Embed Size (px)
Citation preview

The nucleus for accommodation in themidbrain of the macaque
The effect of accommodation, pupillary constriction,and extraocular muscle contraction produced by
stimulation of the oculomotor nucleus on theintraocular pressure
Robert S. Jampel and Joel Mindel*
This paper reports upon: (1) the relationship between iris-bulge and accommodation of the eye;(2) the stereotaxic localization of midbrain sites that produce iris-bulge and changes in theretinoscopic reflex (accommodation); (3) the patterns of response from excitation of sites withinthe oculomotor nuclear complex; and (4) the effect of stimulation of the parasympatheticnucleus on the intraocular pressure.
I t has been generally accepted, in spiteof many decades of controversy, that thenuclei for the intraocular muscles arelocated in the anterior and dorsal regionof the oculomotor complex, probably inthe small-celled nuclei. The first descriptionof the autonomic component of the oculo-motor nucleus was by Edinger1 and West-phal- and bears their names. It consists ofa paired column of relatively small nervecells, extending anteriorly from about the
From the Institute of Ophthalmology, Columbia-Presbyterian Medical Center, New York, N. Y.
Supported by Research Grant NB 04547 from theNational Institute of Neurological Diseases andBlindness, United States Public Health Service,Bethesda, Md.
"Fellow in Neuro-ophthalmology, supported byUnited States Public Health Service Grant 5TINB 5324. Present address: University of Michi-gan Medical Center, Ann Arbor, Mich.
middle of the oculomotor complex, dorsaland medial to the somatic cell columns. Arostral extension of these small cell groupslies close to the midline, anterior and dorsalto the most rostral somatic oculomotor sub-nucleus.3"5 This extension gives the appear-ance of a single median nucleus and hasbeen called the "rostral median nucleus,"the "anteromedian nucleus," or the "rostralpart of the Edinger-Westphal nucleus."There has been some uncertainty in regardto whether the Edinger-Westphal nucleusand the anteromedian nucleus are twoseparate distinct nuclei or represent a con-tinuous cell mass.
Electrical stimulation experiments withhistological verification have proved thatthe Edinger-Westphal nucleus was actuallypart of the visceral efferent component.0"8
Pupillary constriction has served as themain functional indicator and was obtained
40
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/932995/ on 02/12/2018

Volume 6Number 1
Accommodation in the midbrain of the macaque 41
by stimulation of both the body and rostralparts of the Edinger-Westphal nucleus.However, there have been no recent de-tailed studies of the functional representa-tion of accommodation in the parasym-pathetic component and the literature onthis subject is meager. Hensen andVolckers9'xl produced pupillary constric-tion and forward bulging of the iris thatwas due to lens movements from stimu-lating the floor of the third ventricle of thedog. They localized accommodation in themost rostral part of the oculomotor com-plex. Adamiik10 confirmed these observa-tions. Bender and Weinstein7 evoked con-spicuous bilateral bulging forward of theirises on one occasion, in an area 1 mm.below the area from which they obtainedpupillary constriction in the macaque. War-wick'1' r> found cell degeneration in theipsilateral Edinger-Westphal nucleus andeven more marked cell degeneration in theanteromedian nucleus following ciliaryganglionectomy and oculomotor nervedivision. The first purpose of this paperis to report the results of stereotaxicallycontrolled electrical stimulation of thenucleus for accommodation and adjacentareas.
The effects of accommodation on intra-ocular pressure were meticulously studiedby Hess and Heine12 by electrical stimula-tion of the ciliary muscle and ciliary gan-glion in several species, including the catand monkey. They concluded that accom-modation had no effect on intraocularpressure. Greaves and Perkins13 found noincrease in intraocular pressure in the catfrom stimulating the peripheral cut end ofthe oculomotor nerve if contractions of theextraocular muscles were eliminated withdecamethonium iodide. However, Schmerland Steinberg14 reported increased intra-ocular pressure in the rabbit while stimu-lating the apex of the orbit. Also,Armalyiri> 1(i stimulated the ciliary ganglionin sympathectomized cats and recorded asignificant decrease in intraocular pressure.The second purpose of this paper is toreport the effects of accommodation and
pupillary constriction, evoked by electricalstimulation of the parasympathetic com-ponent of the oculomotor nucleus, on intra-ocular pressure.
Materials and methods
Twelve female monkeys (Macaco, mulatto), thatweighed between 2.0 and 2.5 kilograms, were usedin 15 experiments. Various anesthetic techniqueswere used: (1) intraperitoneal or intravenouspentobarbital in doses of from 60 to 125 mg., (2)halothane by inhalation, or (3) 30 to 50 mg. ofintraperitoneal pentobarbital supplemented by 4mg. of intramuscular Sernyl. The last techniquewas most satisfactory. In no discernible way didthe anesthetic technique appear to alter the ex-perimental results, but the most responsive animalswere lightly anesthetized. Procaine 1 per cent wasinjected locally before the skin incisions weremade.
All animals received endotracheal intubation andtheir heads were fixed in a stereotaxic apparatus.The head was shaved and a midline dorsal crani-otomy was performed. Approximately 3 by 7 cm.of parietal bone was removed. The dura was in-cised and carefully retracted so that the superiorsagittal sinus was not damaged. A No. 28 spinalneedle, insulated to the tip with Insul-X** served asa monopolar electrode. The indifterent electrodewas a cylindrical platinum rod placed in therectum.
The stereotaxic atlas devised for the macaqueby Snider and Lee17 was used as a guide for allelectrode placements. The electrode was insertedvertically from about 6 to 8 mm. anterior to thefrontal interaural plane (coordinates An to As)and as close to the midline as practicable (co-ordinates Rt to Li) and formed an angle of about30 degrees with the midbrain axis. The electrodewas moved downward in the midbrain in 0.5mm. steps until the desired response was obtained(anywhere from coordinates H + 3 to H - 5 ) , orthe electrode was withdrawn and reinsertedanterior or posterior to the first tract and theprocess repeated. Electrical stimuli were deliveredby an AEL stimulator, Model No. 104A. Theparameters used were 1 to 9 v., 100 per secondfrequency, and 1 msec, duration. Minimal stimuliwere always employed to obtain the desiredresponse. Stimuli were delivered in trains of upto 100 seconds.
Ocular, accommodative, and pupillary move-ments were observed and recorded cinemato-graphically. The Purkinje images formed on thesurface of the lens by an oblique light were ob-served and photographed. Retinoscopy (skiascopy)
"Insul-X Corporation, Yonkers, N. Y.
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/932995/ on 02/12/2018

42 jampel and Mindel Investigative OphthalmologyFebruary 1967
taneously,was insertedlimbus or at
with a slit retinos,cope was performed when indi-cated by the standard ophthalmic technique.13
Intraocular pressure was recorded manomet-ricallv either in one eye or in. both eyes simul-
A No. 25 gauge hypodermic needleinto the anterior chamber at thethe center of the cornea and con-
nected to a Statham pressure transducer, ModelP23 DC. The baseline pressures were adjusted byraising or lowering a saline reservoir that wasconnected to the system through a three-way stop-cock. Pressure changes were recorded by an ink-writing Grass oscillograph Model 52-925-725.Systemic blood pressure was monitored from thefemoral artery and intravenous drugs were injectedby means of indwelling catheters in the femoralartery and vein. Heparin in small amounts wasused to keep the polyethylene catheters patent.
In some experiments, to eliminate the influenceof the extraocular muscles on intraocular pressure,curare (100 meg, per cubic centimeter) was givenintravenously until respirations ceased. The animalthen received artificial respiration. The amount ofcurare required was usually about 500 meg, Also,in some experiments, phenylephrine 10 per centophthalmic solution was instilled in the eye toproduce mydriasis and facilitate the measurementof accommodation. Atropine 1 per cent and cyclo-pentolate 1 per cent were used as test drugs toproduce paralysis of the ciliary muscle. The roleof the extraocular muscles in producing changesin intraocular pressure was also studied bysequential cutting of their insertions from theglobe during stimulation of the somatic efferentsubnuclei of the oculomotor complex, and, on twooccasions, the .intracranial portion of the oculo-motor nerve. In order to identify the stimulatedpoints, electrolytic lesions were produced in themidbrain with coagulation currents. The animalswere put to death painlessly and perfused by the
intracardiac injection of refrigerated 10 per centformalin. Standard histological techniques wereused to localize the brain lesions.
Drugs used in these experiments were: halo-thane, Sernyl (Parke, Davis & Co.), sodium pento-barbital, procaine hydrochloride, d-tubocurarinechloride (Burroughs Wellcome & Co.), phenyl-ephrine (Winthrop Laboratories), Cyclopentolate(Schieffelin & Co.), and atropine (Alcon Labora-tories, Inc.).
Experimental results
The relationship between "iris-bulge" andaccommodation of the eye. The responsewas evoked in ten experiments by elec-trical stimulation of the midbrain pointsand the intracranial segment of the oculo-motor nerve. It was characterized by aconspicuous forward bulging of the pupil-lary or central portion of the iris whichproduced a marked convexity of the irisdiaphragm and a marked decrease in thedepth of the anterior chamber (Fig. 1).The extent of the bulging appeared directlyproportional to the intensity of the stimu-lating voltage until a maximum responsewas obtained. On observation of the eyefrom the side during iris-bulge, the centralportion of the lens appeared to becomeconical and to move forward into theanterior chamber. The following techniqueswere used to evaluate this phenomenon:
Retinoscopy (skiascopy). A dioptricchange in the power of the lens was ob-served which was dependent on the
Fig. 1. Photographs before and after electrical stimulation of the nucleus for accommodationin the oculomotor nuclear complex (see text). In A, the iris diaphragm is concave; in B, it isconvex. The iris is pushed forward by the bulging of the anterior surface of the lens. Notethat the diameter of the pupil is the same in both pictures.
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/932995/ on 02/12/2018

Volume GNumber 1
Accommodation in the midbrain of the macaque 43
strength of the stimulus. At high voltages(5 to 9 v.) increases in refractive powerof up to 10 diopters were measured.Increases of more than 10 diopters wereprobable in our young macaques. Theseincreases could not be measured accuratelyby retinoscopy because the retinoscopicreflex was obscured by the miosis andocular movements which resulted fromcurrent spread to adjacent areas whenhigh voltages were used. When changes ofless than 3 to 4 diopters were measured,little or no iris-bulge was observed.
Cinematography of the Furkinje images.Phenylephrine 10 per cent was instilledinto the eye in which the iris-bulge wasproduced by stimulation of midbrain sites.After 30 minutes the pupil became widelydilated (diameter about 7 mm.) and themidbrain sites were restimulated. Themovements of the Purkinje images reflectedfrom the anterior and posterior lens sur-faces were observed and photographed.During stimulation the anterior Purkinjeimage became smaller and moved forwardand farther away from the posterior Pur-kinje image. Movements of the anteriorlens surface appeared relatively unaffectedby phenylephrine, while pupillary constric-tion was markedly diminished.
The effect of atropine and cyclopentolateinstilled into the eye on the iris-bulge and,accommodation evoked by electrical stimu-lation. These drugs completely blocked theresponse within 40 minutes after instilla-tion.
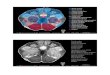
The stereotaxic localization of midbrainsites that produce iris-bulge and changesin the retinoscopic reflex (accommodation).Iris-bulge was obtained in 12 experimentsby stereotaxically controlled monopolarstimulation of midbrain sites that were at-tained during 19 different electrode in-sertions. The sites responsible for accom-modation were localized to the anterodorsalregion of the oculomotor nuclear complexin the so-called anteromedian nucleus (Fig.2). This area extended about 2 to 3 mm.in a rostrocaudal direction and about 1mm. to either side of the midline (co-
ordinates Ao to As and F^ to Lx). Isolatediris-bulge and iris-bulge with pupillary con-striction were evoked more frequently thanisolated pupillary constriction.
The patterns of response from excitationof sites within the oculomotor nuclear com-plex. In two experiments the electrode wasfortuitously inserted in the exact midline(Fig. 2). Low voltages (1 to 3 v.) evokedsymmetrical iris-bulge (accommodation ofmore than 3 diopters) in both eyes withoutassociated pupillary constriction or ocularmovements as the electrode entered therostral portion of the oculomotor nucleus.Gradually increasing the voltage (from 1to 8 v.) produced the following superim-
Fig. 2. Photomicrographs of a coronal sectionthrough the rostral part of the oculomotor nucleus.The electrode tract is below the aqueduct in themidline. A large electrolytic lesion is seen in thedorsomedial portion of the oculomotor nucleus atthe bottom of the electrode tract in the area ofthe anteromedial subnucleus. (A, Nissl, xlO; B,Nissl, x40.)
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/932995/ on 02/12/2018

44 Jampel and Mindel Investigative OphthalmologyFebruary 1967
posed phenomena in sequence: (1) bilat-eral symmetrical pupillary constriction,(2) bilateral symmetrical or asymmetricaldownward ocular movement, (3) bilateralabrupt (phasic) adduction of the eyes,and (4) bilateral lid retraction. When theelectrode was moved downward about 1mm. from the site that produced isolatedbilateral iris-bulge a site was reached inwhich low voltage stimulation producedboth iris-bulge and pupillary constrictionand 0.5 mm. below this site stimulationproduced only bilateral pupillary constric-tion.
In ten experiments the stimulating elec-trode was angulated slightly off the midlinebecause of technical problems or was de-liberately inserted laterally about 1 mm.from the midline. The sequence of events,as the electrode was moved downward,was usually the same as in midline stimu-lation except that the responses were ipsi-lateral, i.e., isolated iris-bulge, iris-bulgeand pupillary constriction, and isolatedpupillary constriction. In some experimentsiris-bulge and pupillary constriction couldnot be dissociated and each phenomenonoccurred to a varying extent at all thesites of stimulation, but in most cases, iris-bulge was more intense than pupillaryconstriction in dorsal and rostral sites andthe reverse was true from more ventraland caudal sites.
When the electrode was reinserted indifferent tracts at intervals of 1 mm. fromleft to right across the midline in a frontalplane (e.g., AG, L^ AG, M(midline); AG,Rt) the sequence of responses to stimula-tion was: (1) left ipsilateral iris-bulge orpupillary constriction or both, (2) bilateraliris-bulge or pupillary constriction or both,and (3) right ipsilateral iris-bulge or pupil-lary constriction or both. If the stimulusvoltages were increased (e.g., 2 to 6 v.) ata lateral site which ordinarily producedipsilateral reactions, responses were ob-tained in the contralateral eye. Also, thesecontralateral responses were of lower am-plitude than the ipsilateral responses.
It was more usual to have ocular move-
ments associated with iris-bulge and pupil-lary constriction then for these responsesto occur as isolated phenomena. In rostraltracts, iris-bulge was associated with down-ward ocular movements that had an in-ward component and in more caudal tractspupillary constriction was associated withocular adduction.
The effect of stimulation of the para-sympathetic nucleus on the intraocularpressure. As described above, stimulationproduced accommodation or pupillary con-striction or both, depending on the posi-tion of the electrode. Accommodative am-plitudes of up to 10 diopters were obtainedwith intermittent stimulation at one persecond and with prolonged stimulation.Even with such strong accommodativemovements, associated with a marked de-crease in the depth of the anterior cham-ber, no significant rise in intraocular tensionwas recorded (Fig. 4, upper trace).Maximal isolated pupillary constriction andpupillary constriction associated with ac-commodation also produced no change inintraocular pressure. Varying the baselineintraocular pressure from 0 to 50 mm. Hghad no effect on the absence of pressureresponse to accommodation or pupillaryconstriction. Prolonged stimulation of upto 100 seconds had no effect on intraocularpressure that could not be accounted forby extraocular muscle contraction or varia-tions in blood pressure due to currentspread into contiguous areas (Figs. 4, 5,and 6), as described below.
A common response from oculomotornucleus stimulation was to obtain accom-modation associated with increased extra-ocular muscle tone even when no ocularmovements were visible. In these cases,there was a steep rise in intraocular pres-sure that appeared at first glance to berelated to the accommodation. However,intravenous curare abolished these in-creases in intraocular pressure even whenvigorous accommodative movements werestill taking place.
During prolonged stimulation in or nearthe oculomotor nucleus, changes in sys-
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/932995/ on 02/12/2018

Volume 6Number 1
Accommodation in the midbrain of the macaque 45
temic blood pressure were observed whichcorrelated in time with changes in intra-ocular pressure (Figs. 4 and 6). A commonresponse was a slight decrease in systemicblood pressure and intraocular pressureupon cessation of the stimulus (Fig. 6).The reverse was also seen, namely, an in-crease in systemic blood pressure and in-traocular pressure following cessation ofthe stimulus (Fig. 4, two lower traces).
In one experiment, intraocular pressuredecay was studied during evoked accom-modation with the opposite eye serving ascontrol: the intraocular pressures in botheyes were elevated to 30 mm. Hg and thegradual decrease in pressure in both eyeswas observed. Accommodation evoked inone eye during this period of intraoculardecay did not affect the slope of the decaycurve of that eye as compared to the non-accommodating eye.
Discussion
Accommodation. The maximum accom-modative range evoked in young macaquesfrom the dorsorostral region of the oculo-motor complex was from 10 to 12 diopters.This range of accommodation was previ-ously found by Hess and Heine12 by stimu-lating the ciliary ganglion. A marked de-crease in the depth of the anterior cham-ber accompanied high amplitudes of ac-commodation. The conical bulging move-ment of the central portion (collarette) ofthe iris, which accompanied these reactions,was a passive motion. It resulted from thecontraction of the ciliary muscle whichproduced a forward movement of the an-terior surface of the lens and an increasein the dioptric power of the eye. Con-spicuous passive iris movement was easilyseen with changes in dioptric power ofthe lens of at least 3 diopters. Accommoda-tion of less than 3 diopters was observedonly by retinoscopy.
Passive iris movements mainly, but alsochanges in the retinoscopic reflex and thePurkinje images, were used as indicatorsfor stereotaxically mapping out the nucleusof accommodation. The concept of the
morphology of the parasympathetic com-ponent of the oculomotor nucleus that weformulated from these experiments was asfollows: The visceral efferent nucleus con-sists of two adjacent elongated cell massesthat lie on both sides of the midline raphein the anterior and dorsal part of the oculo-motor nucleus. These cell masses extendfrom about 1 mm. to either side of themidline and about 3 mm. in a rostrocaudaldirection. The innervation for the ciliarymuscle and for the pupil appear to beentirely from the ipsilateral nucleus. Thereis a discrete cell mass that mediates accom-modation alone, located dorsal and some-what rostral to a discrete cell mass thatmediates pupillary constriction alone. Thesecell masses are contiguous or there is anintermediate zone of admixture. The rostralpart of the parasympathetic nucleus ap-pears intimately associated with the somat-ic subnucleus for the inferior rectus muscle,and the caudal part with the subnucleusfor the medial rectus muscle.
Our experiments support the idea thatthere is a discrete cell mass subservingaccommodation and a discrete cell masssubserving pupillary constriction, but webelieve them to be contiguous or over-lapping in an intermediate zone. We con-cur completely with Warwick,4'5 in thatwe believe the ocular parasympatheticnucleus to be a continuous cell mass andalso that the innervations are ipsilateral.Warwick concluded from retrograde de-generation experiments that 93 per centof cells in the ciliary ganglion innervatedthe ciliary muscle and 3 per cent the irissphincter, a ratio of about 30 to 1 in favorof the ciliary muscle. This is reasonablesince the ciliary muscle is many timeslarger than the iris sphincter. Our observa-tions suggest that there are many morecells subserving accommodation than pupil-lary constriction since accommodation oraccommodation with varying degrees ofpupillary constriction was a much morefrequently evoked response than isolatedpupillary constriction.
Intraocular pressure. Accommodation
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/932995/ on 02/12/2018

46 ]ampel and Mindel In vestiguti va OphthalmologyFebruary 1967
evoked by stimulation of the parasym-pathetic nucleus of the oculomotor complexdid not cause a rise in intraocular pres-sure. This fact was confirmed for inter-mittent stimulation at one per second thatproduced maximum accommodation, inwhich the movements of the iris and lensseemed to resemble the vigorous action ofa pump, as well as for prolonged stimula-tion for about 100 seconds (Fig. 4). Also,centrally evoked accommodation did notaffect the intraocular pressure decay curve,and experimentally changing the baselineintraocular tension did not influence theresults.
Stimulation of the somatic componentof the oculomotor nucleus produced a steeprise in intraocular pressure due to com-pression of the globe, if there was simul-taneous contraction of the extraocular mus-cles, especially the superior and inferiorrectus muscles (Figs. 3, 5, and 6),
Changes in systemic blood pressure were
frequently associated with stimulation ofthe oculomotor complex. These vasculareffects were probably due to spread ofcurrent into adjacent areas during pro-longed stimulation and coincided with vari-ations in intraocular pressure (Figs. 4 and6). A frequent response was to have eithera rise or a fall in blood pressure and intra-ocular pressure following stimulation (Fig.4, second trace from the top, and Fig. 6).
From our experiments we conclude thatthe acts of accommodation and ciliarymuscle contraction in the healthy macaquehave no significant influence on intraocularpressure or on the regulation of aqueoushumor dynamics. The advantages in ourexperimental approach were that we usedan animal with a large accommodative am-plitude (the cat has only about 3 diopteraccommodative amplitude) and producedaccommodation preganglionically by stim-ulating the central nucleus without dis-turbing the orbital contents at all.
Fig. 3. Traces showing the marked increase in intraocular pressure that resulted from elec-trical stimulation of the somatic oculomotor subnuclei. The increased pressure was causedmainly by the simultaneous contraction of the superior and inferior recti as shown in thelowest trace, since cutting the insertion of the superior rectus prevented most of the rise inintraocular pressure. IOP, Intraocular pressure; BP, blood pressure; STIM, stimulus; SR,superior rectus; MR, medial rectus.
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/932995/ on 02/12/2018

Volume 6Number 1
Accommodation in the midbrain of the macaque 47
Changes in the intraocular pressure fromstimulation in or near the oculomotor nu-cleus were associated with simultaneouscontraction of the extraocular muscles thatproduced compression of the globe. In
addition, intraocular pressure changes wereassociated with changes in the systemicblood pressure. In our experiments thesetwo factors were artifacts. However, it ispossible that one or both are important
Fig. 4. Traces depicting the efFect of prolonged stimulation of the nucleus for accommodationthat produced marked iris-bulge. In the upper trace there was no change in the intraocularpressure or in the blood pressure associated with accommodation, which was the usual re-sponse. The stimulus parameters were minimal (2 v.) and the animal responsive. In the othertraces there are variations in blood pressure that appear related to variations in intraocular pres-sure. Note the abrupt rise in intraocular pressure after the cessation of the stimuli in the lowertwo traces. 1OP, Intraocular pressure; BP, blood pressure; STIM, stimulus.
Fig. 5. Midline stimulation of the anteromedian nucleus produced conspicuous bulging of theiris-lens diaphragm in both eyes. During the first two stimuli there was an abrupt rise inintraocular pressure. Although there was only slight eye movement, the rise in intraocularpressure was assumed to be the result of contraction of the extraocular muscles that wascaused by stimulus spread to adjacent somatic nuclear areas. This contention was verifiedby the intravenous injection of curare which abolished the muscular contraction. Subsequentstimulation resulted in marked iris-bulge, but no increase in intraocular pressure or changein blood pressure. BP, Blood pressure; IOP, intraocular pressure] OD} right eye; OS, left eye;STIM, stimulus.
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/932995/ on 02/12/2018

48 Jampel and Mindel Investigative OphthalmologyFebruary 1967
10 P
W W W W W W ^ ^
Fig. 6. Traces of two experiments in which ipsilateral accommodation and pupillary constric-tion were produced by stimulating the anteromedian nucleus of the oculomotor complex. Theincrease in intraocular pressure was produced by stimulus spread to adjacent somatic motornuclei. When curare was given intravenously the rise in intraocular pressure was graduallyeliminated, although marked accommodation and pupillary constriction were still elicited(each arrow in the lower trace represents the injection of 0.1 mg. of curare). Note thetendency for the blood pressure to decrease when the intraocular pressure increases. BP, Bloodpressure; IOP, intraocular pressure; STIM, stimulus.
means by which aqueous humor dynamicsare influenced by the central nervous sys-tem. We obtained rises in intraocular pres-sure produced by tonic extraocular musclecontraction, that were unassociated withvisible ocular movements. Also, as VonSallmann and Lowenstein10 point out, focalvascular changes confined to the intraocularcirculation may result from brain stimula-tion, since they found areas in the dien-cephalon which, when stimulated, pro-duced variations in intraocular pressurethat were unassociated with variations insystemic blood pressure or extraocular mus-
cle activity. It is obvious that there is muchmore to be learned about the relationshipsbetween the intraocular pressure and thecentral nervous system.
In contrast to our results, Armaly15'ie
obtained a decrease in intraocular pressurein sympathectomized cats from stimulatingthe ciliary ganglion. Further work will beneeded to reconcile our work with hisfindings. Perhaps there are neuronal path-ways that reach the ciliary ganglion frombrain areas other than the oculomotor nu-cleus?
In our own thinking on this complex
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/932995/ on 02/12/2018

Volume 6Number 1
Accommodation in the midbrain of the macaque 49
matter we are not surprised at the factthat accommodation and the structuralchanges reported in the chamber angleaccompanying accommodation14-15 do notinfluence intraocular pressure. The eye ap-pears to maintain its pressure equilibriumin spite of rather gross dynamic ciliarymuscle contractions. Imagine the pommel-ing sensitive tissues might be subjected toif this were not the case!
Summary
1. Conspicuous bulging forward of theiris was evoked in the macaque by stereo-taxically controlled electrical stimulation inthe dorsal and rostral part of the oculo-motor nucleus in the so-called anterome-dian nucleus. The iris-bulge was a passivemotion due to a conical forward bulgingof the anterior lens surface which resultedin an increased dioptric power of the eyeand a marked narrowing of the anteriorchamber. Iris-bulge was observed whenthe eye was accommodating more thanthree diopters. Iris-bulge and retinoscopywere used as indicators that the nucleusfor accommodation was being electricallystimulated in the midbrain.
2. As mapped by the responses in ourexperiments, the parasympathetic compo-nent of the oculomotor nucleus appearsto be composed of paired elongated cellmasses in the dorsorostral portion of theoculomotor nucleus that lie close to thernidline. Discrete innervation of the ipsi-lateral ciliary muscle arises from cells thatare located dorsally and somewhat rostrallyto the cells that innervate the ipsilateraliris sphincter. The cells that innervate thesediscrete functions are either contiguous,mixed, or overlap in an intermediate zone.The majority of the cells appear concernedwith accommodation and a minority withpupillary constriction. Warwick's conceptthat the Edinger-Westphal nucleus andanteromedian nucleus form a continuousmass and are integral parts of the para-sympathetic oculomotor nucleus is sup-ported by these experiments.
3. Stimulation of the somatic oculomotor
subnuclei produced a marked increase inthe intraocular pressure which was due tocompression of the globe by the simul-taneous contraction of normally antagonisticmuscles, especially the superior rectus andthe inferior rectus. The increase in intra-ocular tension was eliminated by severingthe extraocular muscles from the globe orby the intravenous injection of curare.
4. Stimulation of the parasympatheticnucleus that evoked accommodation orpupillary constriction or both, producedno change in intraocular pressure or changein the rate of outflow of aqueous humor.This observation was true regardless ofthe baseline intraocular tension and forprolonged stimulation up to 100 secondsas well as for maximum accommodativemovements occurring at a frequency asfast as one per second.
5. Stimulation in or near the oculomotorcomplex frequently caused variations insystemic blood pressure, which coincidedwith small variations in intraocular pres-sure.
REFERENCES1. Edinger, L.: Uber den Verlauf der centralen
Hirnnervenbahnen mit Demonstration vonPraparaten, Neurol. Centralbl. 4: 309, 1885.
2. Westphal, C.: t)ber einen Fall von chronischerprogressive Lahmung des Augenmuskeln(ophthalmoplegia externa) nebst Beschreibungvon Ganglienzellengruppen im Bereich desOculomotoriuskerns, Arch. f. Psychiat. u.Nervenh. 98: 846, 1887.
3. Crosby, E. C , and Woodburne, R. T.: Thenuclear pattern of the non-tectal portions ofthe midbrain and isthmus in primates, J.Comp. Neurol. 78: 441, 1943.
4. Warwick, R.: The ocular parasympatheticnerve supply and its mesencephalic sources,J. Anat. (London) 88: 71, 1954.
5. Warwick, R.: Oculomotor organization, Ann.Roy. Coll. Surgeons (England) 19: 36, 1956.
6. Benjamin, J. W.: The nucleus of the oculo-motor nerve with special reference to innerva-tion of the pupil and fibers from the pretectalregion, J. Nerv. & Ment. Dis. 89: 294, 1939.
7. Bender, M. D., and Weinstein, E. A.: Func-tional representation in the oculomotor andtrochlear nuclei, Arch. Neurol. & Psychiat.49: 98, 1943.
8. Szentagothai, J.: Anatomical aspects of in-hibitory pathways and synapses, in Florey,
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/932995/ on 02/12/2018

50 Jampel and Mindel Investigative OphthalmologyFebruary 1967
editor: Nervous inhibition, Oxford, 1961,Pergamon Press, Ltd., pp. 32-39.
9. Hensen, V., and Volckers, C : Experimen-taluntersuchung iiber den Mechanisrnus derAccommodation, Kiel, 1868, Schwers.
10. Adamiik: Zur Frage iiber den Mechanisrnusder Accommodation, Zentralbl. med. Wiss. 8:292, 1870.
11. Hensen, V., and Volckers, C : t)ber denUnsprung den Accommodationsnerven, Arch,f. Ophth. 24: 1, 1878.
12. Hess, C, and Heine, L.: Arbeiten aus demGebiete der Accommodationslehre, Arch. f.Ophth. 46: 243, 1898.
13. Greaves, D. P., and Perkins, E. S.: Influenceof the third cranial nerve on intraocular pres-sure, Brit. J. Ophth. 37: 54, 1953.
14. Schmerl, E., and Steinberg, B.: The role ofthe ciliary and superior cervical ganglia inocular tension, Am. J. Ophth. 32: 947, 1949.
15. Armaly, M. F.: Studies on intraocular effectsof the orbital parasympathetic pathway. I.Technique and effects on moq)hology, Arch.Ophth. 61: 14, 1959.
16. Armaly, M. F.: Studies on intraocular effectsof the orbital parasympathetics. II. Effect onintraocular pressure, Arch. Ophth. 62: 117,1959.
17. Snider, R. S., and Lee, J. C : A stereotaxicatlas of the monkey brain (Macaca mulatto),Chicago, 1961, The University of ChicagoPress.
18. Johnson, V.: The physiologic optics of retinos-copy, Arch. Ophth. 13: 65, 1935.
19. Von Sallmann, L., and Lowenstein, O.:Responses of intraocular pressure, blood pres-sure, and cutaneous vessels to electric stimu-lation in the diencephalon, Am. J. Ophth.39: 11, 1955.
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/932995/ on 02/12/2018