Embed Size (px)
Citation preview

Mechanisms of Development 138 (2015) 64–72
Contents lists available at ScienceDirect
Mechanisms of Development
j ourna l homepage: www.e lsev ie r .com/ locate /mod
Tetrapod axial evolution and developmental constraints; Empiricalunderpinning by a mouse model
Joost M. Woltering a,1, Denis Duboule a,b,⁎a Department of Genetics and Evolution, University of Geneva, 30 quai Ernest Ansermet, 1211 Geneva, Switzerlandb School of Life Sciences, Ecole Polytechnique Fédérale de Lausanne (EPFL), 1015 Lausanne, Switzerland
⁎ Corresponding author at: Department of GeneticsGeneva, 30 quai Ernest Ansermet, 1211 Geneva, Switzerla
E-mail address: [email protected] (D. Duboule)1 Present address; Chair in Zoology and Evolutionary B
University of Konstanz, Universitätsstraße 10, 78457 Kon
http://dx.doi.org/10.1016/j.mod.2015.07.0060925-4773/© 2015 The Authors. Published by Elsevier Ire
a b s t r a c t
a r t i c l e i n f oArticle history:Received 19 April 2015Received in revised form 19 July 2015Accepted 20 July 2015Available online 31 July 2015
Keywords:Hox genesSacrumHind limbHomeotic transformationTetrapod evolutionDevelopmental constraint
The tetrapod vertebral column has become increasingly complex during evolution as an adaptation to a terrestriallife. At the same time, the evolution of the vertebral formula became subject to developmental constraints actingon the size of the cervical and thoraco-lumbar regions. In the course of our studies concerning the evolution ofHox gene regulation, we produced a transgenicmousemodel expressing fishHox genes, which displayed a reducednumber of thoraco-lumbar vertebrae and concurrent sacral homeotic transformations. Here,we analyze thismutantstock and conclude that the ancestral, pre-tetrapodial Hox code already possessed the capacity to induce vertebraewith sacral characteristics. This suggests that alterations in the interpretation of theHox codemay have participatedto the evolution of this region in tetrapods, alongwith potentialmodifications of theHOXproteins themselves.Withits reduced vertebral number, thismouse stock violates a previously described developmental constraint, which ap-plies to the thoraco-lumbar region. The resulting offset between motor neuron morphology, vertebral patterningand the relative positioning of hind limbs illustrates that the precise orchestration of the Hox-clock in parallelwith other ontogenetic pathways places constraints on the evolvability of the body plan.© 2015 The Authors. Published by Elsevier Ireland Ltd. This is an open access article under the CC BY-NC-ND
license (http://creativecommons.org/licenses/by-nc-nd/4.0/).
‘The whole subject of homology and segmentation is very complex,imperfectly understood, and well worthy of further study’
[Edwin E. Goodrich, 1913]
1. Introduction
The vertebrate spine is built as a sequence of serial homologouselements, the vertebrae, which develop with different morphologies atdifferent positions along the anterior–posterior axis. Various vertebralformulae reflect both the requirements and the constraints associatedwith a skeleton that needs to accommodate protective, respiratoryand locomotor functions (Romer, 1956; Woltering, 2012). The differen-tiation of initially similar somites into distinct type of vertebrae (i.e. cer-vical, thoracic, lumbar, sacral and caudal in mammals) is establishedearly on during embryogenesis, mainly due to a collinear pattern ofHox gene expression (Kmita and Duboule, 2003) along the antero-
and Evolution, University ofnd..iology, Department of Biology,stanz, Germany.
land Ltd. This is an open access articl
posterior axis (Casaca et al., 2014; Deschamps and van Nes, 2005;Mallo et al., 2010;Wellik, 2009). These coordinated expression patternsindeed generate various combinations of HOX proteins at distinct bodylevels (or a ‘Hox code’ (Kessel and Gruss, 1991)), which genetically in-struct somites about their fates in terms of morphology. In addition,the correspondence between particular combinations of HOX proteinsand critical morphological transitions are maintained throughouttetrapods, suggesting an instructive role for these proteins in settingup these boundaries (Burke et al., 1995; Gaunt, 1994). However, thefact that various HOX proteins display some functional hierarchies inthese processes makes a pure combinatorial system unlikely (seeDuboule and Morata, 1994).
The evolution of land vertebrates was paralleled by an increasingcomplexity of axial regionalization, as an adaptation to a terrestriallifestyle: the sacrum evolved during the fish–tetrapod transition asa connection between the pelvic girdle and the axial skeleton(Carroll and Holmes, 2007), whereas the lumbar region firstappeared in mammals as an adaptation to sagittal flexion duringlocomotion as well as to accommodate the diaphragm (Carroll,1988). In mammals, the sacral and lumbar regions are geneticallycharacterized by the transcription of Hox genes belonging toparalogy groups 10 and 11 (Hox10 and Hox11 genes), evolutionaryrelated to the insect posterior gene Abd-B (Izpisua-Belmonte et al.,1991). Functional approaches have revealed that HOX10 proteinssuppress rib formation whereas HOX11 proteins can induce sacral
e under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/4.0/).

65J.M. Woltering, D. Duboule / Mechanisms of Development 138 (2015) 64–72
processes (Carapuco et al., 2005; Wellik and Capecchi, 2003). Inter-estingly, this collinear distribution of Hox transcripts predates theorigin of vertebrates and fish for instance already express Hox10and Hox11 genes in paraxial mesoderm, in spite of the absence ofboth sacral and lumbar regions (Oulion et al., 2011; Prince et al.,1998; van der Hoeven et al., 1996). Therefore, it is likely that theseparticular region-specific morphologies did not arise through merechanges in Hox gene expression domains. Instead, they may haveinvolved concomitant alterations in the activation of downstreamtarget genes, for example via modification in the interpretation ofthe ‘code’ either following changes in the cis-regulatory modulescontrolling these targets, or due to changes in the HOX proteinsthemselves. Altogether, it is currently unknown whether the emer-gence of these particular body regions involved a simple exaptationof a pre-existing Hox pattern, or if it was accompanied by essentialstructural changes in HOX proteins leading to novel functions.
Regardless of which evolutionary mechanism leads to such criti-cal modifications of the tetrapod spine, its realm of action was likelyreduced due to the strong developmental constraints applied to theaxial formula. The existence of developmental constraints appliedto the organization of segmental patterns in animals was recognizedmore than a century ago, by the mere observations of natural ‘rules’governing the formation of metamerized body plans (see forexample the work of Lankester, described in Jeffs and Keynes,1990). Nowadays, such constraints or limitations in the evolutionof otherwise potentially adaptive traits are thought to derive inpart from the way the underlying regulatory processes are implementedand shared between various developmental contexts, leading tosevere pleiotropic effects at least in vertebrates (Duboule and Wilkins,1998; Kirschner et al., 2005).
The comparative analysis of vertebral columns provides many in-stances of such canalized processes (Asher et al., 2011), as for examplethe well-known constraint that fixes the number of cervical vertebraeto seven in all mammals but manatees and sloths (see e.g. Galis, 1999;Galis andMetz, 2007; Varela-Lasheras et al., 2011), even though naturalselection favored in some instances either the increase or the decreasein neck length, as in giraffes and whales, respectively. In such caseshowever, variations occurred through changes in the sizes of vertebraerather than in their number. It was suggested that this constraint wasgenerated by a potential interference with the migration of thediaphragm muscles, thus leading to an impaired respiration(Buchholtz et al., 2012; Hirasawa and Kuratani, 2013). Likewise, in thethoraco-lumbar region, a constraint seems to restrict the overallnumber of vertebrae to 19 or 20 in most mammals (Narita andKuratani, 2005), perhaps associated with the proper implementationof locomotor mechanisms (Buchholtz, 2014; Galis et al., 2014).
Unfortunately, even though the mechanisms underlying both thetime-sequenced production of somites (Pourquie, 2003) and theconcurrent progressive activation of Hox genes (Noordermeer et al.,2014) start to be understood, evolutionary scenarios accounting forthe macroevolution of the axial skeleton remain complex to addressexperimentally and lack empirical support. In this study, we investi-gate the phenotypic abnormalities in a transgenic mouse containinga HoxAa BAC from the pufferfish (Tetraodon nigroviridis) genome.This transgenic line was produced in the aim of studying inter-species regulatory controls (Woltering et al., 2014) and expressesfish HoxAa genes at slightly ectopic positions during the developingmouse body axis. The resulting morphological transformations indi-cate that pre-tetrapodial HOX proteins can induce tetrapod-specificanatomical features. Also, the number of thoracic and lumbar verte-brae obtained scores below the developmental constraint identifiedby Narita and Kuratani (2005). The paraplegia observed in thesemice suggests that the origin of the constraint on the evolutionarybias in TL vertebral number may lie in the mechanistic independencebetween axial patterning by Hox genes, on the one hand, and hindlimb positioning, on the other hand.
2. Results
2.1. A mouse stock with a reduced number of TL vertebrae
In the course of our studies of Hox gene regulation during the fin tolimb transition, a transgenic mouse stock was generated by using aPufferfish (T. nigroviridis) BAC covering part of the posterior HoxAacluster and containing from HoxA9a through HoxA13a (Wolteringet al., 2014). Three F0 males were obtained, which all showed locomo-tory incapacitation of the hind limbs (paraplegia), as well as a trunkshortened along the anterior to posterior axis (Fig. 1A, Supplementarymovies 1–2). One male died shortly after weaning and the cadaverwas lost; a second male died without any apparent pathological cause,at approximately two years of age, but never reproduced; the thirdmale proved capable of reproducing and was used to establish a linethrough natural mating. As further paternal transmission of this trans-genic condition was never achieved in this line, it was maintainedthrough hemizygous maternal crosses. In addition to the male repro-ductive defects, the problems in hind limb coordination with an abnor-mal gate caused by (partial) hind-limb paralysis persisted in this line.Adult animals improved in this respect after 5 months of age.
To try and understand the etiology of these various phenotypes, weinitially analyzed these mice for potential skeletal and/or neuralabnormalities. Alizarin red-alcian blue staining in newborns and adultsrevealedmajor homeotic transformations in the posterior trunk, includ-ing a large anterior shift of the sacrum leading to a reduction of thelumbar region from usually six (sometimes five) lumbar vertebrae inwild-type mice, to only three lumbar vertebrae in the transgenic condi-tion. In addition, the second and third lumbar vertebrae (L2 and L3)showed partial sacral transformation, as shown by clearly broadenedlateral processes (Fig. 1B, C, Supplementary Fig. 1). This phenotypewas also scored in the skeleton of the second F0 male, for which noline could be established (Fig. 1B [tni HoxAa#2]). The early innervationpattern of the hind limbs was investigated in 12.5 days old fetuses(E12.5), using immune-staining of neurofilaments (Fig. 1D). In trans-genic mice, an abnormal truncation of the peroneal nerve, which inner-vates the dorsal aspect of the hind limbs, was observed consistent withthe locomotory abnormalities detected in thesemice. Both the neuronaland reproductive phenotypes proved very similar to those observed forthe loss of function of Hox10 group genes. These latter mutants indeeddisplay a misspecification of the sciatic part of the lumbosacral plexus,which normally innervates the hind limbs (Carpenter et al., 1997;Tarchini et al., 2005; Wu et al., 2008).
2.2. Gain of function of Tetraodon Hoxa11a
In order to associate the phenotypic abnormalities observed in thetransgenic linewith thepotential expression of the transgenic TetraodonHox genes present in the BAC,we performed in situ hybridization for theTetraodon Hoxa9a to Hoxa13a genes (Fig. 2). Interestingly, these geneswere expressed during mouse development with the expected spatialcollinear pattern along the main body axis, with Hoxa9a beingexpressed most anteriorly and Hoxa13a being confined to the posteriortail region. Comparison with the expression of the endogenous mouseHoxa genes however, indicated the presence of transgenic Hoxa11atranscripts at a too anterior position, i.e. about three to four somitesmore anterior than the corresponding pattern for the mouse Hoxa11gene. Such an anteriorized transcriptional pattern associated with aHox11 gene was previously reported, associated with the replacementin vivo of an endogenous Hoxd11 enhancer by its teleost counterpart.Mice carrying this fish enhancer at the correct endogenous positionexpressed their own Hoxd11 gene too anteriorly (Gerard et al., 1997).Therefore, an anterior gain of function of the Tetraodon Hoxa11a geneas reported heremay reflect a specific cis-regulatory difference betweenfish and mammals, as somewhat supported by the high divergence innon-coding DNA sequences between the fish and tetrapods posterior

L1 T13
S1
T13 L1
S1
T13
S1
L1
tni H
oxA
a
wild
type
L3
L6
S3
CD2
tni H
oxA
a
wild
type
dorsal ventral
wildtypezoom dorsal
tni HoxAa
B
A C
D
tni HoxAa #1 tni HoxAa #2 wildtype
wildtype tni HoxAa
thoracic 13 13
lumbar 5/6 3
sacral 5 5
Fig. 1. Paralysis, sacral transformations and absence of the peroneal nerve in the hind limbs of the tniHoxAa transgenic line. A)Habitus ofwild type and tniHoxAa transgenicmice. Note thehind limbparalysis, as visible fromadorsal view. B) Skeletal staining using alizarin red oneitherwild type specimen, tniHoxAa established line (labeled#1) and an independently obtainedF0 transgenic male who never gave rise to offspring (labeled #2). Both transgenic animals displayed a shortening of the lumbar region, from the normal L6 (sometimes L5) formula ob-served inwild typemice (awild type specimenwith L6 phenotype is shown) to a L3 formula. Partial sacral transformations in the 2nd lumbar vertebra are indicatedwith red arrowheads(also present but not indicated for the L3). The observed changes in the axial formula are indicated in the table shown underneath the skeletal preparations. C) Comparison between ver-tebral morphologies shows that the posterior most lumbar vertebrae (L3) in the tni HoxAa skeleton have partial transformations into sacral and/or caudal vertebrae, as indicated by thebroader andmore horizontally oriented lateral processes. D) Neurofilament staining on E12.5 hind limb buds shows the absence of the peroneal nerve in tniHoxAa embryos. The peronealnerve innervates the dorsal aspect of the limbs and is indicatedwith a black arrowhead in the dorsal and the lateral view of the wild type limb bud. Its vacant position is indicated using agray arrowhead in the tni HoxAa transgenic limb buds. The ventral view shows no apparent mis-specification of the tibial nerve, which innervates the ventral aspect of the hind limbs. Azoom on the peroneal nerve and its vacant position in the transgenic limb bud is shown in the lower panel. T13: 13th thoracic vertebra; L1: 1st lumbar vertebra; S1: 1st sacral vertebra; L3:3rd lumbar vertebra; L6: sixth lumbar vertebra; S3: 3rd sacral vertebra; CD2: 2nd caudal vertebra.
66 J.M. Woltering, D. Duboule / Mechanisms of Development 138 (2015) 64–72
HoxA clusters (Supplementary Fig. 2A). This anteriorized expressionwas equally observed at earlier stages (E10.5). In such embryos, thetransgenic Hoxa11a gene was expressed more anteriorly than thethree mouse paralogous genes Hoxa11, Hoxc11 and Hoxd11 (Fig. 3A).
This ‘anteriorized’ Hoxa11a transcript domain coincided in spacewith the observed sacral homeotic transformations of the lumbar re-gion. In addition, the morphological transformations, which werescored precisely within this very region where Hoxa11a was gained,gave the expected phenotype for an ectopic expression of a tetrapodHox11 group gene.Hox11 genes indeed determine sacral vertebral iden-tity (Wellik and Capecchi, 2003) and their expression at ectopic anteriorpositionswas shown to produce sacral transformations (Carapuco et al.,2005; Gerard et al., 1997). Likewise, the ectopic expression of Hoxd11induced themis-specification of themotor neurons innervating thedor-sal aspects of hind limbs (Misra et al., 2009), a nerve pattern normallyspecified by Hox10 genes (Carpenter et al., 1997). Noteworthy, the spa-tial expression patterns of the endogenousHox11 group genes were notnoticeablymodified in transgenic embryos (Fig. 3A), ruling out a poten-tial auto-regulatory interaction between HOX11 proteins and their ownpromoters as a cause of the observed phenotypic alterations. Therefore,we concluded that the mis-expression of the Tetraodon Hoxa11a genewas directly responsible for the severe abnormalities detected in thesetransgenic animals. In this view, the anteriorization of the axial Hox11
domain, due to the expression of the transgenic Tetraodon Hoxa11agene, induced both neuronal and vertebral homeosis.
2.3. Hind-limb bud formation and Hox-mediated axial patterning
The anterior shift of the sacrum observed in our Tetraodon HoxAatransgenic line was accompanied by a more anterior positioning of thehind-limbs along the body axis, as illustrated by the positioning of thehip joint (Supplementary Fig. 1). In newborn mice, an evaluation ofthe approximate hind-limb position indicated a two to three somitesoffset when compared to wild type specimen, consistent with the ex-tent of the shift observed for the sacrum and thus suggesting that theentire posterior part of the animal had been shifted anteriorly. The posi-tion of forelimb buds seems to be determined by the combination ofHOX proteins found in the lateral plate mesoderm (Nishimoto et al.,2014). Also, hind limb buds were shifted two somites backwards inmicemutant for multiple group 9Hox genes (McIntyre et al., 2007). De-spite these reports, however, our observationwas unexpected since thedetermination of hind limb positioning along the trunk was recentlysuggested to be a Hox independent process (Jurberg et al., 2013), asassessed by both loss- and gain of function approaches. In this lattercase, even theHoxa11 genewas unable to shift hind limb buds anterior-ly when overexpressed at an early stage (Jurberg et al., 2013). Potential

Fig. 2. Expression of mouse HoxA and Tetraodon HoxAa genes in either wild type or tni HoxAamice. In situ hybridization was performed on E11.5 wild type and transgenic embryos (ge-notypes are indicated on the right hand side).Wild type embryos were analyzedwithmouse specific probes to visualize the expression of the endogenousHoxa genes andwith Tetraodonspecific probes to exclude potential cross reactivity with the endogenous mouse genes. Tni HoxAa transgenic embryos were processed for Tetraodon specific probes. Probe names areindicated above as well as whether the probe used was for a wild type or transgenic specimen (indicated on the left hand side). The expression pattern of the Tetraodon probes in thetransgenic context shows the expected collinear pattern with Hoxa9a being expressed most anteriorly (although not as far as the endogenous mouse Hoxa9) and Hoxa13a restricted tothe posterior most tail. There is however marginal differentiation, if any, between the anterior expression limits of Hoxa10a and Hoxa11a. A clear difference is observed in expression be-tween mouse Hoxa11, which has an anterior expression limit close to the posterior limit of the hind limb buds, and the Tetraodon Hoxa11a, that has a limit around three to four somitesmore anterior, coinciding with the anterior limit of the hind limb buds (the anterior level of axial expression in both wild type and tni HoxAa panels is indicated with a dotted line). Thisparticular area where the difference is observed is the part of the bodywhere sacral transformations are scored (Fig. 1). The lower row shows that there is no cross reactivity in the in situhybridization between the Tetraodon probes and the endogenous mouse genes.
67J.M. Woltering, D. Duboule / Mechanisms of Development 138 (2015) 64–72
reasons for this apparent discrepancy such as differences in the strengthof transgene expression in the lateral plate mesoderm (Fig. 3A) arediscussed below.
We further investigated if any alteration in hind limb positioningcould already be observed at the stage of limb bud formation (E10.5)and hence whether additional evidence could be found to document adirect link between hind limb induction on the one hand, and the sacral‘Hox code’, on the other hand. To better evaluate this parameter, somiteswere visualized by using DAPI staining in combination with fluorescentin situ hybridization for the MyoD transcripts (Hebrok et al., 1997).Analysis of E10.5 transgenic embryos showed that the hind limb budsdeveloped at an axial position comparable to the wild type situation,i.e. at around somite level 25 (Fig. 3B). Therefore, the anteriorized posi-tioning of hind limbs in adult mice was likely the meremanifestation ofan anterior shift of the entire sacral region, rather than of a local changein the positioning of the limb field, the territory from where limbs budout of the lateral plate mesoderm.
It has been noticed (e.g. Burke et al., 1995) that hind limbs initiallyemerge at a body level corresponding to lumbar somites. Subsequently,these buds adopt a more caudal position, at the level of the sacralsomites, where they will become attached through the pelvic-sacralconnection. This relative shift of the buds with respect to the somitesmay reflect amore general offset between paraxial and lateral plateme-soderm in the growing embryo. The possibility thus exists that the sa-crum itself provides a cue for an arrest of this relative posterior shift ofthe hind limbs. In such a case, animals displaying a genetically ‘sacral-ized’ lumbar region may concomitantly show anteriorized hind limbs,due to a premature termination of this relative transition. In this view,the more anterior location of hind limbs in adult mice would be anindirect result of repositioning the sacrum, rather than be caused bythe direct influence of ectopic Hoxa11 upon hind limb bud induction.
This anteriorization of the hind limbs becomes apparent at E11.5 al-ready, when neuro-filament staining revealed an axial position corre-sponding to the innervation of spinal nerves 22 to 24 in transgenic
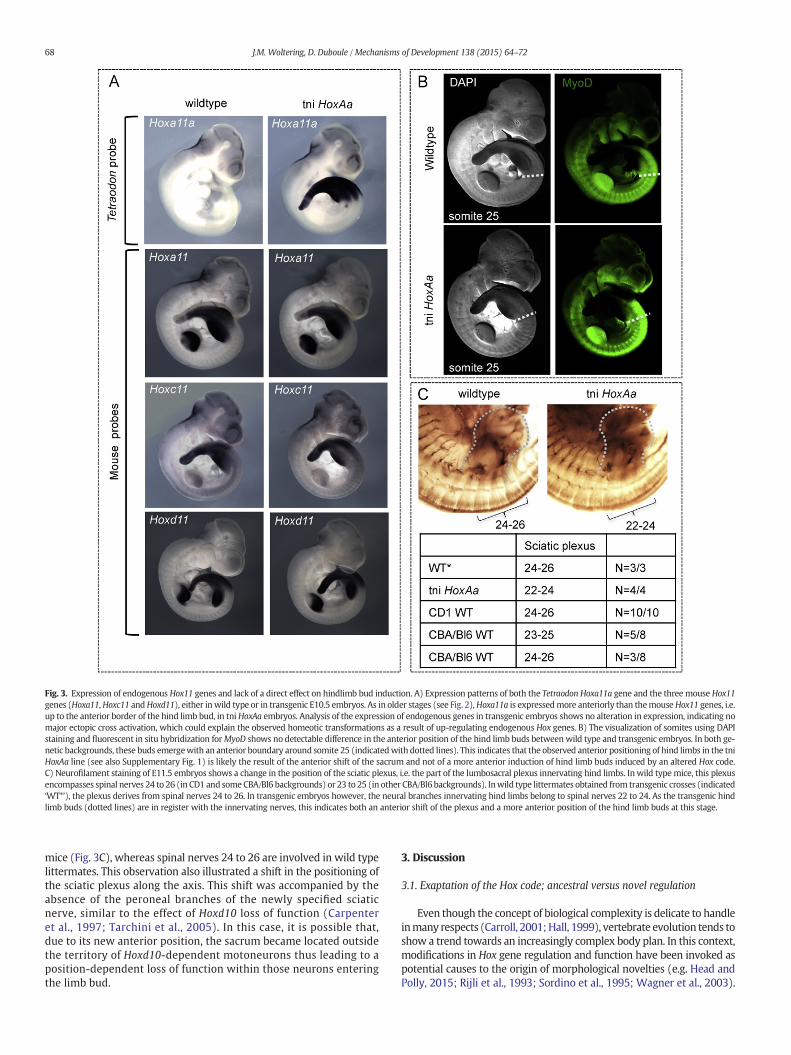
Fig. 3. Expression of endogenous Hox11 genes and lack of a direct effect on hindlimb bud induction. A) Expression patterns of both the Tetraodon Hoxa11a gene and the three mouse Hox11genes (Hoxa11,Hoxc11 andHoxd11), either inwild type or in transgenic E10.5 embryos. As in older stages (see Fig. 2),Hoxa11a is expressedmore anteriorly than themouseHox11 genes, i.e.up to the anterior border of the hind limb bud, in tniHoxAa embryos. Analysis of the expression of endogenous genes in transgenic embryos shows no alteration in expression, indicating nomajor ectopic cross activation, which could explain the observed homeotic transformations as a result of up-regulating endogenous Hox genes. B) The visualization of somites using DAPIstaining and fluorescent in situ hybridization forMyoD shows no detectable difference in the anterior position of the hind limb buds betweenwild type and transgenic embryos. In both ge-netic backgrounds, these buds emergewith an anterior boundary around somite 25 (indicatedwith dotted lines). This indicates that the observed anterior positioning of hind limbs in the tniHoxAa line (see also Supplementary Fig. 1) is likely the result of the anterior shift of the sacrum and not of a more anterior induction of hind limb buds induced by an altered Hox code.C) Neurofilament staining of E11.5 embryos shows a change in the position of the sciatic plexus, i.e. the part of the lumbosacral plexus innervating hind limbs. Inwild type mice, this plexusencompasses spinal nerves 24 to 26 (in CD1 and someCBA/Bl6 backgrounds) or 23 to 25 (in other CBA/Bl6backgrounds). Inwild type littermates obtained from transgenic crosses (indicated‘WT*’), the plexus derives from spinal nerves 24 to 26. In transgenic embryos however, the neural branches innervating hind limbs belong to spinal nerves 22 to 24. As the transgenic hindlimb buds (dotted lines) are in register with the innervating nerves, this indicates both an anterior shift of the plexus and a more anterior position of the hind limb buds at this stage.
68 J.M. Woltering, D. Duboule / Mechanisms of Development 138 (2015) 64–72
mice (Fig. 3C), whereas spinal nerves 24 to 26 are involved in wild typelittermates. This observation also illustrated a shift in the positioning ofthe sciatic plexus along the axis. This shift was accompanied by theabsence of the peroneal branches of the newly specified sciaticnerve, similar to the effect of Hoxd10 loss of function (Carpenteret al., 1997; Tarchini et al., 2005). In this case, it is possible that,due to its new anterior position, the sacrum became located outsidethe territory of Hoxd10-dependent motoneurons thus leading to aposition-dependent loss of function within those neurons enteringthe limb bud.
3. Discussion
3.1. Exaptation of the Hox code; ancestral versus novel regulation
Even though the concept of biological complexity is delicate to handleinmany respects (Carroll, 2001;Hall, 1999), vertebrate evolution tends toshow a trend towards an increasingly complex body plan. In this context,modifications in Hox gene regulation and function have been invoked aspotential causes to the origin of morphological novelties (e.g. Head andPolly, 2015; Rijli et al., 1993; Sordino et al., 1995; Wagner et al., 2003).

69J.M. Woltering, D. Duboule / Mechanisms of Development 138 (2015) 64–72
Within such a conceptual framework, two distinct regulatory levels canbe identified where important changes in the function of Hox genesmay have occurred. The first case is well illustrated by neomorphic struc-tures such as digits and external genitals, which evolved concomitantlypartly via the recruitment of novel Hox expression domains (Dolle et al.,1991; Montavon et al., 2008). These regulatory specificities were ac-quired through both the emergence of new enhancer sequences andthe co-optation of regulatory sequences already used for another purpose(Lonfat et al., 2014; Montavon et al., 2011). In such cases, morphologicalnovelties coincide with regulatory innovations, even when the latter arebased upon pre-existing modules (Lonfat et al., 2014; Woltering et al.,2014). Likewise, modifications in the organization of the spine mayhave relied upon the mere displacement of Hox expression boundaries,as illustrated by the different positions of the cervico–thoracic transitionsin mammals and birds (Burke et al., 1995; Gaunt, 1994).
In an alternative – yet not exclusive – scenario, morphological nov-elties can evolve not only as the result of new regulatory modalitiesbut, rather, through different responses from the systems downstreamof Hox control, for example via the loss or gain of target genes. For in-stance amodification in the response of a target gene following changesin binding sequence can lead to Hox-derived phenotypic alterations(Guerreiro et al., 2013). Also, variations in the HOX protein structurescould potentially lead to important quantitative and/or qualitativemod-ifications in large sets of target loci, as shown for example in insectswhere a modification of the Ubx protein is thought to have accompa-nied the emergence of the hexapod body plan (Ronshaugen et al.,2002). Interestingly, changes in the Hoxa11 coding sequence wereshown to have paralleled the evolution of pregnancy in mammals(Lynch et al., 2008). In addition, sequence analyses of this gene invarious species showed potential signatures of adaptive sequencechange across the fin-to-limb transition (Chiu et al., 2000). The compar-ison between the mouse HOXA11 and Tetraodon HOXA11A sequencesindeed shows a very strong conservation of the homeodomain peptidesequence, whereas the N-terminal parts of the proteins are muchmore divergent (Supplementary Fig. 2B). However, despite this diver-gence in protein sequences outside the homeodomain, the pufferfishHoxa11a protein was clearly capable of inducing the sacrum, a well-defined tetrapod novelty and hence this protein already had the capac-ity to control sacral characteristicswell before the evolution of a sacrum.Consequently, modifications in its coding sequencewere likely not nec-essary for this functional exaptation.
How other features associated with the vertebrates evolved ordisappeared, and to which degree the above patterning concepts mayprovide explanatory frameworks for these key events remains to be in-vestigated. Structures that would be of particular interest in this contextwould be for example the occipital–synarcual complex found in carti-laginous fishes and extinct placoderms (Davis et al., 2012; Soshnikovaet al., 2013), the ostariophysan Weberian vertebrae (Bird and Mabee,2003), the teleost specific homocercal caudal fin vertebrae (Moriyamaand Takeda, 2013), the anuran urostyle (Rockova and Rocek, 2005),the snake's forked lympapophyses (Woltering, 2012) or the enigmaticinterlocking lumbar vertebrae of hero shrew (Stanley et al., 2013).
It is also noteworthy that the general principle of collinear distribu-tion of Hox expression domains along the rostro-caudal body axis ob-served in vertebrates (Gaunt et al., 1988) largely predates theirappearances (Duboule and Dolle, 1989; Graham et al., 1989; Lewis,1978) and thus could not be initially associated with the evolution ofa complex axial skeleton. It is possible that theHox systemwas original-ly linked to the organization of either neural (Deutsch and Le Guyader,1998) or endodermal (Kondo et al., 1996) structures along the APaxis, and was subsequently recruited by mesodermal derivatives dueto its capacity to specialize particular body segments. From there on-wards, modifications in either the regulation and structures of thesegenes, or of their specific targets (either at the regulatory or at the func-tional level) may have produced the variety of vertebral formulaeknown today in extant vertebrates and in the fossil record.
3.2. The Hox constraint on the body plan
This diversity in vertebral formulae is however not infinite and theobserved anatomical bias towards certain prototypes at the detrimentof others, is hard to explain on pure adaptive grounds. Therefore, it islikely that developmental constraints restrict the realm of possibilitiesfor a vertebral column to combine and associate various vertebral types,both in their number and qualities (Asher et al., 2011). The identificationof the developmental processes associated with these constrainedmorphologies has been problematic and without empirical support. Themouse line we describe in this study displays a lumbar region with onlythree vertebraewithout a concomitant increase in the number of thoracicvertebrae. As such, it ‘violates’ the thoracolumbar constraint identified byNarita and Kuratani (2005) andmay help thinking about both the natureof the underlying constraints and the mechanisms involved.
In our mutant mice, the reduction in the number of thoracolumbarvertebrae is caused by the anterior expression of the transgenicHoxa11a gene, presumably as a result of a different interpretation ofcis-regulatory information between the fish and the mouse contexts aspreviously noted (Gerard et al., 1997). The resulting transformation ofthe posterior lumbar region into a sacrum likely induced a morpholog-ical offset between structures and/or cell types, leading to the severephenotypic condition observed. In the case of hind limb positioning,this offset seems to be compensated for, to some extent, since the finalposition of the hind limbs is in register with the anteriorized sacrum.This could be due to an instructive relationship between both struc-tures, the sacrum helping to position the hind limbs. Alternatively, thestrong gain of function of Hoxa11a observed in the lateral plate meso-derm may directly participate in the rostral positioning of the hindlimbs, even though Hoxa11was apparently not able to achieve a similarresult in another experimental context (Jurberg et al., 2013).
In contrast, the differentiation of the appropriate columns of motorneurons innervating the limbs, which is greatly influenced by Hoxgene expression in the developing spinal cord (Jung et al., 2014), maynot have been equally influenced by our gain of function. As a result,the motor neurons innervating the mutant hind limbs may originatefrom a territory of the spinal cord lacking the proper combination ofHOX proteins, in particular HOXD10, the targeted or spontaneousmuta-tion of which generates similar paraplegia phenotypes (Carpenter et al.,1997; Misra et al., 2009; Tarchini et al., 2005; Wahba et al., 2001). Thiscase potentially illustrates the result of interfering with the collineardistribution of HOX proteins, having Hoxa11a transcripts abruptly pro-duced more anteriorly (or at the same body level; see Fig. 2) than thenative Hoxd10mRNAs. The resulting compromised locomotion and ac-companying reproductive incapacitation would of course be stronglyselected against under natural conditions.
3.3. Mechanisms and constraints underlying the phylotypic progression
In vertebrates, the general body architecture (Bauplan) materializesduring late gastrulation,where all vertebrate embryos tend to share im-portant morphological features (the zootype). This ‘phylotypic stage’was associated with the expression of particular transcription factors,including Hox genes (Slack et al., 1993). It was subsequently arguedthat early embryos, regardless of their various modes of gastrulationhad to ‘converge’ towards this particular body plan to set the generalground from which different adult morphological traits can be subse-quently derived. This ‘Hourglass model’ (Duboule, 1994) implies thatembryos progress through a short period (the phylotypic progression)where the underlying developmental mechanisms must be maximallyconstrained, thus making alternative solutions impossible. While theexistence of the developmental hourglass has been recently document-ed in a variety of contexts (see e.g. Irie and Kuratani, 2014) but also(Richardson, 1995) the nature of the constraints responsible for thephylotypic progression (into the bottleneck of the hourglass) is elusive.

70 J.M. Woltering, D. Duboule / Mechanisms of Development 138 (2015) 64–72
Initially, two kinds of constraints were proposed: On the one hand,constraints based on meta-trans regulations (Duboule, 1994), i.e. dueto particular interactions between networks of genes, necessary at thisstage to properly set up the body plan (Raff, 1996). On the other hand,a single mechanism may underlie the passage through the hourglassbottleneck, provided that this mechanism is invariable and requires aparticular context to be implemented. In this view, early meta-transregulations are necessary to bring the developing system into a pointwhere this invariable mechanism can now operate, during thephylotypic period. The fact that this period covers the extension andpatterning of the rostro-caudal axis suggested that genetic mechanismsat work to orchestrate these critical steps might be particularlyconstrained, for some reasons. Hox genes are the major players in thepatterning of the body axis (seeMallo et al., 2010) and possibly in its ex-tension (Denans et al., 2015; Di-Poi et al., 2010; Young et al., 2009) andtheir sequential activation (temporal collinearity) relies upon ameta-cismechanism (the Hox clock) that appears difficult to evolve as it reliesupon a process that reads the linearity of DNA at the Hox loci (seeNoordermeer and Duboule, 2013). On this ground, the Hox clock wasproposed as the major constraint acting during the phylotypic pro-gression (Duboule, 1994). Subsequently, the Hox clock was reportedto closely interact with the segmentation clock, the mechanismwhereby the vertebrate body becomes segmented (Palmeirim et al.,1997). Hox genes indeed can be regulated as a read out of the seg-mentation clock (Zakany et al., 2001) and, conversely, the amountof caudal non-segmented mesoderm available as substrate for thesegmentation clock may be regulated by the Hox clock (Denanset al., 2015). Therefore, a perfect coordination between these twoprecise mechanisms must be secured and hence the vertebrateembryo may have to converge towards this point where both clockswill click in concert for about two days, before the system relaxes,giving more opportunities again for variable interactions betweengene networks.
Offsets between these two clocks can naturally lead to the variety ofvertebral formulae found in vertebrates. Here, we show an example ofhow the anteriorization of a single Hox11 group gene can lead to a com-plete reorganization of the spine. However, this gain of function did notrespect the constraint applied to the system, as it was not accompaniedby a gain of function of the entire set ofHox genes, bringingHoxa11a ex-pression at the rostral position of Hox10 genes. While the effect on thespinemay not have been in itself a cause of counter selection, the inter-actions of body partswith the spinewas affected in spite of some intrin-sic re-organization, such as the anteriorized position of the hind limbs.This suggests the existence of developmental check-points where therelative connections between various structures can be adjusted. Thehind legs were nevertheless not properly innervated, likely due to theabsence of Hox10-positive/Hox11-negative motor neurons, which arerequired for the peroneal branch of the sciatic nerve (Carpenter et al.,1997; Tarchini et al., 2005).
It has also been argued that high developmental constraints mayapply at the phylotypic stage due to the necessity to coordinate interac-tions between the various nascent tissue types (Galis and Metz, 2001;Raff, 1994; Sander, 1983). In other words, to produce a coherent organ-ism, tissues that differentiate within spatially segregated embryonic do-mains during gastrulation (such as for instance neurons, vascularprogenitors or muscles) need to be integrated at this stage. However,a common landmark to the differentiation of these tissues or celltypes, aswell as of the origin of allmajor internal organs, is their originalposition along the developing rostral to caudal axis, suggesting that thislatter information may be the critical determinant. Accordingly, themechanism(s) underlying the organization of the major body axismay be more constrained than others. In this context, our mouse lineillustrates this subtle equilibriumwhereby several patterning processesmust act in concert during thephylotypic stage. The fact thatmotor neu-ron specification, vertebral patterning and hind limb positioning are outof register shows that these processes are not naturally interconnected
with one another, which may re-enforce the constraints acting at thisstage upon the evolvability of the body plan.
Clades outside mammals can show high diversity in both the num-ber of pre-caudal vertebra and the positioning of the posterior pairedappendages along thebody. In this regard, the teleost fish are particular-ly interesting as they can have their pelvic fins positioned as far anterioras their pectoral fins (Murakami and Tanaka, 2011; Murata et al., 2010;Tanaka, 2011). In fishes the initial position of the pelvic fins is specifiedby GDF11, as in vertebrates (see Jurberg et al., 2013), generally at theposition of the trunk to tail transition (Murata et al., 2010). In some spe-cies, however, the pelvic fin buds subsequentlymigrate towards the an-terior, along the trunk. The appropriate innervation of these anteriorlydisplaced pelvic fins occurs through locally exiting motor neurons,which are thus apparently rather independent from the combinationof Hox genes they express (Murakami and Tanaka, 2011; Murata et al.,2010; Tanaka, 2011), perhaps due to a lesser complexity in the realmof movements implemented by these fins. This great flexibility in thefish body plan may be accounted for by a lower interdependence be-tween specific motor neurons and hind limbs such that fishes may notneed to tightly orchestrate the connection between a specific set of neu-rons and the pelvic fins to secure a proper functional outcome. The evo-lution of the sacrum and of a more generic connection between theposterior appendages and the spine, may have introduced yet anotherstrong constraint, thus further decreasing the evolvability of the bodyplan.
4. Materials and methods
4.1. Mouse strains
Themouse tniHoxAa stock (Woltering et al., 2014)was generated bypronuclear injection following well-established procedures. All experi-ments were performed in agreementwith the Swiss law on animal pro-tection (LPA)with the appropriate legal authorization to D.D. Because ofthe severity of the phenotype, this transgenic line is no longer main-tained as living animals.
4.2. In situ hybridizations
Whole mount in situ hybridizations using mouse and Tetraodonprobeswere performed as described previously, using 1.3× SSC concen-tration in the hybridization buffer to prevent cross reactivity betweenmouse RNAs and Tetraodon probes (Woltering et al., 2009, 2014).Unpublished probes for Tetraodon Hoxa9a and Hoxa10a correspond tosequences within exon 1 and were cloned from BAC DNA using the fol-lowing primers:
a9a-FW; ATGTCGACATCCGGAACGCTG
a9a-RV; TTGGATCGAGGCCTGGTTTCTCa10a-FW; ATGGCATGTTCGGACACCC,a10a-RV CTTTGGGGCCCTTGGCTGCAC.
The mouse Hoxc11 probe was cloned from genomic DNA using thefollowing primers:
FW; AACCGGACGAGCTGGGATTC
RV; AGACTAAGACGGATAACGCG.
Fluorescent in situ hybridization for the MyoD probe (Hebrok et al.,1997) was performed using staining with Fluorescein Thyramid Ampli-fication System (Perkin-Elmer). Antibody staining for neurofilamentswas carried out as described previously (Tarchini et al., 2005) usinganti-NF160 (clone NN18, Sigma N-5264) and anti-mouse Ig Fab HRPconjugate (Sigma A-3882). For DAPI staining, embryos were incubatedwith 0.1 mM DAPI in TBS-T after in situ hybridization. Whole mountin situ images in Figs. 2 and 3 were constructed as overlays of gray-

71J.M. Woltering, D. Duboule / Mechanisms of Development 138 (2015) 64–72
scale DAPI fluorescent images and bright field images using AdobePhotoshop.
4.3. In silico sequence analysis
Analysis of non-coding regions in the Hoxa clusters was doneusing LAGAN-VISTA (http://genome.lbl.gov/vista/lagan/submit.shtml)(Frazer et al., 2004). Alignment of HOXA11 and HOXA11A proteinswas done at EMBL (http://www.ebi.ac.uk/Tools/psa/emboss_needle/).Conserved domains were predicted using NCBI's conserved domain da-tabase (CDD) (http://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)(Marchler-Bauer et al., 2015).
Supplementary data to this article can be found online at http://dx.doi.org/10.1016/j.mod.2015.07.006.
Acknowledgments
We would like to thank all members of the Duboule laboratories. Inparticular Joska Zakany for useful discussion and sharing of reagents andFabienne Chabaud and BénédicteMascrez for helpwith animal care andimplementation of legal guidelines.We also thankMoisésMallo for pro-viding constructs. This work was supported by funds (to D.D.) from theUniversity of Geneva, the EPFL, the Swiss National Research Fund(SNSF) (No. 310030B_138662), the European Research Council (ERC;SystemsHox.ch, No. 232790) and the Claraz Foundation.
References
Asher, R.J., Lin, K.H., Kardjilov, N., Hautier, L., 2011. Variability and constraint in themammalian vertebral column. J. Evol. Biol. 24, 1080–1090.
Bird, N.C., Mabee, P.M., 2003. Developmental morphology of the axial skeleton of thezebrafish, Danio rerio (Ostariophysi: Cyprinidae). Dev. Dyn. 228, 337–357.
Buchholtz, E.A., 2014. Crossing the frontier: a hypothesis for the origins of meristicconstraint in mammalian axial patterning. Zoology (Jena) 117, 64–69.
Buchholtz, E.A., Bailin, H.G., Laves, S.A., Yang, J.T., Chan, M.Y., Drozd, L.E., 2012. Fixedcervical count and the origin of the mammalian diaphragm. Evol. Dev. 14, 399–411.
Burke, A.C., Nelson, C.E., Morgan, B.A., Tabin, C., 1995. Hox genes and the evolution ofvertebrate axial morphology. Development 121, 333–346.
Carapuco, M., Novoa, A., Bobola, N., Mallo, M., 2005. Hox genes specify vertebral types inthe presomitic mesoderm. Genes Dev. 19, 2116–2121.
Carpenter, E.M., Goddard, J.M., Davis, A.P., Nguyen, T.P., Capecchi, M.R., 1997. Targeteddisruption of Hoxd-10 affects mouse hindlimb development. Development 124,4505–4514.
Carroll, R., 1988. Vertebrate Paleontology and Evolution. W.H. Freeman and company,New York, pp. 361–400.
Carroll, S.B., 2001. Chance and necessity: the evolution of morphological complexity anddiversity. Nature 409, 1102–1109.
Carroll, R., Holmes, R., 2007. Evolution of the appendicular skeleton of amphibians. In:Hall, B. (Ed.), Fins Into Limbs. The University of Chicago Press, Chicago, pp. 185–224.
Casaca, A., Santos, A.C., Mallo, M., 2014. Controlling Hox gene expression and activity tobuild the vertebrate axial skeleton. Dev. Dyn. 243, 24–36.
Chiu, C.H., Nonaka, D., Xue, L., Amemiya, C.T., Wagner, G.P., 2000. Evolution of Hoxa-11 inlineages phylogenetically positioned along the fin-limb transition. Mol. Phylogenet.Evol. 17, 305–316.
Davis, S.P., Finarelli, J.A., Coates, M.I., 2012. Acanthodes and shark-like conditions in thelast common ancestor of modern gnathostomes. Nature 486, 247–250.
Denans, N., Iimura, T., Pourquie, O., 2015. Hox genes control vertebrate body elongationby collinear Wnt repression. Elife 4.
Deschamps, J., van Nes, J., 2005. Developmental regulation of the Hox genes during axialmorphogenesis in the mouse. Development 132, 2931–2942.
Deutsch, J., Le Guyader, H., 1998. The neuronal zootype. An hypothesis. C. R. Acad. Sci. III321, 713–719.
Di-Poi, N., Montoya-Burgos, J.I., Miller, H., Pourquie, O., Milinkovitch, M.C., Duboule, D.,2010. Changes in Hox genes' structure and function during the evolution of the squa-mate body plan. Nature 464, 99–103.
Dolle, P., Izpisua-Belmonte, J.C., Brown, J.M., Tickle, C., Duboule, D., 1991. HOX-4 genesand the morphogenesis of mammalian genitalia. Genes Dev. 5, 1767–1776.
Duboule, D., 1994. Temporal colinearity and the phylotypic progression: a basis for thestability of a vertebrate Bauplan and the evolution of morphologies throughheterochrony. Development 135–142.
Duboule, D., Dolle, P., 1989. The structural and functional organization of the murine HOXgene family resembles that of Drosophila homeotic genes. EMBO J. 8, 1497–1505.
Duboule, D., Morata, G., 1994. Colinearity and functional hierarchy among genes of thehomeotic complexes. Trends Genet. 10, 358–364.
Duboule, D., Wilkins, A.S., 1998. The evolution of ‘bricolage’. Trends Genet. 14, 54–59.Frazer, K.A., Pachter, L., Poliakov, A., Rubin, E.M., Dubchak, I., 2004. VISTA: computational
tools for comparative genomics. Nucleic Acids Res. 32, W273–W279.
Galis, F., 1999. Why do almost all mammals have seven cervical vertebrae? Developmen-tal constraints, Hox genes, and cancer. J. Exp. Zool. 285, 19–26.
Galis, F., Metz, J.A., 2001. Testing the vulnerability of the phylotypic stage: on modularityand evolutionary conservation. J. Exp. Zool. 291, 195–204.
Galis, F., Metz, J.A., 2007. Evolutionary novelties: the making and breaking of pleiotropicconstraints. Integr. Comp. Biol. 47, 409–419.
Galis, F., Carrier, D.R., van Alphen, J., van der Mije, S.D., Van Dooren, T.J., Metz, J.A., tenBroek, C.M., 2014. Fast running restricts evolutionary change of the vertebral columnin mammals. Proc. Natl. Acad. Sci. U. S. A. 111, 11401–11406.
Gaunt, S.J., 1994. Conservation in the Hox code duringmorphological evolution. Int. J. Dev.Biol. 38, 549–552.
Gaunt, S.J., Sharpe, P.T., Duboule, D., 1988. Spatially restricted domains of homeo-genetranscripts in mouse embryos: relation to a segmented body plan. Development104, 169–179.
Gerard, M., Zakany, J., Duboule, D., 1997. Interspecies exchange of a Hoxd enhancer in vivoinduces premature transcription and anterior shift of the sacrum. Dev. Biol. 190, 32–40.
Graham, A., Papalopulu, N., Krumlauf, R., 1989. The murine and Drosophila homeoboxgene complexes have common features of organization and expression. Cell 57,367–378.
Guerreiro, I., Nunes, A., Woltering, J.M., Casaca, A., Novoa, A., Vinagre, T., Hunter, M.E.,Duboule, D., Mallo, M., 2013. Role of a polymorphism in a Hox/Pax-responsive en-hancer in the evolution of the vertebrate spine. Proc. Natl. Acad. Sci. U. S. A. 110,10682–10686.
Hall, B.K., 1999. Evolutionary Developmental Biology. Kluwer Academic Publishers,Dordrecht, Netherlands.
Head, J.J., Polly, P.D., 2015. Evolution of the snake body form reveals homoplasy inamniote Hox gene function. Nature 520, 86–89.
Hebrok, M., Fuchtbauer, A., Fuchtbauer, E.M., 1997. Repression ofmuscle-specific gene ac-tivation by the murine Twist protein. Exp. Cell Res. 232, 295–303.
Hirasawa, T., Kuratani, S., 2013. A new scenario of the evolutionary derivation of themammalian diaphragm from shoulder muscles. J. Anat. 222, 504–517.
Irie, N., Kuratani, S., 2014. The developmental hourglass model: a predictor of the basicbody plan? Development 141, 4649–4655.
Izpisua-Belmonte, J.C., Falkenstein, H., Dolle, P., Renucci, A., Duboule, D., 1991. Murinegenes related to the Drosophila AbdB homeotic genes are sequentially expressed dur-ing development of the posterior part of the body. EMBO J. 10, 2279–2289.
Jeffs, P.S., Keynes, R.J., 1990. A brief history of segmentation. Semin. Dev. Biol. 1, 77–87.Jung, H., Mazzoni, E.O., Soshnikova, N., Hanley, O., Venkatesh, B., Duboule, D., Dasen, J.S.,
2014. Evolving Hox activity profiles govern diversity in locomotor systems. Dev.Cell 29, 171–187.
Jurberg, A.D., Aires, R., Varela-Lasheras, I., Novoa, A.,Mallo,M., 2013. Switchingaxial progen-itors from producing trunk to tail tissues in vertebrate embryos. Dev. Cell 25, 451–462.
Kessel, M., Gruss, P., 1991. Homeotic transformations of murine vertebrae and concomi-tant alteration of Hox codes induced by retinoic acid. Cell 67, 89–104.
Kirschner, M., Gerhart, J., ebrary Inc., 2005. The Plausibility of Life Resolving Darwin's Di-lemma. Yale University Press, New Haven, p. xiii (314 pp. ill. 24 cm).
Kmita, M., Duboule, D., 2003. Organizing axes in time and space; 25 years of colinear tin-kering. Science 301, 331–333.
Kondo, T., Dolle, P., Zakany, J., Duboule, D., 1996. Function of posterior HoxD genes in themorphogenesis of the anal sphincter. Development 122, 2651–2659.
Lewis, E.B., 1978. A gene complex controlling segmentation in Drosophila. Nature 276,565–570.
Lonfat, N., Montavon, T., Darbellay, F., Gitto, S., Duboule, D., 2014. Convergent evolution ofcomplex regulatory landscapes and pleiotropy at Hox loci. Science 346, 1004–1006.
Lynch, V.J., Tanzer, A., Wang, Y., Leung, F.C., Gellersen, B., Emera, D., Wagner, G.P., 2008.Adaptive changes in the transcription factor HoxA-11 are essential for the evolutionof pregnancy in mammals. Proc. Natl. Acad. Sci. U. S. A. 105, 14928–14933.
Mallo, M.,Wellik, D.M., Deschamps, J., 2010. Hox genes and regional patterning of the ver-tebrate body plan. Dev. Biol. 344, 7–15.
Marchler-Bauer, A., Derbyshire, M.K., Gonzales, N.R., Lu, S., Chitsaz, F., Geer, L.Y., Geer, R.C.,He, J., Gwadz, M., Hurwitz, D.I., Lanczycki, C.J., Lu, F., Marchler, G.H., Song, J.S., Thanki,N.,Wang, Z., Yamashita, R.A., Zhang, D., Zheng, C., Bryant, S.H., 2015. CDD: NCBI's con-served domain database. Nucleic Acids Res. 43, D222–D226.
McIntyre, D.C., Rakshit, S., Yallowitz, A.R., Loken, L., Jeannotte, L., Capecchi, M.R., Wellik,D.M., 2007. Hox patterning of the vertebrate rib cage. Development 134, 2981–2989.
Misra, M., Shah, V., Carpenter, E., McCaffery, P., Lance-Jones, C., 2009. Restricted patternsof Hoxd10 and Hoxd11 set segmental differences in motoneuron subtype comple-ment in the lumbosacral spinal cord. Dev. Biol. 330, 54–72.
Montavon, T., Le Garrec, J.-F., Kerszberg, M., Duboule, D., 2008. Modeling Hox gene regu-lation in digits: reverse collinearity and the molecular origin of thumbness. GenesDev. 22, 346–359.
Montavon, T., Soshnikova, N., Mascrez, B., Joye, E., Thevenet, L., Splinter, E., de Laat, W.,Spitz, F., Duboule, D., 2011. A regulatory archipelago controls Hox genes transcriptionin digits. Cell 147, 1132–1145.
Moriyama, Y., Takeda, H., 2013. Evolution and development of the homocercal caudal finin teleosts. Develop. Growth Differ. 55, 687–698.
Murakami, Y., Tanaka, M., 2011. Evolution of motor innervation to vertebrate fins andlimbs. Dev. Biol. 355, 164–172.
Murata, Y., Tamura, M., Aita, Y., Fujimura, K., Murakami, Y., Okabe, M., Okada, N., Tanaka,M., 2010. Allometric growth of the trunk leads to the rostral shift of the pelvic fin inteleost fishes. Dev. Biol. 347, 236–245.
Narita, Y., Kuratani, S., 2005. Evolution of the vertebral formulae in mammals: a perspec-tive on developmental constraints. J. Exp. Zool. B Mol. Dev. Evol. 304, 91–106.
Nishimoto, S., Minguillon, C., Wood, S., Logan, M.P., 2014. A combination of activation andrepression by a colinear Hox code controls forelimb-restricted expression of Tbx5and reveals Hox protein specificity. PLoS Genet. 10, e1004245.

72 J.M. Woltering, D. Duboule / Mechanisms of Development 138 (2015) 64–72
Noordermeer, D., Duboule, D., 2013. Chromatin architectures and Hox gene collinearity.Curr. Top. Dev. Biol. 104, 113–148.
Noordermeer, D., Leleu, M., Schorderet, P., Joye, E., Chabaud, F., Duboule, D., 2014. Tempo-ral dynamics and developmental memory of 3D chromatin architecture at Hox geneloci. Elife 3, e02557.
Oulion, S., Borday-Birraux, V., Debiais-Thibaud, M., Mazan, S., Laurenti, P., Casane, D.,2011. Evolution of repeated structures along the body axis of jawed vertebrates,insights from the Scyliorhinus canicula Hox code. Evol. Dev. 13, 247–259.
Palmeirim, I., Henrique, D., Ish-Horowicz, D., Pourquie, O., 1997. Avian hairy gene expres-sion identifies a molecular clock linked to vertebrate segmentation andsomitogenesis. Cell 91, 639–648.
Pourquie, O., 2003. The segmentation clock: converting embryonic time into spatialpattern. Science 301, 328–330.
Prince, V.E., Joly, L., Ekker, M., Ho, R.K., 1998. Zebrafish hox genes: genomic organizationand modified colinear expression patterns in the trunk. Development 125, 407–420.
Raff, R.A., 1994. Developmental mechanisms in the evolution of animal form: originsand evolvability of body plans. In: Bengston, S. (Ed.), Early Life on Earth. ColumbiaUniversity Press, New York, pp. 489–500.
Raff, R.A., 1996. The Shape of Life: Genes, Development and the Evolution of Animal.University of Chicago Press, Form.
Richardson, M.K., 1995. Heterochrony and the phylotypic period. Dev. Biol. 172, 412–421.Rijli, F.M., Mark, M., Lakkaraju, S., Dierich, A., Dolle, P., Chambon, P., 1993. A homeotic
transformation is generated in the rostral branchial region of the head by disruptionof Hoxa-2, which acts as a selector gene. Cell 75, 1333–1349.
Rockova, H., Rocek, Z., 2005. Development of the pelvis and posterior part of the vertebralcolumn in the Anura. J. Anat. 206, 17–35.
Romer, S., 1956. Osteology of the Reptiles. Krieger publishing company, Malabar, Florida,pp. 218–297.
Ronshaugen, M., McGinnis, N., McGinnis, W., 2002. Hox protein mutation and macroevo-lution of the insect body plan. Nature 415, 914–917.
Sander, K., 1983. The evolution of patterningmechanisms: gleanings from insect embryo-genesis and spermatogenesis. In: Goodwin, B.C., et al. (Eds.), Development andEvolution. Cambridge University press, Cambridge, pp. 137–159.
Slack, J.M., Holland, P.W., Graham, C.F., 1993. The zootype and the phylotypic stage.Nature 361, 490–492.
Sordino, P., van der Hoeven, F., Duboule, D., 1995. Hox gene expression in teleost fins andthe origin of vertebrate digits. Nature 375, 678–681.
Soshnikova, N., Dewaele, R., Janvier, P., Krumlauf, R., Duboule, D., 2013. Duplications ofhox gene clusters and the emergence of vertebrates. Dev. Biol. 378, 194–199.
Stanley, W.T., Robbins, L.W., Malekani, J.M., Mbalitini, S.G., Migurimu, D.A., Mukinzi, J.C.,Hulselmans, J., Prevot, V., Verheyen, E., Hutterer, R., Doty, J.B., Monroe, B.P.,Nakazawa, Y.J., Braden, Z., Carroll, D., Peterhans, J.C., Bates, J.M., Esselstyn, J.A., 2013.
A new hero emerges: another exceptional mammalian spine and its potentialadaptive significance. Biol. Lett. 9, 20130486.
Tanaka, M., 2011. Revealing the mechanisms of the rostral shift of pelvic fins amongteleost fishes. Evol. Dev. 13, 382–390.
Tarchini, B., Huynh, T.H., Cox, G.A., Duboule, D., 2005. HoxD cluster scanning deletionsidentify multiple defects leading to paralysis in the mouse mutant Ironside. GenesDev. 19, 2862–2876.
van der Hoeven, F., Sordino, P., Fraudeau, N., Izpisua-Belmonte, J.C., Duboule, D., 1996.Teleost HoxD and HoxA genes: comparison with tetrapods and functional evolutionof the HOXD complex. Mech. Dev. 54, 9–21.
Varela-Lasheras, I., Bakker, A.J., van der Mije, S.D., Metz, J.A., van Alphen, J., Galis, F., 2011.Breaking evolutionary and pleiotropic constraints in mammals: on sloths, manateesand homeotic mutations. Evodevo 2, 11.
Wagner, G.P., Amemiya, C., Ruddle, F., 2003. Hox cluster duplications and the opportunityfor evolutionary novelties. Proc. Natl. Acad. Sci. U. S. A. 100, 14603–14606.
Wahba, G.M., Hostikka, S.L., Carpenter, E.M., 2001. The paralogous Hox genes Hoxa10 andHoxd10 interact to pattern the mouse hindlimb peripheral nervous system andskeleton. Dev. Biol. 231, 87–102.
Wellik, D.M., 2009. Hox genes and vertebrate axial pattern. Curr Top Dev Biol 88,257–258.
Wellik, D.M., Capecchi, M.R., 2003. Hox10 and Hox11 genes are required to globallypattern the mammalian skeleton. Science 301, 363–367.
Woltering, J.M., 2012. From lizard to snake; behind the evolution of an extreme bodyplan. Curr. Genom. 13, 289–299.
Woltering, J.M., Vonk, F.J., Muller, H., Bardine, N., Tuduce, I.L., de Bakker, M.A., Knochel,W.,Sirbu, I.O., Durston, A.J., Richardson, M.K., 2009. Axial patterning in snakes and caeci-lians: evidence for an alternative interpretation of the Hox code. Dev. Biol. 332,82–89.
Woltering, J.M., Noordermeer, D., Leleu, M., Duboule, D., 2014. Conservation anddivergence of regulatory strategies at Hox Loci and the origin of tetrapod digits.PLoS Biol. 12, e1001773.
Wu, Y.,Wang, G., Scott, S.A., Capecchi, M.R., 2008. Hoxc10 and Hoxd10 regulate mouse co-lumnar, divisional and motor pool identity of lumbar motoneurons. Development135, 171–182.
Young, T., Rowland, J.E., van de Ven, C., Bialecka, M., Novoa, A., Carapuco, M., van Nes, J., deGraaff, W., Duluc, I., Freund, J.N., Beck, F., Mallo, M., Deschamps, J., 2009. Cdx and Hoxgenes differentially regulate posterior axial growth in mammalian embryos. Dev. Cell17, 516–526.
Zakany, J., Kmita, M., Alarcon, P., de la Pompa, J.L., Duboule, D., 2001. Localized and tran-sient transcription of Hox genes suggests a link between patterning and the segmen-tation clock. Cell 106, 207–217.





























