Embed Size (px)
Citation preview

Neuron, Volume 98
Supplemental Information
The Organization of Projections from Olfactory
Glomeruli onto Higher-Order Neurons
James M. Jeanne, Mehmet Fisxek, and Rachel I. Wilson

0 17spike count
neur
opil
PN
spi
kes
PN
dye
fill
neur
opil
PN
spi
kes
PN
dye
fill
slice 1 (top) slice 2 slice 3 slice 4 slice 5 slice 6
slice 7 slice 8 slice 9 slice 10 slice 11 slice 12 (bottom)
Figure S1: Full response map for example PN. (Related to Figure 1)
The full set of 12 optical slices through the antennal lobe for the example PN shown in Figure 1B-F. Glomerular boundaries are in red with the home glomerulus boundary in yellow. The dye-filled pipette is visible in slices 1-3, and the dye-filled cell body can be seen in slices 1-5.

PN LHN
5 mV0.5 sec
photostimulation
“bleed through” glomeruli (false positives)
**
0 35PN spike count
0 50 12∆ LHN voltage (mV)
0 7.5
PN LHN
“isolated core” glomeruli (false negatives)A B C D
PN spike count ∆ LHN voltage (mV)
neuropil neuropil neuropil neuropil
LHN responses toisolated core voxels
Figure S2: Glomerular hit criteria. (Related to Figures 1 and 2)A) Example of a “bleed through” glomerulus in a PN recording. The PN’s home glomerulus is outlined in yellow and contains many positive voxels (voxels eliciting a PN spike). An adjacent glomerulus also contains several positive voxels. However, note that in the home glomerulus, the center of mass of positive voxels is near the glomerular center, whereas in the adjacent glomerulus, the positive voxels are skewed toward the home glomer-ulus. In the adjacent (red) glomerulus, the positive voxels comprise ≥25% of all voxels, and so this glomerulus would normally be scored as a hit. Because it is a clear case of bleed through, we removed the red glomerulus from the list of glomerular hits in this PN experiment.B) Example of a “bleed through” glomerulus in an LHN recording. In the yellow glomerulus (deemed a true positive), the center of mass of positive voxels is well within the center of the glomerulus. In the red glomerulus (deemed a false positive), the center of mass of positive voxels is skewed towards the yellow glomerulus. Because this was a likely case of bleed through, we removed the red glomerulus from the list of glomerular hits in this experiment. C) Example of an “isolated core” glomerulus in a PN recording. The PN’s home glomerulus (yellow) contains several positive voxels situated near its core, but they comprise < 25% of all voxels (the threshold for calling a glomerulus a hit). Because this was a clear case of an isolated core response, we added this glomerulus to the list of hits in this experiment.D) Example of an “isolated core” glomerulus in an LHN recording. The yellow glomerulus contains two positive voxels situated near its core, but they comprise < 25% of all voxels. The voltage traces for the two positive voxels in the yellow glomerulus (bottom) show clear EPSPs, but the EPSP frequency is relatively low, indicating a low presynaptic PN firing rate (similar to what we observe in a PN in cases where only the voxels near the core are generating any spikes, like the case in C). Because this was a likely case of a true isolated core response, we added this glomerulus to the list of hits in this experiment. In this case, the red outline denotes a second glomerular hit.
VA1d VA1dVA1v
VA6
VA1d
VA1vVA6
VA6VA1v DC2
DC2VA1v
VA1vDC2
0 7.5∆ LHN voltage (mV)
slice 1 (top) slice 2 slice 3 slice 4 slice 5 slice 6
slice 7 slice 8 slice 9 slice 10 slice 11 slice 12 slice 13 (bottom)
neur
opil
∆ LH
N v
olta
gene
urop
il∆
LHN
vol
tage
DC2
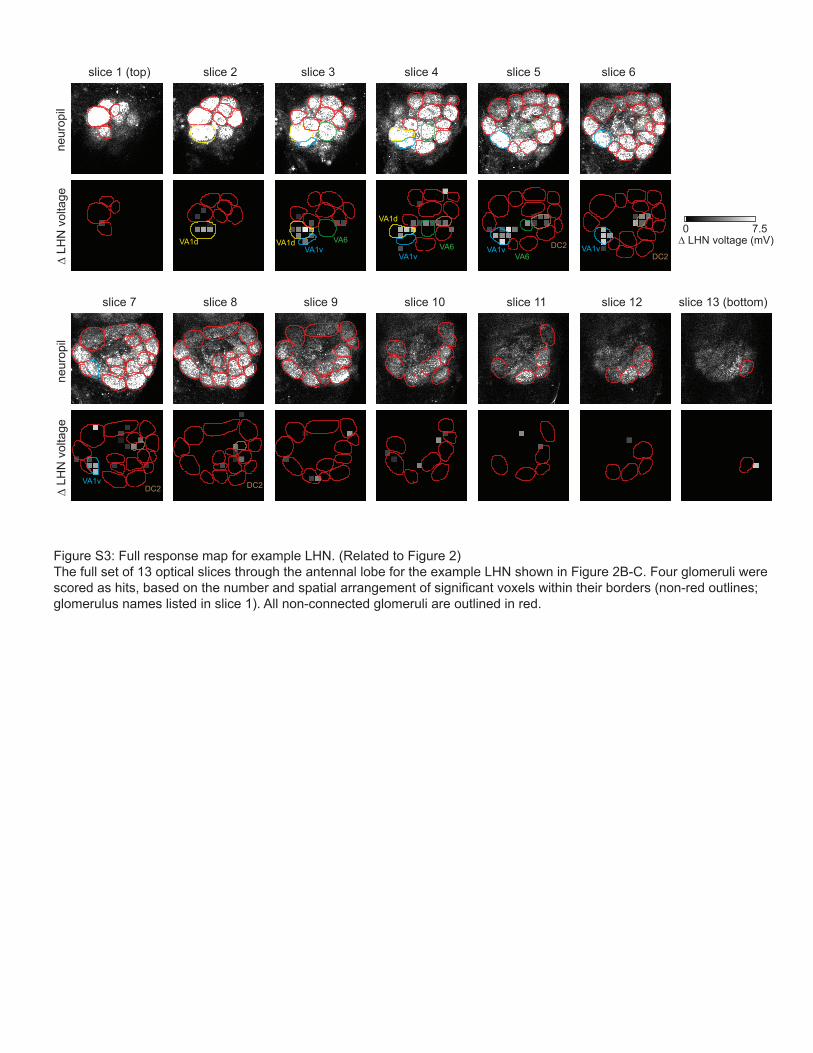
Figure S3: Full response map for example LHN. (Related to Figure 2)The full set of 13 optical slices through the antennal lobe for the example LHN shown in Figure 2B-C. Four glomeruli were scored as hits, based on the number and spatial arrangement of significant voxels within their borders (non-red outlines; glomerulus names listed in slice 1). All non-connected glomeruli are outlined in red.

-10 0 10
1st trial response (z-scored)
-10
0
10
2nd
trial
resp
onse
(z-s
core
d)
-10 0 10
1st trial response (z-scored)
-10
0
10
2nd
trial
resp
onse
(z-s
core
d)
repeated trials shuffledrepeated trials alignedB C
20 µm
30 mV800 msec
A first trialsecond trial
Figure S4: Optogenetic stimulation of voxels is reliable (Related to Figure 2)A) LHN responses to photostimulation of every voxel in an entire z-plane. Responses are displayed within the corre-sponding voxel, with glomerular areas shown in pale gray. After stimulating all voxels (across all z-planes) once, roughly half of the voxels were stimulated again. Voxels that were repeatedly stimulated showed similar responses in this LHN (compare red and black traces). Note that the large responses in the center (corresponding to photostimulation of glomerulus VA6) consist of large EPSPs that do not evoke spikes.B) Mean LHN voltage change on first and second trials for 23,464 voxels from the 53 LHNs with repeated stimulation. Responses for each LHN are reported as z-scores to facilitate comparisons across LHNs. Responses to the first stimula-tion were significantly correlated with responses to the second stimulation (r = 0.49, p < 10-15).C) Same but with voxel identity shuffled between first and second trials within each LHN.

VA6 regular
VL2a regular
D graded
DA1 graded
multi-PN glomeruli single-PN glomeruli
C
DM1 regular
VM4 regular
VA7l regular
VA1v regular
D
0
1
fract
ion
ofco
nnec
tions
A
D 4 0
VA1d 3 0
DA1 8 1+
VA1v 4 2+
DA2 5 0
VA2 1 0
DA3 0
VA3 2 0
DA4l 1 0
VA4 1 0
DC1 1 0
VA5 3 0
DC2 2 0
VA6 1 0
DC3 3.5 0
VA7l 2 0
DL1 2.5 0
VA7m 2.5 0
DL3 5 0
VC1 2 0
DL4 1 0
VC2 3 0
DL5 1 0DM1 1 0
VL2a 1 0
DM2 2 0
VL2p 1 0
DM3 1 0
VM1 2 0
DM4 1 0
VM2 2 0
DM5 2 0
VM3 2 0
DM6 3 0
VM4 1 0
DP1m 1 0
VM7d 3 0VM7v 3 0
0
1
fract
ion
ofco
nnec
tions
B
black = glomeruli with multiple cholinergic PNsblue = glomeruli with a single cholinergic PN
DA1 VA1v5 mV250 msec
chol
iner
gic
PN
sG
AB
Aer
gic
PN
s
glom
erul
i GABAergic PNs → LHNs
cholinergic PNs → LHNs
cholinergic PNs → Kenyon cells
multi-PN glomeruli single-PN glomeruli
regular graded regular graded
regular graded regular graded
VL1 0 1
Figure S5: Characteristics of LHN voltage responses to glomerular photostimulation. (Related to Figure 2)
A) PN numbers per glomerulus. PNs that do not express Gal4 under the control of GH146-Gal4 are not included. For two glomeruli (DA1 and VA1v), GH146+ PNs include both cholinergic PNs and GABAergic PNs. PN counts are taken primarily from Grabe et al. (2016), with additional data from other sources (see STAR Methods). B) Examples of inhibitory LHN responses. Horizontal bar indicates timing of photo-stimulation. We only observed inhibitory responses when we stimulated voxels in either DA1 or VA1v, indicating these are likely monosynaptic responses to GABAergic PN stimulation.C) Examples of excitatory LHN responses. Black: a multi-PN glomeruli typically evoke a graded or irregular responses, which may reflect many the summation of many small EPSPs occurring at irregular intervals. Blue: single-PN glomeruli typically evoke relatively large EPSPs at regular intervals.Bottom: fraction of glomerulus-LHN connections exhibiting regular EPSPs versus graded responses. The observation that graded responses are typical for multi-PN glomeruli is evidence that sister PNs from the same glomerulus form convergent connections onto LHNs, as a general rule.D) Same but for Kenyon cells. Kenyon cells typically receive input from at most one PN in a given glomerulus. All the Kenyon cell responses we observed consisted of regular EPSPs, supporting the conclusion that regular EPSPs arise from single unitary synaptic connections, whereas irregular responses are only observed in LHNs and thus likely arise from converging sister PNs.
LHN voltage responses were classified as regular versus graded by an observer who was blind to the location of the stimulated voxel. This observer was also blind to all information about the recorded LHN. For this classification exercise, voltage traces
were pooled across the entire data set and ordered randomly, so that responses from different LHNs were inter-leaved. A voltage response was classified as “regular" if EPSPs or IPSPs occurred at approximately equal intervals over an epoch lasting at least ~200 ms. Responses consisting of a single EPSP or IPSP were also classified as regular.

V1JJ94
V2JJ79
V3JJ15
local7JJ73
local1JJ102
local6JJ64
local3JJ139
local2JJ142
local4JJ92
local5JJ66
L13JJ107
L15JJ98
L14JJ81CML1
JJ55
CML2JJ60
L1JJ63
LV1JJ95
ML10JJ32
ML11JJ13
MLV1JJ41
L2JJ123
L3JJ82
L5JJ47
L4JJ119
L8JJ117
L9JJ42
L7JJ46
L6JJ83
ML9JJ8
ML8JJ11
ML3JJ133
ML2JJ37
ML4JJ25
ML5JJ27
ML6JJ127
ML7JJ20
50 μm
A
B
ML1JJ126
L11JJ44
L12JJ43
Figure S6: Morphologies of all LHN types identified in this study (Related to Figure 4)A) Morphologies of all 39 LHN types identified in this study, shown as z-projections of coronal stacks. The soma and the primary neurite are typically not retained in the brain after the recording, and so are not shown. Gray region denotes the neuropil boundary of the lateral horn. All exemplars of a given LHN type are superimposed. In each case; a single exemplar is highlighted in red (along with the associated JJ identifier to cross-reference the data in Supplemental Table S2 and Supplemental Data S1), and the remaining exemplars (if any) are in black. The following types resemble LHN types that have been described elsewhere: L12 (called NP6099 by Fişek & Wilson 2014), ML8 (called PD2b1 by Dolan et al. 2017a), ML9 (called Mz671 by Fişek and Wilson 2014 and PD2a1 by Dolan et al. 2017a), and ML2 (cell 121208-1 within the category called aSP-g by Kohl et al. 2013, i.e. the rightmost female LHN in Figure 2B of that study).B) All 89 LHN morphologies represented in the same brain, showing coverage of the entire LH neuropil. The outline of the LH in the opposite hemisphere is shown in red.
methyllaurate
ethylhexanoate
methylacetate
hexanoicacid
V2-LHN odor responses
1 sec odor
0
1
PN responses
∆F/F
(nor
mal
ized
to
glom
erul
ar m
ax)
VA1v
DM2DM1
methyllaurate
odorparaffin oil
A
B
1 sec odor
8 mV
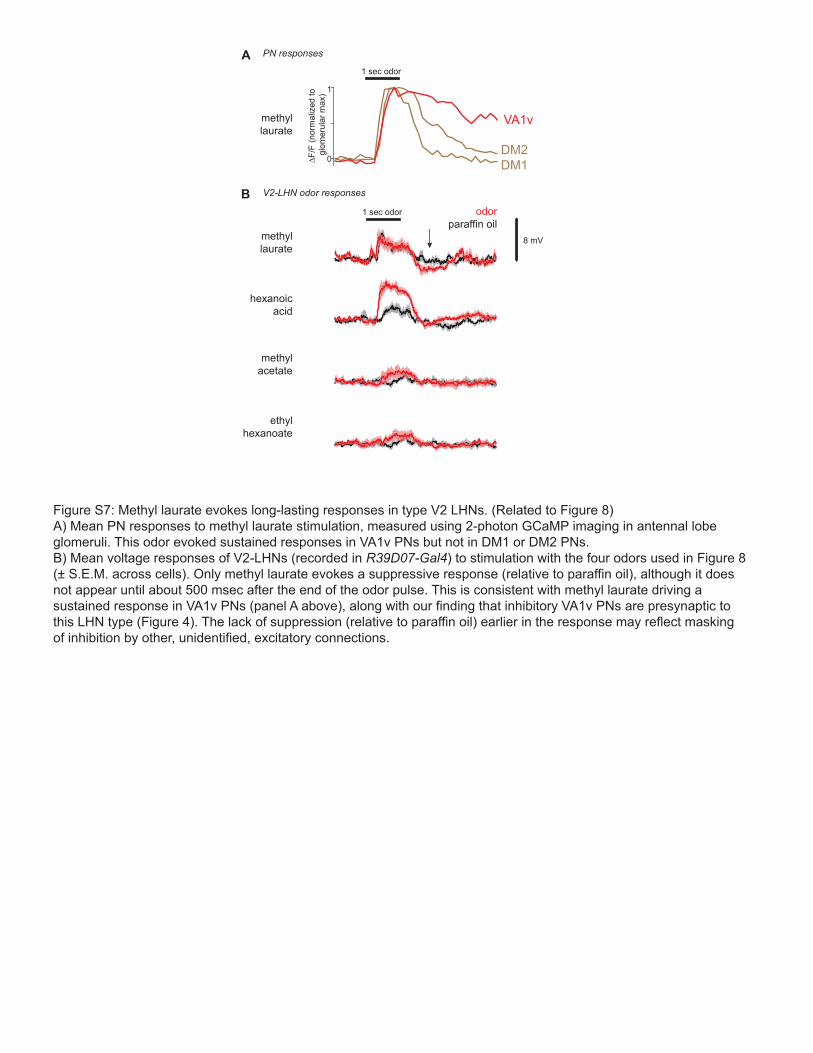
Figure S7: Methyl laurate evokes long-lasting responses in type V2 LHNs. (Related to Figure 8)A) Mean PN responses to methyl laurate stimulation, measured using 2-photon GCaMP imaging in antennal lobe glomeruli. This odor evoked sustained responses in VA1v PNs but not in DM1 or DM2 PNs. B) Mean voltage responses of V2-LHNs (recorded in R39D07-Gal4) to stimulation with the four odors used in Figure 8 (± S.E.M. across cells). Only methyl laurate evokes a suppressive response (relative to paraffin oil), although it does not appear until about 500 msec after the end of the odor pulse. This is consistent with methyl laurate driving a sustained response in VA1v PNs (panel A above), along with our finding that inhibitory VA1v PNs are presynaptic to this LHN type (Figure 4). The lack of suppression (relative to paraffin oil) earlier in the response may reflect masking of inhibition by other, unidentified, excitatory connections.
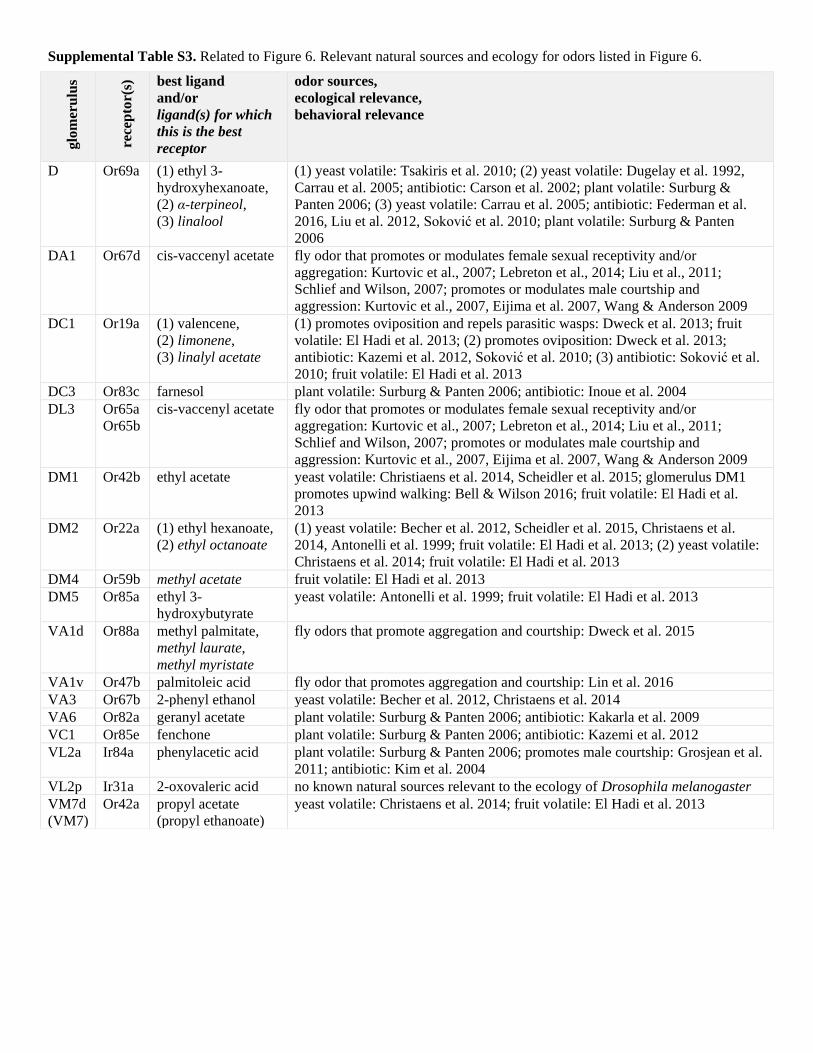
Supplemental Table S3. Related to Figure 6. Relevant natural sources and ecology for odors listed in Figure 6.
glo
mer
ulu
s
rece
pto
r(s)
best ligand
and/or
ligand(s) for which
this is the best
receptor
odor sources,
ecological relevance,
behavioral relevance
D Or69a (1) ethyl 3-
hydroxyhexanoate,
(2) α-terpineol,
(3) linalool
(1) yeast volatile: Tsakiris et al. 2010; (2) yeast volatile: Dugelay et al. 1992,
Carrau et al. 2005; antibiotic: Carson et al. 2002; plant volatile: Surburg &
Panten 2006; (3) yeast volatile: Carrau et al. 2005; antibiotic: Federman et al.
2016, Liu et al. 2012, Soković et al. 2010; plant volatile: Surburg & Panten
2006
DA1 Or67d cis-vaccenyl acetate fly odor that promotes or modulates female sexual receptivity and/or
aggregation: Kurtovic et al., 2007; Lebreton et al., 2014; Liu et al., 2011;
Schlief and Wilson, 2007; promotes or modulates male courtship and
aggression: Kurtovic et al., 2007, Eijima et al. 2007, Wang & Anderson 2009
DC1 Or19a (1) valencene,
(2) limonene,
(3) linalyl acetate
(1) promotes oviposition and repels parasitic wasps: Dweck et al. 2013; fruit
volatile: El Hadi et al. 2013; (2) promotes oviposition: Dweck et al. 2013;
antibiotic: Kazemi et al. 2012, Soković et al. 2010; (3) antibiotic: Soković et al.
2010; fruit volatile: El Hadi et al. 2013
DC3 Or83c farnesol plant volatile: Surburg & Panten 2006; antibiotic: Inoue et al. 2004
DL3 Or65a
Or65b
cis-vaccenyl acetate fly odor that promotes or modulates female sexual receptivity and/or
aggregation: Kurtovic et al., 2007; Lebreton et al., 2014; Liu et al., 2011;
Schlief and Wilson, 2007; promotes or modulates male courtship and
aggression: Kurtovic et al., 2007, Eijima et al. 2007, Wang & Anderson 2009
DM1 Or42b ethyl acetate yeast volatile: Christiaens et al. 2014, Scheidler et al. 2015; glomerulus DM1
promotes upwind walking: Bell & Wilson 2016; fruit volatile: El Hadi et al.
2013
DM2 Or22a (1) ethyl hexanoate,
(2) ethyl octanoate
(1) yeast volatile: Becher et al. 2012, Scheidler et al. 2015, Christaens et al.
2014, Antonelli et al. 1999; fruit volatile: El Hadi et al. 2013; (2) yeast volatile:
Christaens et al. 2014; fruit volatile: El Hadi et al. 2013
DM4 Or59b methyl acetate fruit volatile: El Hadi et al. 2013
DM5 Or85a ethyl 3-
hydroxybutyrate
yeast volatile: Antonelli et al. 1999; fruit volatile: El Hadi et al. 2013
VA1d Or88a methyl palmitate,
methyl laurate,
methyl myristate
fly odors that promote aggregation and courtship: Dweck et al. 2015
VA1v Or47b palmitoleic acid fly odor that promotes aggregation and courtship: Lin et al. 2016
VA3 Or67b 2-phenyl ethanol yeast volatile: Becher et al. 2012, Christaens et al. 2014
VA6 Or82a geranyl acetate plant volatile: Surburg & Panten 2006; antibiotic: Kakarla et al. 2009
VC1 Or85e fenchone plant volatile: Surburg & Panten 2006; antibiotic: Kazemi et al. 2012
VL2a Ir84a phenylacetic acid plant volatile: Surburg & Panten 2006; promotes male courtship: Grosjean et al.
2011; antibiotic: Kim et al. 2004
VL2p Ir31a 2-oxovaleric acid no known natural sources relevant to the ecology of Drosophila melanogaster
VM7d
(VM7)
Or42a propyl acetate
(propyl ethanoate)
yeast volatile: Christaens et al. 2014; fruit volatile: El Hadi et al. 2013











