Embed Size (px)
Citation preview

THE JOURNAL OF BIOLOQICAL C~~EMWCR~ Vol. 237, No. 12, December 1962
Printed in U.S.A.
The Hydrolysis of Long-chain Fatty Acid Esters of Cholesterol with Rat Liver Enzymes*
DANIEL DEYKINt AND DEWITT S. GOODMANI
From the Laboratory of Cellular Physiology and Metabolism, National Heart Institute, National Institutes of Health, Bethesda 14, Maryland
(Received for publication, May 7, 1962)
The liver plays a central role in the metabolism of cholesterol absorbed from the gastrointestinal tract. Most of this cho- lesterol is transported as esterified cholesterol in chylomicrons (2, 3)) by way of intestinal lymph. Recent experiments in this lab- oratory have demonstrated that soon after the intravenous in- jection into rats of chylomicrons containing labeled cholesterol esters, almost 90% of the labeled cholesterol was found in the liver, as esterified cholesterol (4). Slow hydrolysis of the esteri- fied cholesterol then occurred within the liver, followed by grad- ual redistribution of the labeled cholesterol to other tissues.
These findings indicate that liver hydrolysis of chylomicron cholesterol ester is an important component of the metabolism of cholesterol. In addition, the liver participates in the metabo- lism of esterified cholesterol in other ways. Evidence exists that the liver may be the primary source of plasma cholesterol esters (5), and that the individual fatty acid esters of cholesterol are differently metabolized within the liver (6).
Very little information exists concerning the hydrolysis of long- chain fatty acid esters of cholesterol by liver enzymes. Nieft and Deuel (7) demonstrated net hydrolysis of cholesteryl palmi- tate, with the use of a saline extract of rat liver homogenates and a 21..hour incubation. Subsequent investigators, however, have employed short-chain esters, particularly cholesteryl acetate, to study liver cholesterol esterase activity (8,9). It is shown below that this is not a satisfactory substrate for the study of the hy- drolysis of long-chain fatty acid esters of cholesterol.
The present study reports the hydrolysis of long-chain fatty acid esters of cholesterol with enzymes present in the microsomal and soluble fractions of rat liver.
EXPERIMENTAL PROCEDURE
Enzyme Preparations-Fed male Sprague-Dawley rats, weigh- ing 175 to 225 g, were killed by decapitation. The livers were excised and immediately rinsed in ice-cold 0.1 M potassium phos- phate buffer, pH 7.4. The livers were passed through a tissue press, and the pulp was homogenized at ice temperature in a Potter-Elvehjem homogenizer with a loose fitting Teflon pestle, with 2.5 ml of the above buffer per g of liver pulp. After removal of nuclei and cell debris by centrifugation at 2,000 X g for 30 minutes, the supernatant fraction (S-2) was centrifuged at lO,-
* Presented in part at the 1962 Annual Meeting of the Federa- tion of American Societies for Experimental Biology (1).
t Present address: Department of Medicine, Beth Israel Hos- pital and Harvard Medical School, Boston, Massachusetts.
$ Present address: Department of Medicine, Columbia Uni- versity College of Physicians and Surgeons, New York 32, N. Y.
000 X g for 30 minutes to sediment the mitochondria. Micro- somes were sedimented from the 10,000 X g supernatant fraction (S-10) by centrifugation at 104,000 X g for 1 hour. The soluble supernatant fraction (S-104) was then removed, and the micro- somes were washed by suspension in a large volume of buffer followed by centrifugation as before. In early experiments, no efforts were made to remove the layer of free-floating fat from the S-104. In later experiments this layer was removed and discarded, and the remaining supernatant called the “defatted” S-104. The microsomes were finally suspended in a volume of buffer equal to one-tenth the volume of the S-10 from which they had been separated. All centrifugations, and subsequent manip- ulations, were done at 9-5”. The S-104 was fractionated by the addition of solid ammonium sulfate to 30% saturation at 0”. The precipitate that formed was collected by centrifugation at 10,000 x g for 30 minutes, and dissolved in a small volume of water. The dissolved protein was passed over a 2 X 25-cm column of coarse Sephadex G-25 (Pharmacia, Uppsala) to remove the ammonium sulfate. This protein fraction was labeled F030. The Sephadex column was calibrated with solutions of serum al- bumin and ammonium sulfate; the elution of protein was followed by the biuret reaction (lo), and the elution of NHa by reaction with Nessler’s reagent. Elutions were conducted so that 90 to 95% of the protein was collected virtually free of NHd+. After elution, the F,+o was diluted to 5 mg of protein per ml with 0.01 M
phosphate buffer, pH 7.4, and to this was added calcium phos- phate gel, 28 mg per 100 mg of protein. The calcium phosphate gel was kindly provided by Dr. Edward Korn, and was prepared by the method of Dixon and Webb (11). The suspension was agitated at ice temperature for 1 minute, and allowed to stand at least 15 minutes. The gel was separated by centrifugation and discarded. The supernatant fluid was acidified to pH 6.5 with 0.1 N acetic acid, and 100 mg of gel per 100 mg of protein added. After centrifugation, the supernatant (Fx, 3) was separated, and serial elutions (Fx. 4 and Fx. 5) were made from the gel with 1 ml per 10 mg of gel of 0.1 and 0.5 M phosphate buffer, pH 7.4, respectively. Fraction 5 (Fx. 5) contained the partially purified cholesterol esterase activity, and was diluted to 0.1 M with re- spect to phosphate before use. The other fractions obtained during the gel purification displayed only slight hydrolytic ac- tivity for cholesteryl palmitate.
Incubations were carried out in 25-ml Erlenmeyer flasks, at 37”, with shaking, in a metabolic incubator. Air was used as gas phase and the final volume was usually 2.0 ml. No difference was observed when nitrogen was used as gas phase instead of air.
Extraction and Assay of Xterok-Reactions were terminated by
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from

3650 Hydrolysis of Cholesterol Esters Vol. 237, No. 12
transferring the 2-ml incubation-mixtures to CHC13-MeOH (2:1, volume per volume) and washing with CH&-MeOH to a final volume of 50 ml. Each mixture was separated into two phases by the addition of 10 ml of water, and an aliquot of 30 ml was withdrawn from the lower CHCl, phase, total volume 36 ml. Carrier cholesterol and cholesteryl palmitate, 1 mg of each, were added, and the samples evaporated to dryness with a stream of nitrogen.
The free and esterified cholesterol in each sample were sep- arated by chromatography on alumina (Woelm, neutral, grade II activity). Small (lo-mm internal diameter) columns containing 2 g of alumina were charged with the sample dissolved in 1 to 2 ml of light petroleum ether (Fisher reagent grade, b.p. 3656”), and the cholesterol esters eluted with 15 ml of benzene-petroleum ether, 3:7 (volume per volume). Free sterols were then eluted with 10 ml of acetone-ethyl ether, 1: 1 (volume per volume). In later experiments the sterol esters were eluted with 12 ml of benzene-petroleum ether, 1: 1. Various tests showed that this simple procedure separated free and esterified cholesterol with a very high degree of efficiency. Analysis of an incubation mixture containing HQholesteryl pahnitate but no enzyme resulted in the recovery of more than 98% of the H3 in the benzene-petro- leum ether fraction. This radioactivity was not directly precipi- table with digitonin, but became digitonin precipitable after saponification. Furthermore, the small amount of radioactivity (1 to 2%) found in the acetone-ether fraction was also not digi- tonin precipitable until after saponification. Similar results were obtained with heat-inactivated enzyme preparations. These experiments showed that only 1 to 2% of the esterified choles- terol appeared in the acetone-ether eluent, and that nonenzy- matic hydrolysis did not occur during incubation, extraction, and chromatography.
In another experiment, 25y0 of the H3-cholesteryl palmitate was hydrolyzed enzymatically, and the incubation mixture ex- tracted and assayed as described. Only 0.4% of the H3 in the benzene-petroleum ether fraction was precipitable with digitonin in the presence of added unlabeled free cholesterol; the remaining 99.6% of this H3 was digitonin precipitable after saponification. All the H3 found in the acetone-ether eluent was directly precipit- able with digitonin, except for the expected small chromato- graphic “leak” of sterol ester described above. This experiment demonstrated the almost quantitative separation of free and esterified cholesterol achieved during chromatography, and also served to identify the reaction product as free cholesterol, assayed as radioactivity recovered in the acetone-ether eluent. The re- sults given below have all been corrected for the small chromato- graphic contamination of ester cholesterol in the acetone-ether fraction, measured in each experiment by inclusion of a control flask without enzymes.
When esters of labeled fatty acids were used, free fatty acids and fatty acid esters (triglycerides, ethyl palmitate) were sep- arated by the solvent partition method of Borgstriim (12), after extraction of the reaction mixture with the solvent mixture of Dole (13). Control experiments indicated the efficiency of sep- aration to be greater than 99%.
Assay of Radioactivity-All samples were dissolved in 15 ml of 0.5% diphenyloxazole in toluene, and assayed with a Packard liquid scintillation spectrometer. The efficiency for Cl4 was 50 to 55%, and for H3 was 20 to 25%. No quenching was observed.
Other Esterase Activities-The hydrolysis of p-nitrophenyl ace- tate and of p-nitrophenyl phosphate was assayed spectrophoto-
metrically at 400 rnl.c (14). p-Nitrophenyl acetate (m.p. 78.5 to SO”) was synthesized by reacting p-nitrophenol and acetic an- hydride in anhydrous pyridine, and crystallizing the product from ethanol; p-nitrophenyl phosphate was purchased from the Sigma Chemical Company.
Substrate Preparations---(u) Nonradioactive cholesterol esters. Cholesterol (Nutritional Biochemical Corporation) was purified through its dibromide as described by Fieser (15)) followed by two crystallizations from 95% ethanol. Its melting point after puri- fication was 147.7 to 148.9”. Linoleic, oleic, palmitic, and stearic acids were obtained from the Hormel Foundation; all acids were more than 99% pure by gas-liquid chromatography. Cholesteryl linoleate, oleate, palmitate, and stearate were pre- pared by reacting the appropriate acyl chloride with cholesterol in pyridine solution, with a modification of the method of Swell and Treadwell (16). Purified linoleyl and oleyl chlorides were prepared from the corresponding fatty acids by reaction with oxalyl chloride, followed by distillation of the acyl chloride in a vacuum as described by Wood et ~2. (17). Palmitoyl and stearoyl chlorides were prepared by reaction of the fatty acid with thionyl chloride, and were not purified before reaction with cholesterol (16). Cholesteryl palmitate and stearate were purified from the reaction mixtures by two crystallizations from acetone, followed by chromatography on a large column of alumina. Cholesteryl oleate was crystallized once from acetone, followed by chroma- tography on alumina. Cholesteryl linoleate did not crystallize from acetone, but instead came out of solution as an impure oil, which was purified by chromatography on alumina. Approxi- mately 5 g of each ester were synthesized. The column chro- matographies used for purification employed approximately 100 g of alumina (Woelm, grade II). Elution of the product cho- lesterol ester wasachieved in one step, with 75 ml of benzene-light petroleum ether 3:7 (volume per volume) per 10 g of alumina.
The melting points of the cholesterol esters were: linoleate, 41.443.4”; oleate, 49.8-51.2”; palmitate, 76-77.5”; stearate, 81-83”. Gas liquid chromatography of the fatty acid com- ponents indicated that the esters were each more than 97 y0 pure. Infrared spectra obtained with a Perkin-Elmer Model B 21 spec- trophotometer showed no evidence for the presence of trans dou- ble bonds in either cholesteryl oleate or linoleate. Elaidic acid was used as the standard compound for tram double bonds (18), and it was estimated that the presence of 2% tram bonds could have been detected in the range studied. There was complete loss of the absorption peak at 3300 cm-r characteristic of the -OH group, and present in the free sterol. Study of the ultra- violet absorption of cholesteryl linoleate at 220 rnp indicated the presence of approximately 2% of the double bonds as con- jugated diene.
Cholesteryl acetate (m.p. 111.5-114”) and butyrate were pur- chased from Steraloids, Inc. and were chromatographed on alumina before use. The cholesteryl butyrate was found to be impure and suitable only for semi-quantitative work.
(b) Radioactive Cholesterol Esters-7-o-H3-Cholesteryl palmi- tate and stearate (specific activities 48 and 50 mc per mole, respec- tively) were purchased from New England Nuclear Corporation. Both esters were chromatographed on small alumina columns, to remove possible traces of contaminating free sterol. Gas-liquid chromatography of the fatty acid components of a few micro- grams of each revealed the presence of only the expected fatty acid peak, at a sensitivity level where a 2% contamination by another component should have been detected.
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from

December 1962 D. Deykin and D. S. Goodman 3651
H3-Cholesteryl oleate and linoleate were prepared by a micro- modification of the synthesis employed above on a macro scale. To 1 mc of 7-a-H3-cholesterol (New England Nuclear Corpora- tion, 1 mc per 0.48 mg) in a centrifuge tube were added 5.00 mg of carrier cholesterol, 20 ~1 of the appropriate purified acyl chloride, and 0.3 ml of dry pyridine (distilled from BaO). The mixtures were flushed with nitrogen and heated at 50-60” for 3 hours. The reaction mixtures were transferred to cold water, and the products extracted into petroleum ether. The petroleum ether solutions were washed with 0.5 N HCl, with 0.1 N NaOH in 50% ethanol (to extract free fatty acids), and with water. After evaporation of the petroleum ether, the residues were purified by chromatography on 5-g alumina columns. The yields of Ha- cholesteryl linoleate and oleate were 77% and S2%, respectively. Gas-liquid chromatography of the fatty acids from identical micro-syntheses with unlabeled cholesterol revealed the expected fatty acid peak in each case, with unidentified minor peaks com- posing 5% or less of the product.
H3-Cholesteryl acetate and butyrate were also synthesized on a micro scale by reacting 7-a-Ha-cholesterol with reagent grade acetic anhydride and butyryl chloride, respectively, in pyridine solution. The reaction mixtures were transferred to small sep- aratory funnels containing cold water and extracted into petro- leum ether. The petroleum ether was washed with 0.1 N HCI, 0.1 N KHC03, and water, and the products then purified by chromatography on 5-g silicic acid columns, as described by Horning, Williams, and Horning (19).
Other Lipid Substrates. Tripalmitin (m.p. 64.5-65.5”) was a Fisher reagent chemical; it was further purified by silicic acid chromatography (19), with collection and use of the material eluted with a mixture of 30 ml of benzene and 20 ml of hexane. 04..Carboxyl-labeled triplamitin was purchased from Research Specialties Company. It had a specific radioactivity of 7.5 mc per mmole, and was also purified by silicic acid chromatography before use.
Ethyl palmitate was prepared by heating palmitic acid, in ab- solute ethanol containing 0.1 y0 benzene and 2 y0 concentrated HzS04, at 60” overnight. The product was extracted with light petroleum ether after the addition of an equal volume of water, and free fatty acids removed from the petroleum ether solution by washing with 0.05 N KOH in 50% ethanol. Ethyl palmitate- l-Cl4 was similarly synthesized from 1-U4-palmitic acid (Nu- clear-Chicago, 24 mc per mmole). The yields of Cl4 and of un- labeled ethyl palmitate were both more than 95%.
RESULTS
A. Soluble Enzyme
Presentation of Substrate-Initial experiments were conducted to compare several modes of presentation of the water-insoluble lipid substrates to be tested for hydrolysis. Cholesteryl palmi- tate was added to a whole homogenate in three ways: (a) in ace- tone solution, (b) suspended in saline solutions of sodium glyco- cholate or of Tween 20, or (c) suspended in a mixture containing ethanol, acetone, and serum albumin. The data in Table I indi- cate that the substrate was most effectively hydrolyzed when added in acetone solution. Experiments in which identical quan- tities of substrate were dissolved in varying amounts of acetone indicated hydrolysis to be maximum with substrate added in 50 ~1 (final concentration of acetone, 2.5%). Addition of Tween 20, sodium glycocholate or taurocholate, human serum albumin,
TABLE I Effect of mode of presentation of substrate on hydrolysis
of cholesteryl palmitate Each a-hour incubation contained 2 ml of S-2 fraction and 1.25
mpmole (0.011 pc) of HQholesteryl palmitate. The substrate was added as follows. Flask l? dissolved in 50 ~1 acetone; Flask 2, suspended in 50~150/, Tween 20 in 0.15 M N&l; Flask 3, suspended in 100 ~1 of 15 mM sodium glycocholate in 0.15 M NaCl; Flask 4, suspended in 100 ~1 of a solution of 5yo human serum albumin-5% ethanol-5% acetone in 0.15 M NaCl.
Flask Substrate presentation % Hydrolysis
1 Acetone 15.7 2 Tween 20 7.1 3 Sodium glycocholate 2.6 4 Albumin-ethanol-acetone 1.1
TABLE II Subcellular distribution of cholesterol esterase activity
Each 2-ml incubation contained 200 pmoles of potassium phos- phate buffer, pH 7.4, and amounts of enzyme fractions represent- ing equivalent volumes of the whole homogenate. To each mix- ture, 1.0 mpmole (0.046~~) of H3-cholesteryl palmitate was added, and the incubations conducted for 1 hour.
Enzyme fraction
s-2. s-10. S-104 (defatted). Mitochondria . . Microsomes...........
Recovered radioactivity*
I Ester FPX
c.p.m.
17,169 2,620 18,193 2,682 14,126 7,146 18,898 971 20,317 1,135
% Hydrolysis
11.6 11.2 32.0 3.3 3.6
* The c.p.m. listed are the actual values observed whereas the $& hydrolysis in the last column has been corrected for chroma- tographic contamination.
human @lipoprotein, or rat liver polyglycerolphosphatide to in- cubations before the addition of substrate (in 50 ~1 acetone) all inhibited hydrolysis. In the experiments that follow, the sub- strate was always presented in 50 ~1 of acetone solution.
The subcellular distribution of hydrolytic activity is described in Table II. The bulk of activity was associated with the soluble supernatant fraction, although activity was detected in both particulate fractions.
The data in Table III show the amounts of free and esterified cholesterol in the subcellular fractions. Since the S-2, S-10, and particulate fractions contained considerable endogenous cho- lesterol ester, but the defatted S-104 contained none, the specific activity of the substrate being hydrolyzed varied in each fraction to the extent that the endogenous cholesterol ester in the enzyme preparation itself was available for hydrolysis. It is probable, therefore, that the lack of dilution of the substrate in the defatted S-104 fraction was responsible for the greater per cent hydrolysis of the added substrate by the S-104 fraction compared to the S-10 and S-2 fractions.
Purijication of Soluble Enzyme-The hydrolytic activity of the soluble enzyme was almost completely precipitable with am- monium sulfate, at 30% saturation. Subsequent purification
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from
3652 Hydrolysis of Cholesterol Esters Vol. 237, No. 12
TABLE III
Cholesterol content of rat liver homogenate fractions Analyses of fractions prepared as in “Experimental Procedure.”
Lower limit determination ~0.005 mg per ml f0.01.
cholesterol
Fraction
Total I I
FIW Ester
Whole homogenate. 1.02 s-2. . . 0.36 s-10. . 0.24 s-104. . 0.21 S-104 (defatted). . 0.03 Fo30. . 0.01 Fx.~..................... 0 Microsomes (cont. X 10) 1.15 Mitochondria (corm. X 10) 0.76
+%/ml 0.69 0.23 0.16 0.12 0.03 0.01 0 1.02 0.66
0.33 0.13 0.08 0.09 0 0 0 0.13 0.10
TABLE IV
PuriJication of rat liver soluble cholesterol esterase
The l-hour incubations contained amounts of protein approxi- mat,ely prorated to represent equivalent amounts of the whole homogenate.
s-2. s-10. S-104 (defatted). Fo? Fx. 5.
w/ml 40.0 7,600 19.3 36.5 6,132 30.1 27.0 3,699 160.0
5.0 820 338.0 0.3 51 1,321.0
I
* Specific enzyme activity = y0 hydrolysis X 5000 c.p.m. per mg of protein, assuming linearity between amount of protein actually incubated and extent of hydrolysis.
t Total eneyme activity = specific enzyme activity times total protein.
ml
190 168 137 164 171
m !
146,680 184,573 591,840 277,160
67,371
was hindered by rapid loss of activity upon dialysis or lyophiliza- tion. Activity could not be recovered from DEAE-cellulose col- umns by elution with buffers ranging from 0.005 M, pH 7.9, to 1.25 M, pH 6.7. Further purification was effected with the use of Sephadex columns to remove the ammonium sulfate, and sub- sequent calcium phosphate gel adsorption of the Sephadex eluate.
The results of a typical purification experiment are summarized in Table IV. The most purified fraction (Fx. 5) after calcium phosphate gel elution usually displayed an 8 to lo-fold purifica- tion when compared to the S-104. The apparent purification was always very much greater when compared to the S-2 or S-10 fractions (see column headed “Specific enzyme activity” in Table IV). Because of the presence of endogenous esterified cholesterol, estimates of the specific enzyme activity of the initial fractions were in error to the extent that this endogenous material in the enzyme preparation was available as substrate for hydroly- sis. This error is apparent in the paradoxically low total enzyme activity of the S-2 and S-10 fractions, compared to the S-104 (see Table IV), and undoubtedly accounts for the much greater apparent purification of Fx. 5 when compared to the S-2 or S-10 fractions, rather than the S-104 fraction.
o ACE774 JE BUFFER 30 0 K-PO, BUFFER
PH
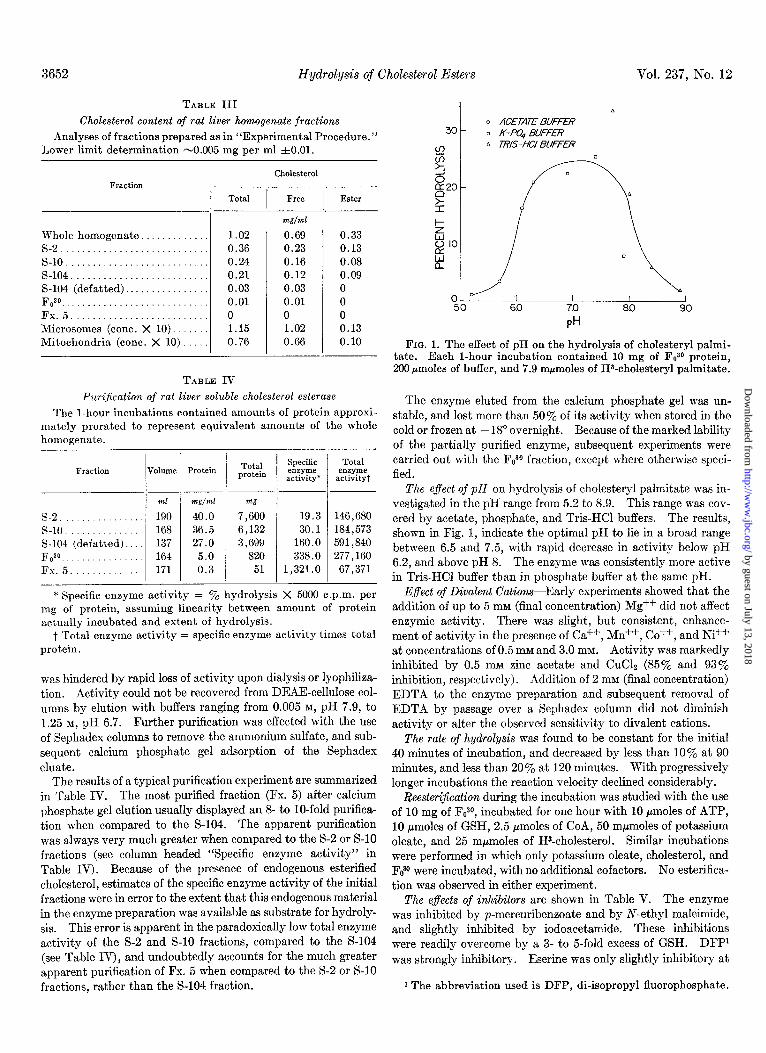
FIG. 1. The effect of pH on the hydrolysis of cholesteryl palmi- tate. Each l-hour incubation contained 10 mg of Faso protein, 200 pmoles of buffer, and 7.9 mpmoles of HQholesteryl palmitate.
The enzyme eluted from the calcium phosphate gel was un- stable, and lost more than 50% of its activity when stored in the cold or frozen at - 18” overnight. Because of the marked lability of the partially purified enzyme, subsequent experiments were carried out with the Fo30 fraction, except where otherwise speci- fied.
The e$ect of pH on hydrolysis of cholesteryl palmitate was in- vestigated in the pH range from 5.2 to 8.9. This range was cov- ered by acetate, phosphate, and Tris-HCl buffers. The results, shown in Fig. 1, indicate the optimal pH to lie in a broad range between 6.5 and 7.5, with rapid decrease in activity below pH 6.2, and above pH 8. The enzyme was consistently more active in Tris-HCl buffer than in phosphate buffer at the same pH.
E$ect of Divalent Cations-Early experiments showed that the addition of up to 5 mM (final concentration) Mg++ did not affect enzymic activity. There was slight, but consistent, enhance- ment of activity in the presence of Ca++, Mn++, Co++, and Ni++ at concentrations of 0.5 mM and 3.0 mM. Activity was markedly inhibited by 0.5 mM zinc acetate and CuClz (85% and 93% inhibition, respectively). Addition of 2 mM (final concentration) EDTA to the enzyme preparation and subsequent removal of EDTA by passage over a Sephadex column did not diminish activity or alter the observed sensitivity to divalent cations.
The rate of hydrolysis was found to be constant for the initial 40 minutes of incubation, and decreased by less than 10% at 90 minutes, and less than 20% at 120 minutes. With progressively longer incubations the reaction velocity declined considerably.
ReesterQkation during the incubation was studied with the use of 10 mg of Fo30, incubated for one hour with 10 pmoles of ATP, 10 pmoles of GSH, 2.5 pmoles of CoA, 50 mpmoles of potassium oleate, and 25 mpmoles of H3-cholesterol. Similar incubations were performed in which only potassium oleate, cholesterol, and Foa were incubated, with no additional cofactors. No esterifica- tion was observed in either experiment.
The effects of inhibitors are shown in Table V. The enzyme was inhibited by p-mercuribenzoate and by N-ethyl maleimide, and slightly inhibited by iodoacetamide. These inhibitions were readily overcome by a 3- to 5-fold excess of GSH. DFPl was strongly inhibitory. Eserine was only slightly inhibitory at
1 The abbreviation used is DFP, di-isopropyl fluorophosphate.
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from

December 1962 D. Deykin and D. S. Goodman 3653
low4 M, a concentration that completely inhibits cholinesterase activity.
E$ect of Enzyme Concentration-The initial rate of hydrolysis of cholesteryl palmitate was linearly related to enzyme concen- tration at concentrations of FP below 4 mg of protein per ml. The rate of hydrolysis was, however, strikingly nonlinear at concentrations of Fo30 in excess of 5 mg of protein per ml. Since the Fo30 fraction contained negligible amounts of cholesterol ester, this could not result from the addition of endogenous sub- strate together with the enzyme fraction. In the various experi- ments presented here, the concentrations of protein employed were in the linear range of the enzyme concentration-activity curve.
The substrate speci$city of the soluble enzyme was studied by comparing the hydrolytic activity of each fraction during the purification procedure with each of three different esters. p-Ni- trophenyl acetate, an ester known to be hydrolyzed by a wide range of esterases, was employed as one model substrate. p-Ni- trophenyl phosphate was used as a measure of phosphatase activity, and cholest,eryl palmitate was used to measure choles- terol esterase activity. The results shown in Fig. 2 demonstrate that a selective purification of the cholesterol esterase activity was achieved with Fx. 5. The microsomal fraction showed striking hydrolytic activity for p-nitrophenyl acetate, whereas Fx. 3 displayed the most phosphatase activity. Fx. 3 also represented the greatest concentration of soluble activity toward p-nitrophenyl acetate. These results suggest that the soluble
TABLE V
Inhibition of cholesteryl palmitate hydrolysis with soluble enzyme
The l-hour incubations in Experiment A each contained 9 mg of Fo30 protein, 200 pmole of potassium phosphate buffer, pH 7.4, and 20 mpmoles (0.020 pc) of substrate. In Experiment B, each one-hour incubation contained the same buffer, Fx. 5,0.75 mg of protein and cholesteryl palmitate, 1.0 mpmole.
Experi- ment
A
B
Additions
Recovered radioactivity
Inhibition
fimoles
None N-ethyl maleimide, 1 N-ethyl maleimide, 6 N-ethyl maleimide 6 + GSH,
20 Iodoacetamide, I Iodoacetamide, 6 p-Mercuribenzoate, 0.8* p-Mercuribenzoate, 2 p-Mercuribenzoate, 2 + GSH,
20 Eserine sulfate, 0.2 Eserine sulfate, 1
c.p.m. 6,897 2,096 7,689 763 7,966 247 6,852 2,461
% 0
68 95
0
6,441 1,913 2 7,342 1,429 33 8,692 324 95 8,672 281 95 6,208 2,503 0
7,217 1,519 28 7,881 798 65
_-
None 5,036 772 0 DFP, 0.01 4,738 671 8
DFP, 0.10 4,943 268 81
DFP, 0.50 5,811 259 89
-
* This experiment was done on a different day and is referred to a different control.
r 2 .80 G a 0 60. -. !k 8 $j 40 w 2 k .20-
O- MICROSOMES S-IO S-104 Fo30 Fx 5
FIG. 2. Relative esterase activities of liver homogenate frac- tions. Cholesteryl palmitate hydrolysis was studied with a-hour incubations containing 1.25 mpmoles of H3-cholesteryl pahnitate. p-Nitrophenyl acetate and phosphate hydrolyses were both meas- ured at pH 7.4. Five rmoles of p-nitrophenyl acetate were added in 0.25 ml of isopropanol to incubations of final volume 5 ml, and the hydrolysis rate measured and corrected for spontaneous hy- drolysis. Two-milliliter incubations containing 2 mg of p-nitro- phenyl phosphate were analyzed for hsdrolssis after 30 minutes. The amounts of protein in the different fractions were approxi- mately nrorated as in Tables II and III. The S-104 was a de- fatted preparation. The data are plotted to indicate the relative activity of each fraction against each ester. The most active fraction for each ester was arbitrarily assigned an activity of 1 .OO
IOr-
mpMOLES INCUBATED
FIG. 3. The hydrolysis of 5 cholesterol esters at varying sub- strate concentrations, with the soluble enzyme. The 30-minute incubations contained 9 mg of FoaO protein, 200 rmoles of potas- sium phosphate buffer, pH 7.4, and varying amounts of substrates. L = cholesteryl linoleate; 0 = cholesteryl oleate; A = cholesteryl acetate; P = cholesteryl palmitate; S = cholesteryl stearate.
cholesterol esterase is a different enzyme from those involved in the hydrolysis of the other esters studied.
The fatty acid speci&ity of the soluble cholesterol esterase was tested in two ways. In one set of experiments, the different cholesterol esters were individually studied at several substrate levels, and substrate-concentration curves were determined. The results, shown in Fig. 3, demonstrate that considerably more hydrolysis was observed with cholesteryl oleate and linoleate than with cholesteryl palmitate. There was very little hydroIysis of cholesteryl stearate. The hydrolysis of cholesteryl acetate was
intermediate between that of oleate and palm&ate; in a single
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from

3654 Hydrolysis of Cholesterol Esters Vol. 237, No. 12
0 s-104 Fo
30 Fx 5
FIG. 4. Hydrolysis of equimolar mixtures of cholesterol esters. Each l-hour incubation contained 5 mpmoles each of unlabeled cholesteryl palmitate, stearate, oleate, and linoleate. In each mixture, 0.3 to 0.5 mpmole of one of the H*-cholesteryl esters was included as follows: Ha-cholesteryl-palmitate = P; -stearate = S; -oleate = 0; -1inoleate = L. Incubations contained the following amounts of protein: S-104 (defatted), 10 mg; F#O, 10 mg; Fx. 5, 0.5 mg. *sample lost.
I I .- 60 7.0 8.0 90
PH
FIG. 5. The effect of pH on the hydrolysis of cholesteryl palmi- tate with microsomes. Each l-hour incubation contained, in 2 ml, 15 mg of microsomal protein, 200 pmoles of Tris-malate buffer, and 0.9 mpmole, Ha-cholesteryl palmitate.
incubation cholesteryl butyrate was hydrolyzed to approximately the same extent as cholesteryl acetate.
A second study was conducted to measure the hydrolysis of each ester in an equimolar mixture of cholesteryl palm&ate, stearate, oleate, and linoleate. To achieve this, four identical acetone solutions were prepared containing equal amounts of the four esters, but with radioactivity in only one of the esters. Each of the four solutions contained radioactivity in a different ester. The results of this study, shown in Fig. 4, also demon- strated a distinctly greater hydrolysis of cholesteryl oleate and linoleate. In contrast to the preceding results, in these experi- ments cholesteryl stearate was approximately as effective a sub- strate as cholesteryl palmitate. Fig. 4 also indicates that the pattern of substrate preference did not change during the purifi- cation of enzyme from S-104 to Fx. 5.
Other Lipid Substrates-The soluble enzyme was extremely active in effecting the hydrolysis of ethyl palmitate. A constant rate was observed, and more than 90 y0 hydrolysis achieved in one hour, with substrate concentrations up to 100 mpmoles in 2 ml (the largest amount tested). Tripalmitin alone, or added with corn oil in an attempt to give a better dispersion, was not effectively hydrolyzed (5.3% hydrolysis of 10 or 20 mpmoles in one hour).
B. Microsomes
Studies were also conducted on the properties of the cholesterol esterase activity of washed microsomes.
Substrate presentation in the microsomal system was studied with the use of cholesteryl palmitate dissolved in acetone, and suspended in Tween 20, sodium taurocholate, and in ethanol- acetone-serum albumin. Optimal hydrolysis was again found with the substrate in acetone solution at a final concentration of acetone of 2.5%. Addition of Tween 20, sodium taurocholate, or human serum albumin to the incubations before the addition of substrate in acetone solution were all inhibitory. There was no requirement for GSH.
The e$ect of pH on the microsomal esterase was studied over the range 5.4 to 9.0. The results showed the optimal pH range to be 6.0 to 6.5, with an abrupt decline in activity below pH 5.8 and a more gradual decline above pH 6.5. As with the soluble enzyme, activity was greater in Tris-KC1 buffer than in phosphate
TABLE VI Inhibition of cholesteryl palmitate hydrolysis with microsomes Each l-hour incubation contained, in 2 ml, 15 mg of microsomal
protein, 200 pmoles of potassium phosphate buffer, pH 6.1, 1 mpmole (0.008 PC) of H3-cholesteryl palmitate, and additions as listed.
Additions I
Inhibition
pmoles % N-ethyl maleimide, 1. ...................... 24 N-ethyl maleimide, 6. ...................... 52 N-ethyl maleimide, 6 + GSH, 20 ............ 3 Iodoacetamide, 1. .......................... 74 Iodoacetamide, 6 ........................... 79 p-Mercuribenzoate, 0.4. ..................... 49 p-Mercuribenzoate, 2 ........................ 98 p-Mercuribenzoate, 2 + GSH, 10. ........... 0 Eserine sulfate,0.2 .......................... 94 Eserine sulfate, 1. .......................... 94 DFP, 0.01.. ................................. 84 DFP, O.l..... ............................... 95 DFP, 0.5 .................................... 95
mpMOLES INCUBATED
FIG. 6. Hydrolysis of 5 cholesterol esters at varying substrate concentrations, with microsomes. The incubations were similar to those of Table VI, with substrates similar to those of Fig. 3.
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from

December 1962 D. Deykin and D. S. Goodman 3655
buffer at the same pH. Experiments with Tris-malate buffer to cover the entire pH range (Fig. 5) clearly demonstrated that the pH optimum was in the range 5.8 to 6.5.
The e.fects of enzyme and of substrate concentration were studied by measuring the hydrolysis of each of 4 different, amounts of substrate (2, 6, 20, and 50 mpmoles), with each of 5 different amounts of microsomal protein (3.5, 6.9, 21, 35, and 69 mg of protein). The results indicated that, in the range of substrates tested, the relationship between enzyme concentration and rate of hydrolysis was never linear.
The effects of inhibitors are shown in Table VI. As with the soluble enzyme, N-ethyl maleimide, iodoacetamide, and p-mer- curibenzoate were all inhibitory; these inhibitions were readily overcome by addition of excess GSH. The microsomal enzyme was much more sensitive to DFP and to eserine than was the soluble enzyme.
The efects of divalent cations (Mg++, Mn++, Ca++, Co++, Ni++, Zn++, and Cu++) were studied at 0.5 mM concentration. Copper and zinc ions were markedly inhibitory, but the other cations tested did not affect hydrolysis.
Fatty acid specificity of the microsomal cholesterol e&erase was investigated by the same techniques used to study the soluble enzyme. Substrate concentration curves for each of 5 different sterol esters are shown in Fig. 6. The rate of hydrolysis of cholesteryl acetate was much greater than that of any of the long-chain fatty acid esters, and was linear throughout the con- centration range tested. This range extended to 50 mpmoles of substrate, although Fig. 6 only shows the results for amounts up to 20 mpmoles. The hydrolysis of cholesteryl butyrate (not shown) was less than that of acetate, but considerably more than that, of linoleate. Of the long-chain esters, linoleate and oleate again served as better substrates than did cholesteryl palmitate and stearate. In addition, the hydrolysis of each component of equimolar mixtures of cholesteryl palmitate, stearate, oleate, and linoleate showed an order of preference similar to that of the soluble enzyme. These experiments were performed in the same manner as those of Fig. 4, and resulted in a 10.0% hydroly- sis of cholesteryl linoleate, 9.8% of cholesteryl oleate, and 6.8% each of cholesteryl stearate and palm&ate. These results, and the data shown in Figs. 3 and 6, indicate that the soluble and microsomal enzymes showed fairly similar patterns of specificity for the four long-chain esters studied, but strikingly different preferences for the short-chain esters.
Other Lipid Substrates-Ethyl palmitate was almost quanti- tatively (97 to 98%) hydrolyzed in 1 hour in concentrations up to 100 mpmoles in 2 ml. Tripalmitin hydrolysis (15% of 20 mpmoles in 1 hour) slightly exceeded that of cholesteryl oleate and linoleate at similar concentrations.
DISCUSSION
These experiments show that rat liver contains enzymes that hydrolyze the long-chain fatty acid esters of cholesterol, and that the bulk of hydrolytic activity is found in the soluble frac- tion of the liver homogenate. The data from Table II indicate that the S-10 fraction hydrolyzed 11.2% of the added substrate, but that an equivalent sample of microsomes hydrolyzed only 3.6% of the added substrate, or 32$& of the amount of hydrolysis (of added substrate) effected by the S-10. Analytic measure- ments (Table III) showed that the endogenous cholesterol ester present in the microsomes was only 0.013/0.08, or less than 9, the total cholesterol ester present in the S-10. It is thus clear
that the lesser activity of the microsomes alone was not, due to the presence of a relatively large pool of endogenous substrate in the microsomes, and that most of the hydrolytic activity of the S-10 was contributed by the soluble fraction. When compared to an equivalent sample of defatted S-104, the microsomes hy- drolyzed only 11 To as much of the added substrate. Therefore, the contribution of the microsomes lies between 11 and 32% of the observed total hydrolysis.
Studies of enzymic reactions involving lipid substrates rou- tinely employ emulsified or finely suspended preparations of the water-insoluble substrates. With emulsions similar to those employed in this study (substrate added in acetone solution), it has been possible to study the characteristics of the enzymic cyclization of squalene (20) and of several reactions involving sterol precursors of cholesterol (21) .2 In addition, other workers have applied kinetic analyses usually used for reactions occurring in true solution to enzymic reactions with steroid substrates added in propylene glycol (22) or in methanol (23) solution. Sarda and Desnouelle (24) and Desnouelle, Constantin, and Baldy (25) have interpreted the typical Michaelis concentration dependence of some reactions with insoluble substrates as the result of dependence on the surface area of substrate available for reaction. Nevertheless, the water insolubility of lipid sub- strates renders difficult. the interpretation of enzymic reaction rates and substrate specificity studies, and conclusions derived from the present study are subject to the uncertainties arising from these difficulties.
When the data shown in Figs. 3 and 6 were plotted according to the method of Lineweaver and Burk (26), straight lines were obtained for each substrate. This concentration dependence suggests that the cholesterol ester suspensions served as the direct substrates for the hydrolytic enzymes. By extrapolation of these plots in the usual way, approximate substrate concentra- tions for half-maximal velocity, for the soluble enzyme, were determined as follows: 14.3 X 10m6 M for cholesteryl acetate, oleate, and linoleate, and 4.8 X low6 M for cholesteryl palmitate. Extrapolated values for maximal velocities of hydrolysis (ex- pressed as mpmoles substrate hydrolyzed per 10 mgm of F,$O per 30 minutes) were: cholesteryl palmitate, 3.1; cholesteryl acetate, 9.1; and cholesteryl oleate and linoleate, 14.3.
The substrate concentration curves with individual cholesterol esters (Figs. 3 and 6) indicate that both the soluble and the microsomal enzymes hydrolyzed the unsaturated fatty acid esters (cholesteryl oleate and linoleate) more rapidly than the saturated ones. It, is possible, however, that this observed difference resulted from different, physical properties of the sub- strate suspensions of the individual cholesterol esters, rather than from true enzymic specificity. To investigate this possibility, experiments were performed with equimolar mixtures of all four long-chain fatty acid esters as substrate. It was felt that the mixed emulsions so formed would minimize any effects dependent, upon different properties of the individual est.er emulsions. That, this in fact occurred can be seen from the hydrolysis of cholesteryl stearate by the soluble enzyme. When presented alone, stearate was not effectively hydrolyzed, but when dis- persed among the other three esters, it was hydrolyzed as effec- tively as pahnitate. The preference for oleate and lmoleate in the mixtures was similar to that observed with individual esters.
2 J. Avigan, D. S. Goodman, and D. Steinberg, J. Biol. Chem., submitted for publication.
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from

3656 Hydrolysis of Cholesterol Esters Vol. 237, No. 12
These results suggest that the more rapid hydrolysis of cholesteryl oleate and linoleate, compared to cholesteryl palmitate and stearate, reflected true enzymic specificity. It is recognized, however, that this conclusion cannot be established with certainty by these experiments.
The data shown in Fig. 4 indicate that the bulk of liver choles- terol esterase activity, present in the soluble fraction of the liver homogenate, was separable from the liver esterase activities involved in the hydrolysis of p-nitrophenylacetate and -phos- phate. The hydrolysis of p-nitrophenylacetate has been found to reflect the activity of a variety of esterases, including pan- creatic cholesterol esterase (27). This would seem not to be true for the liver soluble cholesterol esterase. The microsomes, in contrast, were associated with striking hydrolysis of p-nitro- phenylacetate.
The microsomal cholesterol esterase differed from the soluble esterase in several respects. The microsomal enzyme was in- hibited by eserine in concentrations similar to those effective against cholinesterase; the soluble enzyme was not inhibited at this concentration. In addition, the microsomal enzyme was strongly inhibited by DFP at concentrations below 10e5 M.
The soluble enzyme was also inhibited by DFP, but only when the concentration of inhibitor was an order of magnitude greater. The microsomal pH optimum near 6.1 was well below the op- timum of the soluble enzyme. There was no enhancement of microsomal activity by divalent cations. Tripalmitin, not effec- tively hydrolyzed by the soluble enzyme, was hydrolyzed to a greater extent than either cholesteryl oleate or linoleate in the microsomal system. Most striking was the marked hydrolysis of cholesteryl acetate and butyrate with microsomes, far exceed- ing their relative hydrolysis by the soluble system. In addition, both enzymes displayed great activity toward ethyl palmitate.
The data shown in Figs. 3 and 6 indicate the extent of hydroly- sis of cholesterol esters with soluble enzyme equivalent to 2 to 3 ml of liver whole homogenate, and with microsomes equivalent to 5 to 6 ml of whole homogenate, respectively. From these data, and from similar experiments (e.g. cf. Table II), it can be estimated that the soluble enzyme hydrolyzes 5 to 10 times more cholesteryl palmitate, stearate, oleate, and linoleate, than does the microsomal fraction. Figs. 3 and 6 also demonstrate that experiments based on the hydrolysis of cholesteryl acetate do not reflect the hydrolysis of physiologically significant esters.
Recent experiments have demonstrated that cholesterol esters in chylomicrons are taken up almost quantitatively from the blood by the liver. In the liver a slow net hydrolysis ensues, which is more than 80 ‘% complete in 33 hours (4). Based on the rate of hydrolysis of cholesteryl oleate by the soluble enzyme observed here (Fig. 3), an estimated maximal rate of hydrolysis for a 7- to 8-g rat liver would be of the order of 600 ml.cmoles of cholesteryl oleate hydrolyzed in one hour. This value is in remarkably close agreement with the net rate of hydrolysis observed in viva (4).
SUMMARY
1. The hydrolysis of long-chain fatty acid esters of cholesterol with rat liver homogenates has been shown. The bulk of hy- drolytic activity was associated with the soluble protein fraction, although the microsomes contributed between 11 and 32% of the observed total hydrolysis.
2. The soluble enzyme was partially purified by ammonium sulfate precipitation and calcium-phosphate gel absorption. The
enzyme was unstable ana lost activity after freezing, dialysis, or lyophilization. The soluble cholesterol esterase was separable from the major esterase activities involved in the hydrolysis of p-nitrophenylacetate and p-nitrophenylphosphate.
3. The soluble enzyme displayed a broad pH optimum between pH 6.5 and 7.5; the microsomal enzyme displayed a sharper peak. near pH 6.1. The soluble enzyme did not hydrolyze tripalmitin as effectively as did the microsomes. The soluble enzyme was not inhibited by lo-’ M eserine or by 10m5 M DFP; the micro- somal enzyme was strikingly inhibited at these concentrations.
4. The order of hydrolysis of cholesterol esters by the soluble enzyme was found to be: Cholesteryl linoleate = oleate > ace- tate > palmitate > stearate. The order of hydrolysis of choles- terol esters by the microsomes was found to be: Cholesteryl ace- tate >> linoleate = oleate > palmitate > stearate.
5. The rate of hydrolysis of cholesterol oleate by the soluble enzyme is in close agreement with the net rate of hydrolysis observed in vivo.
Acknowledgment-The expert technical assistance of Miss
Margaret Dedeian is gratefully acknowledged.
1.
2.
3.
4. 5.
6.
7.
8.
9. 10.
11.
12. 13. 14. 15. 16.
17.
18. 19.
20. 21.
22.
23.
24.
25.
26.
27.
REFERENCES
DEYRIN, D., AND GOODMAN, D. S., Federation Proc., 21, 297 (1962).
SWELL, L., TROUT, E. C., JR., HOPPER, J. R., FIELD, H., JR.,. AND TREADWELL, C. R., J. Biol. Chem., 232, 1 (1958).
DASKALAEIS, E. G., AND CHAIKOFF, I. L., Arch. Biockem. Bio-- phys., 68, 373 (1955).
GOODMAN, D. S., J. Clin. Invest., 41, (1962). FRIEDMAN, M., AND BYERS, S. O., J. Clin. Invest., 34, 1369
(1955). KLEIN, P. D., AND MARTIN, R. A., J. Biol. Chem., 234, 1685
(1959). NIEFT, N. L., AND DEUEL, H. J., JR., J. Biol. Chem., 177, 145
(1949). BYRON, J. E., WOOD, W. A., AND TREADWELL, C. R., J. Biol.
Chem., 206, 483 (1953). SASTRY, P. S., AND GANGULY, J., Biochem. J., 80, 397 (1961). GORNALL, A. G., BARDAWILL, C. S., AND DAVID, M. M., J.
Biol. Chem., 1’77, 751 (1949). DIXON, M., AND WEBB, E. C., Enzymes, Longmans, London,
BJ~:T%~‘B Acta Physiol Stand 26 DOLE, V. P.1 J:‘Clin. Invest., Z&i, 150 (i956\.
111 (1952).
HARTLEY, B. S., AND KILBY, B. A., Biochem. J., 60,672 (1952). FIESER, L. F. Org. Syntheses, 36, 43 (1955). SWELL, L., AND TREADWELL, C. R., J. Biol. Chem., 212, 141
(1955). WOOD, T. R., JACKSON, F. L., BALDWIN, A. R., AND LONGEN-
EOEER. H. E.. J. Am. Chem. Sot.. 66,287 (1944). HARTMA~, L., AND SHORLAND, F. B:, Biochem. J., 69,l (1958). HORNING, M. G., WILLIAMS, E. A., AND HORNING, E. C., J.
Lipid Res., 1, 482 (1960). GOODMAN, D. S., J. Biol. Chem., 236, 2429 (1961). AVIGAN, J., GOODMAN, D. S., AND STEINBERG, D. Federation
Proc., 21, 300 (1962). SHARMA. D. C.. FORCHIELLI, E., AND DORFMAN, R. I., J. Biol.
Chem.; 237, i495 (1962). KAWAHARA, F. S., WANG, S.-F., AND TALALAY, P., J. Biol.
Chem.. 237, 1500 (1962). SARDA, i., AND DE.SNOUELLE, P., Biochim. et Biophys. Acta,
30, 513 (1958). DESNOUELLE, P., CONSTANTIN, M. J., AND BALDY, J., Bull.
Sot. Chim. Biol., 37, 285 (1955). LINEWEAVER, H., AND BURE, D., J. Am. Chem. Sot., 66, 658
(1934). MYERS, D. K., in BOYER, P. D., LARDY, H., AND MYRBHCK
K., The enzymes, 2nd ed., vol. IV, Academic Press, N. Y., 1960, p. 476.
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Daniel Deykin and DeWitt S. GoodmanEnzymes
The Hydrolysis of Long-chain Fatty Acid Esters of Cholesterol with Rat Liver
1962, 237:3649-3656.J. Biol. Chem.
http://www.jbc.org/content/237/12/3649.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/237/12/3649.citation.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from





























