Embed Size (px)
Citation preview

ISSN 1062�3590, Biology Bulletin, 2010, Vol. 37, No. 4, pp. 357–362. © Pleiades Publishing, Inc., 2010.Original Russian Text © I.V. Kur’ina, Yu.I. Preis, A.A. Bobrov, 2010, published in Izvestiya Akademii Nauk, Seriya Biologicheskaya, 2010, No. 4, pp. 423–429.
357
INTRODUCTION
The first publications concerning the quantitativeecology of sphagnobiont testate amoebae, commonfor bog ecosystems, appeared in the 1990s (Charman,Warner, 1992, 1997; Warner, Charman, 1994). Theshells of testate amoebae, able to remain in peat sedi�ments for a long time, are used as indicators of cli�matic conditions in palaeoecological studies, includ�ing palaeoclimatic reconstructions (Warner, Char�man, 1994; Booth, 2001).
By now the wetness optimums and tolerance limitsfor testate amoebae have been determined in accor�dance with the subsoil water level and the substratewetness in oligotrophic peat moss bogs of the southernand middle taiga of the European part of Russia(Bobrov and Minaeva, 2000; Bobrov et al., 2002;Mazey et al., 2007), Europe (Tolonen et al., 1992;Woodland et al., 1998; Mitchell et al., 1999; Lamen�towicz and Mitchell, 2005; Blundell and Barber,2005), North America (Booth, 2002; Booth and Zyg�munt, 2005), and New Zealand (Charman, 1997).The relation of testate amoebae to the trophic status oftheir habitats still continues to be practicallyunknown.
In the case of oligotrophic soils of Western Siberia,only a few studies of testate amoebae have been carriedout (Rat’kova, 1971; Bobrov et al., 1995; Rakhleeva,2002). The vast territory of marshlands, occupyingmore than 40% of the total area of Western Siberia,determines the diversity of bog formation conditionsand, therefore, the diversity of habitats for testateamoebae inhabiting different bog types.
The purpose of this study was to reveal quantitativeecological characteristics of testate amoebae in rela�tion to different environmental factors (index of wet�
ness, pH, and the trophic status) in the mires of themiddle taiga of Western Siberia by the example of bogsof the Khanty�Mansi Autonomous Area.
MATERIAL AND METHODS
The material was collected in Kondinskie OzeraNational Park, located at the latitude of 60°58′ northand the longitude of 70°11′ east, and in the Kukush�kino bog, located at the latitude of 60°51′ north andthe longitude of 63°31′ east. Both objects were locatedin the middle taiga subzone of Western Siberia(Khanty�Mansi Autonomous Area).
To analyze the communities of testate amoebae, wecollected 33 surface samples from 6 bogs with differingwater regimes and ash levels; all bogs were located inKondinskie Ozera National Park.
A series of microhabitats was selected on the stud�ied bogs, which included tussocks, inter�tussock hol�lows, flat and wet mossy mats, and mire areas. Eachsample represented a surface specimen of sphagnumfrom 5 to 10 cm in depth, which was divided into threeparts to analyze testate amoebae, determine the ashlevel and botanical composition, and measure the pHvalue.
The index of wetness (IW), calculated according tothe botanical composition of the mossy sample, wasconsidered as the characteristic of the watering level ofhabitats (Elina and Yurkovskaya, 1992). The trophicstatus was determined through the ash level value; todo this, samples were dried and burned at 800°Сaccording to the standard procedure (State Stan�dard…, 1985). The pH of the water extract of a mossysample was measured by a HI�8314 pH meter (HannaInstruments, Germany).
Testate Amoebae Inhabiting Middle Taiga Bogs in Western SiberiaI. V. Kur’inaa, Yu. I. Preisa, and A. A. Bobrovb
a Institute of Monitoring of Climatic and Ecologic Systems, Siberian Branch,Russian Academy of Sciences, Tomsk, 63402 Russia
b Department of Soil Sciences, Moscow State University, Moscow, 119899 Russia;e�mail: [email protected]
Received January 26, 2010
Abstract—The population of testate amoebae from the most typical middle taiga bogs of Western Siberia havebeen studied. More than one hundred (103) species and intraspecific taxons of testate amoebae have beenrevealed in recent surface samples. The relation between ecological characteristics of habitats and the com�position of a Protozoa population has been demonstrated. The ecological preferences of species concerningthe index of wetness, ash level, and acidity have been revealed. Using the correspondence analysis, the eco�logical optimums and the tolerance of species and intraspecific taxons of testate amoebae have been estab�lished.
DOI: 10.1134/S1062359010040059
ZOOLOGY

358
BIOLOGY BULLETIN Vol. 37 No. 4 2010
KUR’INA et al.
Kukushkino bog (samples 1–11). The bog is locatedin the northern part of the Ob–Irtysh interfluve. Thesamples were collected in pine–shrub–sphagnummires (3 samples) and a hummock�ridge complex(8 samples). These habitats are oligotrophic and havelow acidity (pH 3.5–4.9) and ash (2.24–4.38%) levels,and the IW value varies from 4 to 9.
The northeastern shore of Rangetur Lake (samples 12–17). The samples were collected from oligotrophicscheuchzeria–sphagnum mire (one sample), mesotrophicgrass sedge–bogbean–sphagnum mire (one sample),sedge–scheuchzeria–sphagnum–hypnum mire (onesample), and sedge–bogbean–bludderwort mire(three samples). All samples had a low pH value (3.4–4.8); three samples had a low ash value (less than10%), whereas two other samples had a high ash value(up to 37%).
Yuzhnoe bog (samples 18–19). This bog is locatedto the south of Rangetur Lake and represents amesotrophic grassy–sphagnum mire with a runningwater regime. The samples had a low pH value and alow ash value (4%).
Polosatoe bog (samples 20–29). The central hollowmesotrophic–eutrophic part of the aapa complex(8 samples) and the peripheral oligotrophic–mesotrophic part (2 samples) represented the mostvarious habitats, in which pH and ash levels variedfrom 3.8 to 6.5 and from 2.48% to 38.22%, respec�tively. The IW value varied from 2.9 to 9.
Drying aapa complex with dwarf shrubs and a Sph�agnum magellanicum cover (sample 30). The pH valueof the water extract was 5.1.
Zhuravlinoe bog (samples 31–33) represented arunning mesotrophic mire with a sedge–grassy–sph�agnum quagmire. The pH value varied from 4 to 5.1,and the ash level varied from 7.5 to 18.47%.
In general, the examined samples represented aseries of habitats with a wide range of the wetness gra�dient (2.9–9), substrate ash level (2.24–38.33%), andpH values (3.5–6.5).
To reveal the species diversity and to calculate thenumber of testate amoebae, we prepared a water sus�pension of fresh tow samples using the standard proce�dure (Rakhleeva and Korganova, 2005). For eachsample at least 300 shells were determined. We alsodetermined the total density for all shells and for eachspecies separately per 1 g of the oven�dry substance.
The rhizopodium analysis data were treated statis�tically using the Statistica 6.0 and PAST 1.87 software.Samples with an abnormally high ash level and too lowdensity of testate amoebae (nos. 12, 16, 17, 25, 27, 32,33) were excluded from the analysis.
RESULTS
Biodiversity. The analysis of surface sphagnumsamples revealed 103 species, varieties, and forms oftestate amoebae. Each sample contained from 11 to 47
species and forms of Testacea. The dominant complexincluded 13 species and intraspecific forms: Heleoperapetricola, Hyalosphenia papilio, H. elegans, Nebelatincta, Phryganella hemisphaerica, Assulina muscorum,Euglypha cristata, E. rotunda, Sphenoderia lenta,Trinema complanatum, T. enchelys, T. lineate, andAmphitrema flavum. The following species wererevealed in more than 30% of samples, but their abun�dance in communities did not exceed 5%: Bullinulariaindica v. minor, Centropyxis aculeate, Euglypha com�pressa, E. cristata v. decora, E. cuspidate, E. filifera,E. strigosa, Corythion delamarei, C. dubium, andTrinema penardi.
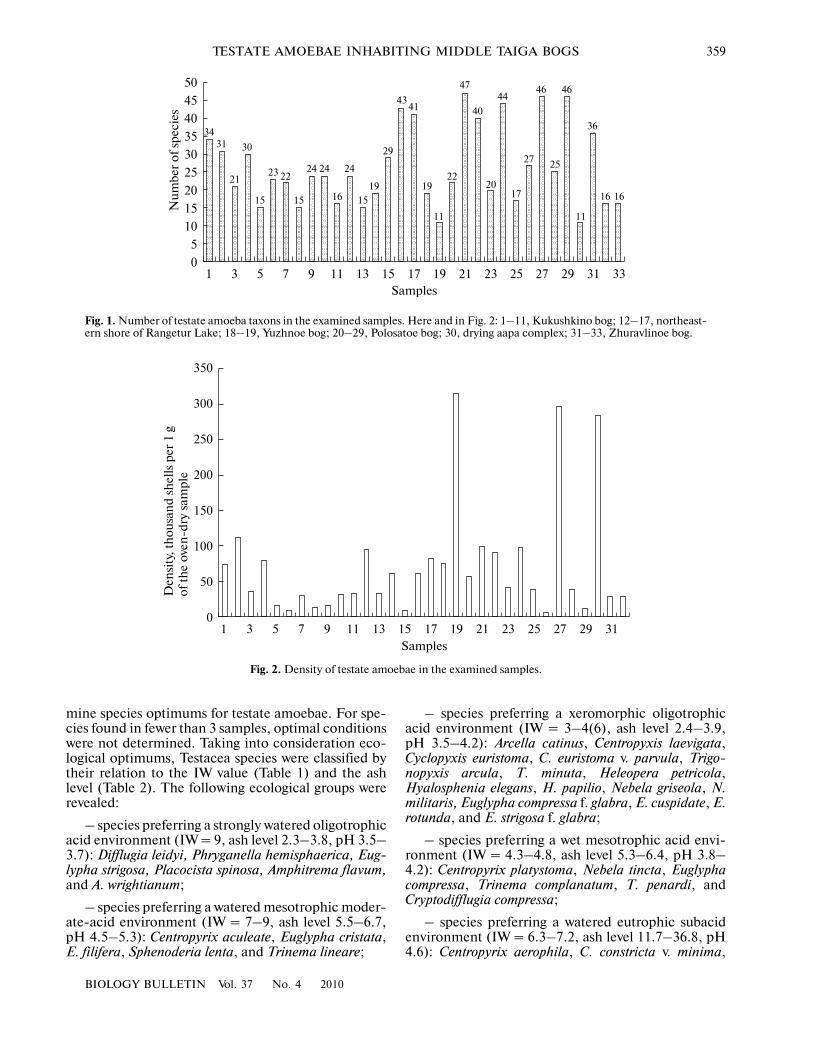
The maximum species diversity was observed forthe oligo� and mesoeutrophic hollows and mires (upto 46–47 species and intraspecific forms). The mini�mum diversity was observed for mesotrophic sphag�num mires with a running water regime and for a mon�odominant sphagnum microstand of a drying aapacomplex (11 species, varieties, and forms; see Fig. 1).
The examined samples did not contain any steno�topic species, except for the rare subspecies C. aculeatev. lata Decloitre.
The density of amoeba populations in the exam�ined samples varied from 7500 to 3 330 400 shells per1 g of the oven�dry substrate (Fig. 2). The maximumdensity was registered in samples with both the lowest(sample nos. 19, 30) and the highest (sample no. 27)species diversity.
A canonical correspondence analysis showed thatthe watering factor, trophic status, and acidity ofbiotopes could explain only 31% of all variations in thestructure of the Testacea community. Only the firstcanonical root was determined as significant (0.976,р = 0.012), and it was closely associated with the IWvalue (the correlation coefficient was 0.98). This resultagrees with the conclusions of other authors that thewatering level of the environment is the main factorinfluencing the community of testate amoebae(Bobrov et al., 2002; Booth, 2001, 2002; Lamentowiczand Mitchell, 2005; Blundell and Barber, 2005; Boothand Zygmunt, 2005).
The dependence of the diversity of Testacea on theacidity and trophic status of their habitats is ratheruncertain. The shell density in the samples did notdepend on the watering level, pH, and trophic status ofthe environment; the correlation coefficients were –0.05, 0.33, and 0.49, respectively (р > 0.05). RegardingpH and ash levels, we observed two groups of Testaceacommunities, one group from high�ash habitats, andanother group from low�ash habitats (Fig. 3). The firstgroup was represented by the samples with unusuallyhigh ash content. In both cases we did not reveal anycorrelation between the composition of Testacea com�munities and changes in the ash content and pH value.
Species optimum and tolerance. In the case of opti�mal environmental conditions, species reach theirmaximum abundance; this fact allowed us to deter�
BIOLOGY BULLETIN Vol. 37 No. 4 2010
TESTATE AMOEBAE INHABITING MIDDLE TAIGA BOGS 359
mine species optimums for testate amoebae. For spe�cies found in fewer than 3 samples, optimal conditionswere not determined. Taking into consideration eco�logical optimums, Testacea species were classified bytheir relation to the IW value (Table 1) and the ashlevel (Table 2). The following ecological groups wererevealed:
– species preferring a strongly watered oligotrophicacid environment (IW = 9, ash level 2.3–3.8, pH 3.5–3.7): Difflugia leidyi, Phryganella hemisphaerica, Eug�lypha strigosa, Placocista spinosa, Amphitrema flavum,and A. wrightianum;
– species preferring a watered mesotrophic moder�ate�acid environment (IW = 7–9, ash level 5.5–6.7,pH 4.5–5.3): Centropyrix aculeate, Euglypha cristata,E. filifera, Sphenoderia lenta, and Trinema lineare;
– species preferring a xeromorphic oligotrophicacid environment (IW = 3–4(6), ash level 2.4–3.9,pH 3.5–4.2): Arcella catinus, Centropyxis laevigata,Cyclopyxis euristoma, C. euristoma v. parvula, Trigo�nopyxis arcula, T. minuta, Heleopera petricola,Hyalosphenia elegans, H. papilio, Nebela griseola, N.militaris, Euglypha compressa f. glabra, E. cuspidate, E.rotunda, and E. strigosa f. glabra;
– species preferring a wet mesotrophic acid envi�ronment (IW = 4.3–4.8, ash level 5.3–6.4, pH 3.8–4.2): Centropyrix platystoma, Nebela tincta, Euglyphacompressa, Trinema complanatum, T. penardi, andCryptodifflugia compressa;
– species preferring a watered eutrophic subacidenvironment (IW = 6.3–7.2, ash level 11.7–36.8, pH4.6): Centropyrix aerophila, C. constricta v. minima,
Num
ber
of s
peci
es
50
45
40
35
30
25
20
15
10
5
01 3 5 7 9 11 13 15 17 19 21 23 25 27 29 31 33
Samples
3431
21
30
15
23 22
15
24
1619
29
4341
11
22
47
40
20
44
17
27
46
25
11
36
16
24 24
15
19
46
16
Fig. 1. Number of testate amoeba taxons in the examined samples. Here and in Fig. 2: 1–11, Kukushkino bog; 12–17, northeast�ern shore of Rangetur Lake; 18–19, Yuzhnoe bog; 20–29, Polosatoe bog; 30, drying aapa complex; 31–33, Zhuravlinoe bog.
Den
sity
, th
ousa
nd
shel
ls p
er 1
g
350
300
250
200
150
100
50
01 3 5 7 9 11 13 15 17 19 21 23 25 27 29 31
Samples
of t
he
oven
�dry
sam
ple
Fig. 2. Density of testate amoebae in the examined samples.

360
BIOLOGY BULLETIN Vol. 37 No. 4 2010
KUR’INA et al.
Difflugia 0bacillifera, D. globulosa, D. pulex,Lesquereusia spiralis, Tracheleuglypha dentate, andTrinema enchelys.
The analysis of the distribution of testate amoebaedepending on different habitat types allowed us toselect five groups of habitats: oligotrophic mossy hum�mocks and hillocks, oligotrophic mossy covers andinterhummock areas, oligotrophic mossy mires,mesoeutrophic mossy mires, and mesoeutrophicgrassy mires.
In the group of oligotrophic mossy hummocks, thedominant complex (the abundance exceeded 10%)included Assulina muscorum, Centropyxis laevigata,Cyclopyxis eurystoma, C. eurystoma v. parvula,
Hyalosphenia papilio, Euglypha rotunda, Nebela mili�taris, Trigonopyxis arcula, T. minuta, and Trinemacomplanatum. All these species, except for the last two,reached their maximum relative abundance in thisgroup.
The dominant complex in the group of olig�otrophic mossy covers included Hyalosphenia elegans,H. papilio, Amphitrema flavum, Assulina muscorum,Nebela tincta, and Phryganella hemisphaerica. Thefirst two species had the maximum abundance in thisgroup of biotopes (the total relative abundance was35–56%). These species were also continually presentin oligotrophic mossy mires, but their total abundancein this case did not exceed 16%.
The dominant complex of oligotrophic mossymires included Amphytrema flavum, Difflugia leidyi,Nebela griseola, Phryganella hemisphaerica, Heleoperapetricola, and Hyalosphenia elegans.
It seems that the watering level preferences ofPhryganella hemisphaerica and Placocista spinosainclude both oligotrophic mossy covers and mires,since these species are abundant in both biotopegroups, whereas in other groups their density was verylow.
The samples from mesoeutrophic mossy miresalways included Euglypha cristata and Hyalospheniapapilio and also many various species from the generaArcella and Centropyxis. The dominants of this groupwere Difflugia globulosa, Euglypha cristata, Sphenode�ria lenta, Trinema complanatum, T. enchelys, T. lin�eare, Euglypha rotunda, Lesquereusia spiralis, Nebelatincta, and Tracheleuglypha dentate.
pH
6
5
4
3
2
1
0 10 20 30 40
Ash level
AB
Fig. 3. Ordination of testate amoeba communities regard�ing the acidity and ash level of their habitats. A, low�ashhabitat group; B, high�ash habitat group.
Table 1. Ordination of testate amoebae regarding the index of wetness (IW)
Species IW optimum
Amphitrema wrightianum, Arcella arenaria, A. discoides v. scutelliformis, A. hemisphaerica, A. intermedia, A. intermedia v. laevis, A. jurassica, Centropyxis aculeata v. minima, C. cassis f. spinifera, C. constricta, Difflu�gia angulostoma, D. bacillariarum, D. bacillifera, D. leidyi, D. pulex, Difflugiella oviformis f. fusca, Euglypha anodonta, E. ciliata, Phryganella hemisphaerica, Lesgueruesia spiralis, Netzelia oviformis
8.52–7.1
Amphitrema flavum, Arcella catinus v. sphaerocysta, A. discoides, A. rotundata v. stenostoma, A. vulgaris, Cen�tropyxis aculeata, C. aerophila, C. aerophila v. sphagnicola, C. constricta v. minima, C. ecornis, Difflugia glob�ulosa, D. pristis, Difflugiella oviformis, Heleopera petricola v. ametystea, Euglypha cristata, E. cristata v. deco�ra, E. filifera, E. rotunda, E. strigosa, Lesquereusia epistomium, Nebela carinata, N. dentistoma, N. griseola, Placocista spinosa, Pseudodifflugia gracilis v. terricola, Sphenoderia lenta, Tracheleuglypha dentata, Trinema complanatum v. globulosa, T. enchelys, T. complanatum v. inequalis, Quadrulella symmetrica
7–6.15
Arcella rotundata v. aplanata, Bullinularia indica v. minor, Centropyxis sylvatica, Corythion dubium, Cyclopy�xis eurystoma v. parvula, Difflugia. globulus, Euglypha compressa, E. compressa f. glabra, E. cuspidata, E. stri�gosa f. glabra, Heleopera sylvatica, Hyalosphenia elegans, H. papilio, H. papilio v. stenostoma, Nebela bohe�mica, N. collaris, N. galeata, N. tincta, Phryganella acropodia, Trinema complanatum, T. lineare, T. lineare v. truncatum, T. penardi
6–5.12
Arcella artocrea, A. artocrea v. aplanata, A. catinus, A. vulgaris v. undulata, Assulina muscorum, A. muscorum v. stenostoma, Bullinularia gracilis, Centropyxis. aculeata v. lata, C. laevigata, C. minuta, C. platystoma, Corythion delamarei, C. dubium v. orbicularis, Cryptodifflugia compressa, Cyclopyxis eurystoma, C. kahli, Cy�phoderia ampulla, Euglypha anodonta v. magna, E. tuberculata, E. tuberculata v. minor, Heleopera petricola, H. rosea, Nebela collaris f. grandis, N. marginata, N. militaris, N. parvula, Trigonopyxis arcula, T. minuta
5–3.88

BIOLOGY BULLETIN Vol. 37 No. 4 2010
TESTATE AMOEBAE INHABITING MIDDLE TAIGA BOGS 361
Such species as Cyphoderia ampulla, Lesquereusiaspiralis, L. epistomium, Nebela collaris, N. collaris f.grandis, N. galeata, N. marginata, Netzelia oviformis,and Quadrulella symmetrica were observed only in thegroup of mesoeutrophic mossy mires.
In the case of mesoeutrophic grassy mires, thedominant complex included Tracheleuglypha dentate,Difflugia oviformis f. fusca, Trinema lineare, T. com�planatum, and T. enchelys; the first three speciesreached their maximum abundance in this group. Thepercentage of Trinema lineare and Difflugiella ovifor�mis f. fusca reached a very high level in several samples(>70% and 99%). The dominant complexes of otherbiotope groups did not include these species, exceptfor T. lineare, which was also abundant (up to 43%) inthe group of mesoeutrophic mossy mires.
Species such as Amphitrema flavum, Assulina mus�corum, Cyclopyxis euristoma v. parvula, Euglypharotunda, Heleopera petricola, Nebela tincta, Trinemacomplanatum, and T. lineare were common for allgroups of biotopes.
DISCUSSION
From the biogeographical point of view, WesternSiberia is a very interesting territory, because it repre�sents one of the largest accumulative plains on theEarth. There were many glaciations on this territory,including the Samarovskoe (230000–170000 years ago)and Zyryanskoe (80000–10000 years ago). According tothe modern concepts, these glaciations formed a con�tinuous ice cover only in the north of Western Siberiawhich made it possible to find any endemic or deletedspecies on this territory. This hypothesis was con�firmed by Rakhleeva’s study (Rakhleeva, 2002), in
which two species rare for Eurasia, Centropyxis patulaStepanek and Quadrulella quadrigera Deflandre, werefound in mesotrophic–eutrophic habitats of valleybogs. However, the search for rare species in uplandbogs seems to be fairly ineffective, since the endemicspecies of acidophilic sphagnobiont testate amoebaeare very rare; the only exception is a Circum�Austra�lian group of Testacea in the southern hemisphere(Smith et al., 2008).
The obtained results were compared with the dataof other authors who studied the bogs of the Europeanpart of Russia (Bobrov et al., 2002; Mazey et al., 2007)and northwestern Poland (Mitchell and Lamentow�icz, 2005). Though the exact comparison of the spe�cies�specific optimums of the environment wateringlevel was impossible because of the use of differentassessment methods, we obtained a similar range ofseries of species to those described by Payne et al.(Payne et al., 2006). Thus, our data confirmed thehypothesis of the similarity of environmental prefer�ences of testate amoebae inhabiting the oligotrophicbogs of the northern hemisphere (Bobrov et al., 2002).
The watering level of a habitat is the main factorinfluencing the communities of testate amoebae.Regarding this factor, three different groups wererevealed among the species with the lowest tolerance:hydromorphic (Arcella discoides v. scutelliformis,A. rotundata v. stenostoma, Difflugia angulostoma,D. bacillariarum, D. pristis, Lesquereusia epistomum,Netzelia oviformis, Tracheleuglypha dentate, Dif�flugiella oviformis, and D. oviformis f. fusca); hygro�morphic (Difflugia globules, Nebela bohemica, N. den�tistoma, N. tincta, Trinema enchelys, and Pseudodifflu�gia gracilis v. terricola); and xeromorphic (Trigonopyxisminuta and Nebela militaris).
Table 2. Ordination of testate amoebae regarding the ash level
Species Ash level optimum, %
Arcella rotundata v. yitenostoma, Centropyxis ecornis, Lesquereusia epistomium, Trinema enchelys 9.39–7.03
Centropyxis aculeata, C. constricta v. minima, C. sylvatica, Difflugia bacillariarum, D. bacillifera, D. globu�losa, D. pulex, Euglypha cristata, E. cristata v. decora, E. filifera, Heleopera petricola v. ametystea, Nebela collaris, N. dentistoma, N. galeata, Sphenoderia lenta, Tracheleuglypha dentata, Trinema penardi, Quadru�lella symmetrica
6.96–6.03
Arcella discoides, Centropyxis aerophila, C. platystoma, Cryptodifflugia compressa, Difflugia angulostoma, Difflugiella oviformis, Heleopera sylvatica, Euglypha anodonta, E. rotunda, E. strigosa, E. tuberculata, Ne�bela bohemica, N. marginata, Pseudodifflugia gracilis v. terricola, Phryganella acropodia, Trinema com�planatum, T. lineare
5.92–5.04
Amphitrema flavum, A. wrightianum, Arcella rotundata v. aplanata, A. jurassica, Assulina muscorum, Corythion dubium, C. dubium v. orbicularis, Hyalosphenia papilio v. stenostoma, Nebela tincta, Euglypha compressa, E. compressa f. glabra, E. cuspidata, E. strigosa f. glabra, E. tuberculata v. minor, Phryganella hemisphaerica, Placocista spinosa
5–4.03
Arcella catinus, A. catinus v. sphaerocysta, A. discoides v. scutelliformis, Bullinularia indica v. minor, Centro�pyxis laevigata, Corythion delamarei, Cyclopyxis eurystoma, C. eurystoma v. parvula, Difflugia leidy, Dif�flugiella oviformis f. fusca, Euglypha ciliata f. glabra, Heleopera petricola, Hyalosphenia elegans, H. papilio, Nebela carinata, N. griseola, N. militaris, Trigonopyxis arcula, T. minuta
4–2.65

362
BIOLOGY BULLETIN Vol. 37 No. 4 2010
KUR’INA et al.
Regarding the trophic status of the environment,two groups of stenotopic indicator species wererevealed: a group preferring oligotrophic conditions(Trigonopyxis minuta, Difflugia leidyi, and Arcella cati�nus), and a group preferring mesoeutrophic condi�tions (Centropyxis platystoma, Difflugia angulostoma,D. globules, Netzelia oviformis, and Difflugiella ovifor�mis).
Regarding the exceptional arrangement in certainbiotopes, we revealed the following species of testateamoebae able to serve as indicator species: (1) Difflugiabacillariarum (mire biotopes) and (2) Difflugia bacil�lifera, Amphitrema wrightianum, Arcella discoides v.scutelliformis, A. jurassica, and Nebela carinata (mossymire biotopes).
Biotope groups segregated according to theirwatering level, trophic status, and substrate type corre�spond to the common elements of boggy microlan�dscapes. Therefore, the revealed structural features ofthe testate amoeba communities inhabiting these ele�ments can be used to reconstruct environmental con�ditions during palaeoecological studies.
REFERENCES
Alekseev, D.A. and Bobrov, A.A., Testaceans of the Euro�pean North of Russia, Tsitologiia, 1992, vol. 34, no. 4, p. 17.
Blundell, A. and Barber, K.A., 2800�Year PalaeoclimaticRecord from Tore Hill Moss, Strathspey, Scotland: theNeed for a Multi�Proxy Approach to Peat�Based ClimateReconstructions, Quaternary Sci. Rev., 2005, no. 24,pp. 1261–1277.
Bobrov, A. and Minaeva, T., Indication Capacity of Testa�ceans Protozoa testacea and Sphagna Species Associations,Proc. 11th Int. Peat Congr., Quebec, 2000, vol. I, pp. 6–17.
Bobrov, A.A., Charman, D., and Warner, B., Ecology ofTestate Amoebae from Oligotrophic Peatlands: SpecificFeatures of Polytypic and Polymorphic Species, Izv. Akad.Nauk, Ser. Biol., 2002, no. 6, pp. 738–751.
Bobrov, A.A., Historical Dynamics of Lake–Bog Ecosys�tems and Succession of Testaceans (Testacea), Zool. Zh.,2003, vol. 82, no. 2, pp. 215–223.
Bobrov, A.A., Yazvenko, S.B., and Warner, B.G., Taxo�nomic and Ecological Implications of Shell Morphology ofThree Testaceans (Protozoa: Rhizopoda) in Russia andCanada, Arch. Protistenkd., 1995, vol. 145, pp. 119–126.
Booth, R.K. and Zygmunt, J.R., Biogeography and Com�parative Ecology of Testate Amoebae Inhabiting Sphag�num�Dominated Peatlands in the Great Lakes and RockyMountain Regions of North America, Diversity, Distribu�tions, 2005, no. 11, pp. 577–590.
Booth, R.K., Testate Amoebae as Paleoindicators of Surface�Moisture Changes On Michigan Peatlands: Modern Ecologyand Hydrological Calibration, J. Paleolimnol., 2002,vol. 28, pp. 329–348.
Booth, R.K., Ecology of Testate Amoebae (Protozoa) inTwo Lake Superior Coastal Wetlands: Implications for Pale�
oecology and Environmental Monitoring, Wetlands, 2001,vol. 21, no. 4, pp. 564–576.
Charman, D.J. and Warner, B.G., Relationship betweenTestate Amoebae (Protozoa: Rhizopoda) and Microenviron�mental Parameters on a Forested Peatland in NorthwesternOntario, Can. J. Zool., 1992, vol. 70, pp. 2474–2482.
Charman, D.J. and Warner, B.G., The Ecology of TestateAmoebae Protozoa rhizopoda in Oceanic Peatlands in New�foundland, Canada: Modeling Hydrological Relationshipsfor Paleoenvironmental Reconstruction, Ecoscience, 1997,no. 4, pp. 555–562.
Charman, D.J., Modelling Hydrological Relationships ofTestate Amoebae (Protozoa: Rhizopoda) on New ZealandPeatlands, J. Royal Soc. (N. Zeal.), 1997, vol. 27, no. 4,pp. 465–483.
Elina, G.A. and Yurkovskaya, T.K., Methods of Determina�tion of Paleohydrological Regimen as a Basis of ObjectiveCauses of Succession of Bog Vegetation, Bot. Zh., 1992,vol. 77, no. 7, pp. 120–124.
GOST 11306�83. Torf i produkty ego pererabotki, Metodyopredeleniya zol’nosti (GOST 11306�83. Peat and Its Pro�cessing Products. Methods of Ash Content Determina�tion), Moscow: Izd. Standartov, 1985.
Lamentowicz, M. and Mitchell, E., The Ecology of TestateAmoebae (Protists) in Sphagnum in North�Western Polandin Relation to Peatland Ecology, Microbial. Ecol., 2005,vol. 50, pp. 48–63.
Mazei, Yu.A., Tsyganov, A.N., and Bubnova, O.A., SpeciesComposition, Distribution, and Structure of the Commu�nity of Testate Amoebae of Peat Bog in the Middle VolgaRegion, Zool. Zh., 2007, vol. 86, no. 10, pp. 1155–1167.
Mitchell, E.A.D., Buttler, A.J., and Warner, B.G., Ecologyof Testate Amoebae (Protozoa: Rhizopoda) in SphagnumPeatlands in the Jura Mountains, Switzerland and France,Ecoscience, 1999, no. 6, pp. 565–576.
Payne, R.J. and Kishaba, K., Blackford 1.J, Mitchell E.Ecology of Testate Amoebae (Protista) in South�CentralAlaska Peatlands: Building Transfer�Function Models forPalaeoenvironmental Studies, Holocene, 2006, vol. 16,pp. 403–414.
Rakhleeva, A.A. and Korganova, G.A., On the Problem ofAssessment of Abundance and Species Diversity of TestateAmoebae (Rhizopoda, Testacea) in Taiga Soils, Zool. Zh.,2005, vol. 84, no. 12, pp. 1427–1436.
Rakhleeva, A.A., Testaceans (Testacea, Protozoa) of TaigaSoils in Western Siberia (Surgut Polesye), Izv. Akad. Nauk,Ser. Biol., 2002, no. 6, pp. 752–762.
Tolonen, K., Warner, B.G., and Vasander, H., Ecology ofTestaceans (Protozoa: Rhizopoda) in Mires in SouthernFinland: I. Autecology, Arch. Protistenkd., 1992, vol. 142,pp. 119–138.
Warner, B.G. and Charman, D.J., Holocene Changes on aPeatland in Northwestern Ontario Interpreted from TestateAmoebae (Protozoa) Analysis, Boreas, 1994, vol. T. 23,pp. 271–279.
Woodland, W.A., Charman, D.J., and Sims, P.C., Quantita�tive Estimates of Water Tables and Soil Moisture inHolocene Peatlands from Testate Amoebae, Holocene,1998, vol. 8, pp. 261–273.













![Intra-Lake Arcellinida (Testate Lobose Amoebae) Response ......curs as rainfall during the summer [30]. Silver Lake is close to four lakes located along the Highway 7 that havebeenpreviously](https://img.dokumen.tips/doc/110x75/61296389b4121c7a8109f7c0/intra-lake-arcellinida-testate-lobose-amoebae-response-curs-as-rainfall.jpg)















