Embed Size (px)
Citation preview

Oecologia (Berl) (1982) 54:236-242 Oecologia �9 Springer-Verlag 1982
The Effect of Vapor Pressure on Stomatal Control of Gas Exchange in Douglas Fir (Pseudotsuga menziesii) Saplings Frederick C. Meinzer* Dept. of Botany, University of Washington, Seattle, WA 98195, USA
Summary. Increasing leaf to air vapor pressure deficit (VPD) caused reductions in stomatal conductance of both current year and previous season needles of Pseudotsuga menziesii saplings. The stomata of current year needles were found to be more responsive to changes in VPD than those of previous season needles. The reductions in stomatal con- ductance of current year needles were not associated with decreases in xylem pressure potential. In fact, the reductions in stomatal conductance of current year needles were some- times sufficient to reduce transpiration and thus raise xylem pressure potential even though VPD was increasing. There was a decline in stomatal responsiveness to VPD in current year needles between early and late summer. Pressure-vo- lume curves determined for different age needles at different times of the year suggested that differences and changes in stomatal responsiveness to VPD may have been caused in part by differences and changes in needle water potential components. Hexane washes of current year needles during the late summer succeeded in partially restoring their VPD sensitivity, suggesting that changes in the water permeabili- ty of the external cuticle during needle maturation may also have played a role in causing the summer decline in VPD responsiveness.
In both current and previous year needles VPD-induced changes in stomatal conductance had a greater relative ef- fect on transpiration (qw) than on net photosynthesis (PhN). In maturing needles the ratio of the sensitivities of tran- spiration and net photosynthesis to changes in stomatal conductance, (~qw/~gs)/(~PhN/~gs), remained nearly con- stant as VPD was varied. This provides experimental sup- port for a recent hypothesis that stomata respond to envi- ronmental fluctuations in such a manner as to maintain the above ratio constant, which optimizes CO2 uptake with respect to water loss.
Introduction
It has become apparent that the stomata of many species are sensitive to changes in the vapor pressure gradient be- tween the leaf and the air (Sheriff 1979; Farquhar 1978). I f stomatal closing in dry air is brought about by a negative feedback system involving changes in bulk leaf water status, then decreases in stomatal conductance should be asso-
* Present address: Departamento de Biologia, Facultad de Cien- cias, Universidad de los Andes, M~rida, Venezuela
ciated with increased leaf water deficits. This type of system appears to operate in a number of species (Sheriff 1977), but in the majority of species in which stomatal response to humidity has been observed, humidity-induced changes in stomatal aperture seem to be independent of bulk leaf water status (Lange et al. 1971; Schulze et al. 1972; Hall and Hoffmann 1976; Sheriff 1977; Edwards and Meidner 1978; Lange and Medina 1979; Ludlow and Ibaraki 1979; Pallardy and Kozlowski 1979).
Various mechanisms have been suggested to account for direct stomatal responses to humidity. Some of these include dehydration of mesophyll cell walls resulting in a decline in mesophyll conductance at high vapor pressure gradients (Whiteman and Koller 1964), and increased evap- oration from internal guard and epidermal cell walls caus- ing toss of turgor in these cells (Sheriff 1977). Indeed, recent experiments (Meidner 1975; 1976a, b) and computer simu- lations (Tyree and Yianoulis 1980) have suggested that the bulk of the evaporation within the substomatal cavity should occur within a small radius from the stomatal pore i.e. the inner guard and epidermal cell walls. However, none of these hypotheses involving sites of evaporation inside the leaf can explain those stomatal responses to humidity characterized by a decrease in transpiration rate with an increase in the vapor pressure gradient. This type of humidi- ty response has been termed a "feedforward" response and is discussed in detail by Farquhar (1978). Peristomatal transpiration, direct evaporation from the external cuticle of guard and subsidiary cells (Seybold 1961, 1962), is the only proposed mechanism consistent with feedforward re- sponses (Sheriff 1979; Farquhar 1978).
It is evident that direct stomatal responses to humidity act to conserve water and inhibit development of severe leaf water deficits, but their full importance in terms of control of water use efficiency cannot be evaluated without simultaneous measurement of CO2 uptake. An improve- ment of water use efficiency resulting from humidity-in- duced stomatal closure has been demonstrated by Schulze et al. (1975).
Total gas exchange measurements in studies of stomatal response to humidity are especially important in view of suggestions that feedforward responses are consistent with behavior predicted by a recent hypothesis of stomatal opti- mization of carbon gain with respect to water loss proposed by Cowan and Farquhar (1977) and Cowan (1977). Accord- ing to this hypothesis, optimal stomatal behavior is that which minimizes loss of water for a given amount of carbon
0029-8549/82/0054/0236/$01.40

237
fixed during a given time period. This would be accom- plished if the ratio of the sensitivities of transpiration (qw) and net photosynthesis (PhN) to changes in stomatal con- ductance (g~)
5 ~ = 5qw/SPh N
remained constant during the time period. Responses of tobacco, hazel (Farquhar et al. 1980) and cowpea (Hall and Schulze 1980) stomata to humidity have been shown to be consistent with this hypothesis. However, its further veri- fication over a wider range of environmental conditions and in a wider range of species has been hindered by the experimental difficulties involved in determining 5PhN/~g ~ when more than one environmental component is allowed to vary simultaneously.
The stomata of Douglas fir (Tan et al. 1977) and other conifers such as Picea engelmannii (Kaufmann 1976), Picea sitchensis (Grace et al. 1975) and Pinus banksiana (Bliss and Mayo 1980) have been reported to be responsive to humidi- ty. In the case of Pieea sitchensis and Pinus banksiana, the response seems to be independent of bulk leaf water status (Watts and Neilson 1978; Bliss and Mayo 1980) but the nature of the humidity response in other conifers is not well studied. To date, there have been no investigations of stomatal response to humidity in conifers with respect to its possible role in the optimization of gas exchange by stomata.
The objectives of this study were to investigate the na- ture of stomatal response in Douglas fir to changes in the external vapor pressure and to examine the influence of observed stomatal responses on total gas exchange. Gas exchange behavior was examined from the standpoint of its degree of conformity to optimal behavior as predicted by the Cowan and Farquhar (1977) hypothesis described above. Foliage of current and previous seasons was exam- ined because field observations (Meinzer, unpublished) sug- gested a marked difference in responsiveness to vapor pres- sure of these two age classes. It is likely that Douglas fir exhibits significant adaptations with respect to regulation of water use economy in response to changing humidity, since it grows in regions in which periods of summer drought and high atmospheric moisture stress may occur.
Materials and Methods
Plant Material
Experiments were carried out on potted Douglas fir (Pseu- dotsuga menziesii (Mirb.) Franco) plants grown from seed of Twin Harbors seed source (030-10-1) sown in spring, 1976 at the Weyerhauser Mima Nursery south of Olympia, Washington. The plants were lifted and potted in March, 1978 and grown outdoors with the pots set in a sawdust- filled cold frame to prevent soil freezing during the winter. The saplings were watered as needed to maintain adequate soil moisture levels and were approximately 80 to 100 cm tall at the time of their use in the experiments. Measure- ments were carried out with either previous season (1979) foliage or foliage produced during the current (1980) grow- ing season. Bud burst occurred between late April and early May and the first experiments with current foliage were not performed until June, after the needles were nearly fully expanded.
Apparatus
Experiments involving the response of whole plants to changes in vapor pressure were carried out in a controlled environment room. Light was furnished by a bank of fluo- rescent tubes. Leaf temperatures at the level of the foliage samples were measured with copper-constantan thermocou- ples held in place on the lower, stomatal surfaces of the needles. Water vapor pressure in the room was measured with an Assmann Psychrometer, and with the humidity sen- sor of a steady-state ventilated diffusion porometer (Meinzer 1981). Stomatal conductance was measured with this porometer which is similar to those described by Beard- sell et al. (1972) and Bingham and Coyne (1977). Leaf water potential was estimated by measuring xylem pressure poten- tial of twigs with a pressure chamber.
Gas exchange rates were measured in attached twigs sealed in thermoelectrically cooled cuvettes (Salo 1974) in- side a controlled environment chamber (Controlled Envi- ronments, Inc. Pembina, North Dakota). Light was pro- vided by a bank of fluorescent tubes. Photon flux density was measured with LI-COR quantum sensors (LI-COR, Inc. Lincoln, Nebraska). Leaf temperature was measured with copper-constantan thermocouples held in place on the lower, stomatal surface of the needles. The cuvettes were equipped with fans to minimize boundary layer influences and to ensure uniform composition of the air. Air was passed through the cuvettes at a rate of 2 1 min- a in an open circuit system. Differences in CO 2 concentration of the ingoing and outgoing air streams were measured with a Hartmann and Braun URAS II infrared gas analyzer. Water vapor differentials were measured with an EG & G Modell 911 dew point hygrometer, or a Vaisala thin film capacitance relative humidity sensor. In the case of the latter, sensor temperature was recorded to permit calcu- lation of absolute humidity. The vapor pressure in the in- going air was controlled by first saturating the air then passing it through a thermoelectrically cooled vapor trap.
Procedures
When responses of whole plants to changes in vapor pres- sure were to be examined, well-watered plants were given an additional watering the day before the experiment. They were placed in the controlled environment room the after- noon prior to the experiment. The photoperiod was compa- rable to the natural photoperiod outside at the time the experiment was carried out. Photosynthetically active pho- ton flux density at the level of the foliage samples used for stomataI conductance measurements was approximately 250 ~tE m-Zs 1. Vapor pressure in the room was set at the initial level to be used in the following day's experiment. Fluctuation in vapor pressure at the initial and subsequent levels was +0.2 kPa. The leaf thermocouples were also at- tached at this time. Leaf temperature of the foliage samples selected for stomatal conductance measurements were taken to be the average of four thermocouples placed on different plants at levels similar to those of the stomatal conductance samples. The difference between thermocouples was never greater than 2 ~ C. The leaf-air vapor pressure difference (VPD) was calculated as the difference between saturation vapor pressure at average leaf temperature and ambient vapor pressure and thus yielded the same value for all of the plants. Chamber air temperature was set such that leaf temperatures were between 25 and 27 ~ C. This was neces-

238
sary in order to attain the desired ranges of VPD. Experi- ments were initiated in the morning at least 2 h after the beginning of the photoperiod. Measurements were started at high ambient vapor pressure which had been set the previous afternoon. Vapor pressure was decreased in steps during the remainder of the experiment. Steady-state sto- matal conductance of the same attached twigs at each vapor pressure was determined by comparing successive (2 to 3) measurements obtained with the porometer. In control ex- periments carried out at constant VPD', the plants main- tained nearly steady stomatal conductance throughout the day. After determination of steady-state stomatal conduc- tance at a given VPD, xylem pressure potential of at least two twigs per plant was measured.
In one controlled environment room experiment, the cuticle was partially removed from needles on selected twigs 7 d prior to the experiment by plunging attached twigs into hexane for approximately 5 sec. The twigs were then with- drawn from the hexane, shaken to remove excess solvent and quickly dried in a stream of compressed air. This proce- dure was repeated twice with each twig. Dead or visibly damaged needles (about 10%) were removed on the day prior to the experiment.
Pressure-volume curves of twigs bearing a single age class of needles were measured in late spring and late sum- mer to determine if there was any relationship between characteristics of stomatal response to humidity and water potential components of the needles. A detailed theoretical treatment of the pressure-volume curve is given by Tyree and Hammel (1972). Briefly, the pressure-volume curve as determined here consists of a plot of total leaf water poten- tial versus the recirprocal of relative water content. Twigs bearing foliage of the desired age were excised under water. The cut ends were maintained under water and the samples were wrapped in a plastic bag and placed in darkness over- night (14-16 h) to permit hydration to near maximum rela- tive water content. The twigs were removed from the water and the ends were quickly trimmed to remove the portion that had been under water. The cut ends were freed of bark. The stems were then fitted into rubber stoppers and the initial weights of the twig-stopper assemblies were re- corded on a Mettler balance to the nearest rag. The twigs were quickly sealed into a pressure chamber and the pres- sure increased slowly to determine the balancing pressure (pressure at which the sap first appeared at the cut surface). The pressure was released slowly and the twig-stopper as- semblies were placed on a table top to allow transpiration to occur. At frequent intervals initially (15 to 30 rain), the twig-stopper assemblies were reweighed and the balance pressure determined. Less frequent measurements (every t to 2 h) were necessary when the twigs were in later stages of dehydration. After the last measurement the stoppers were weighed separately in order to determine twig weights. The twigs were then oven dried for 24 h and the dry weights recorded. Relative water content (RWC) on a decimal basis was defined as
R, _ fresh wt - dry wt W k 5 . . . . .
saturated wt - dry wt
where fresh weight is the weight at a given balancing pres- sure and saturated weight is the initial fresh weight after overnight hydration.
The basic procedure for experiments in the gas exchange system was as described earlier (Meinzer 1982). Experi-
ments were initiated at high ambient vapor pressure and the vapor pressure was reduced in steps allowing sufficient time for rates of gas exchange to become steady after each change. Leaf temperatures were maintained between 25 and 26 ~ C in order to attain the desired levels of VPD.
Stomatal conductance (g~; cm s-1), transpiration rate (qw; mg cln-2s -1) and net photosynthesis (PhN; mg CO2 cm-2s -1) were determined as described earlier (Meinzer 1981 b).
In some gas exchange experiments an analysis was car- ried out to determine whether any of the observed stomatal responses to vapor pressure were consistent with optimiza- tion of carbon dioxide uptake with respect to water loss (Cowan and Farquhar 1977). A simplified procedure was used to calculate Oqw/~PhN (Farquhar et al. 1980). The sen- sitivity of transpiration rate to change in conductance was defined as
~q,v/~g~- AC
where AC is the humidity difference (mg H20 cm -3) be- tween the leaf and the air. This definition of ~qw/Og~ as- sumes that changes in conductance do not change leaf tem- perature significantly, which was the case in the thermo- electrically cooled assimilation chambers in which leaf tem- perature was held constant. The sensitivity of net photosyn- thesis to changes in conductance, ~Phs/~g~, was obtained from a plot of PhN versus gs (Fig. 7). The variations in gs were obtained by varying VPD. The slope of the curve of Ph N versus g~ at a given point gives ~Ph/NOg~ for that value of g~. Nonlinear regression lines were fitted through the data points by computer. The resulting regression equa- tion was differentiated with respect to g~ and was used to calculate ~PhN/~g~ at different values of g~
Results and Discussion
Response of Whole Plants to Changes in Ambient Vapor Pressure
Stomatal responsiveness to changes in vapor pressure var- ied considerably according to the age and developmental stage of the needles. The response of previous season nee- dles to a range of VPD during late March is seen in Fig. 1. Similar response patterns were observed during April and June measurements. Initially, there was no apparent decline in g~ as VPD was increased. There was considerable varia- tion in responsiveness ofg~ to VPD between saplings result- ing in the fairly large standard errors shown. Some saplings maintained a relatively constant gs over the entire range of VPD, while g~ in others declined with each increase in VPD. The effect of the relative insensitivity of gs to VPD in the previous season foliage can be seen in the responses of transpiration and xylem pressure potential to increased VPD (Fig. 1). Transpiration increased sharply and xylem pressure potential decreased by over 0.9 MPa over the range of VPD attained. From this information it is difficult to determine whether the decrease in conductance observed was the result of a direct stomatal response to humidity or a whole leaf negative feedback response to declining water potential.
The effect of increasing VPD on stomatal conductance of nearly fully expanded needles in June is shown in Fig. 2. It is apparent that the stomata of developing needles were much more responsive to changes in VPD than the stomata

239
0.105
0.090
0.075 T
0.060 E
0.045
0.030
0.015
0
0.6
1.0 c~
~ 1,L T
0
I I I I
I I I I
1.0 2.0 VPD (kPa)
1.4
1.2 ~- r03
1.0 T E ( i
0.8 c n E
0.6 T ,:::3
o,z, #
0.2
I
t T
3.0
Fig. 1. The effect of leaf to air vapor pressure difference (VPD) on stomatal conductance (gs;o), transpiration (qw;O) and xylem pressure potential (~up) of previous season needles during late March. All points are means. Vertical bars indicate standard errors
0.090
0.075
% 0.060
E 0.045
N 0.030
0,015
0
i
< I I I I I
6
5
4 E u
s ~ 2 o
l g
0.8
T ' I ' '. ' " 0 10 20 3.0
VPD (kPo)
Fig. 2. The effect of leaf to air vapor pressure difference (VPD) on stomatal conductance (g~;o), transpiration (qw;o) and xylem pressure potential (~%) of current season needles during June. All points are means, Vertical bars indicate standard errors
of previous year needles. In fact, above -~ 1.5 kPa VPD, the decline in gs of young needles was sufficient to maintain transpiration and thus xylem pressure potential constant (Fig. 2). Similar responses have been observed in Picea sit- chensis (Watts and Neilson 1978), Pinus banksiana (Bliss and Mayo 1980) and several nonconiferous species (Sheriff 1979). In some of the plants, transpiration was observed to decline and xylem pressure potential increase with in- creasing VPD (Fig. 3). These types of responses (Figs. 2 and 3) cannot be explained by negative feedback hypothesis involving sites of evaporation inside the leaf and passage of water through the stomatal pore, but rather correspond to feedforward control of stomatal response to humidity difference (Farquhar 1978).
By September, stomata of current foliage were found to be much less responsive to changes in VPD than in June. In contrast to the June experiments, transpiration was ob- served to increase over the entire range of VPD attained and xylem pressure potential showed a marked decline.
Similar declines in stomatal responsiveness to humidity changes, associated with maturation and ageing of foliage have been observed in Picea sitchensis (Neilson and Jarvis 1975) and P. engelmannii (Kaufmann 1979).
In view of the evidence that the stomata responded to ambient vapor pressure via direct external evaporation from the stomatal apparatus (Figs. 2 and 3), two possible mechanisms seemed most likely to explain the decline in stomatal responsiveness to humidity. One possibility was that water potential components of the needles, specifically osmotic potential at incipient plasmolysis, changed during the summer and that some of this change was reflected in the guard cells causing them to be less sensitive to decline in ambient humidity. The second possibility was that there may have been a change in the characteristics of the external cuticle covering the guard and associated cells.
In an effort to investigate the second possibility, the stomatal (lower) surface of needles was examined with a scanning electron microscope. The appearance of the exter- nal surface of current year needles sampled in June and September was very similar. There were substantial deposits of epicuticular wax on the surface of needles sampled in both months, although the deposits on the needles sampled in June appeared to be slightly less densely packed. The external appearance of the cuticle is not always a reliable indicator of its water permeability (Seymour/980), howev- er, and inner layers of cuticle synthesized later in the grow- ing season may have influenced the responsiveness of sto- mata to external humidity by decreasing the water perme- ability of the cuticle.
An additional plant growth room experiment was car- ried out to further examine the effect of the external cuticle on the stomatal response to humidity. Selected twigs were subjected to partial removal of the cuticle as described ear- lier. The effect of increasing VPD on stomatal conductance, transpiration and xylem pressure potential of hexane- treated versus untreated twigs is shown in Fig. 4. Although the sample size was small (3 saplings), differences in re- sponse were apparent. Without exception, stomatal conduc- tance of hexane-treated needles showed a greater percentage decrease than that of untreated needles. The result of this was that transpiration rates of hexane-treated needles at higher VPD's were actually lower than those of untreated needles. No differences in xylem pressure potential between the two treatments were detected.
Needle water potential components were estimated by means of pressure-volume curves to check the possibility that differences in stomatal responsiveness to humidity may have been caused in part by differences in leaf water poten- tial components. There were considerable differences in leaf water potential components depending upon needle age and the time of year (Fig. 5). For example, in the late spring, initial osmotic potential at full turgidity was - 1 . 7 MPa in previous year foliage and -0 .75 MPa in maturing new needles. The osmotic potential at incipient plasmolysis (tur- gor loss point) was about -2 .05 and -1 .50 MPa in the previous year and developing needles respectively. When pressure-volume curves were determined again in late sum- mer (Fig. 5 c and d), it was found that water potential com- ponents within each age class had changed from the late spring values and that there were still differences between age classes. In current season and previous year needles the turgor loss point had dropped by about 0.6 MPa to - 2 . 1 and - 2 . 6 MPa respectively. Similar seasonal changes
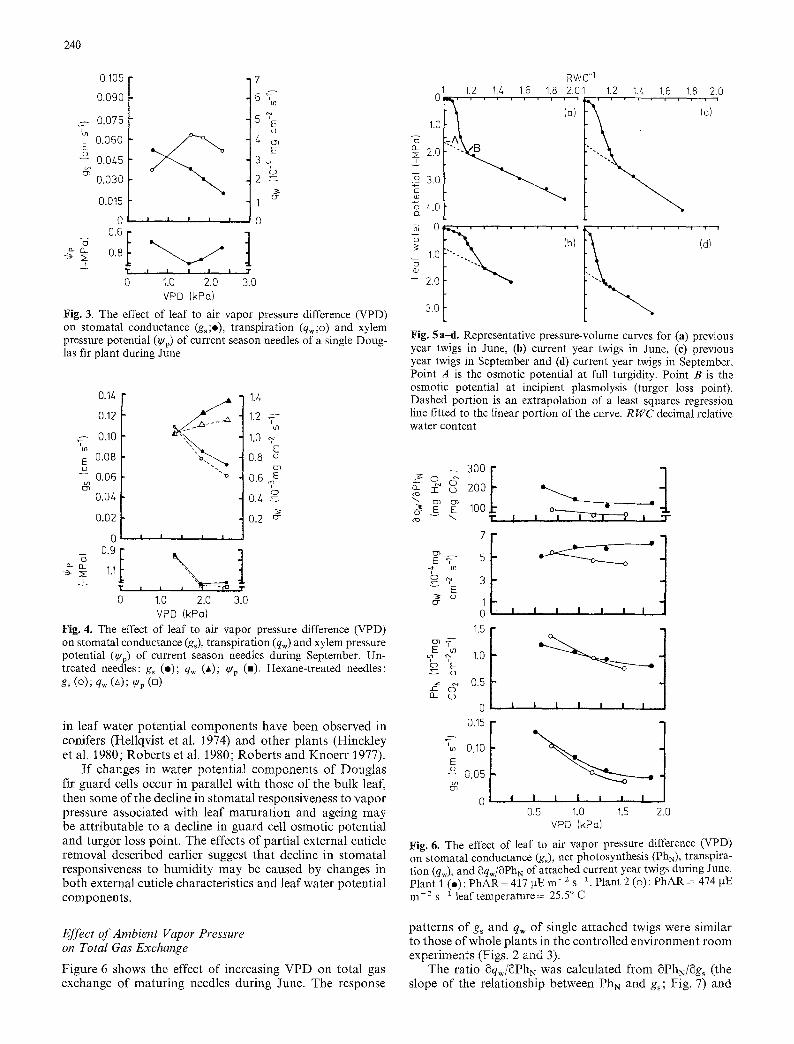
240
0,105 [
0.090
~.- 0.075 tth
0.060 E o
0.0/,5
0.030
0.012
0 "-5 0 . 6 [
ca 0.8
- - "1"
0
xk I I I I I
I i i i i
1.0 2.0 VPD (kPa)
7
6"7 tn e~
5 'E o
4 cn E
3 , , i
o 2
1
T 3.0
Fig. 3. The effect of leaf to air vapor pressure difference (VPD) on stomatal conductance (gs;e), transpiration (qw;O) and xylem pressure potential (~Up) of current season needles of a single Doug- las fir plant during June
0.1.4
0.12
0.10 v E 0.08 u
- - 0.06
0.0L
0.02
0 0.9
1.1
"I"
0
< / " J - ~"
1.4
1.2 ~-
1.0 E
0.8 u
0.6 f ~D
0.4
0.2
I I I I I
a- [3_
I I 1.0 2.0 3.0 VPD (kPo)
Fig. 4. The effect of leaf to air vapor pressure difference (VPD) on stomatal conductance (g~), transpiration (qw) and xylem pressure potential (~,p) of current season needles during September. Un- treated needles: g~ (e); q~ (A); ~tp (i). Hexane-treated needles: g~ (o); qw (zx); gtp (n)
in leaf water potent ia l components have been observed in conifers (Hellqvist et al. 1974) and other plants (Hinckley et al. 1980; Rober ts et al. 1980; Rober ts and Knoer r 1977).
I f changes in water potent ial components o f Douglas fir guard cells occur in parallel with those of the bulk leaf, then some of the decline in s tomatal responsiveness to vapor pressure associated with leaf ma tura t ion and ageing may be a t t r ibutable to a decline in guard cell osmotic potent ial and turgor loss point. The effects of par t ia l external cuticle removal described earlier suggest that decline in s tomatal responsiveness to humidi ty may be caused by changes in both external cuticle characteristics and leaf water potent ia l components .
Effect of Ambient Vapor Pressure on Total Gas Exchange
Figure 6 shows the effect of increasing VPD on total gas exchange of matur ing needles during June. The response
1.0
~: 2.0 i
~d 3.0 a)
o 4.0
0
1.0 (1)
2.(3
3.0
RWC-1 1,2 1.4 1,6 1,8 2.0
' ' ' ' ' ' '/a)' 1.2 1.L 1.6 1.8 2.0
(c)
' ' ' ib)" , , , , , , , i , ,
(d)
F i g . 5 a-& Representative pressure-volume curves for (a) previous year twigs in June, (b) current year twigs in June, (c) previous year twigs in September and (d) current year twigs in September. Point A is the osmotic potential at full turgidity. Point B is the osmotic potential at incipient plasmolysis (turgor loss point). Dashed portion is an extrapolation of a least squares regression line fitted to the linear portion of the curve. R WC decimal relative water content
_ 300 r 7
1 100 cr ~ | T (,~ ~ ~ I I
7 E 1 bq
v E N o
Cr ~ I I I I I I I
1.5 f u
Z c 7 o5 [3- r.D
0 I I I I i I I
0,10 E
0,05
0 I I I I i I I 0.5 1.0 1.5 2.0
VPD (kPa)
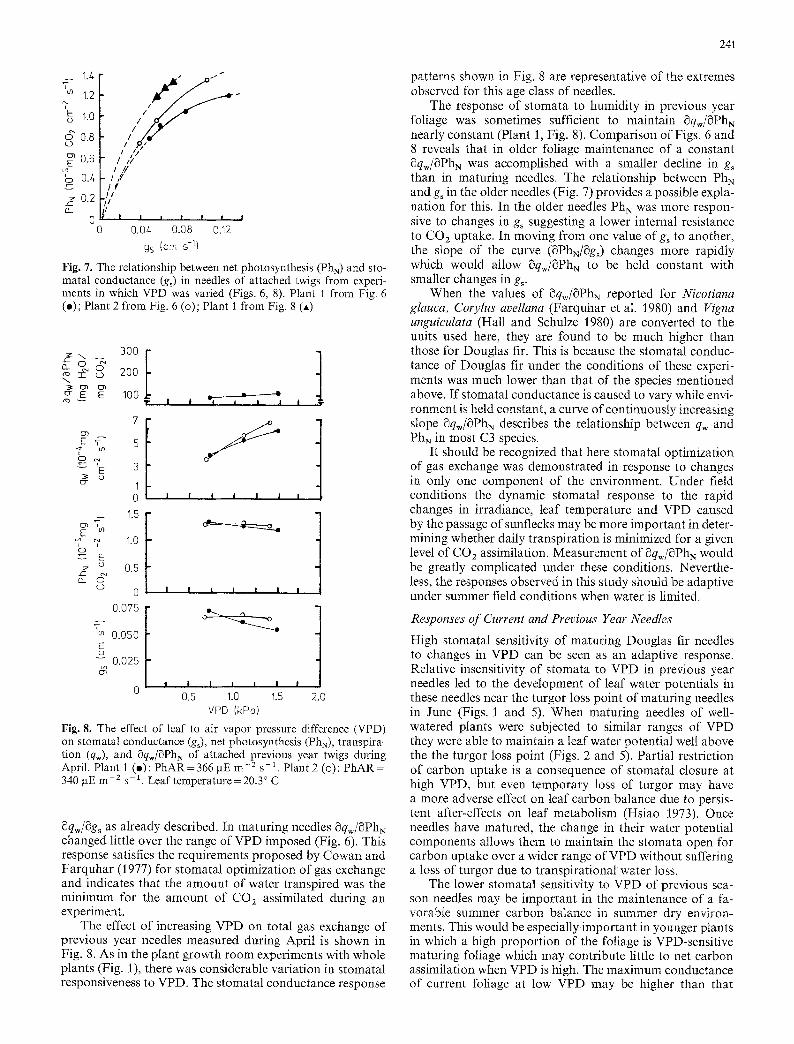
Fig. 6. The effect of leaf to air vapor pressure difference (VPD) on stomatal conductance (g~), net photosynthesis (PhN), transpira- tion (qw), and 8qw/~PhN of attached current year twigs during June. Plant 1 (o): PhAR=417 ~tE m -2 s -1. Plant 2 (o): PhAR= 474 gE m -2 s- ~ leaf temperature = 25.5 ~ C
pat terns of gs and qw of single at tached twigs were similar to those of whole plants in the controlled environment room experiments (Figs. 2 and 3).
The ratio ~qw/~PhN was calculated from ~PhN/Sgs (the slope of the relat ionship between PhN and g~ ; Fig. 7) and
241
1./* 'T
u~ 1.2 i
o E 1.0
cY 0.8 ~D
0.6 m
�89 0.4
0.2 x z
O_
@
/ f Ir II I
I I I I [ [ I
0.04 0.08 0.12
gs (cm s -1) Fig. 7. The relationship between net photosynthesis (PhN) and sto- matal conductance (g~) in needles of attached twigs from experi- ments in which VPD was varied (Figs. 6, 8). Plant 1 from Fig. 6 (o) ; Plant 2 from Fig. 6 (o); Plant 1 from Fig. 8 (A)
o s00! " 200 ' -r 0
o- E E I 00
E'- 7[ T 5 k
~E 3 3= O
u- 1 0
~_ 1.5 f 'I' '7 1.0 o
E Z o 0.5
o 0
~- 0.075 f '~ 0,050
- - 0 .025
0
I I I I I I
~ . ~
! l
I i I I I I I
t I i i i !
0.5 1.0 1.5 2.0
VPD (kPa)
Fig. 8. The effect of leaf to air vapor pressure difference (VPD) on stomatal conductance (gs), net photosynthesis (PhN) , transpira- tion (qw), and 8qw/SPhN of attached previous year twigs during April. Plant 1 (o): PhAR= 366 gEm -2 s -1. Plant 2 (o): PhAR= 340 gEm -2 s -1. Leaf temperature= 20.3 ~ C
8qw/Sgs as already described. In maturing needles 8qw/SPh N changed little over the range of VPD imposed (Fig. 6). This response satisfies the requirements proposed by Cowan and Farquhar (1977) for stomatal optimization of gas exchange and indicates that the amount of water transpired was the mininmm for the amount of CO2 assimilated during an experiment.
The effect of increasing VPD on total gas exchange of previous year needles measured during April is shown in Fig. 8. As in the plant growth room experiments with whole plants (Fig. 1), there was considerable variation in stomatal responsiveness to VPD. The stomatal conductance response
patterns shown in Fig. 8 are representative of the extremes observed for this age class of needles.
The response of stomata to humidity in previous year foliage was sometimes sufficient to maintain 8qw/SPh N nearly constant (Plant 1, Fig. 8). Comparison of Figs. 6 and 8 reveals that in older foliage maintenance of a constant 5qw/SPh N was accomplished with a smaller decline in gs than in maturing needles. The relationship between Ph N and gs in the older needles (Fig. 7) provides a possible expla- nation for this. In the older needles PhN was more respon- sive to changes in g~ suggesting a lower internal resistance to CO2 uptake. In moving from one value o f g S to another, the slope of the curve (SPhN/Sg~) changes more rapidly which would allow 8qw/SPh N to be held constant with smaller changes in g~.
When the values of 8qw/0PhN reported for Nicotiana glauca, Corylus avellana (Farquhar et al. 1980) and Vigna unguiculata (Hall and Schulze 1980) are converted to the units used here, they are found to be much higher than those for Douglas fir. This is because the stomatal conduc- tance of Douglas fir under the conditions of these experi- ments was much lower than that of the species mentioned above. I f stomatal conductance is caused to vary while envi- ronment is held constant, a curve of continuously increasing slope 8qw/SPh N describes the relationship between qw and PhN in most C3 species.
It should be recognized that here stomatal optimization of gas exchange was demonstrated in response to changes in only one component of the environment. Under field conditions the dynamic stomatal response to the rapid changes in irradiance, leaf temperature and VPD caused by the passage of sunflecks may be more important in deter- mining whether daily transpiration is minimized for a given level of CO2 assimilation. Measurement of 5qw/SPhN would be greatly complicated under these conditions. Neverthe- less, the responses observed in this study should be adaptive under summer field conditions when water is limited.
Responses of Current and Previous Year Needles
High stomatal sensitivity of maturing Douglas fir needles to changes in VPD can be seen as an adaptive response. Relative insensitivity of stomata to VPD in previous year needles led to the development of leaf water potentials in these needles near the turgot loss point of maturing needles in June (Figs. I and 5). When maturing needles of well- watered plants were subjected to similar ranges of VPD they were able to maintain a leaf water potential well above the the turgor loss point (Figs. 2 and 5). Partial restriction of carbon uptake is a consequence of stomatal closure at high VPD, but even temporary loss of turgor may have a more adverse effect on leaf carbon balance due to persis- tent after-effects on leaf metabolism (Hsiao 1973). Once needles have matured, the change in their water potential components allows them to maintain the stomata open for carbon uptake over a wider range of VPD without suffering a loss of turgor due to transpirational water loss.
The lower stomatal sensitivity to VPD of previous sea- son needles may be important in the maintenance of a fa- vorable summer carbon balance in summer dry environ- ments. This would be especially important in younger plants in which a high proportion of the foliage is VPD-sensitive maturing foliage which may contribute little to net carbon assimilation when VPD is high. The maximum conductance of current foliage at low VPD may be higher than that

242
of older foliage (Running 1975; Meinzer unpublished) but the significance of this in terms of its contr ibut ion to a positive carbon balance will depend on the photosynthetic capacity of matur ing needles at a particular developmental stage. On clear summer days in the field, higher stomatal conductance in current foliage generally occurs only in the early morning hours when both VPD and irradiance are low (Meinzer, unpublished observations). In mature indi- viduals of Douglas fir and other evergreen conifers several age classes of foliage form a significant port ion of the total leaf surface area and it is necessary to better characterize differences in stomatal behavior resulting from needle age and developmental stage.
Acknowledgements. I wouid like to thank Dr. R.B. Walker for his continued support during the study and Dr. L.C. Bliss for his comments on the manuscript. Funds for construction of the steady-state porometer were provided by the University of Wash- ington Graduate School Research Fund.
References
Beardsell MF, Jarvis PG, Davidson B (1972) A null-balance diffu- sion porometer suitable for use with leaves of many shapes. J Appl Ecol 9: 677-690
Bingham GE, Coyne PI (1977) A portable, temperature-controlled, steady-state porometer for field measurements of transpiration and photosynthesis. Photosynthetica 11 : 148-160
Bliss LC, Mayo JM (1980) An ecophysiological investigation of the jackpine woodland with reference to revegetation of mined sands. Alberta Oil Sands Env Res Program VE 6.1
Cowan IR (1977) Stomatal behavior and environment. Adv Bot Res 4:117-227
Cowan IR, Farquhar GD (1977) Stomatal function in relation to leaf metabolism and environment. Syrup Soc Exp Biol 31:471-505
Edwards ME, Meidner H (1978) Stomatal responses to humidity and water potentials of epidermal and mesophyll tissue. J Exp Bot 29 : 771-780
Farquhar GD (1978) Feedforward responses of stomata to humidi- ty. Aust J Plant Physiol 5 : 787-800
Farquhar CD, Schulze ED, Ktippers M (1980) Responses to hu- midity by stomata of Nicotiana glauca L. and Corylus avellana L. are consistent with the optimization of carbon dioxide up- take with respect to water loss. Aust J Plant Physiol 7:315-327
Grace J, Malcom DC, Bradbury IK (1975) The effect of wind and humidity on leaf diffusive resistance in Sitka spruce seed- lings. J Appl Ecol 12:931-940
Hall AE, Hoffmann GJ (1976) Leaf conductance response to hu- midity and water transport in plants. Agronomy J 68 : 876-881
Hall AE, Schulze ED (1980) Stomatal response to environment and a possible interrelation between stomatal effects on trans- piration and CO2 assimilation. Plant, Cell and Env 3:467-474
Hellqvist J, Richards GP, Jarvis PG (1974) Vertical gradients of water potential and tissue water relations in Sitka spruce trees measured with the pressure chamber. J Appl Ecol 11:637-667
Hinckley TM, Duhme F, Hinckley AR, Richter H (1980) Water relations of drought hardy shrubs : Osmotic potential and sto- matal reactivity. Plant, Cell and Env 3:131-140
Hsiao TC (1973) Plant responses to water stress. Ann Rev Plant Physiol 24:519 570
Kaufmann MR (1976) Stomatal response of Engelmann spruce to humidity, light, and water stress. Plant Physiol 57 : 898-901
Kaufmann MR (1979) Stomatal control and the development of water deficit in Engelmann Spruce seedlings during drought. Can J For Res 9:297 304
Lange OL, L6sch R, Schulz ED, Kappen L (1971) Responses of stomata to changes in humidity. Planta 100:76-86
Lange OL, Medina E (1979) Stomata of the CAM plant Tillandsia recurvata respond directly to humidity. Oecologia 40:352363
Ludlow MM, Ibaraki K (1979) Stomatal control of water loss in siratro (Macroptilium atropurpureum (DC) Urb.), a tropical pasture legume. Ann Bot 43 : 63%647
Meidner H (1975) Water supply, evaporation, and vapor diffusion in leaves. J Exp Bot 26 : 666-673
Meidner H (1976a) Vapor loss through stomatal pores with the mesophyll tissue excluded. J Exp Bot 27:17~174
Meidner H (1976b) Water vapor loss from a physical model of a substomatal cavity. J Exp Bot 27:691-694
Meinzer FC (1981) Water use efficiency in Douglas fir: The nature of plant response to selected environmental variables. Seattle WA, PhD Thesis, University of Washington
Meinzer FC (1982) The effect of light on stomatal control of gas exchange in Douglas fir (Pseudotsuga menziesii) saplings. Oeco- 1ogia (in press)
Neilson RE, Jarvis PG (1975) Photosynthesis in Sitka spruce (Picea sitchensis (Bong.) Carr.) VI. Response of stomata to tempera- ture. J Appl Ecol 12:879-891
Pallardy SG, Koztowski TT (1979) Stomatal response of Populus clones to light intensity and vapor pressure deficit. Plant Physiol 64:112-114
Rawson HM, Woodward RG (1976) Photosynthesis and transpira- tion in dicotyledonous plants I. Expanding leaves of tobacco and sunflower. Aust J Plant Physiol 3 : 247-256
Roberts SW, Knoerr KR (1977) Components of water potential estimated from xylem pressure measurements. Oecologia 28:191-202
Roberts SW, Strain BR, Knoerr KR (1980) Seasonal patterns of leaf water relations in four co-occurring forest tree species: Parameters from pressure-volume curves. Oecologia 46:330 337
Running SW (1976) Environmental control of leaf water conduc- tance in conifers. Can J For Res 6:104~112
Salo, DJ (1974) Factors affecting photosynthesis in Douglas fir. Seattle WA, PhD Thesis, University of Washington
Schulze ED, Lange OL, Buchsbom U, Kappen L, Evenari M (1972) Stomatal responses to changes in humidity in plants growing in the desert. Planta 108:259 270
Schulze ED, Lange OL, Evenari M, Kappen L, Buchsbom U (1975) The role of air humidity and temperature in controlling stoma- tal resistance of Prunus armeniaca L. under desert conditions III. The effect on water use efficiency. Oecologia 19:303-314
Seybold A (1961/62) Ergebnisse und Probleme pflanzlicher Trans- pirationsanalysen. Jh Heidelberger Akad Wiss 6: 5-8
Seymour VA (1980) Leaf cuticle: An investigation of some physical and chemical properties derived from a study of Berberis. Seattle WA, PhD Thesis, University of Washington
Sheriff DW (1979) Stomatal aperture and the sensing of environ- ment by guard cells. Plant, Cell and Env 2:15-22
Sheriff DW (1977) The effect of humidity on water uptake by, and viscous flow resistance of, excised leaves of a number of species: Physiological and anatomical observations. J Exp Bot 28 : 1399-1407
Tan CS, Black TA, Nnymah JU (1977) Characteristics of stomatal diffusion resistance in a Douglas fir forest exposed to soil water deficits. Can J For Res 7 : 595-604
Tyree MT, Hammel HT (1972) The measurement of the turgor pressure and the water relations of plants by the pressure bomb technique. J Exp Bot 23:267-282
Tyree MT, Yianoulis P (1980) The site of water evaporation from sub-stomatal cavities, liquid path resistances, and hydroactive stomatal closure. Ann Bot 46:176-193
Watts WR, Neilson RE (1978) Photosynthesis in Sitka spruce (Pi- cea sitchensis (Bong.) Carr.) VIII. Measurements of stomatal conductance and 14CO2 uptake in controlled environments. J Appl Ecol 15:245-255
Whiteman PC, Koller D (1964) Environmental control of photo- synthesis and transpiration in Pinus halepensis. Israel J Bot 13:166-176
Received September 19, 1981






![Fundy Model Forest · Douglas fir ( Pseudotsuga menziesii [Mirb.] Franco) region of the Pacific Northwest , and noted that seedling establishment and growth were seriously retarded](https://img.dokumen.tips/doc/110x75/5f5dea41236b8e60de46ba44/fundy-model-forest-douglas-fir-pseudotsuga-menziesii-mirb-franco-region-of.jpg)






















