Embed Size (px)
Citation preview

Available online at www.sciencedirect.com
nology 149 (2008) 29–49www.elsevier.com/locate/revpalbo
Review of Palaeobotany and Paly
The diversity of the Carboniferous phytoplankton
G.L. Mullins a,⁎, T. Servais b
a Department of Geology, University of Leicester, University Road, Leicester LE1 7RH, United Kingdomb Laboratoire de Paléontologie et Paléogéographie du Paléozoïque (LP3), UMR 8014 du CNRS, USTL, SN5, F-59655 Villeneuve-d’Ascq Cedex, France
Received 6 March 2007; received in revised form 9 October 2007; accepted 18 October 2007Available online 30 October 2007
Abstract
Here we present a synthesis of the published literature on in situ Carboniferous acritarchs, prasinophyte algae and associated microfossils. TheEarly Carboniferous had a rich acritarch fossil record, with assemblages documented in both equatorial and high latitude regions. However, thedata suggest that there was no relationship between acritarch diversity and palaeolatitude in the Early Carboniferous. Our synthesis reveals a trendof declining acritarch diversity from the Tournaisian to the Visean, and generally very low diversity in the latest Early Carboniferous Serpukhovianand Late Carboniferous. However, fairly diverse acritarch assemblages of up to 13 taxa have been described from the Late Carboniferous ofEngland and South America. They imply that the very low diversity so far documented could result from a lack of research input. However, theseassemblages need to be re-examined as they can be based on a limited number of individuals and taxonomic over-splitting is a possibility.Furthermore, the reworking of older assemblages into these Late Carboniferous strata needs to be thoroughly discounted. The data also suggests atrend of declining prasinophyte diversity through the Carboniferous. However, the very low number of taxa typically reported and their groupingunder open nomenclature (e.g. Cymatiosphaera spp.) makes this trend of questionable robustness. No clear pattern was observed in the diversityof the Zygnemataceae, although more diverse assemblages could be related to increased terrestrial influence or continental deposition. The drop inacritarch diversity in the Early Carboniferous represents a continuation of a decline that began in the Late Devonian. Although a number ofhypotheses have been invoked to account for this Devonian–Carboniferous diversity decline, our synthesis has shown that the phytoplankton fromthis interval are generally poorly studied, with only a handful of investigations having been completed in stratigraphically continuous sequences.Further work is urgently needed if we are to evaluate these hypotheses and understand this fundamental change in the nature of the ocean’sprimary producers.© 2007 Elsevier B.V. All rights reserved.
Keywords: Carboniferous; diversity; acritarchs; Prasinophyceae; Hydrodictyaceae; Zygnemataceae
1. Introduction
From the Cambrian to the Devonian, the acritarchs andprasinophyte algae dominated the marine, organic-walled,microfossil record. The biological affinities of the acritarchsremain poorly understood, but a dinoflagellate, or ancestraldinoflagellate affinity has been suggested on the basis of grossmorphological similarity, comparable patterns of distributionand the co-occurrence of biomarkers (e.g. Moldowan et al.,1996; Moldowan and Talyzina, 1998; Talyzina and Moczy-dlowska, 2000; Servais et al., 2004). The majority of acritarchs
⁎ Corresponding author. Fugro Robertson Limited, Llandudno LL30 1SA,United Kingdom.
E-mail address: [email protected] (G.L. Mullins).
0034-6667/$ - see front matter © 2007 Elsevier B.V. All rights reserved.doi:10.1016/j.revpalbo.2007.10.002
are considered to have been marine, although some aresuggested to be non-marine (e.g. Arabisphaera, Mehlisphaeri-dium). Prasinophyte microfossils represent the reproductivephycomata of green algae with a geological record that extendsfrom the Precambrian to Recent (e.g. Martin, 1993; Colbath andGrenfell, 1995). Furthermore, it is only the modern marine taxathat produce fossilizable phycomata. Some debate surrounds theaffinity of some Carboniferous organic-walled microfossils. Wefollow Guy-Ohlson (1996) and Stephenson et al. (2004) inconsidering Carbaneuletes to be a possible prasinophyte.However, other genera commonly regarded as prasinophytes(e.g. Duvernaysphaera, Leiosphaeridia, Polyedryxium, Qua-draditum) are here placed in the acritarchs (see Colbath andGrenfell, 1995 regarding the affinity Leiosphaeridia). Theacritarchs and prasinophyte algae are often referred to as the“Palaeozoic phytoplankton”, and their analysis has been shown

30 G.L. Mullins, T. Servais / Review of Palaeobotany and Palynology 149 (2008) 29–49
to be important in biostratigraphical, palaeoenvironmentaland palaeobiogeographical studies (e.g. Servais et al., 2003;Mullins et al., 2004; Stricanne et al., 2004, 2006). However, incomparison with the Early Palaeozoic, relatively little is knownabout their diversity and geographical distribution in the LatePalaeozoic.
Our ability to recognize the fossilized remains of theZygnemataceae has improved over the last 25 years (seeColbath and Grenfell, 1995; Grenfell, 1995; van Geel andGrenfell, 1996). These filamentous green algae, which areexclusively freshwater, may produce fossilizable, acid-resistant,zygospores as a part of their sexual reproduction. We havefollowed Grenfell (1995, Table 1) when assigning our fossiltaxa to the zygnemataleans, which have a geological recordfrom the Ordovician to Recent. The colonial Hydrodictyaceaeare also exclusively freshwater, with the walls of the vegetativecell containing a fossilizable, sporopollenin-like material(Colbath and Grenfell, 1995; Batten, 1996). The colonial,brackish or freshwater, alga Botryococcus has also been widelyreported from Carboniferous strata. Although it is beyond thescope of this paper to fully document all of the occurrences ofthis alga, several reports are acknowledged below.
The Carboniferous has been recognized as an importantinterval in Earth’s history. It is marked by a significant changein the distribution of the continents, as Laurussia and Gondwanafused to form Pangaea, and by important palaeoclimaticfluctuations, with significant glacial episodes (Fig. 1; e.g.Veevers and Powell, 1987; Crowell, 1999; Torsvik and Cocks,2004). It is also marked by important biotic events, such as thediversification of the land plants (e.g. Willis and McElwain,2002) and a significant number of palynological studies have
Fig. 1. Palaeogeographical reconstruction for the Early Carboniferous (340 Ma) andthe Late Carboniferous represents glacial deposits. Figure after Torsvik and Cocks,
been completed on Carboniferous spores and pollen. Incomparison, relatively little is known about the acritarchs,prasinophytes and other algal microfossils through this interval.Indeed, it seems to be a common assumption that they are not tobe found, or that their scarcity makes them of little practicalvalue, and thus they are often overlooked or ignored. Here wepresent a compilation of ninety-two publications that havedocumented in situ (i.e. not reworked) microfossils ofacritarchs, Prasinophyceae, Zygnemataceae and Hydrodictya-ceae in Carboniferous strata and we have used these data toassess their diversity through this interval.
The historical basis of the subdivisions of the Carboniferouswere described by Davydov et al. (2004), who documented thefoundation of the traditional Russian, Western European andNorth American classifications; the subdivisions in these areasare summarized on Fig. 2. A global Carboniferous timescalerepresenting a fusion of the three schemes has developed since1978 and the Carboniferous is now subdivided into theMississippian and Pennsylvanian subperiods, and in to sevenstages (Fig. 2; Davydov et al., 2004). We have employed theInternational Commission on Stratigraphy (ICS) subdivisionshere, but in some instances problems have arisen through aninability to accurately determine the exact stratigraphical posi-tion of the studied collections; for example, a flora identified as‘Namurian’ can only be correlated with either the Serpukhovianand/or Bashkirian stages of the ICS scheme (Fig. 2).
2. The Carboniferous algal microfossil record
Ninety-two studies have described acritarchs and prasino-phytes, together with Hydrodictyaceae and Zygnemataceae,
Late Carboniferous (310 Ma) times. The pale area in the southern hemisphere in2004.
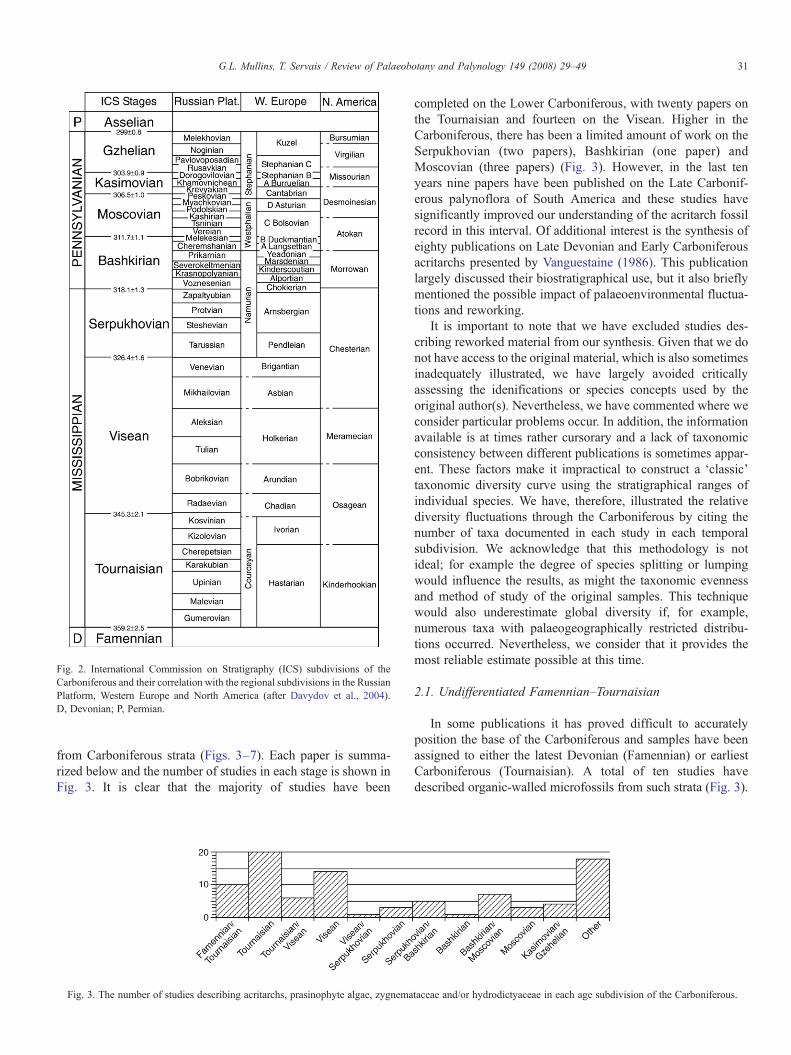
Fig. 2. International Commission on Stratigraphy (ICS) subdivisions of theCarboniferous and their correlation with the regional subdivisions in the RussianPlatform, Western Europe and North America (after Davydov et al., 2004).D, Devonian; P, Permian.
31G.L. Mullins, T. Servais / Review of Palaeobotany and Palynology 149 (2008) 29–49
from Carboniferous strata (Figs. 3–7). Each paper is summa-rized below and the number of studies in each stage is shown inFig. 3. It is clear that the majority of studies have been
Fig. 3. The number of studies describing acritarchs, prasinophyte algae, zygnema
completed on the Lower Carboniferous, with twenty papers onthe Tournaisian and fourteen on the Visean. Higher in theCarboniferous, there has been a limited amount of work on theSerpukhovian (two papers), Bashkirian (one paper) andMoscovian (three papers) (Fig. 3). However, in the last tenyears nine papers have been published on the Late Carbonif-erous palynoflora of South America and these studies havesignificantly improved our understanding of the acritarch fossilrecord in this interval. Of additional interest is the synthesis ofeighty publications on Late Devonian and Early Carboniferousacritarchs presented by Vanguestaine (1986). This publicationlargely discussed their biostratigraphical use, but it also brieflymentioned the possible impact of palaeoenvironmental fluctua-tions and reworking.
It is important to note that we have excluded studies des-cribing reworked material from our synthesis. Given that we donot have access to the original material, which is also sometimesinadequately illustrated, we have largely avoided criticallyassessing the idenifications or species concepts used by theoriginal author(s). Nevertheless, we have commented where weconsider particular problems occur. In addition, the informationavailable is at times rather cursorary and a lack of taxonomicconsistency between different publications is sometimes appar-ent. These factors make it impractical to construct a ‘classic’taxonomic diversity curve using the stratigraphical ranges ofindividual species. We have, therefore, illustrated the relativediversity fluctuations through the Carboniferous by citing thenumber of taxa documented in each study in each temporalsubdivision. We acknowledge that this methodology is notideal; for example the degree of species splitting or lumpingwould influence the results, as might the taxonomic evennessand method of study of the original samples. This techniquewould also underestimate global diversity if, for example,numerous taxa with palaeogeographically restricted distribu-tions occurred. Nevertheless, we consider that it provides themost reliable estimate possible at this time.
2.1. Undifferentiated Famennian–Tournaisian
In some publications it has proved difficult to accuratelyposition the base of the Carboniferous and samples have beenassigned to either the latest Devonian (Famennian) or earliestCarboniferous (Tournaisian). A total of ten studies havedescribed organic-walled microfossils from such strata (Fig. 3).
taceae and/or hydrodictyaceae in each age subdivision of the Carboniferous.

32 G.L. Mullins, T. Servais / Review of Palaeobotany and Palynology 149 (2008) 29–49
In North America, Boneham (1970) reported the occurrenceof the Neogene acritarch Leiosphaeridia plicata Felix (presum-ably a misidentification) and the new species Leiosphaeridialinebacki from the Famennian-early Tournaisian Clegg CreekMember of the New Albany Shale, southern Illinois. However,Boneham (1970) only described the leiospherids and tasmani-tids and it is probable that this only represents the palynoflora at
Fig. 4. A summary of the acritarchs cited to occur in the Carboniferous with the total nto date genus and species combination and might not be the name used in the originalBashkirian; Mosc., Moscovian; Kas., Kasimovian; Gzh., Gzhelian.
certain levels and does not accurately reflect its true com-position. Similarly, the prasinophyte alga Tasmanites noremiEisenack was the only taxon reported from the WoodfordFormation of Pontotoc County, Oklahoma by Eisenack (1962).The sampled strata were originally referred to the Mississippianby Eisenack (1962), but subsequent studies have regarded theWoodford Shale to span the Devonian–Carboniferous boundary
umber of taxa described in each publication. The taxon name given is the most uppublication. Fam., Famennian; Tourn., Tournaisian; Serp., Serpukhovian; Bash.,

Fig. 4 (continued ).
33G.L. Mullins, T. Servais / Review of Palaeobotany and Palynology 149 (2008) 29–49
(e.g. Over, 1992). However, diverse palynofloras are knownfrom the latest Devonian or earliest Carboniferous of NorthAmerica. In Saskatchewan, Canada, sixteen samples from the
undifferentiated Famennian/Tournaisian part of the MiddleSandstone Member of the Bakken Formation yielded a total oftwenty-three acritarchs and three prasinophytes (Playford and

Fig. 4 (continued ).
34 G.L. Mullins, T. Servais / Review of Palaeobotany and Palynology 149 (2008) 29–49
McGregor, 1993, Fig. 4). Thus, this study documents the mostdiverse acritarch and prasinophyte assemblages recorded fromthese equivocal strata worldwide. In addition, the study of
Playford and McGregor (1993) is one of the few phytoplanktonstudies to have examined Devonian–Carboniferous boundary insome detail.

Fig. 4 (continued ).
35G.L. Mullins, T. Servais / Review of Palaeobotany and Palynology 149 (2008) 29–49

Fig. 5. A summary of fhe prasinophyte algae cited to occur in the Carboniferous with the total number of taxa described in each publication. The taxon name given isthe most up to date genus and species combination and might not be the name used in the original publication.
36 G.L. Mullins, T. Servais / Review of Palaeobotany and Palynology 149 (2008) 29–49
Famennian or Tournaisian strata are also known from theeastern Elburz region of northeast Iran, where Coquel et al.(1977) examined the Khosh–Yeilagh Formation. The prasino-phyte Tasmanites spp. (abundant at some levels) and acritarchsGorgonisphaeridium spp., Micrhystridium albertensis Staplin,Stellinium spp. and Veryhachium nasicum (Stockmans andWillière) were reported. It should also be noted that the colonialalga Botryococcus was reported from the younger ViseanMobarak Formation and Cheseghaleh Formation (Coquel et al.,1977).
The acritarchs and prasinophytes of the Fairfield Formation,Canning Basin, Western Australia, were described by Playford(1976). A total of thirty-three samples from four boreholes(BMR 2 Laurel Downs, BMR Lennard River 2, BMRNoonkanbah 4 and Pickands Mather AH 2) were examined,but only eight samples yielded a total of two prasinophyte andnine acritarch taxa. The exact age of the Fairfield Formation isuncertain, however, and they have been assigned to undiffer-entiated Upper Devonian and lower Tournaisian, with theconformably overlying Laurel Formation containing middleTournaisian endothyrids (see Playford, 1976; Edgell, 2004).
In Morocco, Rahmani-Antari (1980) reported the occurrenceof Veryhachium and leiospheres in the Late Devonian/earlyTournaisian (Tn1b) sample BU408 from the Borj d’Erfoud. The
Late Devonian/early or mid Tournaisian part of the DOT 1 well(from a depth of 2586 m) of the Doukkala Basin, central-westMorocco, has also been examined and the occurrence of fiveacritarchs was reported by Rahmani-Antari (1990). Further-more, Rahmani-Antari and Lachkar (2001) defined twopalynozones (ST1 and ST2) in undifferentiated Strunian (i.e.latest Famennian) or Tournaisian strata from four Moroccancores (EBA2, DOT1, MKL102, MKL101) and an outcropsection (BU). The oldest palynozone ST1 yielded eight taxaincluding Maranhites brasiliensis Brito and Maranhites britoiStockmans and Willière and the zygnematalean Horologinella(now Tetraporina) horologia (Staplin). The younger palyno-zone ST2 contained only three species (Rahmani-Antari andLachkar, 2001). According to Rahmani-Antari and Lachkar(2001) acritarchs also occur in the unequivocal Carboniferousstrata of Morocco, but they were in a poor state of preservation.
In Europe, two studies by Turnau (1975) and Stempien-Sałek(1997) described the acritarchs from undifferentiated Famen-nian or Tournaisian strata in Poland. Turnau (1975) examinedthe Famennian and lowermost Tournaisian of the Babilon 1 andRzeczenica 1 boreholes, near Rzeczenica, northern Poland, andreported the occurrence of ten acritarchs and one prasinophyte.Subsequently, Stempien-Sałek (1997) described seven acri-tarchs from strata assigned to the Ra to Ma miospore biozones,

37G.L. Mullins, T. Servais / Review of Palaeobotany and Palynology 149 (2008) 29–49
equivalent to the upper expansa to sandbergi conodontbiozones of the middle Famennian to Tournasian (Tn2), inboreholes in the Kolobrzeg region of Western Pomerania.
2.2. Tournaisian
The Tournaisian contains the most diverse assemblages ofacritarchs known from the Carboniferous. The Yali and Naximformations of Nyalam, Tibet, have yielded the most diverseassemblages, with thirty-six acritarchs and seven prasinophytes(Gao, 1986). The Yali Formation was originally assigned to theTournaisian Vallatisporites vallatus–Aneurospora incohatus(VI) and Cingulizonates bialatus–Auroraspora macra (BM)miospore zones, while the Naxim Formation was determined asbelonging to the Visean Lycospora pusilla (PU) miospore Zone(Gao, 1983, 1984a,b, 1986). However, Zhang and Jin (1976;according to Zhong-Qiang, 2004) studied the brachiopods of thelower Naxim Group in Nyalam County and assigned them to theTournaisian Fusella–Ovatia Assemblage, with the brachiopodsfrom the middle NaximGroup being assigned to the lower ViseanMarginirugus–Syringothyris Assemblage and the upper ViseanDowhatania–Balakhonia Assemblage. Thus, the age determina-tions derived from the brachiopods are significantly differentto those from the sporomorphs. We consider both the Yali
Fig. 6. A summary of fhe zygnemataceae algae cited to occur in the Carboniferous witthe most up to date genus and species combination and might not be the name used
and Naxim formations to be of Tournaisian age, as did Li et al.(2002).
Tournaisian assemblages containing more than 10 species ofacritarchs and/or prasinophytes have also been reported fromBelgium (Stockmans and Willière, 1966a,b), Wales (McNestry,1988), Canada (Playford and McGregor, 1993), Bolivia (Vavr-dová et al., 1993), Spain (Lachkar et al., 1993) and Poland(Filipiak, 2005).
In Belgium, Stockmans and Willière (1966a) describedan assemblage of 17 acritarchs and 1 prasinophyte from 10samples between 212 and 216.5 m in the sondage de l’Asiled’aliénés [the lunatic asylum borehole], Tournai. These levelswere originally assigned to the Tournaisian (Tn1a, i.e. Strunian)and then subsequently to the “schists of Spiriferellina peracuta,that is to say in Tournaisien Tn2a”, by R. Legrand and GMortelmans (pers. comm. in Stockmans and Willière, 1966a).However, N. Hughes (pers. comm. in Stockmans and Willière,1966a) stated that the assemblage of small sporomorphs in thesamples had similarities with those of S.N. Naumova (noreference cited) from the Famennian I. The samples have beenreferred to the lower part of the Tournaisian here. Themajority oftaxa were referred to species of Micrhystridium and Veryha-chium, but specimens of Baltisphaeridium, Cymatiosphaera,Polyedryxium, Solisphaeridium and Tornaciawere also reported
h the total number of taxa described in each publication. The taxon name given isin the original publication.

Fig. 6 (continued ).
Fig. 7. A summary of fhe hydrodictyaceae algae cited to occur in theCarboniferous with the total number of taxa described in each publication. Thetaxon name given is the most up to date genus and species combination andmight not be the name used in the original publication.
38 G.L. Mullins, T. Servais / Review of Palaeobotany and Palynology 149 (2008) 29–49
(Stockmans and Willière, 1966a). Fifteen acritarchs, againprimarily of Micrhystridium and Veryhachium, but with singleSolisphaeridium, Unellium and Cymatiosphaera, were reportedfrom between 573.5 and 589.2 m in the sondage de Vieux Leuze,Hainaut, Belgium (Stockmans and Willière, 1966b). Thesampled levels were attributed to the Tournaisian (Tn2a) by R.Legrand and G. Mortelmans (pers. comm. in Stockmans andWillière, 1966b). However, it is important to note thatVanguestaine (1986) suggested that reworking of older stratawas responsible for the acritarch occurrences in the Tournai andLeuze boreholes of Belgium.
The Lower Limestone Shale Group of the Ewenny and Cwrt-yr-ala boreholes of south Wales, which was originallyconsidered to be of Courceyan age, was examined by McNestry(1988). The strata, of a marine shelf facies, yielded a total oftwenty-five acritarchs and two prasinophytes, in addition to thecolonial alga Botryococcus spp.
Several studies have been undertaken on Canadian Tournai-sian strata and some of these have been directed towardsunderstanding the generation of the oil reserves in this area. Asummary is given here, beginning with the diverse assemblagesrecovered from the Bakken Formation. The Upper ShaleMember of the Bakken Formation of southern Saskatchewanhas yielded ten acritarchs, which represents a significant declinefrom the twenty-seven taxa reported from the underlying
Famennian/Tournaisian Middle Sandstone Member of theBakken Formation (Playford and McGregor, 1993; seeabove). The age of the Upper Shale Member on the basis ofmiospores (assuming no reworking) is Famennian, butconodont data indicate that the Upper Shale Member correlateswith the early, but not earliest, Tournaisian Siphonodellasandbergi and Lower crenulata conodont zones (Playford andMcGregor, 1993, Fig. 4, 11). Additional occurrences in theBakken Formation at the level of genus or morphotype (e.g.

39G.L. Mullins, T. Servais / Review of Palaeobotany and Palynology 149 (2008) 29–49
spiny acanthomorphs) were given in Stasiuk (1996; age given asKinderhookian) and Stasiuk and Fowler (2004); the later alsoexamined the Exshaw Formation. The only other Canadianrecord is that of the type of Gorgonisphaeridium winslowiae inthe Banff Formation (see Staplin et al., 1965).
An early to mid Tournaisian assemblage of eleven acritarchsand one prasinophyte in three samples from the MontalbonAnticline, Teruel, c. 70 km SE of Zaragoza, Spain, was des-cribed by Lachkar et al. (1993). The assemblage was dated byusing the palynomorphs, which were highly thermally altered,but in a generally good state of preservation.
Vavrdová et al. (1993) presented the preliminary results of astudy of the uppermost Colpacucho and lowermost Cumanaformations of the Devonian–Carboniferous boundary in theLake Titikaka region of Bolivia. Miospores indicative of theTournaisian VI miospore Zone were recognized and a total offourteen taxa (comprising 2 prasinophytes and 12 acritarchs)were reported (Vavrdová et al., 1993).
The Devonian–Carboniferous boundary in the Holy CrossMountains of central Poland, was examined in some detail byFilipiak (2005), who noted assemblages of thirteen acritarchsand four prasinophytes in the Tournaisian parts of the Radin andZareby beds. These assemblages are similar to those of theFamennian, but are less diversified with Micrhystridium beingcommon, and generally comprise smaller morphotypes (Fili-piak, 2005). Furthermore, Filipiak (2005) also noted thatassemblages from the Tournaisian were commonly dominatedby a single acritarch genus, with representatives of the poly-gonomorphs and acanthomorphs being particularly common,and that these taxonomic differences between the Famennianand Tournaisian reflects palaeoenvironmental change.
Several other publications have described Tournaisianacritarchs, but all of these documented fewer than ten taxa. InEurope, Higgs et al. (2002) reported Leiosphaeridia and Cy-matiosphaera from the Tournaisian part of the KöprülüFormation, Hakkan Province of Turkey. A low diversity floraof six taxa (2 prasinophytes and 4 acritarchs) was alsodocumented from the Formation l’Huisserie near Beudmontpied de Boeuf, France (Houlgatte et al., 1988). The new taxonBiannulatisphaerities simplex, which may be fungal or algalin affinity, was described from the CM-VF concurrent miosporerange zones (i.e. upper Tournaisian to upper Visean) of Scotlandby Neville in Neves et al. (1973). The brackish-water BallaganFormation of the Midland Valley of Scotland has yielded theacritarch Leiosphaeridia, ?Carbaneuletes sp. A (a possibleprasinophyte), a number of indeterminable algal fossilsdescribed under an informal or open nomenclature, zygnema-taleans and Botryococcus spp. Elsewhere in Europe, the VImiospore Zone of the Late Devonian–Early CarboniferousTercemas and Bordalete formations of the north and south PedraRuiva and Monte do Penedo sections, SW Portugal, wasexamined by Pereira et al. (1996), who reported the occurrenceof five acritarchs and two prasinophytes.
In Africa, a total of five Tournaisian acritarchs and twoprasinophytes, in addition to Horologinella (now Tetraporina)horologia (Staplin), were documented from the IssendielFormation of the Illizi Basin, Algeria (Attar et al., 1980). In
South America, Maranhites brasiliensis Brito was cited asoccurring in the Curua, Langa and Poti formations of the Amazonand Parnaíba basins, Brazil (Daemon, 1974), andMicrhystridiumsp. and Veryhachium spp. both occur in the Formación Zarritas ofSierra de Almeida, Chile (Rubinstein et al., 1996).
2.3. Undifferentiated Tournaisian/Visean
Several publications have described acritarchs from strata thatcannot be accurately determined as either Tournaisian or Visean.Twelve acritarchs and one prasinophyte have been reported fromthe late Tournaisian to early Visean CV (Cordviosporites–Ver-rucosporites) miospore Biozone of central and western Argentina(Césari and Gutiérrez, 2001, p. 133). Other data from SouthAmerica includes that of Dueñas and Césari (2006) on theoccurrence of Gorgonisphaeridium cf. winslowiae Staplin et al.and Maranhites insulatus Burjack and Oliviera in the SM-4borehole (2010–2340 ft), in the Llanos Orientales Basin,Colombia. Rocha-Campos (1979) described the occurrence ofTasmanites in the Moxotó Formation of well IM st-1-PE, JatobáBasin, Brazil. In addition, Pterospermopsis brasiliensis Brito hasbeen reported from the upper part of ZoneO of theWell Izst-1-Maof the Maranhão Basin, Brazil (Brito, 1967). Although originallyassigned to “probably upper Devonian” by Brito (1967, p. 476),the upper part of Zone O was considered to correlate with the lateTournaisian–late Visean in the Parnaíba Basin, Brazil (Grahnet al., 2001, p. 83). Elsewhere, sample BU411 from Borjd’Erfoud, Morocco, was assigned to the Tournaisian (Tn2–Tn3)and Visean (V1-2) by Rahmani-Antari (1980) and it yieldedBaltisphaeridium sp., Leiofusa sp., Stellinium octoaster (Staplin)and four species of Veryhachium. Bajaník and Planderová (1985)reported two acritarchs and Tetraporina glabra Naumova fromthe Tournaisian or Visean Ochtiná Formation of the SlovakRepublic.
2.4. Visean
Acritarchs and prasinophytes have been reported fromseveral Visean sequences, but assemblages are typically of alow diversity. Hemirupta, Micrhystridium, Veryhachium andCymatiosphaera were reported by Filipiak (2005) from hisphytoplankton assemblage III of the Holy Cross Mountains,Poland, but their distribution was not detailed on any strati-graphical log. However, the acritarchs Hemirupta, Leio-sphaeridia, Micrhystridium and Veryhachium, and theprasinophytes Cymatiosphaera, Tasmanites and Dictyotidiumjachowiczii Filipiak were reported from the Zareby Beds,equivalent to the Visean TC and DP miospore biozones, ofthe Galezice IG 3 borehole, Holy Cross Mountains, Poland(Filipiak, 2005). In another detailed study by Utting (1987), fiveacritarchs, three in open nomenclature, and Chomotriletes multi-vittatus, which was considered by Utting (1987) to be an aletemiospore, were described from the Visean (V2–V3) WindsorGroup of Nova Scotia and its equivalents in Newfoundland.
There are only a small number of Visean taxa reported fromelsewhere. Biannulatisphaerites simplex Neville in Neves et al.,a fossil of perhaps algal or fungal affinity, is the only acritarch to

40 G.L. Mullins, T. Servais / Review of Palaeobotany and Palynology 149 (2008) 29–49
have been reported from the Visean of Scotland and England(Neville in Neves et al., 1973). Elsewhere in Scotland, thezygnematalean Tetraporina horologia (Staplin) was reportedfrom the upper Visean between St. Monance and Pittenweem,Fife (Neville, 1968). In Australia, Playford (1978) reported theoccurrence of his new taxon Chomotriletes multivittatusPlayford in the Ducabrook Formation of the DrummondBasin, Queensland. Similarly, the types of the acritarch Retia-letes radforthii Staplin and zygnematalean Azonotetraporina?(now Tetraporina) horologia Staplin come from the ViseanGolata Formation of west-central Alberta, Canada (Staplin,1960; see also Richards et al., 1993).
The lower Visean Radaevian, Bobrikovian and Tulianhorizons, and un-named strata, of the Volga–Ural and MoscowBasin, Russia, were examined by Byvsheva (1967, 1985;see Fig. 2). Byvsheva (1967) reported the acritarchs Azonaletes(now Arabisphaera) mosquensis Naumova and Byvsheva,Azonaletes (now Radialetes) costatus (Playford), Azonaletes(now Retialetes) bobrikii Byvsheva and Azonaletes (now Re-tialetes) radforthii (Staplin), and the zygnematalean Azonaletes(now Hemisphaerium) novus Byvsheva. Furthermore,Byvsheva (1967, p. 45) listed the occurrence of three speciesof Tetraporina, but no precise stratigraphical information wasgiven, while Byvsheva (1980) illustrated Tetraporina spp. inthe Visean. Byvsheva (1985) also listed acritarchs and zyg-nemataleans, including several of the taxa listed by Byvsheva(1967, 1980), in the Volga–Ural region and Moscow Basinof Russia. Similarly, Teteryuk (1976) also reported the oc-currence of Tetraporina spp. in the Visean of the Donets Basin,Russia.
In France, six species of Micrhystridium were describedfrom a lower Visean phosphate nodule from the MontagneNoir (Deflandre, 1946), while an un-named Visean sandstone inwells V1-59 and BBBB2-6, of the Sirte Basin, Libya, alsoyielded a limited assemblage of three acritarchs and oneprasinophyte (Tawadros et al., 2001). The Foram 1 well (1885.8–1892.8m), from theWesternDesert, Egypt, yieldedLeiosphaeridiaspp. and the colonial alga Botryococcus sp. according to Schrank(1984) and the interval was dated as late Visean, but with a midVisean or early Namurian age being possible; Schrank (1987)considered the interval to be Visean.
Kora and Schultz (1987) described two acritarchs and Tas-manites spp. from the upper Visean Abu Thora Formation of theUm Bogma area, SW Sinai, Egypt. However, the specimens ofGorgonisphaeridium cf. plerispinosum Wicander and Ecthy-mapalla cf. retusa Loeblich and Wicander which theyillustrated need to be re-examined to ascertain their relationshipwith the types of these respective taxa. Kora and Schultz (1987)originally regarded the assemblage to be of late Visean age, butthe age is late Visean–early Namurian following Zaslavskayaet al. (1995).
2.5. Undifferentiated Visean/Serpukhovian
Two studies have described material from strata that cannotbe reliably assigned to either the Visean or Serpukhovian.Schrank (1987) described Botryococcus spp. from the Foram 1
well of the Western Desert of Egypt. The other record is of twospecies of Tetraporina in the Mattson Formation of the JackfishRiver, Northwest Territories of Canada (Sullivan, 1965).
2.6. Serpukhovian
There are only two publications that describe Serpukhovianacritarchs. Cox (1986) examined the Chesterian, equivalent tothe Namurian A, Bear Gulch Limestone Member of the HeathFormation, Montana. Several acritarch taxa were reported,including an unnamed sphaeromorph and specimens assignedto Deltotosoma sp., Gorgonisphaeridium sp., Quisquilites? sp.,Leiofusa sp., Dactylofusa? sp. A and Dactylofusa? sp. B.However, with the exception of the sphaeromorph which lookssomewhat laevigate, the other illustrations appear to be vari-ously deformed examples of a single type of ornamented,spherical, organic-walled microfossil. In England, the Chokier-ian to Alportian (Namurian A) Cheddleton Sandstones, of PotBank Quarry, Mow Cop, have yielded specimens assigned toVeryhachium spp. and Micrhystridium spp. (Turner et al.,1994). The only other unequivocal Serpukhovian occurrence isof the zygnematalean Tetraporina, which was reported from thelower Serpukhovian of Russia by Einor (1986).
2.7. Undifferentiated Serpukhovian/Bashkirian
Sequences in South America and China have yieldedacritarchs of an indeterminable Serpukhovian/Bashkirian age.The Namurian to early Westphalian Jejenes Formation of LasLajas Creek, San Juan, Argentina, was examined by Césariand Bercowski (1998), who reported the occurrence of Leio-sphaeridia sp., Gorgonisphaeridium sp. and Veryhachium(Veryhachium) reductum (Deunff) (see also Limarino et al.,2002). The shallow marine Guandacol Formation of the PaganzoBasin, Argentina, which was considered to be Namurian–earliestWestphalian in Limarino et al. (2002; see also Césari andGutiérrez, 2001, text—Fig. 3), has yielded five acritarchs,including indeterminate or un-named taxa,Maranhites insulatus,Navifusa and Portalites gondwanensis Nahuys et al., theprasinophyte Tasmanites and the zygnemataleans Brazilea andTetraporina (Ottone, 1991). The Malanzán Formation of Sierrade Los Llanos, La Rioja, Argentina, which was also considered tobe ofNamurian–earliestWestphalian age inLimarino et al. (2002;see also Césari and Gutiérrez, 2001, text—Fig. 3), yielded Leio-sphaeridia, Cymatiosphaera, Greinervillites sp. and Navifusavariabilis, the zygnemataleans Brazilea and Tetraporina, and thecolonial Botryococcus (Gutiérrez and Limarino, 2001). Asummary of previous studies was presented by Limarino et al.(2002), who documented three taxa in the Malanzan Formation,two taxa in the Guandacol and Jejenes formations, and six taxa inthe upper member of the Cortaderas Formation of the Eastern,Central andWestern domains of Argentina respectively. A total often acritarch and three prasinophyte taxa were reported in theseNamurian–earliest Westphalian post-glacial transgressive strata(Limarino et al., 2002). It should be noted that Césari andGutiérrez (2001, p. 133, text—Fig. 3) placed all of the CortaderasFormation in their late Tournaisian–early Visean CV Biozone

41G.L. Mullins, T. Servais / Review of Palaeobotany and Palynology 149 (2008) 29–49
(see below). Gao (1987) reported a flora comprising sixacritarchs, one prasinophyte, and the zygnematalean Tetrapor-ina, in Jingyuan, Gansu, China, and these were correlatedapproximately with the Namurian (E2-H), SO spore zone, andwith the KD-RK zones.
2.8. Bashkirian
There is only one report of acritarchs from unequivocalBashkirian strata. Medvedeva and Kuzovleva (1971) describedtwo new species of Synsphaeridium from the BalinskoyFormation, of the Volga–Ural region of Russia. However, thematerial of Medvedeva and Kuzovleva (1971) is somewhatpoorly illustrated. In South America, di Pasquo (2002) reportedthe occurrence of the colonial alga Botryococcus in the TupambiFormation, of the Tarija Basin, northern Argentina, which isprobably early Late Carboniferous in age.
2.9. Undifferentiated Bashkirian/Moscovian
In South America, there are several reports of acritarchsfrom undifferentiated Bashkirian or Moscovian strata. A wellpreserved assemblage of Leiosphaeridia sp., Pterospermella sp.and Biannulatisphaerites simplex Neville in Neves et al. (whichis possibly of algal or fungal affinity) was described from thethermally mature Caño Indio and Río Palmar formations,Morrowan–Atokan or Desmoinesian series, Venezuela (McLeanand Harvey, 2002). An additional twelve acritarchs wereconsidered to be, or possibly be, reworked from older strata,(McLean and Harvey, 2002). Elsewhere in South America,Souza (1998) and Souza et al. (2003) described the occurrence ofDeusilites tenuistriatus Gutiérrez in the Westphalian ItarareSubgroup of the Paraná Basin, São Paulo, Brazil, while Woodand Elsik (1999) described the zygnematalean Reduviasporo-nites stoschianus (Balme) from the Copacabana Formation ofPeru, which was considered to be Early–Middle Pennsylvanian,Morrowan–Atokan.
In Israel, the occurrence of Veryhachium longispinumHorowitz, Solisphaeridium rossignoli Gilkson in Horowitz,and Micrhystridium stellatum Deflandre, were documented inthe Westphalian Sa’ad Formation of the Zohar 8 and Avdat 1cores (Horowitz, 1973). It is important to note that the age of thematerial was determined by sporomorphs and the recoveredphytoplankton was also shown to be present in the Permian (seeHorowitz, 1973, p. 184). Furthermore, Zaslavskaya et al. (1995)referred the Sa’ad Formation to the Permian.
In the United Kingdom, a diverse assemblage was describedfrom the Mansfield Marine Band, Westphalian B and C bound-ary, Upper Carboniferous, of the Smeaton and Bilby cores fromnear the English towns of Doncaster and Worksop respectively(Spode, 1964). Thin mudstone bands (c. 1 m thick) containinggoniatites, lamellibranchs, gastropods and lingulids yieldedscarce palynomorphs, including the new speciesMicrhystridiumkeratoides, which was based on 80+ specimens (see Spode,1964, p. 360, 365). However, several other taxa were recognizedon single individuals only. Triangular veryhachids with threeprocesses were assigned to three formae originally described
from the Upper Ordovician and Permian, although this probablyrepresents taxonomic over-splitting. In addition, Spode (1964)reported the occurrence of Micrhystridium nannacanthumDeflandre, Micrhystridium inconspicuum Deflandre, and aspecimen of Micrhystridium cf. recurvatum Valensi, whichwere originally described from the Silurian, Upper Cretaceousand Middle Jurassic respectively. Considering all of thesefactors, it is clear that the collections of Spode (1964) need to bere-examined.
2.10. Moscovian
Peppers (1970) and Ravn (1986) each described one newspecies of the zygnematalaen Maculatisporites from the coalmeasures of the Carbondale Formation and of the Floris andKilbourn formations, Desmoinian Series, of Illinois and Iowarespectively. The only known acritarchs from this level are fromthe Horizonmarin de Petit Buisson,Westphalien C, Boussu, síegeSentinelle des Charbonnages du Borinage, Belgium, where threetaxa, including two new species of Micrhystridium, weredescribed (Stockmans and Willière,1962).
2.11. Undifferentiated Kasimovian/Gzhelian
Only a handful of studies have described palynoflora from thelatest parts of the Carboniferous. In South America theprasinophyte Tasmanites spp. was documented in the Stephanian(Carboniferous) to Kungurian (Permian) Campo Mourão, TacibaandPalermo formations of the ParanáBasin ofBrazil (deQuadros,2002). Additionally, the zygnematalean Tetraporina prima wasrecovered from the Stephanian Chaokochung Formation ofKaiping, Hebei, China (Gao, 1985). The upper half of the TupeFormation of the Paganzo Basin, Argentina, which was correlatedwith the DMb-c biozones of Césari and Gutiérrez (2001) andwhich is regarded as probably Stephanian here, has yielded thezygnematalean Tetraporina horologia according to Ottone(1991).
2.12. Other Carboniferous occurrences
Several studies have described acritarchs from the Carbon-iferous, but the lack of precise stratigraphical informationmeans that they can only be assigned to either the Early or LateCarboniferous.
An un-named Lower Carboniferous unit in borehole ST-8 innorthern Saudi Arabia yielded a diverse, well preservedpalynoflora of nine acritarchs, eleven zygnemataleans, two in-certae sedis, two chlorophytes and the colonial alga Botryo-coccus, but unfortunately precise stratigraphical control islacking and the environment of deposition is unknown (Hemerand Nygreen, 1967; also see comments below). Elsewhere,Playford (1963) described one acritarch and three zygnemata-leans from the Early Carboniferous of Spitzbergen. In addition,Sinha et al. (1983) described two acritarchs from the ChailFormation of the Himalaya, from a level perhaps correspondingto somewhere between the Visean and Namurian, but thespecies identifications imply reworking.

42 G.L. Mullins, T. Servais / Review of Palaeobotany and Palynology 149 (2008) 29–49
In the Late Carboniferous, García (1996) described a total ofthirteen acritarchs, two hydrodictyaceae and one zygnemata-ceae from the El Imperial Formation of the San Rafael Basin,Argentina. Several of the illustrated specimens appear to bepoorly preserved and damaged and the acritarchs, together withthe scolecodonts, comprised some 6.3% of the assemblage(see also Césari and Gutiérrez, 2001, p. 124). These strata wereconsidered to correspond to the Raistrickia–Plicatipollenitessubzone of the Lunbladispora–Potonieisporites Zone and tobe of middle Late Carboniferous age (García, 1996). Subse-quently, the Raistrickia–Plicatipollenites subzone was corre-lated with the A and B subzones of the Namurian–StephanianRaistrickia densa–Convolutispora muriornata (DM) Biozoneby Césari and Gutiérrez (2001, text—Fig. 2). Eleven acritarchs,one prasinophyte, two Hydrodictyaceae and four zygnemata-leans were documented from the DM Biozone by Césariand Gutiérrez (2001; the age of these assemblages is discussedfurther below). As well as these records, di Pasquo andAzcuy (1997) described an assemblage dominated by hydro-dictyaceae, zygnemataleans and Botryococcus from the LateCarboniferous Mandiyutí Group of the Tarija Basin, northernArgentina. In addition, Ottone and García (1996) describedLeiosphaeridia sp. and Veryhachium trispinosum (Eisenack)from the Late Carboniferous Santa Maxima Formation ofArgentina.
In Brazil, the Late Carboniferous Itarare Subgroup ofItaporanga, São Paulo, has yielded the zygnematalean Brazi-lea scissa (Balme and Hennelly), the acritarch Deusilitestenuistriatus Gutiérrez et al. and the colonial alga Botryococ-cus (di Pasquo et al., 2003). Souza et al. (1993), Souza et al.(2000) and Souza (2003) have also described material from theLate Carboniferous of Brazil. Also in South America, theacritarchs D. tenuistriatus and Portalites gondwanensis, andthe prasinophyte Tasmanites sp., were reported from the LateCarboniferous of the Paso de las Bochas of the Río Negro,Uruguay, by Daners et al. (2004). In addition, the colonial algaBotryococcus, the Hydrodictyaceae Quadrisporites horridus(Hennelly) and the zygnematalean Tetraporina punctata(Tiwari and Nevale) were also documented to occur.
In Europe, Ilavská (1964) described four acritarchs from theNiedere Tatra, Slovakia, which is considered to be ofWestphalian C to Stephanian (J. Williams pers. comm.).
The Late Carboniferous–Permian Mansi Salt Beds ofHimachal Pradesh, India, were reported to contain five acritarchtaxa by Srivastava and Venkataraman (1984). However, it isimportant to note that their figured specimens seem to show anumber of indistinct, poorly preserved, sphaeromorph-likefossils lacking any diagnostic features and these records should,therefore, be disregarded.
Kondratyev (1963) reported numerous species of the zygne-matalean Tetraporina from the middle and upper Carboniferousof the Tunguska Basin (sixteen species) and from the LowerCarboniferous of the Moscow Basin (four species). We considerthat these reports probably represent taxonomic over-splitting.
Five studies have reported material from Carboniferous, orpossible Carboniferous, strata. Limestones from Carboniferouslevels in the 1111 Gîrla More borehole, c. 1.5 km northwest of
Gîrla More village in the southwest part of the MoesianPlatform, Romania, have yielded Lophosphaeridium spp. andacritarchs assigned to the ‘form group Veryhachium trispino-sum’ according to Iordan et al. (1984), but no precise age wasgiven. In India, Shrivastava and Venkataraman (1975) reportedon the microfossils from the Blaini Formation, which was datedas Early to Late Carboniferous on the basis of the palynoflora.However, the illustrations of the fossil material show a numberof featureless sphaeromorph-like objects and these records areprobably best disregarded. Three possible Carboniferousoccurrences include that of Vergel (1987), who reported Micr-hystridium sp. and Portalites gondwanensis in the UpperPalaeozoic of Santa Fe, Argentina. Calver et al. (1984) listedVeryhachium sp. from a level below the Early PermianTasmanites Shale in the Douglas River, Tasmania, Australia.The acritarchs Hystrichosphaeridea and Schizosporis gondwa-nensis Hart were reported from the Karoo sediments of SouthAfrica, which are generally regarded to be Permian, but“contain miospores assignable to the Carboniferous Period”according to Hart (1963).
3. Carboniferous acritarch and algal microfossil diversity
Estimating the diversity of Carboniferous acritarchs and algalmicrofossils is problematical, as there is a lack of precise age controlin some instances. Furthermore, taxonomic consistency and/oraccuracy are also sometimes uncertain (see comments above). Inaddition, the environment of deposition will have influenced therelative diversity of the marine acritarchs, and the terrestrialHydrodictyaceae and Zygnemataceae within each assemblage.Moreover, sampling biases caused by the amount of research input(e.g. Fig. 3) and the variable duration of each stage will also effectdiversity calculations. We have therefore illustrated the relativediversity fluctuations through the Carboniferous by documentingthe maximum number of taxa reported by a single study in eachtime bin (Fig. 8; see also comments above).
3.1. Diversity of the acritarchs
Our data suggest that acritarch diversity declined from theTournaisian to the Serpukhovian, with generally low diversityin the Late Carboniferous (Fig. 8). However, some fairly diverseLate Carboniferous assemblages are known (see below). Themost diverse acritarch assemblages occur in the Tournaisian,with 36 taxa being described from the Yali and Naximformations of Tibet (Gao, 1986). Other significant records ofTournaisian acritarchs include those of McNestry (1988), whoreported a total of 25 taxa from the Ewenny (19 taxa) and Cwrt-yr-ala (22 taxa) boreholes of South Wales, and Stockmans andWillière (1966a,b), who documented 17 and 15 taxa in thel’Asile d’aliénés and Vieux Leuze boreholes respectively. Thenumber of acritarch taxa recorded is lower in equivocalTournaisian or Visean strata, with 12 taxa being reported fromthe CV Biozone of Argentina by Césari and Gutiérrez (2001;Fig. 9; see comments on the age of the Cortaderas Formation).Both Deflandre (1946) and Utting (1987) documented six taxafrom unequivocal Visean, while Turner et al. (1994) reported

Fig. 8. Minimum and maximum number of acritarch and prasinophyte algal taxadocumented in each age subdivision of the Carboniferous. F, Famennian;T, Tournaisian; V, Visean; S, Serpukhovian; B, Bashkirian; M, Moscovian;K, Kasimovian; G, Gzhelian.
43G.L. Mullins, T. Servais / Review of Palaeobotany and Palynology 149 (2008) 29–49
just two taxa (Veryhachium spp. and Micrhystridium spp.) fromunequivocal Serpukhovian (see also comments on Cox, 1986above). Strata that are assigned to undifferentiated Serpukho-vian or Bashkirian have yielded slightly more diverseassemblages. Gao (1987) documented six acritarch speciesfrom Chinese strata that we consider to be of a possibleSerpukhovian or Bashkirian age. Similarly, the summary paperof Limarino et al. (2002) documented six acritarchs in theNamurian (=Serpukhovian or Bashkirian) upper member of theCortaderas Formation of Argentina, although this formationwas refered to the late Tournaisian–early Visean by Césari andGutiérrez (2001, p. 133).
There is only one record of unequivocal Bashkirianacritarchs, which comprises poorly illustrated Synsphaeridiumcompactum and S. rarum (Medvedeva and Kuzovleva, 1971).A potentially significant record from undifferentiated Bashkir-ian or Moscovian is the report of 10 acritarchs in the MansfieldMarine Band (Westphalian B–C boundary) of England (Spode,1964). However, this material needs to be re-examined (seecomments above). In the Moscovian, Stockmans and Willière(1962) described the new species Micrhystridium parvidumentiand Micrhystridium westphalienum, together with Veryhachiumwenlockium Downie, from the Westphalian C of Belgium.
As well as the records cited above, two publications havedocumented fairly diverse Late Carboniferous acritarch assem-blages in Argentina. García (1996) reported 13 acritarch taxa,including acanthomorphs, netromorphs and sphaeromorphs, fromthe middle Late Carboniferous El Imperial Formation. Thestratigraphical position of the El Imperial Formation is somewhatequivocal, but it correlates with the A and B subzones of theRaistrickia densa–Convolutispora muriornata (DM) Biozone(see also below; Césari and Gutiérrez, 2001, text—Fig. 2). Césariand Gutiérrez (2001) documented the acritarchs reported from theLate Carboniferous Namurian to Stephanian DM Biozone. Thedata presented by Césari and Gutiérrez (2001) is reproduced here
to more clearly show the number of taxa through the LateCarboniferous in Argentina (Fig. 9). Subzone A of the DMBiozone, from which seven acritarchs were reported, may be ofNamurian–earlyWestphalian age based on its correlationwith theGuandacol and Malanzán formations (see comments above;Césari and Gutiérrez, 2001, text—Fig. 4; Limarino et al., 2002).Subzone B contains considerably fewer acritarchs, with just twospecies of Portalites being reported (Fig. 9). Finally, the diversityincreases to nine acritarchs in Subzone C,whichwe interpret to beof possible late Westphalian–Stephanian age (Fig. 9). Furtherinvestigation is required, however, as the reported occurrence ofthe typically Devonian Stellinium octoaster in the assemblagesdocumented by García (1996) and Césari and Gutiérrez (2001)might imply that some reworking of older material has ocurred. Inadditon, some of the illustratedmaterial is rather poorly preserved.Nevertheless, Césari and Gutiérrez (2001, p. 128) stated:“Although some palynofloras from this [DM Assemblage]Biozone come from deposits influenced by glaciation, we donot see evidence of reworking in the sections.”
It is clear that phytoplankton diversity in the Serpukhovianand Late Carboniferous is significantly reduced when comparedto that observed in the earlier Palaeozoic. Furthermore, thesurvival of acritarchs into the Late Carboniferous appears tohave been highly selective. Many records have documentedspecimens assigned to the genera Micrhystridium and Veryha-chium, although the numerous species recognized probablyreflects inconsistency in the taxonomic nomenclature employedand over-splitting. Similarly, several records have described theoccurrence of species of the sphaeromorph genus Leiosphaeri-dia. Reports of other earlier Palaeozoic genera in the LateCarboniferous should be disregarded or re-examined. Forexample, the Late Carboniferous records of Baltisphaeridium(e.g. Spode, 1964) are probably erroneous, while the presence ofDuvernaysphaera, Gorgonisphaeridium, Maranhites and Stel-linium in glacially influenced Late Carboniferous strata needs tobe re-examined (García, 1996; Césari and Gutiérrez, 2001; seecomments above).
3.2. Diversity of algal microfossils
The prasinophytes, zygnemataleans and hydrodictyaceaereported from the Carboniferous are summarized on Figs. 5–7.The prasinophyte algae may show a declining diversity throughthe Carboniferous, but this is based on a limited number of dataand may be misleading as typically only 1–3 taxa have beendocumented in the majority of studies (Fig. 5). Furthermore, it isdifficult to assess true diversity in several records as the authorshave not documented individual species, but have used opennomenclature as Cymatiosphaera spp., Dictyotidium spp. andTasmanites spp. In the Tournaisian, a maximum of seven taxawere reported from China (Gao, 1986; Fig. 5). In Poland fourspecies were documented in the Tournaisian, declining to threespecies in theVisean (Filipiak, 2005; Fig. 5). In addition, diversityis generally low in the small number of records from the LateCarboniferous.
There is no clear pattern in the diversity of the zygnema-taleans through the Carboniferous (Fig. 6). Furthermore, the

Fig. 9. The number of acritarchs, prasinophytes, zygnemataceae and hydrodictyaceae documented in the Carboniferous of Argentina by Césari and Gutiérrez (2001).The CV (Cordviosporites–Verrucosporites) Biozone equiates to the late Tournaisian–early Visean; Subzone A of the DM (Raistrickia densa–Convolutisporamuriornata) Biozone may be of Namurian–early Westphalian age (see above); Subzone C of the DMBiozone may be of late Westphalian–Stephanian age (see above).
44 G.L. Mullins, T. Servais / Review of Palaeobotany and Palynology 149 (2008) 29–49
most diverse assemblages in the Early and Late Carboniferousare known from strata whose exact age is equivocal. An un-named unit in Saudi Arabia, where Hemer and Nygreen (1967)documented 11 species of Fusilites, Hemisphaerium, Lacuna-lites and Tetraporina, represents the most diverse assemblage,but the exact chronostratigraphical position of these assem-blages remains unknown (Fig. 6; also see above). Thisassemblage was recovered from a dark grey, micaceous,carbonaceous shale within a sandstone and occurred inassociation with a rich microfossil record of vascular landplants. Although the environment of deposition was not stated,the occurrence of numerous zygnemataleans together with Bo-tryococcus and non-marine acritarchs (e.g. Arabisphaera)indicates a significant terrestrial influence. Similarly, diverseassemblages are known from the Late Carboniferous, but againtheir exact chronostratigraphical position remains equivocal. InArgentina, three species of Tetraporina, and Brazilea scissa,were reported from the DM Biozone by Césari and Gutiérrez(2001; Figs. 6, 9, see also above). Similarly, ten taxa assigned toBrazilea, Kagulubeltes, Maculatisporites and Tetraporinawere described from the Late Carboniferous Mandiyutí Groupof Argentina (di Pasquo and Azcuy, 1997; Fig. 6). The diverseassemblages of pollen, spores and algae in diamictitic mud-
stones and siltstones led di Pasquo and Azcuy (1997) to proposea continental palaeoenvironment for the unit. Kondratyev(1963) described a diverse assemblage of sixteen species ofTetraporina from the Tunguska Basin, Russia (Fig. 6), but weconsider that this probably represents taxonomic oversplitting.Other records of zygnemataleans from the Carboniferoustypically describe 1–4 species (Fig. 6) and times of high di-versity probably reflect the dominance of non-marine influ-enced deposition (see above).
It is not possible to say anything about the diversity of thehydrodictyaceae through the Carboniferous, as only two speciesof Quadrisporites have been documented (Fig. 7). Theirdiversity would be influenced by the dominance of terrestrialpalaeoenvironments, and Césari and Gutiérrez (2001) statedthat Santa Máxima and El Imperial formations, which containQuadrisporites, “represent marine-littoral deposits.”
4. Palaeogeographical distribution
The palaeogeographical distribution of the organic-walledmicrofossil assemblages documented above is shown on Fig. 1.It is clear that the majority of records are known from thepalaeoequatorial regions to moderately high palaeolatitudes

45G.L. Mullins, T. Servais / Review of Palaeobotany and Palynology 149 (2008) 29–49
of the southern hemisphere. Only a few northern hemisphereoccurrences are known. In addition, most of the documen-ted localities are situated on the margins of Laurussia andGondwana.
Acritarch assemblages from possible Fammenian or Tour-naisian strata appear to show higher diversites at low latitudesbetween 0 and 30°S (Turnau, 1975; Playford, 1976; Playfordand McGregor, 1993; Stempien-Sałek, 1997) than at higherlatitudes between 30 and 60°S (Coquel et al., 1977; Rahmani-Antari, 1980, 1990). However, this pattern is not observed inassemblages from certain Tournaisian strata, or possibleTournaisian or Visean strata. In certain Tournaisian strata, themost diverse acritarch assemblages have been obtained fromNepal, situated on the margin of Gondwana (Gao 1986; seeFig. 1). Other notably diverse assemblages have come fromLaurussia (Stockmans and Willière, 1966a,b; McNestry, 1988;Playford and McGregor, 1993; Filipiak, 2005), Armorica(Lachkar et al., 1993) and Gondwana (Vavrdová et al., 1993).All of these localities were situated between c. 10°S and 45°S,but it is important to note that other localities from comparablelatitudes have only yielded lower diversity floras (e.g. Houlgatteet al., 1988; Pereira et al., 1996; Rubinstein et al., 1996).Acritarch assemblages of Visean age are known from lownorthern to high southern latitudes (Fig. 1). Although the data issuggestive of higher diversity assemblages at lower latitudes(e.g. Deflandre, 1946; Utting, 1987; Filipiak, 2005), the numberof taxa recovered and the number of productive localities is toolow to be certain. Thus, no consistent relationship betweenacritarch diversity and palaeolatitude can be discerned atpresent.
Of significant interest are the Late Carboniferous floralassemblages that have been documented from South America,which were situated at high latitudes in the southern hemisphere(e.g. Ottone, 1991; García, 1996; di Pasquo and Azcuy, 1997;Césari and Bercowski, 1998; Souza, 1998; Césari andGutiérrez, 2001; Gutiérrez and Limarino, 2001; Souza et al.,2003; Limarino et al., 2002; de Quadros, 2002; Daners et al.,2004). It is clear that these assemblages are temporally andpalaeogeographically associated with the Late CarboniferousGondwana glaciation (e.g. Fig. 1; Isbell et al., 2003, Fig. 1).These assemblages contain many taxa that are found elsewherein the Carboniferous (e.g. Veryhachium, Micrhystridium, Leio-sphaeridia, Tasmanites). However, they also yielded the twospecies of Hydrodictyaceae known from the Carboniferous(Fig. 8). In addition, Portalites and Deusilites are also knownfrom these assemblages, as well as from the Early Carbonif-erous of Saudi Arabia (Hemer and Nygreen, 1967), and thissuggests that these genera may be restricted to the high latitudesof the southern hemisphere.
5. Conclusions
Ninety-two publications have described in situ acritarchs,Prasinophyceae, Zygnemataceae and/or Hydrodictyaceae fromCarboniferous strata. The majority of these publications havedescribed material from the Early Carboniferous, principallyfrom the Tournaisian and Visean. In contrast, relatively little is
known about the phytoplankton of the latest Early Carbonif-erous Serpukhovian and the Late Carboniferous.
The majority of the Carboniferous phytoplankton assemblagesso far documented were located on Laurussia, Gondwana andArmorica, which were situated in the equatorial to high latituderegions of the southern hemisphere (Fig. 1). Only a small numberof records are known from the northern hemisphere. Importantly, asynthesis of the data currently available shows no consistentrelationship between palaeolatitude and the diversity of Carbon-iferous phytoplankton. However, it does show that some LateCarboniferous, high latitude, phytoplankton assemblages aretemporally and palaeogeographically associated with the Gond-wana glaciation (Fig. 1). Furthermore, the genera Portalites andDeusilites may be restricted to these high latitude southernregions, as these genera are only known to occur in South Americaand Saudi Arabia.
The number of taxa documented in each study in each temporalsubdivision has been used to illustrate the relative diversityfluctuations through the Carboniferous. The data indicate thatacritarch diversity declined significantly from the Tournaisian (amaximum of 36 taxa documented in a single study), to equivocalTournaisian or Visean (13 taxa), Visean (6 taxa), and theSerpukhovian (2 taxa). Acritarch diversity in the Late Carbonif-erous is generally very low, with typically only 1–3 species beingdocumented in each assemblage. However, several relativelymore diverse assemblages, with up to 13 taxa, have been docu-mented in the Late Carboniferous of England and South America(Spode, 1964; García, 1996; Césari andGutiérrez, 2001; Limarinoet al., 2002). The presence of fairly diverse acritarch assemblagesin the Late Carboniferous could imply that the very low diversityso far documented results from a lack of research input. However,these assemblages should be re-examined to assess theirsignificance as (1) some of the species documented are based onthe recovery of single specimens, (2) taxonomic oversplitting ofsimple morphologies is evident, (3) the recycling of olderassemblages into these Late Carboniferous strata needs to befully discounted.
The reduction in acritarch diversity through the EarlyCarboniferous represents the continuation of a decline thatbegan in the latest Devonian. Moreover, phytoplankton fossilsare generally absent until the Late Triassic and this period ofscarcity has been termed the “phytoplankton blackout” (Riegel,2001, in press). However, the factor(s) responsible for thedecline in the acritarchs remain controversial and a full debategoes beyond the scope of this paper. Nevertheless, a briefsynopsis is provided.
The rich spore and pollen fossil record present duringthe “blackout”period indicates that the lackof phytoplankton fossilsis unlikely to be a preservational artefact (Riegel, in press).However, numerous other explanations have been invoked. Onepossibility is that the decline in diversity was related to the end offossilizable cyst construction and/or the appearance of a majortaxon of non-encysting algae (e.g. Strother, 1996, in press;Martin, 2003; see also Riegel, in press). The decline has also beenlinked to both increased and decreased nutrient availability (e.g.Martin, 2003; Riegel, in press). Changes in palaeooceanograhy,including the interruption of the circumequatorial current during

46 G.L. Mullins, T. Servais / Review of Palaeobotany and Palynology 149 (2008) 29–49
the collision of Gondwana and Laurussia, the spread of lowsalinity surface waters, and the stratification of the oceans andblack shale formation, have also been cited as possible drivingmechanisms (Riegel, 2001, in press). The absence of phyto-plankton fossils has also been linked to a major period ofglaciation and the dominance of physical (rather than chemical)weathering, to glacio-eustatic sea-level fluctuations, to thediversification of the land plants, and to changes in nutrient fluxrelated to the deposition of sediments in large continental basinsand the deep sea (Riegel, 2001, in press). In another recent study,Strother (in press) has noted that the Devonian–Carboniferousacritarch diversity decline coincided with changing atmosphericand ocean chemistry (a reduction in pCO2 and a change from alow Mg calcite to high Mg calcite+aragonite ocean), and withincreased carbon flux to the oceans related to land plant evolution.Proposing that Palaeozoic phytoplankton may have had ineffi-cient carbon concentration mechanisms, Strother (in press) linkedthe diversity decline to inefficient inorganic carbon assimilationand a loss of cyst formation as a principal mechanism for survivalfollowing a large-scale shift in heterotrophic nutrition. It isimportant to state, however, that our synthesis shows that there is ageneral absence of high-resolution phytoplankton studies acrossthe critical Devonian–Carboniferous boundary interval withwhich to test these hypotheses. Further work is urgently neededto understand the mechanism(s) that drove this fundamentalchange in the nature of the ocean’s primary producers.
With regards the prasinophyte algae, the data suggest apattern of declining diversity through the Carboniferous.However, the robustness of this trend is questionable as thenumber of taxa reported is typically very low and the species areoften grouped in open nomenclature (e.g. Cymatiosphaera spp.etc). Similarly, there is no clear trend in the diversity of theZygnemataceae, with the majority of assemblages yielding onlya small number of taxa. It is also likely that the diversezygnematalean assemblages known in the Early Carboniferous(e.g. Hemer and Nygreen, 1967) and Late Carboniferous (e.g. diPasquo and Azcuy, 1997) reflect increased terrestrial influenceor continetal deposition.
Acknowledgements
We thank John Williams of the Natural History Museum,London, who facilitated access to his database and to many ofthe publications cited in this paper. We also thank two anony-mous referees for their helpful comments. This research wasfunded by the Leverhulme Trust (reference F/00212/F).
References
Attar, A., Fournier, J., Candilier, A.M., Coquel, R., 1980. Étude palynologiquedu Dévonien terminal et du Carbonifère inférieur du bassin d’Illizi (Fort-Polignac), Algérie. Revue de l’Institut Français du Pétrole 35 (4), 585–619.
Bajaník, Š., Planderová, E., 1985. Stratigrafická pozícia spodnej časti ochtinskéhosúvrtvia gemerika medziMagnezitovcami aMagurou. Geologické práce, Správy82, 67–76 Geologický Ústav Dionýza Štúra, Bratislava, 1985.
Batten, D.J., 1996. Chapter 7C. Colonial chlorococcales. In: Jansonius, J.,McGregor, D.C. (Eds.), Palynology: Principles and Applications, 1. AmericanAssociation of Stratigraphic Palynologists Foundation, pp. 191–203.
Boneham, R.F., 1970. Acritarchs (Leiosphaeridia) in the New Albany Shaleof southern Indiana. Proceedings of the Indiana Academy of Science, 79,pp. 254–262.
Brito, I.M., 1967. Silurian and Devonian Acritarcha from Maranhão Basin,Brazil. Micropaleontology 13 (4), 473–482.
Byvsheva, T.V., 1967. K palinologicheskoi kharakteristike i stratigraficheskomuraschleneniyu bobrikovskovo gorizonta nizhnevo karbona Volga–Uralskoioblasti [Palynologic characteristics and stratigraphical subdivision of theBobrikovo horizon of the lower Carboniferous in the Volga–Ural region].Iskopayemyye Spory I Pyl’tsa Rasteniy, Ikh Znacheniye Dlya Stratigrafiches-kogo Raschleneniya Neftegazonosnykh Tolshch Dokembriya, Paleozoya,Mezozoya Yevropeyskoy chasti SSSR i Sredney Azii. Vsesouznyy Naucno-Issledovatelskiy Geologorazvedocnyy Neftanoy Institut (VNIGNI), Geohimi-ceskiy Sbornik, Trudy, 52. Gos. Nauchno-Tekhn, Moscow, USSR, pp. 14–47.
Byvsheva, T.V., 1980. Zonalnye kompleksy spor verkhneturneiskikh otlozheniivostochnykh raionov Russkoi plity [Spore zone complexes of upperTournaisian deposits of the eastern region of the Russian Plate].Palinologicheskie Issledovaniya Proterozoya i Fanerozoya Neftegazonos-nykh Regionov SSSR. Vsesoyuznyi Nauchno-Issledovatelskii Geologor-azvedochnyi Neftyanoi Institut, 217. Trudy, Moscow (VNIGNI), pp. 53–61.
Byvsheva, T.V., 1985. Spory iz otlozhenii turneiskogo i vizeiskogo yarusovRusskoi plity. Atlas Spor i Pyltsy Neftegazonosnykh Tolshch FanerozoyaRusskoi i Turanskoi Plit. Vsesoyuznyi Nauchno-Issledovatelskii Geologor-azvedochnyi Neftyanoi Institut, 253. Trudy, Moscow (VNIGNI), pp. 80–159.
Calver, C.R., Clarke, M.J., Truswell, E.M., 1984. The stratigraphy of a LatePalaeozoic borehole section at Douglas River, eastern Tasmania: a synthesisof marine macro-invertebrate and palynological data. Papers and Proceed-ings of the Royal Society of Tasmania, 118, pp. 137–161.
Césari, S.N., Bercowski, F., 1998. Palinología de la Formación Jejenes(Carbonífero) en la Quebrada de Las Lajas, provincía de San Juan, Argentina,Nuevas inferencias paleoambientales. Ameghiniana 34 (4), 497–509.
Césari, S.N., Gutiérrez, P.R., 2001. Palynostratigraphy of Upper Paleozoicsequences in central western Argentina. Palynology 24, 113–146.
Colbath, G.K., Grenfell, H.R., 1995. Review of biological affinities of Paleozoicacid-resistant, organic-walled eukaryotic algal microfossils (includingacritarchs). Review of Palaeobotany and Palynology 86, 287–314.
Coquel, R., Loboziak, S., Stampfli, G., Stampfli-Vuille, B., 1977. Palynologiedu Dévonien supérieur et du Carbonifère inférieur dans l’Elburz oriental(Iran Nord–Est). Revue de Micropaléontologie 20 (2), 59–71.
Cox, R.S., 1986. Preliminary report on the age and palynology of the Bear GulchLimestone (Mississppian, Montana). Journal of Paleontology 60, 952–956.
Crowell, J.C., 1999. Pre-Mesozoic Ice Ages: Their Bearing on Understandingthe Climate System: Boulder, 192. Geological Society of America Memoir,Colorado, pp. 1–106.
Daemon, R.F., 1974. Palinomorfos-guias do Devoniano superior e CarboniferoInferior das Bacias do Amazonas e Parnaíba. Anais da Academia Brasileirade Ciencias 43 (3/4), 549–807.
Daners, G., de Santa Ana, H., de Souza, P.A., 2004. Nuevos datos palinológicosde la sección basal de la Formación San Gregorio en la localidad de Paso delas Bochas (Pérmico Inferior-Cuenca Norte, Uruguay). Congreso Uruguayode Geologia, Montivideo, Anais.
Davydov, V., Wardlaw, B.R., Gradstein, F.M., 2004. The Carboniferous period.In: Gradstein, F.M., Ogg, J.G., Smith, A.G. (Eds.), A Geologic Timescale2004. Cambridge University Press, pp. 222–248.
Deflandre, G., 1946. Radiolaires et Hystrichosphaeridés du Carbonifère de laMontagne Noir. Comptes Rendus des séances de l’Académie des sciences223 (13), 515–516.
di Pasquo, M., 2002. The Crassispora kosankei–Cystoptychus azcuyi palynozoneof the Upper Carboniferous Tupambi Formation, Tarija Basin, northernArgentina. Review of Palaeobotany and Palynology 118 (1–4), 47–76.
di Pasquo, M., Azcuy, C.L., 1997. Palinología del Grupo Mandiyutí,Carbonífero Superior. VIII Congreso Geológico Chileno (Antofagasta1997). Actas 1, Sesión Temática 3, 475–479 Universidad Catolica delNorte, Departmento de Ciencias Geológicas.
di Pasquo, M., Azcuy, C.L., Souza, P.A., 2003. Palinología del CarboníferoSuperior del Subgrupo Itararé en Itaporanga, Cuenca Paraná, Estado deSão Paulo, Brasil. Parte 1: sistemática de esporas y paleofitoplancton.Ameghiniana 40 (3), 277–296.

47G.L. Mullins, T. Servais / Review of Palaeobotany and Palynology 149 (2008) 29–49
de Quadros, L.P., 2002. Acritarchos e Tasmanites do Permo-Carbonifero dabacia do Paraná. Revista do Instituto Geologico (São Paulo) 23, 39–50.
Dueñas, H., Césari, S.N., 2006. Palynological evidence of Early Carboniferoussedimentation in the Llanos Orientales Basin, Colombia. Review ofPalaeobotany and Palynology 138, 31–42.
Edgell, H.S., 2004. Upper Devonian and Lower Carboniferous foraminiferafrom the Canning Basin, Western Australia. Micropaleontology 50, 1–26.
Einor, O.L., 1986. The former USSR. In: Wagner, R.H., Winkler Prins, C.F.,Granados, L.F. (Eds.), The Carboniferous of the World. III The former USSR,Mongolia, Middle Eastern Platform, Afghanistan, & Iran. Publication, vol. 33.IUGS, pp. 13–407.
Eisenack, A., 1962. Mitteilungen über Leiosphären und über das Pylom beiHystrichosphären. Neues Jahrbuch für Geologie und Paläontologie,Abhandlungen 114, 58–80.
Filipiak, P., 2005. Late Devonian and Early Carboniferous acritarchs andprasinophytes from the Holy Cross Mountains (central Poland). Review ofPalaeobotany and Palynology 134 (1–2), 1–26.
Gao, L., 1983. Discovery of Late Devonian spore assemblages from NyalamCounty, Xizang (Tibet) and their stratigraphic significance. Contribution toGeology Qinghai–Xizang (Tibet) Plateau 8, 183–218.
Gao, L., 1984a. Early Devonian spore assemblages in China. Neuvième CongrèsInternational de Stratigraphie et deGéologie duCarbonifère,Washington,D.C.and Champaign-Urbana, Illinois, May 17–26, 1979, 2, pp. 103–108.
Gao, L., 1984b. Late Devonian and Early Carboniferous miospore zones fromNyalam County, Xizang (Tibet) and the boundary of the Devonian–Carboniferous. Abstracts Himalayan Geol. Int. Symp., (ISGH), p. 3.
Gao, L., 1985. Carboniferous and Early Permian spore assemblages of NorthChina region and the boundary of the Carboniferous and Permian. DixièmeCongrès International de Stratigraphie et de Géologie du Carbonifère,Madrid, 1983, 2, pp. 409–424.
Gao, L., 1986. Late Devonian and Early Carboniferous acritarchs from NyalamCounty, Xizang (Tibet), China. Review of Palaeobotany and Palynology 47,17–30.
Gao, L., 1987. Namurian miospore zonation from Jingyuan, Gansu and the mid-Carboniferous boundary. Bulletin of the Institute of Geology ChineseAcademy of Geological Sciences 16, 193–249.
García, G.B., 1996. Palinología de la Formación El Imperial, PaleozoicoSuperior, Cuenca San Rafael, República Argentina; Parte II, Granos depolen, incertae sedis, acritarcas. Ameghiniana 33 (1), 7–33.
Grahn, Y., Loboziak, S., Melo, J.H.G., 2001. Integrated miospore-chitinozoanbiozonation of the Parnaíba Basin and its correlation with Petrobras(Müller 1962) Silurian–Lower Carboniferous palynozones. In: Melo, J.H.G.,Terra, G.J.S. (Eds.), Correlaçâo de seqüências Paleozóicas Sul-Americanas.Ciência-Técnica-Petróleo, 20. Exploração de Petróleo, Seção, pp. 81–89.
Grenfell, H.R., 1995. Probable fossil zygnematacean algal spore genera. Reviewof Palaeobotany and Palynology 84 (3–4), 201–220.
Gutiérrez, P.R., Limarino, C.O., 2001. Palinología de la Formación Malanzán(Carbonífero Superior), La Rioja, Argentina: nuevos datos y consideracionespaleoambientales. Ameghiniana 38 (1), 99–118.
Guy-Ohlson, D., 1996. Prasinophycean algae. In: Jansonius, J., McGregor, D.C.(Eds.), Palynology: Principles and Applications, 1. American Association ofStratigraphic Palynologists Foundation, pp. 181–189.
Hart, G.F., 1963. A probable Pre-Glossopteris micro-floral assemblage fromLower Karroo sediments. South African Journal of Science 59 (5), 135–146.
Hemer, D.O., Nygreen, P.W., 1967. Algae, acritarchs and other microfossilsincertae sedis from the Lower Carboniferous of Saudi Arabia. Micropale-ontology 13 (2), 183–194.
Higgs, K.T., Finucane, D., Tunbridge, I.P., 2002. Late Devonian and EarlyCarboniferous microfloras from the Hakkari Province of southeasternTurkey. Review of Palaeobotany and Palynology 118, 141–156.
Horowitz, A., 1973. Plantes microfossiles du Carbonifère supérieur de la partiesud d’Israël. Revue de Micropaléontologie 16 (3), 184–193.
Houlgatte, E., LeHérissé, A., Pelhate, A., Rolet, J., 1988. Evolution géodynamiquedu Bassin carbonifère de Laval. Géologie de la France 1, 27–46.
Ilavská, Z., 1964. Sporen und Hystrichosphaerideen aus dem Karbon derNiederen Tatra. Geologicky Sbornik 15, 2.
Iordan, M., Iliescu, V., Visarian, A., Baltres, A., Săndulescu, E., Seiferth, K.1984. Litho- and biostratigraphy of the Paleozoic sequences in the Oprişor
and Gîrla More boreholes (south-western part of the Moesian Platform).D. S. Inst. Geol. Geofiz, v. LXIX/4 (1982), 5–28.
Isbell, J.L., Miller, M.F., Wolfe, K.L., Lenaker, P.A., 2003. Timing of latePaleozoic glaciation in Gondwana: was glaciation responsible for thedevelopment of northern hemisphere cyclothems? Geological Society ofAmerica Special Paper 370, 1–20.
Kondratyev, G.K., 1963. The first finds of Angiospermae type pollen in UpperPaleozoic beds of the Tunguska Basin. Akademie Naukove SSSR, SiberianDivision, Geological Geophysical 12, 63–72.
Kora, M., Schultz, G., 1987. Lower Carboniferous palynomorphs from UmBogma, Sinai (Egypt). Grana 26, 53–66.
Lachkar, G.A., Moreau-Benoit, A., Raymond, D., 1993. Découverte d’unemicroflore tournaisienne dans la chaine Ibérique Orientale (Espagne).Geobios 26, 523–534.
Li, J., Servais, T., Brocke, R., 2002. Chinese Paleozoic acritarch research: reviewand perspectives. Review of Palaeobotany and Palynology 118, 181–193.
Limarino, C.O., Césari, S.N., Net, L.I., Marenssi, S.A., Gutiérrez, R.P.,Tripaldi, A., 2002. The Upper Carboniferous postglacial transgression inthe Paganzo and Río Blanco basins (northwestern Argentina): facies andstratigraphic significance. Journal of South American Earth Sciences 15,445–460.
Martin, F., 1993. Acritarchs: a review. Biological Review 68, 475–538.Martin, R.E., 2003. The fossil record of biodiversity: nutrients, productivity,
habitat area and differential preservation. Lethaia 36, 179–193.McLean, D., Harvey, C., 2002. Palynomorphs from the Late Carboniferous
Caño Indio and Río Palmar formations of Venezuela. In: Hills, L.V.,Henderson, C.M., Bamber, E.W. (Eds.), Carboniferous and Permian ofthe World, 19. Canadian Society of Petroleum Geologists, Memoir, pp.371–381.
McNestry, A., 1988. The palynostratigraphy of two uppermost Devonian–LowerCarboniferous borehole sections in South Wales. Review of Palaeobotanyand Palynology 56, 69–87.
Medvedeva, A.M., Kuzovleva, G.A., 1971. Izuchenie akritarkh iz neftei iporod Volgo–Uralskoi oblasti metodom replik. Spory i Pyltsa vNeftyakh i Porodakh Neftegazonosnykh Oblastei SSSR [Spore andpollen in oils and deposits of oil-and gas-bearing regions of the USSR].K III Mezhdunarodnoi Palinologicheskoi Konferentsii Novosibirsk,Sovetskii Soyuz. Izdatelstvo Nauk, Moskva, pp. 21–27. 1971.
Moldowan, M.J., Dahl, J., Jacobson, S.R., Huizinga, B.J., Fago, F.J., Watt, D.S.,Peters, K.E., 1996. Chemostratigraphic reconstruction of biofacies:molecular evidence linking cyst-forming dinoflagellates with pre-Triassicancestors. Geology 24 (2), 159–162.
Moldowan, J.M., Talyzina, N.M., 1998. Biogeochemical evidence fordinoflagellate ancestors in the Early Cambrian. Science 281, 1168–1170.
Mullins, G.L., Aldridge, R.J., Siveter, D.J., 2004. Microplankton associations,biofacies and palaeoenvironment of the type lower Ludlow Series, Silurian.Review of Palaeobotany and Palynology 130, 163–194.
Neville, R.S.W., 1968. Ranges of selected spores in the upper Visean of the East Fifecoast section between St. Monace and Pittenweem. Pollen et Spores 10 (2),431–462.
Neves, R., Gueinn, K.J., Clayton, G., Ioannides, N.S., Neville, R.S.W.,Kruszewska, K., 1973. Palynological correlations within the LowerCarboniferous of Scotland and northern England. Transactions of theRoyal Society of Edinburgh 69 (2), 23–70.
Ottone, E.G., 1991. Palynologie du Carbonifère supérieur de la coupe de MinaEsperanza, Bassin Paganzo, Argentine. Revue de Micropaléontologie 34,118–135.
Over, D.J., 1992. Conodonts and the Devonian–Carboniferous boundary in theupper Woodford Shale, Arbuckle Mountains, South-Central Oklahoma.Journal of Paleontology 66 (2), 293–311.
Peppers, R.A., 1970. Correlation and palynology of coals in the Carbondale andSpoon formations (Pennsylvanian) of the northeastern part of the IllinoisBasin. Ilinois State Geological Survey, Bulletin 93, 1–173.
Pereira, Z., Clayton, G., Oliveira, J.T., 1996. Palynostratigraphy of theDevonian–Carboniferous boundary in southwest Portugal. Annales de laSociété géologique de Belgique 117 (1), 189–199.
Playford, G., 1963. Lower Carbonifeous microfloras of Spitsbergen. Palaeon-tology 5, 619–678.

48 G.L. Mullins, T. Servais / Review of Palaeobotany and Palynology 149 (2008) 29–49
Playford, G., 1976. Plant microfossils from the Upper Devonian and LowerCarboniferous of the Canning Basin, Western Australia. PalaeontographicaAbteilung B Palaophytologie 158, 1–71.
Playford, G., 1978. The Lower Carboniferous spores from the DucabrookFormation, Drummond Basin, Queensland. Palaeontographica Abteilung BPalaophytologie 167, 105–160.
Playford, G., McGregor, D.C., 1993. Miospores and organic-walled micro-phytoplankton of Devonian–Carboniferous boundary beds (Bakken For-mation), southern Saskatchewan; a systematic and stratigraphic appraisal.Geological Survey of Canada, Bulletin 445, 1–106.
Rahmani-Antari, K., 1980. Etude palynologique dans le dévono-carbonifère de lacoupe du Borj d’Erfoud (Tafilalt). Mines, Géologie et Energie 48, 105–114.
Rahmani-Antari, K., 1990. Etude palynologique et évaluation de l’indiced' altération thermique du Paléozoique du forage DOT 1 (bassin desDoukkala centre-ouest marocain). Review of Palaeobotany and Palynology66 (3–4), 211–227.
Rahmani-Antari, K., Lachkar, G., 2001. Contribution a l’etude biostratigraphi-que du Devonien et du Carbonifère de la plate-forme marocaine; datationet correlations. Revue de Micropaléontologie 44 (2), 159–183.
Ravn, R.L., 1986. Palynostratigraphy of the Lower and Middle Pennsylvaniancoals of Iowa. Technical Papers, 7. Iowa Geological Survey, pp. 1–244.
Richards, B.C., Bamber, E.W., Higgins, A.C., Utting, J., 1993. Carboniferous.In: Stot, D.F., Aitken, J.D. (Eds.), Sedimentary Cover of the Craton inCanada, 5. Geological Survey of Canada, pp. 202–271. Geology of Canada.
Riegel, W., 2001. Phytoplankton development during the Late Palaeozoic. TerraNostra 4, 66–72.
Riegel, W., in press. The Late Palezoic phytoplankton blackout — Artefact orevidence of global change? Review of Palaeobotany and Palynology.
Rocha-Campos, A.C., 1979. Dinantian stratigraphy of Brazil. PalaeontologicalCharacteristics of the main subdivisions of the Carboniferous. Huitième CongrèsInternational de Stratigraphie et du Géologie Carbonifère, Moscow, Sept. 8–13,1973. Compte Rendu, 3. Publishing Office «Nauka», Moscow. 1979.
Rubinstein, C., Niemeyer, H., Urzúa, F., 1996. Primeros resultados palinológicosen la Formación Zorritas, Devónico–Carbonífero de Sierra de Almeida,Región de Antofagasta, Chile. Revista Geológica de Chile 23 (1), 81–95.
Schrank, E., 1984. Paleozoic and Mesozoic palynomorphs from the Foram 1well (Western Desert, Egypt). Neues Jahrbuch für Geologie und Paläonto-logie, Monatshefte 1984 (2), 95–112.
Schrank, E., 1987. Palaeozoic and Mesozoic palynomorphs from northeastAfrica (Egypt and Sudan) with special reference to the Late Cretaceouspollen and dinoflagellates. Berliner geowiss. Abhandlungen 75 (1),249–310.
Servais, T., Li, J., Molyneux, S., Raevskaya, E., 2003. Ordovician organic-walled microphytoplankton (acritarch) distribution: the global scenario.Palaeogeography, Palaeoclimatology, Palaeoecology 195, 149–172.
Servais, T., Stricanne, L., Montenari, M., Pross, J., 2004. Population dynamicsof galeate acritarchs at the Cambrian–Ordovician transition in the AlgerianSahara. Palaeontology 47, 395–414.
Shrivastava, R.N., Venkataraman, K., 1975. Palynostratigraphy of the BlainiFormation. Bulletin Industriel Geological Association 8 (2), 196–199.
Sinha, A.K., Ashgirei, G.D., Umnova, N.I., 1983. Carboniferous palynomorphsfrom Chail Formation of southern Himachal Himalaya, and their tectonicimplications. Himalayan Geology volume 11 (Palaeontology and biostra-tigraphy of the Himalaya). Wadia Institue of Himalayan Geology, DehraDun, pp. 415–423. 1981 (issued 1983).
Souza, P.A., 1998. Palaeoenvironmental considerations about the ItararéSubgroup at Araçoiaba da Serra, State of São Paulo, (Upper Carboniferous,Paraná Basin), Brazil. Ameghiniana 35 (3), 315–320.
Souza, P.A., 2003. New palynological data of the Itararé Subgroup from the BuriCoal (Late Carboniferous, Paraná Basin), São Paulo State, Brazil. RevistaBrasileira de Paleontologia 5, 49–58.
Souza, P.A., Lima, M.R., de, Saad, A.R., 1993. Palinologia dos carvõespaleozóicos do Estado de São Paulo. I–O Carvão de Buri. Revista doInstituto Geológico, São Paulo 14 (1), 5–20.
Souza, P.A., Batezelli, C.V.B., di Pasquo, M., Azcuy, C.L., Saad, A.R.,Perinotto, J.A.deJ., 2000. Ocorrência de Palinomorfos no Subgrupo Itararé(Carbonífero/Permiano da Bacia do Paraná) na Região de Jundiaí (SP,Brasil). Revista Universidade Guarulhos, Geociências 5, 28–32.
Souza, P.A., Petri, S., Dino, R., 2003. Late Carboniferous palynology from theItararé Subgroup (Paraná Basin) at Araçoiaba de Serra, São Paulo State,Brazil. Palynology 27, 39–74.
Spode, F., 1964. A new record of hystrichospheres, from the Mansfield MarineBand, Westphalian. Proceedings of the Yorkshire Geological Society, 34,pp. 357–370.
Srivastava, R. N., Venkataraman, K. 1984. Palynostratigraphy of the MandiSalt Beds of Himachal Pradesh. Symposium on evolutionary botany andbiostratigraphy, February 24-26, 1979, Department of Botany, Univer-sity of Calcutta, published by R. K. Jain, New Delhi, India, 285–289.
Staplin, F.L., 1960. Upper Mississippian plant spores from the Golata Formation,Alberta, Canada. Palaeontographica Abteilung B Palaophytologie 107 (1–3),1–40.
Staplin, F.L., Jansonius, J., Pocock, S.A.J., 1965. Evaluation of someacritarchous hystrichosphere genera. Neues Jahrbuch für Geologie undPaläontologie, Abhandlungen 123 (2), 167–201.
Stasiuk, L.D., 1996. Organic facies in black shale of Devonian–MississippianBakken Formation, southeastern Saskatchewan. Interior plains and ArcticCanada [Plaines interieures et region arctique du Canada]. GeologicalSurvey of Canada, pp. 15–22. Current Research.
Stasiuk, L.D., Fowler, M.G., 2004. Organic facies in Devonian andMississippian strata of Western Canada Sedimentary Basin: relation tokerogen type, paleoenvironment, and paleogeography. Bulletin of CanadianPetroleum Geology 52 (3), 234–255.
Stempien-Sałek, M., 1997. Some acritarchs of the Upper Palaeozoic fromWestern Pomerania (NW Poland). Acta Universitatis Carolinae, Geologica40, 667–673.
Stephenson, M.H., Williams, M., Leng, M.J., Monaghan, A.A., 2004. Aquaticplant microfossils of probable non-vascular origin from the BallaganFormation (Lower Carboniferous), Midland Valley, Scotland. Proceedingsof the Yorkshire Geological Society, 55, pp. 145–158.
Stockmans, F., Willière, Y., 1962. Description de trois Hystrichosphèrestrouvés dans les schistes de l’horizon Marin de Petit Buisson.Observations sur la paléontologie, la lithologie et la stratigraphie duWestphalien B et C dans la Partie Occidentale du Massif du Borinage(district du Couchant de Mons), 5. Publication, Centre national degéologie houillère (Belgique), pp. 11–30.
Stockmans, F., Willière, Y., 1966a. Les Acritarches du Dinantien du sondage del’Asile d’aliénés à Tournai, Belgique. Bulletin de la Société belge degéologie, de paléontologie et d’hydrologie 74 (2), 462–477.
Stockmans, F., Willière, Y., 1966b. Les Acritarches du Dinantien du sondage deVieux Leuze à Leuze (Hainaut, Belgique). Bulletin de la Société belge degéologie, de paléontologie et d’hydrologie 75 (2), 233–242.
Stricanne, L., Munnecke, A., Pross, J., Servais, T., 2004. Acritarch dis-tribution along an inshore–offshore transect in the Gorstian (lowerLudlow) of Gotland, Sweden. Review of Palaeobotany and Palynology130, 195–216.
Stricanne, L., Munnecke, A., Pross, J., 2006. Assessing mechanisms ofenvironmental change: palynological signals across the Late Ludlow(Silurian) positive isotope excursion (δ13C, δ18O) on Gotland, Sweden.Palaeogeography, Palaeoclimatology, Palaeoecology 230, 1–31.
Strother, P.K., 1996. Acritarchs. In: Jansonius, J., McGregor, D.C. (Eds.),Palynology: Principles and Applications, 1. American Association ofStratigraphic Palynologists Foundation, pp. 81–106.
Strother, P.K., in press. A speculative review of factors controlling the evolutionof phytoplankton during Paleozoic time. Revue de Micropaléontolgie.
Sullivan, H.J., 1965. Palynological evidence concerning the regionaldifferentiation of Upper Mississippian floras. Pollen et Spores 7 (3),539–563.
Talyzina, N.M., Moczydlowska, M., 2000. Morphological and ultrastructuralstudies of some acritarchs from the Lower Cambrian Lükati Formation,Estonia. Review of Palaeobotany and Palynology 112, 1–21.
Tawadros, E., Rasul, S.M., El-Zaroug, R., 2001. Petrography and palynology ofquartzites in the Sirte Basin, central Libya. Journal of African Earth Sciences32 (3), 373–390.
Teteryuk, V.K., 1976. Namurian stage analogues in the Carboniferous Period ofthe Donets Basin (based on palynological data). Geological Journal 36,110–122.

49G.L. Mullins, T. Servais / Review of Palaeobotany and Palynology 149 (2008) 29–49
Torsvik, T.H., Cocks, L.R.M., 2004. Earth geography from 400 to 250 Ma: apalaeomagnetic, faunal and facies review. Journal of the Geological Society,London 161, 555–572.
Turnau, E., 1975. Microflora of the Famennian and Tournaisian deposits fromboreholes of Northern Poland. Acta Geologica Polonica 25 (4), 505–528.
Turner, N.E., Spinner, E., Spode, F., Wignall, P.B., 1994. Palynostratigraphy of aCarboniferous transgressive systems tract from the earliest Alportian(Namurian) of Britain. Review of Palaeobotany and Palynology 80, 39–54.
Utting, J., 1987. Palynology of the Lower Carboniferous Windsor Group andWindsor-Canso boundary beds of Nova Scotia, and their equivalents inQuebec, New Brunswick and Newfoundland. Geological Survey of Canada,Bulletin 374, 1–93.
van Geel, B., Grenfell, H.R., 1996. Chapter 7A. Spores of zygnemataceae.1996 In: Jansonius, J., McGregor, D.C. (Eds.), Palynology: Principles andApplications, 1. American Association of Stratigraphic PalynologistsFoundation, pp. 173–179.
Vanguestaine, M., 1986. Late Devonian and Carboniferous acritarch stratigraphyand paleogeography. Late Devonian events around the Old Red Continent;Field Conference in Aachen (FRG), With Excursions in the Ardenne(Belgium), the Rheinisches Schiefergebirge (FRG) and in Moravia (CSSR).Annales de la Société Géologique de Belgique, 109(1), pp. 93–102.
Vavrdová, M., Isaacson, P.E., Diaz, E., Bek, J., 1993. Devonian–Carboniferousboundary at Lake Titikaka, Bolivia: preliminary palynological results.Comptes Rendu XII ICC-P 1, 187–199.
Vergel, M.M., 1987. Palinología del Paleozoica superior en la perforacion YPFSF J (Josefina), Provincia de Santa Fe, Argentina. II Anteturma varriegermi-nantes, groupo Acritarcha e incertae sedis. Ameghiniana 24 (1–2), 67–80.
Veevers, J.J., Powell, C.M., 1987. Late Paleozoic glacial episodes inGondwanaland reflected in transgressive–regressive depositional sequencesin Euramerica. Geological Society of America Bulletin 98, 475–487.
Willis, K.J., McElwain, J.C., 2002. The Evolution of Plants. Oxford UniversityPress, p. 378.
Wood, G.D., Elsik, W.C., 1999. Paleoecologic and stratigraphic importance ofthe fungus Reduviasporonites stoschianus from the Early–Middle Pennsyl-vanian of the Copacabana Formation, Peru. Palynology 23, 43–53.
Zaslavskaya, N., Eshet, Y., Hirsch, F., Weissbrod, T., Gvirtzman, G., 1995.Recycled Lower Paleozoic microfossils (Chitinozoa) in the Carboniferous ofSinai (Egypt) and Permo-Triassic of the Negev (Israel): paleogeographicconsiderations. Newsletters on Stratigraphy 32 (1), 57–72.
Zhang, S.X., Jin, Y.G., 1976. The Upper Paleozoic brachiopods from theMountain Qolmolungma Region. Reports of Scientific Expedition ofMountains Qolmolungma Region. Science Press, Beijing, pp. 159–242.
Zhong-Qiang, 2004. Devonian–Carboniferous brachiopod zonation in theTarim Basin, northwest China: implications for biostratigraphy andbiogeography. Geological Journal 39, 431–458.





























![Hyperspectral Remote Sensing of Phytoplankton Species ...²ˆ芳.pdf · type [7]. In addition, they are also indicative of variability in phytoplankton species diversity in the ocean](https://img.dokumen.tips/doc/110x75/604b1bd50788831e2b23d1b3/hyperspectral-remote-sensing-of-phytoplankton-species-epdf-type-7.jpg)