Embed Size (px)
Citation preview

This article was downloaded by: [129.130.252.222]On: 18 July 2014, At: 14:32Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH,UK
Bulletin de la SociétéBotanique de France. LettresBotaniquesPublication details, including instructions for authorsand subscription information:http://www.tandfonline.com/loi/tabg19
Thallus formation of twocyanobacterial lichens:Peltigera didactyla andPeltigera praetextata, underlaboratory conditionsElfie Stocker-Wörgöttera
a Universität Salzburg, Institut färPflanzenphysiologie, Hellbrunnerstr. 34, 5020Salzburg, Österreich/Autriche, AustriaPublished online: 10 Jul 2014.
To cite this article: Elfie Stocker-Wörgötter (1991) Thallus formation of twocyanobacterial lichens: Peltigera didactyla and Peltigera praetextata, underlaboratory conditions, Bulletin de la Société Botanique de France. Lettres Botaniques,138:3, 179-187, DOI: 10.1080/01811797.1991.10824920
To link to this article: http://dx.doi.org/10.1080/01811797.1991.10824920
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all theinformation (the “Content”) contained in the publications on our platform.However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness,or suitability for any purpose of the Content. Any opinions and viewsexpressed in this publication are the opinions and views of the authors, andare not the views of or endorsed by Taylor & Francis. The accuracy of theContent should not be relied upon and should be independently verified with

primary sources of information. Taylor and Francis shall not be liable for anylosses, actions, claims, proceedings, demands, costs, expenses, damages,and other liabilities whatsoever or howsoever caused arising directly orindirectly in connection with, in relation to or arising out of the use of theContent.
This article may be used for research, teaching, and private study purposes.Any substantial or systematic reproduction, redistribution, reselling, loan,sub-licensing, systematic supply, or distribution in any form to anyone isexpressly forbidden. Terms & Conditions of access and use can be found athttp://www.tandfonline.com/page/terms-and-conditions
Dow
nloa
ded
by [
] at
14:
32 1
8 Ju
ly 2
014

Bull. Soc. bot. Fr., 138, l..ettres bot. (3), 179 -187, (1991).
Thallus formation of two cyanobacterial lichens
Peltigera didactyla and Peltigera praetextata,
under laboratory conditions (*)
hy Elfic STOCKER-WORGOTTER
Universitiit Salzburg, Institut )ùr Pjlanzenphysiologie, llellbrunnerslr .. 14, 5020 Salzburg, Osterreich/Autriche, Austria
Summary.- The formation of mature thalli of Pelrigera didaclyla from soredia occured within an average time span of 5 to 6 months or even Jess in Jaboratory cu~ures. The size and growth forms of the thalli (musselshaped and branched type) were dependent upon the number of the soredia involved in the thallus forming process. Not only soredia amalgamated, but also lobe primordia can fuse during early development to form a joint thallus.
The resynthesis of Peltigera praetextata from its isolated myco- and cyanobiont was achieved in laboratory controlled cultures. Fully developed thalli were only formed on sail substratum alter an incubation lime of at !east 2 years. Culture experiments w~h Peltigera praetextata were aise carried out by Yoshimura and Yamamoto (1990) by the tissue culture method. These resu~s will be discussed.
Résumé.· En ce qui concerne Peltigera didactyla (syn. P. spuria), on obtient, à partir d'ensemencement des sorédies, la formation de thalles adultes, eux-mêmes sorédiés, en moyenne en 5 à 6 mois. La taille et la forme des thalles (en écuelle ou lobés) dépe':ldent du nombre de sorédies qui ont contribué à la formation du thalle. Non seulement les sorédies s'amalgament au début de la culture en formant une masse sorédiée unique, mais plus tard les jeunes lobes qui en sont issus, fusionnent à leur tour pour former un seul thalle adutte. Les problèmes de contamination, qui sont fréquents dans les cultures initiales, disparaissent, en partie, quand on utilise pour l'ensemencement, des sorédies récoltées sur des thalles obtenus en culture.
En ce qui concerne Peltigera praetextata, sa resynthèse à partir de la cyanobactérie et de spores du champignon n'a été réussie au laboratoire que dans le cas où du sol est utilisé comme substrat. Sur agar, la formation de mycélium n'a lieu qu'en présence d'un nombre suffisant de cellules du photosymbiote et la "resynthèse" se réduit à la formation d'une association lâche Nostoc-champignon, sans structure lichénique reconnaissable. Sur sol, les premiers stades de la formation d'un thalle ne sont obtenus qu'après un temps de latence très long, de 2 à 3 mois. Tout d'abord les Nostoc et le champignon s'associent en formant des structures organisées comme des sorédies, ensuite des amas sorédiés plus volumineux se développent, enfin les premiers lobes thalliens apparaissent. Des thalles bien développés, avec des rhizines, et ressemblant à ceux croissant en milieu naturel sont obtenus en deux ou trois ans.
Key words : thallus formation- cyanobacterial lichens- Peltigera species.
* * *
(')Conférence prononcée lors de la séance du i 9 octobre 1990.
©Société botanique de France 1991 - ISSN 0181 - 1797
Dow
nloa
ded
by [
] at
14:
32 1
8 Ju
ly 2
014

180 LETIRES BOTANIQUES
INTRODUCTION Therc have hccn numerous attempts to cultivatc foliose lichens, Lut
as you know, gcnerally with limited sueeess. One of the hest known speeics in this respect is Xanthoria parietina,
which first had bccn rcsynthcsizcd by the French liehcnologist Bonnicr (1886, 1 889) and la ter on studied hy Werner (1927). Bu brick and Gal un (1986) suceeedcd in cultivating Xanthoria parietina from its isolated spores and algae. Although fruiting bodies were formcd, the culture of a weil dcvcloped foliosc thallus was not achievcd.
Foliosc lichens with Trebouxia photobionts have not bcen rcsynthcsizcd until now.
Aecording to Lallcmant and Bernard (1977) and many othcr authors, sorne lichen spores do not germinale or if they germinale they do not form mycelia without the presence of a suitable photobiont.
Although foliosc lichens arc wide-sprcad and weil studicd, there are only fcw investigations on thcir juvenile growth and devclopmcnt.
A first stcp in this direction was made by Sehustcr, Ott and J ahns (1985) who studicd the devclopmcnt of sorne lichen spccics in the natural cnvironmcnt. So sorne fundamental knowledge of a few lichen )ife cycles is availahlc. Thus in sorne cases exists a possibility to compare lichen dcvclopmcnt undcr laboratory eonditions with the devclopmcnt of frecliving forms.
For our culture experimcnts with foliosc lichens wc lookcd for a suitable objeet. Wc scarched for a lichen which probably reprcscnts a simpler system according to the symbiotic rclationship.
First culture cxperimcnts werc stimulated by the observation that the foliose, but eyanohactcrial lichen Peltigera didactyla (former name Pdtigera spuria) can colonizc open, disturhcd sites within sorne months or a half ycar.
ln this respeet Peltigera didactyla represcnts a lichen with a very short life cyde whieh mcans that thallus formation would be observable under laboratory conditions.
First I will tell you from the cultivation of Peltigera didactyla from soredia.
The second part of my lecture will direct your attention to the artifieial recstablishment of the related Peltigera praetextata from its isolated spores and algae.
MATEHIAL AND METIIODS Cultivation of soredia of PeUigera didactyla
First lichen mate rial of P. didactyla was collected at 1300 m elevation in the Rottensteiner Valley, Carinthia, Austria. For later experiments the soredia were laken from cultivated thalli. The culture substratum was sterilized sail from the habitat of the lichen. Culture conditions : 19--20°C, day-night regime 14 : 1 o h, at a light intensity of 60-90 f.LE m's··•. The relative humidity in the culture chamber was 50-60%.
Resynthesis experiments with Peltigera pratextata Thalli with fruiting bodies of P. praetextata were also gathered in the Rottensteiner Valley.
Fungal spores were isolated from washed apothecia (phosphate buffered saline, Bubrick and Galun (1986) and tape water). Cultiva! ion of the fungus was conducted on agarplates in petri dishes or in test tubes.
For isolation of the cyanobiont small thallus pieces were macerated between two glass slides and soma Nostoc cells were transferred by the micropipette method of Ahmadjian (1973) to sterile media. The cyanobiont cells were cultivated on solid and in liquid BBM-medium (Bischoff and Bold, 1963) and purified
Dow
nloa
ded
by [
] at
14:
32 1
8 Ju
ly 2
014

E. STOCKER-WORGOTTER 181
from contamination by successive inoculation to new media by the method of Boissière et al. {1987). Resynthesis cultures were conducted on BBM-Agar {BBM-solution, + 2% agar, + sail extract prepared by the method of Esser, 1976) and on sterilized sail (autoclaved for 3 hours on two successive days) from the natural habitat of the lichen. The cultures were kept either at 19--2o•c or at varying temperature, 10•c at night and 2o•c at day lime. ln the culture room was a daily cycle of 14: 10 h, in the growth chamber with varying temperature a lightdark regime of 12: 12 h maintained. The light sources were flourescent lamps (type Philips 65W/RW) with a photon flux of 100 1-1E m·•s·'. The relative humidity was the same as described above, 50-60%.
The developmental stages were investigated by light microscopy and micro- and macrophotographie methods as described by Stocker-Wiirgiitter and Türk (1990, 1991 ).
RESULTS Cultivation of Thalli of Peltigera didactyla
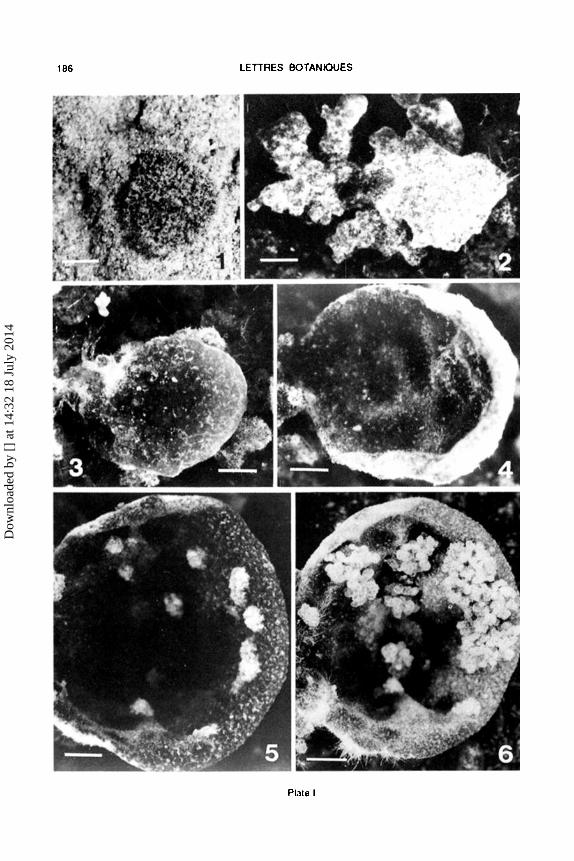
Germination of the soredia took place during a time intervall of 2-4 wccks. Figure 1 shows you a germinated soredium after 21 days in eulturc. Following further growth the soredia were fixed to the substratc by outgrowing hyphae. During the colonization phase neighbouring sorcdia fuscd and formed a basal tissue or clump of tissue. Aftcr the formation of the basal tissues had taken place, thallus tliffercntiation initiatctl.
Aftcr 2-3 months of cultivation the tissue clumps had transformed into small uplifted thallus lobe primordia. Somctimcs, during further growth, whcn thesc lobes came into direct contact with each othcr they fuscd. Figure 2 shows you a lobe primordium during this amalgamating proeess.
ln the final phase of this process mostly single, broad and concave thallus lobes with a smooth surface (Figure 3) had been formed. lly further differentiation eup- or dish-shaped thalli with a weil dcvclopcd cortex cmcrgcd (Figure 4, cup-shaped thallus aftcr 4 months of cultivation). Aftcr 5 months, a high perccntagc of the cultivatcd thalli bcgan to differentiatc soralia (Figure 5). Sorcdia diffcrentiation in culture occurcd very quickly and was mostly tcrminated within 3-4 weeks (Fig. 6, cultivated thallus with sorcdia). During this process the thalli did not grow. Aftcrwards growth somctimcs proeeeded. Soredia produeed hy the cultivated thalli showed the same vitality and germination rate as sorcdia won from nature. They have an important advantagc, they wcre free of contaminants and so we could take them for further eultivation. As thalli grown undcr natural conditions the cultivatcd thalli showed a varicty of growth forms. ln the cultures the mussel-shaped and irregularly branched types werc very common. Although the thalli appearcd hcalthy for a longer timc (up to one year) a dcgcncration of soralia and in the final phase the formation of fruiting bodies did not occur in lahoratory culture.
Rcsynthcsis of thulli of Peltigera praetextata from the isolatcd spores and t~yanobactcria
Cultivation on agar plates Soon it hccamc obvious that the formation of .nycclia by this fungus
eould not be stimulated without cyanobiont eells. These findings agree to carlier observations made by Scott (1959, 1960, 1964 a, b).
After a rclativcly long lag phase of 2-3 months a high amount of the Nostoc colonies had hccn infccted by the lichen fungus. llut the eultivation on agar plates only provided loosc fungal cyanobacterial associations, but no thallus-likc structures.
Dow
nloa
ded
by [
] at
14:
32 1
8 Ju
ly 2
014

182 LETTRES BOTANIQUES
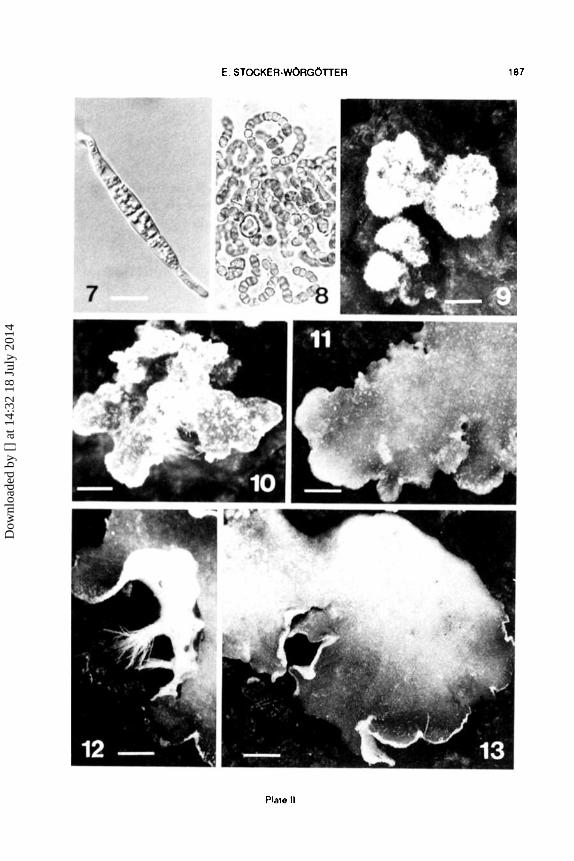
Cultivation on soil substratum First purificd Nostoc-colonies (Figure 8) were transfcrrcd to the soil
and after an incubation time of 2-3 weeks inoculated with mycobiont spores (Figure 7) or hyphae obtained from germinated spores. The recombination of the isolated myco- and cyanobiont was marked by a long initiation phase. During this long !ag-phase nearly the cntire soil substratum was covcrcd by a layer of Nostoc ccli aggregates. Although the fungus was permanently in contact with photobiont cells, it grcw very slowly. The first minute lichenizcd stages emerged after an incubation time of 3 months.
By further growth these associations showed a globose and soredialike arrangement (Figure 9), although they originated from ascosporcs and cyanobacteria. During the next dcvclopmental stcp (5-6 months) neighbouring soredial stages fused and grcw together forming a basal tissue or clump of tissue. Further differentiation principally flollowed the same way as already described for Peltigera didactyla. Amalgamation of lobe primordia was very common (Figure 10). So most of the developing thalli represented joint thalli. After an incubation time of one year nearly 90% of the thallus primordia had transformed into corticated thallus lobes (Figure 11). After two years and six months the thalli (Figure 13) became very similar to young thalli from nature. After this long incubation time sorne of the thalli had formed rhizines (Figure 12). Finally, after three ycars of devclopment, most of the cultured thalli had reached a size of 2 ems.
DISCUSSION Peltigera didactyla
Our investigations with Peltigera didactyla demonstrated that thallus formation and the differentiation of organs for vegetative reproduction oceur routinely under laboratory conditions within a time span of 5-6 months or even Jess.
Earlier cultures in the natural environmcnt and with adult thalli of Peltigera erumpens (former name of P. didactyla) over a time period of more than one year have been reported by Darbishire (1927). Darbishire (1927) found that the thalli which had overgrown musses had formcd soredia. From thesc soredia new small thalli arose within a time interval of further 5 months. The time span observed by Darbishirc (1927) eorresponded very weil with our devclopmental times, both in cultures in the laboratory and also in the natural environmcnt (Stockcr-Worgotter and Türk, 1989).
According to our results, groups or severa) agglomerations of soredia were necessary for thallus formation. Single soredia often degenerated within severa! weeks after germination. During further dcvclopment small lobe primordia could mergc to develop a joint thallus. Schuster et al. (1985) found that not only soredia from one lichen specics fused and formed a sorcdia-ficld, but also whole lobes or even whole plants of different relatcd species e.g. Physcia adscendens and Ph. tenella or Cladonia mitis and Cl. rangiferina can grow together and form one thallus. These authors concluded that this fact provided probably an cxplanation for the great morphologieal variation observed within sorne lichen genera.
Dow
nloa
ded
by [
] at
14:
32 1
8 Ju
ly 2
014

E. STOCKER-W0RG0TTER 183
Referring to our results we have mostly found two different kinds of thallus morphology, the mussel-shaped and the branched thallus type. The later originated from two or three thallus lobes, which had amalgamated during devclopment. For further experiments it would be of great interest to study these fusing processes in detail.
Culture experiments with soredia have been rare until now (e.g. Tabler, 1911 ; Darbishire, 1927, Margot, 1973, Margot and De Sloover, 1974), beeause of contamination problems. ln our cultures contamination could be reduced by using seriai cultivation. For the following experiments soredia were taken from the already cultivated thalli which were very clean (free of dust and moss spores).
This was possible, as the soredia from the cultivated thalli showed the same vitality as the soredia won from natural thalli. Now in one culture series 1 have the third generation of thalli, won from soredia of the cultivated thalli and they arc still alive.
As you know Peltigera didactyla shows two strategies of reproduction, first by sorcdia, but also by spores. ln nature, sorne thalli, on which the sorcdia had dcgencratcd, fonn fruiting bodies within a few months. Until recent timc, in laboratory cultures wc have not observed the formation of fruiting bodies. Wc think that the factors which induce fruitification by P. didactyla arc not undcrstood enough. But from our cxperimcnts wc can assume that besicles internai factors a participation of climatic factors should be very important. Thcre are the scasonal changes, which cannot be easily simulated within a culture chamber.
ln contrast to Ahmadjian (1989) we found that the soil substratum was very suitable for the cultivation of J>eltigera species, the precondition to gct wholly differcntiated thalli. Ail expcriments to cultivate Peltigera didactyla soredia on agarplates or on silica gel (Galun et al., 1972 ; Marton and Galun, 1976) have failed.
Peltigera praetextata ln agreement with Scott (1964a, b) wc found that mycelia of Peltige
ra praetextata only developcd when they came into direct contact with cyanobiont cells or an extract from them. From our point of view, earlier culture failures with P. praetextata were due to the slow growth of this fungus, the long !ag-phase for rcsynthcsis and the lack of favourable media for long tcrm cultivation.
Furthermorc, according to the present results it became obvious that the fungus of P. praetextata did not have special nutritional responscs to the medium.
Earlier culture experimcnts with P. praetextata were carried out by Yoshimura and Yamamoto (1990) and presentcd at the International Symposium on Lichen Biology at Madrid. Yoshimura and Yamamoto achieved the formation of young thallus primordia with a different method, by cultivation of liehcn tissues. Thcsc authors cultivatcd their purified lichen fragments on plain agar (2% watcr agar), which did not contain further nutricnts. They achievcd the dcvclopmcnt of thallus primordia after one year of cultivation. Like the soil substratum the simple agar medium prcvcnted both symbionts from disintegra-
Dow
nloa
ded
by [
] at
14:
32 1
8 Ju
ly 2
014

184 LETIRES BOTANIQUES
tion. The carly developmental stages, Yoshimura and Yamamoto obscrvcd sccm to be idcntical to our rcsynthcsis stages, obtaincd from fungal spores and Nostoc ccli aggregatcs.
Now 1 wanted to cite direetly from thcir work, prescnted at Madrid : "Aggrcgate Nostoc colonies arc surrounded by hyphae of the mycobiont of Peltigera praetextata and they grow bigger and bigger ; then they form globosc primordia of lichen thalli, in whieh Nostoc cclls arc rather poorly composcd with hyphae. Primordia composed of mycobionts and phycobionts (Nostoc) grow gradually to bccome dorsovcntral juvenile lichen th alli ... Lichen th alli grow to scvcral millimctcrs in sizc after nearly one-ycar culture and they are composed of severa] irregular lobes".
It should be noted that the agar mcthod uscd by thcsc authors was not very suitcd for long tcrm cultivation longer thun one ycar, beeause the moisture content of agar plates cannot be regulated. Under this aspect the cultures on agar do not show a high degree of reproducibility. As earlicr culture cxpcrimcnts (e.g. Thomas, 1939; cultures with Cladonia pyxidata) have shown, lichcnizcd stages on agar are often products of chance. To ovcrcomc these difficulties wc used soil for the rcsynthcsis expcriments. A furthcr advantagc of soil substrata is that they only rcquirc addition of watcr. Our experiences from culture studics eorrcsponded very weil with the fincJings of Lange et al. (1986) that lichens containing eyanobactcria nccded watcr in liquid form to achieve an optimal co2 assimilation, whereas lichens with green photobionts arc able to absorb watcr vapour.
Culture cxpcriments with the closcly relatcd Peltigera canina wcrc conductcd by Ahmadjian (1989). Although Ahmadjian obscrvcd the beginnings of contacts bctween the lichen bionts and a stimulation of growth of the fungus hy the isolatcd Nostoc cells, resynthcsis stages wcrc not observed on agar plates or on mica strips dipping into an agar substratum.
Acknowledgements.· Il would like to thank Prof. Letrouit-Galinou and the Société Botanique de France lor the invitation to Paris to give a lecture du ring the "Séance du 19 octobre 1990".
The investigations were supported by a grant of the Fonds zur Fèirderung der wissenschaftlichen Forschung Proj. Nr. P 7470-BIO.
BJBLIOGHAPIIY AHMADJIAN V., 1973.· Methods of isolating and cuhuring lichen symbionts and thalli. ln Ahmadjian V. and
M.E. Hale, The Lichens, New York, London, 653 · 659. AHMADJIAN V .. 1989.· Studies on the isolation and synthesis of bionts of the cyanolichen Peltigera canina
(Peltigeraceae). Pl. Syst. Evol., 165, (1/2), 29-39. BISCHOFF H.W. and H.C. BOLD, 1963.- Some soil algae from enchanted rocks and related species. Univ.
Texas Publ. N°. 6318, Phycol. Studies, 4, 1- 95. BOISSIËRE J.C., M.C. BOISSIËRE, P. CHAMPION-ARNAUD, R. LALLEMANT et J. WAGNER, 1987.- Le
cycle de Nostoc des genres Peltigera et Col/ema en cu hures in vitro el dans le thalle lichénique. Canad. J. Bot., 65, 1468-1477.
BON NIER G., 1886.· Recherches expérimentales sur la synthèse des lichens dans un mil eu privé des germes. Compt. Rend. Acad. Sei. (Paris), 103, 942-944.
BONNIER G., 1889.· Recherches sur la synthèse des lichens. Ann. Sei. nat. bot. (Paris) Sér. VIl, 9, 1-34. BUBRICK P. and M. GALUN, 1986.- Spore to spore resynthesis of Xanthoria parietina. Lichenologist, 18,
47. 49. DARBISHIRE O. V., 1927.· The soredia of Peltigera erumpens Wains. and P. scutata KBR. Trans Brit. Mycol.
Soc. Xli (1927), 52 · 70. ESSER K., 1976.· Kryptogamen: Blaualgen, Algen, Pilze, Flechten. Springer Verlag, Berlin, Heidelberg, New
York, p. 25.
Dow
nloa
ded
by [
] at
14:
32 1
8 Ju
ly 2
014

E. STOCKER-WORGOnER 185
GALUN M., K. MARTON and L. BEHR, 1972.- A method tor the culture of lichen thalli under controlled conditions. Arch. Microbiol., 83, 189 - 192.
LALLEMANT R. et T. BERNARD, 1977.- Obtention de cu~ures pures des mycosymbiontes du Lobaria laetevirens (Lightt.) Zahlbr. et du Lobaria pulmonaria (L.) Hoffm., le rôle des gonidies. Rev. bryol. et lichénol., 43, 303- 308.
LANGE O.L., E. KILIAN and H. ZIEGLER, 1986.- Water vapor uptake and photosynthesis of lichens performance differences in species with green and bluegreen algae as phycobionts. Oecologia, 71, 104-110.
MARGOT J., 1973.- Experimental study of the effects of sulphur dioxide on the soredia of Hypogymnia physodes. ln Ferry B.W. et al., Air pollution and lichens, Athlone Press, London, 314-329.
MARGOT J. et J. DE SLOOVER, 1974.- La culture des sorédies lichénique : un test de la vitalité des thalles soumis à la pollution. Bull. Soc. Roy. Belgique, 107 (1 ), 33-40.
MARTON K. and M. GALUN, 1976.- ln vitro dissociation and reassociation of the lichen Heppia echinulata. Protoplasma, 87, 135- 143.
SCHUSTER G., S. on and H.M. JAHNS, 1985.- Artificial cultures of lichens in the natural environment. Lichenologist, 17, 247-253.
scan G.D, 1959.- Observations on spore discharge and germination in Pelt1gera praetextata (Fik) Vain. Lichenologist, 1, 1 09 - 111.
SCOn G.D .. 1960.- Studies of the lichen symb1osis. 1. The relationship between nutrition and moisture content in the maintenance of the symbiotic state. New Phytologist, 59, 374-381.
SCOn G.D., 1964a.- Studies of the lichen symbiosis. 2. Ascospore germination in the genus Peltigera Zeitschr. Allgem. Mikrobiol., 4 (4), 326-336.
SCOn G.D., 1964 b.- The lichen symbiosis. Adv. of Sei., 21, 244-248. STOCKER-WORGOTIER E. and R. TÜRK, 1989.- Artiticial cultures of the cyanobacterial lichen Peltigera
didactyla in the natural enwonment. Pl. Syst. Evol., 165, 39- 48. STOCKER-WORGbnER E. and R. TÜRK, 1990.- Thallus formation of the cyanobacterial lichen Peltigera
didactyla from Soredia und er laboratory conditions. Botanica Acta, 103, 315 - 321. STOCKER-WORGbnER E. and R. TÜRK, 1991.- Artificial resynthesis of the cyanolichen Peltigera praetextata
under laboratory conditions. Lichenologist, 23 (2), 127- 138. WERNER R.G., 1927.- Recherches biologiques et expérimentales sur les ascomycètes des lichens. Thèse
(Faculté des Sciences, Paris) Braun et c•, Mulhouse. TOBLER F., 1911.-11. Die Entwicklung der Cladonia-Soredien. Jahrb. f. wiss. Bot., 49, 409-417. THOMAS EA, 1939.- Über d1e B1ologie von Flechtenbildnern.- Beitr. z. Kryptogamenfl. der Schweiz, 9 ( 1 ),
1 -200. YAMAMOTO Y., 1990.- Cultured Usnea rubescens tissues thal produced a high leve! of usnic acid. Studies of
Cel/ aggregates and the Production of natural pigments in plant cel/ culture (Chapter Il), 76-87. YOSHIMURA 1. and Y. YAMAMOTO, 1990.- Developments of lichen thalli of Peltigera praetextata in culture.
3'' International Colloquium on Lichen Biology (Lecture), Madrid, April1990.
Plate 1.- Figs. 1~.- Development of Peltigera didactyla from soredia under laboratory conditions. F1g. 1. Germinated sored1um, af1er 3 weeks in culture. bar = 60 11m. Fig. 2.- Early stage of lobe ditterentiation. Thallus 1n1!ial formed by 2-3 amalgamated lobes alter an incubation lime of 3 months; bar= 200 11m. F1g. 3.- Corticated, slightly concave formed thallus primord1um, alter 3 1/2 months in culture. bar= 500 11m. Fig. 4.- Cup-shaped thallus with a weil developed cortex alter a cultivation time of 4 months. bar= 450 11m. Fig. 5.- Thallus at the beginning of soralia formation, 4 1/2 months in culture. bar= 600 ).lm. Fig. 6.- The same thallus as shawn in Fig. 5, 4 weeks later. bar= 700 ).lm.
Plate 11.- Figs. 7- 13.- Resynthesis stages of Peltigera praetextata. F1g. 7.- Germinated fungal spore on agar substratum. bar= 10 )lm. Fig. 8.- Cult1vated Nostoc colonies on agarplate. bar = 40 11m. Fig. 9.- Fused soredial stages, form1ng a basal tissue or basal tissue (5-6 months). bar = 145 11m. Fig. 10.- More ditterentiated thal lus initial with fusod lobes. bar= 50 11m. Fig. 11.- Thal! us alter an incubation time of 20 months. bar= 1 mm. Fig. 12.· Part of thallus with weil developed rhizines. bar= 1.3 mm. Fig. 13.- Thallus, af1er 2 1/2 years in culture. bar= 1.3 mm.
Dow
nloa
ded
by [
] at
14:
32 1
8 Ju
ly 2
014
186 LETTRES BOTANKlUES
Plate 1
Dow
nloa
ded
by [
] at
14:
32 1
8 Ju
ly 2
014
E. STOCKER-W0RG0TIER 187
Plaie Il
Dow
nloa
ded
by [
] at
14:
32 1
8 Ju
ly 2
014





























