Embed Size (px)
Citation preview

ARTICLE IN PRESS
Immunobiology 213 (2008) 377–387
0171-2985/$ - se
doi:10.1016/j.im
Abbrevations
interferon; iNO
dendritic cell; N
oligodesoxynuc
peritoneal exud
TLR, Toll-like�Correspond
Immunology
Wasserturmstra
Tel.: +499131
E-mail addr
www.elsevier.de/imbio
The innate immune response against Leishmania parasites
Jan Liesea, Ulrike Schleichera,b, Christian Bogdana,b,�
aDepartment of Medical Microbiology and Hygiene, Institute of Medical Microbiology and Hygiene, University Clinic Freiburg,
Freiburg, GermanybInstitute of Clinical Microbiology, Immunology and Hygiene, University Clinic of Erlangen, Wasserturmstraße 3-5,
D-91054, Erlangen, Germany
Received 31 October 2007; received in revised form 19 December 2007; accepted 21 December 2007
Abstract
Parasites of the genus Leishmania are the causative agents of cutaneous, mucocutaneous or visceral leishmaniasis.The parasite species and host genetic factors determine the quality of the immune response and thereby the outcome ofthe infection. Here, we summarize previously published and present novel data on several aspects of the early innateimmune reaction to Leishmania (L.) major, L. braziliensis and L. infantum, which cause cutaneous, mucocutaneous orvisceral leishmaniasis, respectively. We will focus on (1) the effector molecules that contribute to the control of theparasite in the skin, lymph nodes and/or spleen; and (2) on the pattern recognition receptors (Toll-like receptors,TLRs), cell types (myeloid dendritic cells, plasmacytoid dendritic cells), cytokines (IL-12, IFN-a/b), and signalingpathways (Tyk2 kinase) that are necessary for the initial sensing of the parasites and the subsequent development of anefficient NK cell response.r 2008 Elsevier GmbH. All rights reserved.
Keywords: Parasite/protozoan infection; Leishmania; NK cells; Toll-like receptors; Type I interferon
Introduction
Leishmania are protozoan parasites that in nature aretransmitted by bites of sand flies. The vectors inject theflagellated, promastigote form of the parasite into the
e front matter r 2008 Elsevier GmbH. All rights reserved.
bio.2007.12.005
: BMMF, bone marrow-derived macrophages; IFN,
S, inducible nitric oxide synthase; mDC, myeloid
K cell, natural killer cell; NO, nitric oxide; ODN,
leotide; pDC, plasmacytoid dendritic cell; PE-MF,ate macrophages; phox, phagocyte NADPH oxidase;
receptor.
ing author at: Institute of Clinical Microbiology,
and Hygiene, University Clinic of Erlangen,
ße 3-5, D-91054, Erlangen, Germany.
852 2552; fax: +499131 852 2573.
ess: [email protected] (C. Bogdan).
dermis of the host, where it is rapidly taken up byphagocytic cells (e.g. neutrophils, macrophages andimmature dendritic cells) and transforms into thereplicative, intracellular form termed amastigotes. De-pending on the parasite species they either elicitcutaneous leishmaniasis (members of the subgenusLeishmania, e.g. L. major, L. tropica, L. mexicana;members of the subgenus Viannia, e.g. L. braziliensis),mucocutaneous leishmaniasis (e.g. L. braziliensis) orvisceral leishmaniasis (e.g. L. infantum (identical to L.
chagasi), L. donovani) (Chappuis et al. 2007; Lukes et al.2007; Pearson and de Queiroz Sousa 1996)
Cutaneous leishmaniasis is characterized by a tran-sient skin lesion (papule or ulcer) that is self-healing inimmunocompetent hosts, although the parasites willpersist in small quantities throughout life (Bogdan et al.

ARTICLE IN PRESSJ. Liese et al. / Immunobiology 213 (2008) 377–387378
1996, 2000; Mendonca et al. 2004; Schubach et al.1998). The mouse model of this infection has beenwidely used in the past to define immunologicalmechanisms that contribute to the control of thisintracellular pathogen. It was shown that immunecells (macrophages, dendritic cells (DCs), natural killer(NK) cells, CD4+ and CD8+ T cells), cytokines(interferon (IFN)-g and interleukin (IL)-12), and effec-tor molecules (nitric oxide (NO) produced by inducibleNO synthase (iNOS)) are key components in theimmune response to the parasite (Bogdan et al. 1993;Solbach and Laskay 2000). The genetic background ofthe murine host has a significant influence on theoutcome of the infection: whereas in most mousestrains (e.g. C57BL/6) the local (i.e. cutaneous) inocula-tion with L. major promastigotes only leads to atemporary swelling of the skin that resolves on itsown, some mouse strains (e.g. BALB/c) develop severenon-healing skin ulcers that are accompanied by arapid systemic spreading of the parasites and anultimately fatal visceral disease (Sacks and Noben-Trauth 2002).
Visceral leishmaniasis of humans can also bemimicked in a mouse model. In both C57BL/6 andBALB/c mice, the intravenous injection of L. infantum
will lead to the enlargement of spleen and liver resultingfrom massive parasite loads in these organs. Whereasthe parasite burden in the liver will eventually decline, itwill remain at very high levels in the spleen (Leclercqet al. 1996; Smelt et al. 1997). Similar to L. major
infections, the control of visceral leishmaniasis isalso dependent on IL-12 and the development ofIFN-g-producing CD4+ T cells, CD8+ T cells andNK cells (Kirkpatrick and Farrell 1982; Murray 1997;Murray et al. 2005).
In our projects on the innate immune response toLeishmania parasites, we utilized the mouse models ofcutaneous and visceral leishmaniasis to study theparameters that are required for an efficient innateimmune response to L. major, L. braziliensis andL. infantum. Specifically, we investigated (1) the effectormolecules (iNOS and phagocyte NADPH oxidase(phox)) that exert the antileishmanial activity ofphagocytes; and (2) the pattern recognition receptors(Toll-like receptors, TLRs), cell types (myeloid dendriticcells (mDCs), plasmacytoid dendritic cells (pDCs)),cytokines (IL-12, IFN-a/b), and signaling pathways(Tyk2 kinase, signal transducer and activator oftranscription 4 (STAT4)) that are necessary forthe development of an efficient NK cell response. Weused a variety of in vitro and in vivo strategies to analyzethe contributions of these single elements to theconcerted immune response against Leishmania. Thepresent article summarizes recently published andpresents previously unpublished data, which are dis-cussed in the context of the current literature.
Materials and methods
Mice
C57BL/6 mice were purchased from Charles RiverBreeding Laboratories (Sulzfeld, Germany). All usedmice were 7–8 weeks of age and held under specificpathogen-free conditions in the animal facilities of ourinstitute. All animal experiments were approved by thegovernmental animal welfare committee of the Regier-ungsprasidium Freiburg.
Isolation of macrophages
Thioglycolate-elicited peritoneal exudate macro-phages (PE-MF) were prepared from the peritonealcavity of C57BL/6 mice 4 days after i.p. injection of 3ml4% Brewer’s thioglycolate broth as described before(Bogdan et al. 1997; Schleicher et al. 2005). Cells wereresuspended in RPMI1640 culture medium (supplemen-ted with 2mM L-glutamine, 23.8mM NaHCO3, 10mMHEPES, 50 mM 2-mercaptoethanol) plus 2.5% FCS(Sigma, Deisenhofen, Germany), and 6� 106 cellswere seeded in 60mm diameter tissue culture dishes(Greiner BioOne, Frickenhausen, Germany). Afterculture at 37 1C in 5% CO2 and 95% humidified airfor 90–120min, nonadherent cells were washed off andthe remaining macrophage monolayer was furtherincubated in fresh medium together with the respectivestimuli (CpG oligodesoxynucleotide (ODN) 1668 (1 mM,Thermo Electron, Ulm, Germany), LPS Escherichia coli
O111:B4 (200 ng/mL; Sigma-Aldrich, Deisenhofen,Germany), poly(I:C) (50 ng/mL, Sigma), rmIFN-g(20 ng/mL, provided by G. Adolf, Vienna, Austria)) asdetailed in the figure legends.
Parasites and infection
The propagation and preparation of promastigotes ofthe L. major strain MHOM/IL/81/FEBNI was de-scribed before (Bogdan et al. 2000). For use in theexperiments, parasites were expanded for several days toreach stationary growth phase.
Cytokine mRNA expression analysis
Total RNA from cells was extracted using theTRIZOL reagent (Invitrogen, Karlsruhe, Germany).Contaminating genomic DNA was removed withDNase I (DNAfree, Ambion, Austin, USA) beforeRNA was reverse transcribed using the high capacitycDNA Archive Kit (Applied Biosystems, Darmstadt,Germany). To determine the amount of target genelevels the HT7900 quantitative PCR system (AppliedBiosystems) was used. Each cDNA was measured in

ARTICLE IN PRESSJ. Liese et al. / Immunobiology 213 (2008) 377–387 379
triplicates with the following gene-specific assays (Taq-Man Gene Expression Assays, Applied Biosystems):IFN-a2 (Mm00833961_s1), IFN-a4 (Mm00833969_s1),IFN-a5 (Mm00833976_s1), IFN-a9 (Mm00833983_s1),IFN-a11 (Mm01257312_s1), IFN-a12 (Mm00616656_s1), IFN-a13 (Mm00781548_s1), and IFN-b(Mm00439546_s1). The gene for mouse hypoxanthineguanine phosphoribosyl transferase-1 (HPRT-1,Mm00446968_m1) was used as an endogenouscontrol for calibration of the mRNA levels. Quantitativeanalysis was performed using the SDS 2.1 soft-ware (Applied Biosystems). mRNA levels were calcu-lated by the following formula: relative expression¼ 2�ðCTðTargetÞ�CTðEndogenous controlÞÞ � f , with f ¼ 104 as anarbitrary factor.
Type I IFN protein assay
IFN-a/b in supernatants was determined with theVSV/L929 bioassay as described previously (Diefenbachet al. 1998). Briefly, L929 fibroblasts were incubatedwith culture supernatants or a standard of purifiedmouse IFN-a/b (generously provided by Dr. I. Gresser,Institute Curie, Paris, France) overnight, followed bytreatment with vesicular stomatitis virus (VSV) for 2days. Subsequently, 3-(4,5-dimethylthiazol-2-yl)-2,5-di-phenyltetrazolium bromide (MTT; Sigma) was addedand cells were lysed with 10% SDS/HCl after 8 h offurther incubation. On the next day absorption atl ¼ 550 nm was determined for triplicates using aSpectraMax 340PC384 ELISA plate reader (MolecularDevices, Ismaning, Germany) and the concentration ofIFN-a/b was calculated by comparison with thestandard curve using the SoftMax Pro 4.6 software(Molecular Devices).
Results and discussion
Effector molecules
Inducible nitric oxide synthase
Production of nitric oxide (NO) by inducible nitric-oxide synthase (iNOS, also referred to as NOS2) isindispensable for the clinical resolution of a cutaneousL. major infection (Diefenbach et al. 1998; Liew et al.1990) as well as for maintaining life-long control ofpersisting Leishmania parasites (Stenger et al. 1996). InL. major-infected mice, we found that the antileishma-nial effect of iNOS was most prominent in the skin,somewhat less impressive in the lymph node, and veryweak in the spleen (Blos et al. 2003). We also madesimilar observations with L. braziliensis-infected mice(Soares Rocha et al. 2007).
In L. donovani-infected mice, iNOS was initiallyreported to be essential for the control of the parasitesin the liver (Murray and Nathan 1999). However, in asubsequent study, in which wild-type and iNOS knock-out mice were both carrying the resistance allele of thesolute carrier family 11a member 1 (Slc11a1, formerlyalso termed natural resistance-associated macrophageprotein-1 or NRAMP-1), iNOS exerted only a minor ofdegree of antileishmanial activity in the liver as well as inthe spleen (White et al. 2005). This is in accordancewith the limited expression of iNOS in the spleen ofL. donovani-infected mice (Melby et al. 1998). Studies, inwhich L. donovani-infected mice deficient for iNOS andthe phagocyte NADPH oxidase (phox) were treatedwith amphotericin B, revealed that the control ofresidual parasites in the liver neither required iNOSnor phox, which indicates the existence of an iNOS/phox-independent macrophage mechanism to restrictreplication of L. donovani in the liver (Murray 2005,2006). Although L. major, L. braziliensis, andL. donovani are clearly susceptible to NO-mediatedkilling by macrophages in vitro (Blos et al. 2003;Diefenbach et al. 1998; Perez et al. 2006; Roach et al.1991; Soares Rocha et al. 2007), the in vivo data stronglysuggest a tissue-specific expression of anti-Leishmania
control mechanisms.In addition to its antimicrobial effector functions
iNOS fulfills numerous signaling functions and con-tributes to the regulation of cytokine gene expressionand cytokine responsiveness (reviewed in Bogdan2001a, b). In the L. major model, we found that iNOS-derived NO functions as a cofactor for the activation ofTyk2 kinase and the induction of NK cell cytotoxicityand IFN-g production by IL-12 (Diefenbach et al. 1998,1999).
The key cytokine for the induction of iNOS is IFN-g(Dalton et al. 1993; Huang et al. 1993; Kamijo et al.1993). However, using a neutralizing sheep-anti-IFN-a/b antiserum IFN-a/b rather than IFN-g was shown toaccount for the initial induction of iNOS in the skin andlymph node at day 1 of infection with L. major
(Diefenbach et al. 1998). NO production was notdetectable in macrophages after stimulation with eitherL. major promastigotes or IFN-a/b alone, but cotreat-ment with both stimuli led to NO release (Mattner et al.2000). Surprisingly, we found that the extent to whichiNOS mRNA and protein is induced by type I IFNand L. major is affected by the sequence of thetwo stimuli: treatment of macrophages with IFN-a/bat the time of infection (co-treatment) or after infectionled to the expression of iNOS and NO. In contrast,treatment of macrophages with IFN-a/b prior toinfection with L. major (pre-treatment) rendered themacrophages iNOS-negative (Mattner et al. 2000),which underlines the potential inhibitory capacities oftype I IFN.

ARTICLE IN PRESSJ. Liese et al. / Immunobiology 213 (2008) 377–387380
Phagocyte NADPH oxidase (phox)
In vitro Leishmania parasites were reported to besensitive to reactive oxygen intermediates generated byneutrophils or macrophages (reviewed in Bogdan 2007).As iNOS was largely dispensable for the control ofL. major or L. brazliensis in the spleen (Blos et al. 2003;Diefenbach et al. 1998; Soares Rocha et al. 2007), weconsidered that the multimeric phagocyte NADPHoxidase (phox), which converts molecular oxygen intosuperoxide anions (O2
�), contributes to the defenseagainst Leishmania in visceral organs. In vitro, we foundthat killing of L. major or L. braziliensis amastigote orpromastigote parasites by bone marrow-derived orperitoneal exudate macrophages that lack the catalyticsubunit of phox (gp91phox�/�) was not impaired (Bloset al. 2003; Soares Rocha et al. 2007). Similarly, theinfection rates of neutrophils, which are known for theirprominent expression of phox, were comparable in wild-type and gp91phox�/� cells (Blos et al. 2003). In vivo
wild-type and knockout mice exhibited skin lesions ofequal severity after subcutaneous infection with L. major
or L. braziliensis. However, long-term relapses ofskin swelling were observed in some gp91phox�/� miceafter high-dose infection with L. major, which didnot occur in wild-type controls or in mice infected withL. braziliensis (Blos et al. 2003; Soares Rocha et al.2007). Compared to wild-type mice the parasite burdensof gp91phox�/� mice in the skin, in the lymph node, and– most prominently – in the spleen, were significantlyelevated during the acute and also during the chronicphase of L. major infection (Blos et al. 2003), which wedid not observe in the case of L. brazilinesis-infectedmice (Soares Rocha et al. 2007). phox deficiency did notalter the generation of a protective immune response asassessed by comparison of wild-type and gp91phox�/�
mice for the cell composition, cytokine production(TNF, IFN-g) and expression of iNOS protein in theskin, lymph nodes and/or spleen (Blos et al. 2003; SoaresRocha et al. 2007). Collectively, our data demonstratethat phox contributes to the reduction of parasitenumbers in L. major infection in a site- and stage-specific manner.
In L. donovani-infected mice phox was largelydispensable for the control of the parasites in the spleenand liver (Murray and Nathan 1999; White et al. 2005).
Activation of natural killer cells
Cytokines and signaling molecules
IL-12. IL-12 is a key cytokine for the generation ofprotective immunity to an infection with L. major. Ininitial studies using neutralizing anti-IL-12 antiseraendogenous IL-12 was shown to acivate NK cells andCD8+ T cells and to drive the development of a Th1type immune response, which leads to the production of
IFN-g (Scharton-Kersten et al. 1995; Trinchieri 2003).Conversely, treatment of mice with IL-12 enhanced theinnate NK cell activity (Laskay et al. 1995; Scharton-Kersten et al. 1995; Schleicher et al. 2004) and protectedBALB/c from an otherwise lethal L. major infection(reviewed in Bogdan et al. 1993; Sacks and Noben-Trauth 2002). Bioactive IL-12p70 protein is a hetero-dimer consisting of the p35 and the p40 subunits.Studies of L. major infection in mice that lack either ofthe two subunits have provided evidence that thepresence of IL-12 is crucial to mount a protectiveimmune response against the parasite and to control andheal an infection (Mattner et al. 1997, 2004). We madecomparable observations with L. braziliensis-infectedmice (Soares Rocha et al. 2007). In IL-12p35�/� orIL-12p35/p40�/� mice the early NK cell response(IFN-g production; cytotoxicity against tumor targetcells) to L. major or L. infantum was absent, whichclearly demonstrates that IL-12 is essential for triggeringNK cell activity in vivo (Liese et al. 2007; Schleicher et al.2007).
In vitro we found that bone marrow-derived mDCsrelease IL-12 p40, but no IFN-a/b, after exposure toL. infantum or L. major (Liese et al. 2007; Schleicheret al. 2007). In the model of visceral leishmaniasis wedemonstrated that splenic DCs become IL-12p40/p70+
after injection of L. infantum (Schleicher et al. 2007).
Tyk2. Tyk2 belongs to the family of Janus tyrosinekinases and is involved in the intracellular signalingpathways of interferons (e.g. IFN-a/b) and cytokines(e.g. IL-12) as it associates with the respective receptors(Schindler et al. 2007). Tyk2-deficient macrophages andmice were found to exhibit an enhanced resistance toLPS, which most likely is due to the fact that LPS effectsare mediated in part by the generation of endogenousIFN-a/b (Kamezaki et al. 2004; Karaghiosoff et al.2000, 2003). Tyk2 deficiency led to an impairment of thedevelopment of Th1 cells, although to a variable extent(Karaghiosoff et al. 2000; Minegishi et al. 2006;Schleicher et al. 2004; Shimoda et al. 2000; Tokumasaet al. 2007). In vitro, the IL-12-driven differentiation ofCD4+ T cells into Th1 cells was delayed in the absenceof Tyk2 (Schleicher et al. 2004). In vivo, indirect effectsmediated by Tyk2-deficient dendritic cells and theirreduced production of IL-12 might also contribute tothe impaired Th1 cell response (Aizu et al. 2006;Tokumasa et al. 2007).
We investigated the role of Tyk2 kinase in the mousemodel of cutaneous leishmaniasis (L. major). Tyk2�/�
mice developed more severe footpad lesions comparedto wild-type controls, which was paralleled by increasedparasite burdens in the skin, draining lymph node, andthe spleen. Nevertheless, the partly necrotizing skinlesions finally healed with scar formation (Schleicheret al. 2004). Similar to IL-12p35�/� or IL-12p35/p40�/�

ARTICLE IN PRESSJ. Liese et al. / Immunobiology 213 (2008) 377–387 381
mice, Tyk2�/� mice were unable to mount a NK cellresponse at day 1 of infection, not even after adminis-tration of exogenous IL-12. In contrast, treatment withIFN-a/b induced activation of NK cells, but not to thesame extent as seen in wild-type mice (Schleicher et al.2004). These data show that both IL-12 and Tyk2 areessential for the early NK cell cytotoxicity and IFN-gproduction in L. major-infected mice. Importantly,however, at later time-points of infection (e.g. day 30,day 250), the production of IFN-g by NK cells becameindependent of Tyk2. Whereas CD4+ T cells taken fromL. major-infected Tyk2�/� mice at different time-pointsshowed a normal response to IL-12 or IL-12 plus IL-18after restimulation in vitro, this was not the case inCD8+ T cells, which failed to express IFN-g and tophosphorylate STAT4 (Schleicher et al. 2004). Wetherefore conclude that the defects observed in NK cellsand CD8+ T cells largely account for the exacerbatedcourse of infection in L. major-infected Tyk2�/� mice,whereas the ultimate clinical healing of these mice ismost likely mediated by IFN-g+ CD4+ T cells.
Type I interferons. A broad range of cells will respondto an appropriate stimulus with the production of type Iinterferons. This includes cells of the immune systemsuch as neutrophils, mDCs, plasmacytoid dendritic cells(pDCs), NK cells, and T cells (reviewed in Bogdan et al.2004). pDCs are somewhat unique inasmuch theyproduce very high amounts of IFN-a/b in men (Cellaet al. 1999; Siegal et al. 1999) and mice (Asselin-Paturelet al. 2001; Nakano et al. 2001). Therefore, these cellsare also referred to as high-IFN-a/b-producing cells(IPCs).
Type I IFNs were first recognized for their role inviral infections, but convincing evidence now exists thatdocuments their function in innate and acquiredimmunity to bacterial and parasitic infections (Bogdanet al. 2004; Decker et al. 2005). One major goal of ourprojects was to analyze the expression and function ofIFN-a/b during the immune response to Leishmania
parasites.
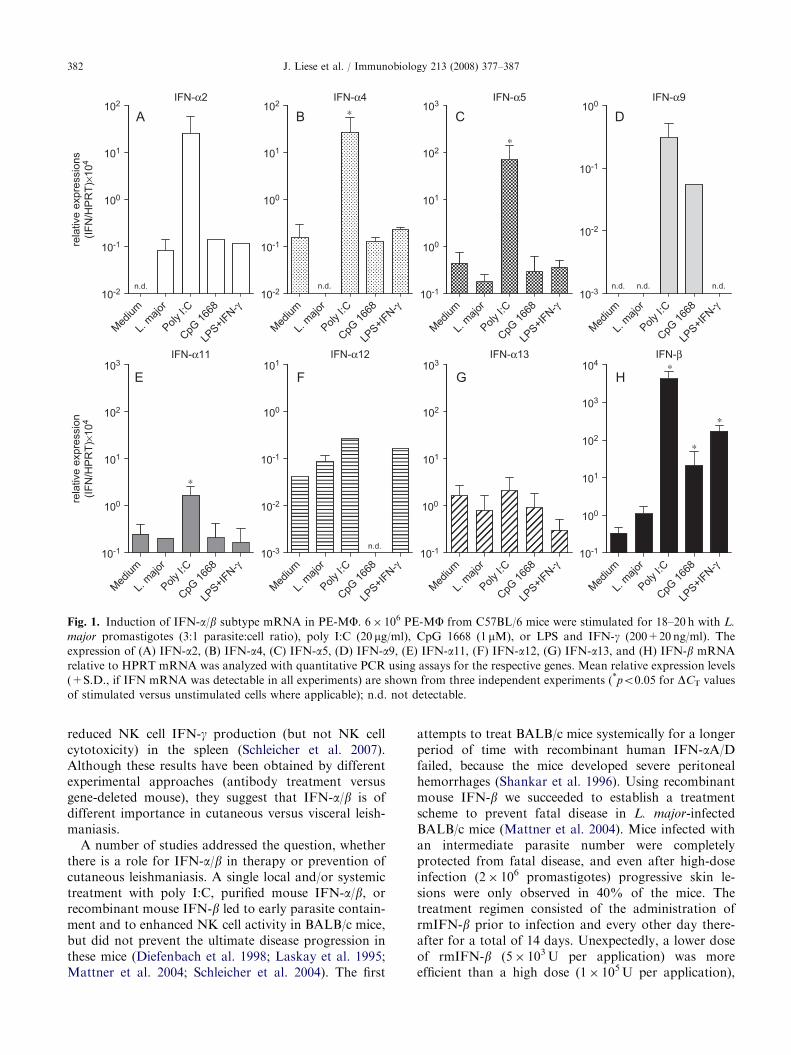
IFN-a/b in vitro. When we stimulated bone-marrowderived macrophages (BMMFs) or peritoneal exudatemacrophages (PE-MF) from C57BL/6 mice withL. major promastigotes for 18 h, we found no consistentand significant induction of the mRNA of the variousIFN-a/b subtypes (with the possible exception ofIFN-a2). In contrast, the cells strongly responded tothe TLR3 ligand poly I:C (up-regulation of IFN-a2, -a4,-a5, -a9, and -b mRNA; weak or no increase of IFN-a11, -a12 and -a13 mRNA) and somewhat less to CpG1668 ODN and LPS (induction or up-regulation of IFN-a2 and IFN-b mRNA, respectively) (Figs. 1, 3A and B).Poly I:C, but not L. major promastigotes, led to theaccumulation of IFN-a/b in the culture supernatants of
macrophages as detected by bioassay (Fig. 2A and B).In a previous study we had detected small quantities ofIFN-a/b bioactivity in the supernatants of PE-MFstimulated with L. major promastigotes for 24 h(Diefenbach et al. 1998). However, in this earlieranalysis, the macrophages were derived from a differentmouse strain (129/SvEv�C57BL/6 mice) and the VSVassay used had a considerably higher sensitivity(ca. 1–10U IFN-a/b/mL versus 10–100U IFN-a/b/mLin the present study) (data not shown). Future studieswill address this discrepancy by (a) direct comparison ofthe effect of L. major promastigotes on macrophagesderived from different mouse strains and (b) byperforming detailed time-course experiments.
During our search for cells that produce IFN-a/b inresponse to Leishmania parasites, we also investigated indetail mDCs and pDCs, both of which were eithergenerated from bone marrow-derived precursor cells in
vitro or directly isolated from the spleens of mice. pDCsreleased very high amounts of IFN-a/b after exposure toL. major, L. braziliensis or L. infantum in the absence ofdetectable parasite endocytosis, whereas mDCs wereunable to do so despite rapid uptake of the Leishmania.The effect of live Leishmania on pDCs could bemimicked by lysates of Leishmania promastigotes aswell as by genomic DNA (but not by kinetoplast DNA)of the parasites (Schleicher et al. 2007). From these datawe conclude that at least in vitro pDCs are a much morepotent source of IFN-a/b than macrophages and mDCs.
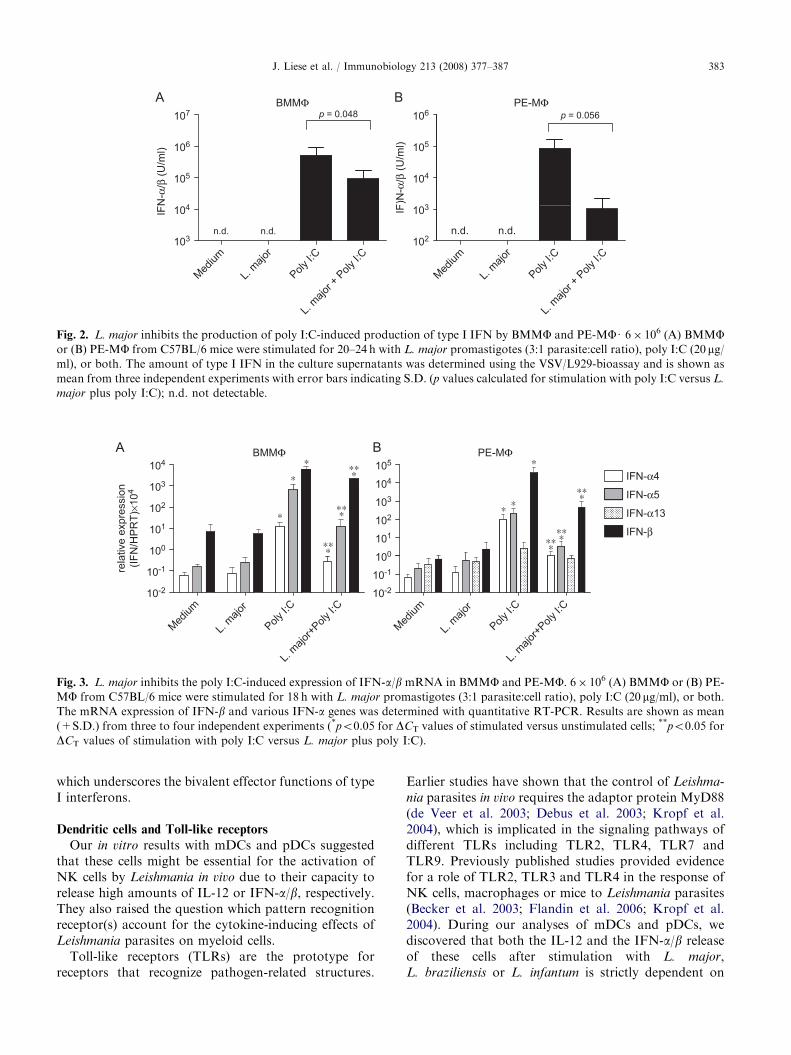
During our experiments with macrophages, we ob-served that L. major promastigotes can also exertinhibitory effects on the release of IFN-a/b. Whenadded to BMMFs or PE-MFs simultaneously with polyI:C, L. major impeded the expression of IFN-a4, -a5,-a13 and -b mRNA and caused a reduced release ofIFN-a/b protein compared to a stimulation with polyI:C alone (Figs. 2A and B, 3A and B).
IFN-a/b in vivo. The first, indirect hints for aprotective effect of IFN-a/b in murine cutaneousleishmaniasis came from the analysis of congenicB6.C-H-28c mice. These mice, which were known toproduce lower amounts of IFN-a/b in response toviruses compared to C57BL/6 wild-type mice, showed amore severe, but nevertheless self-healing course ofinfection with L. major (Shankar et al. 1996). InL. major-infected 129/SvEv�C57BL/6 mice we ob-served that the application of sheep-anti-IFN-a/bneutralizing antibodies abrogated the early NK cellactivity (cytotoxicity and IFN-g expression) in thelymph node and the expression of iNOS protein in theskin. In accordance with these findings, we detectedIFN-a/b-positive cells in the dermis of these mice at day1 of infection (Diefenbach et al. 1998). In the model ofvisceral leishmaniasis (L. infantum), we found thatdeletion of the IFN-a/b receptor I chain partially
ARTICLE IN PRESS
IFN-α2
101
102IFN-α4
101
102∗
IFN-α5
102
103
∗
IFN-α9100
10-1
100
rela
tive
expr
essi
ons
(IFN
/HP
RT)
×104
10-1
100
100
101
10-1
Medium
L. majo
r
Poly I:C
CpG 16
68
LPS+IF
N-γ10-2
n.d.
Medium
L. majo
r
Poly I:C
CpG 16
68
LPS+IF
N-γ10-2
n.d.
Medium
L. majo
r
Poly I:C
CpG 16
68
LPS+IF
N-γ10-1
Medium
L. majo
r
Poly I:C
CpG 16
68
LPS+IF
N-γ
n.d. n.d. n.d.
IFN-α11
102
103IFN-α12
100
101IFN-α13
102
103IFN-β
103
104 ∗
∗
100
101
rela
tive
expr
essi
on(IF
N/H
PR
T)×1
04
∗
10-2
10-1
100
101
100
101
102∗
Medium
L. majo
r
Poly I:C
CpG 16
68
LPS+IF
N-γ10-1
Medium
L. majo
r
Poly I:C
CpG 16
68
LPS+IF
N-γ10-3 n.d.
Medium
L. majo
r
Poly I:C
CpG 16
68
LPS+IF
N-γ10-1
Medium
L. majo
r
Poly I:C
CpG 16
68
LPS+IF
N-γ10-1
10-2
10-3
Fig. 1. Induction of IFN-a/b subtype mRNA in PE-MF. 6� 106 PE-MF from C57BL/6 mice were stimulated for 18–20 h with L.
major promastigotes (3:1 parasite:cell ratio), poly I:C (20 mg/ml), CpG 1668 (1 mM), or LPS and IFN-g (200+20 ng/ml). The
expression of (A) IFN-a2, (B) IFN-a4, (C) IFN-a5, (D) IFN-a9, (E) IFN-a11, (F) IFN-a12, (G) IFN-a13, and (H) IFN-b mRNA
relative to HPRT mRNA was analyzed with quantitative PCR using assays for the respective genes. Mean relative expression levels
(+S.D., if IFN mRNA was detectable in all experiments) are shown from three independent experiments (*po0.05 for DCT values
of stimulated versus unstimulated cells where applicable); n.d. not detectable.
J. Liese et al. / Immunobiology 213 (2008) 377–387382
reduced NK cell IFN-g production (but not NK cellcytotoxicity) in the spleen (Schleicher et al. 2007).Although these results have been obtained by differentexperimental approaches (antibody treatment versusgene-deleted mouse), they suggest that IFN-a/b is ofdifferent importance in cutaneous versus visceral leish-maniasis.
A number of studies addressed the question, whetherthere is a role for IFN-a/b in therapy or prevention ofcutaneous leishmaniasis. A single local and/or systemictreatment with poly I:C, purified mouse IFN-a/b, orrecombinant mouse IFN-b led to early parasite contain-ment and to enhanced NK cell activity in BALB/c mice,but did not prevent the ultimate disease progression inthese mice (Diefenbach et al. 1998; Laskay et al. 1995;Mattner et al. 2004; Schleicher et al. 2004). The first
attempts to treat BALB/c mice systemically for a longerperiod of time with recombinant human IFN-aA/Dfailed, because the mice developed severe peritonealhemorrhages (Shankar et al. 1996). Using recombinantmouse IFN-b we succeeded to establish a treatmentscheme to prevent fatal disease in L. major-infectedBALB/c mice (Mattner et al. 2004). Mice infected withan intermediate parasite number were completelyprotected from fatal disease, and even after high-doseinfection (2� 106 promastigotes) progressive skin le-sions were only observed in 40% of the mice. Thetreatment regimen consisted of the administration ofrmIFN-b prior to infection and every other day there-after for a total of 14 days. Unexpectedly, a lower doseof rmIFN-b (5� 103U per application) was moreefficient than a high dose (1� 105U per application),
ARTICLE IN PRESS
BMMΦ107 p = 0.048
PE-MΦ106 p = 0.056
105
106
104
105
103
104
n.d.n.d.
IFN
-α/β
(U/m
l)
102
103
n.d.n.d.
IF)N
-α/β
(U/m
l)
Medium
L. majo
r
Poly I:C
L. majo
r + P
oly I:C
Medium
L. majo
r
Poly I:C
L. majo
r + P
oly I:C
Fig. 2. L. major inhibits the production of poly I:C-induced production of type I IFN by BMMF and PE-MFU 6� 106 (A) BMMFor (B) PE-MF from C57BL/6 mice were stimulated for 20–24 h with L. major promastigotes (3:1 parasite:cell ratio), poly I:C (20 mg/ml), or both. The amount of type I IFN in the culture supernatants was determined using the VSV/L929-bioassay and is shown as
mean from three independent experiments with error bars indicating S.D. (p values calculated for stimulation with poly I:C versus L.
major plus poly I:C); n.d. not detectable.
BMMΦ104 ∗
∗
PE-MΦ
104
105
IFN-α4∗
100
101
102
103
∗
101
102
103 IFN-α5
IFN-α13
IFN-β
∗ ∗∗∗∗
∗∗∗
∗∗∗
∗∗∗
∗∗∗∗∗∗
10-2
10-1rela
tive
expr
essi
on(IF
N/H
PR
T)×1
04
10-2
10-1
100
Medium
L. majo
r
Poly I:C
L. majo
r+Poly
I:C
Medium
L. majo
r
Poly I:C
L. majo
r+Poly
I:C
Fig. 3. L. major inhibits the poly I:C-induced expression of IFN-a/b mRNA in BMMF and PE-MF. 6� 106 (A) BMMF or (B) PE-
MF from C57BL/6 mice were stimulated for 18 h with L. major promastigotes (3:1 parasite:cell ratio), poly I:C (20 mg/ml), or both.
The mRNA expression of IFN-b and various IFN-a genes was determined with quantitative RT-PCR. Results are shown as mean
(+S.D.) from three to four independent experiments (*po0.05 for DCT values of stimulated versus unstimulated cells; **po0.05 for
DCT values of stimulation with poly I:C versus L. major plus poly I:C).
J. Liese et al. / Immunobiology 213 (2008) 377–387 383
which underscores the bivalent effector functions of typeI interferons.
Dendritic cells and Toll-like receptors
Our in vitro results with mDCs and pDCs suggestedthat these cells might be essential for the activation ofNK cells by Leishmania in vivo due to their capacity torelease high amounts of IL-12 or IFN-a/b, respectively.They also raised the question which pattern recognitionreceptor(s) account for the cytokine-inducing effects ofLeishmania parasites on myeloid cells.
Toll-like receptors (TLRs) are the prototype forreceptors that recognize pathogen-related structures.
Earlier studies have shown that the control of Leishma-
nia parasites in vivo requires the adaptor protein MyD88(de Veer et al. 2003; Debus et al. 2003; Kropf et al.2004), which is implicated in the signaling pathways ofdifferent TLRs including TLR2, TLR4, TLR7 andTLR9. Previously published studies provided evidencefor a role of TLR2, TLR3 and TLR4 in the response ofNK cells, macrophages or mice to Leishmania parasites(Becker et al. 2003; Flandin et al. 2006; Kropf et al.2004). During our analyses of mDCs and pDCs, wediscovered that both the IL-12 and the IFN-a/b releaseof these cells after stimulation with L. major,L. braziliensis or L. infantum is strictly dependent on

ARTICLE IN PRESS
Fig. 4. Model for the TLR9-dependent activation of NK cells by dendritic cells in the spleen of L. infantum-infected mice. mDCs
endocytose L. infantum parasites, the genomic DNA of which gets into contact with TLR9 in the endosomal compartment after
parasite degradation. pDCs are unable to phagocytose L. infantum promastigotes. Possible explanations for the TLR9-dependent
induction of IFN-a/b is the transient expression of TLR9 on the cell surface of pDCs and/or the release of soluble TLR9 ligands by
the parasites which subsequently enter the endosomal compartment of pDCs via fluid-phase endocytosis. X denotes a cell-type that
might compensate for the expression of IFN-a/b in the spleen of mice depleted of pDCs. For further details see text.
J. Liese et al. / Immunobiology 213 (2008) 377–387384
TLR9 signaling (Liese et al. 2007; Schleicher et al. 2007).In accordance with these in vitro findings we observedthat TLR9 is indispensable for the production of IL-12by mDCs and the induction of NK cell IFN-g releaseand cytotoxicity in L. major- or L. infantum-infectedmice (Liese et al. 2007; Schleicher et al. 2007). Finally,deletion of mDCs abolished the NK cell response in thespleen of L. infantum-infected mice (Fig. 4). Impor-tantly, this was not the case after depletion of pDCs. InpDC-depleted mice, the infection with L. infantum led toan unaltered up-regulation of IFN-a4 and IFN-bmRNA, which indicates that splenic pDCs are eithernot a source of IFN-a/b in L. infantum-infected mice orthat other cells (such as macrophages or mDCs; cell-typeX in Fig. 4) compensate for the IFN-a/b production ofpDCs once they are depleted (Schleicher et al. 2007)(Fig. 4). Conversely, in L. infantum-infected mice thatwere devoid of mDCs the mRNA expression of type IIFNs was drastically increased compared to controlmice which clearly demonstrates that IFN-a/b in theabsence of IL-12 is not sufficient to trigger NK cellactivation in visceral leishmaniasis (Schleicher et al.2007). At present, we have not yet formally defined thenature of the cell type that accounts for the massiveexpression of IFN-a/b in the absence of mDCs, butpDCs are certainly a likely candidate.
When we followed the course of L. major infection inTLR9�/� mice, we found that these mice developed amore severe footpad swelling and had increased parasiteloads in the acute phase of the infection compared towild-type mice. This was paralleled by increased mRNAexpression levels of Th2-associated cytokines andeffector molecules (IL-4, IL-13, arginase 1) in the skinand in the draining lymph node, whereas the mRNAexpression of IFN-g, IL-12p35, IL-12p40, and iNOS was
comparable in both strains of mice. Ultimately, theTLR9�/� mice were able to contain the infection (Lieseet al. 2007). These observations demonstrate that TLR9plays an important role in the innate immune responseto L. major, which leads to an aggravation of thedisease, but TLR9-independent mechanisms exist thatconfer the healing phenotype.
Conclusions
The key antileishmanial effector molecules in experi-mental cutaneous and visceral leishmaniasis are reactivenitrogen intermediates (NO and NO-derived metabo-lites) and reactive oxygen intermediates (O2
� andsubsequent metabolites). In cutaneous leishmaniasisdue to L. major, iNOS and phox complement eachother in the skin, lymph node and spleen: whereas iNOSis most important for the control of L. major in thedermis and the draining lymph node, the function ofphox is critical for preventing visceralization of theparasite. In the case of L. donovani infections, the resultsare much less uniform. There is evidence for a variabledegree of iNOS/NO-dependent control of the parasitesin macrophages and in the liver during the acute phaseof infection, but also for iNOS/phox-independentantileishmanial mechanisms during the chronic (latent)phase of infection after chemotherapy.
The activation of NK cells in visceral and most likelyalso in cutaneous leishmaniasis results from the intimateinteraction of NK cells with mDCs, which are triggeredby Leishmania parasites for the production of IL-12 in aTLR9-dependent fashion. The true contribution ofpDCs and their TLR9-dependent release of IFN-a/b in
vivo requires further investigation.

ARTICLE IN PRESSJ. Liese et al. / Immunobiology 213 (2008) 377–387 385
Acknowledgments
The authors wish to thank Andrea Hesse, ClaudiaKurzmann and Rosa Mammato for their skillful andenthusiastic technical assistance throughout the years ofjoint research and Drs. Jochen Mattner and NicoleJusties for their contributions to some of the studiesreviewed. Our work summarized here results from a 6-year research project supported by the German Re-search Foundation within the Priority ProgramSPP1110 ‘‘Innate Immunity’’ (grants Bo996/3-1, 3-2and 3-3 to C.B.).
References
Aizu, K., Li, W., Yajima, T., Arai, T., Shimoda, K., Nimura,
Y., Yoshikai, Y., 2006. An important role of Tyk2 in APC
function of dendritic cells for priming CD8+ T cells
producing IFN-g. Eur. J. Immunol. 36, 3060–3070.
Asselin-Paturel, C., Boonstra, A., Dalod, M., Durand, I.,
Yessaad, N., Dezutter-Dambuyant, C., Vicari, A., O’Gar-
ra, A., Biron, C., Briere, F., Trinchieri, G., 2001. Mouse
type I IFN-producing cells are immature APCs with
plasmacytoid morphology. Nat. Immunol. 2, 1144–1150.
Becker, I., Salaiza, N., Aguirre, M., Delgado, J., Carrillo-
Carrasco, N., Gutierrez-Kobeh, L., Ruiz, A., Cervantes,
R., Perez Torres, A., Cabrera, N., Gonzalez, A., Mal-
donado, C., Isibasi, A., 2003. Leishmania lipophosphogly-
can (LPG) activates NK cells through toll-like-receptor-2.
Mol. Biochem. Parasitol. 130, 65–74.
Blos, M., Schleicher, U., Rocha, F.J., Meissner, U., Rollingh-
off, M., Bogdan, C., 2003. Organ-specific and stage-
dependent control of Leishmania major infection by
inducible nitric oxide synthase and phagocyte NADPH
oxidase. Eur. J. Immunol. 33, 1224–1234.
Bogdan, C., 2001a. Nitric oxide and the immune response.
Nat. Immunol. 2, 907–916.
Bogdan, C., 2001b. Nitric oxide and the regulation of gene
expression. Trends Cell Biol 11, 66–75.
Bogdan, C., 2007. Phagocyte Effector Functions Against
Leishmania Parasites. Landes Bioscience, Austin, TX,
193–206pp.
Bogdan, C., Gessner, A., Rollinghoff, M., 1993. Cytokines in
Leishmaniasis: a complex network of stimulatory and
inhibitory interactions. Immunobiology 189, 356–396.
Bogdan, C., Gessner, A., Solbach, W., Rollinghoff, M., 1996.
Invasion, control, and persistence of Leishmania parasites.
Curr. Opin. Immunol. 8, 517–525.
Bogdan, C., Thuring, H., Dlaska, M., Rollinghoff, M., Weiss,
G., 1997. Mechanism of suppression of macrophage nitric
oxide release by IL-13. J. Immunol. 159, 4506–4513.
Bogdan, C., Donhauser, N., Doring, R., Rollinghoff, M.,
Diefenbach, A., Rittig, M.G., 2000. Fibroblasts as
host cells in latent leishmaniosis. J. Exp. Med. 191,
2121–2129.
Bogdan, C., Mattner, J., Schleicher, U., 2004. The role of type
I interferons in non-viral infections. Immunol. Rev. 202,
33–48.
Cella, M., Jarrossay, D., Fachetti, F., Alebardi, O., Nakajima,
H., Lanzavecchia, A., Colonna, M., 1999. Plasmacytoid
monocytes migrate to inflamed lymph nodes and produce
large amounts of type 1 interferon. Nat. Med. 5, 919–923.
Chappuis, F., Sundar, S., Hailu, A., Ghalib, H., Rijal, S.,
Peeling, R.W., Alvar, J., Boelaert, M., 2007. Visceral
leishmaniasis: what are the needs for diagnosis, treatment
and control? Nat. Rev. Microbiol. 5, 873–882.
Dalton, D.K., Pitts-Meek, S., Keshav, S., Figari, I.S., Bradley,
A., Stewart, T.A., 1993. Multiple defects of immune cell
function in mice with disrupted interferon-g genes. Science
259, 1739–1742.
de Veer, M.J., Curtis, J.M., Baldwin, T.M., DiDonato, J.A.,
Sexton, A., McConville, M.J., Handman, E., Schofield, L.,
2003. MyD88 is essential for clearance of Leishmania major:
possible role of lipophosphoglycan and Toll-like receptor 2
signaling. Eur. J. Immunol. 33, 2822–2831.
Debus, A., Glasner, J., Rollinghoff, M., Gessner, A., 2003.
High levels of susceptibility and T helper 2 response in
MyD88-deficient mice infected with Leishmania major are
interleukin-4 dependent. Infect. Immun. 71, 7215–7218.
Decker, T., Muller, M., Stockinger, S., 2005. The yin and yang
of type I interferon activity in bacterial infection. Nat. Rev.
Immunol. 5, 675–687.
Diefenbach, A., Schindler, H., Donhauser, N., Lorenz, E.,
Laskay, T., MacMicking, J., Rollinghoff, M., Gresser, I.,
Bogdan, C., 1998. Type 1 interferon (IFN-a/b) and type 2
nitric oxide synthase regulate the innate immune response
to a protozoan parasite. Immunity 8, 77–87.
Diefenbach, A., Schindler, H., Rollinghoff, M., Yokoyama,
W., Bogdan, C., 1999. Requirement for type 2 NO-synthase
for IL-12 responsiveness in innate immunity. Science 284,
951–955.
Flandin, J.-F., Chano, F., Descoteaux, A., 2006. RNA
interference reveals a role for TLR2 and TLR3 in the
recognition of Leishmania donovani promastigotes by
interferon-g-primed macrophages. Eur. J. Immunol. 36,
411–420.
Huang, S., Hendriks, W., Althage, A., Hemmi, S., Blue-
thmann, H., Kamijo, R., Vilcek, J., Zinkernagel, R.M.,
Aguet, M., 1993. Immune response in mice that lack the
interferon-g receptor. Science 259, 1742–1744.
Kamezaki, K., Shimoda, K., Numata, A., Matsuda, T.,
Nakayama, K.-I., Harada, M., 2004. The role of Tyk2,
STAT1 and STAT4 in LPS-induced endotoxin signals. Int.
Immunol. 16, 1173–1179.
Kamijo, R., Shapiro, D., Le, J., Huang, S., Aguet, M., Vilcek,
J., 1993. Generation of nitric oxide and induction of major
histocompatibility complex class II antigen in macrophages
from mice lacking the interferon g receptor. Proc. Natl.
Acad. Sci. USA 90, 6626–6630.
Karaghiosoff, M., Neubauer, H., Lassnig, C., Kovarik, P.,
Schindler, H., Pircher, H.-P., McCoy, B., Bogdan, C.,
Decker, T., Brem, G., Pfeffer, K., Muller, M., 2000. Partial
impairment of cytokine responses in tyk2-deficient mice.
Immunity 13, 549–560.
Karaghiosoff, M., Steinborn, R., Kovarik, P., Kriegshauser,
G., Baccarini, M., Donabauer, B., Reichart, U., Kolbe, T.,
Bogdan, C., Leanderson, T., Levy, D., Decker, T., Muller,
M., 2003. Central role for type I interferons and Tyk2 in

ARTICLE IN PRESSJ. Liese et al. / Immunobiology 213 (2008) 377–387386
lipopolysaccharide-induced endotoxin shock. Nat. Immu-
nol. 4, 471–477.
Kirkpatrick, C.E., Farrell, J.P., 1982. Leishmaniasis in beige
mice. Infect. Immun. 38, 1208–1216.
Kropf, P., Freudenberg, M.A., Modolell, M., Price, H.P.,
Herath, S., Antoniazi, S., Galanos, C., Smith, D.F., Muller,
I., 2004. Toll-like receptor 4 contributes to efficient control
of infection with the protzoan parasite Leishmania major.
Infect. Immun. 72, 1920–1928.
Laskay, T., Diefenbach, A., Rollinghoff, M., Solbach, W.,
1995. Early parasite containment is decisive for resistance
to Leishmania major infection. Eur. J. Immunol. 25,
2220–2227.
Leclercq, V., Lebastard, M., Belkaid, Y., Louis, J., Milon, G.,
1996. The outcome of the parasitic process initiated by
Leishmania infantum in laboratory mice. A tissue-depen-
dent pattern controlled by the Lsh and MHC loci.
J. Immunol. 157, 4537–4545.
Liese, J., Schleicher, U., Bogdan, C., 2007. TLR9 signalling is
essential for the innate NK cell response in murine
cutaneous leishmaniasis. Eur. J. Immunol. 37, 3424–3434.
Liew, F.Y., Millott, S., Parkinson, C., Palmer, R.M.,
Moncada, S., 1990. Macrophage killing of Leishmania
parasite in vivo is mediated by nitric oxide from L-arginine.
J. Immunol. 144, 4794–4797.
Lukes, J., Mauricio, I.L., Schonian, G., Dujardin, J.-C.,
Soteriadou, K., Dedet, J.-P., Kuhls, K., Tintaya, K.W.Q.,
Jirku, M., Chocholova, E., Haralambous, C., Pratlong,
F., Obornik, M., Horak, A., Ayala, F.J., Miles, M.A.,
2007. Evolutionary and geographical history of the
Leishmania donovani complex with a revision of current
taxonomy. Proc. Natl. Acad. Sci. USA 104, 9375–9380.
Mattner, F., Di Padova, K., Alber, G., 1997. Interleukin-12 is
indispensable for protective immunity against Leishmania
major. Infect. Immun. 65, 4378–4383.
Mattner, J., Schindler, H., Diefenbach, A., Rollinghoff, M.,
Gresser, I., Bogdan, C., 2000. Regulation of type 2 NO
synthase by type I interferons in macrophages infected with
Leishmania major. Eur. J. Immunol. 30, 2257–2267.
Mattner, J., Wandersee-Steinhauser, A., Pahl, A., Rollinghoff,
M., Majeau, G.R., Hochman, P.S., Bogdan, C., 2004.
Protection against progressive leishmaniasis by IFN-b.J. Immunol. 172, 7574–7582.
Melby, P.C., Yang, Y.-Z., Cheng, J., Zhao, W., 1998. Regional
differences in the cellular immune response to experimental
cutaneous or visceral leishmaniasis with Leishmania dono-
vani. Infect. Immun. 66, 18–27.
Mendonca, M.G., de Brito, M.E.F., Rodrigues, E.H.G.,
Bandeira, V., Jardim, M.L., Abath, F.G.C., 2004. Persis-
tence of Leishmania parasites in scars after clinical cure of
American cutaneous leishmaniasis: is there a sterile cure?
J. Infect. Dis. 189, 1018–1023.
Minegishi, Y., Saito, M., Morio, T., Watanabe, K., Agematsu,
K., Tsuchiya, S., Takada, H., Hara, T., Kawamura, N.,
Ariga, T., Kaneko, H., Kondo, N., Tsuge, I., Yachie, A.,
Sakiyama, Y., Iwata, T., Bessho, F., Ohishi, T., Joh, K., Imai,
K., Kogawa, K., Shinohara, M., Fujieda, M., Wakiguchi, H.,
Pasic, S., Abinun, M., Ochs, H.D., Renner, E.D., Jansson,
A., Belohradsky, B.H., Metin, A., Shimizu, N., Mizutani, S.,
Miyawaki, T., Nonoyama, S., Karasuyama, H., 2006. Human
tyrosine kinase 2 deficiency reveals its requisite roles in
multiple cytokine signals involved in innate and acquired
immunity. Immunity 25, 745–755.
Murray, H.W., 1997. Endogenous interleukin-12 regulates
acquired resistance in experimental visceral leishmaniasis.
J. Infect. Dis. 175, 1477–1479.
Murray, H.W., 2005. Prevention of relapse after chemotherapy
in a chronic intracellular infection: mechanisms in experi-
mental visceral leishmaniasis. J. Immunol. 174, 4916–4923.
Murray, H.W., 2006. Responses to Leishmania donovani in
mice deficient in both phagocyte oxidase and inducible
nitric oxide synthase. Am. J. Trop. Med. Hyg. 74,
1013–1015.
Murray, H.W., Nathan, C.F., 1999. Macrophage microbicidal
mechanisms in vivo: reactive nitrogen vs oxygen inter-
mediates in the killing of intracellular visceral Leishmania
donovani. J. Exp. Med. 189, 741–746.
Murray, H.W., Berman, J.D., Davies, C.R., Saravia, N.G.,
2005. Advances in leishmaniasis. Lancet 366, 1561–1577.
Nakano, H., Yanagita, M., Gunn, M.D., 2001.
CD11c+B220+Gr-1+ cells in mouse lymph nodes and
spleen display characteristics of plasmacytoid dendritic
cells. J. Exp. Med. 194, 1171–1178.
Pearson, R.D., de Queiroz Sousa, A., 1996. Clinical spectrum
of leishmaniasis. Clin. Infect. Dis. 22, 1–13.
Perez, L.E., Chandrasekar, B., Saldarriaga, O.A., Zhao, W.,
Arteaga, L.T., Travi, B.L., Melby, P.C., 2006. Reduced
nitric oxide synthase 2 (NOS2) promoter activity in the
syrian hamster renders the animal functionally deficient in
NOS2 activity and unable to control an intracellular
pathogen. J. Immunol. 176, 5519–5528.
Roach, T.I.A., Kiderlen, A.F., Blackwell, J.M., 1991. Role of
inorganic nitrogen oxides and tumor necrosis factor alpha
in killing Leishmania donovani amastigotes in gamma
interferon/lipopolysaccharide-activated macrophages from
Lshs and Lshr congenic mouse strains. Infect. Immun. 59,
3935–3944.
Sacks, D.L., Noben-Trauth, N., 2002. The immunology of
susceptibility and resistance to Leishmania major in mice.
Nat. Rev. Immunol. 2, 845–858.
Scharton-Kersten, T., Afonso, L.C.C., Wysocka, M., Trinch-
ieri, G., Scott, P., 1995. IL-12 is required for natural killer
cell activation and subsequent T helper 1 cell develop-
ment in experimental leishmaniasis. J. Immunol. 154,
5320–5330.
Schindler, C., Levy, D.E., Decker, T., 2007. JAK-STAT
signaling: from interferons to cytokines. J. Biol. Chem. 282,
20059–20063.
Schleicher, U., Mattner, J., Blos, M., Schindler, H., Rollingh-
off, M., Karaghiosoff, M., Muller, M., Werner-Felmayer,
G., Bogdan, C., 2004. Control of Leishmania major in the
absence of Tyk2 kinase. Eur. J. Immunol. 34, 519–529.
Schleicher, U., Hesse, A., Bogdan, C., 2005. Minute numbers
of contaminant CD8+ T cells or CD11b+CD11c+ NK
cells are the source of IFN-g in IL-12/IL-18-stimulated
mouse macrophage populations. Blood 105, 1319–1328.
Schleicher, U., Liese, J., Knippertz, I., Kurzmann, C., Hesse,
A., Heit, A., Fischer, J.A., Weiss, S., Kalinke, U., Kunz, S.,
Bogdan, C., 2007. NK cell activation in visceral leishma-
niasis requires TLR9, myeloid DCs, and IL-12, but is

ARTICLE IN PRESSJ. Liese et al. / Immunobiology 213 (2008) 377–387 387
independent of plasmacytoid DCs. J. Exp. Med. 204,
893–906.
Schubach, A., Haddad, F., Neto, M.P.-O., Degrave, W.,
Pirmez, C., Grimaldi, G., Fernandes, O., 1998. Detection of
Leishmania DNA by polymerase chain reaction in scars of
treated human patients. J. Infect. Dis. 178, 911–914.
Shankar, A.H., Morin, P., Titus, R.G., 1996. Leishmania
major: differential resistance to infection in C57BL/6 (high
interferon-a/b) and congenic B6.C-H-28c (low interferon-a/b) mice. Exp. Parasitol. 84, 136–143.
Shimoda, K., Kato, K., Aoki, K., Matsuda, T., Miyamoto, A.,
Shibamori, M., Yamashita, M., Numata, A., Takase, K.,
Kobayashi, S., Shibata, S., Asano, Y., Gondo, H.,
Sekiguchi, K., Nakayama, K., Nakayama, T., Okamura,
T., Okamura, S., Niho, Y., Nakayama, K.-I., 2000. Tyk2
plays a restricted role in IFNa signaling, although it is
required for IL-12-mediated T cell function. Immunity 13,
561–571.
Siegal, F.P., Norimitsu, K., Shodell, M., Fitzgerald-Bocarsly,
P.A., Shah, K., Ho, S., Antonenko, S., Liu, Y.-J., 1999. The
nature of the principal type 1-interferon-producing cells in
human blood. Science 284, 1835–1837.
Smelt, S.C., Engweda, C.R., McCrossen, M., Kaye, P.M.,
1997. Destruction of follicular dendritic cells during chronic
visceral leishmaniasis. J. Immunol. 158, 3813–3821.
Soares Rocha, F.J., Schleicher, U., Mattner, J., Alber, G.,
Bogdan, C., 2007. Cytokines, signaling pathways, and
effector molecules required for the control of Leishmania
(Viannia) braziliensis in mice. Infect. Immun. 75,
3823–3832.
Solbach, W., Laskay, T., 2000. The host response to
Leishmania infection. Adv. Immunol. 74, 275–317.
Stenger, S., Donhauser, N., Thuring, H., Rollinghoff, M.,
Bogdan, C., 1996. Reactivation of latent leishmaniasis by
inhibition of inducible nitric oxide synthase. J. Exp. Med.
183, 1501–1514.
Tokumasa, N., Suto, A., Kagami, S., Furuta, S., Hirose, K.,
Watanabe, N., Saito, Y., Shimoda, K., Iwamoto, I.,
Nakajima, H., 2007. Expression of Tyk2 in dendritic cells
is required for IL-12, IL-23, and IFN-g production and the
induction of Th1 cell differentiation. Blood 110, 553–560.
Trinchieri, G., 2003. Interleukin-12 and the regulation of
innate resistance and adaptive immunity. Nat. Rev.
Immunol. 3, 133–146.
White, J.K., Mastroeni, P., Popoff, J.-F., Evans, C.A.W.,
Blackwell, J.M., 2005. Slc11a1-mediated resistance to
Salmonella enterica serovar typhimurium and Leishmania
donovani infections does not require functional inducible
nitric oxide synthase or phagocyte oxidase activity.
J. Leukoc. Biol. 77, 311–320.