was encountered again (Table 2: Buol-5) near a vertical wall at 145 m depth (Table 2: E6). After a while, this individual (ID 3) moved upward about 18 m and stayed behind a large rock (Fig. 3). The observation time was the longest of all surveys with four hours and 16 minutes of recording. The temperature changed up and down several times (Fig. 4). Its difference
between the maximum and the minimum was 6.7 °C, which was the largest range of all surveys.
On the next day, on the 15th December, the ID 3 was observed again under an overhang (Table 2: E7, Buol-6) near other sites where it was previously found at Buol-2 , Buol-3 and Buol-5. Its posture was upside down with its ventral side facing the ceiling of the overhang. Such posture is also known in coelacanths from the Comorian Archipelago (FRICKE et al., 1987). ID 3 was found totally four times in this area in two surveys during the seven months and seems to live in this area.
During the 2007 survey (Table 1: R5) a coelacanth (Table 2: ID 7) was recorded off Manado by 195 m depth, with a temperature of 12.4 °C, the coolest one registered during all surveys.
The field survey in 2009 was carried out off Manado and around Talise and Bangka islands off the northern coast of Sulawesi Island from the 12th September to the 9th October (Table 1: R8). On the 24th September 2009, six individuals (Table 2: E11, ID 9, 10, 11, 12, UN 2, 3) were found under an
overhang off Talise Island between 144 and 150 m deep. There was almost no water current in this area. All individuals were close to each other under the same overhang and most of them stayed still with their head facing downward. Four days later, ID 9 was observed again, alone on a steep slope at 172.9 m depth (Table 2: E12), which was located within several dozen meters far from Talise-2 and 30 m deeper.
Four new coelacanths were recorded from the 29th September to the 6th October 2009 in Talise and Banggka Islands and off Manado (Table 2: E13 and 14, ID 13, 14, 15). ID 15 is the only juvenile coelacanth filmed so far. It was in a small crack between 164.6 and 170.9 m deep in a temperature between 14.5 and 15 °C.
The shallowest record of a coelacanth was at 115.6 m depth off the southern part of Manado (Table 2: E16, Manado-3) on the 9th October in 2009. ID 16 (Table 2) was found at the end of the morning, alone alongside a large rock on the edge of a shelf. The water temperature was about 20.0 ºC.
From the 6th to the 16th November in 2010, the field survey was conducted around Biak Island (Table 1: R9). Adult coelacanths were found at two different sites (Table 2: E17–19). Two individuals (ID 17 and 18) were found under an overhang between 212.5 and 218.9 m deep (Table 2: Biak-1) on the 11th November. Three other ones (ID 19, 20, 21) were registered under an overhang between 193.2 and 195.9 m deep (Table 2: Biak-2) on the 13th November. Two days later, Biak-2 was observed again and only ID 20 stayed there.
Two new coelacanths were recorded off Manado in December 2010 (Table 1: R10; Table 2: E 20 and 21; ID 22 and 23).
On the 4th May 2012, an individual (Table 2: E22, ID 24) was found on a steep slope at 169.4 m depth off Manado (Table 2: Manado-6). It swam up and down 5.4 m between 168.1 and 173.5 m deep for 17 minutes. Thirteen months later (the 9th June 2003), this individual (ID 24) was observed again near a cliff at 152.3 m depth (Table 2: E24, Manado-8), where located on the same slope of Manado-6.
Another new coelacanth (Table 2: E23, ID 25) was recorded alone on a Manado site (Manado-7).
Field surveys were conducted from the 19th to 30th May 2015 off Lolak Island and from the 30th October to the 16th November 2015 off Lolak and Bitung Islands (Table 1; R13 and R14). Three new individuals (Table 2: ID 26–28) were recognized at different places off Lolak Island (Lolak 1-4). All individuals were alone. ID 26 was observed in May and once
again in October 2015 on a steep slope (Table 2: E25, E27). The coelacanth ID 28 was found on the 1st November along one of the large rocks scattered on a gentle slope in a bay of the Lolak Island by 125 m depth.
The encountered Indonesian coelacanths during these surveys were in caves, alongside large rocks, under overhangs or on steep slopes (Table 2; Fig. 2).
Most individuals were found under overhangs or alongside large rocks. Some individuals were encountered not to hide in any shade and stayed just on rocky slopes. In overhangs and cracks, all individuals had their ventral side alongside and close to a rock wall, but without touching it.
Estimated total lengths are shown in Table 3. Most individuals are more than 1 m in total length. ID 15 is a juvenile individual of 31.5 cm and ID 23 is 90 cm in total length. No individual beyond 140 cm has been recorded in Indonesia so far.
DISCUSSION
During the field surveys by underwater ROV recording for Indonesian coelacanths, Latimeria menadoensis, from 2005 to 2015, 30 different individuals were identified and three others were unidentified. Among those, six individuals were observed more than twice: ID 3 at Buol-2, 3, 5, 6 (Table 2: E3, E4, E6 and E7); ID 8 at Talise-1 (E9 and E10), ID 9 at Talise 2 and 3 (E11 and E12), ID 20 at Biak-2 (E18 and E19), ID 24 at Manado-6 and 8 (E22 and E24), ID 26 at Lolak-1 and 3 (E25 and E27). ID 8 was observed on the 14th September 2009 and was encountered again at the same place (Talise-1) the day after. ID 24 was observed on the 4th May 2012 and then 13 months later, on the 9th June 2013 at the same locations of Manado area within several kilometers.
Al l individuals observed more than twice were encountered in the same area, at locations close from each other within hundreds of meters. However, in the Comorian Archipelago, same individuals of L. chalumnae were observed, within a two week period, in several caves distributed in a 8 kilometers wide area (FRICKE et al., 1991).
When individuals of L. menadoensis were encountered, some of them stayed stationary at same place but some individuals such as ID 6 on the 12th December 2006 (E5) and ID 24 on the 4th May 2012 (E22) swam away. It seemed to avoid and escape from the brightness of the light or the sounds
of the propellers of the ROV.Temperature and depth data for each Indonesian
coelacanth record are shown in Fig. 3. Though an average of water temperature on the ocean surface was about 30 °C, the temperature often dropped into about 11 °C at 300 m water depth. The temperatures changed widely even though the changes of depth were small at several places. All Indonesian coelacanths, L. menadoensis, were encountered during day time at a depth between 115.6 and 218.9 m and a temperature between 12.4 and 21.5 ºC. Indonesian coelacanths have exceptionally been recorded in an environment with a temperature exceeding 20 ºC, regardless of the depth. The African coelacanth, L. chalumnae, was observed by submarine vehicles in steep slopes between 150 m and 253 m (FRICKE et al., 1991). Divers recorded the presence of this species at 104 m depth at Sodwana Bay, South Africa (VENTER et al., 2000). Latimeria chalumnae seem to prefer a temperature range of 15–20 °C, where they tend to choose their daytime habitats (FRICKE et al., 1991). The suitable temperature for L. menadoensis seems to be almost the same as L. chalumnae.
Water temperature can increase or drop for more than 5 °C in a very short period of time (about 30 min according to our recordings). The coelacanths however often stayed there without moving away or significantly modifying their behavior. These water currents of different temperatures are caused by the movement of the marine thermocline. During our surveys we observed in some area that as the thermocline
level changed, the water flow was reversed. Coelacanths always kept their head against the water flow, like other observed actinopterygian fishes did. It means that as the direction of the water flow changed, the swimming direction of each coelacanth individual changed accordingly.
On the 24th September 2009 (Table 2: E11) six individuals were recognized under an overhang (Talise-2). The all had about the same size. There was almost no water flow, and each individual was head downward. They were close to each other but no direct contacts or interactions between coelacanths were observed. No evidence of a social behavior has been recognized in the Comorian populations of coelacanths (FRICKE et al., 1991).
According to the observations of the African coelacanths, the species L. chalumnae is nocturnal and hides in cave in daytime but comes out at night, supposedly in search of food (FRICKE et al., 1991). The living geomorphologic environment of L. menadoensis is similar to that of L. chalumnae, with rocky steep slopes with undermarine caves or overhangs. However, Latimeria menadoensis was also observed alongside large rocks or along vertical wall in daytime. Some of these individuals were encountered above 200 m deep, in an environment that remains sl ightly bright . Latimeria menadoensis seems to be less sensitive to the daylight than L. chalumnae.
These first observations are potentially crucial since the habitats of the Indonesian coelacanths could be possibly not as
restricted as those of the African coelacanths, and so a broader distribution of the Indonesian coelacanth could be expected. Occurrences of coelacanths along the eastern coast of the African continent were recorded in South Africa (SMITH, 1939), Comoros (SMITH, 1953), Madagascar (HEEMSTRA et al., 1996), Kenya (DE VOS and OYUGI, 2002), Tanzania (BENNO et al., 2006), and Mozambique (BRUTON et al., 1992). In Indonesia, coelacanths have only been found off the northern coast of the Sulawesi Island and the southern coast of the Biak Island in northern New Guinea.
Field surveys of the Indonesian coelacanths should be continued in all area of Indonesia and expanded to all Southeast Asia in order to increase our knowledge of the distribution of this rare species and to define, in a close future, the most relevant conservation policies of the different known populations.
ACKNOWLEDGMENTS
We are grateful to Dr. Teruya UYENO for his valuable advice, guidance and encouragement throughout the present study. Many thanks go to colleagues of the Faculty of Fisheries and Marine Sciences of the Sam Ratulangi University at Manado, Indonesia, for their generous supports to conduct expeditions. Also we express our thanks to Dr. SUHARSONO, Dr. Zainal ARIFIN, Dr. DIRHAMSYAH and colleagues of the Indonesian Institute of Science for their advice and administrative work to obtain our research permit. Other thanks go to Mr. OPO, Mr. REFRY and colleagues at Murex Dive Resort, and DAUD, AL,
and other staffs for our work on the research boat. Dr. BATUNA and the Murex staffs supported our daily life during the expeditions. We would like to express our sincere thanks to colleagues at Aquamarine Fukushima, Japan, for their understanding and supports. We would like to thank Dr. Gaël CLÉMENT of Muséum national d’Histoire naturelle and Dr. Camila CUPELLO of Universidade do Estado do Rio de Janeiro as a referee for their critical reading of the manuscript and their comments.
Unfortunately one of our authors, Dr. Djoko Hadi KUNARSO has passed away during the process of this contribution. We pray for the response of his soul.
This study was supported in part by funding from Interdisciplinary Collaborative Research program of Atmosphere and Ocean Research Institute, the University of Tokyo.
REFERENCES
BENNO, B., VERHEIJI, E., STAPLEY, J., RUMISHA, C., NGATUNGA, B., ABDALLAH, A. and KALOMBO, H. 2006. Coelacanth (Latimeria chalumnae SMITH, 1939) discoveries and conservation in Tanzania. South African Journal of Science, 102: 486–490.
BRUTON, M. N., CABRAL, A. J. P. and FRICKE, H. 1992. First capture of a coelacanth, Latimeria chalumnae (Pisces, Latimeriidae), off Mozambique. South African Journal of Science, 88: 225–227.
CUPELLO, C., BRITO, P. M., HERBIN, M., MEUNIER, F. J., JA N V I E R, P . , DU T E L, H. and CL É M E N T, G. 2015.
type according to the definition of MEUNIER et al. (2013). This organization is less complex than the three other plicidentine types (polyplocodont, eusthenodont and dendrodont), which are described in most of extinct and extant Sarcopterygii (SCHULTZE, 1969, 1970).The bony plates of the lung
The lung of Latimeria is reduced to a short oesophagus diverticulum (CUPELLO et al., 2015) surrounded by scattered small and thin ovoid plates (Fig. 9A, B). These plates cover the surface of the vestigial lung, and are composed of a fibrous matrix with fibrocytes: it is a cellular bony tissue (CUPELLO et al., 2017). Each plate is enclosed in a membranous bag. These plates look fragile in their middle plane since a central weakness zone opened on the microtome knife, creating an artefactual lumen (Fig. 9C) (CUPELLO et al., 2017). The fibres are collagenous and organized in superposed layers. The cells are true osteocytes (Fig. 9E) with a more or less star-shaped outline, and send cytoplasmic extensions in the thickness of the plate (Fig. 9D, E). Rare lining cells (osteoblasts) are
present on the surface of the plates, and conjunctive fibres (SHARPEY’s fibres) penetrate within the plate (Fig. 9C). The extracellular matrix shows various staining intensities with concentric and parallel (Fig. 9D) or globular shaped lines. These lines are considered to be Liesegang lines that characterize an active spheritic mineralizing process. The lung plates of Latimeria are thus true bony plates, and are homologous with the large bony plates known in the abdominal cavity of fossil actinistians (BRITO et al., 2010; CUPELLO et al., 2017). The calcified walls that surrounded the lung of the Cretaceous coelacanth Axelrodichthys are made of large osseous plates of various thickness (BRITO et al., 2010), as those of other fossil coelacanths (CLEMENT, 1999, 2005). The bony tissue of these plates is a vascularized cellular bone with more or less large vascular cavities and some internal remodelling (BRITO et al., 2010). Moreover the mineralization of the lung ossified plates in Axelrodichthys is spheritic as in Latimeria’s plates (CUPELLO et al., 2017). So there is a continuity of the histological structure of the lung plates in the coelacanths during their
evolution, but with an important reduction of the bony plates in Latimeria linked to the vestigial state of its lung (CUPELLO et al., 2015, 2019).
CONCLUSION
This overview of eighty years (1938–2018) of histological work on Latimeria skeletal tissues allows some anatomical and evolutionary considerations. It can be enlightened a drastic reduction of endochondral ossification during the long evolutionary history of coelacanths. The persistence of large volume of cartilage in the endoskeleton at adult stage can be compared to the “little bone and considerable cartilage” that characterize the skeleton of a number of demersal notothenioid telostean fishes (EASTMAN et al., 2014).
A processus of spheritic mineralization has been recently highlighted in various skeletal elements of Latimeria, by the presence of globular dentine in teeth, in odontodes of the tegumentary skeleton (scales, fin rays), in scales at the interface between the external layer and the basal plate, as well as in lung bony plates. The spheritic mineralization (i.e., a radiating arrangement of hydroxyapatite crystals and of the organic matrix) in vertebrate skeletal tissues is considered as a precursor of inotropic mineralization (specific interactions between collagen fibrils and the mineral phase) that possibly represents a derived evolutionary stage of calcification mechanisms (ØRVIG, 1951, 1968; FRANCILLON-VIEILLOT et al., 1990; ZYLBERBERG et al., 1992). The simultaneous presence of both mineralization processes (spheritic and inotropic) in the mineralized tissues of Latimeria and in other osteichthyans such as teleosteans (ZYLBERBERG and MEUNIER, 2008) is however questionable. The mineralized spherules present at the limit between the external layer and the basal plate of Latimeria scales are considered to be the product of an inotropic mineralization (MEUNIER and ZYLBERBERG, 1999). It thus appears that the state of the mineralized spherules observed in the various skeletal tissues of Latimeria, including the lung bony plates, must be studied with adapted ultrastructural techniques in order to test their true origin: spheritic or inotropic mineralization?
ACKNOWLEDGEMENTS
This contribution is linked to an international roundtable on coelacanths evolution held at the Kitakyushu Museum and at the Aquamarine Fukushima in August 2017. We would like to thank the anonymous reviewers for their constructive comments and valuable suggestions. C.C. was partially financed by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brasil (CAPES) – Finance Code 001 (Programa Nacional de Pós Doutorado-PNPD).
INTRODUCTION
The first study of fossil coelacanths was done by AGASSIZ (1833–44) and was followed by a lot of papers with the discovery of new fossils (see reviews of JANVIER, 1996 and FOREY, 1998). Fossil coelacanths constitute an important monophyletic group composed of more than forty genera, eighty species, and belong to the Sarcopterygii (ANDREWS, 1973; JANVIER, 1996; FOREY, 1998; UYENO and YABUMOTO, 2007; and many others). For a century coelacanths were considered extinct since the end of the Cretaceous time (Fig. 1).
With the discovery of a living coelacanth in 1938 (SMITH, 1939) the scientific community was soon highly interested to compare the incomplete fossil sarcopterygian fishes dataset to the recent anatomical and biological organization of Latimeria chalumnae. Many authors (see THOMSON, 1969; MILLOT and ANTHONY, 1958; MILLOT et al., 1978 FOREY, 1984, 1998; JANVIER, 1996) focused their work on Latimeria as a key-taxon to test or propose various hypotheses on the origin and evolutionary history of the early tetrapods and sarcopterygii in general, and to infer their possible biological characteristics. The fossil sarcopterygian fishes are known by their fossilized mineralized tissues, essentially their bones, scales and teeth. Together with the extant lungfishes, especially Neoceratodus, Latimeria offered a unique access to the skeleton as a whole: with both mineralized and unmineralized skeletal tissues. Here, we aim to present an overview of the last eighty years (1938–2018) of histological studies of Latimeria’s skeleton.
Since the discovery of the Raja Laut (“king of the sea” ), off Sulawesi Island in Indonesia (ERDMANN et al., 1998, 1999), the genus Latimeria comprises two species: L. chalumnae SMITH 1939 and L. menadoensis POUYAUD et al., 1999. However our knowledge of the anatomy and biology of the living coelacanths is so far essentially based on L. chalumnae.
THE LATIMERIA SKELETON
The first anatomical observations on the extant coelacanth were done by SMITH (1940). The anatomy of the Latimeria skeleton has been later described in detail by MILLOT and ANTHONY (1958). The living coelacanth shows a general organization similar to that of the other actinistian fishes (THOMSON, 1969; JANVIER, 1996; FOREY, 1998). The skeleton can be divided in various parts (Fig. 2): skull, axial skeleton, paired and unpaired fins, tegumentary skeleton (scales). The skull, axial skeleton and fins are constituted of endoskeletal elements that are overlain by exoskeletal elements. In the extant Latimeria the endoskeleton of the skull shows an important regression of the bones, especial ly in the neurocranium, which are replaced by cartilaginous tissues, as in its Mesozoic fossil actinistian relatives (ROMER, 1937, 1942; MILLOT and ANTHONY, 1958; BJERRING, 1973; FOREY, 1998). In the same way, the endoskeleton of paired and unpaired fins is essentially constituted of four axial cartilaginous elements (Fig. 2; MILLOT and ANTHONY, 1958), with pre and post-axial
elements and eventually minute superficial endochondral ossification (CASTANET et al., 1975). The axial skeleton is composed of the notochord which is coated by an unmineralized fibrillary sheath and is totally deprived of well developed vertebrae due to the lack of ossified centra, although some discrete neural and haemal arches develop in the anterior part of the notochord (MILLOT and ANTHONY, 1958). The anterior neural and haemal spines are relatively short but they progressively increase in length posteriorly. Importantly neural and haemal spines are composed of perichondral bone surrounding a cartilaginous core (Fig. 2).
The scales belong to the exoskeleton. In both extant coelacanth species they are of elasmoid type, composed of an upper external layer also called “external ornamented layer”, and of a lower thicker layer, called the basal plate, which is stratified and almost totally unmineralized (Fig. 3). The upper external layer is ornamented with radial crests. In the posterior area of the scale these radial reliefs are overlain by numerous denticulations, the odontodes, which can be superposed (Fig. 3). The anterior area of the external layer, which is covered by the anterior scales, shows crests with irregularities that have been interpreted as growth marks and tentatively used for ageing coelacanths (HUREAU and OZOUF, 1977; MILLER, 1979).
The tooth plates of the buccal cavity support series of teeth of various sizes (Fig. 4). They range in three morphotypes: the fangs (7–10 mm in height), middle-sized teeth (3–4 mm in height) and rounded tubercles (MILLOT and ANTHONY, 1958). The teeth of the two first categories are conical and sharp whereas those of the third category have an obtuse tip (Fig. 5). Fangs of Latimeria are inserted into a socket (MILLOT and ANTHONY, 1958; HOBDELL and MILER, 1969; CASTANET et al., 1975) and they have a smooth external surface (MILLOT et al., 1978; MEUNIER et al., 2015).
A particularity of the exoskeleton of Latimeria is the abundance of odontodes at the surface of various skull bones
(MILLOT and ANTHONY, 1958; BERNHAUSER, 1961), on the fin rays (CASTANET et al., 1975) and on the posterior (free) area of scales (Fig. 3) (MILLOT and ANTHONY, 1958; CASTANET et al., 1975; HADIATY and RACHNATIKA, 2003; MEUNIER et al., 2008).
Among the various components of the whole skeleton, and apart the SMITH’s pioneer work (SMITH, 1940), numerous studies have been carried out on the histological organization of scales (ROUX, 1942; SMITH et al., 1972; CASTANET et al., 1975; GIRAUD et al., 1978a; MILLER, 1979; SMITH, 1979; MEUNIER, 1980; MEUNIER and ZYLBERBERG, 1999; HADIATY and RACHMATIKA, 2003; MEUNIER et al., 2008) and teeth (MILLER and HOBDELL, 1968; GRADY, 1970; HOBDELL and MILLER, 1969; CASTANET et al., 1975; SHELLIS and POOLE, 1978;
SMITH, 1978; SASAGAWA et al., 1984; MEUNIER et al., 2015). Contrary to the superficial skeleton, there are clearly less histological studies of the skeletal bony elements (FRANCILLON et al., 1975). Meanwhile, in addition to the usual skeletal elements, specific bony tissues have been described in fossil (BRITO et al., 2010) and extant coelacanths (CUPELLO et al., 2015, 2017), such as the mineralized plates surrounding the lung. These bony plates in actinistians do not belong to the skeleton sensu stricto. They are specific bony specializations in relation to the lung (see below), as is the “rocker bone” in relation to the gas bladder of some ophidiiform teleosts (PARMENTIER et al., 2008). Despite the name, the rocker bone is not true bony tissue (PARMENTIER et al., 2008), contrary to
the bony plates of fossil and extant coelacanths (BRITO et al., 2010; CUPELLO et al., 2017).
ADULT LATIMERIA SKELETAL TISSUES
Bony tissuesCortical areas of the dermal skeleton are constituted of
primary periosteal bony t issue. I ts mineralization is heterogeneous and generally marked by series of growth lines, probably due to alternative physiological cycles linked to seasonal variations (Fig. 6). The cortical primary bone shows vascular canals and numerous SHARPEY’s fibres (Fig. 6B). The centre of these bones is frequently made of a spongiosa with vascular cavities surrounded by secondary bony deposits that result from remodelling processes (Fig. 6). This secondary bone is separated from the primary one by reversal cementing lines that are hypermineralized.
Bone remodelling in adult specimens can be more or less developed according to a bone and within a given bone. Primary bone is destroyed by osteoclasts and the deposition process of the secondary bone results from the activity of osteoblasts that have replaced the osteoclasts. The secondary bone does generally not replaced the whole primary bone areas, which can still be recognizable by the presence of
SHARPEY’s fibres and/or growth marks (Fig. 6).The dermal fin rays that sustain paired and unpaired fins
in Latimeria are true lepidotrichia (CASTANET et al., 1975). They are made of two parallel opposite gutter-shaped hemirays, each of which being a series of hemisegments articulated by a collagenous ligament. Each hemisegment is made of a mineralized collagenous fibrillary matrix with embedded osteocytes. The hemisegments are sometimes fused at the basal part of the rays due to centrifugal deposition of bony laminae around the primary articulation, and eventually to the mineralization of the ligament (see figs. 19, 20 in CASTANET et al., 1975). Odontodes observed on the external convex surface of the rays are similar to those present on the dermal bones of the skull and scales. The distal extremity of the lepidotrichia is overlapped by actinotrichia (GÉRAUDIE and MEUNIER, 1980) that are long tapered rods of elastoidin , a fibrous protein of collagenous nature (FAURÉ-FREMIET, 1936; GARRAULT, 1936). The presence of lepidotrichia with distal actinotrichia in Latimeria, as in the fins of Actinopterygii, is
regarded as a plesiomorphic character for Osteichthyes (GÉRAUDIE and MEUNIER, 1980, 1984).
The upper layer of the Latimeria scale is relatively thin and it is constituted of bony tissue with embedded osteocytes, whereas the basal plate is much thicker and composed of numerous strata made of thick collagenous fibres (Fig. 7). Between two collagenous layers there are star-shaped cells (SMITH et al., 1972, pl. VI; CASTANET et al., 1975, fig. 12), the elasmocytes, which cytoplasmic processes insert between the collagenous fibres. These fibres are set in a very specific network. In each layer, the fibres are parallel to each other and the direction of the fibres change from a layer to another one. This regular organization results in a spatial arrangement termed “twisted plywood” (GIRAUD et al., 1978a,b). The rotation of fibres direction from one layer to the next has a mean angle of 27° (GIRAUD et al., 1978a). This organization is found in each scale of L. chalumnae. In L. menadoensis, the basal plate of the scales is also a twisted plywood but its rotation angle seems to be slightly less regular (MEUNIER et al.,
2008). The basal plate in both species is unmineralized (Fig. 7) excepted at the contact between the superficial layer and the basal plate, where spheritic mineralized granules are seen in the very first layers of the basal plate (MEUNIER, 1980; MEUNIER and ZYLBERBERG, 1999; MEUNIER et al., 2008). There is no pore-canal system so Latimeria’s scales are not cosmoid, contrary to most of palaeozoic sarcopterygian fishes. Due to the presence of an unmineralized stratified basal plate, the scales of Latimeria are defined as elasmoid-like scales. However the plywood-like organization is considered to be homologous with the bony basal plate of cosmoid scales of extinct sarcopterygian fishes (MEUNIER, 1980; SIRE and HUYSSEUNE, 2003; MONDEJAR, 2018; SCHULTZE, 2018).Cartilages
The cartilaginous tissues in Latimeria are characterized by long chondrocytes (Fig. 8) contrary to those of teleostean fishes that are relatively spherical in shape (MEUNIER, 1979; ZYLBERBERG and MEUNIER, 2008). Cartilage tissues can show an endochondral ossification process. This phenomenon has been studied respectively on the urohyal (PEGUETA, 1968), the MECKEL’s cartilage and the proximal piece of the pectoral girdle (FRANCILLON et al., 1975). The endochondral ossification is relatively limited when it occurs and the volume of enchondral bone remains reduced to thin bony layers (Fig. 8). The cartilage is destroyed by chondroclasts in areas where endochondral ossification occurs. Serial chondrocytes, known at the origin of the calcified cartilage, seem to be lacking in the endochondral ossification process in Latimeria (FRANCILLON et al., 1975). This constitutes a true difference with teleostean endochondral ossifications (MEUNIER, 1979; ZYLBERBERG and MEUNIER, 2008). Tooth tissues
The three tooth morphotypes described in Latimeria are fangs (7–10 mm in height), middle-sized teeth (3–4 mm in height) and rounded tubercles (MILLOT and ANTHONY, 1958). These teeth and tubercles are constituted of a cone of orthodentine set around a large pulp cavity and overlain by an external hypermineralized layer considered as true enamel (GRADY, 1970; CASTANET et al., 1975; SHELLIS and POOLE, 1978; SMITH, 1978; SASAGAWA et al., 1984). Various studies have revealed the presence of globular dentine (Fig. 5) at the periphery of the first third of the tooth (MILLER and HOBDELL, 1968; HOBDELL and MILLER, 1969; SHELLIS and POOLE, 1978; SASAGAWA et al., 1984). This typical histological organization characterizes the various teeth of the bucco-pharyngeal cavity as well as the various odontodes of the dermal bony plates, the fin rays and the scales (CASTANET et al., 1975).
In the fang the inner wall of the pulp cavity displays a series of plies (Fig. 5; MEUNIER et al., 2015). These plies start at the base of the tooth and reach at least the first third to the half of the tooth towards its tip (Fig. 5). Such plies are also present but less developed in the small caniniform teeth (MEUNIER et al., 2015). These dentine plies in the pulp cavity characterizes a plicidentine organization of orthodentine type. These primary plies correspond to a simplexodont plicidentine
REFERENCES
AGASSIZ, L. 1833–44. Recherches sur les Poissons fossiles. Imprimerie Petitpierre, Neuchâtel. [5 volumes and atlas.]
ANDREWS, S. M. 1973. Interrelationships of crossopterygians. In: GREENWOOD, P. H., MILES, R. S. and PATTERSON, C. (eds.), Interrelationships of Fishes, Acad. Press, London, pp. 137–177.
BERNHAUSER, A. 1961. Zur Knochen- und Zah, histology von Latimeria chalumnae Smith und einiger Fossilformen. Sber. öst. Akad. Wiss., 170: 119–137.
BJERRING, H. C. 1973. Relationships of coelacanthiforms. In: GREENWOOD P. H., MILES R. S. and PATTERSON C. (eds.), Interrelationships of Fishes, Acad. Press, London, pp. 179–205.
BRITO, P. M., MEUNIER, F. J., CLÉMENT, G. and GEFFARD-KURIYAMA, D. 2010. The histological structure of the calcified lung of the fossil coelacanth Axelrodichthys araripensis (Actinistia: Mawsoniidae). Palaeontology, 53: 1281–1290.
CASANE, D. and LAURENTI, P. 2013. Why coelacanths are not ‘living fossils’. Bioessays, 35: 332–338. DOI 10.1002/bies.201200145.
CASTANET, J., MEUNIER, F., BERGOT, C. and FRANÇOIS, Y. 1975. Données pré l iminaires sur les s t ructures histologiques du squelette de Latimeria chalumnae. I - Dents, écailles, rayons de nageoires. In: Coll. Inter. CNRS, "Problèmes actuels de Paléontologie. Evolution des Vertébrés", Paris, 4–9 Juin 1973, 159–168, 4 Pls.
CLEMENT, G. 1999. The actinistian (Sarcopterygii) Piveteauia madagascarensis LEHMAN from the Lower Triassic of Northeastern Madagascar: a redescription on the basis of new material. Journal of Vertebrate Paleontology, 19: 234–242.
CLEMENT, G. 2005. A new coelacanth (Actinist ia, Sarcopterygii) from the Jurassic of France, and the question of the closest relative fossil to Latimeria. Journal of Vertebrate Paleontology, 25: 481–491.
CUPELLO, C., BRITO, P. M., MEUNIER, F. J., HERBIN, M., JANVIER, P., DUTEL, H. & CLÉMENT, G. 2015. Allometric growth in the extant coelacanth lung during ontogenetic development. Nature Communications, 6: 8222. DOI: 10.1038/ncomms9222.
CUPELLO, C., CLEMÉNT, G., MEUNIER, F. J., HERBIN, M., YABUMOTO, Y. and BRITO, P. M. 2019. The long-time adaptation of coelacanths to moderate deep water: reviewing the evidences. Bulletin of Kitakyushu Museum of Natural History and Human History Series A (Natural History), 17: 29–35.
CUPELLO, C., MEUNIER, F. J., HERBIN, M., JANVIER, P., CLÉMENT, G. and BRITO, P. M. 2017. The homology and funct ion of the lung pla tes in extant and foss i l coelacanths. Scientific Reports , 7 : 9244. DOI: 10.1038/s41598-017-09327-6.
EASTMAN, J. T., WITMER, L. M., RIDGELY, R. C. and KUHN, K.
L. 2014. Divergence in skeleta l mass and bone morphology in antarctic notothenioid fishes. Journal of Morphology, 275: 841–861. DOI: 10.1002/jmor.20258
ERDMANN, M. V., CALDWELL, R. L. and MOOSA, M. K. 1998. Indonesian “king of the sea” discovered. Nature, 395: 335.
ERDMANN, M. V., CALDWELL, R. L., JEWETT, S. L. and TJAKRAWIDJAJA, A. 1999. The second recorded living coelacanth from north Sulawesi. Environmental Biology of Fishes, 34: 445–451.
FAURÉ-FREMIET, M. 1936. La structure des fibres d’éastoidine. Archives d'Anatomie Microscopique, 32: 249–269.
FOREY, P. L.- 1984. The coelacanth as a living fossil. In: ELREDGE N. and STANLEY S.M. (eds.), Living fossils. Springer Verlag, Berlin, pp. 166–169.
FOREY, P. L.- 1998. History of the Coelacanth Fishes. Chapman and Hall, London, 419 pp.
FRANCILLON, H., MEUNIER, F., NGO TUAN PHONG, D. and RICQLÈS, A. (DE). 1975. Données préliminaires sur les structures histologiques du squelette de Latimeria chalumnae. I I- Tissus osseux et cartilages. In: Coll. Inter. CNRS, "Problèmes actuels de Paléontologie. Evolution des Vertébrés", Paris, 4–9 Juin 1973, Centre National de la Recherche Scientifique, pp. 169–174, 2 pls.
FRANCILLON-VIEILLOT H., BUFFRENIL V. DE, CASTANET J., GÉRAUDIE J., MEUNIER FJ., SIRE J-Y., ZYLBERBERG L., a n d R I C Q L È S A . D E 1 9 9 0 . M i c r o s t r u c t u r e a n d mineralization of vertebrate skeletal tissues. In: CARTER, J. G. (ed.), Skeletal Biomineralization: Patterns, Processes and Evolutionary Trends. Vol. I , Van Nostrand, New York, pp. 471–530
GARRAULT, H., 1936. Développement des fibres d’élastoidine (actinotrichia) chez les salmonides. Archives d'Anatomie Microscopique, 130: 105–137.
GÉRAUDIE, J. and MEUNIER, F. J. 1980. Elastoidin actinotrichia in coelacanth fins: a comparative study with teleosts. Tissue and Cell, 12: 637–645.
GÉRAUDIE, J. and MEUNIER, F. J. 1984. Structure and comparative morphology of camptotrichia of lungfish fins. Tissue and Cell, 16: 217–236.
GIRAUD, M. M., CASTANET, J., MEUNIER, F. J. and BOULIGAND, Y. 1978a. The fibrous structure of coelacanth scales: a twisted "plywood". Tissue and Cell, 10: 671–686.
GIRAUD, M. M., CASTANET, J., MEUNIER, F. J. and BOULIGAND, Y. 1978b. Organisation spatiale de l'isopédine des écailles du coelacanthe (Latimeria chalumnae SMITH). Comptes Rendus de l’Académie des Sciences, 287: 487–489.
GRADY, J . E. 1970. Tooth development in Latimeria chalumnae (SMITH). Journal of Morphology, 132: 377–388.
HADIATY, R. K. and RACHMATIKA, I. 2003. Morphological study of the scales of Latimeria menadoensis POUYAUD et al., 1999. Treubia (A Journal on Zoology of the Indo-Australian Archipelago), 33: 1–11.
HOBDELL, M. H. and MILLER, W. A. 1969. Radiographic
anatomy of the teeth and tooth supporting tissues of Latimeria chalumnae. Archives of Oral Biology, 14: 855–858.
HUREAU J.-Cl. and OZOUF, C. 1977. Détermination de l’âge et croissance du cœlacanthe Latimeria chalumnae SMITH, 1939 (Poisson, Crossoptérygien, Coelacanthidé). Cybium, 2: 129–137.
JANVIER, P. 1996. Early Vertebrates. Clarendon Press, Oxford, 393 pp.
MEUNIER, F. J. 1979. Etude histologique et microradiographique du cartilage hémal de la vertèbre de la carpe, Cyprinus carpio L. (Pisces, Teleostei, Cyprinidae). Acta Zoologica, 60: 19–31.
MEUNIER, F. J. 1980. Les relations isopédine-tissu osseux dans le post-temporal et les écailles de la ligne latérale de Latimeria chalumnae (SMITH). Zoologica Scripta, 9: 307–317.
MEUNIER, F. J. and ZYLBERBERG, L. 1999. The structure of the external layer and of the odontodes of scales in Latimeria chalumnae (Sarcopterygii, Actinistia, Coelacanthidae) revisited using scanning and transmission electron microscopy. In: SÉRET, B. and SIRE, J.-Y. (eds.), Proc. 5th Indo-Pacific Fish Conf., Nouméa, 1997, Soc. Fr. Ichtyol., Paris, pp. 109–116.
MEUNIER, F. J., ERDMANN, M. V., FERMON, Y. and CALDWELL, R. L. 2008. Can the comparative study of the morphology and histology of the scales of Latimeria menadoensis and L. chalumnae (Sarcopterygii, Actinistia, Coelacanthidae) bring new insight on the taxonomy and the biogeography of recent coelacanthids? Geological Society, London, Special Publications, 295: 351–360.
MEUNIER, F. J., BRITO, P. M. and LEAL, M.-E. 2013. Quelques données morphologiques et histologiques sur la structure de la plaque dentaire linguale d’Arapaima gigas (Osteoglossidae; Teleostei). Cybium, 37: 263–271
MEUNIER, F. J., MONDEJAR-FERNANDEZ, J., GOUSSARD, F., CLÉMENT, G. and HERBIN, M. 2015. Presence of plicidentine in the oral teeth of Latimeria chalumnae (Sarcopterygii; Actinistia; Coelacanthidae). Journal of Structural Biology, 190: 31–37.
MILLER, W. A. 1979. Observations on the structure of mineralized tissue of the coelacanth, including scales and their associate odontodes. Occasional Papers of the Californian Academy of Sciences, 134: 68–78.
MILLER, W. A. and HOBDELL, M. H. 1968. Preliminary report on the histology of the dental and paradental tissues of Latimeria chalumnae (SMITH) with a note on tooth replacement. Archives of Oral Biology, 13: 1289–1291.
MILLOT, J. and ANTHONY, J. 1958. Anatomie de Latimeria chalumnae. T. 1. Squelette, muscles et formations de soutien. CNRS Edr., Paris. 122 pp., 80 pls.
MILLOT, J., ANTHONY, J. and ROBINEAU, D. 1978. Anatomie de Latimeria chalumnae. T. 3. Appareil digestif, téguments, écailles. CNRS Edr., Paris. pp. 7–28, 135–139.
MONDEJAR-FERNANDEZ, J. 2018. On cosmine: its origins, b io logy and impl ica t ions for sarcopterygian interrelationships. Cybium, 42: 41–65.
ØRVIG T. 1951. Histologic studies of Placoderms and fossil Elasmobranchs. 1: The endoskeleton, with remarks on the hard tissues of lower vertebrates in general. Arkiv för Zoologi, 2: 321–454, 8 pl.
ØRVIG T. 1968. The dermal skeleton; general considerations. In: ØRVIG T. (ed.), Current problems of lower vertebrate phylogeny. Proceedings of the fourth Nobel Symposium, Stockholm, 1967, J. Wiley, New-York. pp. 373–397.
PARMENTIER, E., COMPÈRE, P., CASADEWALL, M., FONTENELLE, N., CLOOTS, R. and HENRIST, C. 2008. The rocker bone: a new kind of mineralized tissue? Cell and Tissue Research, 334: 67–79.
PEGUETA, V. P. 1968. Enchondral ossification in Latimeria chalumnae SMITH. Dopovidi Akademii Nauk Ukrainia SSR., 7: 653–656 (in Russian; English abstr.).
POUYAUD, L., WIRJOATMODIO, S., RACHMATIKA, L., TJAKRAWIDJAJA, A., HADIATY, R. K. and HADIE, W. 1999. Une nouvelle espèce de coelacanthe, preuves génétiques et morphologiques. Comptes Rendus de l’Académie des Sciences, 322: 261–267.
ROMER, A.S. 1937. The braincase of the carboniferous crossopterygian Megalichthys nitidus. Bulletin of the Museum of Comparative Zoology, 824: 1–73.
ROMER, A.S. 1942. Cartilage as an embryonic adaptation. American Naturalist, 76: 394–404.
ROUX, G. H. 1942. The microscopic anatomy of the Latimeria scale. South African Journal of Medical Sciences, 7: 1–18.
SASAGAWA, I., ISHIYAMA, M. and KODERA, H. 1984. Fine structure of the pharyngeal teeth in the coelacanth fish (Latimeria chalumnae). In: FEARNHEAD, R. W. and SUGA, S. (eds.), Tooth enamel IV, Elsevier Science Publishers, Amsterdam, pp. 462–466.
SCHULTZE, H.-P. 1969. Die Faltenzähne der Rhipidistiden Crossopterygier, der Tetrapoden und der Actinopterygi-er-Gattung Lepisosteus nebst einer Beschreibung der Zahnstruktur von Onychodus (Struniiformer Crossoptery-gier). Palaeontologica Italica, New Series, 35(65): 63–137.
SCHULTZE, H.-P. 1970. Folded teeth and the monophyletic origin of Tetrapods. American Museum Novitates, 2408: 1–10.
SCHULTZE, H.-P. 2018. Hard tissues in fish evolution: history and current issues. Cybium, 42(1): 29–39.
SHELLIS, R. P. and POOLE, D. F. G. 1978. The structure of the dental hard tissues of the coelacanthid fish Latimeria chalumnae SMITH. Archives of Oral Biology, 23: 1105–1113.
SIRE, J.-Y. and HUYSSEUNE, A. 2003. Formation of dermal skeletal and dental tissues in fish: a comparative and evolutionary approach. Biological Review, 78: 219–249.
SMITH, J. L. B. 1939. A living fish of the Mesozoic type. Nature, 143: 455–456.
48
SMITH, J. L. B. 1940. A living coelacanth fish from South Africa. Transactions of the Royal Society of South Africa, 28: 1–106, 44 Pls.
SMITH, M. M. 1978. Enamel in the oral teeth of Latimeria chalumnae (Pisces: Actinistia): a scanning electron microscopic study. Journal of Zoology, 185: 355–369.
SMITH, M. M. 1979. Scanning electron microscopy of odontodes in the scales of coelacanth embryo, Latimeria chalumnae SMITH. Archives of Oral Biology, 24: 179–183.
SMITH, M. M., HOBDELL, M. H. and MILLER, W. A. 1972. The structure of the scales of Latimeria chalumnae. Journal of Zoology, London, 167: 501–509.
THOMSON, K. S. 1969. The biology of the lobe-finned fishes.
Biological Reviews, 44: 91–154. UYENO, T. and YABUMOTO, Y. 2007. Origin of extant
coelacanths. The Coelacanth, Fathom the Mystery 2007. Aquamarine Fukushima, pp. 24–26.
ZYLBERBERG, L. and MEUNIER, F. J. 2008. New data on the structure and the chondrocyte populations of the haemal cartilage of abdominal vertebrae in the adult carp Cyprinus carpio (Teleostei, Ostariophysii, Cyprinidae). Cybium, 32: 225–239.
ZYLBERBERG, L., GÉRAUDIE, J., MEUNIER, F. J., & SIRE J. Y., 1992. Biomineralization in the integumental skeleton of the living lower Vertebrates. In: HALL, B. K. (ed), Bone, Volume 4, CRC Press, Boca Raton, pp. 171–224.
François J. MEUNIER, Camila CUPELLO and Gaël CLÉMENT
View publication statsView publication stats







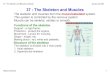



![UvA-DARE (Digital Academic Repository) The ... 1 Introduction ! 2! ... [7]; (E) the CaCO 3 skeleton microstructure ... Any living organism producing its mineralized tissue requires](https://img.dokumen.tips/doc/110x75/5aa952217f8b9a81188c9d2d/uva-dare-digital-academic-repository-the-1-introduction-2-7-e.jpg)