Embed Size (px)
Citation preview

Development 112, 997-1008 (1991)Printed in Great Britain © The Company of Biologists Limited 1991
997
The regulation of the cell cycle during Drosophila embryogenesis: the
transition to polyteny
ALBERT V. SMITH and TERRY L. ORR-WEAVER*
Department of Biology, Massachusetts Institute of Technology and Whitehead Institute, Cambridge, MA 02142, USA
*To whom correspondence should be addressed at: The Whitehead Institute, Nine Cambridge Center, Cambridge, MA 02142, USA
Summary
The process of polytenization plays a crucial role inDrosophila development, and most of the larval tissuesare polytene. By analyzing the pattern of DNAreplication in embryos pulse-labeled with BrdU, weshow that many larval tissues undergo a transition tobegin becoming polytene late in embryogenesis. Ourresults demonstrate that in these larval tissues polytenyresults from a modified cell cycle, the endo cell cycle, inwhich there is only an S (synthesis) phase and a G (gap)phase. A key regulator of the mitotic cell cycle, theproduct of the string gene (the Drosophila homologue ofcdc25), is not required for the endo cell cycle. Thedevelopmental regulation of the endo cell cycle is
striking in that tissue-specific domains undergo polyteneDNA replication in a dynamic pattern at defined times inembryogenesis. During subsequent rounds of the endocell cycle in late embryogenesis and first instar larvaldevelopment, the domains are subdivided and thetemporal control is not as rigid. The length of the Gphase varies among different tissues. By quantifyingDNA content, we show that during the early polytene Sphases the genome is not fully duplicated.
Key words: cell cycle, polytene, Drosophila development,string.
Introduction
The presence of polyploid or polytene cells is wide-spread in nature, in fact some 300 examples have beendiscovered in animals, plants and protozoans (forreviews see Nagl, 1978; Brodsky and Uryvaeva, 1984).Polytene chromosomes are found in cells as diverse asthe macronucleus of ciliates and the trophoblast cellsthat give rise to the mammalian placenta. Many tissuesthat become polytene or polyploid are highly meta-bolically active and synthesize large amounts ofparticular products.
Polyploid or polytene cells increase ploidy bydissociating DNA replication from mitosis, yet little isunderstood about how this is regulated. Polytene cellsare defined by the visible synapsis of the multiple copiesof the chromosomes. The term polyploid has beenapplied to three types of cells: those in which theendoreplicated chromosomes are not synapsed orvisible, those is which cyclic condensation of thechromosomes reveals multiple distinct sets, and in somecases multinucleate cells have been referred to aspolyploid also. This range of polyploid cell typessuggests that there are several levels at which DNAreplication can be uncoupled from mitosis: cytokinesismay be blocked, both cytokinesis and nuclear divisionmay not occur, or chromosome segregation may be
missing as well. Polytene cells are the most deviant fromthe normal diploid cell cycle since they retain somedegree of synapsis of the replicated chromatids andexhibit none of the steps of mitosis. Polyteny couldresult either from continuous DNA replication outsideof a cell cycle, or an altered cell cycle that excludesmitosis.
Dipterans are useful organisms in which to investi-gate the regulation and mechanism of polytenization.They employ extensive use of polyploid and polytenetissues in their developmental strategy; all of the larvaltissues except the nervous system and imaginal tissuesare polytene. Most of the information on polyteneDNA replication has been gained from analysis of thelarval salivary gland of Drosophila. Endoreplication hasbeen analyzed in the larval salivary glands by quantify-ing DNA content at different larval developmentaltimes and by pulse labeling with [3H]thymidine (Rud-kin, 1973; Pearson, 1974; Hammond and Laird, 19856).These studies showed that the DNA content of nucleifell into discrete classes whose mean values differed bya factor of two, and that there were periods when[3H]thymidine was not incorporated into most of thesalivary gland nuclei. This implied that during the larvalperiods polytene DNA replication is cyclic, rather thancontinuous. The quantitation also showed that duringan S phase in the salivary gland cells the total genome is

998 A. V. Smith and T. L. Orr-Weaver
not completely duplicated, and approximately 20-30%is underrepresented (Rudkin, 1969; Hammond andLaird, 1985b). Correlation of the [3H]thymidine label-ing pattern with the Drosophila salivary gland chromo-some banding pattern by in situ analysis suggested thateach S phase has an initial period of discontinuouslabeling, followed by continuous labeling and finallyanother period of discontinuous label. This pattern isreproducible, permitting the positions of the latereplication labeling to be mapped (for review seeRudkin, 1972). Thus the characterization of salivarygland replication in the late larval stages stronglysuggested that replication was cyclic, demonstrated thatthe genome was not fully duplicated and showed atemporal order of replication of genomic regions.However, direct information about the onset ofpolytenization during development of the salivary glandwas lacking, as was the ability to investigate theproperties of a specific S phase in endoreplication.
The process of endoreplication in Drosophila larvaltissues other than the salivary gland has not beeninvestigated extensively, and there is no direct evidenceas to whether the replication is cyclic or when, duringdevelopment, polyteny initiates in these tissues. Analy-sis of the adult polyploid nurse cells and follicle cellsindicated that DNA replication in these tissues wascyclic (Mahowald et al. 1979; Hammond and Laird,1985a). It has been demonstrated that differentpolytene tissues in the larvae attain different levels ofpolyteny. For example the salivary gland becomes1024-2048C (Rudkin, 1972), while Richards has esti-mated the fat body to be 256C (Richards, 1980), Lamband Hochstrasser find the Malpighian tubules to be256C (Lamb, 1982; Hochstrasser, 1987), and Lamb hasreported the midgut to be 64C (Lamb, 1982). This tissuespecificity in the degree of polyteny could be aconsequence of slower replication, of initiating poly-tene DNA replication at different points in develop-ment, or of cessation at different times.
We chose to investigate the DNA replicationoccurring during embryogenesis to determine whenlarval tissues underwent a transition to polyteny. Bypulse labeling staged embryos with BrdU (bromo-deoxyuridine) we were able to visualize when andwhere DNA replicated in whole-mount embryos. Thetime of endoreplication could be correlated withdevelopmental events, and a single specific S phasecould be analyzed for each tissue. Thus we were able totest directly whether DNA synthesis was cyclic in manypolytene tissues and to analyze the tissue specificity ofthe onset of polyteny. Our results establish thatpolyteny arises through cyclic DNA synthesis in whatwe term an endo cell cycle; here we describe thecharacteristics of this novel cell cycle.
Materials and methods
Fly strainsWild-type Drosophila melanogaster embryos were collectedfrom the Canton-S strain. The stg7M53 allele was originallyisolated by Jurgens et al. (1984).
BrdU labeling of embryosDrosophila embryos were labeled with BrdU following amodification of the protocol of Bodmer et al. (1989). Aftercollection for 40 min at 25°C, embryos were incubated at 25°Cuntil they had reached the appropriate developmental stagefor each experiment. The embryos were dechorionated inbleach, rinsed extensively with distilled water, and once withthe Drosophila tissue culture medium M3 (Ashburner, 1989).To allow BrdU uptake, the embryos were permeabilized byincubation in octane (Aldrich) with agitation for 5 min. Thenthey were transferred to M3 with lmgml"1 BrdU (Boehr-inger-Mannheim) and incubated for 30 min at room tempera-ture. In some experiments the BrdU was injected into theembryos. Prior to fixation the labeled embryos were rinsedwith M3. BrdU-labeled embryos that had not yet undergonecuticle deposition were fixed in a 1:1 mixture of 3.7%formaldehyde (in 0.1M KPO4 pH7.5):heptane (Ashburner,1989) and subsequently devitellinized using methanol. Theywere then rehydrated to lxPBS in a methanol series. Allsamples were hydrolyzed and antibody labeled as described(Bodmer etal. 1989). The secondary antibody was a goat anti-mouse antibody conjugated to horseradish peroxidase (Bio-Rad). Ages given for embryos shown in the figures representthe midpoint age of the collection at the time of fixation.
To examine replication in whole-mount embryos that hadsecreted cuticle, multiple holes were made in the cuticle ofembryos, using forceps, to allow antibody entry. To dissecttissues, embryos were transferred to double-stick tapefollowing the labeling period and covered with Drosophilasaline. They were hand devitellinized and dissected with glassneedles. The relevant tissue was transferred to a poly-L-lysinecoated slide and the tissue was fixed in 3:1 (ethanol:aceticacid) for 1 h. The tissue was rehydrated through an ethanolseries. All samples were then hydrolyzed and antibodylabeled as described above.
BrdU labeling of larvaeLarvae were labeled by feeding them BrdU using a protocolsimilar to that previously described (Truman and Bate, 1988).A one hour collection of embryos was allowed to incubate at25°C (approximately 19 h) until larvae hatched. These larvaewere transferred to media containing agar (0.8%), sucrose(5.0%), yeast extract (5.0%), and inactivated yeast (2.0%)and allowed to grow to the appropriate age. They were thenfed on food containing 1 mgml"1 BrdU with lower amounts ofyeast extract (1.0%) and inactivated yeast (0.5%) for lh at25°C. After the labeling period the larvae were dissected inDrosophila saline and the tissues to be examined weretransferred to poly-L-lysine coated slides. The tissues werefixed in 3:1 (ethanol:acetic acid) for lh, rehydrated throughan ethanol series, hydrolyzed, and antibody labeled.
Alternatively, to look at whole-mount larvae, wholelabeled larvae were fixed in 3:1 (ethanol:acetic acid) for lhand subsequently rehydrated through an ethanol series. Whilethe larvae were in lxPBS, extensive holes were made in thelarval cuticle to allow antibody entry. Antibody staining wasas described above.
Staining with anti-tubulin and anti-histone antibodiesA monoclonal antibody against ^-tubulin (Amersham) at a1:2000 dilution was used to examine mitotic figures indissected tissues from 11-13 h embryos. The signal wasdetected with a rhodamine-conjugated goat anti-mouseantibody (Jackson Immuno Research Laboratories).
To ensure that dissected larval tissues were accessible toantibodies they were bound to an anti-histone monoclonal

The endo cell cycle and polyteny 999
antibody (Chemicon) at a 1:500 dilution followed by theHRP-conjugated secondary antibody described above.
MicroscopyA Zeiss Axiophot microscope equipped with Nomarski opticsand fluorescence was used to observe the tissues, embryos andlarvae using Plan-Neofluar lOx, 25x and 40x objectives.
Quantification of DNA contentTissues to be quantified were dissected in Drosophila salineand transferred to poly-L-lysine coated slides. To control forvariation in the fixation and subsequent staining, leg imaginaldiscs from third instar larvae were dissected onto each slide.The tissue was squashed under a coverslip to disperse thecells; the slide was then frozen in liquid nitrogen, and thecoverslip removed. The tissue was fixed in 3:1 (ethanol:aceticacid) for 1 h, rehydrated in an ethanol series, and rinsed twicein lxPBS. Staining was in 0.5/igml"1 DAPI (4,6-Diamidino-2-phenylindole, Sigma) for 5min. The samples were thenwashed once in PBS, and mounted in 70% glycerol. Therelative fluorescence was measured as described in Goff andColeman (1987). A Zeiss photomicroscope interfaced to aZeiss microspectrofluorometer and a Zeiss Plan-Neofluar 25 xobjective were used. A measuring diaphragm of 0.4 mm wasused because it just circumscribed the nuclei being measured.
The values of fluorescence were measured in relativemachine units that are not directly comparable fromexperiment to experiment. For each slide the average value ofthe 2C peak of imaginal cells was used as the standard todetermine the ploidy of the endoreplicated cells. Thestatistical significance was tested using the Wilcoxon-Mann-Whitney rank test which tests whether two distributions havedifferent means (Lindren et al. 1978).
The quantification of DAPI fluorescence is very reproduc-ible in that when a specific nucleus was measured repeatedlythe range in fluorescence measured differed only by 5 %. Inaddition, results obtained in different experiments showedcomparable levels of replication, indicating the reproduc-ibility of these measurements. For both the salivary gland andthe hindgut, the peaks are not made broad by the presence ofreplicating cells, since the tissues were dissected fromembryos at a stage when these tissues were no longerendoreplicating. Replicating cells would also not make asignificant contribution to peak width in the imaginal discmeasurements because only 10 % of leg disc cells are in Sphase at any given time (Graves and Schubiger, 1982).
Results
The transition to polyteny during embryogenesisMost cells undergo endoreplication during larvaldevelopment, but the developmental time of the onsetof polyteny in larval cells had not been ascertained. Thepattern of mitoses during embryogenesis has beendescribed in detail (Campos-Ortega and Hartenstein,1985; Foe, 1989), and it is known when mitotic divisionscease. Thus we could determine whether a transition topolyteny occurred in embryogenesis following thecompletion of the mitotic divisions. We term DNAreplication that is uncoupled from mitosis in cells thatlater in development contain polytene chromosomes as'polytene', even if the ploidy has not reached sufficientlevels for the chromosomes to be visible. In embryogen-
esis there are 13 rapid cleavage divisions that occur in asyncytium; following cellularization most cells that willgive rise to tissues in the larva undergo only threeadditional mitoses that are completed by the sixth hourof embryogenesis. The neural and imaginal tissues arethe sole tissues that continue to grow mitotically duringembryonic and larval development. To determine ifpolytene DNA replication was initiated during embryo-genesis, we tested whether BrdU incorporation couldbe detected in cells at a developmental time after thecompletion of their mitotic divisions. Staged embryos ofthe appropriate age were collected, permeabilized,labeled with BrdU and the incorporated label wasvisualized by antibody staining.
These experiments revealed polytene DNA repli-cation in larval precursor cells after the last mitosis ofembryogenesis. In the period between 8 and 15 h afterfertilization, we observed DNA replication in thesalivary gland, midgut, hindgut, Malpighian tubulesand a group of cells on the dorsal side of the embryo(Fig. 1). The pattern of polytene replication wasstriking in that it occurred in tissue-specific domainsthat replicated at specific developmental times, and thereplication was not continuous (Fig. 2). For example,all of the cells of the salivary gland replicated during alh period beginning at 8.5 h after fertilization(Fig. 1A,B). Next, cells at the anterior and posterior ofthe midgut underwent replication. This replicationinitiated in a group of cells in the anterior and theposterior midgut and spread in a wave anteriorly andposteriorly, respectively (Fig. 1B-D). Replication alsooccurred in a cluster of large, flat cells located betweenthe anterior and posterior midgut (Fig. 1C). From 10 to12 h of embryogenesis the hindgut replicated; this wasfollowed by replication of the Malpighian tubules(Fig. 1C,D). Skaer had previously described replicationin the Malpighian tubules at this time after thecompletion of mitosis (Skaer, 1989). In the two hoursimmediately preceding cuticle deposition, we observeda second round of replication in the midgut (Fig. 1E,F).This replication initiated in a band of cells thatcorresponded to the site of the first constriction of themidgut, and it spread anteriorly and posteriorly so thatcells throughout the midgut replicated. The second andthird constrictions of the midgut were not preceded byreplication in a band of cells. Immediately prior tocuticle deposition BrdU was incorporated into a groupof cells on the dorsal side of the embryo (Fig. IF). TheDNA replication pattern is shown only prior to cuticledeposition in this figure. The replication occurringduring later embryogenesis and early larval stages isdescribed below.
The replication observed in the salivary gland, gutand Malpighian tubules is endoreplication leading topolyteny because periods of replication (see below for adescription of later rounds of replication) are notseparated by mitosis (Fig. 3). These tissues weredissected from 11-13 h embryos and examined formitotic figures by staining them with a monoclonalantibody against /3-tubulin. In agreement with theprevious results of others (Campos-Ortega and Harten-

1000 A. V. Smith and T. L. Orr-Weaver
Fig. 1. Endoreplication during embryogenesis. Wild-type embryos were labeled with BrdU for 30min, fixed and stainedwith an antibody using the HRP reaction. The darkly staining cells are those that incorporated BrdU during the pulse. Allof the embryos in this figure are oriented with their anterior side to the left and the dorsal side up. (A) An 8.5 h oldembryo at the germ band extended stage (stage 11), showing replication in the salivary gland (sg). The other replicatingcells in this embryo are part of the neural tissues which are dividing mitotically. (B) A 9.25 h embryo (stage 12) undergoinggermband retraction and endoreplication in the developing anterior midgut (am), posterior midgut (pm), and the salivarygland (sg). (C) By 9.75h (stage 13) the hindgut (hg) is replicating, while replication persists in the anterior and posteriormidgut. (D) A 10.5 h embryo (stage 14) in which replication has initiated in the Malpighian tubules (mt), as well ascontinuing in the anterior and posterior midgut, and hindgut (hg). (E) A 12 h embryo (stage 15) with a stripe of cellsreplicating in the midgut. (F) A 13.75 h embryo (stage 16) with more extensive labeling throughout the midgut and in agroup of cells on the dorsal side of the embryo. Magnification X190.
stein, 1985), we failed to detect mitotic figures in thesetissues (data not shown). The sensitivity of detection ofBrdU label appears to be such that DNA replication butnot DNA repair was detected. If DNA repair wasobserved, we would have predicted more homogeneouslabeling of the embryos rather than the dynamic patternof domains observed. The domains of DNA replicationare not a consequence of impermeability to BrdU sincelabeling both by permeabilization with octane andinjection of the nucleotide analogue gave the samepattern of replication domains.
In addition to demonstrating that larval precursorcells undergo a transition to polyteny during the latterpart of embryogenesis, our results directly show that
the yolk nuclei are polyploid or polytene and are thefirst nuclei to switch from a mitotic cell cycle topolyteny. After the first nine syncytial divisions themajority of the nuclei migrate to the surface of theembryo (Foe and Alberts, 1983). Approximately 50nuclei are left behind, referred to as the yolk nuclei, andgo through two more mitotic divisions. Based on theincreasing size of the yolk nuclei following their lastmitosis, it has been assumed that they becomepolyploid. We confirmed this by demonstrating thatafter cleavage cycle 11 the yolk nuclei underwent DNAreplication that was uncoupled from mitosis, thusincreasing their ploidy (data not shown).
Cells within single tissues undergoing similar devel-


Fig. 2. The spatial patterning ofendoreplication during embryogenesis. Theendoreplication domains are shown for theembryonic stages between the onset ofpolyteny and cuticle deposition. Thedeveloping gut and salivary gland are shownin grey at the times when these tissues arenot observed to replicate. (A) As the germband begins to retract (stage 11) the salivarygland (green) is the first tissue in whichendoreplication is observed; (B) Duringgermband retraction (stage 12) the anteriorand posterior midgut (red) begin toendoreplicate. The small red circlesrepresent the large replicating nuclei seenbetween the two parts of the developingmidgut; (C) During the dorsal closure stageof embryogenesis (stage 13) the hindgut(yellow) endoreplicates. Shortly thereafterreplication begins in the Malpighian tubules(purple); (D) At head involution (stage 14)replication persists in the developingmidgut, hindgut and the Malpighian tubules.The replication in the midgut proceedsanteriorly in the anterior midgut andposteriorly in the posterior midgut. As thereplication ceases in the anterior andposterior midgut, there is a stripe ofreplication near the middle of the sac-likemidgut (blue); (E) The replication seen atthe middle of the midgut is coincident withthe first constriction of the developingmidgut (stage 15). As the constrictionsappear, the stripe of labeling observed inthe midgut expands anteriorly andposteriorly. Replication is next observed ina group of cells that extend along theposterior part of the dorsal side of theembryo (magenta); (F) As the midgutbecomes more convoluted (stage 16)replication is observed to extend throughthe entire midgut and replication persists inthe dorsal cells.

The endo cell cycle and polyteny 1001
germ bandcellular extension
fertilization blastoderm ,
germ band dorsalretraction closure
NSXVl
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 hours
-•13
cleavagedivisions
salivary gland
midgut
hlndgut
malplghian tubules
post-blastodermdivisions endoreplicatlon
Fig. 3. Regulation of the cell cycle during Drosophila embryogenesis. A time line of embryogenesis until cuticle depositionis shown with morphogenetic events indicated by arrows on the top. For the tissues examined in this study the mitoticdivisions are shown below the line. Thirteen cleavage divisions occur prior to cellular blastoderm formation; the post-blastoderm divisions are indicated by black boxes, (data of Campos-Ortega and Hartenstein, 1985 and Foe, 1989). Skaercites the last division of the Malpighian tubules as being slightly later, between 6-10 h (Skaer, 1989). The periods duringwhich endoreplication occurs (as assayed by BrdU incorporation) are shown by hatched boxes for each of the tissueslabeled on the right. The length of replication in the midgut is shown as long due to the changing pattern of labelingobserved within this tissue (see Fig. 1). Because the origin of the dorsal cells is not known, the post-blastoderm mitoses forthis tissue are not illustrated.
opmental fates underwent endoreplication together as adomain (summarized in Fig. 2). The time of onset ofpolytene DNA replication relative to the last mitoticdivision for each of these lineages varied (Fig. 3). Therewere three hours between the last mitotic division in thesalivary gland precursor cells and endorephcation,while four and a half hours lapsed in the Malpighiantubules. In the salivary gland and Malpighian tubules itwas possible to identify unambiguously all of the cells ofthe tissue, and the vast majority replicated synchron-ously (Fig. 1). However, while some of the polytenedomains included every cell of a tissue, this was notalways the case. For example, a group of cells in theposterior of the hindgut, adjacent to the anal plate, didnot replicate with the other hindgut cells during the firstperiod of replication (Fig. 1). The replication domainsare reminiscent of the domains characterized for thepost-blastoderm mitotic divisions (Foe, 1989). Foeobserved that the 14th mitosis occurs in a temporallyand spatially regulated pattern in which cells undergo-ing a similar developmental fate divide together as adomain.
Although we identified the domains of the salivarygland, gut and Malpighian tubules, it is possible that wedid not identify cells of other tissues that werereplicating individually. In particular it was difficult toidentify unambiguously the cells of the fat body since itis spread throughout the embryo. In addition, we havebeen unable to assign the replicating dorsal cells to apreviously defined tissue. Nevertheless, it is clear that atransition to polyteny is made in the latter half ofembryogenesis and that this transition is developmen-tally regulated.
The nature of the endo cell cycleHaving shown that polytene replication began inembryogenesis, we investigated the regulation of itsonset and the relationship of polytenization to themitotic cell cycle. Our results indicated that polytenyresults from a modified cell cycle as opposed tocontinuous DNA rephcation. Each domain underwentreplication for a specific period of time and replicationwas not observed during other times (Figs 2, 3). Sincereplication occurred in a discrete period rather thancontinuously, polyteny appears to be a consequence ofan altered cell cycle with an S(synthesis) phase followedby a G(gap) phase, the endo cell cycle.
The BrdU labeling patterns provided a qualitativedescription of the periods of polytene replication.However we wished to quantify the actual extent ofreplication during each of these periods to address twoaspects of the endo cell cycle. First, we wanted todetermine whether the genome is replicated only onceduring each period of labeling (the presumed S phase ofthe endo cell cycle). Second, since in the larval stagesthe salivary gland chromosomes have regions that areunderrepresented and the entire genome is not endo-replicated (Rudkin, 1969), we also wanted to testwhether there was underreplication of the chromo-somes during the first round of polytene DNAsynthesis.
To measure the relative DNA content of endorepli-cated cells free from interference of surroundingtissues, it was necessary to dissect the tissues from theembryo. The desired tissue was dissected from stagedembryos and the cells were dispersed into a monolayeron a slide, and the DNA content was quantified by

1002 A. V. Smith and T. L. Orr-Weaver
Imaginal Disc
2C
«c
Imaginal Disc
4C
15 30 45 60
Rolitivt Fluora*cenc« (Uachlna Unit*)
Fig. 4. Quantitation of hindgut DNA content. Imaginaldiscs from third instar larvae and hindgut cells from 13 hembryos were dissected onto slides, flattened to amonolayer, stained with DAPI, and the relativefluorescence was measured on a Zeiss photomicroscopeequipped with a Zeiss microdensitometer. The ploidy ofthe endoreplicated hindgut cells was calculated from theratio of the mean of this peak to the mean of the 2C peakof the imaginal discs. The data were pooled from anexperiment measuring the hind guts of three animals. Datafrom a second experiment showed comparable replicationin the hindgut.
microdensitometry of DAPI-stained cells. As an inter-nal control the DNA content of the endoreplicatingtissue was compared to the DNA content of cells in theimaginal discs, which necessarily have a 2C or a 4Cvalue since they divide mitotically. Because variation inDAPI staining can result from variability in the fixationand subsequent staining, imaginal discs were dissected,fixed and stained together on the same slide as thetissues of interest.
Hindguts were dissected from 13 h embryos, after thefirst period of polytene replication was completed, andthe DNA content of the cells quantified. In the hindguttwo populations of cells were observed, one with aploidy value of 3.4C and another peak coincident withthe 2C peak of imaginal discs (Fig. 4). Therefore inthese cells the genome was replicated only once duringthe observed period of BrdU labeling. In addition, theentire genome was not duplicated in the S phase of theendo cell cycle, since the higher DNA content peak wassignificantly lower than the 4C peak of imaginal discs
10 20 30 40
Salivary Gland
8.2C
Relative Fluorwcenca (Uichlno Units)
Fig. 5. Quantitation of salivary gland DNA content.Imaginal discs from third instar larvae and salivary glandsfrom 13 h embryos were dissected and the DNA contentquantified as in Fig. 4. The data were pooled from anexperiment measuring the salivary glands of two animals.Data from two other experiments showed comparablereplication in the salivary gland.
(Wilcoxon-Mann-Whitney statistic=4.22, P<0.001).This would correspond to underreplication of 30 % ofthe genome. DAPI shows a preference in binding A-Trich DNA, and therefore if the unrepresented DNAwas heterochromatic satellite DNA, our data mayoverestimate the percent underreplication. There arediffering results as to the extent of the bias of DAPIbinding, but even in the most extreme preferencereported, our data would still show a 3.6C value for thehindgut cells, a significant underrepresentation of 20 %(Lin etal. 1977; Ashburner, 1989; Daxhelet etal. 1989).The 2C peak presumably was composed of regions ofthe hindgut that were unreplicated, such as cellsadjacent to the anal plate (Fig. 1) and imaginal cells.Our results further imply that the endoreplicatinghindgut cells had a ploidy of 2C prior to endoreplicationand that these cells entered the endo cell cycle from Gl.
The quantitation of salivary gland DNA content at13 h showed two populations of cells; one with a ploidyvalue of 2C and another with a peak at 6.2C (Fig. 5).The 6.2C peak was made up of cells that definitely hadendoreplicated since their DNA content exceeded the4C value. The 2C cells are most likely the cells of theimaginal ring of the salivary gland because they arelocated near the anterior tip of the gland in our

The endo cell cycle and polyteny 1003
squashes (data not shown). In contrast to the hindgut,the salivary gland achieved a ploidy higher than the 4Cpeak of imaginal disks. This was surprising because inmapping replication domains we only observed oneround of endoreplication in the salivary gland. Thereare three possible explanations for these results: (1) theendoreplication in the salivary gland starts from G2 ofthe mitotic cell cycle; (2) there is an earlier round ofreplication before involution of the salivary gland; or(3) the genome replicates twice during the observedperiod of replication. We favor the first explanationbecause the precursors of the salivary gland divide onlytwice after blastoderm formation, and it has beenshown that during these two mitotic divisions cells enterS phase immediately after the completion of mitosiswithout an intervening Gl phase (Campos-Ortega andHartenstein, 1985; Edgar and O'Farrell, 1990). Thus itis likely the cells of the salivary gland enter the endo cellcycle with a 4C value. However, since we are unable toisolate and quantify the precursor cells for the salivarygland prior to its involution, we cannot rule out theother two alternatives. The 6.2C value observed doesnot correspond to an integral doubling of the genomiccontent. The percent underreplication that this rep-resents would depend on whether two rounds ofendoreplication occurred or whether the salivary glandcells entered the endo cell cycle from G2.
The regulation of the endo cell cycleSince our results showed that polyteny in many tissuesarises from a cycle of alternating G and S phases andthat the transition to the endo cell cycle occurs inembryogenesis, we wanted to determine whether theendo cell cycle is affected by regulators of the diploid,mitotic cell cycle. Several Drosophila genes involved inregulating the entry into mitosis have been identified(Edgar and O'Farrell, 1989; Lehner and O'Farrell,1989; Jimenez et al. 1990; Lehner and O'Farrell, 1990a;Whitfield et al. 1990). In particular, string has beenshown to be a key regulator that drives the postblastoderm divisions (Edgar and O'Farrell, 1989); stringis a Drosophila homolog of the Schizosaccharomycespombe cdc25 gene, a positive regulator of the G2-M
transition (Russell and Nurse, 1986; Edgar and O'Far-rell, 1989). Even though mitosis is not occurring in theendo cell cycle, genes that trigger the G2-M transitioncould also drive the endo cell cycle if the mitoticmachinery was uncoupled from the cell cycle at a pointpast the transition step. If string drives the endo cellcycle then we would not expect to observe endoreplica-tion in string mutant embryos.
To test whether string regulates the endo cell cycle wedetermined whether polytene DNA replication couldbe observed in string mutant embryos. Staged stringembryos were pulse-labeled with BrdU, and the onequarter of the embryos that were string mutants wereidentified by their altered morphology. The midgutunderwent DNA replication at times when endoreplica-tion occurs in wild-type embryos (Fig. 6). To be surethat these replicating cells were not yolk nuclei, wemapped the position of yolk nuclei in string mutants byusing an enhancer trap line that labels yolk nuclei (datanot shown). Replication was also observed in othertissues that become polytene, such as the hindgut andMalpighian tubules. A group of cells at the position ofthe salivary gland replicated at the correct developmen-tal time for salivary gland replication. Moreover, thereplication observed in string embryos showed a spatialand temporal pattern similar to the pattern in wild-typeembryos, and the midgut appeared to go through tworounds of endoreplication prior to cuticle deposition.
Our experiment clearly shows that endoreplicationoccurs in the absence of string product. This result isconsistent with the observations of Edgar and O'Farrellthat the string gene is no longer expressed in wild-typeembryos when these cells are undergoing an endo cellcycle. Two other genes essential for the G2-Mtransition, cyclinA and cyclinB, also are not expressedin these tissues as they become polytene (Lehner andO'Farrell, 1989; Lehner and O'Farrell, 1990ft; Whitfieldetal. 1990).
An additional conclusion arose from our analysis ofendoreplication in string mutant embryos. Although the14th through 16th mitotic divisions are not completed instring mutants, we still observed the onset of the endocell cycle in the gut at the appropriate stage ofdevelopment. Thus the completion of all 16 mitotic
Fig. 6. BrdU labeling of astring mutant embryo.Replicating cells can be seen inthe gut of this homozygousstg7**53 mutant embryo at thesame developmental time atwhich the midgut replicates inwild-type embryos.Magnification x330.

1004 A. V. Smith and T. L. Orr-Weaver
divisions is not a requirement for the establishment ofthe endo cell cycle, and it is not necessary to titrate outmitotic factors to enable the transition to endoreplica-tion to occur.
The endo cell cycle in late embryonic and early larvaldevelopmentWe wanted to determine the length of the G phase ofthe endo cell cycle, and to derive this information weneeded to know when subsequent rounds of replicationoccurred-in the endoreplicating tissues. In addition, thefull characterization of subsequent rounds of replicationwould reveal if endoreplication persisted in domains.The only tissue in which we saw a second period ofreplication prior to cuticle deposition was the midgut, inwhich some cells reinitiated replication by 12.5h ofembryogenesis. For the specific set of midgut cells thatbegan a second round of endoreplication the G phasewas 3 h, and this second round initiated within a specificsubdomain of the midgut and then spread throughoutthe midgut.
To analyze the next round of endoreplication inadditional tissues necessitated examining embryos aftercuticle deposition. We took two approaches to over-come the problem of limited access of the antibody: (1)we dissected out tissues from BrdU-labeled animals andperformed the antibody incubation on the isolatedtissue; and (2) we opened the cuticle at multiple sites inwhole animals to allow antibody entry. To test that anyvariation in BrdU labeling was not due to differentialaccessibility to the antibody, dissected first instar larvaltissues were labeled with an anti-histone antibody. Allthe cells within the tissues were observed to stain withthis control antibody (data not shown).
We did not observe another period of replication inthe salivary gland during embryogenesis in eitherdissected tissue or in whole-mount animals (Fig. 7).This was confirmed by quantification of the DNA
Fig. 7. The salivary gland doesnot endoreplicate during lateembryogenesis.This salivarygland with adherent fat bodywas dissected from an embryothat was labeled from the 14thhour of embryogenesis to the20th (near the time ofhatching). While replicationoccurs in the fat body, thesalivary gland does notundergo an additional round ofendoreplication during lateembryogenesis. Magnificationx470.
content of the salivary gland at hatching; the DNAcontent was identical to that of salivary glands from 15 hembryos. However, the fat body cells next to thesalivary gland did endoreplicate during late embryogen-esis (Fig. 7). The salivary gland replicated again in thefirst instar larva, 7h after hatching. Thus there was agap phase in the salivary gland of 18 h, a gap muchlonger than that observed for cells in the midgut.Therefore, the endo cell cycle differs from tissue totissue in the length of the G phase.
Endoreplication continued in developmentally regu-lated domains in the gut after cuticle deposition in theembryo and during the first larval instar, but thesedomains were smaller subdivisions of the first endo cellcycle domains. Examples of such subdomains in themidgut are shown in Fig. 8. The hindgut endoreplicatedat all times of first instar development that weexamined; the subdomains consisted of anterior-posterior stripes of cells (Fig. 8A). In addition to thesesubdomains, isolated cells replicated in the midgut,hindgut and gastric caeca (Fig. 8A,B). In some tissues,most notably the gastric caeca, isolated cells were seento replicate prior to larger numbers of cells replicatingin domains. The most extreme difference from the earlyembryonic replication domains occurred in the cells ofthe Malpighian tubules, where single cells replicatedindependently both in late embryogenesis and the firstinstar larval stage (Fig. 8A,B, and data not shown).Later in larval development replication was observed inthe gastric caeca as a domain, as well as in othersubdomains of the midgut (Fig. 8C). The subdomainsobserved in these later developmental stages comprisedthe majority of cells within a region, but not always allof the cells. Subdivision of domains as tissues differen-tiate has been observed for the post-blastoderm mitoticdomains as well (Foe, 1989).
The striking difference between the early endoreplcation domains in the embryo and those in the larva wathat the larval domains did not exhibit the same degre
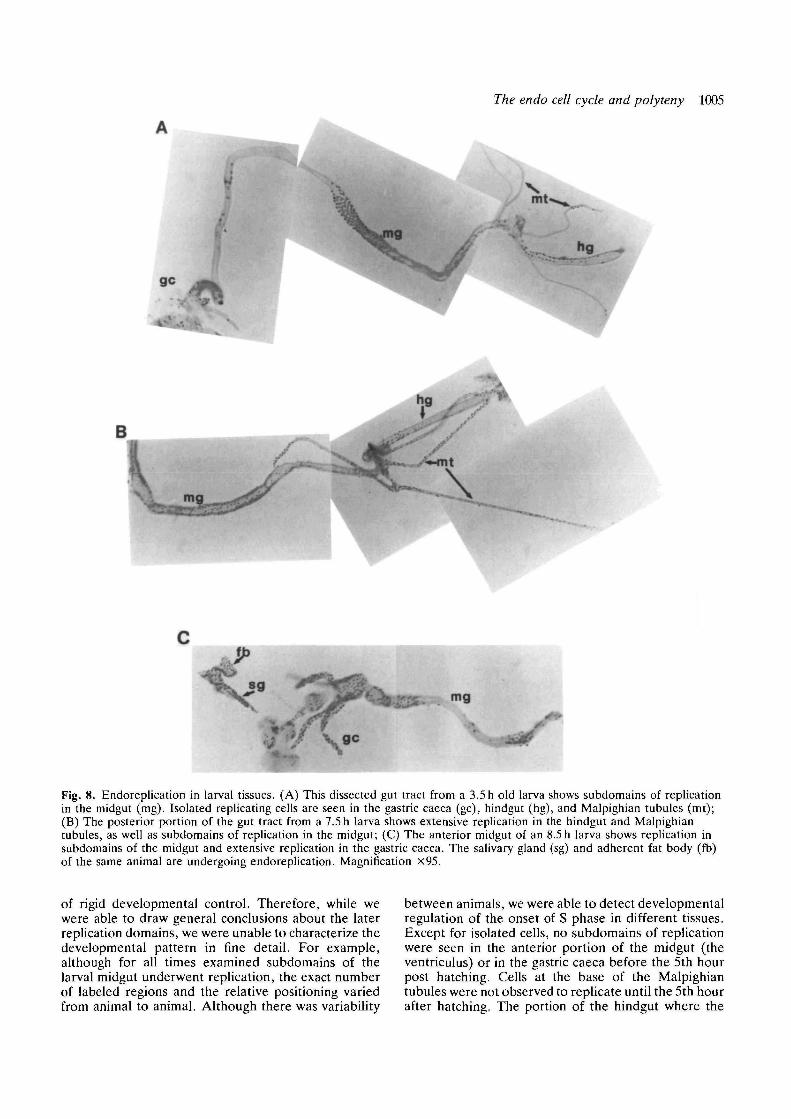
The endo cell cycle and polyteny 1005
gc
mt«
hg . -*
Fig. 8. Endoreplication in larval tissues. (A) This dissected gut tract from a 3.5h old larva shows subdomains of replicationin the midgut (mg). Isolated replicating cells are seen in the gastric caeca (gc), hindgut (hg), and Malpighian tubules (mt);(B) The posterior portion of the gut tract from a 7.5 h larva shows extensive replication in the hindgut and Malpighiantubules, as well as subdomains of replication in the midgut; (C) The anterior midgut of an 8.5 h larva shows replication insubdomains of the midgut and extensive replication in the gastric caeca. The salivary gland (sg) and adherent fat body (fb)of the same animal are undergoing endoreplication. Magnification x95.
of rigid developmental control. Therefore, while wewere able to draw general conclusions about the laterreplication domains, we were unable to characterize thedevelopmental pattern in fine detail. For example,although for all times examined subdomains of thelarval midgut underwent replication, the exact numberof labeled regions and the relative positioning variedfrom animal to animal. Although there was variability
between animals, we were able to detect developmentalregulation of the onset of S phase in different tissues.Except for isolated cells, no subdomains of replicationwere seen in the anterior portion of the midgut (theventriculus) or in the gastric caeca before the 5th hourpost hatching. Cells at the base of the Malpighiantubules were not observed to replicate until the 5th hourafter hatching. The portion of the hindgut where the

1006 A. V. Smith and T. L. Orr-Weaver
Malpighian tubules attach was not observed to replicateuntil the 7th hour after hatching, while the salivarygland initiated replication as a domain at approximatelythe same time. After this time the amount of replicationin these areas was observed to increase and reach amaximum 9h after hatching. The variability betweenanimals that we observed may result partially from thedifficulty in staging development after cuticle formationdue to the absence of morphological criteria toascertain independently developmental age. However,we think this is not the sole explanation for the variancebecause the asynchrony is observed so soon after theunambiguous developmental landmark of hatching.Environmental factors could well influence replication;such effects have been documented for salivary glandreplication in the later instars (Rudkin, 1973).
Discussion
The process of polytenization plays a significant role inDrosophila development. Previous work had estab-lished that most larval tissues are polytene andindicated that polytene DNA replication in the salivarygland was cyclic. Our results show that the cells that willconstitute many polytene larval tissues begin becomingpolytene midway through embryogenesis. PolyteneDNA replication occurs in domains that are spatiallyand temporally regulated. Moreover, the polytenereplication was cyclic in all of the tissues that weobserved. Thus the transition to polyteny involves achange from a diploid cell cycle with a Gl, S, G2 and Mphase to an endo cell cycle, in which the S phase ofDNA synthesis alternates with a G phase.
The developmental specificity of the transition topolyteny was very striking. The endo cell cycle initiatesin domains that are regulated developmentally in adynamic pattern, and after the initial rounds ofendoreplication the domains are subdivided. Althoughwe cannot be certain that the subdomains observedafter cuticle deposition correspond to regions of specificfunction, it seems likely that they reflect furtherdifferentiation of these tissues. For example, onesubdomain that we observed in the midgut is a band ofcells at which the first constriction of the gut appears.The tissue specificity and temporal regulation of thesereplication domains raises the question of whetherendoreplication is a prerequisite for or a consequenceof cell differentiation. While our data do not addressthis directly, we argue for the latter, based on recentdata on the post-blastoderm mitotic domains. The threepost-blastoderm mitotic divisions occur in domains inwhich cells that are differentiating into the same tissuesundergo mitosis together as a domain (Foe, 1989).When the string gene is placed under the control of thehsp70 promoter and induced by heat shock, the mitoticdomains are overridden and mitosis is inducedsynchronously throughout the embryo. In this situation,the embryo nevertheless develops normally, indicatingthat the mitotic domains reflect the developmental fateof cells rather than directing it (Edgar and O'Farrell,1990).
We showed that during the initial S phases of theendo cell cycle in the hindgut and salivary gland thegenome is not fully duplicated. It had been demon-strated previously that during the larval stages, thesalivary gland DNA is not fully represented; under-representation also occurs in several other polyploid orpolytene tissues (for review see Spradling and Orr-Weaver, 1987). If the underrepresented DNA is due tounderreplication, it most likely results from thedifferential activation of replication origins in particulargenomic regions, in the case of the salivary gland, in thecentric heterochromatin and euchromatin. The mech-anism by which these replication origins are shut offwith the first cycles of endoreplication remains to beelucidated. However, it has been proposed recentlythat the underrepresented DNA in polytene tissues hasbeen somatically eliminated rather than differentiallyreplicated (Karpen and Spradling, 1990). If so, ourresults show that this elimination would have to occurduring a brief interval in embryogenesis. In ourexperiments, we directly measured the extent of DNArepresentation after the first round of endoreplication.In experiments in Drosophila nasutoides Zachariasmeasured DNA content at multiple points during larvaldevelopment and by quantifying the rate of increasewas able to extrapolate that the hindgut beganunderreplication in the first endo cell cycle, the salivarygland in the second, and the Malpighian tubules in thefirst or the second endo cell cycle (Zacharias, 1986).
Polyteny arises from a cell cycle with solely an S and aG phase. Is this endo cell cycle actively regulated, or isit a default consequence of failure to complete mitosisproperly? A precedent for the latter idea is theobservation that a mutant with defective mitoticspindles in Drosophila does not arrest the cell cycle andproduces polyploid cells (Ripoll etal. 1985). The precisedevelopmental regulation of the endo cell cycle arguesfor its active regulation. Moreover, the observation thatendoreplication occurs with proper developmentaltiming in string mutants shows that the endo cell cycle isindependent of G2-M regulation and indicates that itcannot be solely a consequence of defective mitosis or acytoskeletal alteration. Certainly cytoskeletal changesmust accompany the transition to the endo cell cycle,but we assert that these changes are downstream fromthe regulatory steps as opposed to triggering the events.A predicted key cytoskeletal alteration would bedissociation of the centrosomes from the nuclearenvelope. The centrosomes have been well character-ized in the polyploid follicle cells of the Drosophilaadult, and they have been observed to disappear, butnot until several rounds of endoreplication haveoccurred. However, several changes in the morphologyof the centrosomes are coincident with the onset of theendo cell cycle, including a closer association with theplasma membrane than with the nuclear envelope, anda randomization of the orientation of the two centrioleswithin each follicle cell (Mahowald et al. 1979).
It is possible that the endo cell cycle may becontrolled by critical regulatory steps of the mitotic cellcycle. The two key regulatory points of the mitotic cell

The endo cell cycle and polyteny 1007
cycle are the Gl-S and G2-M transitions. The genesthat trigger either of these cell cycle decision pointscould be responsible for the cyclic nature of polyteneDNA replication, provided mitosis was subsequentlyuncoupled from S phase. A key regulator of both theGl-S and G2-M transitions is the kinase encoded bythe cdc2 gene, initially identified in S. pombe (Nurseand Bissett, 1981; Simanis and Nurse, 1986). Thespecificity is conferred by G2-M specific and "Gl-Sspecific cyclins (Hagan et al. 1988; Richardson et al.1989; Wittenberg et al. 1990). Our results show directlythat the key regulator of the G2-M transition inDrosophila, the product of the string gene and anactivator of cdc2, is not involved in the regulation of theendo cell cycle. Moreover, both of the G2 cyclins, cyclinA and cyclin B, are not expressed in cells undergoingpolyteny, and it is therefore unlikely that they play aregulatory role in this cycle (Lehner and O'Farrell,19906; Whitfield et al. 1990). Recent evidence suggeststhat cdc2 is also not involved in the endo cell cycle. Afunctional homologue of cdc2 that is able to comp-lement the S. pombe and S. cerevisiae mutations hasbeen cloned from Drosophila (Jimenez et al. 1990;Lehner and O'Farrell, 1990a). There is an additionalgene in the Drosophila genome that has 53 % homologyto cdc2 (Lehner and O'Farrell, 1990a). Interestingly, ithas been reported that neither of these cdc2 homol-ogues is transcribed during embryogenesis in polytenecells (Jimenez etal. 1990; Lehner and O'Farrell, 1990a).
Two alternatives remain. A novel set of regulatorygenes unique to the endo cell cycle may exist, oradditional, as yet unidentified, regulators of the mitoticcell cycle may act on both this cell cycle and the endocell cycle. The phenotype of string mutant embryos,arrested at mitosis of the first post-blastoderm division,indicates strongly that it is the limiting trigger respon-sible for entry into mitosis in Drosophila (Edgar andO'Farrell, 1989). The genetics of the Gl-S transitionhas not been investigated in Drosophila, so there existsthe possibility of regulators for this transition inaddition to cdc2 and the Gl-S cyclins. In either case,the isolation of mutations affecting the endo cell cyclewill be critical to elucidating its regulation. Ourdescription of the developmental timing of the onset ofthe endo cell cycle will be useful in screening for suchmutations.
In addition to the overall change in the cell cycleduring polytenization, feedback controls that serve ascheckpoints in the diploid cell cycle must be altered. Inparticular, the controls that ensure that another roundof DNA replication is not initiated prior to mitosis mustbe circumvented in the endo cell cycle. Blow andLaskey have shown that nuclear envelope permeabiliz-ation is sufficient to permit reinitiation of DNAreplication in a cell-free system in Xenopus (Blow andLaskey, 1988). Thus one can hypothesize that thenormal block to reinitiation could be overcome in theendo cell cycle by cyclic alterations in the nuclearenvelope. It is interesting that the pore complexes ofthe nuclear envelope of yolk nuclei go through cyclicchanges synchronously with the blastoderm nuclei,
even when the yolk nuclei are becoming polyploid(Stafstrom and Staehelin, 1984). Clearly further investi-gation of the structure of the nuclear envelope duringthe endo cell cycle is warranted.
While the employment of polyteny as a developmen-tal strategy is most pronounced in the Diptera, manyorganisms have polytene or polyploid cells. It is likelythat the novel cell cycle that we have characterized inDrosophila also functions in these other organisms. Theinsights gained from characterization of the endo cellcycle that leads to polyteny will provide a frameworkfor understanding the cell cycles that regulate poly-ploidy. Moreover, the identification of regulators forthe endo cell cycle in Drosophila will provide candi-dates for the regulation of polyteny and polyploidy inother organisms.
We thank Mike Bate, Volker Hartenstein, Jose Campos-Ortega and Norbert Perrimon for many stimulating dis-cussions and for instruction on embryo dissection andanatomy. Annette Coleman provided invaluable assistance inmicrodensitometry and access to her equipment. We acknowl-edge the advice of Rolf Bodmer on embryo permeabilizationand BrdU labeling. We are grateful to Walter Olbricht for anilluminating discussion on statistics. We thank Bruce Edgarand Pat O'Farrell for communication of results prior topublication and Allan Spradling, Ruth Lehmann, BobHorvitz, Sima Misra and members of this laboratory forhelpful comments on the manuscript. A.S. was supported byNIH predoctoral training grant GM07287 and by a grant fromthe Lucille P. Markey Charitable trust. T.O-W. was sup-ported by NIH grant GM39341, the Searle Scholars Program/The Chicago Community Trust, and by a grant from theLucille P. Markey Charitable Trust.
References
ASHBURNER, M. (1989). Drosophila. A Laboratory Manual. ColdSpring Harbor, New York: Cold Spring Harbor LaboratoryPress.
BLOW, J. J. AND LASKEY, R. A. (1988). A role for the nuclearenvelope in controlling DNA replication within the cell cycle.Nature 332, 546-548.
BODMER, R., CARRETTO, R. AND JAN, Y. N. (1989). Neurogenesisof the peripheral nervous system in Drosophila embryos: DNAreplication patterns and cell lineages. Neuron 3, 21-32.
BRODSKY, V. Y. AND URYVAEVA, I. V. (1984). GenomeMultiplication in Growth and Development: Biology of Polyploidand Polytene Cells. New York: Cambridge University Press.
CAMPOS-ORTEGA, J. A. AND HARTENSTEIN, V. (1985). The
Embryonic Development of Drosophila melanogaster. NewYork, Tokyo: Spnnger-Verlag.
DAXHELET, G. A., COENE, M. M., HOET, P. P. AND Cocrro, C. G.
(1989). Spectrofluorometry of dyes with DNAs of different basecomposition and conformation. Analyt. Biochem. 179, 401-403.
EDGAR, B. A. AND O'FARRELL, P H. (1989). Genetic control ofcell division patterns in the Drosophila embryo. Cell 57,177-187.
EDGAR, B. A. AND O'FARRELL, P. H. (1990). The threepostblastoderm cell cycles of Drosophila embryogenesis areregulated in G2 by string. Cell 62, 469-480.
FOE, V. E. (1989). Mitotic domains reveal early commitment ofcells in Drosophila embryos. Development 107, 1-22.
FOE, V. E. AND ALBERTS, B. M (1983). Studies of nuclear andcytoplasmic behaviour during the five mitotic cycles that precedegastrulation in Drosophila embryogenesis. J. Cell Sci. 61, 31-70.
GOFF, L. J. AND COLEMAN, A. W. (1987). The solution to thecytological Paradox of Isomorphy. J. Cell Biol. 104, 739-748.

1008 A. V. Smith and T. L. Orr-Weaver
GRAVES, B. J. AND SCHUBIGER, G. (1982). Cell cycle changesduring growth and differentiation of imaginal leg discs inDrosophila melanogaster. Devl Biol. 93, 104-110.
HAGAN, I., HAYLES, J. AND NURSE, P. (1988). Cloning and
sequencing of the cyclin-related cdcl3+ gene and a cytologicalstudy of its role in fission yeast mitosis. / . Cell Sci. 91, 587-595.
HAMMOND, M. P. AND LAIRD, C. D. (1985a). Chromosome
structure and DNA replication in nurse and follicle cells ofDrosophila melanogaster. Chromosoma 91, 267—278.
HAMMOND, M. P. AND LAIRD, C. D. (19856). Control of DNA
replication and spatial distribution of defined DNA sequences insalivary gland cells of Drosophila melanogaster. Chromosoma91, 279-286.
HOCHSTRASSER, M. (1987). Chromosome structure in four wild-typepolytene tissues of Drosophila melanogaster. The 87A and 87Cheat shock loci are induced unequally in the midgut in a mannerdependent on growth temperature. Chromosoma 95, 197-208.
JIMENEZ, J., ALPHEY, L., NURSE, P. AND GLOVER, D. M. (1990).
Complementation of fission yeast cdc2u and cdc25u mutantsidentifies two cell cycle genes from Drosophila: a cdclhomologue and string. EMBO J. 9, 3565-3571.
JURGENS, G . , WlESCHAUS, E . , NuSSLEIN-VoLHARD, C. AND
KLUDING, H. (1984). Mutations affecting the pattern of thelarval cuticle in Drosophila melanogaster: II. Zygotic loci on thethird chromosome. Wilhelm Roux' Arch, devl Biol. 193,2S3-295.
KARPEN, G. H. AND SPRADUNG, A. C. (1990). Reduced DNA
polytemzation of a minichromosome region undergoing position-effect variegation in Drosophila. Cell 63, 97-107.
LAMB, M. J. (1982). The DNA content of polytene nuclei inmidgut and malpighian tubule cells of adult Drosophilamelanogaster. Wilhelm Roux' Arch, devl Biol. 191, 381-384.
LEHNER, C. F. AND O'FARRELL, P. H. (1989). Expression and
function of Drosophila cyclin A during embryonic cell cycleprogression. Cell 56, 957-968.
LEHNER, C. F. AND O'FARRELL, P. H. (1990a). Drosophila cdclhomologs: a functional homolog is coexpressed with a cognatevariant. EMBO J. 9, 3573-3581.
LEHNER, C. F. AND O'FARRELL, P. H. (1990ft). The roles of
Drosophila cyclins A and B in mitotic control. Cell 61, 535-547.LIN, M. S., COMINGS, D. E. AND ALFI, O. S. (1977). Optical
studies of the interaction of 4'-6-diamidino-2-phenyhndole withDNA and metaphase chromosomes. Chromosoma 60, 15-25.
LINDREN, B. W., MCELRATH, G. W. AND BERRY, D. A. (1978).
Probability and Statistics. New York: Macmillan.MAHOWALD, A. P., CAULTON, J. H., EDWARDS, M. K. AND FLOYD,
A. D. (1979). Loss of centrioles and polyploidization in folliclecells of Drosophila melanogaster. Expl Cell Res. 118, 404-410.
NAGL, W. (1978). Endopolyploidy and Polyteny in Differentiationand Evolution: Towards an Understanding of Quantitative andQualitative Variation of Nuclear DNA in Ontogeny andPhytogeny. NY: North-Holland Publishing Co.
NURSE, P. AND BISSETT, Y. (1981). Gene required in G] for
commitment to cell cycle and in G2 for control of mitosis infission yeast. Nature 292, 558-560.
PEARSON, M. J. (1974). Polyteny and the functional significance ofthe polytene cell cycle. J Cell Sci 15, 457-479.
RICHARDS, G. (1980). The polytene chromosomes in the fat bodynuclei of Drosophila melanogaster. Chromosoma 79, 241-250.
RICHARDSON, H. E., WITTENBERG, C , CROSS, F. AND REED, S. I.
(1989). An essential G l function for cyclin-like proteins in yeast.Ce//59, 1127-1133.
RlPOLL, P . , PlMPINELU, S., VALDIVTA, M. M. AND AVILA, J. (1985).
A cell division mutant of Drosophila with a functionallyabnormal spindle. Cell 41, 907-912.
RUDKIN, G. T. (1969). Non replicating DNA in Drosophila.Genetics Supplement 61, 227-238.
RUDKIN, G. T. (1972). Replication in Polytene Chromosomes. InResults and Problems in Cell Differentiation: DevelopmentalStudies with Giant Chromosomes, 4 (eds: W. Beermann, H.Ursprung and J. Reinert), 59-85. NY: Spnnger-Verlag.
RUDKIN, G. T. (1973) Cyclic synthesis of DNA in polytenechromosomes of Diptera. In The Cell Cycle in Development andDifferentiation, (ed. Balls and Billett), pp. 279-292.Philadelphia: Institute for Cancer Research.
RUSSELL, P. AND NURSE, P. (1986) cdc75+ functions as an inducerin the mitofic control of fission yeast. Cell 45, 145-153.
SIMANIS, V. AND NURSE, P. (1986). The cell cycle control genecdc2+ of fission yeast encodes a protein kinase potentiallyregulated by phosphorylation. Cell 45, 261-268.
SKAER, H. (1989). Cell division in Malpighian tubule developmentin D. melanogaster is regulated by a single tip cell. Nature 342,566-569.
SPRADLING, A. AND ORR-WEAVER, T. (1987). Regulation of DNA
replication during Drosophila development. A. Rev. Genet 21,373-403.
STAFSTROM, J. P. AND STAEHELIN, L. A. (1984). Dynamics of the
nuclear envelope and of nuclear pore complexes during mitosisin the Drosophila embryo. Eur. J. Cell Biol. 34, 179-189.
TRUMAN, J. W AND BATE, M. (1988). Spatial and temporalpatterns of neurogenesis in the central nervous system ofDrosophila melanogaster. Devl Biol. 125, 145-157.
WHITFIELD, W., GONZALEZ, C , MALDONADO-CODINA, G. AND
GLOVER, D. (1990). The A- and B-type cyclins of Drosophila areaccumulated and destroyed in temporally distinct events thatdefine separable phases of the G2 - M transition. EMBO J. 9,2563-2572.
WITTENBERG, C , SUGIMOTO, K. AND REED, S. I. (1990). Gl-specific
cyclins of S. cerevisiae: cell cycle periodicity, regulation bymating pheromone, and association with the p34CDC29 proteinkinase. Cell 62, 225-237.
ZACHARIAS, H. (1986). Tissue-specific schedule of selectivereplication in Drosophila nasutoides. Roux's Arch, devl Biol.195, 378-388.
(Accepted 16 April 1991)





























