Embed Size (px)
Citation preview

Tt
Da
b
c
a
ARR2A
KGPPPU
1
mtgtbuawa
ltFstel
0d
Environmental and Experimental Botany 70 (2011) 251–258
Contents lists available at ScienceDirect
Environmental and Experimental Botany
journa l homepage: www.e lsev ier .com/ locate /envexpbot
he role of proline-associated pentose phosphate pathway in cool-seasonurfgrasses after UV-B exposure
ipayan Sarkara,b, Prasanta C. Bhowmika, Young-In-Kwonb,c, Kalidas Shettyb,∗
Department of Plant, Soil, and Insect Sciences, Stockbridge Hall, University of Massachusetts, Amherst, MA 01003, USADepartment of Food Sciences, Chenoweth Lab, University of Massachusetts, Amherst, MA 01003, USADepartment of Food and Nutrition, Hannam University, Daejeon 305811, South Korea
r t i c l e i n f o
rticle history:eceived 25 February 2010eceived in revised form3 September 2010ccepted 28 September 2010
a b s t r a c t
Physiological adjustments of cool-season turfgrasses were investigated to determine the role of proline-associated pentose phosphate pathway for phenolic biosynthesis and stimulation of antioxidant responsesystem following UV-B exposure. Creeping bentgrass, Kentucky bluegrass, tall fescue, and perennial rye-grass plugs were subjected to artificial UV-B exposure (biologically effective UVBBE radiation 8 kJ m−2 d−1.)for one week with 10-h photoperiod. Significant accumulation of phenolics and stimulation of antioxi-
eywords:lucose-6-phosphate dehydrogenaseentose phosphate pathwayhenolicrolineV-B stress
dant enzyme activity was observed in all investigated cool-season turfgrasses after UV-B exposure andthis induction corroborated with higher glucose-6-phosphate dehydrogenase activity and high accumu-lation of proline. Guaiacol peroxidase activity also increased in all investigated cool-season turfgrassspecies after UV-B exposure. In this study, the shift of carbon flux from glycolysis to pentose phosphatepathway following UV-B exposure and as a result of that, the higher accumulation of phenolics and stim-ulation of antioxidant response system provides an insight to understand a probable defense mechanismof cool-season turfgrasses against UV stress.
© 2010 Elsevier B.V. All rights reserved.
. Introduction
The depletion of stratospheric ozone concentration due to man-ade ozone-destroying pollutants (chlorofluorocarbon) is leading
o an increase in ultraviolet-B radiation (UV-B 290–320 nm) atround level (Shindell et al., 1998). It is estimated that ozone inhe stratosphere will continue to fall, with lowest levels projectedetween 2010 and 2019 in the northern hemisphere that may resultp to 50–60% increase in UV-B radiation in springtime (Taalas etl., 2000). This increase is likely to continue into foreseeable futureith potentially deleterious consequence for agriculture, forestry
nd horticulture (Jordan et al., 1994).Ultraviolet-B radiation either alone or in combination with
ow soil moisture or high temperature causes severe damageo turfgrass (Ervin et al., 2004a; Schmidt and Zhang, 2001).
requently, increased heating from respiratory function duringtorage, transport, and subsequent exposure to high UV-B radia-ion during transplanting can cause shock to turfgrass sod (Gieset al., 1997). The shock often results in bleached, inactive turfgrasseaves.∗ Corresponding author. Tel.: +1 413 545 1022; fax: +1 413 545 1262.E-mail address: [email protected] (K. Shetty).
098-8472/$ – see front matter © 2010 Elsevier B.V. All rights reserved.oi:10.1016/j.envexpbot.2010.09.018
Turfgrasses are also exposed to UV-B radiation during variousstages of growth, under present environmental conditions. Plantsbetter adapted to UV-B induced photo-bleaching usually containmore robust screening (pigment) and scavenging (antioxidant) pro-tection systems (Mackerness, 2000). Schmidt and Zhang (2001)found significant reduction of photochemical activity (19.2%), andchlorophyll content (18.5%) with UV radiation in creeping bentgrass(Agrostis stolonifera L.). The reduction of visual quality and photo-chemical efficiency of Kentucky bluegrass (Poa pratensis L.) plugand sod was also observed by Ervin et al. (2004a,b,c). Resistance toUV-B damage of turfgrass most likely involves both avoidance andtolerance mechanisms.
UV-B radiation can induce a wide range of responses in plants,including increased concentrations of protective UV-B-absorbingpigments in leaves and reductions in rates of CO2 assimilation andplant growth (Kostina et al., 2001; Teramura and Sullivan, 1994).UV-B exposure is believed to enhance the level of reactive oxygenspecies (ROS), although the mechanism of their generation is notknown (Rao et al., 1996). Plant metabolizes ROS through antioxi-
dant defense system. This defense system consists of low molecularweight antioxidants such as ascorbate, glutathione, �-tocopherol,and carotenoids, as well as several enzymes such as superoxide dis-mutase (SOD), catalase (CAT), glutathione reductase, and ascorbateperoxidase (Alscher and Hess, 1993; Bowler et al., 1994).
2 Exper
Tcabanicpsp1pap1
ap1d(fprpprtWirfcpsp
tutlfcsProecdltat(ptctcvsgu
52 D. Sarkar et al. / Environmental and
Role of phenolics as antioxidant has been well documented.he structural chemistry of polyphenols suggests their free radi-al scavenging properties (Rice-Evans et al., 1997). The activity ofntioxidant is determined by its reduction potential, ability to sta-ilize and delocalize the unpaired electron, reactivity with otherntioxidants, and transition metal chelating potential. Polyphe-ols have all the above characters and thus play significant role
n antioxidative defense systems in plants. Many phenylpropanoidompounds such as flavonoids, isoflavonoids, anthocyanins andolyphenols are induced in response to wounding, nutritionaltress, cold stress, and high visible light. UV radiation also induceshenolic antioxidant in plants (Christie et al., 1994; Graham,991; Hahlbrock and Scheel, 1989). Various phenolic compounds,articularly flavonoids, may counter UV radiation (Dhariwal etl., 1998). Antioxidants, like carotenoides and polyphenol com-ounds may also protect plants from UV-B damage (Borman et al.,997).
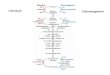
Stimulation of protective secondary metabolite pathways, suchs biosynthesis of phenolic antioxidants is a natural response oflant to biotic and abiotic stresses (Rice-Evans et al., 1997; Shetty,997, 1999, 2004). Induction of phenylpropanoid pathway and pro-uction of proline in plants during abiotic stress is well documentedChristie et al., 1994; Dorffling et al., 1997). Proline is synthesizedrom glutamate by a series of reduction reaction. In this synthesisrocess, proline and pyrroline-5-carboxylate (P5C) may regulateedox and hydride ion-mediated stimulation of pentose phosphateathway (Hagedorn and Phang, 1983; Phang, 1985). During res-iration, oxidation reactions produce hydride ions, which helpeduction of P5C to proline into the cytosol. Through the reac-ion of proline dehydrogenase, proline can enter the mitochondria.
ithin the mitochondria, instead of NADH, proline acts as a reduc-ng equivalent and can support oxidative phosphorylation. Theeduction of P5C in cytosol provides NADP+, which is a co-factoror glucose-6-phosphate dehydrogenase (G6PDH). G6PDH playsrucial role, by catalyzing first rate limiting step of the pentosehosphate pathway. Phang (1985) first proposed this model andtated its role in stimulation of purine metabolism via ribose-5-hosphate in animal cells.
On the basis of this hypothesis, Shetty (1997) proposed a modelhat proline-associated pentose phosphate pathway could stim-late both the shikimate and phenylpropanoid pathways, andherefore, the modulation of this pathway could lead to the stimu-ation of phenolic phytochemicals. This model has been proposedor the mode of action of phenolic metabolites based on theorrelation between stress stimulated phenolic biosynthesis andtimulation of antioxidant enzyme response pathways in plants.henolic antioxidants can behave as antioxidants by trapping freeadicals in direct interactions or scavenge them through a seriesf coupled, antioxidant enzyme defense system reactions (Bowlert al., 1994; Pinhero et al., 1997; Rao et al., 1996). It is clear fromurrent theories that antioxidant response pathway in plants isependent upon key NADPH-requiring enzymes, similar to pro-
ine biosynthesis. This defensive mechanism are believed to workhrough antioxidant response pathways involving peroxidases,s well as biosynthesis of polymeric phenolics that lead to pro-ective lignification of smaller polymers that act as antioxidantsShetty, 2004). We hypothesized that proline-associated pentosehosphate pathway may play a crucial role under UV-B stress inurfgrass species and as a result of this; phenolic phytochemicalsan effectively counter oxidative stress within cells. In earlier inves-igations, turfgrasses under UV-B stress showed higher antioxidant
oncentrations in leaf tissues (Ervin et al., 2004a,b,c). But there isery limited understanding regarding physiological mechanisms ofuch behavior in cool-season turfgrasses. So, our overall researchoal was to evaluate whether UV-B exposure could cause anpregulation of the pentose phosphate pathway and stimulationimental Botany 70 (2011) 251–258
of phenolic-linked antioxidant enzyme response in cool-seasonturfgrasses.
2. Materials and methods
2.1. Plant material and growth conditions
Creeping bentgrass (‘Penncross’), perennial ryegrass (Loliumperenne L.) (‘Palmer II’), Kentucky bluegrass (‘Baron’), and tall fes-cue (Festuca arundianacea Schreb.) (‘Kentucky 31’) plugs (2.5 cm)were collected from Joseph Troll Turf Research Center, Universityof Massachusetts, South Deerfield. All turfgrass plugs were pottedin 4.5 cm diameter 6 cm tall pots with soil-sand mixture. All plantswere initially grown in growth chamber with controlled tem-perature (20 ◦C) and photoperiod (10-h) under fluorescent whitelight (300 �mol m−2 s−1; 400–700 nm). Plants were fertilized andmowed once a week for the duration of study.
2.2. UV-B treatment
After maintaining in proper growth condition under growthchamber for two months, turfgrass plugs from each specieswere then transferred into another chamber that supplied anequal amount of photosynthetically active radiation supple-mented by radiation from UV-B tube (two 30 W fluorescent tubes,70 �mol m−2 s−1, UVB-313, Cleveland, Ohio) for 1 week. UV-B 313lamp did not produce any additional photon flux in the 400–700 nm(PAR) range. The pots were placed evenly (3.0 cm), kept 0.6 m belowthe UV-B source. The tubes were first preburned 24 h to stabilizethe UVB output. The level of biologically effective UVB (UVBBE)radiation was 8 kJ m−2 d−1. The UVBBE radiation was measuredby using an IL1700 Research Radiometer with calibrated photo-detector/filters (International Light, Newburyport, MA, USA). TheUVBBE was estimated via Caldwell (1971) generalized plant damageaction spectrum, normalized to 300 nm. Plants were exposed with10-h photoperiod. Control was carried out without UV-B exposurefor each species under fluorescent white light (300 �mol m−2 s−1,400–700 nm, Philips lighting, Somerset, NJ). All treatments werereplicated 5 times (5 pots from each species) and entire experimentrepeated two times. Shoot samples were collected for biochemicalanalysis.
2.3. Enzyme extraction
Turfgrass leaf tissue (200 mg) was collected and thoroughlymacerated by using a cold pestle and mortar with cold enzymeextraction buffer [0.5% polyvinylpyrrolidone (PVP), 3 mmol L−1
EDTA, and 0.1 mol L−1 potassium phosphate buffer of pH 7.5]. Theextracted sample was centrifuged at 10,188 gn for 10 min at 2–5 ◦Cand stored in ice. Supernatant was used for further chemical anal-ysis.
2.4. Glucose-6-phophate dehydrogenase (G6PDH) assay
A modified method originally described by Deutsch (1983) wasused. The enzyme reaction mixture containing 5.88 �mol L−1 ß-NADP, 88.5 �mol L−1 magnesium chloride (MgCl2), 53.7 �mol L−1
glucose-6-phosphate, and 0.77 mmol L−1 maleimide was prepared.This mixture was used to obtain baseline (zero) of the spectropho-tometer reading at 340 nm wavelength. To 1 mL of this mixture,
100 �L of the extracted enzyme sample was added. The rate ofchange in absorbance per minute was used to quantify the enzymeactivity in the mixture with the help of the extinction co-efficientof NADPH (6.22 mmol L−1 cm−1) and expressed as nanomoles permg protein.
Exper
2
wt(nrT33ip
2
(psopstoe(
2
ept9Treo
2
ahCptass2iCH
2
xloatc(
D. Sarkar et al. / Environmental and
.5. Proline dehydrogenase (PDH) assay
A modified method described by Costilow and Cooper (1978)as carried out to assay the activity of PDH. The enzyme reac-
ion mixture containing 100 mmol L−1 sodium carbonate bufferpH 10.3), 20 mmol L−1 L-proline solution and 10 mmol L−1 mMicotinamide adenine dinucleotide (NAD) was used. To 1 mL of thiseaction mixture, 200 �L of extracted enzyme sample was added.he increase in absorbance was measured at 340 nm for 3 min, at2 ◦C. The absorbance was recorded at zero time and then aftermin. In this spectrophotometric assay, one unit of enzyme activ-
ty is equal to the amount causing an increase in absorbance of 0.01er min at 340 nm (1.0 cm light path).
.6. Succinate dehydrogenase (SDH) assay
To assay the activity of SDH a modified method of Bregman1987) was used. The assay mixture containing 1.0 mL of 0.4 mol L−1
otassium phosphate buffer (pH 7.2), 40 �L of 0.15 mol L−1 sodiumuccinate (pH 7.0), 40 �L of 0.2 mol L−1 sodium azide, and 10 �Lf 6.0 mg mL−1 2,6-dichlorophenolindophenol (DCPIP) was pre-ared. This mixture was used to obtain baseline (zero) of thepectrophotometer reading at 600 nm wavelength. To 1.0 mL ofhis mixture, 200 �L of the enzyme sample was added. The ratef change of absorbance per minute was used to quantify thenzyme in the mixture using the extinction co-efficient of DCPIP19.1 mmol L−1 cm−1).
.7. Guaiacol peroxidase (GPX) assay
The activity of GPX was assayed by a modified method (Lalouet al., 1997). The enzyme reaction mixture containing 0.1 mol L−1
otassium phosphate buffer (pH 6.8), 56 mmol L−1 guaiacol solu-ion, and 50 mmol L−1 hydrogen peroxide (H2O2) was used. To90 �L of this reaction mixture, 10 �L of enzyme sample was added.he absorbance was recorded at zero time and then after 5 min. Theate of change in absorbance per minute was used to quantify thenzyme in the mixture by using the extinction coefficient of thexidized product tetraguaiacol (26.6 mmol L−1 cm1).
.8. Catalase (CAT) assay
The CAT activity was assayed according to the methods of Beersnd Sizer (1952). To 1.9 mL of distilled water 1 mL of 0.059 mol L−1
ydrogen peroxide (Merck’s Superoxol or equivalent grade, Mercko. & Inc., Whitehouse Station, NJ) in 0.05 mol L−1 potassiumhosphate, pH 7.0 was added. This mixture was incubated in a spec-rophotometer for 4–5 min to achieve temperature equilibrationnd to establish blank rate. To this mixture 0.1 mL of diluted enzymeample was added and the disappearance of peroxide was followedpectrophotometrically by recording the decrease in absorbance at40 nm for 2–3 min. The change in absorbance �A240/min from the
nitial (45 s) linear portion of the curve was calculated. One unit ofAT activity was defined as amount that decomposes 1 �mol of2O2.
.9. Superoxide dismutase (SOD) assay
A competitive inhibition assay was performed that usedanthine-xanthine oxidase generated superoxide to reduce nitrob-ue tetrazolium (NBT) to blue formazan. Spectrophotometric assay
f SOD activity was carried out by monitoring the reduction of NBTt 560 nm (Oberley and Spitz, 1984). The reaction mixture con-ained 13.8 mL of 50 mmol L−1 potassium phosphate buffer (pH 7.8)ontaining 1.33 mmol L−1 diethylenetetraaminepentaacetic acidDETAPAC); 0.5 mL of 2.45 mmol L−1 NBT; 1.7 mL of 1.8 mmol L−1imental Botany 70 (2011) 251–258 253
xanthine and 40 IU mL−1 catalase. To 0.8 mL of reagent mixture100 �L of phosphate buffer and 100 �L of xanthine oxidase wasadded. The change in absorbance at 560 nm was measured every20 s for 2 min and the concentration of xanthine oxidase wasadjusted to obtain a linear curve with a slope of 0.025 absorbanceper min. The phosphate buffer was then replaced by the enzymesample and the change in absorbance was monitored every 20 s for2 min. One unit of SOD was defined as the amount of protein thatinhibits NBT reduction to 50% of the maximum.
2.10. Total protein assay
Protein content was determined by the method of Bradfordassay (Bradford, 1976). One parts of dye reagent (Bio-Rad proteinassay kit II, Bio-Rad Laboratory, Hercules, CA) was diluted with 4parts of distilled water. A volume of 5 mL of diluted dye reagentwas added to 50 �L of the turfgrass tissue extract. After vortexingand incubating for 5 min, the absorbance was measured at 595 nmagainst a blank (5 mL reagent and 50 �L buffer solution) by usinga UV–VIS Genesys spectrophotometer (Milton Roy, Inc., Rochester,NY).
2.11. ABTS [2,2′-azino-bis(3-ethylbenzthiazoline-6-sulphonicacid)] cation radical and antioxidant activity assay
The total antioxidant activity of turfgrass leaf extract was mea-sured by the ABTS+ radical cation-decolorization assay involvingpreformed ABTS+ radical cation (Pellegrini et al., 1999). ABTS (SigmaChemical Co, St. Louis, MO) was dissolved in water to a 7 mmol L−1
concentration. ABTS+ radical cation was prepared by reacting 5 mLof 7 mmol L−1 ABTS stock solution with 88 �L of 140 mmol L−1
potassium persulphate, and mixture was allowed to stand in thedark at room temperature for 12–16 h before use. Prior to assayABTS+ stock solution was diluted with 95% ethanol (ratio 1:88) togive an absorbance at 734 nm of 0.70 ± 0.02, and was equilibratedto 30 ◦C. One millilitre ABTS was added to glass test tubes contain-ing 50 �L of each tissue extract, and mixed by vortex mixer for30 s. After 2.5 min incubation, mixtures were read at 734 nm. Thereadings were compared with controls, which contained 50 �L of95% ethanol instead of the extract. The Trolox reference standardfor relative antioxidant activities was prepared with 5 mmol L−1
stock solution of Trolox in ethanol for introduction into the assaysystem at concentrations within the activity range of the assay(0–20 �mol L−1 final concentration) for preparing a standard curveto which all data were referenced. The percent inhibition was cal-culated by:
% inhibition = [Acontrol734 − Aextract
734 ])
[Acontrol734 ]
× 100
2.12. Total soluble phenolic assay
The total soluble phenolics were determined by an assay modi-fied from Shetty et al. (1995). A quantity of 50 mg (fresh weight-FW)turfgrass leaf tissue was immersed in 2.5 mL of 95% ethanol andkept in the freezer for 48-h. After 48-h, the sample was homoge-nized and centrifuged at 12,225 gn for 10 min. Then 0.5 mL of samplesupernatant was diluted with 0.5 mL of distilled water and trans-ferred into a test tube with 1 mL of 95% ethanol and 5 mL of distilledwater. In each sample, 0.5 mL of 50% (v/v) Folin-Ciocalteu reagent
was added and mixed. After 5 min, 1 mL of 5% Na2CO3 was added tothe reaction mixture and allowed to stand for 60 min. A blank wasprepared with 0.5 mL distilled water instead of sample. Absorbancewas noted at 725 nm after 1-h. The absorbance values were con-verted to total phenolics and were expressed in mg equivalents of
2 Experimental Botany 70 (2011) 251–258
gu
2
pTtCTm2atbr
2
bM(fl2
2
ww8ndp
3
3at
ihigctsc
TMg(
N
54 D. Sarkar et al. / Environmental and
allic acid per g FW of the sample. Standard curves were establishedsing various concentrations of gallic acid in 95% ethanol.
.13. HPLC analysis of proline
High performance liquid chromatography (HPLC) analysis waserformed using an Agilent 1100 liquid chromatograph (Agilentechnologies, Santa Clara, CA) equipped with a diode array detec-or (DAD 1100). The analytical column was reverse phase Nucleosil18, 250 nm × 4.6 mm with a packing material of 5 �m particle size.he extract samples were eluted out in an isocratic manner with aobile phase consisting of 20 mmol L−1 potassium phosphate (pH
.5 by phosphoric acid) at a flow rate of 1 mL/min and detectedt 210 nm. l-Proline (Sigma chemicals, St. Louis, MO) dissolved inhe 20 mmol L−1 potassium phosphate solution was used to cali-rate the standard curve. The amount of proline in the sample waseported as mg of proline per g FW.
.14. Photochemical efficiency
Photochemical efficiency of turfgrass shoots was measuredy using a fluorometer (OS1-FL; Opti-Sciences, Tyngsboro,A). The test was carried in dark adapted mode and Fv/Fm
Fv/Fm = [Fm − Fo]/Fm the ratio of variable fluorescence to maximaluorescence) ratio was calculated. Plants were kept in dark at least-h before the measurement.
.15. Statistical analysis
Experiments were carried out in completely randomized designith five replications. The effect of UV-B exposure on turfgrassesas determined by the analysis of variance (ANOVA) of SAS (version
.2; SAS Institute, Cary, NC). Differences among UV-B treated andon treated turfgrass species at the end of UV-B exposure wereetermined by the least significant difference (LSD) test at the 0.05robability level.
. Results
.1. Changes in total soluble phenolics and free radical-linkedntioxidant activity of cool season turfgrasses following UVreatment
A significant increase in total phenolic content was observedn all UV-B exposed cool-season turfgrasses (Fig. 1) (Table 1). Theigher increment of total soluble phenolic was found in UV-B
nduced creeping bentgrass (152%), followed by Kentucky blue-
rass (122%), tall fescue (100%), and perennial ryegrass (41%) overontrol of the respective species. Similar trend was observed inhe total free radical scavenging-linked antioxidant activity of cool-eason turfgrasses after one week of UV-B treatment. Exposure ofool-season turfgrass plants to UV-B radiation significantly stim-able 1ean squares (MS) from ANOVA of total soluble phenolics (TSP), total antioxidant activity
enase activity (SDH), total proline content, proline dehydrogenase (PDH) activity, superoGPX), and photochemical efficiency (PE) as influenced by cool-season turfgrass species (
ANOVA TSP ABTS G6PDH SDH
Source df
S 3 NS NS * NSUV 1 ** * * *
S × UV 3 NS NS NS NS
S: Nonsignificant.* Significant at P ≤ 0.05.
** Significant at P ≤ 0.01.
Fig. 1. Total soluble phenolics (mg g−1 F W) in UV-B (1 week) treated and untreatedcreeping bentgrass (CB), Kentucky bluegrass (KB), tall fescue (TF) and perennial rye-grass (PR). Vertical bars are LSD values (P ≤ 0.05) for UV-B treated and untreatedspecies comparisons in four cool-season turfgrasses.
ulated the total antioxidant activity (ABTS assay) (Fig. 2). Higherantioxidant activity was observed in UV-B induced Kentucky blue-grass (49%), followed by tall fescue (47%), creeping bentgrass (47%),and perennial ryegrass (43%). But higher percentage of stimulationof total antioxidant activity due to UV-B exposure was found increeping bentgrass (17% increase) as compared to control.
3.2. Glucose-6-phosphate dehydrogenase (G6PDH) and succinatedehydrogenase (SDH) activity
Significantly higher G6PDH activity was observed in all UV-Binduced turfgrass plants compared to control (Fig. 3) (Table 1).The stimulation of G6PDH activity was more prominent in creep-ing bentgrass (more than double), whereas the magnitude of thisstimulation was lower in case of tall fescue. Results indicated thatactivity of succinate dehydrogenase reduced significantly in all UV-B induced turfgrass plants as compared to control (Fig. 4) (Table 1).The higher reduction was observed in creeping bentgrass plantsafter one-week exposure to UV-B radiation.
3.3. Changes in proline content and proline dehydrogenase (PDH)activity
Exposure to UV-B radiation significantly enhanced the total pro-line content in all cool-season turfgrass species (Fig. 5) (Table 1).Total proline content was significantly higher in UV-B inducedcreeping bentgrass, followed by perennial ryegrass, Kentucky blue-
grass, and tall fescue as compared to their respective control.Proline dehydrogenase activity also increased in cool-season tur-fgrasses after UV-B exposure (Fig. 6) (Table 1). The stimulation ofPDH activity was higher in Kentucky bluegrass when compared toother cool-season turfgrass species in this study.(ABTS), glucose-6-phosphate dehydrogenase activity (G6PDH), succinate dehydro-xide dismutase activity (SOD), catalase activity (CAT), Guaiacol peroxidase activity
S) and UV-B treatment (UV).
PROLINE PDH SOD CAT GPX PE
* * NS NS NS NS** * NS NS ** *
NS NS NS NS NS NS

D. Sarkar et al. / Environmental and Experimental Botany 70 (2011) 251–258 255
Fig. 2. Total antioxidant activity (%) (ABTS assay) in UV-B (1 week) treated anduntreated creeping bentgrass (CB), Kentucky bluegrass (KB), tall fescue (TF) andperennial ryegrass (PR). Vertical bars are LSD values (P ≤ 0.05) for UV-B treated anduntreated species comparisons in four cool-season turfgrasses.
Fig. 3. Glucose-6-phosphate dehydrogenase (G6PDH) (nmol mg−1 protein) activityin UV-B (1 week) treated and untreated creeping bentgrass (CB), Kentucky blue-grass (KB), tall fescue (TF) and perennial ryegrass (PR). Vertical bars are LSD values(P ≤ 0.05) for UV-B treated and untreated species comparisons in four cool-seasonturfgrasses.
Fig. 4. Succinate dehydrogenase (SDH) (nmol mg−1 protein) activity in UV-B (1week) treated and untreated creeping bentgrass (CB), Kentucky bluegrass (KB), tallfescue (TF) and perennial ryegrass (PR). Vertical bars are LSD values (P ≤ 0.05) forUV-B treated and untreated species comparisons in four cool-season turfgrasses.
−1
Fig. 5. Total proline content (mg g F W) in UV-B (1 week) treated and untreatedcreeping bentgrass (CB), Kentucky bluegrass (KB), tall fescue (TF) and perennial rye-grass (PR). Vertical bars are LSD values (P ≤ 0.05) for UV-B treated and untreatedspecies comparisons in four cool-season turfgrasses.3.4. Changes in superoxide dismutase (SOD), catalase (CAT) andguaiacol peroxidase (GPX) activity
The activity of SOD changed differently among the cool-seasonturfgrasses after UV treatment. The SOD activity slightly increasedin Kentucky bluegrass, and in tall fescue after exposure of theseplants to UV-B radiation, but it decreased in case of creepingbentgrass, and in perennial ryegrass (Table 2). The CAT activ-ity of cool-season turfgrasses also showed similar inconsistenttrend after UV treatment. Activity of this enzyme decreased increeping bentgrass after UV-B induction, whereas same treatmentcaused slight stimulation of CAT activity in other three cool-seasonturfgrass species (tall fescue, perennial ryegrass, and Kentuckybluegrass) (Table 2). Different turfgrass species did not show anysignificant differences in SOD and CAT activity (Table 1) after UV-B exposure, but it showed interesting overall metabolic trends inall turfgrass species to counter oxidative stress and maintainingcellular homeostasis. Significant stimulation of GPX activity wasobserved in all cool-season turfgrass plants after UV treatment.The higher stimulation of GPX activity was found in creeping bent-
grass compared to other turfgrass species (Table 2). This higher GPXactivity was directly proportional to increased G6PDH activity andinduction of phenolic biosynthesis in turfgrass plants after theirUV-B exposure.Fig. 6. Proline dehydrogenase (PDH) (Unit mg−1 protein) activity of UV-B (1 week)treated and untreated creeping bentgrass (CB), Kentucky bluegrass (KB), tall fescue(TF) and perennial ryegrass (PR). Vertical bars are LSD values (P ≤ 0.05) for UV-Btreated and untreated species comparisons in four cool-season turfgrasses.

256 D. Sarkar et al. / Environmental and Experimental Botany 70 (2011) 251–258
Table 2Superoxide dismutase (SOD) (units mg−1 protein), catalase (CAT) (units mg−1 protein) and guaiacol peroxidase (GPX) (nmol mg−1 protein) activity of creeping bentgrass (CB),Kentucky bluegrass (KB), tall fescue (TF) and perennial ryegrass (PR) after one week of UV-B treatment along with controls.
Treatments SOD (units mg−1 protein) CAT (units mg−1 protein) GPX (nmol mg−1 protein)
Creeping BentgrassUV-B treated 9.34 cda 52.43 d 58.64 aControl 10.64 bc 65.53 c 20.33 c
Kentucky bluegrassUV-B treated 12.45 a 72.34 b 32.46 bControl 11.86 ab 68.94 bc 13.54 d
Tall fescueUV-B treated 10.85 ab 76.32 a 30.54 bControl 10.65 bc 70.56 b 18.65 cd
Perennial ryegrassUV-B treated 7.65 d 70.87 b 28.54 bControl 8.94 cd 63.54 c 14.45 d
Tnt at P
3
wderoTft
4
oeitUaUrcople
Fcgs
his test was preceded by an ANOVA and each mean was based on five replicates.a Values followed by same letter within each column are not significantly differe
.5. Photochemical efficiency
The photochemical efficiency of cool-season turfgrass plantsere measured by using chlorophyll fluorescence technique andata represented as Fv/Fm ratio to understand the photochemicalfficiency of these turfgrass species after their exposure to UV-Badiation. After one week of UV treatment a significant reductionf Fv/Fm ratio was observed in all turfgrass plants (Fig. 7) (Table 1).he reduction of photochemical efficiency and discoloration of tur-grass leaves were more profound in creeping bentgrass, whereashe change of this quality parameter is less prominent in tall fescue.
. Discussion
Significant accumulation of total soluble phenolics wasbserved in all investigated cool-season turfgrasses after UV-Bxposure. Accumulation of soluble phenolics during UV-B stressn these cool-season turfgrasses suggests the probable multifunc-ional roles of phenolics in plant cells. Phenolics can either absorbV-B radiation in leaves (Smith and Markham, 1998) or it can acts antioxidant by scavenging reactive oxygen species induced byV-B radiation (Natori and Ueno, 1987). It may also stimulate the
esponse of antioxidant enzymes such as superoxide dismutase,atalase and guaiacol peroxidase to scavenge peroxide or other
xygen radicals during UV-B stress. This UV-B radiation inducedhenolic biosynthesis in cool-season turfgrasses support the ear-ier findings of phenolic stimulation in other plant species (Lavolat al., 1997; Rosa et al., 2001).
ig. 7. Photochemical efficiency (Fv/Fm ratio) in UV-B (1 week) treated and untreatedreeping bentgrass (CB), Kentucky bluegrass (KB), tall fescue (TF) and perennial rye-rass (PR). Vertical bars are LSD values (P ≤ 0.05) for UV-B treated and untreatedpecies comparisons in four cool-season turfgrasses.
= 0.05.
Stimulation of total antioxidant activity due to UV-B radiationwas observed in all of these investigated turfgrass species. Higherinduction of total soluble phenolics and total antioxidant activity increeping bentgrass and in Kentucky bluegrass after UV-B exposureprovide an insight to understand the possible biochemical mecha-nism of these species against UV-B stress.
The other interesting finding of this study is the high glucose-6-phosphate dehydrogenase (G6PDH) activity of cool seasonturfgrasses following UV-B exposure. G6PDH is the first rate lim-iting enzyme of pentose phosphate pathway (PPP), and higheractivity of this enzyme reflects the higher role of pentose phos-phate pathway in the cool-season turfgrasses during UV-B stress.Higher G6PDH activity of UV-B induced creeping bentgrass alsocorroborates with its phenolic content and antioxidant activitywhich emphasizes the role of pentose phosphate pathway to sup-port anabolic need of this turfgrass species under UV-B stress. Onthe contrary, succinate dehydrogenase (SDH), a key enzyme of tri-carboxylic cycle that provides NADH for mitochondrial oxidativephosphorylation was reduced during UV-B treatment in all investi-gated turfgrass species. It reflects that carbon flux may be divertedfrom catabolic tricaroxylic acid cycle to anabolic pentose phos-phate pathway in the cool-season turfgrasses to deal with UV-Binduced oxidative stress. The pathway shift was more prominentwith creeping bentgrass (high baseline SDH) and with perennialryegrass (low baseline SDH).
Analysis of proline content also supports the above findings, ashigh proline accumulation of UV-B induced cool-season turfgrassesalso indicate that carbon flux diverted from TCA cycle to prolinesynthesis from glutamate through �-ketoglutarate. High prolinecontent in these turfgrasses also reflects the role of this impor-tant amino acid in the cellular system during UV stress. HigherG6PDH activity along with higher proline content in these cool-season turfgrasses during UV treatment may suggest a probablecoupling of proline synthesis with pentose phosphate pathwayin the cytosol under stress induced condition. Proline dehydro-genase (PDH) activity also increased after UV-B exposure in allinvestigated turfgrass species. The activity of this enzyme washigher in case of Kentucky bluegrass, which indicates a probableproline oxidation in Kentucky bluegrass to support mitochon-drial oxidative phosphorylation. This metabolic role of prolineis not clear in other investigated turfgrass species after UV-Bexposure.
Antioxidant enzymes activity after UV-B treatment showedinconsistent results among these turfgrass species. The activity ofsuperoxide dismutase (SOD) either reduced or almost unchanged
in all these turfgrass species after UV-B exposure. Suppression ofSOD due to UV treatment was also observed in other plant speciesin earlier investigations (Murphy and Huerta, 1990; Willekens etal., 1994). The activity of other important enzyme, catalase also
Exper
siwloslbAsscemasitiwSsLobdapltc
ammU(wIistwtwpsit
5
atrtsdTdAtsc
D. Sarkar et al. / Environmental and
howed inconsistent results in these turfgrass species. The activ-ty of this enzyme reduced significantly in creeping bentgrass,
hereas it increased in tall fescue and in perennial ryegrass fol-owing UV-B exposure. This indicates a species specific responsef these antioxidant enzymes in cool-season turfgrasses during UVtress. Polle (1997) suggested that higher UV-B doses may stimu-ate pathogenesis-related defense pathways in plants, presumablyy suppression of key defense enzymes such as SOD or catalase.lthough interactions and responses of individual species to UV-Btress related to SOD and CAT were not significant (Table 1), but ithowed interesting overall metabolic trends (Table 2) to maintainellular redox environment among four turfgrass species after UV-Bxposure. This information is needed to study metabolic adjust-ents of turfgrass species in more details at individual species
nd at individual cultivar levels for further understanding andtudies. On the contrary to SOD and CAT responses, activity of gua-acol peroxidase (GPX) showed different trend in these cool-seasonurfgrasses after UV-B treatment. The GPX activity significantlyncreased after UV treatment in all investigated turfgrass species,
here highest activity was observed in case of creeping bentgrass.timulation of peroxidase activity was also observed in Arabidop-is thaliana (Rao et al., 1996) and in sunflower (Helianthus anus.) (Costa et al., 2002) after UV-B exposure. Rao et al. (1996) alsobserved that UV-B radiation enhanced activated oxygen speciesy increasing membrane-localized NADPH oxidase activity andecreased catalase activity in Arabidopsis thaliana. As high GPXctivity corroborates with higher phenolic content, it indicates theossible cross linking of phenolic moieties for the biosynthesis of
ignins and lignans in the plant cell wall. Our findings indicate thathese types of peroxidases probably play a multipurpose role inool-season turfgrasses during UV-B stress.
Another important response of plants to UV-B radiation is dam-ge of photosynthetic apparatus. The disruption of chloroplastembranes by UV-B radiation was observed in soybean (Glycineax L.) (Campbell, 1975), and in Rumex patientia (Campbell, 1975).V-B radiation was suggested as an inhibitor of photosystem-II
PSII), or some specific step in the electron transport associatedith PSII apart from the resultant effects of chloroplast disruption.
wanzik et al. (1983) showed that the impairment of PSII activ-ty by UV-B radiation is due to blockage of PSII reaction centers inpinach (Spinacea oleracea L.). In our study, the maximum quan-um efficiency of PSII photochemistry of cool-season turfgrassesas measured by estimating Fv/Fm ratio. The significant reduc-
ion of Fv/Fm ratio was observed in all turfgrass plants after oneeek UV-B exposure. This low Fv/Fm ratio suggests an induction ofhotoinhibition in turfgrass leaves after UV-B radiation. As photo-ynthetic activity after UV treatment was highest in tall fescue, itndicates a probable avoidance of UV stress by this species throughhe reduction of photoinhibition.
. Conclusions
In summary, all these investigated turfgrass species have shownsignificant role of phenolics and antioxidants response sys-
em during UV-B stress. To counter oxidative stress due to UV-Badiation, these turfgrass species may adapt an energy efficient sys-em through proline-associated pentose phosphate pathway whichupport needs of anabolic pathways and also support mitochon-rial oxidative phosphorylation by helping energy (ATP) synthesis.his study can provide further clues to understand a probable
efense mechanism of cool-season turfgrasses during UV-B stress.dditional investigation with different cultivars of cool-seasonufgrasses and examination of different exogenous antioxidanttimulators would provide a detailed understanding of UV-B stressountering mechanism of cool-season turfgrasses.
imental Botany 70 (2011) 251–258 257
References
Alscher, R.G., Hess, J.L., 1993. Antioxidants in Higher Plants. CRC Press, Boca Raton,Florida.
Beers, R.F., Sizer, I.W., 1952. A spectrometric method for measuring the break downof the hydrogen peroxide by catalase. J. Biol. Chem. 195, 133–140.
Borman, J.F., Reuber, S., Weissenbock, G., Cen, Y.-P., 1997. Ultraviolet radiation as astress factor and the role of protective pigments. In: Lumsden, P.J. (Ed.), Plantsand UV-B. Responses to Environmental Change. Cambridge Univ. Press, UK, pp.157–168.
Bowler, C., Van Camp, W., Van Montagu, M., Inze, D., 1994. Superoxide dismutase inplants. CRC Crit. Rev. Plant Sci. 13, 199–218.
Bradford, M., 1976. A rapid and sensitive method for the quantification of micro-gram quantities of protein utilizing the principle of protein-dye binding. Anal.Biochem. 72, 248–254.
Bregman, A.A., 1987. Laboratory Investigations in Cell Biology. John Wiley and SonsInc., New York.
Caldwell, M.M., 1971. Solar UV irradiation and the growth and development ofhigher plants. In: Giese, A.C. (Ed.), Photophysiology, vol. 6. Academic Press, NewYork.
Campbell, W.S., 1975. Ultraviolet-induced ultrastructural changes in mesophyll cellsof Glycine max. In: Nachtwey, D.W., Caldwell, M.M., Biggs, R.H. (Eds.), Impactsof Climatic Change on Biosphere. Part I. Ultraviolet Radiation Effects. Monogr.U.S Dept. of Transportation. Nat. Tech Info. Ser, Springfield, VA, pp. 167–176.
Christie, P.J., Alfenito, M.R., Walbot, V., 1994. Impact of low-temperature stress ongeneral phenylpropanoid and anthocyanin pathways: enhancement of tran-script abundance and anthocyanin pigmentation in maize seedlings. Planta 194,541–549.
Costa, H., Gallego, S.M., Tomaro, M.L., 2002. Effect of UV-B radiation on antioxidantdefense system in sunflower cotyledons. Plant Sci. 162, 939–945.
Costilow, R.N., Cooper, D., 1978. Identify of proline dehydrogenase and pyrroline-5-carboxylic acid reductase in Clostridium sporogenes. J. Bacteriol. 134, 139–146.
Deutsch, J., 1983. Glucose-6-phosphate dehydrogenase. In: Bergmeyer, H.V. (Ed.),Methods of Enzymatic Analysis, 3rd ed. Verlag Chemie, VCH Publishers, DeerfieldBeach, FL, pp. 191–197.
Dhariwal, H.S., Kawai, M., Uchimiya, H., 1998. Genetic engineering for abiotic stresstolerance in plants. Rev. Plant Biotechnol. 15, 1–10.
Dorffling, K., Dorffling, H., Lesselich, G., Luke, E., Zimmerman, C., Melz, G., Jurgens,H.U., 1997. Heritable improvement of frost tolerance in winter wheat by in vitroselection of hydroxyproline-resistant proline overproducing mutants. Plant Mol.Biol. 23, 221–225.
Ervin, E.H., Zhang, X., Fike, J.H., 2004a. Ultravolet-B radiation damage on Kentuckybluegrass I: antioxidant and colorant effects. HortScience 39, 1465–1470.
Ervin, E.H., Zhang, X., Fike, J.H., 2004b. Ultravolet-B radiation damage on Kentuckybluegrass II: hormone supplement effects. HortScience 39, 1471–1474.
Ervin, E.H., Zhang, X., Fike, J.H., 2004c. Ultravolet-B radiation damage on Kentuckybluegrass III: cultivar effects. HortScience 39, 1475–1477.
Giese, M.S., Gaussoin, R.E., Shearman, R.C., Riordan, T.P., 1997. Sod production char-acteristics of turf-type Buchloe dactyloides. Int. Turfgrass Soc. Res. J. 8, 455–465.
Graham, T.L., 1991. A rapid, high resolution high performance liquid chromatogra-phy profiling procedure from plant and microbial secondary metabolites. PlantPhysiol. 95, 584–593.
Hagedorn, C.H., Phang, J.M., 1983. Transfer of reducing equivalents into mito-chondria by the interconversions of proline and pyrroline-5-carboxylate. Arch.Biochem. Biophys. 225, 95–101.
Hahlbrock, K., Scheel, D., 1989. Physiology and molecular biology of phenyl-propanoid metabolism. Plant Mol. Biol. 40, 347–369.
Iwanzik, W., Telvini, M., Dohnt, G., Voss, M., Weiss, W., Graber, P., Renger, G., 1983.Action of the UV-B radiation on photosynthetic primary reactions in spinachchloroplasts. Plant Physiol. 58, 401–407.
Jordan, B.R., James, P., Strid, A., Anthony, R., 1994. The effect of ultraviolet-B radi-ation on gene expression and pigment composition in etiolated and green pealeaf tissue: UV-B induced changes are gene-specific and dependent upon thedevelopmental stage. Plant Cell Environ. 17, 45–54.
Kostina, E., Wulff, A., Julkunen-Titto, R., 2001. Growth, structure and stomatalresponses and secondary metabolites of birch seedlings (Betula pendula) underelevated UV-B radiation in the field. Trees 15, 483–491.
Laloue, H., Weber-Lofti, F., Lucau-Danila, A., Gullemat, P., 1997. Identification ofascorbate and guaiacol peroxidase in needle chloroplasts of spruce trees. PlantPhysiol. Biochem. 35, 341–346.
Lavola, A., Julkunen-Titto, R., Aphalo, P., De La Rosa, T., Lehto, T., 1997. The effectof UV-B radiation on UV-absorbing secondary metabolites in birch seedlingsgrown under simulated forest soil conditions. New Phytol. 137, 617–621.
Mackerness, S.A.H., 2000. Plant response to ultraviolet-B (UV-B:280-320 nm) stress:what are the key regulators? Plant Growth Regul. 32, 27–39.
Murphy, T.M., Huerta, A.J., 1990. Hydrogen peroxide formation in cultured rose cellsin response to UV-C radiation. Plant Physiol. 78, 247–253.
Natori, S., Ueno, I., 1987. Flavonoides Bioact. Mol. 2, 53–85.Oberley, L.W., Spitz, D.R., 1984. Assay of SOD activity in tumor tissue. Methods in
Enzymology, vol. 105. Academic Press, pp. 457–461.
Pellegrini, N., Re, R., Yang, M., Rice-Evans, C., 1999. Screening of dietarycarotenoids and carotenoid-rich fruit extracts for antioxidant activities applying2, 2′-azinobis [3-ethylenebenzothiazoline-6-sulfonic acid] radical cation decol-orization assay. Method Enzymol. 299, 379–389.
Phang, J.M., 1985. The regulatory functions of proline and pyrroline-5-carboxylicacid. Curr. Top. Cell Regul. 25, 91–132.

2 Exper
P
P
R
R
R
S
S
S
58 D. Sarkar et al. / Environmental and
inhero, R.G., Rao, M.V., Paliyath, G., Murr, D.P., Fletcher, R.A., 1997. Changesin activities of antioxidant enzymes and their relationship to genetic andpaclobutrazol-induced chilling tolerance of maize seedlings. Plant Physiol. 114,695–704.
olle, A., 1997. Defense against photooxidative damage in plants. In: Scandalios, J.G.(Ed.), Oxidative Stress and the Molecular Biology of Antioxidant Defenses. ColdSpring Harbor Laboratory Press, Plainview, NY, pp. 623–666.
ao, M.V., Paliyath, G., Ormrod, D.P., 1996. Ultraviolet-B and Ozone-induced bio-chemical changes in antioxidant enzymes of Arabidopsis thaliana. Plant Physiol.110, 125–136.
ice-Evans, C.A., Miller, N.J., Paganga, G., 1997. Antioxidant property of phenoliccompounds. Trends Plant Sci. 2, 152–159.
osa, T.D., Julkunen-Titto, R., Lehto, T., Aphalo, P.J., 2001. Secondary metabolites andnutrient concentrations in silver birch seedlings under five levels of daily UV-Bexposure and two relative nutrient addition rates. New Phytol. 150, 121–131.
chmidt, R.E., Zhang, X., 2001. Alleviation of photochemical activity decline of tur-
fgrasses exposed to soil moisture stress or UV radiation. Int. Turfgrass Res. J. 9,340–346.hetty, K., 1997. Biotechnology to harness the benefits of dietary phenolics; focuson Lamiaceae. Asia Pac. J. Clin. Nutr. 6, 162–171.
hetty, K., 1999. Phytochemicals: biotechnology of phenolic phytochemicals for foodpreservatives and functional food applications. In: Francis, F.J. (Ed.), Encyclo-
imental Botany 70 (2011) 251–258
pedia of Food Science and Technology, 2nd ed. Wiley Wiley, New York, pp.1901–1909.
Shetty, K., 2004. Role of proline-linked pentose phosphate pathway in biosynthesisof plant phenolics for functional food and environmental applications: a review.Process Biochem. 39, 789–803.
Shetty, K., Curtis, O.F., Levin, R.E., Witkowsky, R., Ang, W., 1995. Prevention of virti-fication associated with in vitro shoot culture of oregano (Origanum vulgare) byPseudomonas spp. J. Plant Physiol. 147, 447–451.
Shindell, D.T., Rind, D., Lonergan, P., 1998. Increased polar stratospheric ozonelosses and delayed recovery owing to increasing greenhouse-gas concentrations.Nature 392, 589–592.
Smith, G.J., Markham, K.R., 1998. Tautomerism of flavonol glucosides-relevance toplant UV protection and flower color. J. Photochem. Photobiol. 118, 99–105.
Taalas, P., Kaurola, J., Kylling, A., 2000. The impact of greenhouse gases and halo-genated species on the future solar UV radiation doses. Geograph. Res. Lett. 27,1127–1130.
Teramura, A.H., Sullivan, J.H., 1994. Effects of UV-B radiation on photosynthesis andgrowth of terrestrial plants. Photosynth. Res. 39, 463–473.
Willekens, H., Van Camp, W., Van Montagu, M., Inze, D., Langebartels, C., Sander-mann, H., 1994. Ozone, sulfur dioxide, and ultraviolet B have similar effects onmRNA accumulation of antioxidant genes in Nicotiana plumbaginifolia L. PlantPhysiol. 106, 1007–1014.