Embed Size (px)
Citation preview

Human Cancer Biology
The IL-18 Antagonist IL-18–Binding Protein Is Produced inthe Human Ovarian Cancer Microenvironment
Grazia Carbotti1, Gaia Barisione1, Anna Maria Orengo1, Antonella Brizzolara1, Irma Airoldi3,Marina Bagnoli4, Patrizia Pinciroli4, Delia Mezzanzanica4, Maria Grazia Centurioni2,Marina Fabbi1, and Silvano Ferrini1
AbstractPurpose: Interleukin (IL)-18 is an immune-enhancing cytokine, which induces IFN-g production,
T-helper 1 responses, and antitumor effects. In turn, IFN-g stimulates IL-18–binding protein production,
which blocks IL-18 activity. In view of the potential use of IL-18 in epithelial ovarian cancer (EOC) immuno-
therapy, here, we studied IL-18BP expression and its regulation by cytokines in EOC cells in vitro and in vivo.
Experimental Design: Expression and production of IL-18BP in EOC cell lines, primary ovarian
carcinomas, and the corresponding normal tissues, patients’ serum, and ascites were investigated by
immunochemistry, ELISA, screening of gene expression profiles, and reverse-transcription PCR.
Results: Analysis of gene expression profiles revealed that IL18BP mRNA is increased in EOC tumors
compared with normal ovary cells. Release of IL-18BP was detectable in EOC sera and to a greater extent
in the ascites, indicating production at the tumor site. Indeed, immunochemical analyses on cells
isolated from the ascites and on tumor sections indicated that IL-18BP is expressed in both tumor cells
and tumor-associated leukocytes, which displayed a CD3�CD20�NKp46�CD13þCD14low phenotype.
EOC cell lines do not constitutively express IL-18BP. However, its release is inducible both by IFN-gstimulation in vitro and by xenotransplantation of EOC cells in immune-deficient mice, suggesting a role
for the microenvironment. In vitro experiments and immunochemistry indicated that IL-27 is also
involved in IL-18BP upregulation in EOC cell lines and primary cells through STAT1 activation. Together,
these data indicate that IL-18BP, which is produced in EOC in response to microenvironmental factors,
may inhibit endogenous or exogenous IL-18 activity. Clin Cancer Res; 19(17); 4611–20. �2013 AACR.
IntroductionEpithelial ovarian cancer (EOC) represents 80% of all
ovarian malignancies, is frequently diagnosed at advancedstages, andhas apoor 5-year survival rate (1). Evidence fromseveral studies indicates that T lymphocytes capable ofrecognizing EOC cells are present at the tumor site (2, 3)
and that T-cell–mediated immunitymay have an impact onclinical course of EOC (3). However, similar to othertumors, EOC has the ability to escape from immune systemcontrol through several mechanisms (4, 5).
Several cytokines and chemokines were found to be ele-vated in serum and in the ascites of patients with EOC andhave been implicated in tumor development, angiogenesis(6, 7), progression (8), drug resistance (9), or immunesuppression (10). Some of these cytokines have been con-sidered as serologic biomarkers of EOC (11). Among them,interleukin (IL)-18wasproposed as a potential biomarker bygene expression profiling (12) and, indeed, IL-18 levels wereelevated in serum and ascites of patients with EOC (12, 13).
IL-18 is a proinflammatory and immune-enhancing cyto-kine, which induces IFN-g production by T cells and naturalkiller (NK) cells, mediates T-helper 1 cell (TH1) polariza-tion, and is involved in the defense from pathogens (14–16). IL-18 is synthesized as an inactive precursor (pro-IL-18), which is converted to a mature form (mat-IL-18) bycaspase-1 (17). IL-18 is released from cells as both pro- andmat-IL-18 forms, but only mat-IL-18 can bind to IL-18R.Moreover, in preclinical tumormodels,mat-IL-18 exhibitedantitumor properties through the induction of an immuneresponse, whereas pro-IL-18 had no activity (18, 19).
Authors' Affiliations: Departments of 1Integrated Oncological Therapiesand 2Surgery, IRCCS AOU San Martino-IST Istituto Nazionale per laRicerca sulCancro, and 3Laboratory ofOncology, IRCCS IstitutoG.Gaslini,Genoa, Italy; and 4Department of Experimental Oncology and MolecularMedicine, Fondazione IRCCS Istituto Nazionale dei Tumori, Milan, Italy
Note: Supplementary data for this article are available at Clinical CancerResearch Online (http://clincancerres.aacrjournals.org/).
M. Fabbi and S. Ferrini contributed equally to this work.
G. Carbotti is enrolled in the Doctorate School of Genetics, University ofGenoa, Italy.
Corresponding Authors: Silvano Ferrini, UOC Terapia Immunologica,IRCCS AOU San Martino-IST Istituto Nazionale per la Ricerca sul Cancro,Largo R. Benzi 10, 16132 Genova, Italy. Phone: 0039-010-555-8372; Fax:0039-010-555-8374; E-mail: [email protected]; and Marina Fabbi,E-mail: [email protected]
doi: 10.1158/1078-0432.CCR-13-0568
�2013 American Association for Cancer Research.
ClinicalCancer
Research
www.aacrjournals.org 4611
on May 16, 2020. © 2013 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 19, 2013; DOI: 10.1158/1078-0432.CCR-13-0568

EOCcell lines release pro-IL-18, but notmat-IL-18, due todefective caspase-1 expression or activation, whereas nor-mal ovarian epithelial cells secrete mat-IL-18 (20). Indeed,IL-18 present at high levels in EOC ascites is predominantlythe inactive pro-IL-18 (13), although we cannot excludethat low levels of mat-IL-18, eventually present, may con-tribute to the immune response.
IL-18 binding protein (IL-18BP) is an inhibitor of IL-18activity, as it binds the mature form of this cytokine andblocks its interaction with IL-18R (21) and its isoform "a" isthe mostly expressed form (22). IL-18BP is produced bymonocytes and macrophages (23) and by prostatic (24)and colorectal tumor cells (25) in response to IFN-g stim-ulation. IFN-g is the physiologic inducer of IL-18BP, whichin turn inhibits IL-18 biologic activity in a negative feed-back loop. IL-18BP accumulates in the serum in chro-nic renal failure, in which it may contribute to the defectiveimmune response (26).
To further address the biologic role of endogenousIL-18 in human EOC, we conducted an integrated ana-lysis of IL-18 and IL-18BP in patients with EOC. Thestudy of IL-18BP also seemed relevant, as IL-18–basedimmunotherapy is being evaluated in patients with EOC(refs. 27, 28; NCT00659178) and high levels of IL-18BPmay interfere with this treatment. We found that, indeed,IL-18BP levels are elevated in the serum and particularlyin the ascites of patients with EOC and that IL-18BPis expressed by EOC cells and by reactive leukocytes. Ourdata also suggest that IL-18BP production may be theoutcome of cross-talk between tumor cells and the micro-environment, involving IFN-g and IL-27, a member ofthe IL-12 cytokine family (29, 30). Together, these datasupport an immune-regulatory role for IL-18BP in EOC,as it may limit the activity of endogenous or therapeuticIL-18.
Materials and MethodsCells and cell treatments
The human EOC cell lines SKOV3 (ATTC), A2780(ICLC), A2774 (IST, Genoa, Italy), and OVCAR5 (INT,Milan, Italy) were grown in RPMI-1640, with L-glutamine,
10% FCS, and antibiotics (Lonza). NK-92 cells (ATCC)were grown in medium containing 600 IU IL-2 (Novartis).A vial of each cell line master stock was recently genotypedusing the Cell ID System (Promega) and GeneMappersoftware, version 4.0.
Cells (50 � 103/well) were seeded in 24-well plates inculture medium. The following day, culture medium wasreplaced with medium with or without human recombi-nant IFN-g (PeproTech), human recombinant IL-27 (R&DSystem) or human recombinant IL-35 (Enzo Life Sciences).For IL-27R blocking experiments, an anti-gp130 antibodywas added [monoclonal antibody (mAb) 228, R&D Sys-tems)]. Treatment was carried out for 48 hours. Condi-tionedmedia were then collected, centrifuged, and used forIL-18BP detection.
RT-PCR analysis of IL18BP mRNA expressionCells were detached by trypsin and washed, and total
RNA was isolated by the NucleoSpin RNA II kit (Macherey-Nagel) and reverse-transcribed using the SuperScript IIReverse Transcriptase (Invitrogen). Quantitative RT-PCRanalysis was conducted using the following primers:POLR2A upper primer GACAATGCAGAGAAGCTGG, lowerprimer GCAGGAAGACATCATCATCC; GAPDH upper prim-er GAAGGTGAAGGTCGGAGT, lower primer CATGGGTG-GAATCATATTGGAA; IL18BP upper primer GTGTCCAGC-ATTGGAAGTGACC, and lower primer GGAGGTGCTCAA-TGAAGGAACC. Amplification was carried out by the Mas-tercycler ep realplex instrument (Eppendorf International)using the iQ� SYBR Green Supermix system (Bio-RadLaboratories). Relative quantification of mRNAs was calcu-lated by the DDCt method. For semiquantitative RT-PCR,2 mL of cDNA was separately amplified with 0.25 U ofTaq DNA Polymerase (Roche) in the presence of 1 mmol/Lof the following primers: IL18BP upper primer ACCATG-AGACACAACTGGACACCAG, lower primer TTAACCCTG-CTGCTGTGGACTGCTG; housekeeping gene ACTB upperprimer GGCATCGTGATGGACTCCG, lower primer GCTG-GAAGGTGGACAGCGA in an Eppendorf Mastercycler epGradient S. PCR products were analyzed on 1% agarose gelstained with ethidium bromide.
PatientsClinical samples were obtained upon written informed
consent and previous approval by the InstitutionalReview Board from patients and tumor-free, age-matched(median¼ 60 years; range¼ 43–75)women. All 55patientsshowed evidence of disease and untreated or off treatmentfor at least 2 months (Table 1). Ascitic fluids were collectedduring surgical procedures. Tumor histopathology, grade,and stage were assigned according to the InternationalFederation of Gynecology and Obstetrics (FIGO) criteria.
EOC xenotransplant modelFemale homozygous nonobese diabetic/severe com-
bined immunodeficient (NOD/SCID) mice (The JacksonLaboratory) were bred in-house. Nude mice were fromJanvier. The experiments were carried out according to the
Translational RelevanceInterleukin (IL)-18 is an immune-enhancing cyto-
kine, which is being studied in clinical trials of immu-notherapy. IL-18BP is a natural antagonist of IL-18and limits IL-18 biologic activity. Here, we show thatIL-18BP is produced by both tumor cells and tumor-associated leukocytes. Expression of IL-18BP in ovariancancer cells in vitro is induced by cytokines such asIFN-g and IL-27, which may play a role in the tumorenvironment. The high local levels of IL-18BP at thetumor sitemay limit the inductionof an efficient immuneresponse by either endogenous or therapeutic IL-18.
Carbotti et al.
Clin Cancer Res; 19(17) September 1, 2013 Clinical Cancer Research4612
on May 16, 2020. © 2013 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 19, 2013; DOI: 10.1158/1078-0432.CCR-13-0568
National Regulation on Animal Research Resources andapproved by the Institutional Review Board. Six-week-oldNOD/SCID animals were injected intraperitoneally with5 � 106 SKOV3 or A2774 cells. When ascites developed,animals were euthanized and blood and ascites collected.Nu/Numicewere surgically implanted in the left ovarywith2 � 106 SKOV3 cells. Tumor masses were excised and fixedin 10% buffered formalin.
ELISA for IL-18, IL-18BP, and IFN-gSamples were tested with commercially available ELISA
kits for human IL-18 (MBL), IL-18BPa (DuoSet R&DSystems), and IFN-g (Quantikine, R&D Systems). Assayswere conducted in duplicate and background values weresubtracted.
ImmunochemistryImmunochemistry detection of IL-18BP was conduct-
ed on sections of formaldehyde-fixed paraffin-embeddedcell pellets or tumors explanted from mice or on com-mercially available tissue microarrays of patients withEOC (Super Bio Chips). Antigen retrieval was done with
high-pH citrate buffer in a microwave oven. The sectionswere immunostained using rabbit anti-IL-18BP (cloneEP1088Y, Epitomics), anti-EBI3 (Novus Biologicals Eur-ope), or anti-IL27A (LifeSpan BioSciences) overnight at4�C. The antibody complex was revealed withthe EnVisionþ System-Peroxidase (Dako) and 3-amino-9-ethylcarbazole (AEC, Calbiochem). The sections werecounterstained with modified Mayer hematoxylin andmounted in PermaFluor (Thermo Scientific). The sectionswere observed with a Nikon Eclipse 80i light microscopeequipped with a color camera imaging head, using a �40objective.
Public EOC datasets of gene expressionWe explored IL18BP gene expression in our dataset
(Iorio, GSE19532) and in public EOC datasets pro-cessed through the Affymetrix HG U133 Plus 2.0 arrays(Tothill, GSE9891 and Anglesio GSE12172). Only type IItumors (31) were considered for all three datasets whenexploring pattern of correlation among different relevantgenes. We considered only the platforms includingprobes 222868_s_at because this is the only probe thatcomprises the IL-18BPa isoform. The Tothill dataset con-sists of 18 low malignant potential (LMP), 10 type I and210 type II EOC; the Anglesio dataset consists of 30 LMPand 58 type II EOC cases; and the Iorio dataset includes 17EOC high-grade tumors, two preparations of normalovarian surface epithelial cells (OSE), and six EOC celllines. Raw data were downloaded from GEO and normal-ized through the RMA algorithm (with Expression Con-sole software, Affymetrix) except for the Iorio dataset, forwhich the processed matrix was downloaded from GEO.
Statistical analysisThe normal distribution of the data was verified before
applying parametric tests by transforming data to loga-rithms. The one-way ANOVA and appropriate multiplecomparison tests were used to compare expressionlevels between patients and control subjects. The pairedStudent t test was used when appropriate. Parametricmethods were used to examine the correlation amonggene or protein expression levels (Pearson correlationcoefficient). An a level of 0.05 was used for all statisticaltests. GraphPad Prism 5.0d software and R statisticallanguage (http://www.R-project.org; version 12.2) wereused.
Western blot analysisFor the analysis of phosphorylated proteins, 107 EOC
cells were incubated for 10 minutes at 37�C with orwithout 20 ng/mL of rIL-27 in 0.5 mL of medium. Cellswere lysed in 100 mL of buffer containing 1 mmol/Lsodium orthovanadate. Lysates were resolved underreducing conditions by 10% SDS-PAGE and analyzedby Western blotting using rabbit anti-phospho-STAT1(pY701) anti-serum (Cell Signaling Technology) oranti-b actin or tubulin mAb (Sigma) and chemilumines-cence detection.
Table 1. Distribution by tumor characteristicsfor EOC patients with evidence of disease
Cases atdiagnosis, n
Cases atrelapse, n
Total 48 7
Age at diagnosis, y
>55 35 5
�55 12NA 1 2
Stagea
I 15II 7III 22 3
IV 3 3
NA 1 1
Histotype
Serous 30 4
Endometrioid 10 1
Mucinous 3Others 3 1
NA 2 1
Grade
1 132 19 3
3 15 3
Borderline
NA 1 1
Abbreviation: NA, not available.aStage at diagnosis.
IL-18BP in Ovarian Cancer
www.aacrjournals.org Clin Cancer Res; 19(17) September 1, 2013 4613
on May 16, 2020. © 2013 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 19, 2013; DOI: 10.1158/1078-0432.CCR-13-0568

ResultsGene expression of IL18BP in EOC tumors
IL18BP mRNA was first to be analyzed in our geneexpression dataset (Iorio) including primary EOC tumors,tumor cell lines, and normal ovarian surface epithelial(OSE) cells. A significantly higher expression intensity ofIL18BP was observed in tumor samples than in EOC celllines (P < 0.0001) and in OSE cells (P ¼ 0.01; Fig. 1A). Inaddition, IL18BP expressionwas significantly higher in typeII (high-grade) tumors (31) relative to low malignantpotential tumors, in two independent datasets (Tothill andAnglesio, P¼ 0.0043 and P < 0.0001, respectively), suggest-ing a possible relationship of high IL18BP expression withmalignancy (Fig. 1B).
IL-18BP levels are elevated in EOC sera and ascitesWe then tested IL-18BP serum levels in 48 patients with
EOC (Table 1), all with evidence of disease and untreated,in 7 patients at relapse, and in 13 age-matched healthywomen by ELISA for IL-18BPa, the major isoform of IL-18BP (22). IL-18BP serum levels were higher in bothuntreated (onset, mean � SD ¼ 11.06 � 5.7; median ¼9.45 ng/mL, P ¼ 0.03) and in patients with relapsing EOC(mean � SD ¼ 11.76 � 3.2; median ¼ 11.21 ng/mL, P ¼0.01) than in healthy women (mean � SD ¼ 7.4 � 3.5;median ¼ 6.86 ng/mL; Fig. 1C). By stratifying patients instage I/II and stage III/IV, no significant difference
between the two groups was observed (P ¼ 0.43), suggest-ing that IL-18BP is altered at early stages of EOC and thatIL-18BP serum levels are independent from tumor burden(Fig. 1C). In addition, when patients were stratified intype I and in type II, according to a recent classification(31), no difference was observed between the two groups(data not shown). Consistently, no differences wereobserved by stratifying patients in accordance with tumorgrading (data not shown). Performance of serum IL-18BPas classifier, evaluated by Receiver Operating Characteris-tic analysis, yielded an area under the curve of 0.734,suggestive of a poor performance (Supplementary Fig. S1).
To gain information on the possible tumor origin ofthe high serum IL-18BP levels, we tested 18 EOC serumand ascites pairs, collected at the same time. By pairedanalysis, IL-18BP levels were significantly higher in theascites than in serum of the same patients (mean � SD ¼31.9 � 14.7, median 32.8 vs. mean � SD ¼ 11.7 � 5.9,median 10.5 ng/mL, P < 0.0001; Fig. 1D), thus suggestingthat IL-18BP derives from the tumor site, where it couldlimit the IL-18–driven immune response.
Analyses of IL-18BP correlation with IL-18 and IFN-gIt is known that IL-18BP inhibits IL-18 as the result of a
negative feedback loop mediated by IL-18–induced IFN-g(21, 23). In agreement with our previous data (13), IL-18ELISA levels were elevated in EOC serum and ascites (Sup-plementary Fig. S2A and S2B).
Figure 1. Expression of IL18BP mRNA and secretion of IL-18BP protein in EOC. A, normalized IL18BP gene expression levels in EOC tumors, cell lines,and OSE. Expression levels were significantly higher in tumor samples (one-way ANOVA and Bonferroni post-test). B, IL18BP mRNA expression wassignificantly higher in high-grade tumors (type II) relative to low malignant potential tumors (LMP), in two independent datasets by two-sided unpairedStudent t test (Anglesio dataset) and one-way ANOVA and Tukey post-test (Tothill dataset). Boxes and whiskers represent median and quartileswith minimum and maximum. C, IL-18BP ELISA levels were significantly higher in sera from EOC patients at diagnosis (n ¼ 48) or at relapse (n ¼ 7)than in age-matched female controls (n ¼ 13) by Kruskal–Wallis test. IL-18BP levels were similar in stage I/II and stage III/IV patients. D, IL-18BPlevels were higher in the ascites than in sera simultaneously collected from the same patient (by two-sided paired Student t test). Mean and SD areindicated in C and D.
Carbotti et al.
Clin Cancer Res; 19(17) September 1, 2013 Clinical Cancer Research4614
on May 16, 2020. © 2013 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 19, 2013; DOI: 10.1158/1078-0432.CCR-13-0568
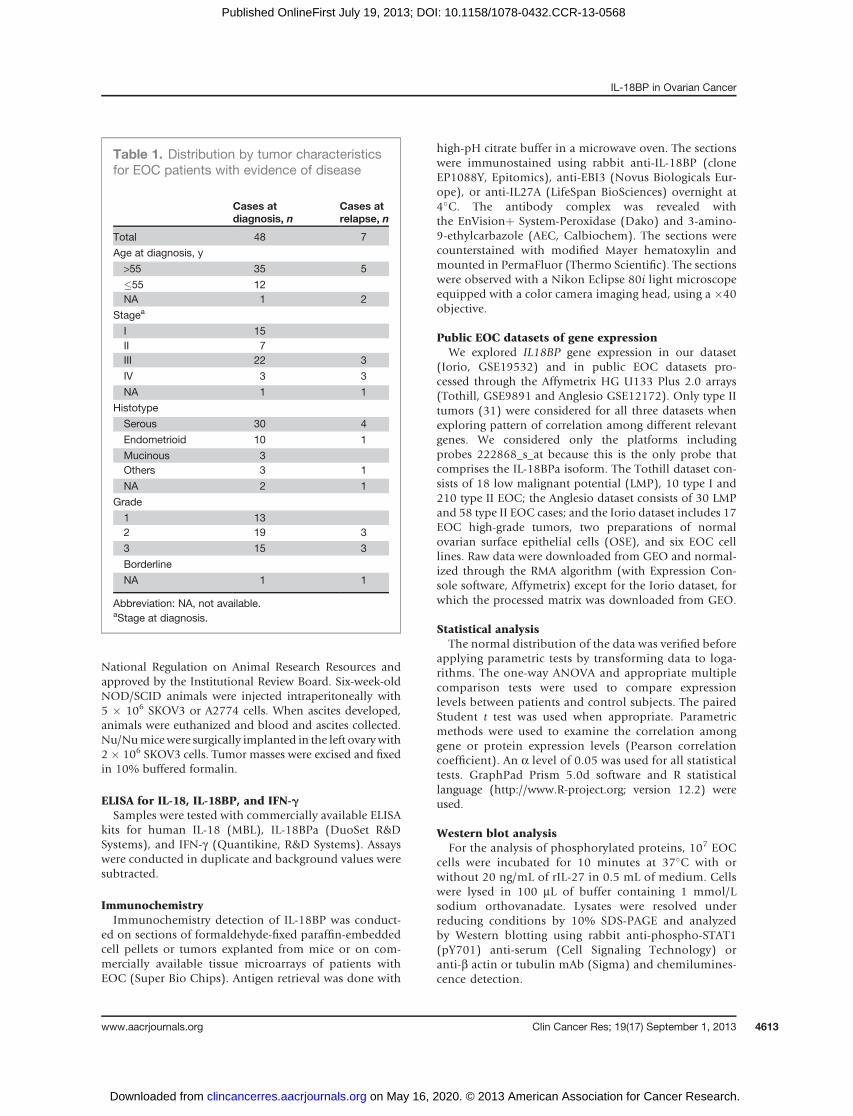
We therefore first evaluated whether there was anyrelationship between IL18 and IL18BP mRNA in twopublic access datasets of ovarian cancers and found amoderate, yet significant correlation (Anglesio dataset,P ¼ 0.0056; Tothill dataset, P < 0.0001; Fig. 2A). In addi-tion, a correlation was found for IFNG and IL18BP andfor IFNG and IL18 in the Tothill dataset (IFNG vs. IL18,P < 0.0001; IL18BP vs. IFNG, P < 0.0001), although onlya minority of patients showed elevated levels of IFNGexpression (Fig. 2B).However, when we investigated possible correlations at
the protein level, we found no significant correlationbetween IL-18 and IL-18BP in the sera (r ¼ 0.12, P ¼ nsby Pearson correlation) and in the ascites (r¼ 0.3, P¼ns) ofpatients with EOC (Supplementary Fig. S2A and S2B).Moreover, IFN-g levels were low or undetectable in ascitesand sera from patients with EOC, in agreement with pre-vious reports, and showed no correlation with IL-18 andIL-18BP (Supplementary Fig. S2C).Although microarray data suggest that the IL-18/IFN-g
axismaybe involved in the regulationof IL-18BP expressionin the tumor tissue of a few patients, no evidence of thisregulation was found in sera and ascites.
Both EOC cells and reactive cells from themicroenvironment express IL-18BPTo address the possible cellular origin of the elevated
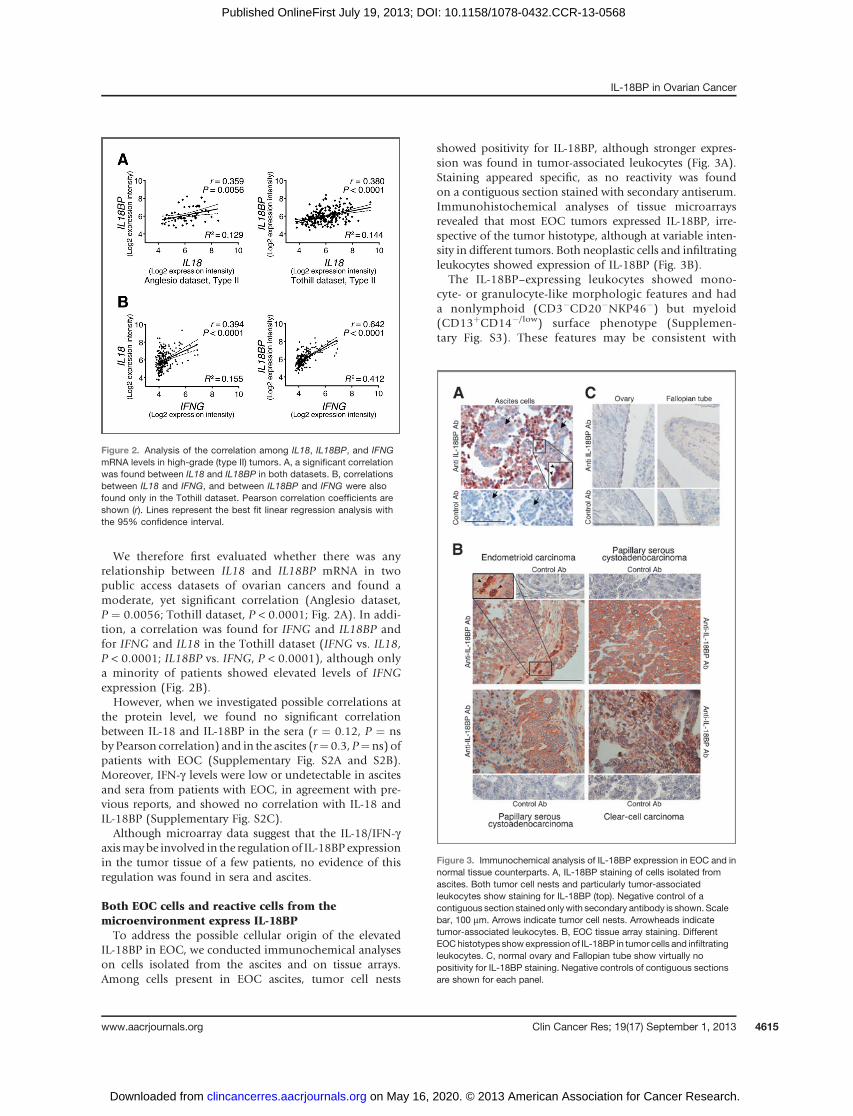
IL-18BP in EOC, we conducted immunochemical analyseson cells isolated from the ascites and on tissue arrays.Among cells present in EOC ascites, tumor cell nests
showed positivity for IL-18BP, although stronger expres-sion was found in tumor-associated leukocytes (Fig. 3A).Staining appeared specific, as no reactivity was foundon a contiguous section stained with secondary antiserum.Immunohistochemical analyses of tissue microarraysrevealed that most EOC tumors expressed IL-18BP, irre-spective of the tumor histotype, although at variable inten-sity in different tumors. Both neoplastic cells and infiltratingleukocytes showed expression of IL-18BP (Fig. 3B).
The IL-18BP–expressing leukocytes showed mono-cyte- or granulocyte-like morphologic features and hada nonlymphoid (CD3�CD20�NKP46�) but myeloid(CD13þCD14�/low) surface phenotype (Supplemen-tary Fig. S3). These features may be consistent with
Figure 2. Analysis of the correlation among IL18, IL18BP, and IFNGmRNA levels in high-grade (type II) tumors. A, a significant correlationwas found between IL18 and IL18BP in both datasets. B, correlationsbetween IL18 and IFNG, and between IL18BP and IFNG were alsofound only in the Tothill dataset. Pearson correlation coefficients areshown (r). Lines represent the best fit linear regression analysis withthe 95% confidence interval.
Figure 3. Immunochemical analysis of IL-18BP expression in EOC and innormal tissue counterparts. A, IL-18BP staining of cells isolated fromascites. Both tumor cell nests and particularly tumor-associatedleukocytes show staining for IL-18BP (top). Negative control of acontiguous section stained onlywith secondary antibody is shown. Scalebar, 100 mm. Arrows indicate tumor cell nests. Arrowheads indicatetumor-associated leukocytes. B, EOC tissue array staining. DifferentEOChistotypes show expression of IL-18BP in tumor cells and infiltratingleukocytes. C, normal ovary and Fallopian tube show virtually nopositivity for IL-18BP staining. Negative controls of contiguous sectionsare shown for each panel.
IL-18BP in Ovarian Cancer
www.aacrjournals.org Clin Cancer Res; 19(17) September 1, 2013 4615
on May 16, 2020. © 2013 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 19, 2013; DOI: 10.1158/1078-0432.CCR-13-0568

"myeloid-derived suppressor cells" (32). Together, thesedata indicate that different cell populations express IL-18BP in the EOCmicroenvironment. In contrast, normalovary and Fallopian tube tissue showed no expressionof IL-18BP by immunohistochemistry (Fig. 3C).
IFN-g and IL-27 upregulate IL-18BP in human EOC celllines
Unlike EOC cells present in tumor specimen, four EOCcell lines showed no constitutive expression of IL-18BPmRNA or protein (Fig. 4). However, culture in the presenceof IFN-g increased IL-18BP protein secretion (Fig. 4A) andIL18BP mRNA (Fig. 4B) expression in EOC cell lines. Inaddition, although human IL-18BP was undetectable in
sera, it was found in the ascites of nude mice bearingorthotopic xenotransplants of the human A2774 andSKOV3 cell lines (Fig. 4C), further suggesting that EOCcells can contribute to IL-18BP production in vivo. Indeed,A2774 and SKOV3 cells grown in immune-deficient miceshowed IL-18BP expression by immunohistochemistry,whereas IL-18BP was virtually undetectable in the sameEOC cell lines in vitro (Supplementary Fig. S4). Thesefindings suggest that factor(s) present in the microenviron-ment are responsible for production of IL-18BP in vivo.Although IFN-g mediates IL-18BP expression in EOC celllines in vitro, it should not be involved in vivo, as it wasvirtually undetectable in human ascites from patients and,moreover, mouse IFN-g is inactive on human cells. Theseconsiderations prompted us to examine whether othercytokines, known to be elevated in patients with EOC, suchas IL-6, TNF-a, VEGF-A, EGF, IL-18, and IL-8, couldmediateIL-18BP expression in EOC cell lines, but none provedactive (data not shown).
A recent report indicated that the heterodimeric cyto-kine IL-27, consisting of EBI3 and IL-27 chains, couldmediate IL-18BP production by human keratinocytesin an IFN-g–independent manner (33). We then testedwhether IL-27 or the related cytokine IL-35 could medi-ate IL-18BP expression in EOC cells. Indeed, IL-27induced IL-18BP secretion (Fig. 5A; S5A and B) in fourEOC cell lines in a dose-dependent fashion, whereasIL-35 showed no activity (not shown). IL-27 also increas-ed IL18BP mRNA expression (Supplementary Fig. S5C)Importantly, the IL-18BP–containing supernatant ofIL-27–stimulated A2780 cells significantly inhibited IL-18 bioactivity in a concentration-dependent manner, asdetected through IFN-g release by the human NK cell lineNK-92. Controls such as IL-27–containing medium orthe supernatant from unstimulated EOC cells producedno inhibition (Fig. 5B).
Because IL-18BP expression is activated through theSTAT1 pathway, we also analyzed STAT1 activation by IL-27 in EOC cells. Indeed, IL-27 activates STAT1 signaling inEOC cell lines (Fig. 5C and Supplementary Fig. S5D), asreported for other IL-27–sensitive cell types (33, 34). Inaddition, Western blot analysis showed that STAT1 wasconstitutively tyrosine-phosphorylated in cells isolatedfrom ascites ex vivo and was further activated by in vitrotreatment with IL-27 (Fig. 5C). Confocal microscopy con-firmed constitutive STAT1 phosphorylation, which increas-ed with IL-27 stimulation, in both EPCAM-positive EOCand EPCAM-negative inflammatory cells (SupplementaryFig. S5E). Consistently, cells isolated from ascites showedspontaneous IL-18BP secretion in culture, which could befurther enhanced by in vitro IL-27 stimulation (Fig. 5A).
Because IL-27 activity is mediated through a heterodi-meric receptor consisting of gp-130 and WSX-1 molecules,we asked whether antibodies neutralizing gp-130 could in-hibit the effect of IL-27. Anti-gp130 mAb significantlyinhibited IL-27–mediated IL-18BP production in two dif-ferent EOC cell lines, further supporting the involvement ofthe IL-27/IL-27R pathway in IL-18BP regulation (Fig. 5A).
Figure 4. IL-18BP expression by EOC cell lines. A, IL-18BP secretion isdetected by ELISA in culture supernatants following EOC cell treatmentwith IFN-g (100 and 1,000 U/mL for 24 hours. �, P < 0.05; ��, P < 0.01 bytwo-sided unpaired Student t test). B, IFN-g–induced IL18BP mRNAexpression in EOC cell lines as detected by RT-PCR analysis. C, HumanIL-18BP ascites levels were measured in NOD/SCID mice bearingorthotopic xenotransplants of the SKOV3 (n¼ 5) and A2774 (n¼ 2) EOCcell lines.
Carbotti et al.
Clin Cancer Res; 19(17) September 1, 2013 Clinical Cancer Research4616
on May 16, 2020. © 2013 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 19, 2013; DOI: 10.1158/1078-0432.CCR-13-0568

IL-27A and EBI3 are expressed in EOC tissuesFurther analyses of two microarray datasets of EOC indi-
cated a correlation between the expression of EBI3 andIL18BP mRNA in EOC primary tumors (SupplementaryFig. S6A). In addition, although IL18BP mRNA expressionshowed no significant correlation with outcome (notshown), high levels of EBI3 expression correlated with ashorter relapse-free survival (Supplementary Fig. S6B). Thecorrelation between IL18BP and EBI3 gene expression sug-
gested a possible paracrine loop of IL-18BP inductionin vivo. This hypothesis was also suggested by the use ofan anti-IL-27A (p28) specific antibody in immunochem-istry, which revealed IL-27 expression predominantly in afraction of tumor-associated leukocytes isolated from theascites and in tissue microarrays, whereas tumor cell nestsappeared negative (Fig. 5C and D). Also, EBI3 proteinshowed a similar distribution both in ascites (Supplemen-tary Fig. S7A) and within tumor tissues (Supplementary
Figure 5. Involvement of IL-27 in IL-18BP expression in EOC cells. A, IL-18BP secretion is detected by ELISA in culture supernatants followingA2780 and A2774 EOC cell stimulation with IL-27 (20 ng/mL for 48 hours, P < 1E-05 vs. untreated control). Anti-gp130 mAb (from 0.5 to 500 ng/mL)significantly inhibits IL-18BP secretion mediated by IL-27 stimulation (�, P < 0.05; ��, P < 0.001; ���P < 1E-05). Black bars show constitutive andIL-27–induced IL-18BP in vitro secretion by cells isolated from patients' ascites A98, representative of three cases with similar results. B, theconditioned medium of IL-27–stimulated A2780 cells containing 2 ng/mL IL-18BP (BPþcm) significantly inhibits IL-18–induced IFN-g release by theNK-92 cells in a concentration (50, 25, or 12%v/v)-dependent fashion (�, P < 0.05; ��, P < 0.001). As controls, conditioned medium of unstimulatedcells (ctrl cm) and IL-27–containing medium (IL-27þ cm) were used. C, Western blot analysis of tyrosine-phosphorylated STAT1 protein in A2780cells and in cells isolated from three different ascites unstimulated or stimulated for 10 minutes with IL-27 (20 ng/mL); b-actin was used as loadingcontrol. D, tumor-associated leukocytes show staining for IL-27A, whereas tumor cell nests show no reactivity (top). Negative control of a contiguoussection is shown. Scale bar, 100 mm. Arrows indicate examples of negative tumor cell nests. E, EOC tissue array staining. Different EOC histotypesshow expression of IL-27A in infiltrating leukocytes (enlarged in the inset).
IL-18BP in Ovarian Cancer
www.aacrjournals.org Clin Cancer Res; 19(17) September 1, 2013 4617
on May 16, 2020. © 2013 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 19, 2013; DOI: 10.1158/1078-0432.CCR-13-0568

Fig. S7B). Together, our data are consistent with a para-crine activation of IL-18BP expression in the tumormicroenvironment.
DiscussionIn this study, we show that IL-18BP levels are elevated in
the serum of patients with EOC and are even higher in theascites, reaching four-fold higher levels than those found innormal serum. This finding suggested a local production ofIL-18BP in the microenvironment of EOC. Indeed, immu-nochemical analyses showed that IL-18BP is expressed byneoplastic cells of different EOC histotypes and by tumor-associated leukocytes with myeloid features. Therefore, thehigh concentration of IL-18BP in the tumor environmentof EOC may limit the effect of endogenous or exogenouslyadministered IL-18. On the other hand, IL-18BP also bindsthe anti-inflammatory cytokine IL-37 (35), which maysuppress the host immune response (36), and this mayresult in a beneficial effect for the host. However, to ourknowledge, no evidence for IL-37 expression in ovariancancer has been provided to date.
Elevated IL-18BP levels were recently described also inserum of patients with prostatic (24) and pancreatic cancer(37). In the latter, the concomitantly elevated levels of free-IL-18 in the serum suggested the existence of a biologicparadox, in view of the immune-enhancing properties ofIL-18. Indeed, increased levels of both IL-18 and its naturalinhibitor IL-18BP were also found in patients with systemiclupus erythematosus, inwhombiologically active free IL-18was still higher than in controls and was amarker of diseaseactivity and a potential contributor to autoimmunity (38).
It was previously shown that high levels of IL-18 arepresent in serum and ascites of patients with EOC (12)and that pro-IL-18 is largely predominant (13). In fact,although IL-18 ELISA preferentially recognized mat-IL-18in sera, this assay also detected pro-IL-18, albeit with areduced sensitivity. Therefore, the presence of "free IL-18"in EOC could be explained by the predominance of pro-IL-18, which is unable to bind IL-18BP and is detected by IL-18ELISA. It is likely that a similar situation may occur in othertumors in which alterations of IL-18 processing have beenreported (39).
Because the presence of mat-IL-18 in the ascites of EOCcould not be formally excluded andmat-IL-18 is an inducerof IL-18BP, via IFN-g production (25, 40), we exploredpossible correlations between immune-reactive IL-18 andIL-18BP or IFN-g levels. No significant correlation wasfound in the ascites and serum of patients with EOC, andIFN-g levels were very low to undetectable in the ascites.However, a correlation between IL18 and IL18BP mRNAlevels was found in two independent datasets of EOC geneexpression profiles. Moreover, IFNG mRNA showed a cor-relation with IL18BP in one dataset, although IFNGmRNAwas elevated only in a minority of cases. These data suggestthat the IL-18/IFN-g loop may be active in the tumor tissuemicroenvironment in somepatients and that other factor(s)may participate in the induction of IL-18BP expression.
Evidence exists to suggest that IL-18BP production is aresult of the interaction between EOC cells and the micro-environment. Human EOC cell lines do not produceIL-18BP in culture but, once grafted in immune-deficientmice, they display IL-18BP expression, suggesting a rolefor factor(s), which function across the species. This is notthe case for IFN-g , in view of its species specificity. Becauseother cytokines, which are elevated in the ascites of EOCfailed to induce IL-18BP expression in vitro, we focused onIL-27, which was recently shown to stimulate IL-18BPexpression in human keratinocytes (33).
IL-27 is a member of the IL-12 family that may have pro-or anti-inflammatory properties in different systems(29, 30). IL-27 is a heterodimeric cytokine, composed ofp28 and EBV-induced gene 3 (EBI3), which upregulatesIL-12R expression and is relevant for TH1 polarization(41, 42). However, the precise contribution of IL-27 toimmune response, inflammation, and cancer is still poorlyunderstood. On one hand, IL-27 has proinflammatoryeffects through the induction of CXCL10 in macrophagesin inflammatory skin disorders (43). However, IL-27may limit the proinflammatory and immune-enhancingactivities of IL-18 in the skin through IL-18BP induction(33) and may dampen autoimmunity, as Il27�/� micewere more susceptible to experimental autoimmune en-cephalomyelitis (44).
Here, we show that IL-27 induces the expression ofIL-18BP mRNA and protein in human EOC cell lines inculture and activates STAT1 signaling in these cells, whereasIL-35, another EBI3-containing cytokine (30), was inactive.Such activity was specifically induced through the IL-27Rcomplex as indicated by the significant inhibition ofIL-18BP induction upon treatment with a neutralizingantibody against the gp130 chain. A potential role ofIL-27 in vivo was suggested by the expression of IL-27A andEBI3 found by immunochemistry in tumor-associated leu-kocytes in both ascites and tumor tissues and by the cor-relation between EBI3 and IL18BP mRNA expression intwo different EOC datasets. Interestingly, high EBI3 expres-sion correlated with a shorter progression-free survival intype II tumors. The finding that IL18BP gene expressionhad no significant correlation with relapse-free survival ofpatients with type II tumors may reflect the multiplicity ofcomponents driving clinical outcome. A correlation withIL27A mRNA expression could not be found (data notshown), but this may relate to technical limitations, as onlyone probeset was present in the arrays. The possible role ofIL-27 in vivo was reinforced by the detection of constitutiveSTAT1 activation in both neoplastic and reactive cells iso-lated from the ascites, in the absence of measurable IFN-glevels. Moreover, these ascites cells showed spontaneoussecretion of IL-18BP in culture, which could be furtherenhanced by the addition of exogenous IL-27. Consistently,STAT1 phosphorylation also was increased by IL-27.
Our study may open new perspectives on understandingthe role of IL-27 in cancer, as its involvement in theantitumor immune response is still poorly understood. Insome hematologic neoplasia, including multiple myeloma
Carbotti et al.
Clin Cancer Res; 19(17) September 1, 2013 Clinical Cancer Research4618
on May 16, 2020. © 2013 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 19, 2013; DOI: 10.1158/1078-0432.CCR-13-0568

(45) and acute leukemias (46, 47), IL-27 displays direct andindirect antitumor effects.We thus hypothesize that in EOCIL-27may be part of an immune-regulatory network, whichlimits the induction of TH1 responses and IFN-g productionin the microenvironment by inhibiting IL-18 activity. Insupport of this concept, studies inmurinemodels highlight-ed a predominant role of IL-27 as an immune-regulatoryand anti-inflammatory agent that generates and maintainsT-regulatory cell functions (48) and induces IL-10 produc-tion by T lymphocytes (49).Although the role of IL-18 in tumor cell biology has
been debated (50), preclinical studies indicated that IL-18displays antitumor activity through its ability to triggerIFN-g production and to favor the induction of a TH1response (18, 19). Therefore, recombinant IL-18 is under-going testing in clinical trials of cancer immunotherapy(27, 28) and in particular a clinical phase I study of IL-18and doxorubicin in advanced-stage EOC (NCT00659178)has recently concluded patient recruitment. It is possiblethat the high local levels of IL-18BP present in EOC maylimit the biologic effects of low levels of endogenous IL-18 or of therapeutically administered IL-18, particularlyat the tumor site.
Disclosure of Potential Conflicts of InterestNo potential conflicts of interest were disclosed.
Authors' ContributionsConception and design: M. Fabbi, S. FerriniDevelopment of methodology: G. Carbotti, G. Barisione, A. BrizzolaraAcquisitionofdata (provided animals, acquired andmanagedpatients,provided facilities, etc.): G. Carbotti, A.M. Orengo, A. Brizzolara, M.G.CenturioniAnalysis and interpretation of data (e.g., statistical analysis, biosta-tistics, computational analysis): I. Airoldi, M. Bagnoli, P. Pinciroli,D. Mezzanzanica, M. FabbiWriting, review, and/or revision of the manuscript: G. Carbotti, I. Airoldi,M. Bagnoli, P. Pinciroli, D. Mezzanzanica, M. Fabbi, S. FerriniAdministrative, technical, or material support (i.e., reporting or orga-nizing data, constructing databases): I. AiroldiStudy supervision: M. Fabbi, S. Ferrini
Grant SupportThis studywas supported by AIRC (Associazione Italiana per la Ricerca sul
Cancro, IG5509, IG13018 and IG13518), Ministry of Health Project"Tumori Femminili," and Compagnia di San Paolo.
The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby markedadvertisement in accordance with 18 U.S.C. Section 1734 solely to indicatethis fact.
Received February 27, 2013; revised June 21, 2013; accepted July 8, 2013;published OnlineFirst July 26, 2013.
References1. Martin LP, Schilder RJ. Management of recurrent ovarian carcinoma:
current status and future directions. Semin Oncol 2009;36:112–25.2. Ferrini S, Biassoni R, Moretta A, Bruzzone M, Nicolin A, Moretta L.
Clonal analysis of T lymphocytes isolated from ovarian carcinomaascitic fluid. Phenotypic and functional characterization of T-cellclones capable of lysing autologous carcinoma cells. Int J Cancer1985;36:337–43.
3. Nelson BH. The impact of T-cell immunity on ovarian cancer out-comes. Immunol Rev 2008;222:101–16.
4. Frumento G, Piazza T, Di Carlo E, Ferrini S. Targeting tumor-relatedimmunosuppression for cancer immunotherapy. Metab Immune Dis-ord Drug Targets 2006;6:233–7.
5. Preston CC, Goode EL, Hartmann LC, Kalli KR, Knutson KL. Immunityand immune suppression in human ovarian cancer. Immunotherapy2011;3:539–56.
6. Nilsson MB, Langley RR, Fidler IJ. Interleukin-6, secreted by humanovarian carcinoma cells, is a potent proangiogenic cytokine. CancerRes 2005;65:10794–800.
7. Leinster DA, Kulbe H, Everitt G, Thompson R, Perretti M, Gavins FN,et al. The peritoneal tumour microenvironment of high-grade serousovarian cancer. J Pathol 2012;227:136–45.
8. Berek JS, Chung C, Kaldi K, Watson JM, Knox RM, Martinez-Maza O.Serum interleukin-6 levels correlatewith disease status in patientswithepithelial ovarian cancer. Am J Obstet Gynecol 1991;164:1038–43.
9. Duan Z, Feller AJ, Penson RT, Chabner BA, Seiden MV. Discovery ofdifferentially expressed genes associated with paclitaxel resistanceusing cDNA array technology: analysis of interleukin (IL) 6, IL-8, andmonocyte chemotactic protein 1 in the paclitaxel-resistant phenotype.Clin Cancer Res 1999;5:3445–53.
10. Curiel TJ, Coukos G, Zou L, Alvarez X, Cheng P, Mottram P, et al.Specific recruitment of regulatory T cells in ovarian carcinoma fostersimmune privilege and predicts reduced survival. Nat Med 2004;10:942–9.
11. Lambeck AJ, Crijns AP, Leffers N, Sluiter WJ, ten Hoor KA, Braid M,et al. Serum cytokine profiling as a diagnostic and prognostic tool inovarian cancer: a potential role for interleukin 7. Clin Cancer Res2007;13:2385–91.
12. LePageC,Ouellet V,Madore J,Hudson TJ, Tonin PN,ProvencherDM,et al. From gene profiling to diagnostic markers: IL-18 and FGF-2complement CA125 as serum-based markers in epithelial ovariancancer. Int J Cancer 2006;118:1750–8.
13. Orengo AM, Fabbi M, Miglietta L, Andreani C, Bruzzone M, Puppo A,et al. Interleukin (IL)-18, a biomarker of human ovarian carcinoma, ispredominantly released as biologically inactive precursor. Int J Cancer2011;129:1116–25.
14. Okamura H, Tsutsui H, Komatsu T, YutsudoM, Hakura A, Tanimoto T,et al. Cloning of a new cytokine that induces interferon-gamma. Nature1995;378:88–91.
15. Takeda K, Tsutsui H, Yoshimoto T, Adachi O, Yoshida N, Kishimoto T,et al. Defective NK cell activity and Th1 response in IL-18-deficentmice. Immunity 1998;8:383–90.
16. Dinarello CA. IL-18: a Th1-inducing, proinflammatory cytokine andnew member of the IL-1 family. J Allergy Clin Immunol 1999;103:11–24.
17. Ghayur T, Banerjee S, Hugunin M, Butler D, Herzog L, Carter A,et al. Caspase-1 processes IFN-gamma-inducing factor and reg-ulates LPS-induced IFN-gamma production. Nature 1997;386:619–23.
18. Heuer JG, McClung CT, Hock RA. Neuroblastoma cells expressingmature IL-18, but not pro-IL-18, induce a strong and immediateantitumour immune response. J Immunother 1999;22:324–35.
19. Nakamori M, Iwahashi M, Nakamura M, Ueda K, Zhang X, Yamaue H.Intensification of antitumor effect by T helper 1-dominant adoptiveimmunogene therapy for advanced orthotopic colon cancer. ClinCancer Res 2003;9:2357–65.
20. WangZY,GaggeroA, Rubartelli A, RossoO,Miotti S,MezzanzanicaD,et al. Expression of interleukin-18 in human ovarian carcinoma andnormal ovarian epithelium: evidence for defective processing in tumorcells. Int J Cancer 2002;98:873–8.
21. Novick D, Kim SH, Fantuzzi G, Reznikov LL, Dinarello CA, RubinsteinM. Interleukin-18 binding protein: a novel modulator of the Th1 cyto-kine response. Immunity 1999;10:127–36.
22. KimSH, EisensteinM, Reznikov L, Fantuzzi G, NovickD, RubinsteinM,et al. Structural requirements of six naturally occurring isoforms of
IL-18BP in Ovarian Cancer
www.aacrjournals.org Clin Cancer Res; 19(17) September 1, 2013 4619
on May 16, 2020. © 2013 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 19, 2013; DOI: 10.1158/1078-0432.CCR-13-0568

the IL-18 binding protein to inhibit IL-18. Proc Natl Acad Sci U S A2000;97:1190–5.
23. Veenstra KG, Jonak ZL, Trulli S, Gollob JA. IL-12 inducesmonocyte IL-18 binding protein expression via IFN-gamma. J Immunol 2002;168:2282–7.
24. Fujita K, Ewing CM, Isaacs WB, Pavlovich CP. Immunomodulatory IL-18 binding protein is produced by prostate cancer cells and its levels inurine and serum correlate with tumor status. Int J Cancer 2011;129:424–32.
25. Paulukat J, BosmannM,NoldM,GarkischS, K€ampfer H, Frank S, et al.Expression and release of IL-18 Binding Protein in Response to INF-g.J Immunol 2001;167:7038–43.
26. Dinarello CA, Novick D, Rubinstein M, Lonnemann G. Interleukin 18and interleukin 18 binding protein: possible role in immunosuppres-sion of chronic renal failure. Blood Purif 2003;21:258–70.
27. Robertson MJ, Mier JW, Logan T, Atkins M, Koon H, Koch KM, et al.Clinical and biological effects of recombinant human interleukin-18administered by intravenous infusion to patients with advancedcancer. Clin Cancer Res 2006;12:4265–73.
28. Robertson MJ, Kirkwood JM, Logan TF, Koch KM, Kathman S, KirbyLC, et al. A dose-escalation study of recombinant human interleukin-18 using two different schedules of administration in patients withcancer. Clin Cancer Res 2008;14:3462–9.
29. Trinchieri G, Pflanz S, Kastelein RA. The IL-12 family of heterodimericcytokines: new players in the regulation of T cell responses. Immunity2003;19:641–4.
30. Vignali DA, Kuchroo VK. IL-12 family cytokines: immunological play-makers. Nat Immunol 2012;13:722–8.
31. Kurman RJ, Shih IeM. Molecular pathogenesis and extraovarian originof epithelial ovarian cancer-Shifting the paradigm. Hum Pathol 2011;42:918–31.
32. Ostrand-Rosenberg S, Sinha P. Myeloid-derived suppressor cells:linking inflammation and cancer. J Immunol 2009;182:4499–506.
33. Wittmann M, Doble R, Bachmann M, Pfeilschifter J, Werfel T, M€uhl H.IL-27 regulates IL-18 binding protein in skin resident cells. PLoS ONE2012;7:e38751.
34. Bachmann M, Paulukat J, Pfeilschifter J, M€uhl H. Molecular mechan-isms of IL-18BP regulation in DLD-1 cells: pivotal direct action ofthe STAT1/GAS axis on the promoter level. J Cell Mol Med 2009;13:1987–94.
35. Bufler P, Azam T, Gamboni-Robertson F, Reznikov LL, Kumar S,Dinarello CA, et al. A complex of the IL-1 homologue IL-1F7b andIL-18-binding protein reduces IL-18 activity. Proc Natl Acad Sci U S A2002;99:13723–8.
36. Nold MF, Nold-Petry CA, Zepp JA, Palmer BE, Bufler P, Dinarello CA.IL-37 is a fundamental inhibitor of innate immunity. Nat Immunol2010;11:1014–22.
37. Carbone A, Vizio B, Novarino A, Mauri FA, Geuna M, Robino C, et al.IL-18 paradox in pancreatic carcinoma: elevated serum levels of
free IL-18 are correlated with poor survival. J Immunother 2009;32:920–31.
38. Migliorini P, Anzilotti C, Pratesi F, Quattroni P, Bargagna M, Dinar-ello CA, et al. Serum and urinary levels of IL-18 and its inhibitor IL-18BP in systemic lupus erythematosus. Eur Cytokine Netw 2010;21:264–71.
39. Pag�es F, Berger A, Henglein B, Piqueras B, Danel C, Zinzindohoue F,et al. Modulation of interleukin-18 expression in human colon carci-noma: consequences for tumour immune surveillance. Int J Cancer1999;84:326–30.
40. Fantuzzi G, Reed D, Qi M, Scully S, Dinarello CA, Senaldi G. Role ofinterferon regulatory factor-1 in the regulation of IL-18 production andactivity. Eur J Immunol 2001;31:369–75.
41. Pflanz S, Timans JC, Cheung J, Rosales R, Kanzler H, Gilbert J, et al.IL-27, a heterodimeric cytokine composed of EBI3 and p28 protein,induces proliferation of naive CD4(þ) T cells. Immunity 2002;16:779–90.
42. Lucas S, Ghilardi N, Li J, de Sauvage FJ. IL-27 regulates IL-12responsiveness of naive CD4þ T cells through Stat1-dependent and-independent mechanisms. Proc Natl Acad Sci U S A 2003;100:15047–52.
43. Shibata S, Tada Y, Kanda N, Nashiro K, Kamata M, KarakawaM, et al.Possible roles of IL-27 in the pathogenesis of psoriasis. J InvestDermatol 2010;130:1034–9.
44. Batten M, Li J, Yi S, Kljavin NM, Danilenko DM, Lucas S, et al.Interleukin 27 limits autoimmune encephalomyelitis by suppressingthe development of interleukin 17-producing T cells. Nat Immunol2006;7:929–36.
45. Cocco C, Giuliani N, Di Carlo E, Ognio E, Storti P, Abeltino M, et al.Interleukin-27 acts as multifunctional antitumor agent in multiplemyeloma. Clin Cancer Res 2010;16:4188–97.
46. Zorzoli A, Di Carlo E, Cocco C, Ognio E, Ribatti D, Ferretti E, et al.Interleukin-27 inhibits the growth of pediatric acute myeloid leu-kemia in NOD/SCID/Il2rg�/� mice. Clin Cancer Res 2012;18:1630–40.
47. Canale S, Cocco C, Frasson C, Seganfreddo E, Di Carlo E, OgnioE, et al. Interleukin-27 inhibits pediatric B-acute lymphoblasticleukemia cell spreading in a preclinical model. Leukemia 2011;25:1815–24.
48. Wojno ED, Hosken N, Stumhofer JS, O'Hara AC, Mauldin E, Fang Q,et al. A role for IL-27 in limiting T regulatory cell populations. J Immunol2011;187:266–73.
49. AwasthiA,Carrier Y, Peron JP,Bettelli E, KamanakaM, Flavell RA, et al.A dominant function for interleukin 27 in generating interleukin 10-producing anti-inflammatory T cells. Nat Immunol 2007;8:1380–9.
50. Vidal-Vanaclocha F, Mendoza L, Telleria N, Salado C, Valc�arcel M,Gallot N, et al. Clinical and experimental approaches to the patho-physiology of interleukin-18 in cancer progression. Cancer MetastasisRev 2006;25:417–34.
Carbotti et al.
Clin Cancer Res; 19(17) September 1, 2013 Clinical Cancer Research4620
on May 16, 2020. © 2013 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 19, 2013; DOI: 10.1158/1078-0432.CCR-13-0568

2013;19:4611-4620. Published OnlineFirst July 19, 2013.Clin Cancer Res Grazia Carbotti, Gaia Barisione, Anna Maria Orengo, et al. Human Ovarian Cancer Microenvironment
Binding Protein Is Produced in the−The IL-18 Antagonist IL-18
Updated version
10.1158/1078-0432.CCR-13-0568doi:
Access the most recent version of this article at:
Material
Supplementary
http://clincancerres.aacrjournals.org/content/suppl/2013/07/23/1078-0432.CCR-13-0568.DC1
Access the most recent supplemental material at:
Cited articles
http://clincancerres.aacrjournals.org/content/19/17/4611.full#ref-list-1
This article cites 50 articles, 15 of which you can access for free at:
Citing articles
http://clincancerres.aacrjournals.org/content/19/17/4611.full#related-urls
This article has been cited by 1 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://clincancerres.aacrjournals.org/content/19/17/4611To request permission to re-use all or part of this article, use this link
on May 16, 2020. © 2013 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 19, 2013; DOI: 10.1158/1078-0432.CCR-13-0568





























