Embed Size (px)
Citation preview

33
The Orientation of the Optic Axis of Spicules ofLeucosolenia complicata
By WALTER CLIFFORD JONES, M.A., PH.D.(From the Department of Zoology, Cambridge, and the Department of Zoology, University
College of North Wales, Bangor)
SUMMARY
The orientation of the optic axes of the spicules of Leucosolenia complicata has beenstudied by using isolated spicules and longitudinal sections of the oscular tubes. Fortri- and quadri-radiates the mean angle of inclination of the optic axis to the tubewall varies with the distance of the spicule from the oscular edge, particularly over thefirst 200 jx. The spicule form likewise varies with position, but is not closely corre-lated with the orientation of the optic axis: two spicules can have the same optic anglebut different shapes, or the same shape and yet different optic angles. For curvedmonaxons the optic axis is directed almost at right angles to the spicule axis at thedistal end, and the spicule curvature is independent of the position in the tube.Slender monaxons, on the other hand, have an optic axis coincident with the spiculelength. It seems a general rule, at least for spicules of L. complicata and L. coriacea,that the optic axis of the first formed calcite rudiment lies in the direction of the celldivision (or cell association) that directly precedes spicule formation.
A mechanism is suggested for the orientation of the optic axis. This is explained interms of an oriented overgrowth of the calcite upon an organic precursor, which isstructurally fixed and oriented within the formative cell complex. The arrangement ofthis complex, by mechanical factors arising during the development of the osculartube, results in the orientation of the optic axis of the developing spicule. The hypo-thesis is in accordance with the observations of Minchin on spicule formation.
CONTENTSPAGE
I N T R O D U C T I O N . . . . . . . . . . . . . 3 3M E T H O D S . . . . . . . . . . . . . 3 4
D e t e r m i n a t i o n o f t h e d i r e c t i o n o f t h e o p t i c a x i s . . . . . . . 3 4S e c t i o n s . . . . . . . . . . . . . 3 5
I s o l a t e d s p i c u l e s . . . . . . . . . . . . 3 6R E S U L T S 3 6
A p p e a r a n c e o f s p i c u l e s i n s u r f a c e - v i e w . . . . . . . . 3 6O p t i c a n g l e s o f t r i - a n d q u a d r i - r a d i a t e s . . . . . . . - 3 7C u r v e d m o n a x o n s . . . . . . . . . . . . 4 1S l e n d e r m o n a x o n s . . . . . . . . . . . 4 2
D I S C U S S I O N 4 4
M e c h a n i s m o f o r i e n t a t i o n o f t h e o p t i c a x i s . . . . . . . 4 4R E F E R E N C E S . . . . . . . . . . . . . 4 7
INTRODUCTION
THE optical properties of calcareous spicules have been known for manyyears as a result of the work of Sollas (1885) and von Ebner (1887). These
authors independently observed that the spicules contain calcite, which givesthem birefringent properties revealed by the use of the polarizing microscope.[Quarterly Journal of Microscopical Science, Vol 95, part 1, pp. 33-48, March 1954.]
£421.1 D

34 Jones—The Optic Axis of Spicules of Leucosolenia
The spicules in fact behave optically as though chiselled out from a singlehomogeneous crystal, and von Ebner was able to classify the forms of spiculesof different species in terms of the relations of the spicule rays to the principalcrystallographic axes of this crystal. Each spicule thus possesses an optic axis(the trigonal axis of symmetry of the calcite crystal), which has been shown forspecies of Sycon to be oriented with respect to the sponge substance by vonEbner (1887) and Bidder (1898). For example, the optic axis of the triradiatesis nearly parallel to the axis of the radial tubes of Sycon raphanus, except attheir free conical ends, where it is nearly perpendicular to the surface (Bidder).
The structure of the homocoelous ascon sponges is a great deal simpler thanthat of the heterocoelous Calcarea because their oscular tubes are compara-tively free of diverticula, and for this reason species of Leucosolenia have beenused by the author for the investigation of the mechanism of spicule orienta-tion and the factors controlling spicule form, accounts of which will bepublished in due course. A study of the orientation of the optic axis of thespicules is an essential part of this research and the results are presented inthis paper.
The spicules occurring in Leucosolenia complicata (Minchin) have alreadybeen described by Minchin (1904), as seen when lying in isolation upon aslide. There are four main types: quadriradiates, triradiates, curved mon-axons, and slender monaxons. The triradiates are essentially similar to thequadriradiates but lack the fourth, or 'gastral', ray that projects into thespongocoel; the other three rays are embedded in the mesogloea of the tubewall and comprise a 'basal1 ray, pointing to the base of the oscular tube, andpaired, or 'oscular' (previously termed 'oral') rays diverging from the oscularend of the basal ray. The monaxons are single-rayed, some being stout, curvedand fitted with lance-heads, others similar, but lacking the lance-heads, andthe remainder, which are numerous, being straight, slender, and sharplypointed. Fig. 7 summarizes the orientation of these spicules (see p. 43).
METHODS
Determination of the direction of the optic axisWhen a spicule is mounted in balsam and rotated between crossed nicols,
the spicule alternately glows brilliantly and darkens completely four times in acomplete revolution of 360°, provided the optic axis of the calcite does notcoincide with the microscope axis. The darkening occurs when the optic axislies in the plane of vibration of the polarized light, or in the vertical plane atright angles to this, and the two planes are easily distinguishable by removingthe analyser nicol: the spicule then appears faint when the optic axis lies in thefirst plane, but very distinct when in the plane at right angles. This arisesfrom the greater difference between the refractive indices of calcite and balsamwhen only the ordinary ray (i.e. the ray that always vibrates at right angles tothe optic axis) is being transmitted through the spicule. When the optic axiscoincides with the microscope axis, these phenomena are not apparent, for

Jones—The Optic Axis of Spicules of Leucosolenia 35
both ordinary and extraordinary rays travel along the optic axis at the samevelocity, and the refractive index of the calcite becomes that for the ordinaryray. In these circumstances the spicule remains invisible at all settingsbetween crossed nicols, and there is no change in distinctness on rotation,when the analyser nicol is withdrawn.
By such means it can be shown that the optic axis of the tri- and quadri-radiates of L. complicata lies in the plane containing the basal ray and bisectingthe angle between the oscular rays, as shown by von Ebner and Sollas for otherspecies. The angle between the optic axis and the basal ray can thus be deter-mined by examining the spicule in side-view, when this plane is perpendicularto the line of vision, with a polarizing microscope. The angle taken, hence-forth termed the 'optic angle', is defined for tri- and quadri-radiates as thesmaller angle of inclination of the optic axis to the line passing through thecentral points at either end of the basal ray (fig. 5, A; see p. 41). This anglewas measured to an accuracy of within ± f ° , three readings on either side ofthe appropriate plane of complete extinction of each spicule being made andthe average taken.
Sections
The spicules can be examined in side-view and in position in the tube wallby cutting thick longitudinal sections that lie in or near an axial plane of thetube, i.e. a plane including the tube's central axis. Sections of 30 fj, thicknesswere found most convenient, for thinner sections give too many broken rays,while spicule overlap obscures the optical phenomena to a great extent inthicker sections. Since even in 30 \i sections the oscular rays are bound to bebroken (their tips are nearly 200 \x apart), it is essential to have the spiculesfirmly embedded, and the Peterfi celloidin-paraffin method was found to besatisfactory. The material used must of course be uncontracted, necessitat-ing its immediate fixation after collection. A neutral 5 per cent, solution offormaldehyde in sea-water was employed as fixative and the specimens werestored in 90 per cent, alcohol to prevent spicule corrosion. After dewaxing,the unstained sections were mounted in balsam or D.P.X.
Only unbroken basal rays which were in focus all along their lengths wereselected for the measurements of optic angle, and in the case of the quadri-radiates there was the additional condition that the gastral ray had also to beunbroken and in the same focal plane as the basal ray. In this way it wasensured that the optic axis lay in a plane perpendicular to the axis of themicroscope.
To obtain some idea of the form of the spicules occurring in the tube sec-tioned, part of the material was left uncut and, after dewaxing, temporarilymounted, choanoderm uppermost, in 90 per cent, alcohol. By suitably tiltingthe piece of wall and using a modified camera lucida (see p. 36), spicules fromdifferent levels could then be drawn in 'surface-view aspect', i.e. with the basalray perpendicular to the line of sight and the two oscular ray tips in the samefocal plane of the microscope. Since all parts of the spicule were brought into

36 Jones—The Optic Axis of Spicules of Leucosolenia
focus before being drawn, the drawing represents the form of the spicule asprojected into a plane perpendicular to the line of vision, and such drawingsenable one to identify approximately the original position in the tube of subse-quently isolated spicules, because the spicule form varies continuously withthe distance from the oscular edge. Furthermore they enable the optic anglesmeasured to be correlated with the shapes of the spicules as seen in surfaceview.
Isolated spicules
Spicules were isolated from several distinct pieces of wall by boiling for afew minutes with 5 per cent, potash solution, washing several times in distilledwater with the help of a centrifuge, and then suspending the spicules in abso-lute alcohol and allowing a drop of the suspension to spread and dry on aslide. Selected specimens were then picked up under a i^-inch objective uponthe tips (50 JU. thick) of finely drawn glass needles which had been lightlysmeared with balsam, and the needles were mounted on a slide by means ofa piece of plasticine. The spicules must of course be taken up in a suitableorientation to be of use, and the basal ray affords a convenient handle, sinceit enables the spicule to be set for both side- and surface-views if it is in linewith the needle. The surface-view aspects of the spicules could thus be drawnas before, while their optic axes could be determined after setting them forside-view observation. Confirmation of the measurements of optic anglederived from the sections, and of the correlation between optic angle andspicule form, was thereby obtained.
A modified camera lucida was employed for the drawings, to avoid changesin magnification as the microscope tube was racked up and down. Themirror of the manufactured article was removed and replaced by a large planemirror which was clamped at an angle of 45° to the bench by means of a retortstand. Since the mirror was not attached to the microscope, nor moved rela-tively to it, the picture was always projected the same distance and hence themagnification factor was constant. This is important because excessive rack-ing was often necessary to bring the isolated spicules, perched on their needles,into focus in the required orientation. The arrangement must of course bedistortionless and this was checked by drawing part of a squared eyepiecemicrometer with the apparatus and then measuring the sides and angles of thedrawing. A X 40 objective and a X 10 eyepiece gave the most convenientmagnification.
RESULTS
Appearance of spicules in surface-viewIn fig. 1 superimposed drawings of the surface-view aspects of tri- and
quadri-radiates derived from successive levels of two oscular tubes are de-picted. It is obvious at a glance that the form and size vary continuously fromthe oscular edge to the base of the tube, and such an arrangement enables oneto determine approximately the original position of subsequently isolated

Jones—The Optic Axis of Spicules of Leucosolenia 37
spicules. However, only those spicules that are in contact with the choano-derm give such a consistent picture; if 'outer' as well as these 'inner' spiculesare included, the arrangement is not quite so perfect. The reason is that thespicules grow in contact with the choanoderm (Minchin, 1908) and leave it onlywhen fully grown, so that only the inner spicules are of similar ages and formedunder a given set of conditions; the outer spicules were formed earlier, whenthe dimensions and possibly the rate of growth of the tube were different.
FIG. 1. Superimposed surface-view drawings of inner tri- and quadri-radiates of two osculartubes, A and B, derived from different specimens of L. complicata. For each drawing theoscular rays are symmetrically disposed on each side of the mid-line. The distances of thespicules from the oscular edge are indicated. Note the dependence of the oscular ray curvature
on the distance.
Optic angles of tri- and quadri-radiates
When the optic angles of tri- and quadri-radiates are plotted against thedistance of the spicules from the oscular edge, the points form the patternshown in fig. 2. At the edge itself high readings are obtained, but at a dis-tance of only 200 p. behind they are several degrees lower, and thereafterdecline gradually towards the base of the tube. The results from two tubes(A and E) are given to show the variation. The tubes were taken from speci-mens collected at Plymouth in different years and while the patterns of pointsare similar, that of tube B is about 30 lower than that of tube A, and a checkof some of the measurements has shown that the difference is not due to asystematic error of measurement. Three other tubes from the first specimengave similar points to those of tube A, though their patterns are less well-defined because fewer readings could be taken; the number of spicules whichsatisfy the conditions for measurement depends on the orientation of the

30
-
z&
o s3
% &
oo 4
0 »
a °k
•
1
i o°0
1
e • •
° *o
0 • k
oe
\ •
*
0
I
o
.
• o
S»e
o
• .
«o
•
I
0 0
o e
1
o0
o•o
t0
12
3 4
5 6
7di
stan
ce
From
the
osc
ular
edg
e (m
m)
FIG
. 2.
Gra
phs
show
ing
the
rela
tion
betw
een
the
opti c
ang
le a
nd t
he d
ista
nce
from
th
e os
cula
r ed
ge f
or t
ri-
and
quad
ri-r
adia
tes
of t
ube
A(a
bove
) an
d tu
be B
(be
low
), as
det
erm
ined
by
usin
g lo
ngitu
dina
l se
ctio
ns o
f th
e os
cula
r tu
be.
Inne
r qu
adri
radi
ates
, O
; qu
adri
radi
ates
with
bas
alra
ys le
ss th
an 7
5 /*
, 0;
oute
r qu
adri
radi
ates
, •;
inne
r tr
irad
iate
s, A
; tr
irad
iate
s w
ith b
asal
ray
s le
ss t
han
75 /*
, A
; o
uter
tri
radi
ates
, A
.

Jones—The Optic Axis of Spicules of Leucosolenia 39
material for sectioning, and even with straight tubes it is not always possibleto orient the sectioning as accurately as desired. However, measurements madewith isolated spicules confirm the pattern, as indicated, for example, by thegraph in fig. 3 (tube C). This tube was derived from the same specimen astube A.
35-
3O?
ena 25-
20-
0-5 10distance from oscular edge (mm.)
1-5
FIG. 3. Graph showing the relation between the optic angle and the approximate distancefrom the oscular edge for tri- and quadri-radiates of tube C, as determined by using isolated
spicules. Quadriradiates, O; triradiates, A.
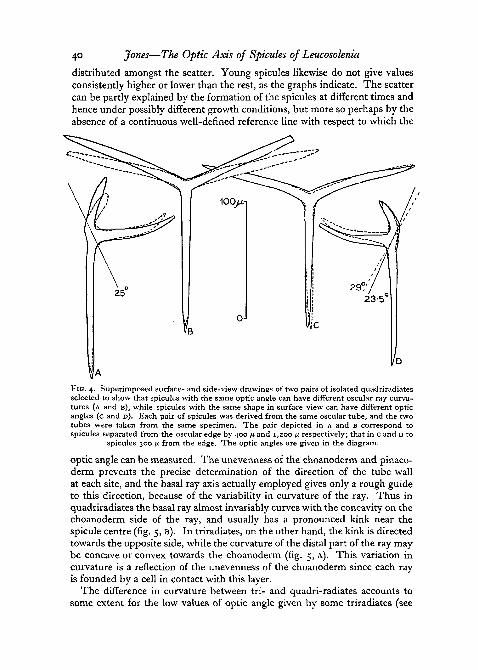
It might have been expected that the surface-view drawings of the spiculesof tubes A and B would reflect the difference between the two patterns ofpoints, but this is not the case (fig. 1). The drawings are quite comparable,the only important difference being one of size; the spicules of tube B aresmaller than those of tube A, a fact to be correlated with the smaller girth ofthe former tube. This was approximately 750 ju. in diameter in the middle,whereas tube A, which was elliptical in cross-section, had large and smalldiameters of 1,700 fx, and 750 /x respectively in the same region. The similarityof the surface-view aspects and the opposing dissimilarity of the optic anglesof tubes A and B reveal that the optic angle cannot be the sole factor influen-cing the curvature of the oscular rays, and this is supported by the fact that theform changes markedly over the first 1,500 fx, whereas the optic angle hasreached a comparatively steady level after only 200 /x. Furthermore, the studyof isolated spicules has shown that two spicules with the same surface-viewappearance can have different optic angles, while two with the same angle canhave different shapes as seen in surface-view (fig. 4). The optic angle is hencenot closely correlated with the curvature of the oscular rays.
A considerable amount of scatter occurs on the graphs, which cannot beexplained by errors of measurement (made to within ±i°) , or by the fact thatsome 'outer' spicules were measured, since these spicules give points evenly
40 Jones—The Optic Axis of Spicules of Leucosolenia
distributed amongst the scatter. Young spicules likewise do not give valuesconsistently higher or lower than the rest, as the graphs indicate. The scattercan be partly explained by the formation of the spicules at different times andhence under possibly different growth conditions, but more so perhaps by theabsence of a continuous well-defined reference line with respect to which the
FIG. 4. Superimposed surface- and side-view drawings of two pairs of isolated quadriradiatesselected to show that spicules with the same optic angle can have different oscular ray curva-tures (A and B), while spicules with the same shape in surface-view can have different opticangles (c and D). Each pair of spicules was derived from the same oscular tube, and the twotubes were taken from the same specimen. The pair depicted in A and B correspond tospicules separated from the oscular edge by 400 fi and 1,200 p respectively; that in c and D to
spicules 300 ft from the edge. The optic angles are given in the diagram.
optic angle can be measured. The unevenness of the choanoderm and pinaco-derm prevents the precise determination of the direction of the tube wallat each site, and the basal ray axis actually employed gives only a rough guideto this direction, because of the variability in curvature of the ray. Thus inquadriradiates the basal ray almost invariably curves with the concavity on thechoanoderm side of the ray, and usually has a pronounced kink near thespicule centre (fig. 5, B). In triradiates, on the other hand, the kink is directedtowards the opposite side, while the curvature of the distal part of the ray maybe concave or convex towards the choanoderm (fig. 5, A). This variation incurvature is a reflection of the unevenness of the choanoderm since each rayis founded by a cell in contact with this layer.
The difference in curvature between tri- and quadri-radiates accounts tosome extent for the low values of optic angle given by some triradiates (see

Jones—The Optic Axis of Spicules of Leucosolenia 41
figs. 2 and 3), for their basal ray axes do not coincide with the inner (gastral)edge of the distal seven-eighths of the ray, which would give a better indica-tion of the direction of the tube wall; and their angles would be increased by1 or 2 degrees if this edge were taken as the reference line. In the quadri-radiates the corresponding edge does in fact coincide in most cases with thebasal ray axis used. Thus in fig. 5, B the side aspects of a triradiate and a quadri-radiate from tube C having the same surface-view appearance (corresponding
FIG. 5. A, superimposed side-view drawings of a tri- and quadri-radiate having the sameoptic angle. The difference in curvature of the basal rays is typical, oa, optic axis; br, basalray axis; triradiate in broken outline. B, side-view drawings of a tri- and quadri-radiate super-imposed with the gastral surfaces of the basal rays coinciding. The optic axes are parallel, butthe measured optic angles differ by two degrees. The low values of optic angle for some of thetriradiates plotted on figs. 2 and 3 would likewise be increased if measured with respect to thegastral surface of the basal ray, which gives a better indication of the direction of the choano-
derm. oa, optic axis; triradiate in broken outline.
to a position of 1,200 /x from the oscular edge) are superimposed so that thegastral surfaces of the basal rays coincide; their optic axes have becomeparallel, but the measured optic angles were respectively 23 ° and 25° (as maybe seen in fig. 3). It thus seems likely that the optic axes of tri- and quadri-radiates of similar age and situation are alike and that the optic axis is directlyrelated to the direction of the tube wall.
There is a possible difference between tri- and quadri-radiates, however, inthat values for the former have never exceeded 2 8°, even when the angle ismeasured against the gastral edge of the basal ray; but these spicules are un-common within 20011 of the oscular edge (only three have been measured fromthis region) and possibly a sufficient number has not been examined.
Curved monaxons
Lance-headed monaxons are stout, uni-rayed spicules which curve in asingle plane containing the optic axis, as Sollas and von Ebner showed for

42 Jones—The Optic Axis of Spicules of Leucosolenia
various species. Because of the curvature, the optic axis makes a continuouslyvarying angle with the direction of the ray, but at the lance-head (the firstformed part (Minchin, 1908)), it is directed at right angles (at least to within50) to the axis of the tip. Fig. 6 shows some examples drawn from different
distance From oscular edge (mm.)2-3 30 3-1 3-3 4-0 40 44 5-0
FIG. 6. Drawings of lance-headed monaxons belonging to the same oscular tube, showing thatthe curvature is independent of the distance from the oscular edge (indicated by the figures)
and that the optic axis (oa) is nearly perpendicular to the axis of the lance-head.
parts of an oscular tube. The curvature is independent of the distance fromthe oscular edge.
The curved monaxons which lack the lance-heads, or have them poorlydeveloped, have optic axes corresponding to those of the lance-headed type.
Curved monaxons are oriented with their plane of curvature in an axialplane of the oscular tube and their free ends pointing away from the tubebase (fig. 7).
Slender monaxons
The slender monaxons are straight, pointed at each end and slender, usuallywith a slight thickening towards the embedded end. At and near the oscular

Jones—The Optic Axis of Spicules of Leucosolenia 43
edge they are oriented to form a crown, but over the greater part of the tubethey are haphazardly directed outwards. Possibly they tend to grow perpen-dicularly to the surface, as Urban (1906) describes for L. eleanor, and aresubsequently disturbed by contractions (or fixation) of the tube. For syconoidspecies Woodland (1905) mentions that the monaxons are irregular below theoscular rim, whereas Bidder (1898) states that the slender monaxons are
-SURFACE VIEW •
FIG. 7. Diagram summarizing the orientation of the spicules and their optic axes (shown asbroken lines) with respect to an oscular tube of L. complicate; sm, slender monaxon; cm,curved monaxon without well-defined lance-head; Im, curved monaxon with lance-head;
g, quadriradiate; *, triradiate; L.S., longitudinal section.
perpendicular to the surface. Much would thus seem to depend on theobserver and the conditions under which the material has developed or beenfixed.
In the slender monaxons of L. complicate/, the optic axis coincides exactlywith the axis of the spicule, and Bidder (1898) records a similar observationfor the slender monaxons of Sycon raphanus. Von Ebner's statement (1887)that in all monaxons the optic axis is inclined to the spicule axis is thusincorrect.
The diagram in fig. 7 summarizes the orientation of the spicules and theiroptic axes.

44 Jones—The Optic Axis of Spicules of Leucosolenia
DISCUSSION
Before discussing the mechanism whereby the optic axes become oriented,a brief account of spicule development and structure are necessary. Accord-ing to Minchin (1908, 1909), each spicule ray (monaxons included) issecreted by a pair of cells, which probably arise by the division of a calcoblastmother-cell. The basal and oscular rays of tri- and quadri-radiates are thusformed by a sextet of cells, of which three are in close contact with the choano-derm. Spicule formation begins with the secretion of three non-birefringentrodlets, each between a pair of cells, which join up centrally to form a three-rayed primordium upon which calcite is subsequently deposited. The innercell of each ,pair then acts as 'founder', secreting material at the tip of thedeveloping ray, while the outer one only later passes along the ray, thickeningit as it proceeds. For monaxons the development is essentially similar to thatof a ray of a triradiate.
As regards spicule structure, Minchin and Reid (1908) have shown that thespicules of L. complicate/, (and other species) contain along the centre of eachray a stainable axial filament, which remains when the calcite has been cor-roded away. This settles the old controversy, summarized by Minchin (1909),between von Ebner, who concluded that the filament was an optical illusioncaused by the presence of a central strand of less pure calcite, and Sollas (forexample), who believed in the presence of organic matter, largely because thespicule chars internally when ignited. Further evidence is afforded by Maas(1904), who decided that an organic network permeated the whole spicule sub-stance, because concentrated potash solution breaks up the spicule into minutecalcite rhombohedra, which he believed were liberated by the dissolution ofthe organic matter holding them together. He also found (1906) that whensyconoid sponge larvae were grown in sea-water containing no lime, calcareousspicules were not formed but were replaced by organic 'spiculoids', secreted bythe same type of cell in approximately the same triradiate or needle-like form.Such spiculoids suggested an over-secretion of the organic matter of the normalspicule in the absence of calcium carbonate. All these facts support the conclu-sion that organic material is present.
Mechanism of orientation of the optic axis
In Leucosolenia [ = Clathrina] coriacea the sextet of formative calcoblastshas a regular arrangement (Minchin 1898), three of the cells being exactlysuperimposed on the other trio in contact with the choanoderm. The tri-radiates formed by these sextets are 'perregular' (von Ebner, 1887), i.e. haveequal rays meeting centrally to form three equal angles and an optic axis setperpendicular to the plane of the three ray tips; and since the optic axis isalmost at right angles to each ray and one ray points to the base of the tube(as in L. complicata), it follows that the optic axis is very nearly perpendicularto the surface of the tube. The optic axis thus coincides closely with thetrigonal axis of symmetry of the sextet (fig. 8, A and B).

Jones—The Optic Axis of Spicules of Leucosolenia 45
In L. complicata, on the other hand, the arrangement of the calcoblasts isdescribed as irregular by Minchin (1908), but a close examination of Minchin'sown drawings in this paper indicates that in four sextets out of the five por-trayed (plate 17, nos. 19, 20, 21; plate 20, no. 90) the outer three cells are dis-placed forwards (i.e. towards the osculum) with respect to the inner trio, asshown by the orientation of the contained spicule primordium. Such a dis-placement would have tilted the axis of the sextet forwards (see fig. 8, E and F)
FIG. 8. Diagrams derived partly from Minchin's drawings of spicule development showingthe tendency for the optic axis of the first-formed calcite rudiment to lie in the direction ofcell division (or cell association), A, B, surface- and side-views respectively of a sextet ofLeucosolenia [= Clathrina] coriacea; E, F, the same for L. complicata; C, early stage in theformation of a lance-headed monaxon; D, the same for a slender monaxon. The optic axis is
indicated by the broken line.
and if once again the optic axis of the calcite coincided with this axis, the opticaxis would have the orientation found in the developing tri- and quadri-radiates of this species. Minchin's own description of the arrangement asirregular could have arisen from disturbance in some cases during the processof fixation, or more probably, from the lack of sufficient time for some youngsextets to become oriented before fixation. The presence of gastral ray calco-blasts beneath many of the sextets may also have confused the picture to someextent.
For the monaxons the optic axis also tends to coincide with the long axisof the cell pair (fig. 8, c and D). This is not so evident in the one case depictedby Minchin for the lance-headed monaxon, but the calcite mass is quite largein this case, and the arrangement of the cells suggests that at an earlier stagein development the tendency would be more marked. A law can thus betentatively stated: for spicules of Leucosolenia coriacea and L. complicata the

46 Jones—The Optic Axis of Spicules of Leucosolenia
optic axis of the first formed calcite rudiment lies in the direction of the celldivision (or cell association) that directly precedes its formation.
Two outstanding problems remain, namely, how does the optic axis cometo coincide with the axis of the sextet, and how is the outer trio displacedforwards relative to the inner trio in L. complicata} As regards the latter,experimental evidence in support of a hypothesis of 'jelly-shear' has beenobtained by the author (Jones, 1952) and will be published shortly. Briefly,the sextet is anchored to the inner cell-layer by the inner trio, and is orientedand tilted forwards by the slow shearing creep of the viscous mesogloeal jellypast it, arising from the secretion of the jelly in the more basal feeding partsof the tube (the choanocyte zone) and the extension by growth of the innercell-layer at mainly the oscular end of the tube. The variation in the opticangle with position in the tube is the result of the different amount of shearingat each site, depending on the relative rates of growth in the different regions,while the difference between L. coriacea and L. complicata sextets is accountedfor by the much slower growth of the oscular tubes of the former as comparedwith the latter (Urban, 1906); the shear would hardly be appreciable duringthe early stages of spicule development after the formation of the sextet inL. coriacea.
The problem of the coincidence of the optic and trigonal sextet axes canlargely be solved by considering the organic constituent of the spicule.Minchin (1898) observed that the first lump of calcite is deposited upon thenon-birefringent rudiment, later identified with the axial filament, and it seemsplausible to assume that the calcite ions are chemically bonded to those of theprecursor. The orientation of the calcite optic axis would then depend on thecrystalline structure of the precursor and the way this is oriented in the cell-sextet, and there seems no reason why it should not be appropriately orientedin the direction of the prior cell division, particularly as the mitotic spindleand the presumed cell membrane between the daughter cells are orientedstructures capable perhaps of functioning as precursors. Oriented over-growth is a well-established process. Thus Schmidt (19326) gives a diagramof oriented crystals of calcium iodide grown on a mica plate and remarks thatcrystalline dye particles can be oriented by the 'righting power' of the micellaeon which they crystallize. Oriented crystals of (organic) urea can also form onthe cubic crystals of (inorganic) ammonium chloride.
Without the concept of oriented overgrowth it is difficult to understandhow the first-formed calcite primordium is oriented and held in the requisiteorientation. Schmidt (1932a) has given a brief review of the relation betweenthe optic axis and spicule form, and points out that mechanical orientingagencies can act solely by virtue of the form and are not concerned with theoptical properties of the material, while, on the other hand, though an electro-magnetic field could orient the optic axis, it would not directly influence theform. Hence Schmidt comes to the view that there must be a relation betweenthe form and the optic axis, depending on a tendency for the mineral to crystal-lize more easily in certain directions. Thus within the calcoblast (he refers

Jones—The Optic Axis of Spicules of Leucosolenia 47
here to echinoderm spicules) the growth of calcite apparently goes on mosteasily in a direction perpendicular to the optic axis, and the branching of therays at 1200 is also a consequence of the properties of the crystallizing sub-stance ; should, therefore, mechanical conditions set the spicule, the optic axiswould likewise become oriented, or should the optic axis be directly oriented,so would become the form also.
Now the assumption that the crystallizing activity of calcite determines therelation between the optic axis and the spicule shape cannot apply to spongespicules, for, as shown above, the variation in spicule form is not closely corre-lated with the variation in optic angle. The basal ray of L. complicata developsprecociously (Minchin, 1908) and could perhaps be oriented by mechanicalfactors, but the angle which the optic axis makes with this varies continuouslyfrom about 18° to 350, and hence surely cannot be determined by the processof crystallization alone, since this generally involves development in the direc-tion of particular crystal faces or corners. To assume that slight variations inthe composition of the solution bathing the ray tip are responsible for suchslight differences in the direction of growth relative to the optic axis seemsuntenable, especially as the solution would not only have to vary with the posi-tion of the sextet in the oscular tube, but also during the development of thespicule itself, for the basal ray is often kinked.
The alternative suggestion, that an electromagnetic field orients the crystalprimordium, is likewise incredible, because, while bio-electric gradients arebelieved to exist in species of Leucosolenia (Hyman and Bellamy, 1922), theydo not nearly approach the large field strengths required for the orientationof crystals of polaroid, say, or for even the imperfect orientation of moleculesof glass or certain liquids that can be obtained by using a Kerr cell.
It may be concluded then that the optic axis is oriented, not by the bio-electric field or the crystallizing activity of calcite, but by a process of orientedovergrowth upon an organic precursor which is structurally bound andoriented within the formative cell complex. The cells and precursor can per-haps be regarded as a single complex organic crystal within which the calciteis chemically bonded so that its optic axis lies in the direction of the cell divi-sion (or cell association) preceding its secretion; the orientation of the opticaxis relative to the sponge wall then arises from the orientation by mechanicalprocesses of the formative cell complex.
I wish to thank Professor James Gray, F.R.S., for guidance and advice, andthe Staff of the Department of Zoology, Cambridge, particularly Dr. C. F. A.Pantin, F.R.S., and Dr. L. E. R. Picken, for friendly help and encouragement.I am also much indebted to Professor F. W. Rogers Brambell, F.R.S., andthe Staff of the Zoological Department, University College of North Wales,Bangor, for affording me the facilities to complete this work.
REFERENCESBIDDER, G. P., 1898. Proc. roy. Soc, 64, 61.EBNEH, V. v., 1887. S. B. Akad. Wiss. Wien, 95, 55.

48 Jones—The Optic Axis of Spicules of Leucosolenia
HYMAN, L. H., and BELLAMY, A. W.( 1922. Biol. Bull. Wood's Hole, 43, 313.JONES, W. C , 1952. Ph.D. thesis, Cambridge.MAAS, O., 1904. Verh. dtsch. zool. Ges., 14, 190.
1906. Arch. Entw. Mech. Org., 22, 581.MINCHIN, E. A., 1898. Quart. J. micr. Sci., 40, 469.
1904. Proc. zool. Soc. Lond., 2, 349.1908. Quart. J. micr. Sci., 52, 301.1909. Ergebn. Zool., a, 171.
MINCHIN, E. A., and REID, D. j . , 1908. Proc. zool. Soc. Lond., 661.SCHMIDT, W. J., 1932a. Handb. biol. ArbMeth., Abt. V, Teil 2/H, 1357-
19326. Ibid., p. 1835.SOLLAS, W. J., 1885. Sci. Proc. R. Dublin Soc, 4, 374.URBAN, F., 1906. Arch. Naturgesch., 1, 33.WOODLAND, W., 1905. Quart. J. micr. Sci., 49, 231.





























