Embed Size (px)
Citation preview

BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions, researchlibraries, and research funders in the common goal of maximizing access to critical research.
The Genus Hypnodontopsis (Bryopsida, Rhachitheciaceae) in Baltic and SaxonAmberAuthor(s): Jan-Peter FrahmSource: The Bryologist, 108(2):228-235. 2005.Published By: The American Bryological and Lichenological Society, Inc.DOI: http://dx.doi.org/10.1639/9URL: http://www.bioone.org/doi/full/10.1639/9
BioOne (www.bioone.org) is a nonprofit, online aggregation of core research in the biological, ecological, andenvironmental sciences. BioOne provides a sustainable online platform for over 170 journals and books publishedby nonprofit societies, associations, museums, institutions, and presses.
Your use of this PDF, the BioOne Web site, and all posted and associated content indicates your acceptance ofBioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use.
Usage of BioOne content is strictly limited to personal, educational, and non-commercial use. Commercial inquiriesor rights and permissions requests should be directed to the individual publisher as copyright holder.

0007-2745/05/$0.95/0
The Bryologist 108(2), pp. 228 235Copyright q 2005 by the American Bryological and Lichenological Society, Inc.
The Genus Hypnodontopsis (Bryopsida, Rhachitheciaceae) inBaltic and Saxon Amber
JAN-PETER FRAHM
Nees Institut fur Biodiversitat der Pflanzen, Rheinische Friedrich-Wilhelms-Universitat, Meckenheimer Allee 170,53115 Bonn, Germany
Abstract. The genus Hypnodontopsis is represented by three extant species, all with a verylimited distribution. One was described as H. mexicana (Ther.) Robins., only known from singlelocalities in Mexico and Uganda, another is H. apiculata Iwats. and Nog., only known from asmall area in Japan, and the last is H. spathulata H. Akiyama and A. Tanaka, recently describedfrom Burma. The genus was, however, present in the Eocene (ca 45–58 myr bp) with five species;one of which is identical to the extant H. mexicana and is the most common moss in Baltic amber.All fossil species are illustrated and described. They are distinguished by variation of characterstates such as apiculate, lingulate, and piliferous leaves. The fact that the genus consisted of five(known) species in Tertiary, some of them very common, but only three extant species, which arequite rare, shows a decline of species number and frequency.
Keywords. Baltic amber, Hypnodontopsis, Paleobotany, Saxon amber, Tertiary.
For more than 140 years the knowledge of moss-es from Baltic amber (Eocene, ca 45 myr bp) hasbeen based on only a few publications (Caspary1907; Dixon 1922; Goeppert 1853; Goeppert &Berendt 1845; Magdefrau 1957). The first fossilmosses from Baltic amber were described by Goep-pert & Berendt (1845) in such form genera as Mus-cites hirsutissimus Goeppert & Berendt, Muscitesapiculatus Goeppert & Berendt, and Muscites du-bius Goeppert & Berendt. As expressed by the lastspecies name, it was not clear whether these frag-ments belonged to mosses. Later, Goeppert (1853)described 19 species of mosses as extinct speciese.g., Dicranum subscoparium Goeppert & Menge,Dicranum subflagellare Goeppert & Menge, Grim-mia subelongata Goeppert & Menge, and Polytri-chum suburnigerum Goeppert & Menge, all nominanuda, in order to express the similarity of thesefossils with extant species, or even such extant spe-cies as Phascum cuspidatum Hedw. or Rhytidi-adelphus squarrosus (Hedw.) Warnst. He alsotransferred species described by Goeppert and Ber-endt in form genera to extant genera or species e.g.,Muscites apiculatus to Barbula subcanescensGoeppert & Berendt and Muscites confertus toHymnostomum microstomum (Hedw.) R. Brown.All of these identifications were dubious and mostof the specimens cited by Goeppert (1853) orGoeppert and Berendt (1845) no longer exist. An-other disadvantage was that all earlier authors werenot bryologists. Furthermore, the knowledge ofmosses in Baltic amber was based on no more than
30 specimens. This gave the impression that only afew mosses are present in Baltic amber. Indeed,99% of all organisms preserved in amber are ani-mals, mostly insects (Weitschat & Wichard 1998).Of the 1% plant fossils, only a small percentage offossils consist of mosses. In addition, collections inmuseums are relatively poor and most of the fossilsembedded in amber are in private collections.Thanks to several amateur collectors from Germa-ny, I was able to study numerous specimens of fos-sil mosses, and this increased our knowledge of themosses of the Eocene markedly and resulted in sev-eral publications (Frahm 1994; 1996a,b; 1999a,b;2000a,b; 2001a,b; 2004a,b). About 150 specimensof mosses embedded in amber have been studiedsince 1994, and these are attributed to more than50, partly extant and partly extinct, taxa.
It is not exactly known how amber accumulatedin the Oligocene and Miocene in eastern Prussiaand Saxony (Schlee 1990; Weitschat & Wichard1998). There are two hypotheses for the origin ofBaltic amber (Schubert 1961). According to thefirst hypothesis, the resin was transported bystreams into the ocean; however, it is not knownexactly how the resin got from the trees into thestreams in such masses. According to another hy-pothesis, the ‘‘amber forests’’ were inundated by atransgression during which the trees were drowned,the resin was washed out by tides, transported byocean current, and deposited in marine clay sedi-ments.
Baltic amber is found in large masses in marine

2005] 229FRAHM: HYPNODONTOPSIS
deposits from the Upper Eocene and lower Oligo-cene near Kaliningrad. It is also found in smallquantities in Quaternary deposits from northeasternGermany to the German Sea, moved by glaciersduring the ice age, and in the Baltic Sea. It wasused in prehistoric times for burning fires (German‘‘Bernstein’’ means burnstone), and since Romantimes as gemstones.
Saxon amber is found in deposits from the UpperOligocene and Lower Miocene near the town ofBitterfeld, eastern Germany, where it was exploitedfor the petrochemical industry during the time ofthe German Democratic Republic. Small amountsof high quality amber were sorted out and sold inthe GDR along the coast of the Baltic Sea to tour-ists as Baltic amber. After the unification of Ger-man states, the exploitation stopped and the quarrydrowned, and also thousands of tons of amber. It isnot known exactly whether the Saxon amber is justa secondary deposit of Baltic amber or of indepen-dent origin. An argument for an independent originis that the amber was, according to IR-spectograph-ic studies, not produced by Pinus succinifera, butby another conifer, Cupressospermum saxonicumMai (Schubert 1961).
The increased knowledge of fossil mosses froman oak-pine forest of the Eocene in present Scan-dinavia now allows a first impression of the mossflora of the amber forest. New records of specimenswith sporophytes enable genera to be identified.Better preserved specimens showing structures thatuntil now were not visible, have given the oppor-tunity to identify specimens, which until now havebeen attributed only tentatively to families or gen-era and this has resulted in permanent changes ofnames.
The records of many specimens of various spe-cies of the same genus in Baltic and Saxon amberallow the first conspectus of all known fossil spe-cies of certain genera. As a first genus, Atrichumwas treated (Frahm 2004a). This study revealedthat all three species known from the amber forestare extinct species, which interestingly belong to agroup of species with small, non-undulate leaves;non-serrate abaxial costae; and simple, uniseriateteeth at the leaf margins. The nearest relatives forma group of three species with very limited rangesthat are found in pine-oak-forests in Mexico, PuertoRico, and China, and could be derived from theseEocene progenitors. All are very rare today.
As a second genus for a survey of all speciesand records, Hypnodontopsis has been chosen, be-cause it is the most common genus in Baltic amber.Twenty-five out of the 150 specimens of mossesthat were studied during the past 10 years, belongto this genus. They belong to five species. In con-trast, there are only three species known at present;
one only from the type locality, one from two re-cords worldwide, and another from a small area inJapan. For the first time the relatively rich fossilrecords of Hypnodontopsis allow the addition of athird dimension — the time dimension, with a win-dow that opens 45 m yr ago. This gives us theopportunity to see which species became extinctover this time period, which survived for a longtime, and how the species were morphologicallystructured at the time.
Hypnodontopsis is a genus of the Rhachitheci-aceae, a family with several genera (RhachitheciumLe Jol., Jonesiobryum Allen & Pursell, Rhachithe-ciopsis P. de la Varde, Tisserantiella P. de la Varde,and Hypnodontopsis Iwatsuki & Noguchi), all ofwhich have few species. The family was revised byGoffinet (1997). The genus Hypnodontopsis wasoriginally monotypic and based on the descriptionof H. apiculata Iwats. & Nog., endemic to Japanand described only about 40 years ago. Later, Rob-inson (1964) placed Oreas mexicana Theriot (Ther-iot 1928) in this genus. This species has only beenrecorded once outside of Central America, namelyfrom Uganda (Hodgetts & Goffinet 1998). Finally,a third extant species, H. spathulata, was discov-ered in Burma (Akiyama & Tanaka 2002), and it isso far only known from the type locality.
HISTORY OF DISCOVERY OF HYPNODONTOPSIS IN
BALTIC AND SAXON AMBER
Hypnodontopsis was first reported from the Eo-cene by Frahm (2000a) based on a specimen fos-silized in Saxon amber, with diagnostic sporo-phytes: a twisted and cygneous seta of 1.5–2.0 mmlength, an ovate capsule with 16 longitudinal ribs,and a peristome in which the teeth are united ineight pairs. The perichaetial leaves are differenti-ated and sheathing. It seemed, however, not to beidentical with either of the two extant species, be-cause papillae characteristic for these species couldnot be observed in the fossil specimen, and wastherefore described as new as H. fossilis J.-P.Frahm.
Another similar sporophyte with twisted, cyg-neous seta, but shorter capsule was later found ona plant in Baltic amber (Frahm 2001b). Sterileplants of this species were, however, already known150 years before and described as Muscites confer-tus by Goeppert and Berendt (1845). Klebs (in Cas-pary 1907) called it the most common moss in am-ber. The fossil material usually consists of comaltufts, which apparently serve for vegetative propa-gation. The leaves are narrow linear, have a per-current costa and laminal cells conspicuously ar-ranged in only 5–6 rows. Based on the discoveryof the sporophyte, the new combination Hypnodon-

230 [VOL. 108THE BRYOLOGIST
FIGURES 1–5. Hypnodontopsis casparyi. — 1. Illustrations of syntypes of Dicranites casparyi (figs. 42–45) and thetype of Dicranites obtusifolius (fig. 46) from Caspary (1907). The specimens 42, 44, and 45 have much shorter leavesand are referred to Muscites confertus (Hypnodontopsis conferta). Figure 43 has been chosen as the lectotype. It is

2005] 231FRAHM: HYPNODONTOPSIS
←
apparently identical to the type of Dicranites obtusifolius (fig. 46), a name which has been rejected. — 2. Hypnodon-topsis casparyi, type of Dicranites grollei. — 3. Hypnodontopsis casparyi, plants in situ on a piece of bark (Velten 9).— 4 Hypnodontopsis casparyi, part of Figure 3 showing the narrow lamina with the rounded-quadrate laminal cells indistinct rows. — 5. Sterile plant (Hoffeins 874).
topsis conferta (Goeppert & Berendt) J.-P. Frahmwas made. At that time it was already supposed thatthe fossil plants could perhaps be identical with theextant H. mexicana (Ther.) Robins. that resembledthe fossil material but had distinct pluripapilloselaminal cells. Shortly after a piece of amber wasdiscovered, in which the leaves of the plant of Hyp-nodontopsis conferta were decolorized allowingdistinct papillae to be recognized (Frahm 2004b),and supporting the hypothesis that this specimen isidentical to the extant species. Since the name ofthe fossil plant was much older, the name of thefossil species was chosen. However, according to§11(7) of the ICBN, the names of fossil speciesantedating extant species have no priority. There-fore this species must be renamed H. mexicana.
The recognition of Hypnodontopsis by fertilespecimens revealed that at least some species canbe recognized in sterile condition by the distinctrows of roundish-quadrate upper and median lam-inal cells (see Sharp et al. 1994, fig. 447). Thisdraws attention to additional fossil material withsuch laminae. Thus, the species described as Di-cranites casparyi Klebs also has only 5–7 rows ofroundish-quadrate cells but much longer leaves.Therefore a third species was added to the genus(Frahm 2004b), and the new combination Hypno-dontopsis casparyi (Klebs) J.-P. Frahm was made.
Before the discovery of Hypnodontopsis in am-ber, another fossil moss with twisted, sinuose seta;short capsule; and eight peristome pair teeth hadbeen described as Dicranites grollei J.-P. Frahm(Frahm 1999b). A comparison of the leaves showedthat these resemble those of H. casparyi, which wasonly known in sterile condition. Therefore D. grol-lei was placed in synonymy with H. casparyi(Frahm 2004b). Later, another fossil was foundwith the typical laminal cells arranged in five rows,however, with leaves ending in a long hairpoint;this was newly described as H. pilifer (Frahm2004b). Recently, another specimen was discoveredwith the same arrangement of laminal cells; how-ever, with lingulate leaves resembling the extant H.apiculata, but with no apiculus (Frahm, in press);this was described new as H. lingulata. The genericplacement of these two species is less sure becausethey are only known from one specimen each insterile condition.
KEY TO THE EXTANT AND EXTINCT (†) SPECIES
OF HYPNODONTOPSIS
In total, five species of Hypnodontopsis are known fromBaltic and Saxon amber; these can be distinguished by thefollowing. The extant species (lingulata and spathulata)are included in the key to show their systematic affiliation.Hypnodontopsis mexicana is known from both fossil andextant material.
1. Costa excurrent in long hairpoint -------------- H. pilifer†1. Costa ending in or below leaf tip, or excurrent as
short apiculus -------------------------------------------------------------- 2.2. Leaves short, lingulate, broadly obtuse ----------- 3.2. Leaves lanceolate, acute ------------------------------------ 5.
3. Leaf tips with ashort apiculus ---------------------------------- 4.3. Leaf tips obtuse --------------------------------------- H. lingulata†
4. Leaves less than 1 mm long. Costa ceasingbefore apex -------------------------------------- H. spathulata
4. Leaves longer than 1 mm. Costa excurrent --
----------------------------------------------------------------H. apiculata5. Leaves 10–20 times as long as wide. Capsule
ovate ---------------------------------------------------------- H. casparyi†5. Leaves 5–10 times as long as wide. Capsule ovate
or cylindric ------------------------------------------------------------------ 6.6. Capsule cylindric. Seta short, only 0.5 mm
long ------------------------------------------------------- H. fossilis†6. Capsule ovate. Seta 1.5–2.0 mm long -------
--------------------------------------------------------------- H. mexicana
HYPNODONTOPSIS CASPARYI (Klebs) J.-P. Frahm,Palaeontographica in press. FIGS. 1–5
Dicranites casparyi Klebs, Abh. K. Preuss. Geol. Lan-desanst. n. ser. 4: 52. 1907.
Dicranites obtusifolius Caspary & Klebs, Abh. K.Preuss. Geol. Landesanst. n. ser. 4: 54. 1907.
Dicranites grollei J.-P. Frahm, Haussknechtia 9: 129.1999.
Plants to 4 mm high. Basal leaves 1.6 mm, upperleaves 2 mm long. Leaves narrowly lanceolate, 10–20 times longer than wide, margins slightly serrateat leap apex. Laminal cells round to quadrate, ar-ranged in 5–10 rows on both sides of costa. Costacomparably wide, filling ¼ of leaf width, ending inleaf apex or shortly below. Seta to 2 mm long,twisted, sinuose. Capsule ovate, 0.4 mm long, con-tracted below mouth. Peristome teeth broadly lan-ceolate.
The description of Dicranites casparyi by Klebsin Caspary (1907) was based on two specimens(‘‘Ehemals Kunow’sche Sammlung Nr. 79, 153)that were illustrated in figs. 42–45. The types arestill in the palaeontological collection of the Natur-kundemuseum in Berlin. It is obvious from the fig-

232 [VOL. 108THE BRYOLOGIST
ures that these syntypes belong to different species;one with relatively short leaves (figs. 42, 44, 45)and one with very long and narrow leaves (fig. 43).Klebs placed both in the same species because theywere present in the same piece of amber. The plantswith shorter leaves, however, look much the sameas those plants described as Muscites confertus, andtherefore D. casparyi was lectotypified based on thefigure with long leaves (Frahm 2001a). It is alsoobvious from Caspary (1907) that the illustrationsof Dicranites obtusifolius resemble that of D. cas-paryi. According to the description of D. obtusifol-ius, this species has 7–9 rows of laminal cells,which strongly supports this species belonging toHypnodontopsis. Therefore Dicranites obtusifoliuswas placed in synonymy with H. casparyi (Frahmin press). The name of Dicranites obtusifolius wasrejected because this species has extremely narrowleaves that are not at all blunt.
The fact that both, H. casparyi and H. conferta,were found in the same specimen (Kunow 151 inCaspary 1907) is an indication that both specieslived at the same time together in the same region,maybe even on the same tree, as species of Ortho-trichum do at present. The specimen, Velten 9, in-cludes a piece of bark with a low turf of this species(in which a plant of H. lingulata is intermixed).This in situ enclosure demonstrates that both spe-cies were epiphytes.
Specimens seen.—B: (5 Baltic amber): Kunow 153(lectotype of H. casparyi, Caspary 1907); Kunow 151(type of Dicranites obtusifolius, Caspary 1907); Glink 1(Frahm 1999b, type of Dicranites grollei); Teuber 1316(Frahm 1999a); Velten 9, Grohn X2 (Frahm in press); Hof-feins 874 (Frahm 2000a as Dicranites casparyi); Glink 2(holotype of Dicranites grollei, Frahm 1999b);. S: Kutsch-er 11, 14 (Frahm 1999a); Kutscher 2c, 3b (Frahm 2000).
HYPNODONTOPSIS FOSSILIS J.-P. Frahm, Lindbergia25: 36. 2000. FIGS. 10–11
Plants to 3 mm high. Leaves narrowly lanceolateto linear, up to 1.2 mm long, 10–20 times as longas wide, acute. Costa percurrent. Laminal cellsround, shortly rectangular only at leaf base. Seta0.5 mm long, twisted. Capsule 0.5 mm long, cylin-dric, with 8 longitudinal furrows. Peristome teeth16, in pairs.
This species is characterized by much longer andnarrower capsules as compared with the other spe-cies of the genus, which have capsule four timeslonger than broad. The capsules gradually mergeinto the seta and are not abruptly delimited.
Specimens seen.—B: Hoffeins 1417/2, Grabenhorst La7, 14, Grohn 2042 as H. conferta (Frahm 2004b). S: (5Saxon amber): Kutscher s.n. (holotype, Frahm 2000a).
HYPNODONTOPSIS LINGULATA J.-P. Frahm, Palaeon-tographica (in press). FIG. 12
Plants minute, without stem, sparsely foliate.Leaves lingulate, obtuse or broadly triangular atapex. Costa ending below leaf tip, reaching 4/5 ofthe leaf length. Laminal cells in 6 rows besidenerve, mamillose.
This species is known from only a single plantthat grew intermixed in a tuft of Hypnodontopsiscasparyi. Therefore the variation of characters isnot known. It resembles very much the extant H.apiculata from Japan in plant size and leaf shape.The latter however has a costa which is shortly ex-current. The extant species seems therefore to be aclose relative of the fossil species.
Specimens seen.—B: Velten 7 (Frahm 2005).
HYPNODONTOPSIS MEXICANA (Ther.) Robins., THE
BRYOLOGIST 67: 449. 1964. (FIGS. 6–9).
Oreas mexicana Ther., Smiths. Misc. Coll. 81: 2. 1928.Muscites confertus Goeppert & Berendt, Bernst. Org.
Reste Vorwelt 1: 112. 1845.Hypnodontopsis conferta (Goeppert & Berendt) J.-P.
Frahm, Trop. Bryol. 20: 80. 2001. [The gender of Hyp-nodontopsis is feminine. Therefore the species must bewritten conferta and not confertus as in Frahm (2001b)].
Plants small, to 5 mm tall. Leaves in rosettes,erect-patent, narrowly lanceolate, lower ones 1 mm,upper ones to 1.5–1.8 mm long. Perichaetial leavesdifferentiated, sheathing. Costa percurrent or end-ing shortly before apex, thick, dark brown, filling¼ of leaf width, protuberant at back of leaf. Lam-inal cells roundish-quadrate, thick-walled, mamil-lose, in 5–6 longitudinal rows at each side of costa.Seta 1.5–2.0 mm long, sigmoid, twisted. Capsule0.4 mm long, ovate, contracted at mouth. Peristometeeth erect, broadly lanceolate, in 8 pairs.
The fossil consists mostly of low rosettes ofleaves, about 3 mm high that seemed to be detachedleaf rosettes of the apical parts of the plants andmay serve for vegetative propagation, perhapsblown into the resin of the amber pine. Sometimesup to five of such fragments are found in one pieceof amber (e.g., Kutscher 5, Frahm 1999a). Thewhole plants are not much larger, as shown by piec-es of bark on which small mats of plants are pre-served. Theses pieces of bark demonstrate that thisspecies was an epiphyte.
The laminal cells have been described as smoothby Frahm (1999a). In fact, they are pluripapillose,which cannot be seen in most fossil specimens pos-sibly because the space between the papillae isfilled with resin.
Hypnodontopsis mexicana is not only the mostcommon, but is also a very dubious taxon. De-
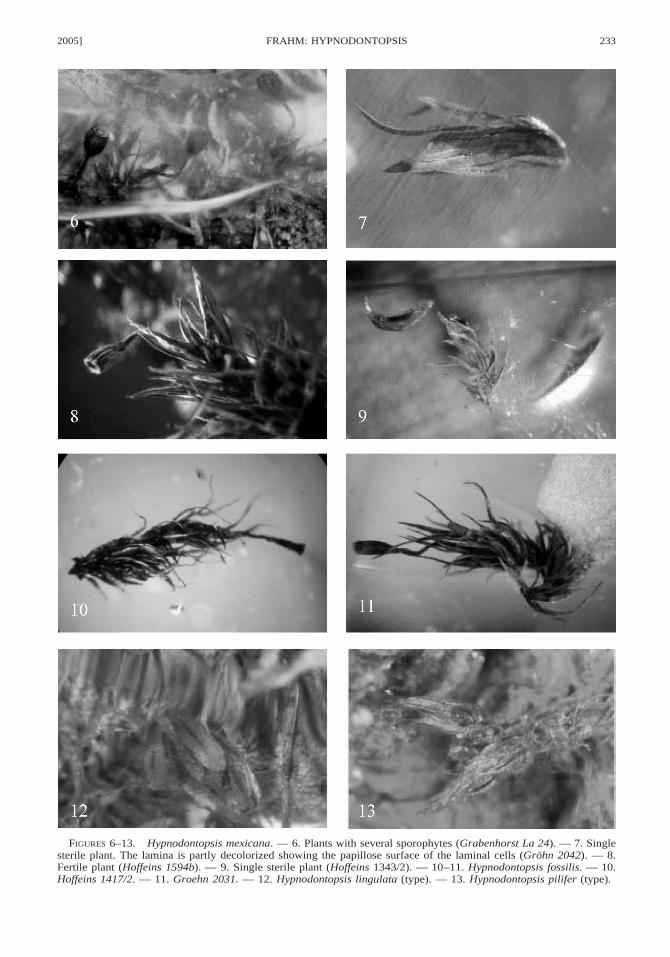
2005] 233FRAHM: HYPNODONTOPSIS
FIGURES 6–13. Hypnodontopsis mexicana. — 6. Plants with several sporophytes (Grabenhorst La 24). — 7. Singlesterile plant. The lamina is partly decolorized showing the papillose surface of the laminal cells (Grohn 2042). — 8.Fertile plant (Hoffeins 1594b). — 9. Single sterile plant (Hoffeins 1343/2). — 10–11. Hypnodontopsis fossilis. — 10.Hoffeins 1417/2. — 11. Groehn 2031. — 12. Hypnodontopsis lingulata (type). — 13. Hypnodontopsis pilifer (type).

234 [VOL. 108THE BRYOLOGIST
scribed as Muscites confertus, Goeppert (1853)synonymized it with Hymenostomum microstomum(Weissia brachycarpa [Nees & Hornsch.] Jur.). Thelatter has, incurved leaf margins and is thereforenot correct. It is also a species of bare soil. Ac-cording to the illustrations of the types of Dicran-ites casparyi, part of these syntypes also belong toMuscites confertus.
Hypnodontopsis mexicana is distinguished fromH. fossilis by shorter and broader capsules and lon-ger setae. These differences make it impossible todifferentiate sterile plants, and all sterile plants areattributed to H. mexicana. Dicranites grollei wasplaced in synonymy with H. conferta (Frahm2004b) since it has same shape of capsule; however,it has longer leaves. When it became apparent thatDicranites casparyi is also a species of Hypnodon-topsis, it was transferred to H. casparyi (Frahm, inpress).
Specimens seen.—B: Goeppert & Berendt (1845) fig.28, type of Muscites confertus; Caspary (1907) fig. 44,45,syntypes von Dicranites casparyi; Caspary (1907) as Hy-menostomum microstomum; Grohn 119 (Frahm 1999a asakrokarpes Laubmoos 1); Teuber 1674 (Frahm 2001a asMuscites confertus), Wichard 20 (Frahm 2001a as Hyp-nodontopsis confertus); Hoffeins 1594 (Frahm 2001b);Grohn 616, Hoffeins 1417/1, Grabenhorst La 23, 24(Frahm 2004b); Sterile material that can only tentativelybe attributed to H. conferta: Hoffeins 1343/2,4,5,6, Grohn2031, Grabenhorst 6,8,9,16 (Frahm 2004b). S: Kutscher4,5,12 (Frahm 1999a als akrokarpes Laubmoos 1).
HYPNODONTOPSIS PILIFER J.-P. Frahm, Journal ofPalynology and Palaeobotany 129: 86. 2004.
FIG. 13.
Plant with several branchlets, 1–2 mm high.Leaves narrowly linear, longly apiculate, marginsnarrowly recurved. Lamina consisting of 5–6 rowsof round, mamillose cells arranged in distinct lon-gitudinal rows. Costa excurrent as long, straighthair.
The amber includes a relatively large piece ofbark–3.5 3 2.0 cm–that is fully covered with acrustaceous lichen with ‘‘lecanoroid’’ apothecia.Two plants of an epiphytic hepatic (Frullania sp.)are on the lichen. This confirms that this species ofHypnodontopsis was also an epiphyte.
A similar fossil moss with such narrow linearleaves and long hairpoint was described as Musci-tes pilifer (Frahm 1999b). In the type specimen ofthe latter species, the shape of the laminal cells isnot visible, however, a similar plant described byFrahm (2004b) shows elongate laminal cells.
Specimens seen.—B: Witsch 1 (type, Frahm 2004b).
DISCUSSION
The genus Hypnodontopsis is comprised of sevenspecies, three extant species, one of which is alsoknown from Baltic amber, and four extinct species.Of the extant species, H. apiculata is confined toJapan and is so rare that it was included in the‘‘World Red List of Bryophytes’’ (Geissler et al.1997). Hypnodontopsis mexicana is even rarer andis known only from one locality in Mexico (typeof Oreas mexicana) and a recent collection fromUganda (Hodgetts & Goffinet 1998). The rarestspecies is the recently described H. spathulata,which is only known from the type locality in Bur-ma. In contrast, the genus was much more commonin the Eocene as indicated by the fossils in Balticand Saxon amber. The fossil records of H. mexi-cana are much more numerous than the two extantrecords today, although the chances of finding fos-sil specimens are much smaller than living speci-mens. Hypnodontopsis casparyi, H. fossilis, H. lin-gulata, and H. pilifer are only known from Balticand Saxon amber. Within the genus, the fossil spe-cies are more numerous than extant species, and itcan be argued that the genus Hypnodontopsis ap-proaches extinction today. Extinction is a normalphenomenon in evolution. It can be sudden (as forinstance, caused by disasters e.g., at the end of theMesozoic) or slow. Hypnodontopsis is an exampleof such slow extinction, although all (fossil and ex-tant) species are epiphytes that are usually regardedas more recent taxa. It can be postulated that thegenus was perhaps even richer in species in theTertiary, as our knowledge of the genus is limitedby comparably few fossil records. Four of five spe-cies have undergone extinction since that time; onlyone remains and has been found in two very distantplaces. The two remaining extant species (H. api-culata and H. spathulata) seem to be derived froma fossil ancestor close to H. lingulata. The fact thatthe Rhachitheciaceae include seven genera, butonly 15 species (Goffinet 1997) suggests that thisfamily was richer in species in the past.
The fossil species of Hypnodontopsis show abroad range of variation of characters, mainly inleaf shape. The leaves are either piliferous or not,linear, lanceolate, or lingulate. The sporophyticcharacters with short, twisted, cygneous setae anda peristome with paired teeth (known from onlythree species) are, in contrast, very constant. Onlythe shape of the capsule varies. Finally, the Eoceneamber forest was a pine-oak forest. Interestingly,one of the extant species, H. spathulata, is knownfrom a pine forest (Akiyama & Tanaka 2002).
ACKNOWLEDGMENTS
I wish to thank Bernard Goffinet and an unknown re-viewer for important additions and helpful remarks.

2005] 235FRAHM: HYPNODONTOPSIS
LITERATURE CITED
AKIYAMA, H. & A. TANAKA. 2002. Hypnodontopsis spa-thulata H. Akiyama & A. Tanaka (Rhachitheciaceae;Musci), a new species from Myanmar (Burma). Bry-ological Research 8: 131–136.
CASPARY, R. 1907. Die Flora des Bernsteins. Abhandlun-gen der preußischen geologischen Landesanstalt N.F.4. Berlin.
DIXON, H. N. 1922. Notes on a moss in amber. Journal ofBotany 60: 149–151.
FRAHM, J.-P. 1994. Die Identitat von Muscites hauchecor-nei Caspary & Klebs (Musci) aus Baltischem Bern-stein. Nova Hedwigia 58: 239–243.
———. 1996a. Laubmoose aus Baltischem Bernstein. Pa-laeontographica Abt. B 241: 127–135.
———. 1996b. Mosses newly recorded from Saxonianamber. Nova Hedwigia 63: 525–527.
———. 1999a. Die Laubmoosflora des Baltischen undBitterfelder Bernsteins. Mitteilungen des Geologisch-Palaontologischen Staatsinstitut 83: 219–238.
———. 1999b. Neue bemerkenswerte Laubmoosfundeaus Baltischem Bernstein. Haussknechtia 9: 129–132.
———. 2000a. New and interesting records of mossesfrom Baltic and Saxonian amber. Lindbergia 25: 33–39.
———. 2000b. Neue Laubmoosfunde aus BaltischemBernstein. Cryptogamie, Bryologie 21: 121–132.
———. 2001a. Neue Laubmoosfunde aus Sachsischemund Baltischem Bernstein. Nova Hedwigia 72: 271–281.
———. 2001b. Hypnodontopsis confertus from Balticamber. Tropical Bryology 20: 79–82.
———. 2004a. The genus Atrichum in Baltic amber. Jour-nal of the Hattori Botanical Laboratory 95: 219–227.
———. 2004b. A new contribution to the moss flora ofBaltic and Saxon amber. Review of Palaeobotany andPalynology 129: 81–101.
———. 2005. Neue Laubmoosfunde in Baltischem Bern-stein. Palaeontographica (in press).
GEISSLER, P., B. TAN, T. HALLINGBACK. 1997. Additions tothe World Red List of bryophytes. Bryological Times93: 1–7.
GOEPPERT, H. R. 1853. Uber die Bernsteinflora. Monats-berichte der Koniglich preußischen Akademie derWissenschaften zu Berlin 1853: 450–477.
——— & G. C. BERENDT. 1845. Der Bernstein und diein ihm befindlichen Pflanzenreste der Vorwelt. Berlin.
GOFFINET, B. 1997. The Rhachitheciaceae: revised circum-scription and ordinal affinities. THE BRYOLOGIST 100:425–439.
HODGETTS, N. G. & B. GOFFINET. 1998. Hypnodontopsismexicana (Ther.) H. Rob., a genus and species new toAfrica. Journal of Bryology 20: 251–252.
MAGDEFRAU, K. 1957. Flechten und Moose in BaltischemBernstein. Berichte der Deutschen Botanischen Ge-sellschaft 70: 433–435.
ROBINSON, H. 1964. New taxa and new records of bryo-phytes from Mexico and Central America. THE BRY-OLOGIST 67: 469–458.
SCHLEE, D. 1990. Das Bernsteinkabinett. Stuttgarter Bei-trage zur Naturkunde Serie C 28: 1–100.
SCHUBERT, K. 1961. Neue Untersuchungen uber Bau undLeben der Bernsteinkiefern. Berichte des Niedersach-sichen Amtes fur Bodenforschung 1–149, 21 Tafeln.
SHARP, A. J., H. CRUM, & P. ECKEL. 1994. The moss floraof Mexico. Memoirs of the New York Botanical Gar-den 69: 1–580.
THERIOT, I. 1928. Mexican mosses collected by BrotherArsene Brouard. Smithsonian Miscellaneous Collec-tions 81: 1–26.
WEITSCHAT, W. & W. WICHARD. 1998. Atlas der Pflanzenund Tiere im Baltischen Bernstein. (Dr. Pfeil) Mun-chen.
ms. received July 7, 2004; accepted Jan. 25, 2005.





























