Embed Size (px)
Citation preview

I'
PHYSIOL. PLANT. 55: 136-142. Copenhagen 1982
The inhibitory effect of gibberellic acid on flowering in Citrus
J. L. Guardiola, C. Monerri and M. Agusti
Guardiola, J. L., Monerri, C. and Agusti, M. 1982. The inhibitory effect of gibbercll acid on flowering in Cilrus. - Physiol. Plant. 55: 136-142. It
The application of gibberellic acid (GAJ ) at any time from early November until bud sprouting, resulted in a significant inhibition of flowering in the sweet orange II.. sinensis (L.) Osbeck] and the Satsuma (c. unshiu Marc.) and Clementine (C.,.. licu/ata Blanco) mandarins. Two response peaks were evident: the first occurrc when the application was timed to the translocation of an unknown f1uwering signal from the leaves tu the buds. The second occurred during bud sprouting, at the tim.: the flower primordia were differentiating. From the pattern of flowering, it appcall that the mechanism of inhibition was similar irrespective of the timing of GAJ applt cation. There was an initial reduction in bud sprouting affecting selectively those bulb originating leafless inflorescences. An additional inhibitiun resulted in a reductiunm the number of leafy influrescences with an increase in the number uf vegetaliH shoots, suggesting the reversion of a floral to a vegetative apex. The inhibited bulb sprouted readily in vitro but invariably vegetative shoots were formed. A continUllU\ influence of the sustaining branch is necessary to keep the flowering commitmcnll'( I the buds; irreversible commitment occurs when the petal primordia are well dit ferentiated.
Additional key words - Bud dormancy, bud sprouting, Citrus reticulata, Citrus si"c'r.· sis, Citrus unshiu.
J. L. Guardiola et al., Ciltedra de Fisiologia Vegetal, Escuela Tecnica Superi,,"', /ngenierus Agronomos, Univ. Po!itecnica, Valencia, Spain.
Introduction
As in many fruit trees, flowering in Citrus is inhibited by the application of exogenous GAJ . Whilst a practical use of this property is being made to regulate flowering in sweet orange (Moss and Bevington 1977), the mechanism of inhibition is not fully understood. An interference with flower initiation has been suggested (Monselise and Halevy 1964, Moss 1970), since GAJ is most effective during the winter rest period, at t.he time when some flowering factor is translocated from the leaves to the buds (Furr and Armstrong 1956, Ayalon and Monselise 1960, Sanchez-Capuchino and Casanova 1973), or during a period of water stress, which readily induces flowering in lemon (Nir et al. 1972). However, later applications of GAJ , timed to bud sprouting when flower differentiation is detectable under the microscope, are also effective (Nir et al. 1972, Goldschmidt
Received 27 October, 1981; revised 16 February, 1982
136 0031-9317/82/060136-07 $03.00/0 © 1982 Physiologia Plantarum PhysioL Plant. 55. I
and Monselise 1972, Guardiola et al. 1980). This reo sponse may involve the reversion of a flower bud to vegetative apex, and the reduction in flowcrin achieved is related to the increase in shoot length. contrast, the earlier GAJ applications do not influence vegetative growth.
Thus, there is a possibility that GAJ may inhibit flo"' ering by acting at two different stages of flower forma· tion. This has been tested in the present study by com' paring the response of three Citrus species to GAl applications from early November to bud sprouting. Spe' cial attention was paid to the anatomical changes occurring in the buds and to their behaviour when ex' cised from the tree in order to gain more insight into tilt mechanism of action of GA3 at the bud level.
Abbreviations - 2,4-D, 2,4-dichlorophenoxyacetic acid; GA, gibberellic acid.
\Iaterials aJ
plant materia
Three separ i11l1uenceo f
lJi tlowenng ,(itTl/S sim.'!; I . linc (c. re/I,
lI11shiu Mal 15-25 years llr;tnge root~
,prays were early Noven treatment c( or 6 replicv hlllck desigl WitS 100 mg mg \-1 for t
(Mojafor, m lilill of 0.1 'Yr, having no d
Bud sprot days before bUled unifor 2,000 buds initiated we! ported (Gua ~hoots devcl< the total nu results were nodes to cm hranehes sell than 24 mon de red for th, trihution of (' rhe influenc determined i
An additi· I avclate ora times of max ,prouting, at I I. respectiv< the rest of th<
null sprou', orange. Twer 10() mg 1-1 Gf October 20. I were periodic untreated cor fore plantin o
dipped for aO; Only the six I
in this experil with bUds of t tree.
Phl'siol. Plant. :

rering in Citrus
'he inhibitory effect of gihher lit 16-142,
~ from early November until ,wering in the swcet orange I Marc.) and Cleme?tine (c. ,(.
vere eVIdent: the fIrst OCCUlTCl!
of an unknown flowering sig urIng bud sproutlOg, at the tillle IJattern of flowering, it appean
:lIve of the lImlOg of GAJ apflj g affecting selectively those buds ibition resulted in a reduetillnia se in the number of vegetati ~tatlve apex. The inhibited budl oots were formed. A continullU! ep the flowering commitment 0/ Ie petal primordia are well wi
g, Cilrus relicu/ala, Cilmf silltn,
tI, Escue/a Tecnicil Supai", tit Spain.
rdiola et al. 1980). This It version of a flower bud to te reduction in f10wcrin increase in shoot length. I Ipplications do not influcnCt
y that GA3 may inhibit flow, rent stages of flower forma' n the present study by com· e Citrus species to GAl apnber to bud sprouting. Speo the anatom 'cal chang :0 their behaviour when et· to gain more insight inlll ll1e
~3 at the bud level.
hlorophenoxyacetic acid: GA
n i'hY'Sio!. PI.nt. ;5. 1982Physio!. PI.nl. 55. I
'als and methods,tatcO
terial and experimental design pl~nl PIa
separate experiments were conducted to test the lhrc~ ce of the time of GA3 application on the degree '1111ucn ., h W h' I' . 1 ert'ng reductIOn m teas mgton nave orange,Ilow
sinensis (L.) Osbeck) and the seedless ClemenIl( I/f/I.I
.
) d' ( .reticulala Blanco an Owan Satsuma Citrusline (C. .
,/'1 Marcovitch) mandanns. Adult trees about 'III.\/ll .'1,-15 years old growmg out~oors and bud?ed on sour . g' rootstocks were used m every expenment. GA3,lrall C " .
'S were penodlcally applted to the whole tree from ,prr November until the time of bud sprouting. Each car) d f . I GA I' d 5nc;ttment consiste 0 a smg e 3-spray app Ie to
6 replicates of whole-tree plots m a randomIzed "r . f G .block design. The concentratIOn 0 the A 3 solutIOn lIas lOll mg 1-1 in the sweet orange experiment and 10 IIlg I-I for the mandarins. A no~-ionic wetting agent (~l()jafor, manufactured by QUlmmor), at a concentralIon of D.I % (v/v) was included, this wetting agent itself h,\\'ing no effect on flowering.
Dud sprouting was measured in early spring, a few days before flower opening. Six to ten branches distri bll;l~d uniformly around the tree and having more than , ilDli buds were selected beforehand, and the shoots ~;itiated were counted and classified as previously repurled (Guardiola et al. 1977). From the number of ,hoots developed and the number of flowers per shoot, the lotal number of flowers could be calculated. The results were cxpressed in shoots and flowers per 100 nodes to compensate for the differences in size of the branches selccted for counting. Only the buds being less Ihan 24 months old at the time of sprouting were considered for the countings and calculations, since the contrihution of older buds to the spring flush was negligible. I hc influence of the age of the bud on sprouting was determined in the sweet orange experiment.
An additional experiment was performed using :\avclate orange trees (Tab. 2). GA3 was tested at the times of maximal sensitivity, in mid-November and at 'prouting, at lower concentrations, 25 mg 1-1 and 10 mg 1'1, respectively. Ten whole tree replicates were used, the rcst of the experimental conditions being as above.
Bud sprouting in vitro was studied in the Navelate urange. Twenty trees were sprayed three times with a 100 mg 1-1 GA3 solution at monthly intervals starting on October 20, Branches from the autumn flush of growth lIere periodically sampled from these trees and from the untreated controls and their buds cultured in vitro. Before planting, half of the buds from the controls were dipped for a few seconds in a 100 mg 1-1 GA3 solution, Only. the six more apical buds of the branch were used In thiS experiment, and their behaviour was compared IIlth bUds of the same age and position sprouting on the Iree.
Bud sprouting in vitro
Bud explants, consisting of the whole bud without the sustaining leaf together with a 15 mm long flap of bark and wood, were planted under sterile conditions on a solid (2% agar) Murashige and Skoog (1962) medium and grown at 26 ± 1°C under continuous illumination (circa 25 W m-2
) provided by Sylvania Gro-Iux lamps. In view of the inhibitory effect of high temperatures on flower development (Moss 1969), growth at a lower temperature (19 ± 1°C) was tried as well. A 2 cm2 piece of leaf lamina was kept in some explants to determine its influence on bud sprouting.
While this procedure was adequate to compare the characteristics of the shoots sprouted, the large number of contaminated explants made the determination of the time-course of sprouting uncertain. Neither the use of a more drastic procedure for surface-sterilization (Giladi et al. 1979), nor the previous application to the tree of antibiotics and fungicides proved useful with our plant material. Finally, the time course of sprouting was followed by forcing the buds on a bed of sterile moist vermiculite, At variance with the observations of Altman and Goren (1977), we did not find sucrose essential for the initial bud emergence. The stage of bud dormancy was estimated from the time necessary to reach 50% sprouting, In the case where some buds had already initiated growth at the time of planting, these were considered to have sprouted in 0 days in the calculations.
Histological observations
Coinciding with the GArtreatments, five branches per tree from the autumn flush of growth were collected and fixed in ethyl alcohol:glacial acetic acid (3: 1, v/v). The five most apical buds were excised and sectioned with a freezing microtome or embedded in paraffin prior to cutting (Johansen 1940), and stained with safran in and fast green,
Results
Anatomical changes during bud sprouting
The resting bud is a complex structure coflsisting normally of three buds of different sizes with well differentiated leaf primordia surrounded by prophylls, and usually of a thorn. The biggest bud, of 250 ± 10 f.lm long up to the top of the apex and with 7-8 leaf primordia which covered completely the apex, was invariably the first to sprout on each node, and the one used in the measurements. Neither increase in size nor indication of flower differentiation could be detected for the first three samplings (Fig. 1). By the fifth sampling the most terminal bud had enlarged significantly. In later samplings sprouting gradually moved towards the base of the branch.
137

I '" Pbysiol. Plant
l11i1orescen'c hoots sioni o
30 u 0 lJ..
The differentiation of the flower parts was related to o 90 Llh, 3, The
the size of the sprouting shoot. The primordia of the .lppli~atiol1s
olf Ilmvers at <.? 70
sepals were clearly visible when the shoots were 650 ~m
long, and at that moment the biggest leaf primordia had z rrCall11~llt
enlarged up to about 1 mm. Petal primordia were well >::J o
differentiated on 1.3 mm long shoots, the biggest leaf g: 50
primordia being at least 2-3 mm long. (J)
>o Control (unto
11\" De~. I~ 30GAJ effects on flowering 11\" Nov. 2
o 0 z
The application of GA3 at any time from late October « up to the bursting of buds reduced flowering signific <.?
z antly in the three species studied (Fig. 2). Two response '" 70
w peaks were evident. The first occurred during the winter :;:
o rest period and it was earlier for C. sinensis and C. --'
lJ.. 50 tional expe haviour of
reticu[ata (end of November and early December) than for C. unshiu (end of December and early January). At
~lative shoethat time the dormancy of the buds was deepest as 30 measured by the time it took to sprout in vitro (Fig. 1). amll'. Furtl
number ofMicroscopic examination did not show evidence of or Control egrowth nor flower differentiation. The second response ircatmentspeak occurred during bud sprouting, when the de
veloping shoots were about 1.0 mm long; shortly afterwards they became insensitive to GA3 as regards flowering (not shown).
" r'lieu/alii
C. III/shill
Species
C. lincl/sis
IJh. 2. Th{ ,prouting (l JIlJ 5 trees tll)lVcrcd (13 ,jngle flowc
C. reticulata
i", ~f~1-1/'\0""'0 9-0
°
! j "'",C'"
\-f/')'''',j/\
c. unshiu
--?
NOV DEC JAN FEB
Fig. 2. The influence of the time of GAJ application all sh~ sprouting (0) and flowering (.0.) in the Washington n3' orange and the Satsuma and Clementine mandarins. Th~ application was timed to bud sprouting. The standard ~rr r the mean is given for the number of shoots developed.
o
(J) 50 --' o
'" >-
is
70
90
GAJ effects on sprouting
Following a GA3 application there was a reductio the number of shoots developed during the nat
n ,
spring flush, peak effects coinciding with the effect ~ flowering (Fig. 2). This effect was mainly due to a. duct ion in the proportion of buds sprouted. The inn ence of GA3 on the number of shoots developed sprouted bud was much smaller and not signifi (Tab. 1). This inhibition of bud sprouting was obsell irrespective of bud age, a factor which had a mar~
influence on natural sprouting. A similar pattern of inhibition was obtained irrcsPtt
tive of the timing of the GA3 application (Tab. 2). previously reported for C. sinensis (Guardiola et 1977), this inhibition was maximal for leafless shu(X and for leafy multiflowered ones. Some reduction in l~
number of leafy single flowered shoots was observed C. unshiu, a response which we have confirmed in add;
JAN.DEcNOVOCT
/.~.
.~/ \ \
0........-----"---""'-------"-.:.·---I
(J)
~ 2 o
1.5 E ..§
>o o I (J)
0.5
Ol-"'----........--.........--........----l
Fig. I. Seasonal change in dormancy, as measured by the time to sprout in vitro, and natural bud emergence as affected by the position of the bud on the branch, in the Navelate sweet orange. The buds have been numbered from the apex of the branch. Mean data for GAJ-treated and untreated trees, the effect of the hormone on these parameters not being significant.
Physiol. Plant. 5;· 138

The influence of bud age on spring sprouting and its inhibition by GAJ in the Washington navel orange. Buds located on fJr I. f rmed during the spnng (12 months old), summer (8 months old) and autumn (5 months old) flushes of growth. Only 'rJoches t~d buds were used to calculate the number of shoots developed. NS not significant. 'h~ :,prou ":-_----------------------------------------- Bud age at sprouting
5 months old 8 months old 12 months old
Sprouted Shoots per Sprouted Shoots per Sprouted Shoots per buds (%) sprouted bud buds (%) sprouted bud buds (%) sprouted bud -l nlreated 86 1.86 52 1.35 34 1.35
GA,-treated .. 73 1.84 36 1.25 25 1.20
l.c\d of signIfIcance P~O.Ol NS P~O.Ol NS P~O.OI P~0.10
I h 1. The pattern of shoot sprouting in untreated and GAJ-treated trees during the winter peak of sensitivity (I) and at J ;uting (II). Results, expressed as number of shoots per 100 nodes ± SE are the mean from 10 (C. sinensis), 6 (c. reticulata) ':; 5 trees (c. unshiu). The following shoot classes have been considered: vegetative (BV), bearing only leaves; leafy single ~\)\\ered (BC), with a sin~le flower on terminal p?si~!on; leafy multiflowered (BM); leafless multiflowered (RF); and leafless \JI1gle flowered (FS), consIsting of a Single flower. - = not present.
Treatment Number of shoots sprouted
,n there was a reduction eloped during the nat 'inciding with the effects :ct was mainly due to a Ito
f buds sprouted. The infW, er of shoots developed ;maller and not signifi JlQ bud sprouting was obse factor which had a rna ing. 'ition was obtained irres A3 application (Tab.2).
sinensis (Guardiola et maximal for leafless sh
ones. Some reduction in lIIc ered shoots was observed . we have confirmed in adll-
BV BC BM RF FS Total
Flowers Shoots
Vegetative Mixed-type Generative Total
114±9 3.5±OA 9.3±O.6 31.8±4.2 44.6±3.9 80±5 3.7±O.2 9.3±1.0 17.2±1.1 30.2±2.1 62±4 5.1±O.7 7.7±O.7 14.2±O.9 27.0± 1.2
heatment
('onlrol (untreated) (jA" Dec. l5 GA" Nov. 29
lab. 3. The influence of the level of flowering on the pattern of shoot sprouting in GA3-treated Navelate orange trees. GA3 .lpplicalions as a full coverage spray of the whole tree at a concentration of 25 mg I-Ion Nov. 29 or Dec. 15. Results, in thousands III nOlyers and shoots per tree, are the mean from 9 replications ± SE.
C, reticulala
C. lillCIlSis GArtreated I 6.5±0.5 3.8±OA 3.9±0.8 3.7±O.2 2.7±O.6 20.6± 1.9
--f~ GArtreated II 7.5±O.6 3.7±OA 3A±O.7 1.5±O.3 3.9±O.5 20.0± 1.7 Untreated 5.3±1.1 3.0±O.5 7.I±O.9 9.8± 1.4 7.5±O.8 32.7±2A
/'\~ (' rt'Iiwllllll GAJ-lreated I 5.7±O.3 8A±0.3 2.1 ±O.5 2.I±O.6 9.1±1.0 27A±1.6
GA,-treated II 6.1±1.2 8A±O.8 O.9±O.1 1.1±0.2 8.8±O.6 25.3± 1.9 Untreated 3.5±0.6 7.6±O.6 4.2±O.8 4.5±O.8 19.0± 1.9 38.8±2.3
C. IImhiu GAJ·treated I 11.3±2.9 4.8±I.O 5.1±1.8 21.2±1.6• GArtreated II 7A±1.1 7A±1.2 11.7± 1.7 26.5±2.2
Untreated 6.8±2.0 7.6±1.3 29.6±7.0 44.0±5.6
t c. sinensis
/) /\• •
Bud sprouting in vitro
139
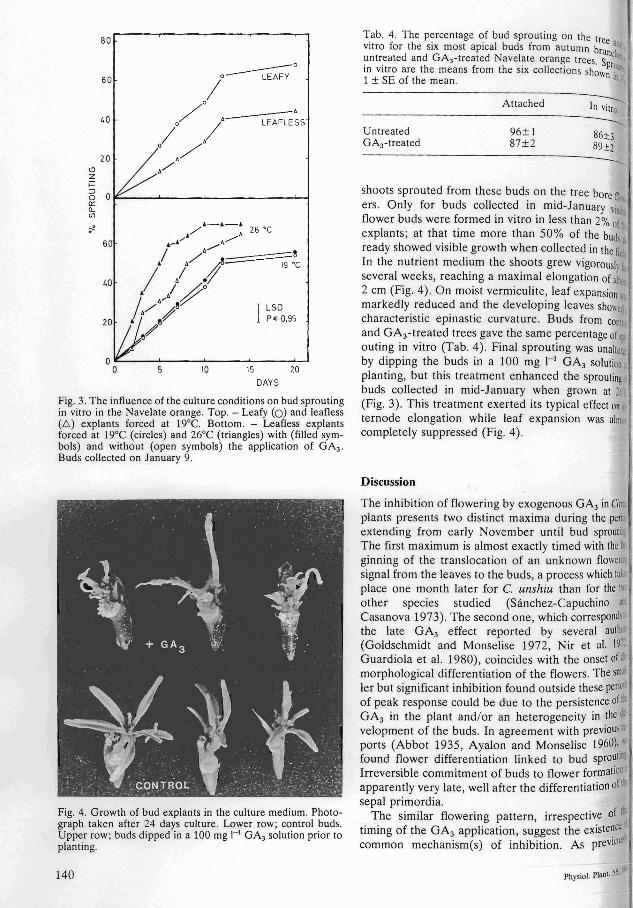
Excised buds readily sprouted in vitro, the time required to achieve 50% sprouting depending on the time of collection (Fig. 1). For the buds collected in mid-January bud sprouting was enhanced and the final percentage of sprouting increased by raising temperature from 19°C to 26°C and by the presence of the leaf lamina in the explant (Fig. 3). The latter also increased slightly the number of shoots initiated per bud explant (data not shown). Almost invariably vegetative growth was obtained in vitro, although more than 95% of the
tiollal experiments and which contrasts with the be-h<lli f .. Our 0 the other two species. The number of vegetative shoots increased slightly and sometimes significdlllIy. Further tests showed that the increase in the "~rnbcr of vegetative shoots was related to the degree (l C{)ntrol exerted on flowering. Only the most effective rcatrne t h' n n s, w lch suppressed the development of leafy
ill ores .h ~ences, Increased the number of vegetative oats significantly (Tab. 3).
Physiol. Plant 55. I
EC
ime of GA3 application on ~ ~ (.6) in the Washington n'llIClementine mandarins. The sprouting. The standard error
nber of shoots developed.
JAN FEB
,hv",n kc ti
[I1'ch,\ 'lnJil,lf
t~I11pt:1
Ih~ nL ((.lb..
.hoolS
1)nlp,lf
,doprll I o~p':I1J
01 cjng th;111 th dative I (his all(onl rast \\hok l.
l'm),C
hd.:rogc nnr.:scer
A seC( wh.:n flo ([('as.: in whik tilt
"n.:.:s dt 1(111 supp Il"w.:r III I
,\Ionst:lisl 'lipport I biggest b ,PflIUI, an :1 rev.:rslo
"ilion. as 1111111, is lih
Th.: in v h.:tween II 10 llo\Veril
eontinuous Ihe Iloweri
1\ Jifkrenl lallie beh,,, 111 October leaf slillluiu Iree is not hranch and (jAj'iIlUUCl
hanJ, the ( \prolliing ir Usually in hil db:t could
96±1 87±2
Attached
Untreated GA3-treated
Tab. 4. The percentage of bud sprouting on the tree _ vitro for the six most apical buds from autumn branc·· untreated and GA3-treated Navelate orange trees. S . in vitro are the means from the six collections show~rl . 1 ± SE of the mean. In
Discussion
shoots sprouted from these buds on the tree bore fl ers. Only for buds collected in mid-January viI flower buds were formed in vitro in less than 2% of, explants; at that time more than 50% of the bUd ready showed visible growth when collected in the fit In the nutrient medium the shoots grew vigoroush several weeks, reaching a maximal elongation of al', 2 em (Fig. 4). On moist vermiculite, leaf expansion markedly reduced and the developing leaves ShOIl(, characteristic epinastic curvature. Buds from conn and GA3-treated trees gave the same percentage ofl1 outing in vitro (Tab. 4). Final sprouting was unahcr. by dipping the buds in a 100 mg I-I GA3 solution planting, but this treatment enhanced the sproutin~
buds collected in mid-January when grown at 2;' (Fig. 3). This treatment exerted its typical effect on ternode elongation while leaf expansion was aim, completely suppressed (Fig. 4).
--In Vitro --------------------. 86±3 89±2 ---------------------.
The inhibition of flowering by exogenous GA3 in Ciu plants presents two distinct maxima during the per extending from early November until bud sproulir The first maximum is almost exactly timed with the ginning of the translocation of an unknown flowen signal from the leaves to the buds, a process which tal. place one month later for C. unshiu than for the I
other species studied (Sanchez-Capuchino J'
Casanova 1973), The second one, which correspond' the late GA3 effect reported by several auth, (Goldschmidt and Monselise 1972, Nir et aL If Guardiola et al. 1980), coincides with the onset oi:' morphological differentiation of the flowers, The SRI
ler but significant inhibition found outside these penl of peak response could be due to the persistence of' GA) in the plant and/or an heterogeneity in the 1.\ velopment of the buds, In agreement with previousr,
LSD p" 0,95
105
80
60
40
20 <:> z >= :::J
00
"" CL <fl
~
60
40
15 20
DAYS
Fig. 3. The influence of the culture conditions on bud sprouting in vitro in the Navelate orange. Top. - Leafy (0) and leafless (L) explants forced at 19°C. Bottom. - Leafless explants forced at 19°C (circles) and 26°C (triangles) with (filled symbols) and without (open symbols) the application of GA3 .
Buds collected on January 9.
ports (Abbot 1935, Ayalon and Monselise 1960).. l.ewis aile! B
found flower differentiation linked to bud sprout!'· IprOllling, si
Irreversible commitment of buds to flower formatior. bseiscd wi t
apparently very late, well after the differentiation oft' \ersion of t
Fig. 4. Growth of bud explants in the culture medium. Photograph taken after 24 days culture. Lower row; control buds. Upper row; buds dipped in a 100 mg I-I GA3 solution prior to planting.
140
sepal primordia, , The similar flowering pattern, irrespective of l:
timing of the GA3 application, suggest the existene<: viou'common mechanism(s) of inhibition. As pre I
Physiol. Plant. ;;.
\\oulu expla characterisll elalive butls Guardiola J

, bud sprouting on the tree :al buds from autumn bra j Navelate orange trees. S m the six collections sho\l'~ .
Attached
96±1 87±2
ese buds on the tree bore llected in mid-January d in vitro in less than 2"10 or nore than 50% of the bUill )wth when collected in the the shoots grew vigorous~ a maximal elongation of vermiculite, leaf expansion :he developing leaves sho curvature. Buds from
;ave the same percentage of . Final sprouting was una" a 100 mg 1-1 GA3 solutiOll
lent enhanced the sproutin~
January when grown at exerted its typical effect on
ile leaf expansion was all1lll :Fig.4).
ing by exogenous GA3 inC, :inct maxima during the (lI:lW
~ovember until bud sproul" most exactly timed with the Ition of an unknown flowe , the buds, a process which I for C. unshiu than for the :d (Sanchez-Capuchino cond one, which corresponds reported by several aut
lselise 1972, Nir et al. 19 coincides with the onset of* iation of the flowers. The lion found outside these pe be due to the persistence of or an heterogeneity in the In agreement with previoUS 'alon and Monselise 1960),. ation linked to bud sprou
It of buds to flower formatiOl II after the differentiation of
g pattern, irrespective of ication, suggest the existe~ce , of inhibition. As preVl
. weet orange (Guardiola et al. 1977) the ",n ~o.r ~ibition of bud sprouting by GA3 is the basic
..:1,:':!1\~ 111 at hioh levels of flowering (Tab. 2), and a '1I301sm ' to .
11~" mechanism appears to be operative for the ,'J1l,IJr e_cootrolled flowering (Moss 1976). Neither
'ratUr :·miX b'r of shoots developed per sprouted bud h• nuOl t;; .'.\ lor the number of lateral organs In these ! I . I). I ). I d b GA . . (Guardiola et al. 1977 IS a tere y 3 as h. ll~ 'd with the untreated control trees. Bud de
.1 10r'lr~ t is either suppressed or unaltered, which may d"rrn~11 . . f h b d B ../ n an internal condition 0 t e u. uds pro!'pcnu 0 f' .,L I "Ifless inflorescences are ar more sensitiveu':lng t;;, . ' •
h Ise developll1g mto leafy mflorescences. Veg:h.tn t {. .., . GA h'
. bud sprouting is insensitive to 3 at t IS stage. eIJU\t;; d d I . . Ihi, all-or-none effect of GA3 on bu eve opment IS m "lnlrast to the quantitative control o.f flowering for t.he
I I.: tr.:e (Moss 1969, Goldschmidt and Monsehse 1\ 1ll . d'ff b h' . h:'172). The reaSon for this ..1 erent e aVIO~r IS t. e hd.:rogcneity in bud senSitiVity to GA3 wIthm an mt!ore"ence type (Guardiola 1981).
",ecollll mechanism of flower inhibition is operative I\hen !lowering is further reduced, and involves an in.lease in the number of vegetative shoots developed I\hilc the number of both leafy and leafless in flores.elKes decreases. The activation of a subapical merislelll suppressing competitively the development of the llu\wr Illeristem has been suggested (Goldschmidt and \I11nselise 1972), but we could not find histological 'uppurt for this suggestion (data not shown). The hl~ge~t hud of every node is invariably the first to ,Plout. and quite often the only one, so the possibility of .11~\CI·sion of the apical meristems to a vegetative con.\ition. as demonstrated by Nir et al. (1972) for C. le
"''''', is likely. I he in vitro tests showed a complex inter-relationship
I'l:l\\~en the bud and its supporting branch with regard hI nuwering, sprouting and the response to GA3 . A "lntinUllus intluence of the branch is necessary to keep th"l1owering commitment of the buds, and this stimulus I' different from that translocated from the leaves. The '.ll11e behaviour was observed in vitro for buds sampled 111 Oetllher or in late January, and at this last date the leaf stimulus has been transported and flowering on the Ir~c is not inhibited by defoliation. Furthermore, the h~al1ch and not the leaves plays a central role in the ~'Al.induced inhibition of bud sprouting; on the other ,Ind. the presence of the leaf lamina promoted bud
'pruuting in vitro, an unexpected observation since it u ually inhibits axilar bud growth (Saunders 1978). This elre~t ' ld b[ . cou· e hormonal in nature as suggested by • •.... IS and Bakhshi (1968) and acts at the first stages of
J~r.(~~t:ng, ~ince in 50% of the explants the leaf lamina \ .s~d wlthlll 6 days of being transplanted. The ren\IOn of the excised buds to a vegetative condition
~h()Uld explain their lack of response to GA3 , since it is a aract·· .
. enstlC of the flower buds not shared by the vegc' JtI\e bC uds (see above) nor by the seeds (Zabala and Juardiola 1974).
.. I PI'n!. 55. 1982
Flowering appears to be a complex phenomenon which may involve hormone and metabolite levels in the buds as well as an internal condition possibly related to its location on the tree. Morphological differences are not detectable until the development of flower primordia. Well developed leaf primordia are present even in those buds forming leatless shoots, detectable growth occurring before their final abortion. GA3 may interfere with flower development at two different stages at least, although its role may not be unique since it is mimicked by 2,4-D, an auxin-like growth regulator (Guardiola et al. 1977). Our results are best explained if it is assumed that the potential development of the bud is committed before the period of time studied. The stimulus coming from the leaf allows, and the application of GA3 interferes with, the expression of the bud potential. Any interference in the development of the buds results in the inhibition of sprouting and, for the less sensitive buds, in a reversal to the vegetative condition. Evidence for this hypothesis comes from the time-course of flower inhibition by the fruit (Guardiola, J. L., Almela, V. and Agustf, M., unpublished results). Thus, gibberellin levels in the plant, whether endogenous or exogenous, may determine the intensity of flowering as suggested (Goldschmidt and Monselise 1972), but the shoot type to be formed would depend on another kind of endogenous control.
Acknowledgements - This research was supported by the Comile de Gesti6n para la Exporraci6n de los Agrios, the Comisi6n Asesora de Iovestigaci6n Cientffica y Tecnica (Granl 463 [/79), and the Excma. Diputaci6n Provincial de Valencia. Thanks are due to Mr. Joaquin Llusar and the Sociedad Naranjera de Exportaci6n nO I de Nules for the orchard facilities provided, and Mr. J. Barbera for technical assistance.
References
Abbot, C. E. 1935. Blossom bud differeniiation in citrus lrees. - Am. J. Bot. 22, 476-485.
Altman, A. & Goren, R. 1977. Differenlial effects of sucrose, abscisic acid, and benzyladenine on shoot growth and callus formation in the abscission zone of excised citrus buds. - Plant PhysioI. 59, 1161-1164.
Ayalon, S. & Monselise, S. P. 1960. Flower bud induction and differentiation in the Shamouti orange. - Proc. Am. Soc. Hortic. Sci. 75, 216-221.
Furr, J. R. & Armstrong, W. W. 1956. Flower induction in Marsh grapefruit in the Coachella valley, California. Proc. Am. Soc. Horric. Sci. 67,176-182.
Giladi, 1., Altman, A. & Goren, R. 1979. A method for aseptic culture of bud explants from citrus trees. - Sci. Hortic. 10, 357-362.
Goidschmidt, E. E. & Monselise, S. P. 1972. Hormonal control of flowering in Citrus and some other woody perennials. In Plant Growth Substances 1970 (D. J. Carr, ed.) pp. 758-766. Springer-Verlag, Berlin, ISBN 3-540-05850-8.
Guardiola, J. L. 1981. Flower initiation and development in Citrus. - Proc. Int. Soc. Citriculture. (In press).
- ,Agustf, M. & Garcia-Man, F. 1977. Gibberellic acid and \. flower bud development III sweet orange. - Proc. Int. Soc. . Citriculture, 696--699.
141

, Agusti, M., Barbera, J. & Garcia-Mari, F. 1980. Influencia de las aplicaciones de acido giberelico durante la brotacion en el desarrollo de los agrios. - Rev. Agroquim. Tecnol. Alim. 20, 139-143.
Johansen, D. A. 1940. Plant microtechnique. - McGraw Hill, New York.
Lewis, L. N. & Bakhshi, J. C. 1968. Interaction of indoleacetic acid and gibberellic acid in leaf abscission control. - Plant. Phy~ol. 43, 351-358.
Monselise, S. P. & Halevy, A. H. 1964. Chemical inhibition and promotion of Citrus flower bud induction. - Proc. Am. Soc. Hortic. Sci. 84, 141-146.
Moss, G. 1. 1969. Influence of temperature and photoperiod on flower induction and inflorescence development in sweet orange (Citrus sinensis L. Osbeck). - J. Hortic. Sci. 44,311-320. 1970. Chemical control of flower development in sweet orange (Citrus sinensis). - Aust. J. Agric. Res. 21, 233-242. 1976. Temperature effects on flower initiation in sweet orange (Citrus sinensis). - Aust. J. Agric. Res. 27, 399-407.
- & Bevington, K. B. 1977. The use of gibberellic., control alternate cropping of Late Valencia Sweet - Aust. J. Agric. Res. 28,1041-1054. 0
Murashige, T. & Skoog, F. 1962. A revised medium fo • growth and bioassays with tobacco tissue cult:' Physio!. Plant. 15,473-479. r
Nir, 1., Goren, R. & Leshem, B. 1972. Effects of Water '. gibberellic acid and 2-chloroethyltrimethylammo· chloride (Ccq on flower differentiation in Eureka t trees. - J. Am. Soc. Hortic. Sci. 97,774-778.
Sanchez-Capuchino, J. A. & Casanova, R. 1973. Ind floral en mandarinos Clementina sin hueso y Salsu Congreso Mundial de Citricultura, 2, 223-226. lilI
Saunders, P. 1978. Phytohormones and bud dormanw Phytohor~ones and Related Compounds: A Comprt~ slve Treatise. (D. S. Letham, P. B. Goodwin and T J Higgins, eds), Vol. 2, 423-445. Elsevier/North-Hull Amsterdam. ISBN 0-444-80054-9.
Zabala, G. & Guardiola, J. L. 1974. Influencia de las cubl,seminales en la germinacion de semillas de citra: Troyer. - Ann. Aula Dei 12, 188-201.
Biolog 6·(2,3~
produ,
J. Van Stad
Introduction
It is almost 1\
l-(.l-hydrox) lin), was isol. ct al. 1<)64).. in cultured c;
X 10-11 M I
active than a 1lJ67, Skoog many other sized, and th gated. In a n 10 the effeci ' N6_position (Matsubara I POrtanee of I carboxyl grot -,3-pOSition. the 3-positio Correct SlerCI he double bl
Received 20 A
", I. Plant. ~5. I'
Edited by L.E.
142 Physio!. Plant. 55.





























