Embed Size (px)
Citation preview

REVIEWS OF GEOPHYSICS, SUPPLEMENT, PAGES 1277-1286, JULY 1995 U.S. NATIONAL REPORT TO INTERNATIONAL UNION OF GEODESY AND GEOPHYSICS 1991-1994
The iron hypothesis: Basic research meets environmental policy
Sallie W. Chisholm Department of Civil and Environmental Engineering, and Department of Biology Massachusetts Institute of Technology, Cambridge, Massachusetts
Introduction In October 1993, an interdisciplinary group of scientists
spread 445 kg of iron over a 64 km 2 patch of surface ocean in the eastern equatorial Pacific. They monitored the patch for 9 days, comparing its biology and chemistry to the unfertilized surrounding waters. They found that the phytoplankton (microscopic plants which comprise the base of the food web in the oceans) were three times more productive in the patch than outside it. In other words, the cells were photosynthesiz-ing—i.e., converting C 0 2 to organic carbon—at a significantly enhanced rate. The group concluded that iron was the limiting factor in this ecosystem, solving one of the more perplexing puzzles in biological oceanography in recent years. This event marked not only the first unenclosed ocean ecosystem experiment, but also the climax to a complex and fascinating evolution of ideas in biological oceanography. The climax was bittersweet, however, as John Martin, the designer of the experiment and originator of the hypothesis, died shortly before the expedition.
The story behind the iron enrichment experiment is interesting not only for the scientific results per se, but also because it reveals why straightforward answers to simple questions are difficult to come by in biological oceanography. More importantly, the results of this experiment, and the events that led up to it, contain important lessons for those working at the interface between science and public policy.
Phytoplankton and the Biological Pump Phytoplankton are responsible for approximately 40% of
the planet's total annual photosynthetic (i.e., 'primary') production. The magnitude of their contribution to global primary production is often unappreciated because they represent such a small fraction of the total photosynthetic biomass on earth. Their high output per unit biomass is possible because of their high surface to volume ratio, because they are free-floating and thus have no supporting structures to maintain, and because they are embedded in a very dynamic ecosystem where time scales of environmental forcing are on the order of days.
Light and nutrients are the primary factors regulating phytoplankton growth. Light attenuates dramatically with depth, penetrating roughly 200m in the open ocean, thus illuminating only a tiny fraction of the total ocean volume. Solar energy heats the surface waters, generating a temperature gradient
Copyright 1995 by the American Geophysical Union.
Paper number 95RG00743. 8755-1209/95/95RG-00743S15.00
(the thermocline) which, along with the salinity gradient (the halocline) generated by surface rainfall, creates a density gradient (the pycnocline) that effectively isolates the surface wind-mixed layer from the waters below. Inorganic nutrients such as nitrogen and phosphorus, central to the cells' biochemistry, are constantly stripped from the surface waters by the growing phytoplankton. Most of the phytoplankton are eaten on the spot and the nutrients comprising their biomass are regenerated at the surface and are available for another round of production. Some fraction of the primary production, however, finds its way to the deep sea, either through the settling of dead cells and fecal matter or through advection in the course of global circulation patterns. Most of the organic carbon which finds its way to the deep sea is ultimately assimilated by bacteria, which regenerate it (and N and P) into inorganic forms (Fig. 1). The collective action of this so-called "biological pump" over time scales of centuries is to create a sharp concentration gradient of nutrients with depth (Fig. 2a). The gradient is larger in the Pacific than the Atlantic because deep ocean currents flow from the Atlantic to the Pacific, collecting as they travel the "rain" of nutrient-rich particles from the productive surface waters above.
This biochemical pump—comprised of phytoplankton cells, their animal predators, and the bacteria that assimilate their waste — plays a central role in the global carbon cycle. Because the elemental composition of phytoplankton is C:N:P = 106:16:1 (commonly referred to as the "Redfield Ratio"— see Redfield, 1934, 1958), about 100 units of carbon are delivered to the deep sea for every unit of phosphorus. As such, the biological pump delivers carbon from the atmosphere to the deep sea, where it is concentrated and sequestered for centuries (Fig. 2b). Superimposed on the biochemical pump is a "solubility pump" which also serves to concentrate dissolved inorganic carbon ( C 0 2 plus bicarbonate and carbonate ions) in the deep ocean. The driving force of this pump is increased solubility of C 0 2 in the cold surface waters at high latitudes where the water sinks to join the deep ocean currents. Although estimates of the relative importance of these two pumps have a wide range, the most recent estimate (Sarmiento and Bender, 1994) suggests that 75% of the difference in dissolved inorganic carbon concentration between the surface and deep oceans is due to the biological pump. If it were eliminated tomorrow, and the dynamics of the oceanic carbon cycle were driven solely by physical and chemical processes, the amount of carbon released from the deep ocean as it equilibrated with the atmosphere would more than double the C 0 2 concentration in the atmosphere.
Although there are many cyclic processes in the surface waters which are coupled to the biological pump, the net flux of biologically-produced materials is one way — from the surface to the deep ocean. So how do the nutrients find their way
1277
1278 CHISHOLM: THE IRON HYPOTHESIS
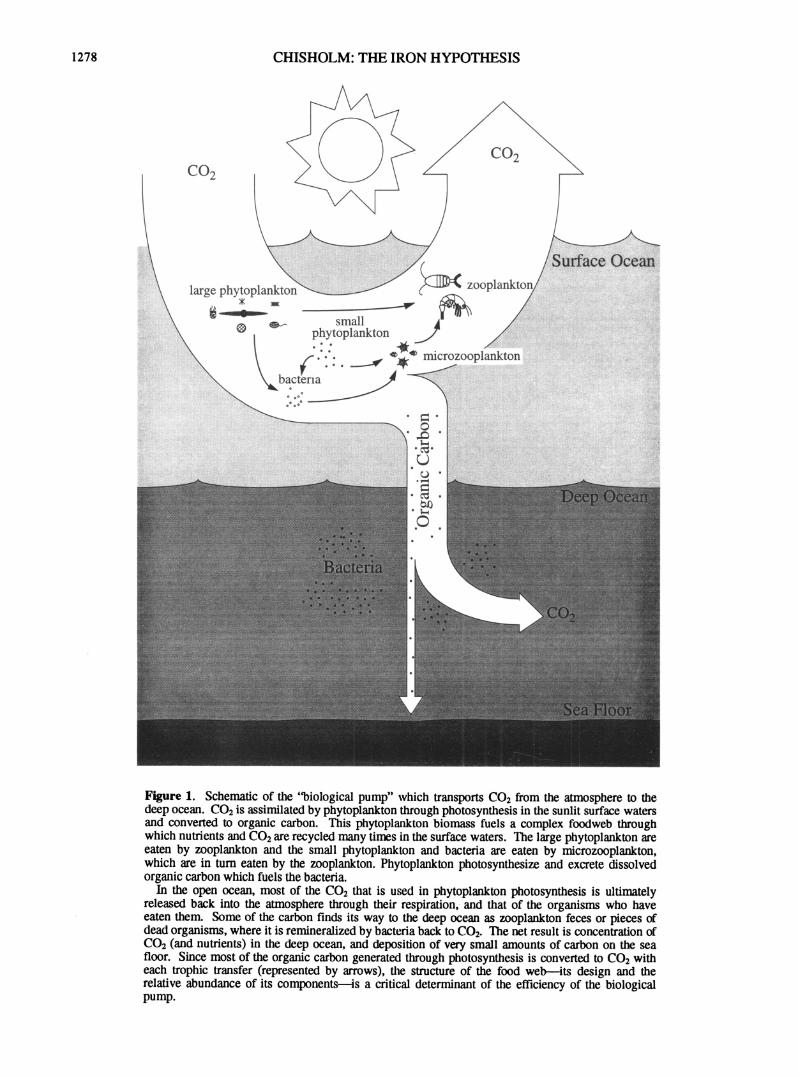
Figure 1. Schematic of the "biological pump" which transports C 0 2 from the atmosphere to the deep ocean. C 0 2 is assimilated by phytoplankton through photosynthesis in the sunlit surface waters and converted to organic carbon. This phytoplankton biomass fuels a complex foodweb through which nutrients and C 0 2 are recycled many times in the surface waters. The large phytoplankton are eaten by zooplankton and the small phytoplankton and bacteria are eaten by microzooplankton, which are in turn eaten by the zooplankton. Phytoplankton photosynthesize and excrete dissolved organic carbon which fuels the bacteria.
In the open ocean, most of the C 0 2 that is used in phytoplankton photosynthesis is ultimately released back into the atmosphere through their respiration, and that of the organisms who have eaten them. Some of the carbon finds its way to the deep ocean as zooplankton feces or pieces of dead organisms, where it is remineralized by bacteria back to C0 2 . The net result is concentration of C 0 2 (and nutrients) in the deep ocean, and deposition of very small amounts of carbon on the sea floor. Since most of the organic carbon generated through photosynthesis is converted to C 0 2 with each trophic transfer (represented by arrows), the structure of the food web—its design and the relative abundance of its components—is a critical determinant of the efficiency of the biological pump.

CHISHOLM: THE IRON HYPOTHESIS 1279
N 0 3 - N ( /zg-at EC0 2, mol m"3
20 40 2.0 2.2 2.4
Figure 2. Depth profiles of (A) inorganic nutrients and (B) dissolved inorganic carbon in the Atlantic and Pacific Oceans. Nutrients are essentially depleted in the surface waters where they limit phytoplankton production. Largely through the mechanism of the biological pump, both nutrients and C 0 2 are concentrated in the deep sea. (A: Adapted from Richards, 1968; B: Adapted from Broecker, 1974).
back to the surface waters to refuel the pump? The answer is critical because the rate of resupply of nutrients from the deep water to the surface regulates the rate of phytoplankton production globally on the time scales relevant to anthropogenic change. There are essentially three mechanisms by which the nutrients are returned to the surface: Upwelling of the nutrient-rich deep water (e.g., along the western coasts of continents); diffusion across the thermocline (e.g., the central ocean gyres); and seasonal, wind-driven deep mixing which erodes the thermocline and entrains the deep water to the surface (e.g., the N. Atlantic). As the phytoplankton pump is refueled by N and P to generate more organic carbon through photosynthesis, C 0 2 from the deep water is being supplied as well. Put quite simply, inorganic N and P are delivered to the surface waters in the biologically dictated Redfield Ratio (along with C0 2 in excess), are converted to organic compounds (i.e„ biomass) in the Redfield proportions of C:N:P, are recycled in the surface waters many times in those proportions, and are ultimately re-pumped to the deep sea—in those proportions. Thus, on a global scale, any changes in upwelling and the supply of nutrients to the surface waters will not influence the biological pump's role in the annual atmospheric carbon budget, so long as the supply and assimilation of N, P, and C are tightly coupled.
In some areas of the world's oceans the supply and assimilation of N, P and C are not tightly coupled, even on an annual average, because N and P do not appear to be limiting phytoplankton growth. These are the so called "high-nutrient, low-chlorophyll" (HNLQ waters of the subarctic Pacific, the Southern ocean around Antarctica, and the equatorial Pacific (see Cullen, 1991). Here the biological pump is inefficient: the phytoplankton standing stocks are not large enough to assimilate the N and P in the surface waters fast enough to deplete them at any time throughout the year. These regions, particularly the Southern ocean, are of major interest to paleo-oceanographers because they could have played a role in past climate change (Sarmiento and Togweiler, 1984). The cold surface waters of the Southern ocean steadily sink to the abyss as part of the global ocean circulation and, in today's ocean, carry with them a large reservoir of unused nutrients—a po
tential carbon sink of some magnitude. The actual significance of this potential sink in the global scheme of things will be discussed below. The more immediate question is: What factor(s) are responsible for the inefficiency of the biological pump in these regions? Why can't the phytoplankton use up the nutrients, as they do so effectively in the other regions of the world's oceans?
Many hypotheses have been proposed, which primarily involve light limitation or control by zooplankton grazing (see Cullen, 1991). Each has merit and describes important factors controlling phytoplankton in these regions. The most provocative, and now compelling, hypothesis in recent years, however, is the suggestion that the supply of iron to these regions is regulating primary productivity.
The Iron Hypothesis As early as the 1930's, the potential role of iron as a limit
ing factor in phytoplankton productivity was appreciated (Gran, 1931; Hart, 1934; Harvey, 1938). These early workers recognized the extreme insolubility of iron in today's oxygenated oceans, and deduced that it must be in very short supply in areas where it was not readily replenished from the land. Although they also recognized the analytical difficulties in actually measuring iron concentrations in the sea, they would be shocked to learn that their estimates of offshore iron concentrations were high by two orders of magnitude (Martin, 1992).
Because of the difficulties of working with trace levels of iron, direct evidence of its role in phytoplankton ecology has always been difficult to obtain. Its potential as a limiting factor becomes clear, however, when one recognizes that the early evolution of the biochemical pathways that drive phytoplankton physiology occurred when the ocean and atmosphere were anoxic. Because of its high solubility under these reduced (oxygen free) conditions, iron was extremely abundant relative to other trace nutrients (Table 1) and biochemical pathways were not under evolutionary pressure to be efficient with respect to iron. When oxygenic photosynthesis evolved around 3 billion years ago, iron was rapidly oxidized, becom-

1280 CHISHOLM: THE IRON HYPOTHESIS
Table 1. Relative molar concentrations of trace elements in today's oceans (normalized to P), and in the anoxic ocean characteristic of early evolutionary times (molar concentrations), com-
ared with the molar ratios of these elements in today's phytoplankton (modified from Brand, 991; Martin et al 1993). Comparing the availability of the various elements in the water to
day with that in the phytoplankton reveals that iron is in short supply relative to the requirements of the cells. In the ancient ocean when biochemical pathways were evolving, iron was readily available relative to other trace elements.
P Fe Zn Mn Cu
Average Plankton Composition (molar ratios)
1000 10 2 0.4 0.6
Average Seawater Atlantic Composition (molar ratios) Pacific
1000
1000
0.45
0.2
1.7
3.0
0.58
0.075
1.7
1.5
Ancient (anoxic) Oceans (molar concentrations)
io-3 10" IO"5 10"
ing significantly less available to the biota. Since evolution cannot rewrite its story when the environment changes, many of the biochemical pathways that sustain life today carry designs inspired by ancient environments. This appears to be the case for iron (Table 1).
In the late 1980's John Martin began to weave together several independent threads of evidence, revitalizing the early idea that iron plays a major regulatory role in phytoplankton productivity (Martin, 1992). First, he recognized that the primary source of iron to the surface waters of the oceans is from the land, either via atmospheric dust deposition in offshore areas, or direct depositions from land masses. Second, his lab was able to demonstrate, for the first time, that dissolved iron concentrations in offshore areas are indeed extremely low (Martin and Gordon, 1988). In the Drake Passage of the Antarctic, for example, the concentration in the surface waters is 1.6 x 10"10 mole Fe kg"1 (or 3.2 mg in 3 ) . Using "Redfieldian" reasoning, he calculated that this amount of iron was one-tenth that required to allow the phytoplankton to assimilate the ambient nitrate in the surface waters. Third, he observed
that atmospheric dust deposition in the two major HNLC areas—the Antarctic and equatorial Pacific oceans—are the lowest in the world (Prospero, 1981; Uematsu, 1987). Conversely, in the equatorial N. Atlantic, which receives large amounts of dust from the Saharan desert (Prospero, 1981), iron concentrations are sufficient for the complete assimilation of available nitrates and phosphates.
Martin and his team buttressed their argument with experiments. They filled bottles with surface waters from the HNLC regions and incubated them at "simulated in situ" light and temperature for roughly a week. To half of the bottles they added iron, and the other half were left alone as control bottles. They monitored phytoplankton abundance in the bottles by various means, and also nutrient (nitrates and phosphates) concentrations, to see if the addition of iron allowed the phytoplankton to assimilate additional nutrients. The general results were always the same: the total chlorophyll (a measure of phytoplankton biomass) in the iron-enriched bottles was always higher than in the control bottles at the end of the experiment, and nitrates were always more depleted in the
^ 6 'o S
3> 4
2
control
\ +Fe
1 1 i i i i 2 4 6
D a y
Figure 3. 'Typical" results from iron-enrichment bottle incubation experiments—in this case from the subarctic Pacific (after Martin & Fitzwater, 1988). Chlorophyll (A), and nitrate (B) concentrations in bottles incubated with (open circles) and without (closed circles) added iron (5 nmol kg"1) for 6 days. The iron-enriched bottles show dramatically increased chlorophyll concentrations and decreased nitrate and phosphate relative to the control bottles. Examination of the phytoplankton community in the bottles revealed substantial growth of pennate diatoms in the iron enriched bottles, which were a minor component of the phytoplankton community initially.
CHISHOLM: THE IRON HYPOTHESIS 1281
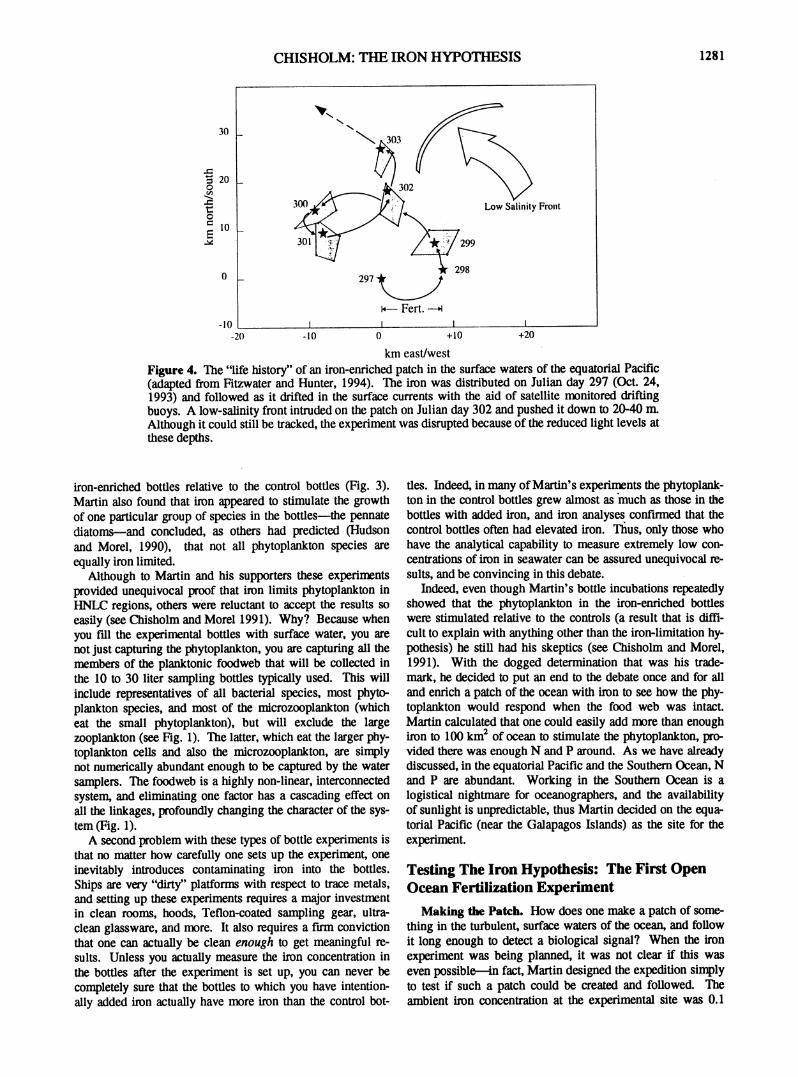
km east/west Figure 4. The "life history" of an iron-enriched patch in the surface waters of the equatorial Pacific (adapted from Fitzwater and Hunter, 1994). The iron was distributed on Julian day 297 (Oct. 24, 1993) and followed as it drifted in the surface currents with the aid of satellite monitored drifting buoys. A low-salinity front intruded on the patch on Julian day 302 and pushed it down to 20-40 m. Although it could still be tracked, the experiment was disrupted because of the reduced light levels at these depths.
iron-enriched bottles relative to the control bottles (Fig. 3). Martin also found that iron appeared to stimulate the growth of one particular group of species in the bottles—the pennate diatoms—and concluded, as others had predicted (Hudson and Morel, 1990), that not all phytoplankton species are equally iron limited.
Although to Martin and his supporters these experiments provided unequivocal proof that iron limits phytoplankton in HNLC regions, others were reluctant to accept the results so easily (see Chisholm and Morel 1991). Why? Because when you fill the experimental bottles with surface water, you are not just capturing the phytoplankton, you are capturing all the members of the planktonic foodweb that will be collected in the 10 to 30 liter sampling bottles typically used. This will include representatives of all bacterial species, most phytoplankton species, and most of the microzooplankton (which eat the small phytoplankton), but will exclude the large zooplankton (see Fig. 1). The latter, which eat the larger phytoplankton cells and also the microzooplankton, are simply not numerically abundant enough to be captured by the water samplers. The foodweb is a highly non-linear, interconnected system, and eliminating one factor has a cascading effect on all the linkages, profoundly changing the character of the system (Fig. 1).
A second problem with these types of bottle experiments is that no matter how carefully one sets up the experiment, one inevitably introduces contaminating iron into the bottles. Ships are very "dirty" platforms with respect to trace metals, and setting up these experiments requires a major investment in clean rooms, hoods, Teflon-coated sampling gear, ultra-clean glassware, and more. It also requires a firm conviction that one can actually be clean enough to get meaningful results. Unless you actually measure the iron concentration in the bottles after the experiment is set up, you can never be completely sure that the bottles to which you have intentionally added iron actually have more iron than the control bot
tles. Indeed, in many of Martin's experiments the phytoplankton in the control bottles grew almost as much as those in the bottles with added iron, and iron analyses confirmed that the control bottles often had elevated iron. Thus, only those who have the analytical capability to measure extremely low concentrations of iron in seawater can be assured unequivocal results, and be convincing in this debate.
Indeed, even though Martin's bottle incubations repeatedly showed that the phytoplankton in the iron-enriched bottles were stimulated relative to the controls (a result that is difficult to explain with anything other than the iron-limitation hypothesis) he still had his skeptics (see Chisholm and Morel, 1991). With the dogged determination that was his trademark, he decided to put an end to the debate once and for all and enrich a patch of the ocean with iron to see how the phytoplankton would respond when the food web was intact. Martin calculated that one could easily add more than enough iron to 100 km 2 of ocean to stimulate the phytoplankton, provided there was enough N and P around. As we have already discussed, in the equatorial Pacific and the Southern Ocean, N and P are abundant. Working in the Southern Ocean is a logistical nightmare for oceanographers, and the availability of sunlight is unpredictable, thus Martin decided on the equatorial Pacific (near the Galapagos Islands) as the site for the experiment.
Testing The Iron Hypothesis: The First Open Ocean Fertilization Experiment
Making the Patch. How does one make a patch of something in the turbulent, surface waters of the ocean, and follow it long enough to detect a biological signal? When the iron experiment was being planned, it was not clear if this was even possible—in fact, Martin designed the expedition simply to test if such a patch could be created and followed. The ambient iron concentration at the experimental site was 0.1

1282 CHISHOLM: THE IRON HYPOTHESIS
Figure 5. Comparison of phytoplankton activity inside and outside the patch during the iron fertilization experiment. (A) Average chlorophyll concentrations (a measure of photosynthetic biomass) and (B) average primary production (the rate of photosynthesis) inside and outside the patch between Julian day 299 and 302. The mixed layer, and hence the iron enriched waters, extended down to about 40 m. (Data courtesy of R. Bidigare and R. Barber; after Martin et al., 1994)
nM Fe (= 10"10 moles per liter), and the target concentration for the patch was 4 nM Fe. To this end, 7,800 moles of Fe (as FeS0 4) were added to 15,600 liters of seawater with enough concentrated HC1 to bring it to pH 2 (iron is very insoluble in seawater). This mixture was dispensed into the ship's propeller wash as it steamed back and forth at 9 km hr'1, creating a 64 km 2 patch over a 24-hour period. Horizontal mixing homogenized the surface streaks of iron within one day, and convective overturn of the mixed layer supplied the additional force necessary to mix the iron down to 35 m in the first 24 hours of the experiment (Martin et al., 1994).
The patch steadily drifted in absolute position, thus a buoy equipped with a Global Positioning System (GPS) receiver was deployed at the center of the patch, and served as the ship's reference point throughout the experiment (Fig. 4). In addition, 4 drifter buoys were deployed in the four corners of the patch to keep track of the periphery (Fig. 4). Although these buoys served as a crude reference, they could not be relied upon as indicators of when one was "in" and "out" of the iron-enriched area, because it was impossible to know a priori how much lateral mixing would take place. Since iron was consumed by the phytoplankton throughout the experiment, it could not be used as a tracer for the patch. Thus the inert tracer, sulfur hexafluoride (SF6), was added to the propeller wash along with the iron (Martin et al., 1994).
The patch was followed for 9 days. During the first five days it maintained its integrity surprisingly well (Fig. 4). On the 6th day a low salinity front moved in from the east, subducting the patch to 20 to 40 m. Although it was still track-able under the surface, the experiment was compromised because the subduction subjected the phytoplankton to a light-shift, thus introducing a new variable (Martin et al., 1994).
The Fate of the Added Iron. Iron is very insoluble in seawater, thus precipitates readily, and it is not entirely clear
what forms are "bioavailable"—i.e., available for transport into the phytoplankton cells. "Dissolvable iron," which is operationally defined and easily measured on board ship, was measured regularly in the patch to monitor the fate of at least one fraction of the iron pool over the course of the experiment. The highest concentration in the patch the day after it was initiated was 3.6 nM—in good agreement with the target concentration. This value decreased steadily each day, and had dropped below the detection limit of the method (2 nM) by the 4th day, apparently reflecting a net loss of available iron from the system (Martin et al., 1994). The detailed results of this aspect of the story await the time-consuming analysis of samples for the other iron fractions.
The Response. Three basic measures were used to monitor the response of the bulk phytoplankton community to the addition of iron: (1) Total chlorophyll concentration, which can increase either because the total number of cells has increased, or because the chlorophyll per cell has increased; (2) Primary productivity, which is a measure of the rate of photosynthesis of the collected phytoplankton assemblage; and (3) Photochemical energy conversion efficiency, a relatively new technique that gives us a "snapshot" of the efficiency with which the photosynthetic mechanisms of the phytoplankton are functioning (and by inference, whether or not the cells are iron starved) (Martin et al., 1994; Kolber et al., 1994).
All three measures revealed quite clearly that the phytoplankton in the iron-enriched patch were stimulated when compared with the assemblages outside the patch (Figs 5 & 6). Moreover, the results indicate that all size categories of the phytoplankton were stimulated by the added iron, and stimulated within the first 24 hours of iron addition (Fig. 6). This timing and overall response contrasts strikingly with the results reported from the bottle incubation experiments described above, in which the response always appeared to be
Chlorophyll (mg m"3) Primary Productivity (mg C m~3d~ l) 0.2 0.4 20 40 60

CHISHOLM: THE IRON HYPOTHESIS 1283
0-1 1-5 5-10 >10 Total
Size Fractions (/xm)
Figure 6. Response of various size fractions of the phytoplankton community to the addition of iron compared with those outside the patch, two days following the beginning of iron enrichment. (A) Increases in F v / F m , which is a measure of the quantum efficiency of part of the photosynthetic apparatus in phytoplankton and reaches a maximum of 0.65 under
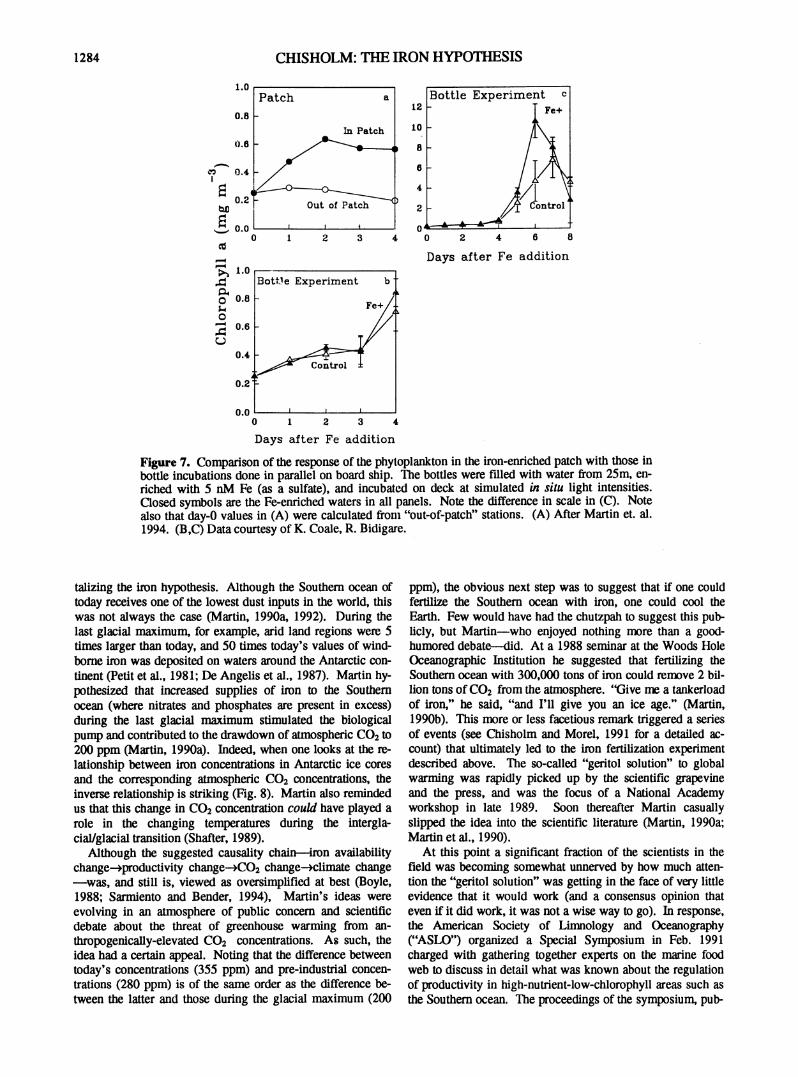
delayed by several days (Fig. 3) and was usually dominated by one particular group—namely the pennate diatoms. Moreover, although the stimulation in the patch was almost instantaneous, the total increase in chlorophyll was rather modest, and appeared to level off after the first two days (Fig. 7a). The total drawdown of C 0 2 in the iron-enriched patch was only 10% of that which should have been possible had all the N and P been assimilated (Watson et al. 1994). As Dr. Richard Barber, the chief scientist on the cruise, so nicely put it, "Apparently the phytoplankton in the patch hadn't read the literature."
Iron-enrichment bottle experiments were done in parallel with the patch experiment, and some striking differences in the phytoplankton response were observed (Fig. 7). Over the first three days of the bottle experiments (Fig. 7b) the control bottles responded to the same degree as the iron-enriched bottles (suggesting that it was inadvertently contaminated with iron), and the magnitude of the response in both treatments was roughly equivalent to that in the patch (Fig. 7a). On the fourth day, however, the chlorophyll in the bottles began to "take off, as has been seen repeatedly in previous experiments (Fig. 3), whereas the chlorophyll concentrations in the patch leveled off at 0.6 mg m*3. After the fourth day we see explosive growth in the bottles (Fig. 7c),
with chlorophyll in the iron-enriched bottle increasing over 10-fold, and reaching more than twice the levels in the control.
One cannot help but wonder if the tremendous growth observed in the bottles by day 6 would have been observed in the patch if it had not been subducted. We suspect not, for several reasons. We know that iron was steadily lost from the patch, and it is likely that the supply ran out before a major bloom could develop. This is supported by the apparent leveling off of chlorophyll concentration in the patch after the second day, and the fact that nitrate concentrations in the water were not depleted significantly. Also, there is reason to suspect that macrozooplankton grazing (which was absent, or significantly reduced, in the bottles) was very active in the patch, repressing the development of higher chlorophyll levels (Martin et al., 1994). The importance of the comparison of the bottle and patch experiments—which is to my mind one of the most significant dimensions of this work—lies in its ability to reveal the degree to which bottle incubations distort our image of how the natural ecosystems function. Much of what we have learned about primary productivity in the sea has come from bottle experiments, thus these comparisons are critical to our understanding the biases in our data.
An expedition is planned for May 1995 in which the IRONEX experiment will be repeated and expanded —continuously fertilizing the patch with iron this time, and carefully examining the role of zooplankton grazing in regulating phytoplankton biomass. We are optimistic that a more complete picture of the dynamics of this ecosystem will emerge with this revised experimental design. The bottom line at this point, however, is that the addition of iron to the equatorial Pacific ecosystem stimulates phytoplankton productivity^—thus the basic premise of the iron hypothesis has been validated.
Science, the Press, and Public Policy This story, as I have told it to this point, is a straightfor
ward account of a recent advance in our understanding of how the oceans work. But there is a very rich subtext accompanying this tale—with lessons for scientists, the agencies that fund them, and the public officials who rely on them for setting public policy.
The part of the story that I have left out has its origins in paleooceanography. By measuring the composition of materials trapped in ice cores from the Antarctic, including the C 0 2
content of ancient air, paleooceanographers can deduce the chemistry of the oceans and atmosphere of past eras. These studies have revealed that during the last glacial maximum, 18,000 years B.P. (before present), C 0 2 levels in the atmosphere were only 200 ppm. This contrasts with today's (pre-industrial) atmospheric concentrations of 280 ppm, and previous interglacial atmospheres (Raynaud et al. 1993) which had similar concentrations. Although most people agree that the deep ocean C 0 2 reservoir must have played a role in glacial/interglacial transitions in atmospheric C0 2 , the mechanism is unclear (Sarmiento and Bender, 1994). One of the leading theories suggests that changes in primary productivity in the Southern ocean played a key role (Knox and McElroy, 1984; Sarmiento and Toggweiler, 1984; Siegenthaler and Wenk, 1984), but explanations for what triggers the changes in productivity have been lacking (Keir, 1988; Toggweiler and Sarmiento, 1985).
These observations were not lost on Martin as he was revi-
1284 CHISHOLM: THE IRON HYPOTHESIS
O O
1 . 0
O.B
0 .6
0 . 4
0 . 2
0 . 0
i
1 . 0
0 . 6
0 . 6
0 . 4
0 . 2 '
0 . 0
Patch
In Patch
Out of Patch
BoUTe E x p e r i m e n t b" t
Fe+/ -
-
^ s ^ * Control
1 2
1 0
8
6
4
2
0
Bottle Experiment c Fe+
0 2 4 6 8
Days after Fe addition
0 1 2 3 4
Days after Fe addition
Figure 7. Comparison of the response of the phytoplankton in the iron-enriched patch with those in bottle incubations done in parallel on board ship. The bottles were Filled with water from 25m, enriched with 5 nM Fe (as a sulfate), and incubated on deck at simulated in situ light intensities. Closed symbols are the Fe-enriched waters in all panels. Note the difference in scale in (C). Note also that day-0 values in (A) were calculated from "out-of-patch" stations. (A) After Martin et. al. 1994. (B,C) Data courtesy of K. Coale, R. Bidigare.
talizing the iron hypothesis. Although the Southern ocean of today receives one of the lowest dust inputs in the world, this was not always the case (Martin, 1990a, 1992). During the last glacial maximum, for example, arid land regions were 5 times larger than today, and 50 times today's values of wind-borne iron was deposited on waters around the Antarctic continent (Petit et al., 1981; De Angelis et al., 1987). Martin hypothesized that increased supplies of iron to the Southern ocean (where nitrates and phosphates are present in excess) during the last glacial maximum stimulated the biological pump and contributed to the drawdown of atmospheric C 0 2 to 200 ppm (Martin, 1990a). Indeed, when one looks at the relationship between iron concentrations in Antarctic ice cores and the corresponding atmospheric C 0 2 concentrations, the inverse relationship is striking (Fig. 8). Martin also reminded us that this change in C 0 2 concentration could have played a role in the changing temperatures during the intergla-cial/glacial transition (Shatter, 1989).
Although the suggested causality chain—iron availability change-»productivity change-»C0 2 change-Klimate change —was, and still is, viewed as oversimplified at best (Boyle, 1988; Sarmiento and Bender, 1994), Martin's ideas were evolving in an atmosphere of public concern and scientific debate about the threat of greenhouse warming from an-thropogenically-elevated C 0 2 concentrations. As such, the idea had a certain appeal. Noting that the difference between today's concentrations (355 ppm) and pre-industrial concentrations (280 ppm) is of the same order as the difference between the latter and those during the glacial maximum (200
ppm), the obvious next step was to suggest that if one could fertilize the Southern ocean with iron, one could cool the Earth. Few would have had the chutzpah to suggest this publicly, but Martin—who enjoyed nothing more than a good-humored debate—did. At a 1988 seminar at the Woods Hole Oceanographic Institution he suggested that fertilizing the Southern ocean with 300,000 tons of iron could remove 2 billion tons of C 0 2 from the atmosphere, "Give me a tankerload of iron," he said, "and I'll give you an ice age." (Martin, 1990b). This more or less facetious remark triggered a series of events (see Chisholm and Morel, 1991 for a detailed account) that ultimately led to the iron fertilization experiment described above. The so-called "geritol solution" to global warming was rapidly picked up by the scientific grapevine and the press, and was the focus of a National Academy workshop in late 1989. Soon thereafter Martin casually slipped the idea into the scientific literature (Martin, 1990a; Martin et al., 1990).
At this point a significant fraction of the scientists in the field was becoming somewhat unnerved by how much attention the "geritol solution" was getting in the face of very little evidence that it would work (and a consensus opinion that even if it did work, it was not a wise way to go). In response, the American Society of Limnology and Oceanography ("ASLO") organized a Special Symposium in Feb. 1991 charged with gathering together experts on the marine food web to discuss in detail what was known about the regulation of productivity in high-nutrient-low-chlorophyll areas such as the Southern ocean. The proceedings of the symposium, pub-

CHISHOLM: THE IRON HYPOTHESIS 1285
300
> 250
6 d CM
O 200
Mean Age of Ice (x 10 3y BP) 2 0 4 0 6 0 8 0 100 1 2 0 140 160
150
1 1 1 1 1 1 1 1 i i i i i i i / »
\ 1 V f |
• V * A
C O . i i 2 | i / 1 1 \A K y a i . \A
K M ' v V -
F e I \ » \ ( \ 1 I\i V\ - 1 \ / v ' i l l ' A \
1 A v M M / ' ^ ^ Y^S, r ^ . / ^ N / 1
2.0
1.5 • (30
fe 1.0 o
6
0.5
10 15 20
Depth (x 100m) Figure 8* Iron and C 0 2 levels in an ice core from the Antarctic, showing an inverse correlation through two glacial/interglacial transitions. Martin hypothesized that the elevated iron concentrations stimulated primary productivity in the waters around the Antarctic, thus removing significant amounts of C 0 2 from the atmosphere (adapted from Martin, 1992). Ice core ages are in years before present".
lished as a special issue of Limnology and Oceanography (Chisholm and Morel, 1991), reveal the true complexity of these ecosystems, and the difficulties involved in arriving at a consensus about what regulates them. The group concluded that the evidence for iron playing a role in regulating productivity in the HNLC areas of the oceans was compelling enough to warrant a small-scale fertilization experiment. They also drafted and adopted a resolution urging "...all governments to regard the role of iron in marine productivity as an area for further research and not to consider iron fertilization as a policy option that significantly changes the need to reduce emissions of carbon dioxide."
Thus the stage was set for what has come to be called the "IRONEX" experiment, the results of which I have summarized above. Martin had to secure the funding for the expedition, and there is no doubt that having the endorsement of the broader oceanographic community smoothed the way for this. Also, since the idea of an ocean fertilization experiment was no longer enmeshed in the more loaded issue of climate engineering, it could develop independently of the potential consequences of its success.
Looking back, however, we cannot help but wonder if this experiment would have come about had John Martin not thrown the climate change question into the public eye. His casting the hypothesis in a context relevant to public concerns catalyzed the scientific community—not the public—to take careful stock of our understanding of the regulation of open ocean ecosystems. This drew the attention of the federal funding agencies (who were involved in the ASLO Symposium), giving them an opportunity to witness the broadest possible "peer review" before endorsing the experiment. The experiment was a success beyond everyone's expectations (except, perhaps, John Martin's), and the issue of engineering climate has receded from view, for now. I am sure the debate will be rekindled sometime in the future, but by then our understanding of how the marine food web is regulated will be more advanced—through insights gained from this, and other, ocean fertilization experiments.
Acknowledgments. This paper is dedicated to the late John Martin, whom I thank for his friendship, inspiration, and wonderful good humor throughout all the struggles. I thank Sara Tanner and Steve Htzwater for sampling help. The bottle experiments reported from the IRONEX cruise were designed and orchestrated by Kenneth Coale, and the pigment data were supplied by Robert Bidigare. Thanks also to Zackary Johnson and Kent Bares for help with the data analysis and graphics, and Edmund Carlevale for editorial advice. This effort was supported in part by grants from NSF, ONR, and the EPA Global Change Research Program (IN-0826-NAEX).
References Boyle, E.A., Vertical oceanic nutrient fractionation and gla
cial/interglacial C0 2 cycles, Nature, 331, 55-56,1988. Brand, L.E, Minimum iron requirements of marine phytoplankton
and the implications for the biogeochemical control of new production, Limnol Oceanogr., 36,1756-1771,1991.
Broecker, W.S., Chemical Oceanography, Harcourt, Brace and Jova-novich, Orlando, Fla., p. 39,1974.
Chisholm, S.W. and F.M.M. Morel, (Eds.), What controls phytoplankton production in nutrient-rich areas of the open sea? Limnol Oceanogr. (Special Issue), 36(8), 1991.
Cullen, J., Hypotheses to explain high-nutrient conditions in the open sea, Umnol Oceanogr., 36,1579-1599,1991.
De Angelis, M., N.I. Barkov, and V.N. Petrov, Aerosol concentrations over the last climatic cycle (160 kyr) from an Antarctic ice core, Nature, 325,318-321,1987.
Htzwater, S. and C. Hunter, John Martin's Legacy: An open-ocean test of the iron hypothesis, USJGOFSNews, 5(3), 1,1994.
Gran, H.H, On the conditions for the production of plankton in the sea, Rapp. Proc. Verb. Cons. Int. Explor. Mer., 75,37-46,1931.
Hart, T.J., On the phytoplankton of the southwest Atlantic and the Bellingshausen Sea 1921-31, Discovery Reports, VIII, 1-268, 1934.
Harvey, H. W., The supply of iron to diatoms, J. Mar. Biol. Assoc. U.K., 22,205-219,1938.
Hudson, R.J.M. and F.M.M. Morel, Iron transport in marine phytoplankton: Kinetics of cellular and medium coordination reactions, Limnol. Oceanogr., 35,1002-1020,1990.
Keir, R.S., On the late Pleistocene ocean geochemistry and circulation, Paleoceanography, 3,413-445,1988.
Knox, F. and M. McElroy, Changes in atmospheric C0 2, influence of marine biota at high latitudes, J. Geophys Res., 89, 4629-4637, 1984.

1286 CHISHOLM: THE IRON HYPOTHESIS
Kolbcr, Z.S., R.T. Barber, K.H. Coale, S.E Rtzwater, RM. Greene, K.S. Johnson, S. Lincfley, and P. G. Falkowski, Iron limits photosynthetic energy conversion efficiency in the equatorial Pacific Ocean, Nature, 371,145-148,1994.
Martin, J.H, Glacial-Interglacial C0 2 change: The iron hypothesis, Paleoceanography, 5,1-13,1990a.
Martin, J.H., A new iron age, or a ferric fantasy, U.S. JGOFS News, 1(4) 5,11,1990b.
Martin, J.H., Iron as a limiting factor, in Primary Productivity and Biogeochemical Cycles in the Sea, edited by P.G. Falkowski and A. Woodhead, pp. 123-137, Plenum Press, New York, NY., 1992.
Martin, J.H., S.E. Fitzwater, R.M. Gordon, C.N. Hunter, and S.J. Tanner, Iron, primary production and carbon-nitrogen flux studies during JGOFS North Atlantic bloom experiment, Deep Sea Res. II, 40,115-134,1993.
Martin, J.H. and R.M. Gordon, Northeast Pacific iron distributions in relation to phytoplankton productivity, Deep Sea Res., 35, 177-196,1988.
Martin, J.H. and S.E. Rtzwater, Iron deficiency limits phytoplankton growth in the north-east Pacific subarctic, Nature, 331, 341-343, 1988.
Martin, J.H., K.H. Coale, K.S. Johnson, S.E. Fitzwater, R.M. Gordon, S.J. Tanner, C.N. Hunter, V.A. Elrod, J.L. Nowicki, T.L Coley, R.T. Barber, S. Lindley, A.J. Watson, K. Van Scoy, CS. Law, M.I. Liddicoat, R. Ling, T. Stanton, J. Stockel, C. Collins, A. Anderson, R. Bidigare, M. Ondrusek, M. Latasa, F.J. Millero, K. Lee, W. Yao, J.Z. Zhang, G. Fredrich, C. Sakamoto, F. Chavez, K. Buck, Z. Kolber, R. Green, P.G. Falkowski, S.W. Chisholm, F. Hoge, R. Swift, J. Yungle, S. Turner, PI Nightingale, A. Hatton, P. Liss, N.W. lindale, Testing the iron hypothesis in ecosystems of the equatorial Pacific, Nature, 371, 123-129, 1994.
Martin, J.H., M. Gordon and S. Fitzwater, Iron in Antarctic waters, Nature, 345,156-158,1990.
Petit, J-R., M. Briat and A. Royer, Ice age aerosol content from East Antarctic ice core samples and past wind strength, Nature, 291-294,1981.
Prospero, J.M., Eolian transport to the world oceans, in The Sea, v. 7, edited by C. Erniliani, pp. 801-874, Wiley, New York, N.Y., 1981.
Redfield, A.C., On the proportions of organic derivations in sea water and their relation to the composition of plankton, James Johnston Memorial Volume, pp. 176-192, Liverpool, 1934.
Redfield, A.C., The biological control of the chemical factors in the environment, Amer. Sci., 46(3), 1-18,1958.
Richards, F.A., Chemical and biological factors in the marine environment, in Ocean Engineering, edited by J.F. Brahtz, pp. 259-303, Wiley, New York, NY., 1968.
Raynaud, D., J. Jouzel, J.M. Barnold, J. Chappellaz, R.J. Delmas, and C. Lorius, The ice record of greenhouse gasses, Science, 59, 926-934,1993.
Sarmiento, J.L. and J.R. Toggweiler, A new model for the role of the oceans in determining atmospheric P002, Nature, 308, 621-624, 1984.
Sarmiento, J.L. and M. Bender, Carbon biogeochemistry and climate change, Photosyn. Res., 39,209-234,1994.
Shatter, G., A model of biogeochemical cycling of phosphorus, nitrogen, oxygen, and sulfur in the ocean: One step toward a global climate model, J. Geophys. Res., 94,1979-2004,1989.
Siegenthaler, U. and T. Wenk, Rapid atmospheric CO2 variations and ocean circulation, Nature, 308,624-625,1984.
Toggweiler, J.R. and J.L. Sarmiento, Glacial to interglacial changes in atmospheric carbon dioxide: The critical role of ocean surface water in high latitudes, in Natural Variations Archean to Present Geophys. Monogr, v. 32, edited by E.T. Sundquist and W.S. Broecker, pp. 163-184, AGU, Washington, D.C, 1985.
Uematsu, M., Study of the continental material transported through the atmosphere to the ocean, J. Oceanogr. Soc. Japan, 43, 395-401,1987.
Watson, AJ., CS. Law, K.A. Van Scoy, F.J. Millero, W. Yao, G.E Friederich, M.I. Liddicoat, RH. Wannikhof, R.T. Barber, and K.H. Coale, Minimal effect of iron fertilization on sea-surface carbon dioxide concentrations, Nature, 371,143-145,1994.
S.W. Chisholm, 48-425, Massachusetts Institute of Technology, Cambridge, MA 02139. (e-mail:[email protected])
(Received August 24,1994; accepted December 23,1994.)





























