Embed Size (px)
Citation preview

The Possible Role of EWS-Fli1 in Evasion of Senescence in
Ewing Family Tumors
Tomoya Matsunobu,1Kazuhiro Tanaka,
1Tomoyuki Nakamura,
1Fumihiko Nakatani,
1
Riku Sakimura,1Masuo Hanada,
1Xu Li,
1Takamitsu Okada,
1Yoshinao Oda,
2
Masazumi Tsuneyoshi,2and Yukihide Iwamoto
1
Departments of 1Orthopaedic Surgery and 2Anatomic Pathology, Graduate School of Medical Sciences, Kyushu University, Fukuoka, Japan
Abstract
The chromosomal translocation t(11;22) yields the EWS-Fli1fusion gene and is associated with oncogenesis of Ewing familytumors (EFT). In this study, using the RNA interferencemethod,we show that EWS-Fli1–targeting small interfering RNAs(siRNA) depleted EWS-Fli1 protein and caused growth inhibi-tion in EFT cells with the accumulation of p27 protein andthe down-regulation of Skp2 protein in dose-dependent, time-dependent, and sequence-specific manners. Depletion of EWS-Fli1 subacutely elicited a senescence-like phenotype, but notapoptosis, in EFT cells. Furthermore, not only the knockdownof p27, but also the forced expression of Skp2, reduced theexpression levels of p27 protein and partially rescued senes-cence-like phenotype caused by EWS-Fli1–targeting siRNAs.The accumulation of p27 protein in EWS-Fli1–depleted cellsinhibited cdk2 kinase activity and was related to the stability ofp27 protein, which resulted from a decrease in Skp2 protein.Immunohistochemical analysis of p27 and Skp2 proteins inEFT samples revealed that there was an inverse relationshipbetween the expression profiles of p27 and Skp2 proteins.These findings indicate an important role of EWS-Fli1 in theprevention of senescence, leading to the unlimited growth andoncogenesis of EFT cells through a decrease in the stability ofp27 protein due to increased action of Skp2-mediated 26Sproteasome degradation. (Cancer Res 2006; 66(2): 803-11)
Introduction
Ewing family tumors (EFT) are rare tumors and account forf2%of childhood cancers (1). A specific translocation between chromo-somes 11 and 22 is found in over 85% of EFTs (2), which results in thegeneration of the EWS-Fli1 chimeric gene (3). The chimeric geneproduct, EWS-Fli1 protein, functions as an aberrant transcriptionfactor. A number of studies have suggested that EWS-Fli1 can act asan oncogene (4, 5). The possible roles of EWS-Fli1 in the cell cycle,differentiation, and apoptosis have already been reported (6–14). It iswidely appreciated that the program of apoptosis is retained inmanytypes of leukemias and solid tumors and that apoptosis contributesto the tumor response to anticancer agents. It has been widelyaccepted that a disturbance of the apoptotic pathway is implicatedin the acquisition of a malignant phenotype. On the other hand, it
would seem that sufficient attention has not been paid to therelationship between senescence and neoplastic transformation,especially in fusion gene–related tumors.Previously, we have reported that the use of antisense oligode-
oxynucleotides to suppress EWS-Fli1 expression significantlyreduced the growth of tumor cells both in vitro and in vivo andthat EWS-Fli1 modulates the cell cycle regulatory genes, such ascyclin D1, cyclin E, p21 , and p27 , whereas the inhibition of EWS-Fli1expression resulted in arrest in the G1 phase of the cell cycle (6–9, 14).Notably, a recent study indicated that p27 plays an important role inthe inhibition of tumor development and shows the characteristicsof a tumor suppressor gene (15). Moreover, p27 has been reported tobe related with cellular senescence, especially premature senescence(16–18). Because we have previously reported the prognostic andtherapeutic relevance of p27 in EFTs (8), it is very interesting toinvestigate whether EWS-Fli1 could be intimately related to p27 orsenescence.Herein, we propose a new concept that EWS-Fli1 has an
antisenescent function. In this study, we report that small interferingRNAs (siRNA) against the breakpoint of EWS-Fli1 mRNA might be avery efficient agent with which to inhibit the expression of EWS-Fli1and the growth of EFTcells, and that EWS-Fli1 might have functionsthat prevent the induction of senescence in cells through thepromotion of Skp2-mediated and 26S proteasome–dependentdegradation of p27 protein. To improve our understanding of therole and clinical relevance of p27 and Skp2 in EFTs, we immuno-histochemically studied their expression in 25 patients. The presentstudy provides a new insight suggesting that the evasion ofsenescence is one of the important mechanisms of oncogenesis ofEFTs by EWS-Fli1.
Materials and Methods
Construction of siRNA duplexes. We purchased 21 nucleotides of siRNAs
corresponding to the sequence of the breakpoint of EWS-Fli1 type I (siBPEFI;
5V-GCAGCAGAACCCUUCUUAU) and type II (siBPEFII; 5V-GGCAGCAGAGUU-CACUGCU), 21 nucleotides of siRNA corresponding to the sequence withinthe 5Vupstream half of EWS-Fli1 mRNA (siEWS1; 5V-GCCCACUGAGACUAGU-CAA), siRNA for p27 (sip27; 5V-GGAGCAAUGCGCAGGAAUAUU), and the
scrambled siRNA (siScr) wasused as a control fromDharmacon (Lafayette, CO).
Transfection of siRNA was carried out with OligofectAMINE (Invitrogen,Rockville, MD) according to the protocol of the manufacturer. Cotransfection
of expression vectors and siRNAs was carried out with LipofectAMINE 2000
(Invitrogen) as described by themanufacturer for EFTcells or HEK 293 cells. To
ensure that the target molecules were efficiently depleted, cells for apoptosis-related assays and senescence-associated h-galactosidase staining assay weretransfected twice with the corresponding siRNA, with the second transfection
4 days after the first transfection.Cell lines and cell culture. The EFT cell lines SK-N-MC, WE-68, PNKT-1,
and RD-ES were cultured as described previously (8, 19). SK-N-MC, PNKT-1,
and WE-68 have EWS-Fli1 type I fusion, whereas RD-ES has the type II
Note: Supplementary data for this article are available at Cancer Research Online(http://cancerres.aacrjournals.org/).
Requests for reprints: Kazuhiro Tanaka, Department of Orthopaedic Surgery,Graduate School of Medical Sciences, Kyushu University, 3-1-1 Maidashi, Higashi-ku,812-8582 Fukuoka, Japan. Phone: 81-92-642-5488; Fax: 81-92-642-5507; E-mail:[email protected].
I2006 American Association for Cancer Research.doi:10.1158/0008-5472.CAN-05-1972
www.aacrjournals.org 803 Cancer Res 2006; 66: (2). January 15, 2006
Research Article
Research. on September 7, 2018. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from

fusion. Human embryonic kidney HEK-293 cells and a breast carcinoma cellline MCF-7 obtained from the American Type Culture Collection (Rockville,
MD) were cultured at 37jC, 5% CO2 in DMEM (Nissui, Tokyo, Japan)
supplemented with 10% fetal bovine serum (Invitrogen). The subtypes of
EWS-Fli1 in the EFT cells were determined by reverse transcription-PCR(RT-PCR) and sequencing. The number of cells was counted using a
COULTER Hematology Analyzer (Beckman Coulter, Fullerton, CA).
Microscopy. For immunofluorescence microscopy, the cell cultured on
the chamber slides were fixed with 4% formaldehyde in PBS for 10 minutes
at room temperature and permeabilized with 0.1% Triton X-100 (Sigma,
St. Louis, MO) in PBS, and then filamentous actin was stained for 1 hour
with 10 AL of rhodamine-phalloidin (Invitrogen) in 200 AL PBS (0.3 Amol/L
final concentration), and washed extensively with PBS as described
previously (20). The cells were examined using confocal laser scanning
microscopy (Olympus, Tokyo, Japan).
Western blot analysis, immunoprecipitation, and cdk2 kinaseactivity assay. Western blot analysis, cell extract preparation, immuno-
precipitation, and cdk2 kinase activity assay using histone-H1 as a
substrate were carried out exactly as described previously (6–9). Nuclear
extracts were prepared as described previously (9). The antibodies used
were as follows: mouse monoclonal anti-human p27 (Kip1; Transduction
Laboratories, Lexington, KY), rabbit polyclonal anti-human p27, phospho-
specific (Thr187; Calbiochem, San Diego, CA), rabbit polyclonal anti-human
Fli1 (Santa Cruz Biotechnology, Santa Cruz, CA), rabbit polyclonal
antipoly(ADP ribose) polymerase (Roche, Mannheim, Germany), rabbit
polyclonal anti-human cdk2 (Santa Cruz Biotechnology), rabbit polyclonal
anti-human Skp2 (Santa Cruz Biotechnology), mouse monoclonal antiactin
(Chemicon, Temecula, CA), and horseradish peroxidase–conjugated anti-
mouse or anti-rabbit secondary antibodies (Santa Cruz Biotechnology).
Real-time quantitative RT-PCR. Real-time quantitative RT-PCR(TaqMan PCR) was done as described previously (8). The primers used for
the detection of EWS-Fli1 transcripts were the forward primer 5V-GGCA-GCAGCCTCCCACTAG-3Vand the reverse primer 5V-CCATGCTCCTCTTCT-GAC-TGAGT-3V. The sequence of the TaqMan probe used to quantify theRT-PCR products of EWS-Fli1 was 5V-(Fam)CCACCCCAAACTGGATCCTA-
CAGCC(TAMRA)-3V. The standard DNA templates containing EWS-Fli1 type
I and type II were generated by PCR and subcloned into the pCR 2.1 TOPOvector (Invitrogen). The mixtures of the primers and the probe for
glyceraldehyde-3-phosphate dehydrogenase (GAPDH) as an internal control
were purchased from PE Applied Biosystems (Foster City, CA). The relative
amount of EWS-Fli1 was standardized against the amount of GAPDHmRNA (8).
Senescence-associated B-galactosidase staining. Cells were seeded
onto the slide chamber in appropriate media, transfected with siRNAs
(100 nmol/L). Senescence-associated h-galactosidase staining was doneas described previously (21). Senescence was scored by determining the
percentage of the population that exhibited a senescence-associated
h-galactosidase activity (21).
Vectors. The expression vectors containing human wild-type p27 cDNAand p27 mutant cDNA, in which methionine at amino acid position 187 and
isoleucine at the position 188 are substituted for threonine and proline,
respectively (p27mt), were purchased from InvivoGen (San Diego, CA). Anexpression vector containing human Skp2 cDNA (22), a gift from Dr. Masaki
Mori (Departments of Molecular and Surgical Oncology, Medical Institute
of Bioregulation, Kyushu University, Beppu, Japan), was digested with
EcoRI/XhoI and the excised cDNA was ligated to the KpnI/EcoRI site of theXpress-tagged expression system pcDNA3.1/His, containing an amino-
terminal Xpress-epitope (Invitrogen). Expression vectors for EWS-Fli1 were
gifts from Dr. C.T. Denny (Gwynne Hazen Cherry Memorial Laboratories,
University of California at Los Angeles, Los Angeles, CA).Degradation of p27 protein. WE-68 cells 16 hours after transfection of
siScr or siBPEFI were treated with 10 Ag/mL cycloheximide for various
times. Cell lysates were immunoblotted for EWS-Fli1, p27, and actin protein.Densitometry was done by means of the NIH Image version 1.61 to quantify
relative amounts of protein detected on Western blot analysis.
Statistics. With regard to statistics about senescence-associated
h-galactosidase–associated experiments in vitro and relationship between
Skp2 protein and p27 protein expression in EFTs (Table 1), P valuecalculations were done with Mann-Whitney test and Fisher’s exact
test, respectively (StatView). Differences were considered significant when
P < 0.05.
Results
Reduction of EWS-Fli1 protein and mRNA expression bysiRNA molecules in dose-dependent, time-dependent, andsequence-specific manners. To knock down the expression ofEWS-Fli1 mRNA in EFT cell lines, we designed 21-nucleotidesiRNAs, corresponding to the sequence of the breakpoint of EWS-Fli1 type I (siBPEFI) or type II (siBPEFII). We also designed siEWS1,which is a 21-nucleotide siRNA corresponding to the sequencewithin the 5Vupstream half of EWS-Fli1 mRNA (i.e., 21-nucleotidesiRNAs corresponding to EWS mRNA) because we had previouslyfound that antisense oligodeoxynucleotides directed against thesequence within the 5Vupstream half of EWS-Fli1 mRNA was moreeffective for inhibiting EFT cell growth than that corresponding tothe sequence of the breakpoint of EWS-Fli1 type I mRNA and thatsuch EWS-targeting antisense oligodeoxynucleotides did not affectthe growth of cells without the EWS-Fli1 chimeric gene (14).To examine whether siRNAs would abolish the expression of
EWS-Fli1, the effects of siRNAs on the protein and mRNA levels ofEWS-Fli1 were examined by Western blot analysis and real-timequantitative RT-PCR (TaqMan PCR assay), respectively. Treatmentwith siBPEFI decreased the EWS-Fli1 protein expression in EFTcells with the type I fusion in both a time- and dose-dependentmanner (Fig. 1A, left and middle , respectively). Treatment withsiBPEFII decreased the EWS-Fli1 expression in RD-ES harboringthe type II fusion, whereas siEWS1 decreased EWS-Fli1 proteinexpression much more effectively than siBPEFII did (Fig. 1A, right).The protein levels of EWS-Fli1 started to decrease within 12 hoursafter the transfection of siBPEFI and had been suppressed over theentire 8 days (Supplementary Fig. S1B and C). Consistent with ourprevious experiments using antisense oligodeoxynucleotides (7, 8),the suppression of EWS-Fli1 expression using siRNAs in WE-68,SK-N-MC, PNKT-1, and RD-ES resulted in the accumulation of p27protein, a cyclin-dependent kinase inhibitor, in both dose- andtime-dependent manners (Fig. 1A ; see also Fig. 4D). The expressionlevel of p27 protein started to increase at least at 16 hours after thetransfection of siBPEFI and continued to be high over the entire8 days (Supplementary Fig. S1B and C). For EWS-Fli1 chimerictranscripts in EFT cells, real-time quantitative RT-PCR revealedthat the level of EWS-Fli1 mRNA decreased to <20% of that of thecontrol and had been suppressed over the entire 8 days (Fig. 1B ;Supplementary Fig. S1D). Consistent with the data from Western
Table 1. Relationship between Skp2 protein and p27protein expression in EFTs
p27 LI Skp2 LI
Negative Positive
Negative 4 10Positive 9 2
NOTE: P = 0.0154, Fisher’s exact test.
Abbreviation: LI, labeling index.
Cancer Research
Cancer Res 2006; 66: (2). January 15, 2006 804 www.aacrjournals.org
Research. on September 7, 2018. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from

blot analysis, siEWS1 suppressed EWS-Fli1 mRNA expression inRD-ES cells much more effectively than siBPEFII did; however, thedifference was not significant. The results for Western blot analysisand RT-PCR in siScr-transfected cells were almost the same asthose in untreated cells throughout the experiments. These resultsindicate that EWS-Fli1–targeting siRNAs suppress EWS-Fli1expression in dose-dependent, time-dependent, and sequence-specific manners, while also up-regulating p27 expression.Knockdown of EWS-Fli1 caused inhibition of EFT cell
proliferation and elicited senescence-like phenotype. We nextinvestigated the effects of siRNA on the growth of EFT cells becausewe had previously reported that antisense oligodeoxynucleotidesagainst EWS-Fli1 mRNA inhibited the growth of EFT cells bothin vitro and in vivo but not that of non-EFT cells (14). siBPEFIspecifically and dose-dependently inhibited the proliferation ofWE-68 with EWS-Fli1 type I but not that of RD-ES with the type IIfusion (Fig. 2A and B). We obtained similar results using two other
EFT cells with the type I fusion, SK-N-MC and PNKT-1 (datanot shown). On the other hand, siBPEFII specifically inhibited theproliferation of RD-ES but not the proliferation of WE-68 or anyother cells expressing EWS-Fli1 type I (Fig. 2B). siEWS1 was alsoeffective in inhibiting EFT cell growth (Fig. 2B). Although siEWS1lost the specificity for the fusion types, siEWS1 effectively inhibitedthe growth of RD-ES and WE-68 cells compared with siBPEFI andsiBPEFII, respectively (Fig. 2B). None of the EWS-Fli1–targetingsiRNAs affected the growth of MCF-7, a breast carcinoma cell linewithout the EWS-Fli1 fusion gene (data not shown). siScr used asnegative control did not affect the proliferation of the EFT cells(Fig. 2A and B). We concluded that both the siRNA that wasdirected against the EWS gene and those that were directedagainst the breakpoint of the fusion gene were able to inhibit theproliferation of EFT cells through the suppression of EWS-Fli1expression.It remains an unsettled question as to why the knockdown of
EWS-Fli1 expression by siRNAs could result in the inhibition of EFTcell growth. Because there have been some reports suggesting thatinhibition of the expression of another oncogenic fusion gene,BCR-ABL , leads to the induction of apoptosis in chronic myeloidleukemia (23), we examined whether the knockdown of EWS-Fli1could also result in apoptosis in EFT cells. To detect apoptotic celldeath in EFT cells, we carried out a DNA fragmentation assay,Western blot analysis for the detection of cleaved PARP protein,and the differential staining of WE-68 cells with Annexin V proteinand propidium iodide through dual-color ‘‘bivariate’’ flow cytom-etry. None of these three methods detected the cell death of EFTcell lines treated with EWS-Fli1–targeting siRNAs (SupplementaryFig. S1A, B , and E). Overall, we concluded that, by itself, theknockdown of EWS-Fli1 expression would not elicit apoptotic celldeath in our EFT cells.Whereas apoptotic cells show distinct morphologic features,
including membrane blebbing, chromatin condensation, and cellshrinkage (24), the WE-68 cells treated with siBPEFI did not displaysuch apoptosis-related features. Rhodamine-labeled phalloidinstaining revealed that WE-68 and SK-N-MC treated with siBPEFIand RD-ES treated with siEWS1 for 3 days displayed a flattenedand enlarged morphology, which could imply a senescence-likephenotype (Fig. 2C ; Supplementary Fig. S2A). The siScr-treatedcells did not display any morphologic changes. To confirm whetherthese morphologic changes are associated with senescence-likephenotype, we carried out senescence-associated h-galactosidasestaining, a well-established biomarker of senescence (21). TheWE-68 cells treated with siBPEFI started to be positively stained forsenescence-associated h-galactosidase 3 days after the transfectionin a dose- and time-dependent manner, whereas the cells treatedwith siScr did not show the expression of senescence-associatedh-galactosidase (Figs. 2D and 3E ; Supplementary Fig. S2B). Weobtained similar results using PNKT-1 (Supplementary Fig. S2D).The cells transfected with siBPEFI did not proliferate over theentire 8 days (Supplementary Fig. S2C). Taken together, thesesubacute, morphologic, and biochemical changes imply thatEWS-Fli1–targeting siRNAs caused EFT cells to fall into prematuresenescence, thereby suggesting that EWS-Fli1 plays an importantrole in preventing this from happening.p27-Dependent senescence induced by EWS-Fli1–targeting
siRNA in EFT cells. In previous reports, we revealed that thedown-regulation of EWS-Fli1 expression resulted in the accumu-lation of p27 protein in several EFT cells (7, 8). Furthermore,because p27 has been reported to be related to cellular senescence,
Figure 1. A, lysates from EFT cells treated with various concentrationsof siRNAs for the required time were prepared. Time course (left) anddose-dependent (middle ) expressions of EWS-Fli1 and p27 protein in WE-68after transfection of siRNAs were examined by Western blot analysis. In thedose-dependent experiment, lysates from WE-68 cells treated with variousconcentrations of siRNAs were prepared 48 hours after transfection. Theexpression of EWS-Fli1 and p27 protein in RD-ES harboring the EWS-Fli1 type IIfusion gene was examined 48 hours after transfection with 100 nmol/L siRNAs(right ). Actin protein was used as loading control throughout the experiments. B,relative amount of EWS-Fli1 mRNA to GAPDH mRNA. RNAs from EFT cellstreated with various concentrations of siRNAs for the required time as indicatedin the figure were prepared and subjected to real-time quantitative PCR asdescribed in Materials and Methods. Time course expressions of EWS-Fli1mRNA in WE-68 after transfection with 100 nmol/L of siRNAs (left) werequantified. Dose-dependent expression of EWS-Fli1 mRNA in WE-68 (middle )and the expression of EWS-Fli1 mRNA in RD-ES cells (right ) were alsoquantified 48 hours after transfection of siRNAs. Columns, amount of EWS-Fli1mRNA extracted from cells treated with siScr (black ) and EWS-Fli1–targetingsiRNAs (white ).
EWS-Fli1 Prevents Cells from Senescence
www.aacrjournals.org 805 Cancer Res 2006; 66: (2). January 15, 2006
Research. on September 7, 2018. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from

especially premature senescence (16–18), we addressed the role ofp27 in inducing premature senescence in EFT cells treated withEWS-Fli1–targeting siRNAs.Because p27 is a potent inhibitor of cdk2 and has been linked
to cell cycle arrest (25), we examined whether an increase in theexpression of p27 protein caused by the depletion of EWS-Fli1expression could suppress cdk2 kinase activity. The protein level ofp27 was readily increased in siBPEFI-treated WE-68 cells whencompared with PBS-treated or siScr-treated cells, whereas the totalintracellular net amount of cdk2 protein did not change with thesetreatments (Fig. 3A). However, an immunoprecipitation assay usinga specific antibody against cdk2 revealed that the cdk2 complexcontained p27 protein in siBPEFI-treated WE-68 cells, but notin PBS- or siScr-treated WE-68 cells (Fig. 3B). Furthermore, thesubsequent cdk2 kinase activity assay using histone-H1 as thesubstrate revealed that in comparison with the controls, cdk2kinase activity was completely abolished 3 days after thetransfection of siBPEFI in WE-68 cells (Fig. 3C).Next, we designed a 21-nucleotide siRNA against human p27
mRNA (sip27) and introduced it into EFT cells. Whereas siBPEFIalone depleted EWS-Fli1 protein and caused accumulation of p27
protein 3 days after transfection in WE-68, the cotransfection ofsip27 with siBPEFI prevented the accumulation of p27 protein in adose-dependent manner (Fig. 3D). As we expected, the cotrans-fection of sip27 with siBPEFI decreased WE-68 and PNKT-1cells, which were positively stained for senescence-associatedh-galactosidase assay in a dose-dependent manner 8 days aftertransfection, whereas sip27 alone did not make these cellssenescent (Fig. 3E and F). These findings suggest that EWS-Fli1protein might be able to evade premature senescence, at leastin part, via the suppression of p27 protein expression. Takentogether, these data indicate that transition from a proliferativeto a quiescent phenotype apparently requires at least the abolitionof cdk2 kinase activity by p27 protein.EWS-Fli1 promotes the proteolysis of p27 protein via an
Skp2-mediated mechanism. Our next concern is how EWS-Fli1regulates p27 expression in EFT cells. In our previous study, wereported that EWS-Fli1 promotes the 26S proteasome-dependentdegradation of p27 protein (8). Recently, it was found that p27protein levels in breast cancer cells can depend on its subcellularlocalization, which would be regulated through the phosphoryla-tion status of p27 protein (26, 27). Therefore, we examined the
Figure 2. Induction of senescence by EWS-Fli1–targeting siRNAs. A, WE-68 cells harboring the EWS-Fli1 type I fusion gene were transfected with PBS, siScr,or siRNA against the breakpoint of EWS-Fli1 type I mRNA (siBPEFI ). Columns, number of cells, counted 3 days after the transfection; bars, SD. The experimentswere triplicated and repeated thrice. B, PBS, siScr, siRNAs against the breakpoint of EWS-Fli1 type I or type II (siBPEFI or siBPEFII, respectively), or a siRNAagainst the sequence of EWS mRNA (siEWS1 ) were introduced into RD-ES with EWS-Fli1 type II or WE-68 with EWS-Fli1 type I. The number of cells was counted3 days after the transfection. C, microscopic photographs of WE-68 3 days after the transfection of siRNAs. The filamentous actin was stained with rhodamine-phalloidin. Cells were examined using confocal laser scanning microscopy. Top, transverse views of the cells; bottom, sagittal sections of the cells. Bars, 10 Am. D, thepercentages of WE-68 cells that were positively stained for senescence-associated h-galactosidase. The cells transfected with PBS or various concentrationsof siRNAs for 8 days were analyzed for senescence-associated h-galactosidase staining. Columns, percentage of cells positive for senescence-associatedh-galactosidase staining; bars, SD.
Cancer Research
Cancer Res 2006; 66: (2). January 15, 2006 806 www.aacrjournals.org
Research. on September 7, 2018. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from

localization and the phosphrylation status of p27 in EFT cellsafter the treatment of EWS-Fli1–targeting siRNAs. In the result,Western blot analysis of fractionated cytoplasmic and nuclearlysates of EFT cells transfected with PBS, siScr, or EWS-Fli1–targeting siRNAs for 3 days revealed that depletion of EWS-Fli1protein resulted in the accumulation of p27 protein at not only thenuclear fraction but also the cytoplasmic fraction (Fig. 4A).Subsequently, Western blot analysis of the phosphorylationof p27 on Thr187 in WE-68 cells using a commercially available
antibody revealed that depletion of EWS-Fli1 protein in WE-68resulted in a marked accumulation of p27 protein; however, it didnot affect the amount of phosphorylated p27 protein (Fig. 3A).Next, we examined whether the phosphorylation of p27 on Thr187
residue would be required for the EWS-Fli1 effect because ithas been reported that mutation of amino acids Thr187/Pro188 toMet187/Ile188 in p27 protein (p27mt) is resistant to proteindegradation in lung cancer cells (28). HEK293 cells cotransfectedwith EWS-Fli1 and wild-type p27 expression vectors (p27WT)
Figure 3. Cellular senescence caused by depletion of EWS-Fli1 in EFT cells is dependent on p27 protein. A, Western blot analysis of lysates from cells transfectedwith siRNAs for 3 days. PBS or 100 nmol/L siScr or siBPEFI were transfected into WE-68, and the expression levels of the indicated proteins were detected byWestern blotting. B, Western blot analysis of the cdk2 complex. Cdk2 immunoprecipitates were separated in a 4% to 12% SDS-PAGE gel and immunoblotted withcdk2- and p27-specific antibodies. Equal loading was verifed by incubation with cdk2 antibodies. C, autoradiography for the kinase activity of cdk2. Cdk2 complexeswere immunoprecipitated from WE-68 cells transfected with siRNAs for 3 days and assayed for their activity using histone-H1 as the substrate as described in Materialsand Methods. D, Western blot analysis of lysates from cells transfected with siRNAs for 3 days. PBS, 100 nmol/L of siScr, or siBPEFI with or without variousconcentrations of sip27 were cotransfected into WE-68, and the expression levels of the indicated proteins were detected by Western blotting. E, representativemicrographs of WE-68 stained for senescence-associated h-galactosidase to determine senescent cells. Senescence-associated h-galactosidase staining was carriedout 8 days after cotransfection into WE-68 cells of PBS, 100 nmol/L of siScr, or siBPEFI with various concentrations of sip27. Bars, 50 Am. F (columns ), percentagesof cells that were positively stained for senescence-associated h-galactosidase shown in (E); bars, SD.
EWS-Fli1 Prevents Cells from Senescence
www.aacrjournals.org 807 Cancer Res 2006; 66: (2). January 15, 2006
Research. on September 7, 2018. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from

reduced the expression level of p27 protein compared with thosecotransfected with mock and p27WT expression vectors (Fig. 4B).Furthermore, p27mt partially prevented its degradation by EWS-Fli1, compared with p27WT (Fig. 4B). These data indicate that thephosphorylation of p27 protein on Thr187 would be required for itsdegradation by EWS-Fli1.Because the ubiquitin-proteasome pathway mainly regulates the
p27 protein level and the ubiquitination of p27 protein leads to itsdegradation, we next examined the stability of p27 protein in WE-68cells after the transfection of siRNAs by the cycloheximide inhibitionof protein synthesis (Fig. 4C, left). Quantitative analyses showed thatthe half-life of p27 protein was dramatically prolonged in siBPEFI-transfected cells, compared with that seen in siScr-transfected cells(Fig. 4C, right).Phosphorylation of p27 protein on Thr187 in vivo is required for
ubiquitination by F-box protein Skp2 and subsequent degradationof p27 protein in the late G1 phase of the cell cycle (29). It isreasonable to suppose that the mechanism to ubiquitinate p27protein after its phosphorylation on Thr187 would be disrupted inEWS-Fli1–depleted EFT cells. Therefore, we next examined theexpression levels of Skp2 protein in EFTcells. Surprisingly, depletionof EWS-Fli1 protein resulted in a reduction in Skp2 proteinexpression in all of the EFTcells tested. The expression level of Skp2protein was positively correlated with that of EWS-Fli1 protein butwas inversely correlated with that of p27 protein (Fig. 4D and E).
Collectively, we concluded that EWS-Fli1 might promote theproteolysis of p27 protein via the Skp2-mediated mechanism.Skp2 mediates p27-related premature senescence in EFT
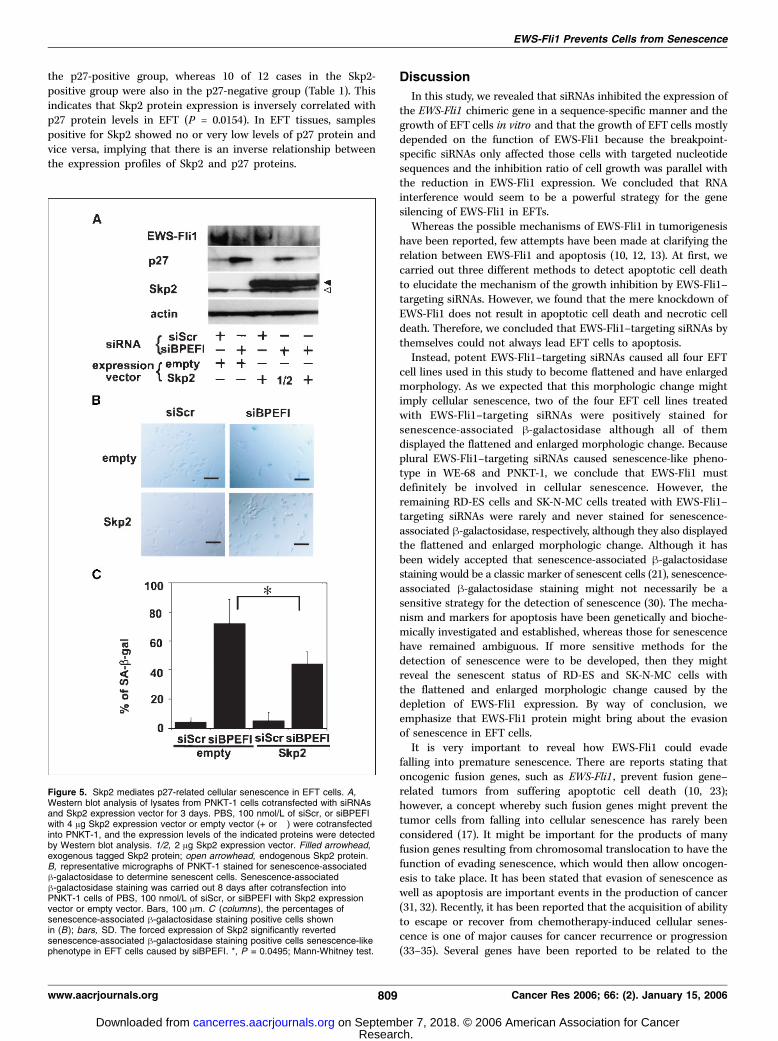
cells. To assess whether Skp2 can mediate premature senescencein EFT cells, we attempted to change the expression level of Skp2protein in EFT cells. To increase Skp2 protein in EFT cells, weintroduced an Skp2 expression vector into EFT cells. The forcedexpression of Skp2 resulted in a decrease in the p27 protein level inPNKT-1 cells and prevented the accumulation of p27 protein inEWS-Fli1–depleted cells (Fig. 5A). A senescence-associated h-galactosidase assay revealed that the forced expression of Skp2could partially, but significantly, revert senescence-like phenotypein EFT cells caused by EWS-Fli1–targeting siRNAs (Fig. 5B and C).These data suggest that Skp2 would mediate the prevention ofpremature senescence in EFT cells through the degradation of p27protein.Inverse patterns of Skp2 and p27 protein expression in
EFTs. To examine the clinical importance of Skp2 proteinexpression, we analyzed the expressions of Skp2 protein andp27 protein using an immunohistochemical staining method intissue samples obtained from primary EFTs. The relationshipbetween p27 protein expression and Skp2 protein expression wasexamined in 25 cases of EFTs. The typical immunohistochemistrypatterns for p27 and Skp2 in EFTs are shown in SupplementaryFig. S3. Nine of 13 cases in the Skp2-negative group were also in
Figure 4. EWS-Fli1 promotes theproteolysis of p27 protein via Skp2-mediatedmechanism. A, immunoblot analysis ofWE-68 cells. Fractionated cytoplasmic andnuclear lysates of WE-68 cells 3 days aftertransfection of PBS, 100 nmol/L of siScr,or siBPEFI were immunoblotted with theappropriate antibodies. B, HEK293 cells weretransfected with an expression vector thatexpresses EWS-Fli1 (lanes 5-7) or thecorresponding empty vector (lanes 2-4)together with expression vectors thatexpress p27-WT or p27mt (+ or �). Lysateswere prepared and then aliquots wereimmunoblotted with anti-Fli1, anti-p27, orantiactin antibodies. C, measurement of thedegradation of p27 protein after inhibition ofprotein synthesis by cycloheximide (CHX ).Left, WE-68 cells 16 hours after transfectionof siScr or siBPEFI were treated with10 Ag/mL cycloheximide for the indicatedtimes, and EWS-Fli1 and p27 protein levelswere monitored by Western blot analysis.Right, densitometry was done by means ofthe NIH Image 1.61 software to quantifyrelative amounts of protein detected onWestern blots (left ). D, immunoblot analysisof EWS-Fli1, Skp2, p27, and actin protein.Lysates from EFT cells 3 days aftertransfection of 100 nmol/L of siRNAs weresubjected to Western blotting. S, siScr; B,siBPEFI; E, siEWS1. E, lysates from WE-68cells 3 days after transfection of PBS orvarious concentrations of siRNAs wereimmunoblotted with the appropriateantibodies (left ).
Cancer Research
Cancer Res 2006; 66: (2). January 15, 2006 808 www.aacrjournals.org
Research. on September 7, 2018. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from
the p27-positive group, whereas 10 of 12 cases in the Skp2-positive group were also in the p27-negative group (Table 1). Thisindicates that Skp2 protein expression is inversely correlated withp27 protein levels in EFT (P = 0.0154). In EFT tissues, samplespositive for Skp2 showed no or very low levels of p27 protein andvice versa, implying that there is an inverse relationship betweenthe expression profiles of Skp2 and p27 proteins.
Discussion
In this study, we revealed that siRNAs inhibited the expression ofthe EWS-Fli1 chimeric gene in a sequence-specific manner and thegrowth of EFT cells in vitro and that the growth of EFT cells mostlydepended on the function of EWS-Fli1 because the breakpoint-specific siRNAs only affected those cells with targeted nucleotidesequences and the inhibition ratio of cell growth was parallel withthe reduction in EWS-Fli1 expression. We concluded that RNAinterference would seem to be a powerful strategy for the genesilencing of EWS-Fli1 in EFTs.Whereas the possible mechanisms of EWS-Fli1 in tumorigenesis
have been reported, few attempts have been made at clarifying therelation between EWS-Fli1 and apoptosis (10, 12, 13). At first, wecarried out three different methods to detect apoptotic cell deathto elucidate the mechanism of the growth inhibition by EWS-Fli1–targeting siRNAs. However, we found that the mere knockdown ofEWS-Fli1 does not result in apoptotic cell death and necrotic celldeath. Therefore, we concluded that EWS-Fli1–targeting siRNAs bythemselves could not always lead EFT cells to apoptosis.Instead, potent EWS-Fli1–targeting siRNAs caused all four EFT
cell lines used in this study to become flattened and have enlargedmorphology. As we expected that this morphologic change mightimply cellular senescence, two of the four EFT cell lines treatedwith EWS-Fli1–targeting siRNAs were positively stained forsenescence-associated h-galactosidase although all of themdisplayed the flattened and enlarged morphologic change. Becauseplural EWS-Fli1–targeting siRNAs caused senescence-like pheno-type in WE-68 and PNKT-1, we conclude that EWS-Fli1 mustdefinitely be involved in cellular senescence. However, theremaining RD-ES cells and SK-N-MC cells treated with EWS-Fli1–targeting siRNAs were rarely and never stained for senescence-associated h-galactosidase, respectively, although they also displayedthe flattened and enlarged morphologic change. Although it hasbeen widely accepted that senescence-associated h-galactosidasestaining would be a classic marker of senescent cells (21), senescence-associated h-galactosidase staining might not necessarily be asensitive strategy for the detection of senescence (30). The mecha-nism and markers for apoptosis have been genetically and bioche-mically investigated and established, whereas those for senescencehave remained ambiguous. If more sensitive methods for thedetection of senescence were to be developed, then they mightreveal the senescent status of RD-ES and SK-N-MC cells withthe flattened and enlarged morphologic change caused by thedepletion of EWS-Fli1 expression. By way of conclusion, weemphasize that EWS-Fli1 protein might bring about the evasionof senescence in EFT cells.It is very important to reveal how EWS-Fli1 could evade
falling into premature senescence. There are reports stating thatoncogenic fusion genes, such as EWS-Fli1 , prevent fusion gene–related tumors from suffering apoptotic cell death (10, 23);however, a concept whereby such fusion genes might prevent thetumor cells from falling into cellular senescence has rarely beenconsidered (17). It might be important for the products of manyfusion genes resulting from chromosomal translocation to have thefunction of evading senescence, which would then allow oncogen-esis to take place. It has been stated that evasion of senescence aswell as apoptosis are important events in the production of cancer(31, 32). Recently, it has been reported that the acquisition of abilityto escape or recover from chemotherapy-induced cellular senes-cence is one of major causes for cancer recurrence or progression(33–35). Several genes have been reported to be related to the
Figure 5. Skp2 mediates p27-related cellular senescence in EFT cells. A,Western blot analysis of lysates from PNKT-1 cells cotransfected with siRNAsand Skp2 expression vector for 3 days. PBS, 100 nmol/L of siScr, or siBPEFIwith 4 Ag Skp2 expression vector or empty vector (+ or �) were cotransfectedinto PNKT-1, and the expression levels of the indicated proteins were detectedby Western blot analysis. 1/2, 2 Ag Skp2 expression vector. Filled arrowhead,exogenous tagged Skp2 protein; open arrowhead, endogenous Skp2 protein.B, representative micrographs of PNKT-1 stained for senescence-associatedh-galactosidase to determine senescent cells. Senescence-associatedh-galactosidase staining was carried out 8 days after cotransfection intoPNKT-1 cells of PBS, 100 nmol/L of siScr, or siBPEFI with Skp2 expressionvector or empty vector. Bars, 100 Am. C (columns ), the percentages ofsenescence-associated h-galactosidase staining positive cells shownin (B); bars, SD. The forced expression of Skp2 significantly revertedsenescence-associated h-galactosidase staining positive cells senescence-likephenotype in EFT cells caused by siBPEFI. *, P = 0.0495; Mann-Whitney test.
EWS-Fli1 Prevents Cells from Senescence
www.aacrjournals.org 809 Cancer Res 2006; 66: (2). January 15, 2006
Research. on September 7, 2018. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from

senescence of fibroblasts, e.g., p53, p16 , and MAPK family genes(36–38). Recently, of the two senescence-inducible EFT cells, PNKT-1 does not have functional p53 (8, 19) and WE-68 with wild-typep53 has a deletion mutation in the p16 gene (39). We have alsorevealed that EWS-Fli1–targeting siRNAs did not affect thephosphorylation status of p38 protein, a member of the MAPKfamily, in EFT cells (data not shown).In this study, we addressed the role of p27 in inducing cellular
senescence in EFT cells treated with EWS-Fli1–targeting siRNAsbecause p27 has been reported to be related with cellularsenescence, especially premature senescence (16–18), and becausep27 is one of the cyclin-dependent kinase inhibitors that cause cellcycle arrest. We revealed that p27 protein, which was accumulatedby depletion of EWS-Fli1, interacted with cdk2 and interfered withthe function of cdk2 and that the down-regulation of p27 protein,which was caused by sip27 and Skp2, could induce, at least in part,EFT cells with a senescent phenotype to revert to type. Thesefindings suggest that p27 might have the important role of inducingor maintaining cellular senescence. We propose that by activelyreducing the level of p27 protein in EFT cells, EWS-Fli1 might beable to alleviate the effect of p27 on its major targets, cyclins/cdk2complexes. As such, the increased cyclin/cdk2 activities mightprompt the progression of the cell cycle and prevent EFT cells fromfalling into senescence. This led us to arrive at our conclusion thatEWS-Fli1 might evade premature senescence through the suppres-sion of p27 expression.We have reported that EWS-Fli1 might attenuate the p27 protein
level via the activation of the proteasome-mediated degradationpathway (8); however, the precise mechanism is still unclear. Manyresearchers, including ourselves, have reported that the concentra-tion of p27 protein in cells is mainly regulated by posttranslationalmechanisms (8, 40–43). F-box protein Skp2, the substrate-specificubiquitin ligase of the SCF complex (Skp1/Cul1/F-box), isresponsive for ubiquitin-dependent degradation of p27 proteinboth in vitro and in vivo . Binding of Skp2 to p27 protein requiresphosphorylation of p27 on Thr187 residue by cyclin-cdk2 complex(29, 40, 42). In this report, we found that down-regulation of p27protein by EWS-Fli1 depends on the phosphorylation of p27 onThr187 residue. We have also found that antiparallel with theaccumulation of p27 protein, the expression of F-box protein Skp2,the substrate-specific E3 ubiquitin ligase of the SCF complex, wasselectively down-regulated in all the EWS-Fli1–depleted EFT cellstested, which implies that EWS-Fli1 might up-regulate theexpression of Skp2 or stabilize Skp2 protein. Presumably, EWS-Fli1 might increase and permit Skp2 protein to accelerate theubiquitination of p27, which is a prerequisite step for the targeteddegradation of p27 protein by the 26S proteasome pathway. In EWS-Fli1–depleted EFT cells, a reduction in Skp2 protein might fail topolyubiquitinate p27 protein. Accordingly, p27 protein can beconsidered to be very stable and accumulated in EWS-Fli1–depleted
EFT cells, which would result in the inhibition of cyclins/cdk2kinase activity. The precise process of regulation of Skp2 expressionby EWS-Fli1 warrants further investigation.In senescence-related experiments, Skp2 plays an important
role in the prevention of premature senescence by EWS-Fli1 in EFTcells. Skp2 reduced the expression of p27 protein and partiallyprevented EFT cells from premature senescence caused by EWS-Fli1–targeting siRNAs. We suppose that Skp2 could be substitutedfor EWS-Fli1. Taken together, it seems that EWS-Fli1 promotes thedegradation of p27 protein and prevents cellular senescencethrough the Skp2-mediated and 26S proteasome–dependentmechanism in EFT cells.It has been reported that an inverse correlation between Skp2
and that p27 was found in several malignant tumors (44, 45). In thepresent study, we also showed that Skp2 protein levels wereinversely correlated with p27 protein levels in EFTs in this report.Although the survival curves drawn according to the Kaplan-Meiermethod showed that patients with EFTs, which were positive forSkp2, tend to have a worse prognosis than patients whose tumorswere negative for Skp2, an analysis of survival using the log-ranktest revealed that the difference was not significant (data notshown). Skp2 is irrefutably linked to proteolysis of p27 protein, butit is not the only molecule needed for the regulation of p27 protein(46). Therefore, we concluded that Skp2 might have an importantconsequence for EFTs patients; however, other mechanisms wouldbe involved in the proteolysis of p27 protein.In summary, the present study showed that EWS-Fli1
selectively accelerates p27 protein degradation by the Skp2-dependent 26S proteasome pathway and finally prevents EFTcells from falling into premature senescence. The findings of thisreport are very significant, not only because of the important roleof EWS-Fli1 in the evasion of premature senescence but alsobecause of the novel concept that oncogenic fusion genes have animportant role to play in the prevention of senescence, as well asapoptosis. Insight into the mechanism behind senescence mayprovide a promising lead to the development of therapeuticstrategies for the treatment of EFTs.
Acknowledgments
Received 6/7/2005; revised 9/29/2005; accepted 11/10/2005.Grant support: Scientific Research grants 14207057 and 17591580 from the Japan
Society for the Promotion of Science, and grant-in aid for Clinical Cancer Researchand grants-in-aid for Cancer Research (14S-4 and 14S-5) from the Ministry of Health,Labour and Welfare, Japan.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.
We thank Dr. F. van Valen (Department of Orthopaedics, University Hospital,Munster, Germany) for VH-64 and WE-68, Dr. Yoshihiko Maehara (Department ofSurgery and Science, Graduate School of Medical Sciences, Kyushu University,Fukuoka, Japan) for the ABI PRISM 7700 Sequence Detection System, and HirokoEguchi and Yuko Yagawa for their technical assistance.
References
1. Grovas A, Fremgen A, Rauck A, et al. TheNational Cancer Data Base report on patterns ofchildhood cancers in the United States. Cancer 1997;80:2321–32.
2. Turc-Carel C, Philip I, Berger MP, Philip T, Lenoir GM.Chromosome study of Ewing’s sarcoma (ES) cell lines.Consistency of a reciprocal translocation t(11;22)(q24;q12). Cancer Genet Cytogenet 1984;12:1–19.
3. Delattre O, Zucman J, Plougastel B, et al. Gene fusionwith an ETS DNA-binding domain caused by chromo-some translocation in human tumours. Nature 1992;359:162–5.
4. Ouchida M, Ohno T, Fujimura Y, Rao VN, Reddy ES.Loss of tumorigenicity of Ewing’s sarcoma cellsexpressing antisense RNA to EWS-fusion transcripts.Oncogene 1995;11:1049–54.
5. May WA, Gishizky ML, Lessnick SL, et al. Ewingsarcoma 11;22 translocation produces a chimeric
transcription factor that requires the DNA-bindingdomain encoded by FLI1 for transformation. Proc NatlAcad Sci U S A 1993;90:5752–6.
6. Li X, Tanaka K, Nakatani F, et al. Transactivation ofcyclin E gene by EWS-Fli1 and antitumor effects ofcyclin dependent kinase inhibitor on Ewing’s familytumor cells. Int J Cancer 2005;116:385–94.
7. Matsumoto Y, Tanaka K, Nakatani F, Matsunobu T,Matsuda S, Iwamoto Y. Downregulation and forcedexpression of EWS-Fli1 fusion gene results in changes in
Cancer Research
Cancer Res 2006; 66: (2). January 15, 2006 810 www.aacrjournals.org
Research. on September 7, 2018. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from

EWS-Fli1 Prevents Cells from Senescence
www.aacrjournals.org 811 Cancer Res 2006; 66: (2). January 15, 2006
the expression of G(1)regulatory genes. Br J Cancer 2001;84:768–75.
8. Matsunobu T, Tanaka K, Matsumoto Y, et al. Theprognostic and therapeutic relevance of p27kip1 inEwing’s family tumors. Clin Cancer Res 2004;10:1003–12.
9. Nakatani F, Tanaka K, Sakimura R, et al. Identifi-cation of p21WAF1/CIP1 as a direct target of EWS-Fli1 oncogenic fusion protein. J Biol Chem 2003;278:15105–15.
10. Prieur A, Tirode F, Cohen P, Delattre O. EWS/FLI-1silencing and gene profiling of Ewing cells revealdownstream oncogenic pathways and a crucial role forrepression of insulin-like growth factor binding protein3. Mol Cell Biol 2004;24:7275–83.
11. Rorie CJ, Thomas VD, Chen P, Pierce HH, O’Bryan JP,Weissman BE. The Ews/Fli-1 fusion gene switches thedifferentiation program of neuroblastomas to Ewingsarcoma/peripheral primitive neuroectodermal tumors.Cancer Res 2004;64:1266–77.
12. Soldatenkov VA, Albor A, Patel BK, Dreszer R,Dritschilo A, Notario V. Regulation of the humanpoly(ADP-ribose) polymerase promoter by the ETStranscription factor. Oncogene 1999;18:3954–62.
13. Soldatenkov VA, Trofimova IN, Rouzaut A,McDermott F, Dritschilo A, Notario V. Differentialregulation of the response to DNA damage in Ewing’ssarcoma cells by ETS1 and EWS/FLI-1. Oncogene 2002;21:2890–5.
14. Tanaka K, Iwakuma T, Harimaya K, Sato H,Iwamoto Y. EWS-Fli1 antisense oligodeoxynucleotideinhibits proliferation of human Ewing’s sarcoma andprimitive neuroectodermal tumor cells. J Clin Invest1997;99:239–47.
15. Nakayama K, Ishida N, Shirane M, et al. Mice lackingp27(Kip1) display increased body size, multiple organhyperplasia, retinal dysplasia, and pituitary tumors. Cell1996;85:707–20.
16. Collado M, Medema RH, Garcia-Cao I, et al.Inhibition of the phosphoinositide 3-kinase pathwayinduces a senescence-like arrest mediated by p27Kip1.J Biol Chem 2000;275:21960–8.
17. Martinez N, Drescher B, Riehle H, et al. Theoncogenic fusion protein RUNX1-CBFA2T1 supportsproliferation and inhibits senescence in t(8;21)-positiveleukaemic cells. BMC Cancer 2004;4:44.
18. Yoon G, Kim HJ, Yoon YS, Cho H, Lim IK, Lee JH. Ironchelation-induced senescence-like growth arrest inhepatocyte cell lines: association of transforming growthfactor h1 (TGF-h1)-mediated p27Kip1 expression.Biochem J 2002;366:613–21.
19. Tanaka K, Iwamoto Y, Noguchi Y, Oda Y, Sugioka Y.The establishment and characterization of a peripheralneuroepithelioma cell line in soft tissue of extremity.Lab Invest 1995;72:237–48.
20. Matsumoto Y, Tanaka K, Harimaya K, Nakatani F,Matsuda S, Iwamoto Y. Small GTP-binding protein, Rho,both increased and decreased cellular motility, activa-tion of matrix metalloproteinase 2 and invasion ofhuman osteosarcoma cells. Jpn J Cancer Res 2001;92:429–38.
21. Dimri GP, Lee X, Basile G, et al. A biomarker thatidentifies senescent human cells in culture and in agingskin in vivo . Proc Natl Acad Sci U S A 1995;92:9363–7.
22. Masuda TA, Inoue H, Sonoda H, et al. Clinical andbiological significance of S-phase kinase-associatedprotein 2 (Skp2) gene expression in gastric carcinoma:modulation of malignant phenotype by Skp2 over-expression, possibly via p27 proteolysis. Cancer Res2002;62:3819–25.
23. Smetsers TF, Skorski T, van de Locht LT, et al.Antisense BCR-ABL oligonucleotides induce apoptosisin the Philadelphia chromosome-positive cell lineBV173. Leukemia 1994;8:129–40.
24. Walker NI, Harmon BV, Gobe GC, Kerr JF. Patterns ofcell death. Methods Achiev Exp Pathol 1988;13:18–54.
25. Polyak K, Kato JY, Solomon MJ, et al. p27Kip1, acyclin-Cdk inhibitor, links transforming growth factor-hand contact inhibition to cell cycle arrest. Genes Dev1994;8:9–22.
26. Viglietto G, Motti ML, Bruni P, et al. Cytoplasmicrelocalization and inhibition of the cyclin-dependentkinase inhibitor p27(Kip1) by PKB/Akt-mediatedphosphorylation in breast cancer. Nat Med 2002;8:1136–44.
27. Liang J, Zubovitz J, Petrocelli T, et al. PKB/Aktphosphorylates p27, impairs nuclear import of p27 andopposes p27-mediated G1 arrest. NatMed 2002;8:1153–60.
28. Park KH, Seol JY, Kim TY, et al. An adenovirusexpressing mutant p27 showed more potent antitumoreffects than adenovirus-p27 wild type. Cancer Res 2001;61:6163–9.
29. Montagnoli A, Fiore F, Eytan E, et al. Ubiquitinationof p27 is regulated by Cdk-dependent phosphorylationand trimeric complex formation. Genes Dev 1999;13:1181–9.
30. Kwak IH, Kim HS, Choi OR, Ryu MS, Lim IK. Nuclearaccumulation of globular actin as a cellular senescencemarker. Cancer Res 2004;64:572–80.
31. Cao L, Li W, Kim S, Brodie SG, Deng CX. Senescence,aging, and malignant transformation mediated by p53 in
mice lacking the Brca1 full-length isoform. Genes Dev2003;17:201–13.
32. Campisi J. Cancer, aging and cellular senescence. InVivo 2000;14:183–8.
33. Wang Y, Zhu S, Cloughesy TF, Liau LM, Mischel PS.p53 disruption profoundly alters the response of humanglioblastoma cells to DNA topoisomerase I inhibition.Oncogene 2004;23:1283–90.
34. Roberson RS, Kussick SJ, Vallieres E, Chen SY, Wu DY.Escape from therapy-induced accelerated cellular se-nescence in p53-null lung cancer cells and in humanlung cancers. Cancer Res 2005;65:2795–803.
35. Elmore LW, Di X, Dumur C, Holt SE, Gewirtz DA.Evasion of a single-step, chemotherapy-induced senes-cence in breast cancer cells: implications for treatmentresponse. Clin Cancer Res 2005;11:2637–43.
36. Itahana K, Dimri G, Campisi J. Regulation of cellularsenescence by p53. Eur J Biochem 2001;268:2784–91.
37. Brookes S, Rowe J, Ruas M, et al. INK4a-deficienthuman diploid fibroblasts are resistant to RAS-inducedsenescence. EMBO J 2002;21:2936–45.
38. Iwasa H, Han J, Ishikawa F. Mitogen-activatedprotein kinase p38 defines the common senescence-signalling pathway. Genes Cells 2003;8:131–44.
39. Kovar H, Jug G, Aryee DN, et al. Among genes involvedin the RB dependent cell cycle regulatory cascade, the p16tumor suppressor gene is frequently lost in the Ewingfamily of tumors. Oncogene 1997;15:2225–32.
40. Nakayama KI, Hatakeyama S, Nakayama K. Regula-tion of the cell cycle at the G1-S transition by proteolysisof cyclin E and p27Kip1. Biochem Biophys Res Commun2001;282:853–60.
41. Sherr CJ, Roberts JM. Inhibitors of mammalian G1
cyclin-dependent kinases. Genes Dev 1995;9:1149–63.42. Vlach J, Hennecke S, Amati B. Phosphorylation-dependent degradation of the cyclin-dependent kinaseinhibitor p27. EMBO J 1997;16:5334–44.
43. Alessandrini A, Chiaur DS, Pagano M. Regulation ofthe cyclin-dependent kinase inhibitor p27 by degrada-tion and phosphorylation. Leukemia 1997;11:342–5.
44. Gstaiger M, Jordan R, Lim M, et al. Skp2 is oncogenicand overexpressed in human cancers. Proc Natl AcadSci U S A 2001;98:5043–8.
45. Osoegawa A, Yoshino I, Tanaka S, et al. Regulation ofp27 by S-phase kinase-associated protein 2 is associatedwith aggressiveness in non-small-cell lung cancer. J ClinOncol 2004;22:4165–73.
46. Kamura T, Hara T, Matsumoto M, et al. Cytoplasmicubiquitin ligase KPC regulates proteolysis of p27(Kip1)at G1 phase. Nat Cell Biol 2004;6:1229–35.
Research. on September 7, 2018. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from

2006;66:803-811. Cancer Res Tomoya Matsunobu, Kazuhiro Tanaka, Tomoyuki Nakamura, et al. Ewing Family TumorsThe Possible Role of EWS-Fli1 in Evasion of Senescence in
Updated version
http://cancerres.aacrjournals.org/content/66/2/803
Access the most recent version of this article at:
Material
Supplementary
http://cancerres.aacrjournals.org/content/suppl/2006/01/24/66.2.803.DC1
Access the most recent supplemental material at:
Cited articles
http://cancerres.aacrjournals.org/content/66/2/803.full#ref-list-1
This article cites 46 articles, 20 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/66/2/803.full#related-urls
This article has been cited by 8 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. (CCC)Click on "Request Permissions" which will take you to the Copyright Clearance Center's
.http://cancerres.aacrjournals.org/content/66/2/803To request permission to re-use all or part of this article, use this link
Research. on September 7, 2018. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from





























