Embed Size (px)
Citation preview

UNIVERSIDAD AUTÓNOMADE BAJA CALIFORNIA SUR
`rea Interdisciplinaria de Ciencias del Mar
Departamento de Biología Marina
Evaluación de TØcnicas ElectroforØticas
Para la Caracterización y Deterrnmación
Taxonómica de Levaduras Marinas de la
Costa Occidental de Baja California Sur
TESIS
QUE COMO REQUISITO PARA OBTENER
EL T˝TULO DE
BiÓLOGO MARINO
PRESENTA
JOSÉ ARTURO S`NCHEZ PAZ
La Paz B e s JW1io de 1998

BlBI10TE
lì
oJI í
L
40432

A mispadres
JosØ Arturo y María Eugenia
por su apoyo en los momentos mÆs
ilnp011antes demi vida
A mi esposa
Adriana
por su amor ypor sermi mejor motivo
de superación

Agradecimientos
Este trabajo es el producto del apoyo de muchas personas De modo que llegó la
hora de dar las gracias a todos aquellos con los que he adquirido deudas intelectuales y
emocionales amigos compaæeros y profesores Inevitablemente solo puedo
reconocer una pequeæa parte de todos aquellos que han tenido alguna influencia en mi
desarrollo No es necesario aæadir que ellos son los responsables de todo lo bueno que
pueda encontrarse en este trabajo
En pnmer lugar deseo agradecer al Dr JosØ Luis Ochoa por su apoyo y
paciencia inagotable paciencia que ha demostrado hacia mi persona
Igual de importante ha sido la ayuda que recibí de mis AMIGOS las mayœsculas
nunca les demostrarÆn lo que los aprecio Norma HernÆndez Saavedra y Arturo Sierra
los defectos contenidos en este trabajo habrían abundado sin su intervención
Al comitØ revisor del Departamento de Biología Marina Hermilo Santoyo David
Siqueiros Aurora Rebolledo y Víctor Carrasca
A mi compaæero de laboratorio Martín Rarnírez ojalÆ y sigamos compartiendo
mucho tiempo mÆs el laboratorio de Levaduras Marinas del CIBNOR
A Eduardo Ruiz PyJon Dariel Tovar y a Ariel Cruz por sus juegos chistes y
travesuras cada vez que existió el peligro inminente de irrupción de la cordura
A todos mis maestros de la Universidad Autónoma de Baja California Sur
Al Centro de Investigaciones Biológicas del Noroeste donde se desarrollaron los
experimentos y donde me he desarrollado profesionalmente

A mis hermanos Claudia y Pedro por su amor y constante interØs
A Daniel HernÆndez Siendo honestos tœ me metiste en este lío de las levaduras
A todos aquellos que no menciono les pido una disculpa pero sirva como excusa
la falta de espacio Si hiciera mención de todos los involucrados podría escribir todas las
tesis del mundo y aœn así permanecería en deuda con alguien

Contenido
PÆgina
lndice General 1
Indice de Tablas IV
lndice de GrÆficas V
Indice de Esquemas y Figuras V
Glosario vii
Abreviaturas de gØneros x
Resumen xi
1 Introducción 1
l Generalidades 1
2 Antecedentes 5
MØtodos de Identificación y Clasificación 6
TaxonoIlÚa numØrica 8
n Objetivo general 10
III Objetivos específicos 10
IV Materiales y mØtodos 10
l Organismos a estudiar y organismos de referencia 10
2 Reactivación de las cepas 12
3 Caracterización de cepas mediante morfología celtøar y colonial 12
Morfología celular 12
Morfología colonial 12
4 Pruebas de asimilación de fuentes carbono y nitrógeno 14
4 1 Preparación del inóculo 14
4 2 Asimilación de carbohidratos 14
Preparación del medio de cultivo 14

4 3 Asimilación de fuentes de nitrógeno 15
Preparación del medio de cultivo 15
5 ElectroforØsis de proteínas solubles totales 16
5 1 Producción de biomasa 16
5 2 Obtención de proteínas solubles totales 16
5 3 ElectroforØsis en gel de poliacrilamida PAGE 17
5 4 Densitometría 18
6 AnÆlisis de los datos 18
7 ElectroforØsis de ADN genómico 19
7 1 Cepas utilizadas 19
7 2 Digestión del ADN con enzimas de restricción y electrofo
rØsis en geles de agarosa 20
7 3 PCR y electroforØsis en gel de agarosa 22
7 4 AnÆlisis de los datos 23
V Resultados 24
l AnÆlisis numØrico de las características morfológicas y fisiológicas 24
2 Contenido de proteínas totales 32
3 Características generales de los patrones de proteínas totales 34
4 AnÆlisis numØrico de los patrones electroforØticos de proteínas
solubles totales 34
5 AnÆlisis de las cepas A18 A18c A23 A43 002 006 A75 B12 B13
B17 C12 C22 y C49 44
5 1 Con base en las pruebas morfológicas y fisiológicas 44
5 2 Con base en los patrones electroforØticos de proteínas 45
6 Concentración de Æcidos nucleicos 47
7 Características generales de patrones de restricción 48
11

8 AnÆlisis numØrico de los patrones electroforØticos de ADN 49
Bam HI 49
EcoRI 50
Hinf1 51
Pst 1 52
9 Características generales de las amplificaciones al azar de
ADN polimórfico RAPDs 54
10 AnÆlisis de los patrones electroforØticos generados por RAPDs
Oligonucleótido 1 secuencia TCACGGTGCA 54
Oligonucleótido 2 secuencia GTCGCCGAC 56
Oligonucleótido 3 secuencia GTAGACGAGC 57
Oligonucleótido 4 secuencia AAACGTCGGG 58
Oligonucleótido 5 secuencia ACTGACTGCC 59
VI Discusión 61
VII Conclusiones 69
VIII Bibliografía 72
IX ApØndices 82
ApØndice l Medios de cultivo 82
ApØndice 2 Determinación de proteínas por el mØtodo de
Bradford 1976 83
ApØndice 3 Reactivos y electroforØsis de proteínas en gel
desnaturalizante SDS PAGE basado en el mØtodo deLaenunli 1970 84
ApØndice 4 TØcnicas para la extracción de Æcidos nucleicos de
levaduras 87
ApØndice 5 Buffer TBE 10x 89
III

lndice de Tablas
Tabla l Clasificación taxonómica de levaduras 3
Tabla 2 Cepas de la Colección de Levaduras Marinas II
Tabla 3 Fuentes de carbono utilizadas en las pruebas de asimilación 15
Tabla 4 EstÆndares de peso molecular en electroforØsis de proteínas 17
Tabla 5 Secuencias de reconocimiento de las enzimas de restricción 21
Tabla 6 Marcadores moleculares utilizados en electroforØsis de ADN 22
Tabla 7 Secuencia de los primers utilizados en el ensayo de RAPD 22
Tabla 8 Programa de corrimiento del PCR 23
Tabla 9 Levaduras que integran el grupo 1 Basado en sus características
morfológicas y fisiológicas 26
Tabla lO Levaduras que integran el grupo n Basado en sus características
morfológicas y fisiológicas 29
Tabla 11 Levaduras que integran el grupo III Basado en sus características
morfológicas y fisiológicas 31
Tabla 12 Biomasa obtenida y contenido de proteínas de los extractos de
cada cepa 33
Tabla 13 Levaduras que integran el grupo 1 Basado en el patrón
electroforØtico de proteínas solubles totales 36
Tabla 14 Levaduras que integran el grupo n Basado en el patrón
electroforØtico de proteínas solubles totales 39
Tabla 15 Levaduras que integran el grupo III Basado en el patrón
electroforØtico deproteínas solubles totales 42
Tabla 16 Levaduras que integran el grupo N Basado en el patrón
electroforØtico de proteínas solubles totales 42
Tabla 17 Resultados de la cuantificación de ADN 47
IV

lnclice de GrÆficas
GrÆfica l Grupo 1 Con base enpruebas convencionales 27
GrÆfica 2 Grupo n Con base en pruebas convencionales 30
GrÆfica 3 Grupo III Con base en pruebas convencionales 32
GrÆfica 4 Grupo 1 Con base en patrones electroforØticos de proteínas 37
GrÆfica 5 Grupo n Con base en patrones electroforØticos de proteínas 40
GrÆfica 6 Grupo IV Con base en patrones electroforØticos de proteínas 43
lnclice de Esquemas y Figuras
Esquema l Diagrama de flujo de la metodología empleada 13
Figura l Dendograma obtenido mediante pruebas convencionales 25
Figura 2 ElectroforØsis de proteínas de 7 cepas de Levaduras Marinas 34
Figura 3 Dendograma obtenido mediante patrones electroforØticos de
proteínas solubles totales 35
Figura 4 Dendograma de las cepas de Rhodotorula y Debaryomyces
vanrijii vanrijii con base en las pruebas convencionales 45
Figura 5 Dendograma de las cepas de Rhodotorula y Debaryomyces
vanrijii vanrijii con base en los patrones electroforØticos de proteínas 46
Figura 6 ElectroforØsis del ADN genórnico 48
Figura 7 Dendograma de las cepas de Rhodotorula y Debaryomyces
vanrijii vanrijii a partir de la digestión de ADN por la enzima Bam H1 49
Figura 8 Dendograma de las cepas de Rhodotorula y Debaryomyces
vanrijii vanrijii a partir de la digestión de ADN por la enzima Eco RI 51
Figura 9 Dendograma de las cepas de Rhodotom1a y Debaryomyces
vanrijii vanrijii apartir de la digestión de ADN por la enzima Hinfl 52
Figura 10 Dendograma de las cepas de Rhodotorula y Debaryomyces
vanrijii vanrijii a partir de la digestión de ADN por la enzima Pst1 53
v

Figura 11 Patrón electroforØtico de las cepas de RhodotoruJa y
Debaryomyces vamijii vamijii obtenido por PCR utilizando el oligonuc1eótido
l 55
Figura 12 Patrón electroforØtico de las cepas de RJlOdotoruJa y
Debaryomyces vanrijii vamijii obtenido por PCR utilizando el oligonuc1eótido
2 56
Figura 13 Patrón electroforØtico de las cepas de RhodotoruJa y
Debaryomyces vanrijii vanrijii obtenido por PCR utilizando el oligonuc1eótido
3 57
Figura 14 Patrón electroforØtico de las cepas de Rhodotorula y
Debaryomyces vamijii vamijii obtenido por PCR utilizando el oligonuc1eótido
4 58
Figura 15 Patrón electroforØtico de las cepas de RhodotoruJa y
Debaryomyces vamijii vanrijii obtenido por PCR utilizando el oligonuc1eótido
5 59
VI

Glosario
Aloenzima Una o mÆs variantes de una enzima usualmente variantes de movilidad
identificadas por electroforØsis codificados por diferentes alelos en el mismo locus del
gen
Biodiversidad TØrmino ampliamente utilizado contracción de diversidad biológica
que puede referirse a la diversidad nœmero de taxa y o sus variaciones genØticas
Cariotipo El complemento cromosomal de una cØlula deun organismo
Dendograma Cualquier diagrama ramificado con taxa terminales cuya estructura
representa similitudes fenotípicas o genotípicas entre los taxa mas que una relación
evolutiva
ElectroforØsis Procedimientos para separar molØculas en un medio de apoyo con
base en su carga elØctrica en combinación con otros factores vØase SDS PAGE
Enzima de restricción Una enzima que corta ADN de doble hebra reconociendo
secuencias específicas generalmente han sido aisladas de bacterias
Esferoplasto CØlulas de microorganismos cuya pared celular ha sido digerida por
medios enzimÆticos
Fenón Se refiere a los grupos de unidades taxonómicas operacionales OTUs
obtenidas a partir de estudios fenØticos
Filogenia Lahistoria evolutiva de un taxa
VIl

Isoenzima Cualquiera de las formas de una enZIma codificadas por un gen en
diferente locus Se pueden originar por ejemplo a partir de una duplicación de genes
PAGE ElectroforØsis en Gel de Poliacrilamida Cualquier separación electroforØtica
normalmente de proteínas realizada en un gel de poliacrilamida como medio de
apoyo
PCR polymerase chain reaction Procedimiento para amplificar selectivamente
produciendo un gran nœmero de copias idØnticas pequeæas muestras de ADN sin la
necesidad de clonar Ampliamente utilizada cuando el material es limitado por ejemplo
con microorganismos o material fósil
PlÆsmido Una molØcula pequeæa de ADN circular separada de los cromosornas pero
que se replica con el resto del material genØtico Muy comœn en procariotas y
ampliamente utilizada en manipulaciones genØticas
Polimorfísmo 1 La ocurrenCIa en una especIe de mÆs de un estado discreto
fenotípico 2 la ocurrencia de dos o mÆs estados de un carÆcter entre los miembros de
un taxón
Primer Una pequeæa sección de ADN de una sola hebra o de ARN que al alinearse a
una hebra complementaria mÆs larga se extiende por la ADN polimerasa a travØs de la
longitud de la hebra de ADN mÆs larga Cebador
RAPD random amplified polymorphic DNA Amplificación al azar de secuencias de
ADN usando oligonucleótidos con secuencias elegidas al azar
Vlll

RFLP restriction fragment length polymorphisms La ocurrencia en un taxón de mÆs
de una forma tipo definida por la presencia o ausencia de un sitio particular de
reconocimiento de una enzima de restricción
SDS Dodecil Sulfato de Sodio Detergente que se liga a las proteínas permitiendo su
separación electroforØtica con base en su peso molecular
Secuencia palindrómica Secuencia de nucleótidos complementaria en sí misma es
decir se lee igual del extremo 3 5 al extremo 5 3 Palindromos perfectos ocurren con
frecuencia en sitios de reconocimiento para enzimas restrictivas
Serovar o Serotipo 1 En taxonomía bacterial un rango infraespecífico sm
reconocimiento oficial que denota a una cepa distinguible en base a sus propiedades
antigØnicas 2 en taxonomía general un grupo de OTUs identificados por una
característica inmunológica particular
SistemÆtica El estudio científico de las clases y la diversidad de los organismos y de
sus interrelaciones incluyendo aspectos de filogenia evolución biogeografia y
genØtica
Sitio de restricción Secuencia corta de ADN tipicamente palindrómica reconocida
por enzimas de restricción de tipo n las cuales cortan el ADN adyacente a este sitio
Taxón Cualquier unidad taxonómica definible descrita o no por ejemplo subespecies
especies tribu gØnero familia etc plural taxa o taxones
Taxonomía El estudio teórico de la clasificación incluyendo bases principios
procedimientos y reglas
IX

Abreviaturas de gØneros
Reconocidos por la CBS Yeast Division of the Centraalbureau voor
Schirnrnelcultures laboratory of Microbiology Technical University Julianalaan 67 A
Delit Netherlands Kreger van Rij I 1984
Candida C
Debaryomyces Deb
Hansenula H
K1uyveromyces K
Leucosporidium Leu
Metschnikowia M
Pichia P
Rhodotom1a Rh
Saccharomyces Sacch
Sporobolomyces Sp
Zygosaccharomyces Z
x

Abstract
Classification and identification of yeasts and yeast like organisms relies heavily
on morphological and physiological characteristics however the problem of using such
criteria frequently suffers from a lack of sensitivity and specificity Whole cell protein
electrophoresis restriction endonuclease analysis of genomic DNA RFLP and random
amplified polymorphic DNA RAPD were used to distinguish different strains of marine
yeasts collected on the West Coast of Baja California Sur Taken together these results
indicate that tbis techniques will be useful tools for routine identification of marine yeasts
Noteworthy of the techniques assayed RAPD proved to be the most reliable for
taxonomic purposes
Resumen
La clasificación e identificación de las levaduras se ha basado principalmente en
características morfológicas y fisiológicas sin embargo el uso de estos criterios ha
generado problemas como falta de sensibilidad y especificidad En el presente estudio
se realizaron anÆlisis electroforØticos de proteínas solubles totales digestión de ADN
genómico mediante enzimas de restricción RFLP Y amplificaciones al azar de ADN
polimórfico RAPD para distinguir entre diferentes cepas de levaduras marinas
colectadas en la Costa Occidental de Baja California Sur En conjunto estos resultados
indican que estas tØcnicas son œtiles para la identificación rutinaria de levaduras
marinas Sin embargo de las tØcnicas ensayadas la del RAPD probó ser la mÆs œtil para
fines taxonómicos
Xl

Evaluación de TØcnicas ElectroforØticas para la
Caracterización y Determinación Taxonómica de
Levaduras Marinas de la Costa Occidental de Baja
California Sur
Taxonornies are not neutral hat racks for the pristine facts of nature
They are theories that create and reflect the deep structure
ofscience and hurnan culture
S J Gould 1985
1 INTRODUCCION
l Generalidades
La mayor parte de la diversidad genØtica del planeta se ubica en las formas
microbianas sin embargo se conoce relativamente poco de la diversidad de los
microorganismos mismos que son potencialmente utilizables en aplicaciones
biotecnológicas Nuestro conocimiento de la biodiversidad microbial es limitado debido
a que al utilizar tØcnicas estÆndar no se pueden cultivar la mayoría de los organismos
que ocurren naturalmente 99 al no ser cultivados no pueden estar disponibles
para su utilización Hugenholtz y Pace 1996 Lamentablemente se desconoce como se
han visto afectadas las comunidades de microorganismo s marinos por las actividades
humanas pese a que se reconoce su gran importancia ecológica y potencial
biotecnológico
Actualmente se reconoce a la conservación como una de las principales
actividades en los programas de investigación y educación En este contexto los
microorganismos tienen un particular e importante potencial La Convención para la

Biodiversidad celebrada en Río de aneiro en 1992 y a la que asistieron mas de 160
países Iwu 1996 ha dado especial ímpetu a la preservación de la diversidad de
especies en una escala global al desaparecer plantas y animales a tasas sin
precedentes una proporción de los microorganismos asociados desaparecen con ellos
El problema es cómo medir esta diversidad y desarrollar metodologías para
caracterizar y preservar aquellas especies que aœn no han sufrido el impacto de las
actividades humanas particularmente de aquellas que puedan tener un valor
económico
Cualquier tipo de anÆlisis acerca del valor de preservar la biodiversidad
requiere la atención de varias disciplinas Una de ellas debe ser necesariamente la
microbiología marina Sin embargo para poder profundizar en cualquier tema
relacionado a la importancia de la preservación de la diversidad biológica y
especialmente de los microorganismos marinos se deberÆ realizar un estudio
taxonómico Esta información serÆ de valor para las industrias que utilizan levaduras ya
sea en la bœsqueda de nuevas enzimas rutas metabólicas saborizantes o productos
farmaceœticos
Es evidente que una porción altamente significativa de la diversidad biológica en
el planeta se perderÆ en la primera mitad del próximo siglo EstÆ claro tambiØn que
esta pØrdida podrÆ tener un muy serio impacto negativo en la sociedad El nœmero de
especies actualmente disponibles para su uso se podrÆ reducir tanto por la pØrdida del
germoplasma como de la variabilidad genØtica impidiØndonos la futura explotación de
recursos nuevos y potenciales Wilson y Petee 1986
Es un buen momento para reconocer el papel que desempeæan las comunidades
de microorganismos marinos Es factible el proponer un estudio para conocer y
determinar la diversidad biológica de estas comunidades probablemente muchos de
los ecosistemas en los que viven se encuentran parcial o totalmente daæados El
descubrimiento de nuevos organismos permitirÆ alcanzar un mejor entendimiento del
ecosistema global el cual se basa despuØs de todo en el mundo microbiano Las
comunidades microbianas marinas ofrecen una riqueza de biodiversidad que podemos
comenzar a cosechar Hugenho1tz y Pace 1996
2

Las levaduras son un grupo de microorganismos eucariotes filogenØticamente
diverso Kurtzman y Phaff 1987 que pertenecen al Reino Fungi En un estadio de su
ciclo vital aparecen como cØlulas simples y se pueden reproducir por fisión o
gemación o una combinación de ambas Kreger van Rij 1969 Esta definición incluye
un gran nœmero de organismos que no forman un grupo taxonómico natural debido a
los diferentes ciclos de vida que presentan Davenport 1980 Se incluyen en la división
Eurnycota segœn la clasificación de Ainsworth 1973 y dadas sus características de
reproducción sexual se agrupan en tres subdivisiones Tabla 1 La primera subdivisión
Ascomycotina incluye a las levaduras que pueden formar esporas de origen sexual
dentro de ascas ascosporas en virtud de esto dichas levaduras pertenecen a la clase
Ascomycetes Kreger van Rij 1984 La segunda subdivisión Basidiomycotina estÆ
integrada por levaduras capaces de formar esporas externas en basidios o esterígmas
y comprende a los organismos agrupados en la clase Basjdjomycetes La tercera
subdivisión Deuteromycotina la integran levaduras que no presentan un estadio sexual
en su ciclo vital por lo que este grupo estÆ conformado por los denominados hongos
imperfectos pertenecientes a la clase Deuteromycetes
Tabla l Clasificación taxonómica de levaduras Martínez 1989
RŒmtiJ I Wí IIIJllliI rW@ ftCmH @@ @@ oltiif FiHfdtii tt
Fungii Eumycota Ascomycotina Ascomycetes Endomycetales Sphermophtoraceae
Saccaromycetaceae
Basidiomycotina Basidiomycetes Ustilaginales Filobasidiaceae
Tremellales Sirobasidiaceae
Tremellaceae
Deuteromycotina Deuteromycetes Blastomycetales Cryptococcaceae
Sporobolomycetaceae
Las levaduras poseen un amplio espectro de producción de metabolitos que
pueden ser utilizados por el hombre por ejemplo vitaminas aminoÆcidos enzimas
polisacÆridos extracelulares proteínas etc Por otra parte se ha intentado mejorar el
rendimiento de las levaduras para desarrollar niveles mÆs altos de producción de estos
3

metabolitos utilizÆndolas como agentes transferentes o receptores de genes Botstein y
Fink 1988 ampliando así sus posibilidades de comercialización Martínez 1989
En los œltimos 40 aæos se han reunido evidencias suficientes para establecer que
las levaduras son parte constitutiva de las poblaciones microbianas marinas Esta
aseveración es el resultado de la observación de levaduras en el mar desde los inicios
de la microbiología marina Durante la Øpoca de los sesentas el interØs por la detección
selectiva de levaduras en substratos marinos se incrementó a tal grado que
microbiólogos de la URSS India Japón Alemania y Portugal y otros países comenzaron
a realizar estudios de este tipo gracias a ello se adquirieron algunos conocimientos
sobre la distribución de algunas poblaciones de levaduras marinas y su composición de
especies HernÆndez Saavedra 1990
Las levaduras tienen una amplia distribución ya que han sido aisladas de
diversos tipos de substratos correspondientes a un variado tipo de hÆbitats dentro de
los que se incluyen los ambientes marinos levaduras marinas Dentro de este grupo
se incluyen a todas las levaduras y organismos levaduriformes que son capaces de
construir y perpetuar poblaciones en el medio marino o aquellas en las que su
reproducción y crecimiento ocurren preferentemente en el mar u óptimamente a la
concentración normal de sales en el mar 2 4 4 0 de cloruro de sodio
principalmente HernÆndez Saavedra 1990
Se ha observado que las poblaciones de levaduras son mÆs abundantes en aguas
costeras encontrÆndose una gran variedad de especies que se incluyen en las tres
divisiones de hongos superiores En ocØano abierto en donde los desechos orgÆnicos
son menores se han aislado en un menor nœmero disminuyendo tambiØn la variedad
de especies encontradas
Se han enlistado 177 especies de levaduras marinas aisladas a partir de agua
sedimentos plantas animales y detritos del medio marino Uden y Fell 1968
Kohlmeyer y KohIrneyer 1979 Sin embargo la mayoría son formas halotolerantes de
origen teïrestre v gr Candida Torulopsis Cryptococcus Trichosporon y
Saccharomyces Las œnicas especies estrictamente marinas son Metzchnikowia zobeDii
y M kIissii Rheiheimer 1974 Kohlmeyer y Kohlmeyer 1979
4

2 Antecedentes
No ha sido sino hasta hace algunos aæos que se ha despertado el interØs de los
científicos en MØxico hacia el estudio de los microorganismo s que viven y se
desarrollan en aguas marinas A nivel internacional estas investigaciones han estado
principalmente dirigidas al estudio de la ecología genØtica y evolución de las bacterias
marmas y mÆs recientemente a su aprovechamiento biotecnológico
HernÆndez Saavedra 1990
HernÆndez Saavedra realizó estudios sobre la distribución de levaduras marinas
en la costa occidental de Baja California Sur en los que reportó el aislamiento de 216
cepas de levaduras correspondientes a 14 gØneros 26 especies y dos variedades
HernÆndez Saavedra 1990 Al correlacionar los parÆmetros fisicoquímicos
temperatura salinidad y oxígeno disuelto con la distribución de cada especie se
encontró que cada una estÆ irœluenciada de manera muy particlÙar
HernÆndez Saavedra etaJ 1992 1995
Los microbiólogos han estudiado la taxonoIlÚa de las levaduras por mÆs de un
siglo pero aœn y cuando se ha obtenido un progreso considerable no se ha podido
desarrollar un sistema de clasificación adecuado Kurtzman y Phaff 1987 La ubicación
taxonóI1lÌca y clasificación de las levaduras reslÙta aœn confusa DespuØs de que
Leeuwenhoek las describió por vez primera en 1680 se obtuvo muy poca información
adicional sobre la morfología y no fue sino hasta 1825 que Starting Cagniard de la
Tour Kutzing y Schwann observaron que las levaduras se reproducen por gemación
HernÆndez Saavedra 1990
El sistema de taxonoIlÚa usado en la actualidad es el reslÙtado del desarrollo e
integración subsecuente de tres importantes aportaciones al problema de la
identificación y clasificación de las levaduras La primera de estas aportaciones ocurrió
en 1870 cuando Reess intentó clasificar a las levaduras hasta entonces conocidas con
base en su morfología cellÙar y a su forma de reproducción La segunda contribución
fimdamental ocurrió en 1883 como reslÙtado de los estudios de Hansen sobre ClÙtivos
puros Con estos se observó que las levaduras difieren en sus habilidades para
s

fermentar carbohidratos La prÆctica moderna de emplear características fisiológicas
para diferenciar especies se sostiene en este descubrimiento La tercera contribución
aconteció a principios del siglo XX cuando Guillermond realizó estudios sobre la
reproducción sexual y la filogenia de las levaduras ascosporógenas van der Walt
1970
El primer esquema completo de clasificación para levaduras esponøadas fue
preparado por Stelling y Dekker en 1931 En 1970 se lograron unificar los criterios de
clasificación Lodder determinó que para entonces el nœmero de especies de levaduras
era 341 vanderWalt 1970
En la actualidad el compendio de clasificación y taxonomía mÆs completo para
levaduras es el elaborado en 1983 por Lodder y Kreger van Rij HernÆndez Saavedra
1990 Hasta la fecha se han descrito alrededor de 600 especies sin embargo se
considera que esto es tan sólo una pequeæa fracción del total de especies existentes
Roberts y Wildman 1995
MØtodos de Clasificación e Identificación
La clasificación de las levaduras se ha basado principalmente en criterios
morfológicos y fisiológicos Sin embargo utilizar exclusivamente este tipo de pruebas
tiene la desventaja de que cualquier variación en la reacción a la prueba puede reslÙtar
en una identificación errónea Kurtzman y Phaff 1987 por lo que varios autores han
discutido su valor como herramienta taxonómica Winge y Roberts 1949 Fell et al
1973 Phaff Y Price 1977 Yamazaki y Komagata 1981 Kurtzman y Phaff 1987 Por lo
tanto se han considerado otros criterios como la composición química de la pared
cellÙar la serología la secuencia de Æcidos nucleicos y la determinación del contenido
de G C Storck et al 1969 No obstante dichas tØcnicas frecuentemente carecen de
sensibilidad y especificidad Lee etal 1985
Las primeras investigaciones en sistemÆtica que utilizaron molØclÙas fueron
desarrolladas con proteínas Las primeras aplicaciones de electroforØsis revelaron una
riqueza de variación proteica en y entre especies Lewontin y Hubby 1966 Este
acercamiento electroforØsis de isoenzimas ha generado una base enorme de datos
6

comparativos y es comœnmente el mÆs usado en sistemÆtica moleclÙar Hillis y Moritz
1990
La electroforØsis de aloenzimas se basa en la idea de que la diversidad genØtica
se desarrolla con el tiempo Esto implica que organismos genØticamente distantes
tendrÆn mayores diferencias en la composición de sus proteínas y que los organismos
mÆs emparentados tendrÆn un alto grado de similitud
Con el paso de los aæos otros criterios han demostrado ser importantes
auxiliares en la clasificación e identificación de especies Para estudios taxonómicos de
algunas especies de Candida se han aplicado varias tØcnicas especializadas Por
ejemplo espectro de resonancia magnØtica de protones de mananas y polisacÆridos
que contienen manosa estudios inmunoelectroforØticos de antígenos patrones de
bandeo de ADN cromosómico mitocondrial y ARN ademÆs del anÆlisis de Æcidos
graso s de cadena larga Bak y Stenderup 1969 Vancanneyt etaJ 1991
El estudio de cariotipos mediante tØcnicas electroforØticas ha mostrado ser una
herramienta fundamental y mÆs reproducible para la identificación de agentes
etiológicos tales como Candida albicans que las tØcnicas fisiológicas y bioquímicas
Mahrous etaJ 1990
Recientemente el anÆlisis de ADN genómico de levaduras con enZlmas de
restricción restriction endonuclease fingerprinting ha demostrado ser una herramienta
efectiva en la diferenciación de algunas cepas de levaduras Lee et al 1985 Por
ejemplo con este mØtodo ha sido posible identificar diferentes grupos de especies
dentro del gØnero Saccharomyces
Sin embargo en vista de la rÆpida tasa de evolución del ADN mitocondrial
ADNmt el estudio de los patrones electroforØticos del ADNmt generados con enzimas
de restricción es una herramienta mÆs sensible en la diferenciación de cepas de
levaduras que la digestión de ADN genómico total
Los experimentos de hibridación deADN ADN que determinan la fidelidad de la
complementación de los Æcidos nucleicos de diferentes cepas son valiosos para
delimitación de especies Olsen 1990 Se pueden dar varios ejemplos para demostrar
la validØz de la homología de ADN ADN respecto a la de experimentos de taxonoIlÚa
7

tradicional En los gØneros Torulopsis y Candida se ha acumulado evidencia de que la
presencia o ausencia del desarrollo hifal no es necesariamente una buena característica
para diferenciación de especies sin embargo la homología del ADN ha demostrado
serlo Olsen 1990
El anÆlisis de amplificación al azar de ADN polimórfico RAPD es un mØtodo
rÆpido y sencillo para obtener patrones de ADN utilizados para la identificación de
bacterias de interØs mØdico Listería spp Helicobader pyloJi y Haemolimphus
somnus Sin embargo para obtener resultados óptimos antes deberÆn probarse un
gran nœmero de oligonucleótidos Abed et al 1995 Esta es una herramienta que
consume menos tiempo para la identificación de organismos y se ha demostrado que es
œtil para la identificación de diferentes especies de levaduras Se ha descrito el uso de
esta tØcnica para discriminar entre especies de Saccharomyces cereVlSlae
Zygosaccharomyces baiJjjy Z rouxii Baleiras Couto etal 1994
Por œltimo con pocas especies se han hecho estudios de homología del ARN
ribosomal y ADN rARN ADN Estos estudios son muy prometedores en cuanto a la
demostración de distancias intergenØricas y relaciones mÆs lejanas entre especies de
levaduras Olsen 1990
TaxonoIIÚa numØrica
El desarrollo de la taxonomía numØrica tiene sus orígenes con dos publicaciones
simultÆneas en 1957 una de H A Sneath y la otra de Charles D Michener y Robert R
Sokal La taxonoIIÚa numØrica propone basar las clasificaciones enteramente en las
semejanzas definiendo una clasificación natural como aquella cuyos taxa poseen
miembros en cierto sentido mÆs similares entre sí de lo que son comparados con
miembros de otros taxa
En la taxonoIIÚa numØrica los objetos a clasificar son denominados unidades
taxonómicas operacionales UTO Las clasificaciones basadas en la taxonomía
numØrica se basan en una gran cantidad de medidas que se representan
numØricamente o se codifican de tal forma que las diferencias entre ellas son
proporcionales a las diferencias entre las UTOs
8

Todos los caracteres y las unidades taxonómicas a clasificar se arreglan en una
matriz de datos y las similitudes entre todos los pares posibles de UTO se computan
con base en todos los caracteres Una forma de representar las semejanzas es la
distancia entre las UTO en un espacio multidimensional de modo que los objetos que
tengan mayor similitud se encontrarÆn muy cercanos en la grÆfica objetos diferentes
tendrÆn distancias mayores entre sí
Las similitudes entre pares de UTO son evaluadas generando una matriz de
similitud que muestra el valor de la similitud de cada UTO con respecto a las demÆs
Sokal 1966 Para obtener clasificaciones mÆs precisas se han desarrollado una
variedad de procedimientos de agrupamiento numØrico sin embargo no existe un
mØtodo de agrupamiento generalmente aceptado Algunos de los mÆs comœnmente
aceptados son
a Ligamiento s˝mple TambiØn conocido como el vecino mÆs cercano
b Ligamiento completo Conocido como el vecino mÆs distante
c Ligamientopromedio no ponderado UPGMA unweighted pair group
method using arithmetic averages
d Ligamiento promedio ponderado VVPGMA weighted paJrgroup
method using arithmetic averages
e MØtodo de Ward
Los resultados de una clasificación numØrica generalmente se representan por
medio de un fenograma Estos diagramas arboriformes indican la similitud entre las
UTO Debido a que los fenogramas colapsan relaciones multidimensionales en dos
dimensiones existe una apreciable distorsión de las relaciones originales que se
muestran en la matriz de similitud Para prevenir esto al realizarse estudios
taxonómicos mediante la taxonomía numØrica se hacen estimados del grado de
distorsión en un fenograma
9

De acuerdo con lo anteriormente expuesto se pretende estandarizar un mØtodo
auxiliar para la caracterzación e identificación de Levaduras Marinas que sea eficaz y
rentable
lI OBJETNO GENERAL
A Determinación de la tØcnica electroforØtica mÆs eficaz para la caracterización de 78
cepas de la Colección de Levaduras Marinas del CIBNOR
lII OBJETNOS ESPECIF1COS
A Caracterizar la Colección de Levaduras Marinas utilizando los criterios de
clasificación tradicional
B Caracterizar la Colección de Levaduras Marinas utilizando como criterio el
patrón electroforØtico deproteínas solubles totales
C Evaluar la posibilidad de utilización de patrones electroforØticos de ADN
genómico obtenidos mediante digestión con enzimas de restricción como herramienta
taxonómica
D Evaluar la posibilidad de utilización de patrones electroforØticos de ADN
generados a travØs de la tØcnica de RAPD como herramienta taxonómica
E Correlacionar los patrones electroforØticos obtenidos de proteínas y ADN con
la ubicación taxonómica de cada cepa obtenida mediante mØtodos convencionales
N MATERIALES Y METODOS
l Organismos a estudiar y organismos de referencia
Las cepas estudiadas Tabla 2 pertenecen a la Colección de Levaduras Marinas
del Centro de Investigaciones Biológicas del Noroeste CIBNOR constituida apartir del
trabajo de investigación realizado por HernÆndez Saavedra 1990 Se utilizaron como
cepas de referencia Saccharomyces cereviseae cepa X2l80 y Schizosaccharomyces
pombe cepa M40
10
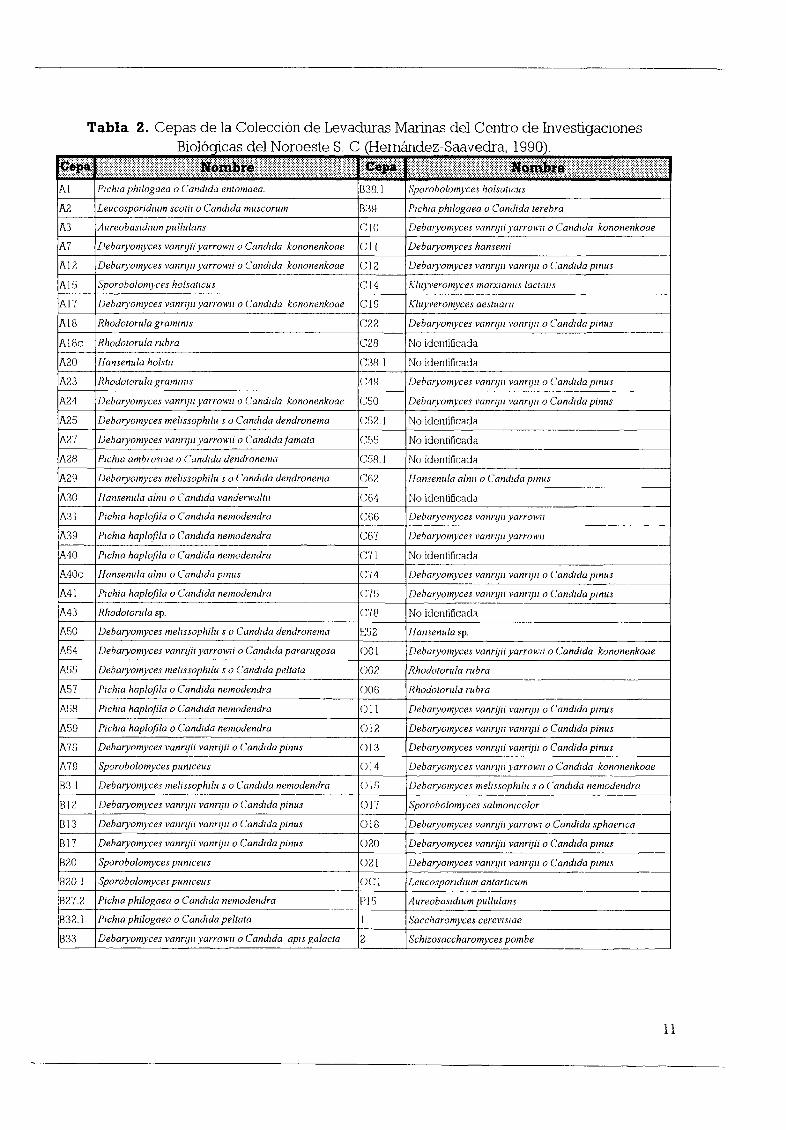
Tabla 2 Cepas de la Colección de Levaduras Marinas del Centro de InvestigacionesB l d 1 N t S C H d S dr 199010 o91cas e oroes e eman ez aave a
Þ iW j j Ij I j j j jI j li f j j j j j j j j j j j j m ïiI j j j j j j j liEti I j I j j j jAl Plchw phllogaea oCandlda entomaea 838 1 Sporobolomyces holsarlcus
A2 LeucospondlUm scorll oCandlda muscorum 839 Plchw phllogaea oCandida rerebra
A3 Aureobasldlllmpululans CJO Debaryomyces vanrljll yarrowlI oCandlda kononenkoae
A7 IJebaryomyces vanrljll yarrowlI oCandlda kononenkoae CII Debaryomyces hansemi
AI2 Debaryomyces vanrljll yarrowlI oCandida kononenkoae C12 Debaryomyces vannJII vanrljll oCandlda pnus
A15 Sporobolomyces holsar cus CI4 Kluyveromyces marxwnus lactatls
Al7 Debaryomyces vanrljll yarrowlI oCandlda kononenkoae CI9 Kluyveromyces aesruam
AI8 Rhodotorula grOlmms C22 Debaryomyces vannJII vanrljll o Candlda pnus
A18c Rhodororula rubra C28 No identificada
A20 Hansenula holsrll C3fl No identificada
A23 Rhodotorula gramlllls C49 Debaryomyces vanrljll vanrlI oCandlda pnus
A24 Debaryomyces vannJII yarrowlI oCandlda kononenkoae C50 Debaryomyces vannJII vannJII oCandlda pnus
A25 Debaryomyces mel1ssophllu s oCandlda dendronema C52 1 No identificada
A27 Debaryomyces vannJII yarrowlI oCandldafamata C55 No identificada
A28 Plchw ambroswe oCandlda dendronema CS8 Noidentilicada
A29 Debaryomyces mel1ssophllu s oCandlda dendronema C62 Iansenllla alnll oCandlda pnIlS
A30 Iansenllla alnll oCandlda vanderwaltll C64 No identificada
A31 P chw haplofila oCandlda nemodendra CGG Deboryomyces vanrljll yarrowlI
A39 Plchw haplofila oCand da nemodendra CG7 Debaryomyces vanrljll yarrowlI
MO Plchw haplofila oCandida nemodendra C71 No identificada
MOc Hansenula alnll oCandida pnIlS C74 Debaryomyces vanrljll vanrljll oCandido pnus
MI P chia haplofila oCandida nemodendra C75 Debaryomyces vanrljll vanrljll oCandlda pnIlS
M3 Rhodotorula sp C78 No identificada
A50 DebOlomyces mel1ssophlll1 s oCandlda dendronema 52 ansenllla sp
A54 Debaryomyces vanrljˇ1 yarrowll oCandlda pararugosa 001 Debaryomyces vanrljiiyarrowlI oCandida kononenkoae
A5S Debaryomyces lIIel1ssophllu s o Candlda peltata 002 Rhodotorula rubra
A57 Plchw haplofila oCandida nemodendra 006 Rhodororllla rubra
A58 Pichia haplofila oCandlda nemodendra 11 DebOlomyces vanrljii vannJII oCandida pnIlS
A59 Pichia haplofila o Candlda nemodendra 012 Debaryomyces vanrljll vanrljll oCandlda pinus
A75 Debaryomyces vannJii vannJii oCandida pinus 3 Debaryomyces vannli vanrljil oCandida pinlls
A79 Sporobolomyces pumceus 014 Debaryomyces vannJII yarrowlI oCandlda kononenkoae
B3 Debaryomyces mel1ssophllu s oCandlda nemodendra 015 DebOlomyces mel1ssophllu s oCandlda nemodendra
812 Debaryomyces vanrll vanrljll oCandlda pinlls 017 Sporobolomyces salmomcolor
B13 Debaryomyces vanrljˇ1 vanrljll oCandlda pnus 018 Debaryomyces vanrljll yarrowl oCandida sphaenca
BI7 Debaryomyces vanrlI vanrljil oCandida pnus 020 Debaryomyces vanrljˇ1 vanrljii oCandlda pmlls
B20 Sporobolomyces pumceus 21 Debaryomyces vannJII vanrlI oCandlda pinus
820 1 Sporobolomyces pumcells Cl LellcospondlUm antartlcllm
B272 Pichia phllogaea oCandlda nemodendra P S AureobasidlUm pululans
B32 PIChlO phllogaea oCandlda peltara l Saccharomyces cerevlSae
B33 Debaryomyces vanrlIyarrowlI oCandlda apls galacta 2 Schizosaccharomyces pombe
11

2 Reactivación de las cepas
Las cepas preservadas en glicerol al 30 a 70oC Esquema 1 nœmero 1 se
descongelaron a temperatura ambiente y se inocularon en matraces Erlenmeyer de 125
mL conteniendo 50 mL de medio M l líquido ApØndice 1 Los cultivos se incubaron a
temperatura ambiente en agitación orbital constante a 100 rpm para su reactivación
Esquema 1 nœmero 2 Una vez que se obtuvo crecirrøento cada cepa se sembró por
estría cruzada en cajas de Petri con medio M l agar ApØndice 1 para verificar su
pureza Las placas se incubaron invertidas a temperatura ambiente durante 48 h
Esquema 1 nœmero 3
Una vez que se verificó la ausencia de contaminación las placas se utilizaron a
para la caracterización de las cepas por mØtodos convencionales morfología celular y
colonial asimilación de fuentes de carbono y nitrógeno y b para preparar preinóculos
para preservar los cultivos y para la obtener la biomØlsa utilizada en los protocolos de
extracción de proteínas y de ADN
3 Caracterización de las cepas mediante su morfologíl celular y colonial
Morfología celular De las placas de medio M lagar Esquema 1 nœmero 3 se
tomó una asada y se transfirió a un tubo de ensayo conteniendo 2 mL de medio
YFD líquido ApØndice 1 DespuØs de 48 h de incllbación a temperatura ambiente
Esquema 1 nœmero 4 se tomó una gota de cada Cullivo y se observó al microscopio
determinÆndose las siguientes características forma de la cØlula redonda ovalada
apiculada triangular elongada y tipo de gemación polar bipolar multilateral etc En
los cultivos de 2 mL medio YFD que se utilizaron para revisar la morfología celular se
observaron las siguientes características presencia o ausencia de sedimento tipo de
crecimiento homogØneo o heterogØneo formación de anillo completo o incompleto
formación de película
Morfología colonial Las cepas se sembraron en placas de medio YFD agar
ApØndice 1 por estría cruzada Esquema 1 nœmero 5 DespuØs de 48 h de
incubación a temperatura ambiente se evaluaron las siguientes características de
12

sooOvv
g mtIl
OI m
E OE tIl oo
@o łlO
c
E f0 z2 M
EO m
O I tIlO m
Q tIl s t32 g cO VoOJ
łl
Πz
0O
loe
Eo
O1
0o
o g tIl
Z mo MO 104 o
S ł c
tr @ o O OJ
0ŒOI VI
1 Eo
o
sE 00O M
E g ªQ O E OO O ID O
M m stIl E O v o
oV Qo mm E vf s
s m
O OO m C
tIl s ï Õ Og s
o VtIl 0tIl Q Qm v
Mm tiloVM Sm mO Q m
M OJO E
88 8
oot
I
m
oC
m
oM
OV
t1I
sO
m tIlm
s M
o oc
ao
OJ
ðt1Imo t1I
m8M O
oO
O g1O I
o
OZ Q
041
o E
e
tIlms tIl
O0
m
8 Õ0
Ó IDO
oc
INOm
E tIl
00tO
cd
Q
Q
m
51o
ÕÖo
Q
z SO m
Þ QÖ
O oO Ss q
Qo ÖV mV
ŒmM
m
0
@t
lll
S
iCIl1tr11I
W
Þti
Þ
OłlM
OO
Qv trm
Gl O
Ec
EN

colonias aisladas tamaæo color tipo de superficie lisa rugosa elevación cóncava
convexa umbonada tipo de bordes completos incompletos brillo textura dura
cremosa butirosa y aspecto hœmedo seco algodón
4 Pruebas de asimilación de fuentes carbono y nitrógeno
4 1 Preparación del inóculo
De placas de medio YFD agar Esquema 1 nœmero 5 se tomó una asada y se
transfirió a tubos de ensayo que contenían 3 mL de agua destilada estØril Esquema 1
nœmero 6 El inóculo se mantuvo en estas condiciones durante 5 días con la finalidad de
agotar las reservas de las cØlulas y forzarlas a utilizar los nutrientes a probar A este
proceso se le conoce como dieta Una vez transcurrido este tiempo se procedió a
sembrar las cepas en las placas que contenían cada una de las diferentes fuentes de
carbono y de nitrógeno
4 2 Asimilación de carbohidratos
Preparación del medio de cultivo Se pesaron 13 4 9 de medio YNB yeast
nitrogen base DIFCO y se disolvieron en 200 mL de agua destilada esterilizÆndose por
filtración Sterile Acrodisc de 045 lm libre de pirógenos de este modo se obtuvo una
solución stock de medio de cultivo 10x
A continuación se prepararon 30 matraces Erlenmeyer de 125 rnL conteniendo lo
siguiente 1 9 de agar 0 5 9 del carbohidrato a probar y 45 rnL de agua destilada Este
medio se esterilizó en autoclave por 10 a 121 oe Una vez que la temperatura del medio
alcanzó aproximadamente 500e se adicionaron 5 rnL del stock YNB 10x en condiciones
de esterilidad
Posteriormente se procedió al vaciado en placa Como control negativo se
utilizaron placas consistentes en medio YNB sin fuente de carbono los diferentes
carbohidratos utilizados se enlistan en la Tabla 3
14

Tabla 3 Fuentes de carbono utilizadas en las pruebas de asimilación
li D Glucosa D Galactosa L Sorbosa D Manosa D Fructosa L Fucosa
I Ribosa D Xilosa D Arabinosa
1 11 11 D Melezitosa Sacarosa D Maltosa D Trehalosa
f lìigBI Salicina
lliiliB i D Melibiosa
1 i IIBiii Lactosa
li II i i i Glicerol D Sorbitol lnositol
lliill Etanol Metanol
1 1 I˛I i Almidón
lsII˛ Succínico Cítrico
IBí j j L Ranmosa D Rafínosa
4 3 Asimilación de fuentes de nitrógeno
Preparación del medio de cultivo Se disolvieron 1 17 g de medio YCB yeast
carbon base DIFCO mÆs 0 078 g de la fuente de nitrógeno a utilizar en 50 m1 de agua
destilada estØril Esta solución 2x se esterilizó por filtración Sterile Acrodisc de 0 45
lm libre de pirógenos
A parte se disolvió 1 g de agar en matraces Erlenrneyer de 125 m1 conteniendo
50 m1 de agua destilada mismos que se esterilizaron en autoclave por 20 a 1210C Una
vez que la temperatura del medio alcanzó aproximadamente los 500C se mezclaron las
dos soluciones y se procedió avaciar el medio en placas
Las fuentes de nitrógeno utilizadas en este ensayo fueron nitrato de sodio nitrito
de sodio y L lisina El control negativo se preparó mezclando 25 m1 de stock YCB 2x y
25 m1 de agar
La tØcnica de siembra de las placas de YCB y YNB fue por goteo una gota
corresponde a aproximadamente 50 fJL mediante una pipeta de transferencia estØril
Samco Scientific Inc A las 48 h de incubación a temperatura ambiente se revisó el
crecimiento considerÆndolo como positivo o negativo con respecto a la placa de
control negativo Esquema 1 nœmero 7
15

5 ElectroforØsis de proteínas solubles totales
5 1 Producción de biomasa
Para la producción de biomasa se preparó un preinóclÙo a partir de un clÙtivo
en agar inclinado con medio MI crecido durante 24 h a temperatura ambiente el cual
se resuspendió en un matraz Erlenmeyer de 125 m1 conteniendo 50 m1 de medio
MI liquido Esquema 1 nœmero 11 De esta suspensión cellÙar se tomaron 5 m1 que se
utilizaron para inoclÙar un matraz Erlenmeyer de 125 m1 conteniendo 50 m1 de medio
MI Este clÙtivo se incubó durante 18 h para alcanzar la fase exponencial media bajo
agitación orbital constante a 150 rpm a temperatura ambiente
En la propagación se utilizó 1 m1 del preinóclÙo para inOClÙar matraces
Erlenmeyer de 125 m1 conteniendo 50 m1 del medio de clÙtivo MI Los clÙtivos se
incubaron a temperatura ambiente bajo agitación orbital constante a 150 rpm Esquema
1 nœmero 12
DespuØs de 48 h de incubación los 50 m1 del clÙtivo se cosecharon por
centrifugación a 12 100 x g rmax durante 10 minutos a 40C en tubos Beckman de
policarbonato de 25 m1 Esquema 1 nœmero 13
5 2 Obtención de proteínas solubles totales
Los paquetes cellÙares se lavaron tres veces con 10 m1 de una solución
amortiguadora de fosfatos 50 rnM pH 7 8 Se determinó el peso hœmedo de los
paquetes cellÙares en una balanza analítica Ohaus Analytical Plus igualÆndose la
biomasa de todas las cepas a0 9 g
En tubos Eppendorf de 1 7 m1 se colocaron 0 9 9 de biomasa hœmeda 200 IJL de
perlas de vidrio y 200 IJL solución amortiguadora de fosfatos 50 rnM pH 7 8
agitÆndose en vortex durante 15 mina 40C
Una veztranscurrido este tiempo las muestras se centrifugaron a 17 800 x g rmax
durante 5 minutos a temperatura ambiente en una microcentrífuga Beckman E
Posteriormente se recuperó el sobrenadante y se calentó a ebullición en baæo María
por 4 mino Este homogeneizado se centrifugó nuevamente para retirar restos de
paredes cellÙares A los sobrenadantes recuperados se les agregaron 3 volœmenes de
16

acetona fría para precipítar las proteínas incubÆndose 15 minutos a 20oC Una vez
hecho lo anterior se centrifugó a 17 800 x g por 5 min Esquema 1 nœmero 13 El
precipitado se disolvió en 70 fJL de solución amortiguadora de fosfatos 50 rnM pH 7 8
Y se procedió a cuantificar las proteínas totales mediante el mØtodo de Bradford 1976
ApØndice 2
5 3 ElectroforØsis en gel de poliacrilamida PAGE
A 60 fJL de las muestras de proteínas totales 600 fJg se les adicionaron 15 fJL de
Buffer de carga ApØndice 3 y se calentaron a ebullición en baæo María durante 4
minutos
Se prepararon geles discontinuos de acrilamida bisacrilamida SDS 3 5 y 12
segœn la tØcnica de Laemrrøi 1970 ApØndice 3 Cada pozo se cargó con 400 fJg de
proteína 40 fJL Y la electroforØsis se realizó en una cÆmara Miniprotean II Bio Rad a
100 V a temperatura ambiente Como estÆndares de peso molecular se usaron los Wide
Range Molecular Weight de Bio Rad cuya composición se muestra en la Tabla 4
Tabla 4 Marcadores de peso molecular utilizados como estÆndares en las
electroforØsis
i i I ˛II I ii ii i iii i i i ii iii i I ii ii i i i i i i i i i iMlii ltiilllMiosina Mœsculo esquelØtico de ratón 200 000
ß galactosidasa E coli 1 16
250Fosforilasa BMœsculo de ratón97
400Albœmina SØrica Bovina 66
200 Ovoalbœmina Clara dehuevo de gallina 45
000Anhidrasa carbonica Bovina 3 1
000Inhibidor de tripsina Soya 2 1
500 Lisosima Clara dehuevo de gallina 14
400 Aprotinina PÆncreas bovino 6
500 Los gelessetiæeron por inmersión enuna solución deAzul Brillante de
Coomasie G 250 yelexceso decolorante seretiró mediante lavados repetidos consolución
para desteæir ApØndice
317

5 4 Densitometría
Los geles se sometieron a densitometría Beckman Appraise Junior bajo las
3iguientes condiciones de barrido longitud de onda 600 nm longitud del barrido 60
rrun ancho del barrido 3 rnm
En cada densitograma se determinaron las distancias de migración de cada pico
considerando al origen como punto de aplicación De este modo y despuØs de haber
aplicado un anÆlisis de regresión lineal a los estÆndares de peso molecular de cada gel
se calculó la masa molecular de cada banda de proteína
6 AnÆlisis de los datos
Los datos referentes a la morfologia celular y colonial se analizaron junto con los
relativos a la asimilación de fuentes de carbono y nitrógeno mediante anÆlisis de
taxonomía numØrica de forma tal que se construyó una matriz de similitud que engloba
las características que convencionalmente son utilizadas para la taxonomía de levaduras
En el caso del anÆlisis de los datos de proteínas totales se desarrolló una matriz
de similitud exclusivamente con datos relativos a las masas moleculares de las bandas
proteicas
Adicionalmente se realizó un anÆlisis de similitud de las cepas A18 A18c A23
A43 002 006 A75 B12 B13 E17 C12 C22 Y C49 gØnero RhodotoruJa y
Debmyomyces vanriþi vanrijil tanto de las pruebas convencionales como de los
patrones electroforØticos de proteínas
El anÆlisis numØrico se basó en el coeficiente de correlación Pearson l r y el
agrupamiento de las cepas se llevó a cabo por el mØtodo de ligamiento promedio no
ponderado UPGMA Todos los cÆlculos el anÆlisis para el agrupamiento la matriz de
similitud y el dendograma se realizaron por medio del programa Statistica para
Windows versión 4 5 de StatSoft en una computadora Exacto 486
18

7 ElectroforØsis de ADN genómico
7 1 Cepas utilizadas
Debido a que se pretende evaluar la sensibilidad de las tØcnicas electroforØticas
de ADN se escogieron œnicamente dos grupos de cepas A18 A18c A23 A43 002 006
del gØnero RhodotoruJa y A75 B12 B13 B17 C12 C22 C49 que corresponden a
Debaryomyces vannjii vanrijii Se eligió al primer grupo para este ensayo debido a su
heterogeneidad se incluyen varias especies dentro de un mismo gØnero mientras
que el segundo grupo se eligió por la homogeneidad que presenta en cuanto a los
resultados obtenidos apartir de la caracterización morfológica y fisiológica
De una caja de Petri con medio YPD agar Esquema 1 nœmero 5 se tomó una
colonia y se transfirió a un tubo de ensayo que conteIÚa 5 m1 de medio YFD líquido
Esquema 1 nœmero 8 incubÆndose en agitación orbital constante a 150 rpm por 24 h
a temperatura ambiente
Del preinóculo anterior se utilizaron los 5 m1 para inocular matraces Erlenmeyer
de 500 m1 conteniendo 150 m1 del medio de cultivo YPD líquido que se incubaron a
temperatura ambiente bajo agitación orbital constante 150 rpm DespuØs de 24 h de
incubación los cultivos se cosecharon por centrifugación a 2 060 x g rmax durante 10
minutos a 40C en tubos Falcon de 50 m1 estØriles Esquema 1 nœmero 9
Para la extracción de ADN Esquema 1 nœmero 10 se utilizaron el mØtodo de
Rose y Broach 1991 para la producción de esferoplastos y una modificación del
mØtodo de Hoffman y Winston 1987 para el aislamiento de ADN total ApØndice 4
Todas las soluciones y material que se utilizó se esterilizó en autoclave a 1210 C por 20
min salvo en los casos que se indique
Para observar la proporción de ADN que se extrajo en relación con el ARN se
corrió una electroforØsis en gel de agarosa al 0 8 El gel se preparó disolviendo por
calentamiento 0 8 g de agarosa en 100 m1 de buffer TBE ApØndice 5 Una vez que se
alcanzó aproximadamente una temperatura de 450C se agregaron 3 fJL de una solución
10 mM de bromuro de etidio La solución se homogeneizó y se vació en el molde Una
19

vez que el gel polimerizó se sumergió en la cÆmara de electroforØsis Mini Sub Cell
Bio Rad que contenía 500 m1 de buffer TBE
A 5 flL de la muestra se le agregaron 5 flL de agua destilada y 5 flL de LB
ApØndice 5 posteriormente se cargaron en los pozos del gel La electroforØsis se
corrió a 70 V por 45 minutos Una vez que transcurrió este tiempo se retiró el gel y se
observó en un transiluminador con luz U V UVP lnc Modelo TM 36 con un pico de
longitud de onda de 302 nm Posteriormente se eliminó el ARN de las muestras
ApØndice 4 verificÆndose mediante una nueva electroforØsis Los resultados se
documentaron mediante fotografia bajo luz UV
Para la cuantificación del ADN purificado se preparó 1 m1 de una dilución 1 500
de cada una de las muestras Las muestras se leyeron a 260 280 320 nm en una cuveta
de cuarzo usando un espectrofotómetro Beckman segœn el mØtodo de
Warburg Christian 1942 Este mØtodo tambiØn incluye las razones entre las longitudes
de onda 260 280nm y 280 260 nm La longitud de 260 nm detecta Æcidos nucleicos la
de 280 detecta proteínas basÆndose en la presencia de aminoÆcidos aromÆticos La
longitud de onda de 320 nm se utiliza como corrección de fondo
De cada muestra se prepararon 200 flL de una solución de ADN conteniendo
1 flg flL
7 2 Digestión del ADN mediante enzimas de restricción y electroforØsis en geles
de agarosa
La digestión del ADN se realizó en un tubo Eppendorf en el cual se preparó una
mezcla de reacción conteniendo 1 flg de ADN 1 U de enzima ver tabla 5 2 5 flL de
buffer lOx ver tabla 5 llevÆndose a un volumen final de 25 flL con agua destilada
estØril En el caso de la enzima Kpn 1 en el tubo Eppendorf ademÆs del ADN la enzima
y el buffer se adicionaron 2 5 flL de un stock 10x de albœmina sØrica bovina metilada
Las enzimas que se utilizaron fueron Alu l Bam Hl Eco RI HinfI Kpn l y PstI
Boehringer Marmheim en la Tabla 5 presenta las secuencias que reconocen cada una
de estas enzimas y el buffer de incubación que se utilizó en cada caso
20

Las reacciones se incubaron durante 2 h a 370C Una vez transcurrido este
tiempo las muestras se almacenaron a 20oC hasta su anÆlisis
Tabla 5 Secuencias de reconocimiento de las enzimas utilizadas buffers de
incubación y volumen de actividad respectivos
II II iB II ii I i lii lIi I IAluI 5 AG CT 3 A 10
Bam HI 5 G GATCC 3 M 10
EcoRI 5 G AATTC 3 H 10
HinfI 5 G ANTC 3 H la
Kpn I 5 GGTAC C 3 L 10
PstI 5 CTGCAG 3 H 10
El ADN de cada cepa digerido por cada enzima se analizó en geles de agarosa
al 1 De cada reacción se tomaron 15 IJL Y se le agregaron 2 IJL de LB Cada muestra
se cargó en el gel corriØndose la electroforØsis a 70 V durante 180 mino Posteriormente
se procedió a observar el gel en un transiluminador con luz UV UVP Inc Modelo
TM 36 con un pico de longitud de onda de 302 nm y se fotografió para anÆlisis
posteriores Como estÆndares se utilizaron los DNA Molecular Weight Marker II
À ADN digerido con Hind IlI y DNA Molecular Weight Marker VIII una mezcla de
ADN de pUCBM21 digerido con Hpa II y ADN de pUCBM21 digerido con Dra I y Hind
IlI Boehringer Mannheim en la Tabla 6 se presentan los tamaæos de los fragmentos
respectivos Como control positivo se utilizó el plÆsmido pGEM 3Z
4043
21

Tabla 6 Marcadores de peso molecular utilizados en las electroforØsis de ADN
11 11n o
¸æ ìiI 61111 11111111 1
1 23 1 30
2 94 1 6
3 6557
4 436 1
5 232 2
6 202 7
7 564
tt 1lilitii III I
1 1 1 14
2 900
3 692
4 501
5 489
6 404
7 320
8 242
9 190
1 O 1 47
1 1 1 24
1 2 1 10
13 67
1 4 37
1 5 34
1 6 26
1 7 19
7 3 PCR de las muestras y electroforØsis en gel de agarosa
A cada tubo se le agregaron 50 ng de ADN 2 5 I1L de una solución stock 20
I1g rnL 1 I1L oligonuc1eótidos 1M Tabla 7 40 8 I1L H20 5 I1L Buffer PCR lOx 2 I1L
dNTP s 0 2 I1L Taqpolimerasa Finalmente se adicionaron 30 I1L de aceite mineral
Tabla 7 Secuencias de los oligonucleótidos generados al azar utilizados en el
ensayo de RAPD
llllilliiìI191 TCACGGTGCA
2 GTCGCCGCA
3 GTAGACGAGC
4 AAACGTCGGG
5 ACTGACTGCC
22

Las muestras se pusieron en un PCR Cyclogene de Techne bajo el programa
que se presenta en la tabla 8 Una vez concluido el programa las muestras se
almacenaron a 40C hasta ser analizadas
Tabla 8 Programa de utilizado para RAPDs
lCillpRilli 11011 91919 50 C 5 mm 1 ciclo
930 C 1 rrun
3 6 o C 1 5 rrun 35 ciclos
7 2 o C 3 rrun
72 o C 3 rrun 1 ciclo
40 C 1 O mm 1 ciclo
Las muestras se analizaron en geles de agarosa al 1 De cada muestra se
tomaron 15 flL Y se le agregaron 2 flL de LB La electroforØsis se corrió a 70 V durante
180 mino Como estÆndares se utilizaron los DNA Molecular Weight Marker II y VIII
Boehringer Mannheim Al tØrmino de la electroforØsis el gel de agarosa se sometió a
iluminación U v en un transiluminador y se fotografió para su anÆlisis posterior
7 4 AnÆlisis de los datos
Los datos obtenidos mediante digestión por enzimas de restricción así como por
PCR se analizaron de la siguiente manera a cada una de las fotografias se les determinó
la distancia de cada banda obtenida en relación al frente de aplicación de la muestra Se
obtuvieron los tamaæos de los fragmentos por medio de una regresión lineal calculada
con base en los estÆndares utilizados Con los datos obtenidos se construyó una matriz
de similitud de la que se obtuvo un dendograma El anÆlisis estadístico se realizó
mediante el coeficiente de correlación Pearson l r y el agrupamiento de las cepas se
llevó a cabo por el mØtodo de ligamiento promedio no ponderado UPGMA Todos los
cÆlculos el anÆlisis para el agrupamiento la matriz de similitud y el dendograma se
realizaron por medio del programa Statistica para Windows versión 4 5 de StatSoft en
una computadora Exacto 486
23

V RESULTADOS
l AnÆlisis numØrico de las características morfológicas y fisiológicas
Como se observa en la figura 1 en el dendograma se pueden delinear
claramente tres grupos a partir de un porcentaje de similitud aproximado de 30 De
este modo se puede decir que el 97 44 de las cepas se encuentran integradas dentro
de algœn grupo y 2 cepas 2 56 no se integran El grupo 1 contiene 42 cepas que
pertenecen a 6 gØneros el grupo Il esta formado por 17 cepas pertenecientes a 6
gØneros y una cepa no identificada el grupo III lo integran 10 cepas que pertenecen a
4 gØneros y 6 cepas sin identificar
Grupo 1
42 cepas integran este grupo lo que representa un 53 85 del total de levaduras
estudiadas y se distribuyen de la siguiente forma 21 son del gØnero Debaryomyces 9
pertenecen al gØnero Pichia 6 al gØnero Rhodotorula 3 al gØnero HansenuJa 2 al
gØnero Leucosporidium y 1 al gØnero Saccharomyces La tabla 9 contiene las especies
que conforman el grupo 1
GØnero Debaryomyces
Cabe mencionar que 21 cepas del gØnero Debaryomyces 60 se encuentran
dentro de este grupo y se distribuyen de la siguiente forma 15 cepas de la especie
Deb vanrijii vanrijii 100 4 de las 13 30 77 cepas de la especie Deb Vann ll
yarrowiiy 2 de las 6 33 33 cepas de Deb melissophilus
AdemÆs se debe seæalar que las cepas B12 B13 C12 C22 Deb vanrijii vanrijiJ
presentan un lOO de homología formando así el fenón a Otras cepas de la misma
especie idØnticas son C74 y C75 012 y 013 020 Y 021 Sin embargo se observa que
las 15 cepas de esta especie divergen a partir una similitud del 65 quedando
integradas en un solo grupo que incluye tambiØn las cepas A28 A40c y C62
24

Jm
CIlm
g1mþll
lTIJm
lo
OJ Ul
tt
lm t5
k ıJ JoLlrd
m
u
o
Ulrdu
51o
oUltl
Ulrdu
51o
@o
I00
oO oO
00
I
1 1
I
oCJ
o
oj
o
OE
Ul
Q
OcU
oæ
tO
Q
oOJ
oo
OQ o
Q lo
Zo 1ii l
O
s01r
ct1
6go Q
ªí P
6 Q
ELlUl
O ou
51 JlQ9 tJO ol1
Q
00
Q
51 iio
o2tClo Qo o
6 o
Qo
p oU
tíct1cl
5o
r
oo
oroU
oct1
U rrict1 U olo l oQ o 1
alotætnoct1 l
9 ffial ii
U 6l o QQ lo lo
o oo
o
ii O
III Q o
@6
tn ot1lo
25
3 ð 58 6º 5Gð 8665SÔ R eGG ð ª6 2o 5

Tabla 9 Lista de especies de levaduras marinas que integran el grupo 1 Basado
en sus caracteristicas morfológicas y fisiológicas
1 I t i i i i i il i I tti i i i i u I I i jAl Pichia philogaea C entomaea CIO Deb C kononenkoaeo vannJllyarrOWll o
A2 Leucosporidiwn scott1ˇ o C muscorum Cl2 Deb vannJll vannJll o C pmus
Al8 Rhodotorula graminis C22 Deb vannJll vannJll o e pmus
A18c Rhodotorula rubra C49 Deb vannJll vannJll o e pmus
A23 Rhodotorula grammlS C50 Deb vanrlll vanrl l1 o C pmus
A25 Deb melissophilus o C dendronema C62 Hansenula aJnji o C pmus
A28 Pichia ambrosiae o C dendronema C74 Deb vanrlll vanrlll o C pmus
A29 Deb melissophilus o C dendronema C75 Deb vanrlJ11 vanrlll o C pmus
A31 Pichia haplofila o C nemodendra E52 Hansenula sp
A39 Pichia haplofila o C nemodendra 001 Deb vanriþi yarrowii o C kononenkoae
A40 Pichia haplofila o e nemodendra 002 Rhodotorula rubra
A40C Hansenula aJnji o e pmus 006 Rhodotorula rubra
A41 Pichia haplofila o C nemodendra 011 Deb vanrlll vanrlll o C pmus
A43 Rhodotorula sp 012 Deb vanrlJll vanrll1 o C pmus
A57 Pichia haplofila o C nemodendra 013 Deb vanrlll vanrlJ11 o C pmus
A58 Pichia haplofila o e nemodendra 014 Deb vanriþï yarrowii o C kononenkoae
A59 Pichia haplofila o C nemodendra 018 Deb vanriþï yarrowii o C sphaerica
A75 Deb vanrlll vanrlll o C pmus 020 Deb vanrlll vanrlll o C pmus
B12 Deb vanrlll vanrlll o C pmus 021 Deb vanrll1 vanrlll o C pmus
B13 Deb vanrlll vanrlll o C pmus OC1 Leucospondiwn antarticwn
B17 Deb vanrlll vanrlJll o C pmus 1 Saccharomyces cereVlseae
GØnero Hansenula
Por otro lado 3 de las cepas de Hansenula 60 que se encuentran en la
Colección se integran dentro de este grupo 2 pertenecen a la misma especie
Hansenula alniJ I Y una mÆs cuya especie no ha sido determinada E52
GØnero Flema
El gØnero Piema incluye a 12 cepas dentro de la Colección de las cuales 9 75
se encuentran en este grupo Las 7 cepas 100 de la especie P haplofila se ubican
26

dentro de este grupo de igual forma que una de las 4 cepas 25 de la especie P
philogaea y la œnica cepa 100 de la especie P ambrosiae
Resulta interesante el hecho de que las 7 cepas de la especie P haplofila se
integran en un grupo que diverge a partir del 85 de similitud De estas cepas solo 2
son idØnticas A39 y A40 La grÆfica 1 muestra los porcentajes de participación de cada
gØnero dentro del grupo
I Grupo 1 I50 0
D Debaryomyees 60
liiiI Piehia 75
liiI RhodotoruJa 1002 4 ïI HansenuJa 60
4 8 liiiI Leueosporidium 100
ID Saeeharomyees l00
14 3
GrÆfica l Porcentajes de representación por gØnero dentro del grupo 1 El
anÆlisis se basó en las características morfológicas y fisiológicas En el recuadro
se presenta el porcentaje en referencia al total de cepas de cada gØnero en la
Colección en este grupo
GØnero Rhodotorula
Las 6 cepas de este gØnero se encuentran ubicadas dentro de este grupo
100 y se distribuyen de la siguiente forma 2 cepas de la especie Rh graminis 3
cepas de la especie Rh rubra y una cuya especie no se ha determinado RhodotoruJa
sp
21

En relación a este gØnero se puede observar que estas 6 cepas divergen a partir
de una similitud del 62 Considerando este nivel de similitud tambiØn se incluye en
este grupo la cepa de Saeeharomyees eerevisiae X2180
GØnero Leueosporidium
El 100 de las especies de este gØnero se encuentran dentro del grupo 1 ambas
cepas pertenecen a especies diferentes Leu seotiiy Leu antartieum
Grupo n
Este grupo lo forman un total de 18 cepas lo que representa un 23 08 del total
de las levaduras estudiadas De estas cepas 17 han sido identificadas y solamente una
no C28 Las 17 cepas se distribuyen de la siguiente forma en 6 gØneros 2
Aureobasidium 8 Debaryomyees 1 Hansenula 2 K1uyveromyees 2 Fiehia y 2
Sporobolomyees En la tabla 10 se presenta un listado de las especies que se
encuentran en este grupo
GØnero Debaryomyees
De las 35 cepas del gØnero Debaryomyees 8 se encuentran dentro del grupo n
lo que representa un 22 86 y se distribuyen de la siguiente forma 4 cepas de Deb
melissophilus 66 67 3 cepas de la especie Deb vanrijii yarrowii 23 08 y la œnica
cepa existente 100 de la especie Deb hansenii
GØnero Aureobasidium
Con respecto a este gØnero cabe menCIOnar que las 2 cepas 100 de
Aureobasidium pullulans se encuentran ubicadas en este grupo Ambas cepas A3 y
P15 son idØnticas
GØnero Kluyveromyees
Las 2 cepas de este gØnero en la Colección K marxianus lactatis y K aestuariJ
se encuentran dentro de este grupo 100 Estas dos cepas tienen un 69 de similitud
L28

Tabla lO Lista de especies de levaduras marinas que integran el grupo 11
Basado en sus características morfológicas y fisiológicas
11IItll I IIIII I i llili I II il illl I lil l 1 1 li I I iii I I 11 111111 1 11 1 IIIII li i III III II II I I lll lllllllI11B39 Plchla philogaea o e terebra
C 11 Deb hansenli
A3
AIS
A20
ASO
AS4
ASS
B3 1
B32 1
B38 1
AureobasldlwnpulJuJans
Sporobolomyces holsatlcus
HansenuJa holstl1 CI4 Kluyveromyces marxlanus lactatls
CI9 Kluyveromyces aestuar11
C28 No ldentlflcada
C66 Deb Vann llyarrowll
C67 Deb vannjÙ yarrow11
015
PIS
Deb melIssophilus o e dendronema
Deb vanrlpl yarrowli o epararugosa
Deb mellssophilus o e peltata
Deb meJ1ssophilus oe nemodendra
Plchla philogaea o e peltata
Sporobolomyces holsatlcus
Deb mellssophilus o e nemodendra
AureobasldlwnpulJuJans
GØnero Pichia
De las 12 cepas de este gØnero sólo 2 16 67 se ubican en este grupo
mostrando 100 de similitud
GØnero Sporobolomyces
En la Colección de Levaduras del CIBNOR se cuenta con 6 cepas del gØnero
Sporobolomyces de las cuales 2 se encuentran en este grupo 33 33 Estas cepas
que presentan un 41 de similitud son la A15 y la B38 l ambas identificadas como Sp
holsaticus
GØnero Hansenula
La œnica cepa de la especie Hansenula holstii se encuentra ubicada en este grupo
representando un 20 de las cepas de este gØnero que forman parte de la Colección
La grÆfica 2 muestra los porcentajes departicipación de cada gØnero dentro del grupo
29

I Grupo II I
Aureobasidium 100
DDebaryomyces 22 86
Hansenula 20
liIK1uyveromyces 1005 6 IiïPichia 1667
DSporobolomyces 3333
No Identificadas 14 3
111
GrÆfica 2 Porcentajes de representación por gØnero dentro del grupo n El
anÆlisis se basó en las caracteristicas morfológicas y fisiológicas En el recuadrose presenta el porcentaje en referencia al total de cepas de cada gØnero en la
Colección en este grupo
Grupo III
Este grupo esta formado por 10 cepas que se dividen en 4 gØneros y 6 cepas no
identificadas 37 5 De las 10 cepas identificadas 5 pertenecen al gØnero
DebØllYomyees 3 al gØnero Sporobolomyees 1 al gØnero Hansenula y 1 al gØnero
Piehia En la tabla 11 se presenta un listado de las especies que conforman este grupo
Como se puede observar en la figura 1 las cepas que pertenecen al gØnero
DebØllYomyees se identificaron como la misma especie Deb vanrijii yarrowii las 3
cepas del gØnero Sporobolomyees pertenecen a la especie Sp punieeus mientras que
la œnica cepa del gØnero Hansenula a la especie H aJnji Y la de Plehia a la especie P
philogaea
La grÆfica 3 muestra los porcentajes de participación de cada gØnero dentro del
grupo
30

Tabla 11 Lista de especies de levaduras marinas que integran el grupo III
B fl rasado en sus caractensticas morfologlcas y lS10 Oqlcas
talB II j j j I RíBiI j j j lqI1 j NßiBgj j j j j j A7
Deb vann l1yarrowl1oe kononenkoae B20 1Sporobolomyces puniceusA12
Deb vann l1yarrOWl1oe kononenkoae B27 2Pichia philogaeao e nemodendra Al
7Deb vann l1yarrOWl1oe kononenkoae C52 1 No identificada A24
Deb vann l1yarrowl1oe kononenkoae CSS No identificada A27
Deb vann l1yarrowl1oe kononenkoae CS8 1No identificada A30
Hansenula alniioe vanderwaltii C64 No identificada A79
Sporobolomyces puniceusC7 1No identificada B20
Sporobolomyces puniceusC78 No identificada GØnero
Debaryomyces De
las 35 cepas de este gØnero 5 se encuentran eneste grupo 14 29 Estas 5cepas
forman parte de las 13 que se identificaron como la especie Deb vanrijii yarrowii
lo que correspondea un 38 46 Estas
cepas divergena partir de un 75 5 de similitudy de ellas 3 son idØnticas A7
A12 Y A17 las cepasA24yA27 muestran unporcentajede similitud de92 GØnero
Sporobolomyces3
de las 6 cepas existentes en la Colección 50 de este gØnero se encuentran ubicadas
eneste grupo Estas 3cepas pertenecenala misma especie Sporobolomyces pw1iceus
De ellas solamente2presentan un 100 de similitud B20 y B20 1 GØnero
Hansenula De
las 5 cepas del gØnero HansenuJa1se integra en este grupo 20 Y es una de
las 3 cepas dela especie HansenuJa a1niiEsta cepa presenta una similitud con las cepas
deDebaryomycesdeun 70 31

GØnero Pichia
Este gØnero incluye a 12 cepas de las cuales 1 forma parte de este grupo
8 33 Esta cepa es una de las 3 que pertenecen a la especie Pichia philogaea y su
similitud con las cepas de Sporobolomyces es de un 40
GØnero No Identificado
De las 7 cepas no identificadas 6 se integran en este grupo 85 71 de entre
ellas las cepas C55 y C58 1 son idØnticas
I Grupo III I45 5
o Debaryomyces 14 29
Sporobo1omyces 50
lIiI Hansenu1a 20
Pichia 833
liiI NoIdentificadas 85 71
9 1
9 1
GrÆfica 3 Porcentajes de representación por gØnero dentro del grupo III El
anÆlisis se basó en las caracteristicas morfológicas y fisiológicas En el recuadro
se presenta el porcentaje del total de cepas de cada gØnero en la Colección en
este grupo
2 Contenido de proteínas totales
En la tabla 12 se presentan los datos de la biomasa obtenida de cada cepa a las
36 horas de cultivo y el contenido de proteína de los extractos
32

Tabla 12 Biomasa obtenida gr a partir de lli1 cultivo de 50 rnl de medio MI
despuØs de 36 horas de incubación a temperatura ambiente contenido de
proteínas lJg rnl de los extractos de cada cepa 1 Saccharomyces cerevisiae 2
Schizosaccharomyces pombe
A57 1 1 1463 1
A58 1 5418
Al 0 9 799 7
A2 1 2 1 63 7
A3 0 9 2 3434
A7 0 9 445 5
A12 0 9 288 5
A15 1 2 906 5
A17 0 9 275 1
A18 1 2 782
A18c 0 9 3 025 9
A20 0 9 474 5
A23 0 9 2 500 8
A24 1 186 5
A25 1 1 168 2
A27 1 250 6
A28 1 358
A29 0 9 399
A30 0 9 379 6
A31 0 9 652 1
A39 1 1 562 3
A40 0 9 1 678 2
A40c 1 954
A41 1 796 8
A43 0 9 1 975 6
A50 1 482 1
A54 1 7 18 4
A55 1 144 5
A59
A75
A79
B3 1
B12
B13
B17
B20
B20 1
B27 2
B32 1
B33
B38
B39
ClO
CIl
C12
C14
C19
C22
C28
C38 1
C49
C50
1
1 1
1
1
0 9
1
1
0 9
1 1
1
1 1
1 1
1 1
1 1
1 1
1 123 6
851 9
983
2519
738 9
961
1 322 6
264 1
1 650 3
1 337
861 5
332 9
1 903 9
980 8
371 7
453 5
1 550 6
389 7
606 7
1 746 6
3 828 6
4 2419
1 2214
560 6
191 1 1 111 1111 llil I II tI˛ lllill I III I 11 11111111111
C62 1 770 1
C64 1 1 551
C66 1 1 665 8
C67 1 383 5
C7 1 1 3 198 5
C74 1 672 4
C75 1 1 563 5
C78 1 2 663 6
E52 1 2 292 1
001 1 549 6
002 1 1 1 979 9
006 1 2493 9
01 1 1 2 464 2
012 1 874 9
O 1 3 1 1 825
014 1 1 380 3
O 15 1 2 1 1 6
O 1 7 0 9 2 248 1
O 18 1 3234
020 1 7524
021 1 954 2
OCl 1 1 2 563 9
Pl 5 1 1 877 6
1 1 5 2 698 8
2 1 2 9034
33

3 Características generales de los patrones de proteínas totales
A partir de los patrones electroforØticos en geles de poliacrilamida Figura 2 se
encontraron 129 bandas diferentes de proteínas con pesos moleculares que oscilan
entre los 467 7 kD Y los 12 6 kD La banda de 17 4 kD es compartida por 33 cepas
diferentes 43 4
1 2 3 4 6 7 kD
45 0
310
21 5
14 4
Figura 2 Patrones electroforØticos de 7 cepas Solo se incluye una colonia tipopor cepa Carrill cepa A3l carril 2 cepa A39 carril 3 cepa A40 carril 4 cepa
A4l carril 5 cepaA57 carril 6 cepaA58 carril 7 cepaA59
4 AnÆlisis numØrico de los patrones electroforØticos de proteínas solubles totales
Como se puede observar en la figura 3 las 77 cepas que se estudiaron forman 5
grupos claramente definidos a partir de una sirrœlitud del 22 El grupo I contiene 28
cepas que pertenecen a 6 gØneros y una cepa no identificada el grupo II contiene 16
cepas identificadas dentro de 7 gØneros y 4 cepas no identificadas el grupo III esta
formado por 17 cepas en su totalidad del gØnero Debaryomyces el grupo N lo
componen 8 cepas pertenecientes a 3 gØneros y finalmente el grupo V esta formado
œnicamente por 2 cepas del gØnero Debaryomyces Cabe mencionar que la cepa C62
no forma parte de ninguno de los grupos anteriores
34

tim
8Hm
timH
fjm 5
ElCl 2l 0
QCl d
d tio
ti Slrd j
H
Clu 2tl u
tl JlQ
m tiH Qm
gm
ro
Sm
odfCl
Q
1
I I
lS I1
n1
1
i R i i aDO I C y a R aD aIRo r miofj 18 1Zl e o lJooB J C ín u w n j o o o u 2 ı ı ü
V o
o
len
en
lC
CC
D
BOf
en
DcU
@en
aOJen
enen
fC
COJ
Oo
o
o Odd
8O o
6 u
wmrri tO o
al u
Õ Od
tilO O
o U
Ulitil Ord o
S u
2IDo
od
d N
tilao
Sl
200
2ID
otildUl
O rd oH 8o O
H d toªí Olo OUlUl oso
o sm
@oii5J Ul
d d0O åJUl 6 H
ooo H OUoH
alOUlo
o
d ffird 65J o
tJH
O Ul
tJ o
rd ID 5JO 62 g
o o til
d iio
Ol
O
C ol
gt rd@
O Otr dP
35

Grupo 1
Las 29 cepas que constituyen este grupo representan el 37 66 del total de
levaduras estudiadas Las 28 cepas identificadas se distribuyen de la siguiente forma 12
pertenecen al gØnero Piema 7 al gØnero Debaryomyees 3 al gØnero Sporobolomyees
3 al gØnero RhodotoruJa 2 al gØnero Kluyveromyees 1 al gØnero Hansenula y una
restante no se ha identificado En la tabla 13 se presentan las especies que conforman el
grupo 1 La grÆfica 4 presenta los porcentajes de participación de cada gØnero dentro
de este grupo
Como se puede observar dentro del grupo existen 2 fenones con representantes
que poseen características idØnticas en cuanto a su patrón electroforØtico de proteínas
totales un fenón formado por las cepas A31 A39 A40 A41 A57 A58 y A59 Piema
haplofila o Candida nemodendra y otro formado por las cepas C50 C74 C75
011 012 013 Y 020 Debaryomyees vanrijii vanrijii o Candidapinus
Tabla 13 Lista de especies de levaduras mannas que integran el grupo 1B d 1 tr 1 t ti d 1001 alasa o en e pa on e eetro ore eo e protemas so es tot es
111 111 1 11 1 1 11111 1 1 1 11 1 1 1 11 111 1 11 11 111111 11 11 1 11 1 1 11111 1 111111111 11 1 111 1 111 11111 11 1 1 1 111 11 1 1 1 1 1 111 1 11 111 11 1 111 11 11 1 1 1 1111 111 11 1 1 1IIII II IIIt r j j j
Al Pichia phiJogaea o e entomaea B38 1 Sporobolomyces holsaticus
A28 Pichia ambrosiae o e dendronema B39 Pichia phiJogaea o e terebra
A30 Hansenula alnii o e vanderwaltii C14 K1uyveromyces marxianus lactatis
A31 Pichia haplofiJa o e nemodendra C19 K1uyveromyces aestuarii
A39 Pichla haplofiJa o e nemodendra C38 1 No identificada
A40 Pichia haplofiJa o e nemodendra CSO Deb vannJll vannJll o e pmus
A41 Pichia haplofiJa o e nemodendra C74 Deb vannJll vannJll o e pmus
A43 Rhodotomla sp C7S Deb vannJll vannJll o e pmus
AS7 Pichia haplofiJa o e nemodendra 002 Rhodotomla rubra
AS8 Pichia haplofiJa o e nemodendra 006 Rhodotorula rubra
AS9 Pichia haplofiJa o e nemodendra 011 Deb vannJll vannJll o e pmus
B20 Sporobolomycespuniceus 012 Deb vannJll vanr1Jll oepmus
B20 1 Sporobolomycespzmiceus 013 Deb vanr1Jll vanr1Jll o e pmus
B27 2 Pichia phiJogaea o e nemodendra 020 Deb vanr1Jll vanr1Jll oe pmus
B32 1 Pichia phiJogaea o e peltata
36
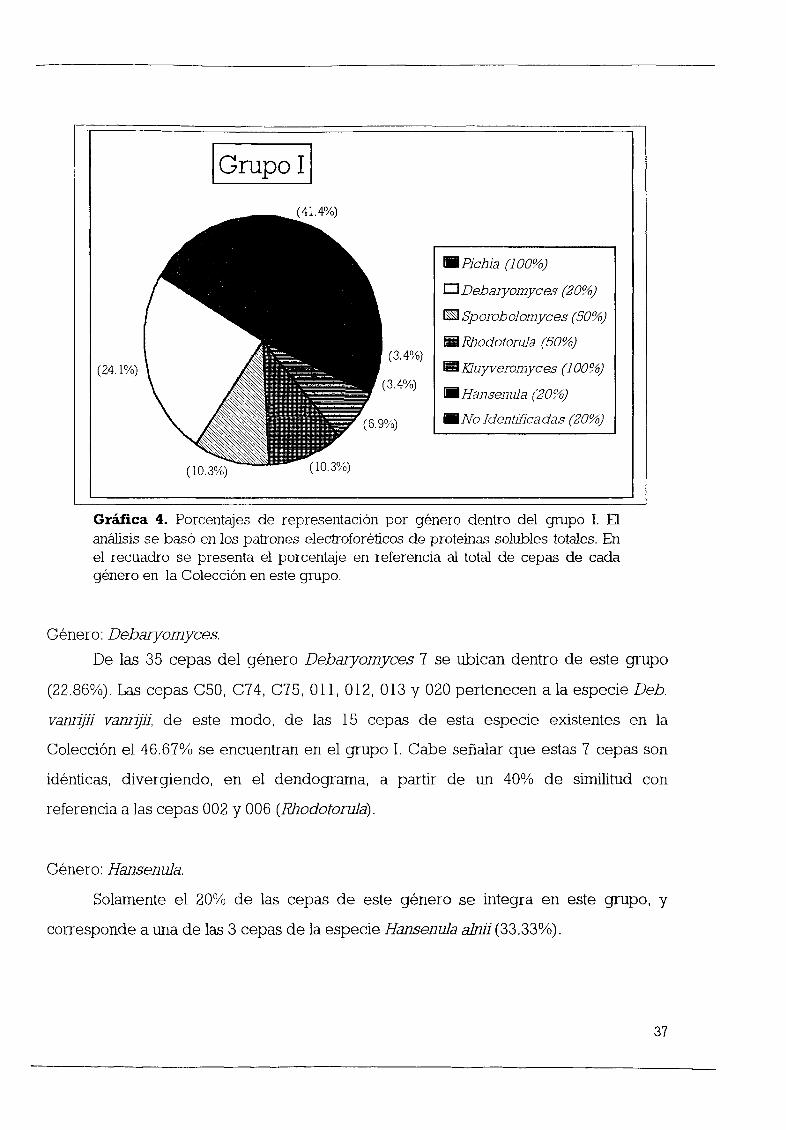
I Grupo 11
24 1
3 4
IIiiIPichia 100
oDebaryomyces 20
Sporobolomyces 50
iïiRhodotorula 50
IiiKluyveromyces 100
IIiiIHansenula 20
IiiINo Identificadas 20
GrÆfica 4 Porcentajes de representación por gØnero dentro del grupo 1 El
anÆlisis se basó en los patrones electroforØticos de proteínas solubles totales Enel recuadro se presenta el porcentaje en referencia al total de cepas de cada
gØnero en la Colección en este grupo
GØnero DebaryomycesDe las 35 cepas del gØnero Debaryomyces 7 se ubican dentro de este grupo
22 86 Las cepas C50 C74 C75 011 012 013 Y 020 pertenecen a la especie Deb
vamijil vanrijil de este modo de las 15 cepas de esta especie existentes en la
Colección el 46 67 se encuentran en el grupo 1 Cabe seæalar que estas 7 cepas son
idØnticas divergiendo en el dendograma a partir de un 40 de similitud con
referencia a las cepas 002 y 006 RhodotoruJa
GØnero HansenuJa
Solamente el 20 de las cepas de este gØnero se integra en este grupo y
corresponde a una de las 3 cepas de la especie HansenuJa alnii 33 33
37

GØnero Kluyveromyees
El 100 de las cepas de este gØnero 2 se encuentran en este grupo la cepa
e 14 se identificó como Kluyveromyees marxianus lactatis mientras que la cepa e 19 se
identificó como Kluyveromyees aestuarii mostrando un 45 de similitud
GØnero Piema
El grupo 1 engloba el 100 de las cepas de este gØnero Las 12 cepas se dividen
en tres especies diferentes 7 pertenecen a la especie P haplofjJa 4 a la especie P
philogaea y una mÆs a la especie P ambrosiae Las 7 cepas de la especie P haploæla
mostraron patrones electroforØticos idØnticos En relación a las cepas de la especie P
philogaea 3 divergen a partir de un 51 de similitud Al B32 1 Y B39 mientras que
las cepas B32 1 y B39 son idØnticas La cepa restante de esta especie es segœn el
anÆlisis mÆs parecida a las cepas B20 y B20 1 del gØnero Sporobolomyees Finalmente
la cepa A28 P ambrosiae presenta un 35 de similitud con la cepa A43 RhodotoruJa
sp
GØnero RhodotoruJa
El 50 de las cepas del gØnero Rhodotorula estÆn representadas en este grupo
las cepas 002 y 006 RhodotoruJa rubra comparten un 47 de similitud mientras que la
cepa A43 muestraun 35 de similitud con la cepa A28 Piema ambrosiae
GØnero Sporobolomyees
2 cepas de la especie Sp punieeus 66 7 componen este grupo ambas son
idØnticas por otro lado la cepa B32 1 Sp holsatieus es idØntica a la cepa B39
identificada como Piemaphilogaea
Grupo n
20 cepas integran este grupo lo que corresponde a un 26 del total de cepas
estudiadas De las 20 cepas 4 no han sido identificadas mientras que las 16 restantes se
distribuyen de la siguiente forma 2 pertenecen al gØnero Aureobasidium 3 al gØnero
38
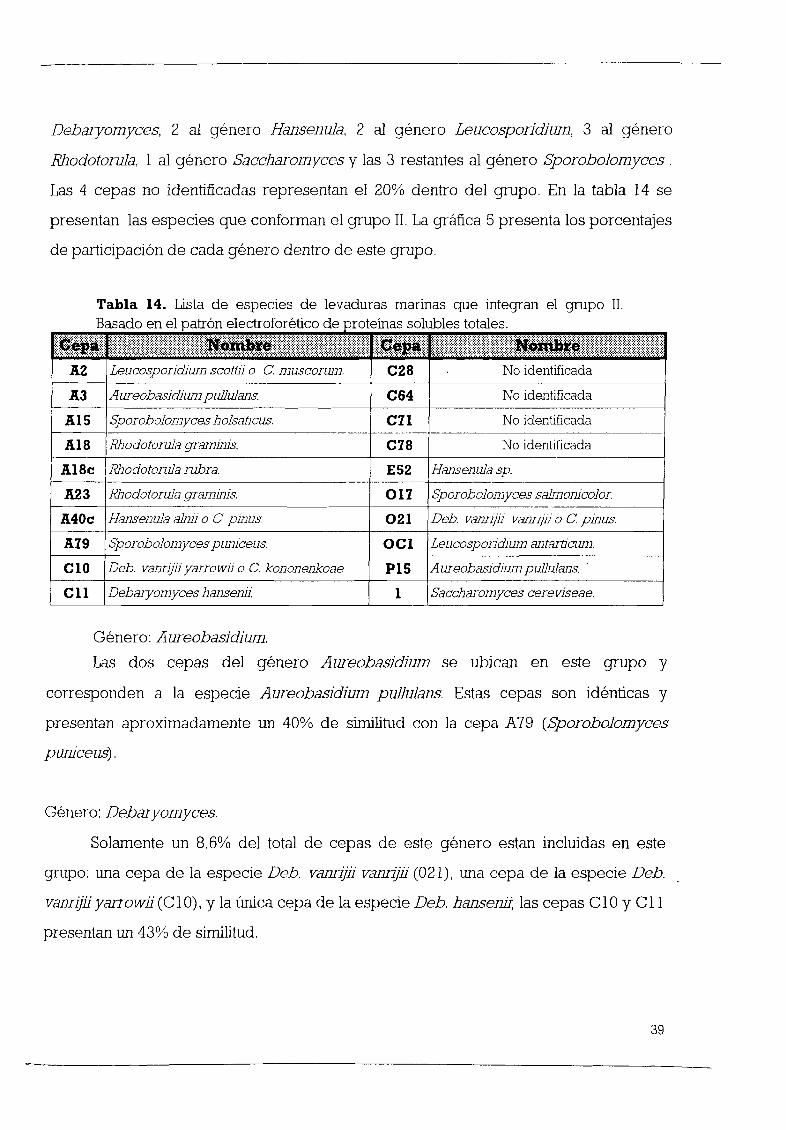
Debaryomyces 2 al gØnero Hansenula 2 al gØnero Leucosporid˝lzm 3 al gØnero
RhodotoruJa 1 al gØnero Saccharomyces y las 3 restantes al gØnero Sporobolomyces
Las 4 cepas no identificadas representan el 20 dentro del grupo En la tabla 14 se
presentan las especies que conforman el grupo n La grÆfica 5 presenta los porcentajes
de participación de cada gØnero dentro de este grupo
Tabla 14 Lista de especies de levaduras marinas que integran el grupo n
Basado en el patrón electroforØtico de proteínas solubles totales
jllllj i j j j j j j j j j j j j j j j j j j jli iljj j j j j j j j j j j jj ji j j j j j IIIIj j j j j j j j j j j j j j jjIIDiijj j j j i j j j j j i i j j j i i i j j jA2 Leucosporidium scottJi o e muscorum C28 No identificada
A3 AureobasidiumpuDulans C64 No identificada
AIS Sporobolomyces holsaticus cn No identificada
AI8 Rhodotorula grammlS C78 No identificada
AI8c Rhodotorula mbra ES2 Hansenula sp
A23 Rhodotomla grammlS 017 Sporobolomyces salmonicolor
A40c Hansenula alnJi o e pmus 021 Deb VanIlJll VanIlJll o e pmus
A79 Sporobolomycespuniceus OCI Leucospondium antarticum
CIO Deb VanIlJll yarrowll o e kononenkoae PIS AureobasidiumpuDulans
CI I Debaryomyces hansenii I Saccharomyces cereVlseae
GØnero Aureobasidiwn
Las dos cepas del gØnero Aureobasidiwn se ubican en este grupo y
corresponden a la especie Aureobasidiwn puUulans Estas cepas son idØnticas y
presentan aproximadamente un 40 de similitud con la cepa A79 Sporobolomyces
puniceus
GØnero Debaryomyces
Solamente un 8 6 del total de cepas de este gØnero estan incluidas en este
grupo una cepa de la especie Deb vanrijii vanrijii 021 una cepa de la especie Deb
vanrijii yarrowii CIO y la œnica cepa de la especie Deb hansenii las cepas CIO y CIl
presentan un 43 de similitud
39

I Grupo II I
15 8
Aureobasidiwn l00
DDebaryomyces 857
10 5 fíijHansenula 40
liiI Leucosporidiwn l00
Rhodotorula 50
líiiiiiI Saccharomyces 100
Sporobolomyces 50
IïiiiINo Identificadas 80
10 5
GrÆfica 5 Porcentajes de representación por gØnero dentro del grupo 11 El
anÆlisis se basó en los patrones electroforØticos de proteínas solubles totales Enel recuadro se presenta el porcentaje en referencia al total de cepas de cada
gØnero en la Colección en este grupo
GØnero Hansenula
El 40 de las cepas de este gØnero que existen en la Colección deLevaduras se
integran en este grupo una corresponde a la especie Hansenula aJnjj y la otra cepa
cuya especie no se ha identificado E52 ambas comparten un 27 de similitud
GØnero Leucosporidiwn
Las dos cepas del gØnero Leucosporidium forman parte de este grupo
presentando un 30 de similitud Es necesario mencionar que la cepa OC 1 presenta una
similitud del 50 7 con las cepas del gØnero RhodotoruJa aparentemente esta cepa
resulta ser mas similar a cepas del gØnero Rhodotorula que a otras de su mismo gØnero
Lo mísmo ocurre con la cepa A2 que presenta una similitud del 43 9 con la cepa 021
Deb vanrijiii vanriji1
40

GØnero RhodotoruJa
El 50 de las cepas de este gØnero se situan dentro de este grupo las cepas A18
y A18c identificadas como Rh graminis y como Rh rubra respectivamente son
idØnticas mientras que la cepa A23 presenta una distancia de similitud de 68 3 en
relación a ellas
GØnero Saccharomyces
La œnica cepa de este gØnero se incluye en este grupo Esta cepa es muestra una
similitud de 48 8 con la cepa E52 HansenuJa sp
GØnero Sporobolomyces
El 50 de las cepas de este gØnero se integran en este grupo las cepas A15 y
017 presentan un 50 de similitud mientras que respecto a la cepa A79 un 32 7
Cepas no identificadas
De las 5 cepas no identificadas que se estudiaron 4 se integran en este grupo
80 Todas divergen a partir de un 30 de semejanza
Grupo IlI
Este grupo esta integrado exclusivamente por cepas del gØnero Debaryomyces
lo que corresponde al 22 de las levaduras estudiadas En la tabla 15 se presentan las
especies que conforman el grupo IlI
GØnero Debaryomyces
El 48 6 de las cepas de este gØnero forman el grupo IlI En relación al nœmero
de cepas por especie 10 cepas identificadas como Deb vanrijii yarrowii se integran en
este grupo asi como las 15 cepas identificadas como Deb vanrijii vanrijii
De la especie Deb vanrijii yarrowii 9 cepas se integran en un fenón mostrando
100 de similitud mientras que la cepa B33 comparte un 38 de similitud con las cepas
de Deb vanrijii vanrijii Por otro lado las 7 cepas de la especie Deb vanrijii vanrijii
41
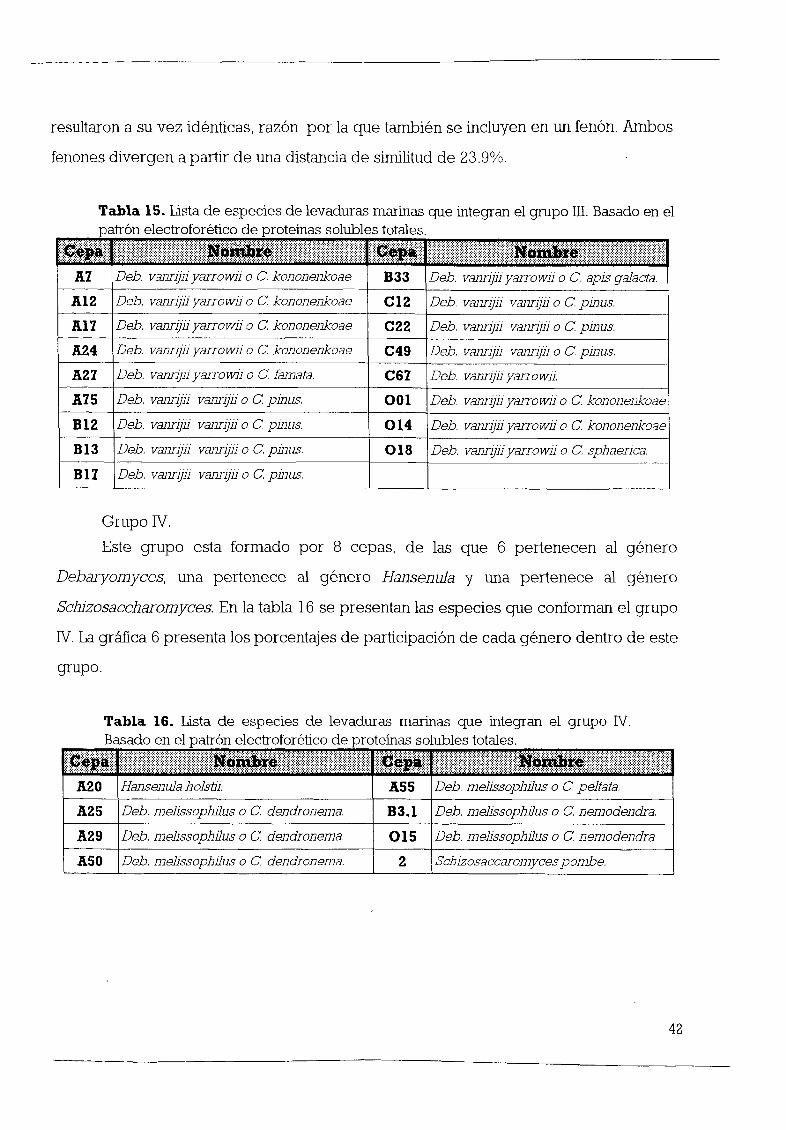
resultaron a su vez idØnticas razón por la que tambiØn se incluyen en un fenón Ambos
fenones divergen a partir de una distancia de similitud de 23 9
Tabla 15 Lista de especies de levaduras marinas que integran el grupo 1lI Basado en el
patrón electroforØtico de proteínas solubles totales
B II li 11 II ftA7 Deb vanrlJllyarrowll o e kononenkoae B33 Deb vanrlJllyarrowll o e aplS galacta
Al2 Deb vanrll1 yarrowll o e kononenkoae Cl2 Deb vanrl l1 vanrl l1 o epmus
A17 Deb vanrll1 yarrOWl1 oe kononenkoae C22 Deb vanrll1 vanrll1 o e pmus
A24 Deb vanrlJllyarrowll o e kononenkoae C49 Deb vannJ11 vanrlJ11 o e pmus
A27 Deb vanrll1 yanowll o e famata C67 Deb vanrll1 yanOWll
A75 Deb vanrlJ11 vanrll1 o epmus 001 Deb vanrll1 yarrowll o e kononenkoae
B12 Deb vanrlJll vanrlJ11 o e pmus 014 Deb vanrlJ11 yarrowll o e kononenkoae
B13 Deb vanrl l1 vanrll1 o e pmus 018 Deb vanrll1 yarroWl1 o e sphaerica
B17 Deb vanrll1 vanrll1 o e pmus
Grupo N
Este grupo esta formado por 8 cepas de las que 6 pertenecen al gØnero
Debaryomyces una pertenece al gØnero Hansenula y una pertenece al gØnero
Schizosaccharomyces En la tabla 16 se presentan las especies que conforman el grupo
N La grÆfica 6 presenta los porcentajes de participación de cada gØnero dentro de este
grupo
Tabla 16 Lista de especies de levaduras marinas que integran el grupo N
Basado en el patrón electroforØtico de proteínas solubles totales
@11I I˛ ii 1111 j lillii jA20 Hansenula holstii A55 Deb melissophilus o e peltata
A25 Deb melissophilus o e dendronema B3 1 Deb melissophilus o e nemodendra
A29 Deb melissophilus o e dendronema o 15 Deb melissophilus o e nemodendra
ASO Deb melissophilus o e dendronema 2 Schizosaccaromyces pombe
42

GØnero DebaryomycesLas 6 cepas estudiadas de este gØnero que forman parte de este grupo son
idØnticas y se identificaron como la misma especie Deb melissophilus Las cepas A25 y
A29 comparten un 48 de similitud con la cepa 015
I Grupo mi
oDebaryomyces 1714
liiiiI Hansenu1a 20
liI Scizosaccharomyces 100
12 5
GrÆfica 6 GrÆfica de los porcentajes de representación por gØnero dentro del
grupo IV El anÆlisis se basó en lospatrones electroforØticos de proteínas solubles
totales En el recuadro se presenta el porcentaje en referencia al total de cepas de
cada gØnero en la Colección en este grupo
GØnero HansenuJa
Una de las 5 cepas del gØnero HansenuJa 20 esta integrada en el grupo IV
esta cepa se identificó como HansenuJa holstii y es la œnica de esta especie en la
Colección Esta cepa presenta una similitud del 35 con la cepa B3 l Deb
melissophilus De igual forma la œnica cepa del gØnero Schizosaccharomyces se
incluye en este grupo y mostró un 37 4 de similitud con la cepa ASO Deb
melissophilus
43

Grupo V
Este grupo solamente esta compuesto œnicamente por dos cepas A54 y C66
que se identificaron como Debaryomyces vanrijÙ yarrowii ambas cepas divergen del
resto a partir del 17 de similitud sin embargo sonsimilares entre sí en un 98
5 AnÆlisis de las cepas A18 A18c A23 A43 002 006 A75 B12 B13 B17 C12 C22 y
C49
5 1 Con base en las pruebas morfológicas y fisiológicas
De acuerdo a los resultados obtenidos de las pruebas morfológicas y fisiológicas
Figura 1 y a los patrones electroforØticos de proteínas solubles Figura 3 se
seleccionaron algunas de las cepas de Debmyomyces vanrijÙ vanrijÙ y del gØnero
Rhodotorula debido que en los dendogramas derivados de los diferentes anÆlisis se
obtiene un agrupamiento muy similar y persistente lo cual es un punto de partida ideal
para determinar la sensibilidad de las tØcnicas que consideran el uso de ADNcomo son
la amplificación de secuencias al azar RAPD y los anÆlisis de restricción mediante el
uso de endonucleasas
Considerando el anÆlisis numØrico de las pruebas taxonómicas convencionales
se observa en la figura 4 que el índice de similitud de algunas cepas de Debaryomyces
vanrijÙ vanrijii es del 100 mientras que las cepas del gØnero Rhodotorula mostraron
mÆs heterogeneidad similitud 90 Claramente se observa la formación de 2
grupos uno incluye al gØnero Rhodotorula y el otro a las cepas de Debaryomyces
vamijÙ vanrijii ambos grupos divergen a partir de un 35 de similitud
Como se puede observar las cepas del gØnero Rhodotorula presentan una
divergencia entre ellas que se origina a partir de un 57 Sin embargo las cepas 002 y
006 ambas de la especie Rhodotorula rubra son las que se encuentran mÆs
distanciadas del grupo con un 76 en cuanto a las cepas A18 y A18c son idØnticas
considerando sus características morfológicas y fisiológicas
44

Las cepas del gØnero Debæyomycesdivergen a partir de un 80 de similitud 4
de ellas son idØnticas B12 B13 C12 y C22 mientras que la cepa B17 muestra un 94
de similitud Las cepas A75 Y C49 muestran un 86 de similitud
A1SA1ScA43A23002006A75C49812813C12C22817
100 ffJ 00 40 20
Figura 4 Dendograma generado de las cepas A18 A18c A23 A43 002 006
A75 B12 B13 B17 C12 C22 y C49 obtenido en base a las características
morfológicas y fisiológicas de las cepas El mØtodo de anÆlisis fue elligamientopromedio no ponderado upGMA utilizando el coeficiente de correlación de
Pearson l r
5 2 Con base en los patrones electroforØticos de proteínas
La figura 5 muestra el dendograma generado al analizar los datos obtenidosa
partir de los patrones electroforØticos de proteínas totales de las cepas A18 A18c A23
A43 002 006 A75 B12 B13 B17 C12 C22 y C49 mediante taxonomía numØrica
Como se puede observar claramente se delinean dos grupos lUlO que incluye a
las cepas del gØnero RhodotoruJa y otro de las cepas del gØnero Debaryomycesambos grupos divergen a partir de un 10 71 de similitud
Las cepas del gØnero RhodotoruJa se separan a partir del 25 71 de similitud la
cepa A43 es la que esta separada de las demÆs por esta misma distancia Las cepas A18
y A18c que son idØnticas comparten con la cepa A23 el 67 14 de similitud Mientras
que las cepas 002 y 006 muestran un índice de similitud de 45 71
En lo referente a las cepas del gØnero Debæyomycesresulta muy interesante el
hecho de que mediante esta tØcnica todas queden incluidas en un fenón mostrando ser
idØnticas
45

A1eA1SoA23002006A43A75B12B13B17C12C22C49
too 80 60 40 20
Figura 5 Dendograma generado de las cepas Al8 Al8c A23 A43 002 006A75 Bl2 Bl3 Bl7 Cl2 C22 y C49 obtenido en base a sus patroneselectroforØticos de proteínas solubles totales El mØtodo de anÆlisis fue el
ligarniento promedio no ponderado upGMA utilizando el coeficiente de
correlacion de Pearson l r
46

6 Concentración de Æcidos nuc1eicos
En la tabla 17 se presentan los datos de ADN obtenido a partir de clÙtivos de las
13 cepas de levaduras analizadas despues de 24 horas de incubación En la tabla se
incluye tambiØn la absorbancìa a 260 nmy la razón de la absorbancìa entre las lecturas
a 260 y 280 nm el cual se utiliza para indicar el grado de pureza del ADN obtenido a
280 mn se identifican las proteínas En la figura 6 se muestra la fotografí a obtenida del
gel con el ADN purificado
Tabla 17 Absorbancia a 260 nro relación de las absorbancias de 260 y 280 nro
y cantidad de ADN obtenida de las 13 cepas que se estudiaron Los resultados
que se presentan se obtuvieron apartir de una dilución 1 500
iillilllllliiAl 8 0 2 1 9 10 1
Al 8c O 4 2 1 8
A23 0 4 2 1 7
A43 0 3 2 1 4 7
002 0 2 1 9 8 5
006 0 4 2 1 7 1
A75 O 3 2 1 5 4
B 1 2 O 5 2 20 1
B 1 3 0 4 2 1 5 5
B 1 7 O 3 2 1 4 5
C 1 2 O 5 2 2 1 6
C22 0 4 2 1 7 1
C49 O 2 2 1 0 7
47

Rhodotom1a D—baryomyces
1 2 3 4 5 6 7 8 9 1112 13
Figura 6 Gel de agarosa al 0 8 en el que se observan las bandas de ADN
genómico la IJg purificado de las muestras Carriles 1 cepa A18 2 cepa A18c
3 cepa A23 4 cepa A43 5 cepa A75 6 cepa B12 7 cepa Bl3 8 cepa B17 9
cepa C12 la cepa C22 11 cepa C49 12 cepa 002 13 cepa 006
7 Características generales de patrones de restricción
Como ya se mencionó con anterioridad para realizar este anÆlisis se utilizaron 6
enzimas de restricción Alu 1 Bam HI Eco RI HinfI Kpn 1 y Pst I de las cuales Alu 1 y
Kpn 1 no generaron patrones electroforØticos bien definidos sin embargo se observa
una mancha que abarca la línea de corrimiento de la muestra que no permite que se
observe un patrón mÆs detallado Esto se puede explicar si consideramos que la
enzima Alu 1 reconoce la secuencia 5 AG CT 3 La probabilidad de encontrar en al
ADN genómico esta secuencia es muy alta la enzima reconoce estas secuencias y las
corta generando así un patrón de restricción con tan alto nœmero de bandas que se
observa un barrido El caso de la enzima Kpn 1 no es diferente esta enzima reconoce
la secuencia 5 GGTAC C 3 6 pares de bases Probablemente esta secuencia es muy
abundante en el genóma de estas levaduras por lo que de igual forma que Alu 1 se
genera un barrido mÆs que un patrón bien definido
Por otro lado el resto de las enzimas presentaron las siguientes características
con la enzima HinfI fue con la que se obtuvo un mayor nœmero de bandas de diferente
48

tamaæo 23 bandas y con la enzima Eco RI se obtuvieron solamente 11 El patrón
electroforØtico mÆs claro que se obtuvo fue el que se generó con las digestiones
utilizando la enzima Pst I aunque cabe aquí mencionar que en todos los geles se
observó un barrido que no permitió una definición muy clara
8 AnÆlisis numØrico de los patrones electroforØticos deADN
Bam HI
La digestión del ADN con esta enzima produce 14 bandas que oscilan entre los
4 9 Kb Y los 12 Kb En la figura 7 se puede observar el dendograma que se generó
utilizando los datos referentes a la enzima de restricción Bam HI en el se distinguen
claramente dos grupos uno que hace referencia a las cepas del gØnero Rhodotorula y
el otro que reœne a las cepas del gØnero Debaryomyces ambos grupos divergen a
partir del 8 72 de similitud
A1BA43A18c
A23002
008A75812
817C12C22C49
813
100 ro 60 40 20 o
Distancia deSimilitud
Figura 7 Dendograma generado a partir de la digestión de ADN de las cepasA18 A18c A23 A43 002 006 A75 B12 B13 B17 Cl2 y C22 por la enzima de
restricción Bam HI El mØtodo de anÆlisis fue el ligarniento promedio no
ponderado upGMA utilizando el coeficiente de correlación de Pearson l r
GØnero Rhodotorula
Las 6 cepas de este gØnero divergen a partir del 18 97 de similitud
produciØndose 2 fenones uno que contiene a las cepas A18 A43 A18c Y A23 y otro
49

que integra las cepas 002 y 006 En el primer fenón se observa que las 4 cepas
divergen a partir de una similitud de 56 92 Dentro de este fenón las cepas A18c y A23
son idØnticas mientras que las cepas A43 y A18 presentan una similitud del 78 46 En
cuanto al otro fenón ambas cepas 002 y 006 son idØnticas
GØnero Debaryomyces
Este grupo presenta una mayor homología que el anterior ya que las cepas que
lo integran divergen a partir de una distancia de similitud del 46 15 En este gØnero
tambiØn se observan dos fenones el primero que incluye a las cepas A75 B12 B17
C12 C22 Y C49 y el segundo en el que se encuentra solamente a la cepa B13 El
primer fenón diverge a partir de una distancia de similitud del 64 62 de este fenón las
cepas A75 y B12 presentan una similitud de 90 26 mientras que las cepas B17 C12
C22 y c49 son idØnticas
EcoRI
La digestión del ADN con la enzima de restricción Eco RI produjo 11 bandas que
oscilan entre 10 9 y 2 2 Kb En la figura 8 se presenta el dendograma generado a partir
de los datos referentes a esta enzima En el dendograma se distinguen dos grupos uno
que hace referencia a las cepas del gØnero Debaryomyces pero que incluye a la cepa
A18 Rh gramims y el otro que reœne a las 5 cepas restantes del gØnero RhodotoruJa
ambos grupos divergen a partir del 15 9 de similitud
GØnero 17ebaryomyces
En este grupo se observan dos fenones el primero que incluye exclusivamente a
la cepa A18 y otro fenón en el que las cepas A75 B12 B17 C12 C22 Y C49 son
idØnticas divergiendo de la cepa B 13 a partir de una distancia de similitud de 85 1
GØnero RhodotoruJa
Las 5 cepas restantes de este gØnero divergen a partir del 43 6 de similitud
produciØndose 2 fenones uno que contiene a las cepas A18c A23 y A43 Y otro que
50

integra las cepas 002 y 006 las cuales son idØnticas En el primer fenón se observa que
las 3 cepas divergen a partir de una distancia de similitud del 72 31 Dentro de este
fenónlas cepas A23 y A43 presentan una similitud del 100
A18A75812817C12C22
IC43813A19cA23 I IA43 I
I002 I006 I
1m ro 60 40 20 o
Distancia de Similitud
Figura 8 Dendograma generado a partir de la digestión de ADN de las cepasA18 A18c A23 A43 002 006 A75 Bl2 B13 B17 C12 y C22 por la enzima de
restricción Eco Rl El mØtodo de anÆlisis fue elligamiento promedio no ponderadoupGMA utilizando el coeficiente de correlación de Pearson l r
HinfI
Al digerir el ADN de las 13 cepas estudiadas con esta enzima de restricción se
originaron 23 bandas que oscilan entre 104 y 0 82 Kb La figura 9 representa el
dendograma generado a partir de los datos referentes a esta enzima Como en los
casos anteriores en el dendograma se distinguen dos grupos uno que hace referencia a
las cepas del gØnero Rhodotorula y el otro que reœne a las 7 cepas del gØnero
Debaryomyces ambos grupos divergen a partir deuna distancia de similitud de 3 6
GØnero Rhodotorula
Las 6 cepas estudiadas de este gØnero divergen segœn el anÆlisis realizado a
partir del 14 9 de similitud Del mismo modo que en los casos anteriores este gØnero
se divide en dos fenones uno que contiene a las cepas A18 A18c y A23 y otro que
integra las cepas A43 002 Y 006 En el primer fenón se observa que las 3 cepas
51

divergen a partir de una distancia de similitud de 564 Las cepas A18 y A18c
comparten una similitud de 72 8 Dentro del otro fenón la cepa 006 diverge de las
cepas A43 y 002 a partir de una distancia de similitud de 26 7 y estas dos comparten
una similitud de 56 9
A18A180A23A43002006A75C22C49817CI2813812
100 ro 60 40 20 o
Distancia de Similitud
Figura 9 Dendograma generado a partir de la digestión de ADN de las cepasA18 A18c A23 A43 002 006 A75 B12 B13 B17 C12 y C22 por la enzima derestricción Hinf 1 El mØtodo de anÆlisis fue elligamiento promedio no ponderadoupGMA utilizando el coeficiente de correlación de Pearson l r
GØnero DebaryomycesEste genero diverge a partir de una distancia de similitud de 32 8 En este caso
se observan dos fenones el primero que incluye las siguientes cepas A75 C22 C49
B 1 7 C12 y B 13 Y otro fenón cuyo œnico integrante es la cepa B 12 En cuanto al primer
subgrupo la cepa B13 difiere de las restantes a partir del 364 de similitud Las cepas
A75 C22 y C49 se separan de las cepas B17 y C12 a partir de una distancia de similitud
de 63 Las cepas C22 y C49 son idØnticas y presentan una similitud con la cepa A75
de 74 9 Por otro lado las cepas B17 y C12 son homólogas en un 908
PstI
La digestión del ADN con esta enzima produce 13 bandas cuyo tamaæo molecular
oscila entre 315 y 26 9 Kb En la figura 10 se puede observar el dendograma que se
52
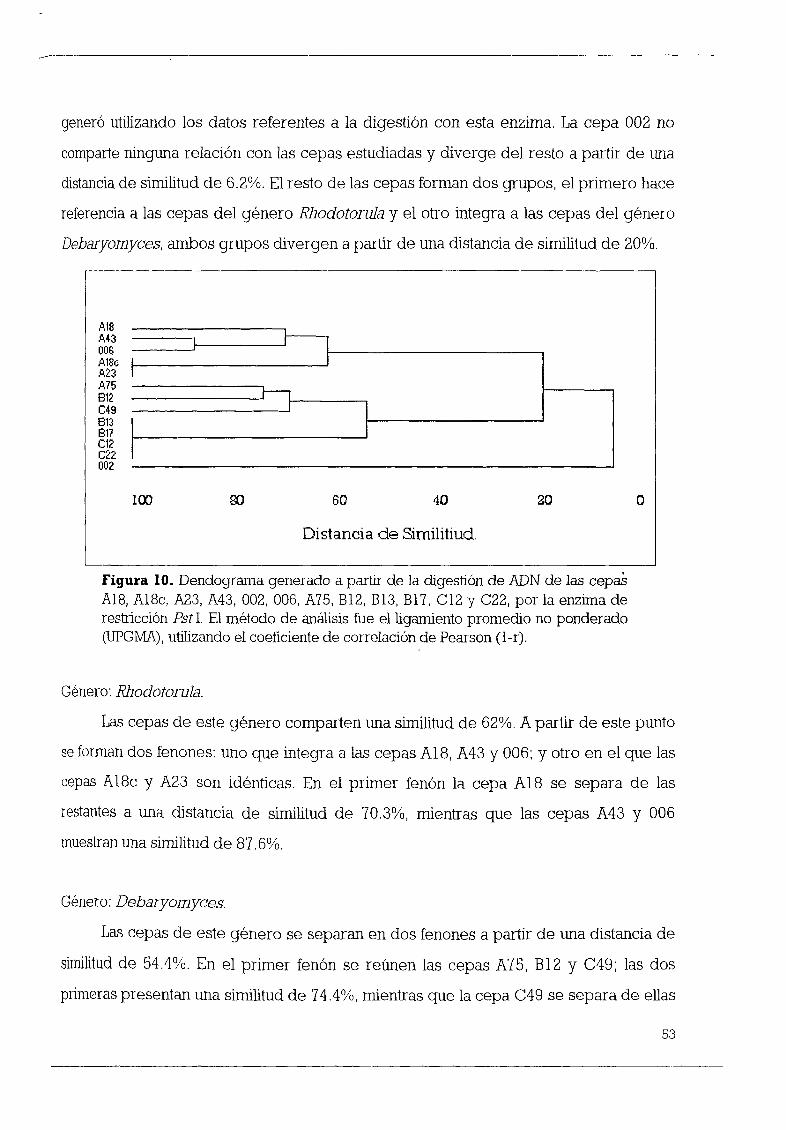
generó utilizando los datos referentes a la digestión con esta enzima La cepa 002 no
comparte ninguna relación con las cepas estudiadas y diverge del resto a partir de una
distancia de similitud de 6 2 El resto de las cepas formandos grupos el primero hace
referencia a las cepas del gØnero Rhodotorula y el otro integra a las cepas del gØnero
Debaryomyces ambos grupos divergen a partir de una distancia de similitud de 20
A18A43006A18cA23A75812C49813817C12C22002
100 ro 60 40 20 o
Distancia de Sirnilitiud
Figura 10 Dendograma generado a partir de la digestión de ADN de las cepaSA18 A18c A23 A43 002 006 A75 Bl2 B13 Bl7 C12 y C22 por la enzima derestricción Pst1 El mØtodo de anÆlisis fue elligamiento promedio no ponderadoupGMA utilizando el coeficiente de correlación de Pearson l r
GØnero Rhodotorula
Las cepas de este gØnero comparten una similitud de 62 A partir de este punto
se forman dos fenones uno que integra a las cepas A18 A43 Y 006 Y otro en el que las
cepas A18c y A23 son idØnticas En el primer fenón la cepa A18 se separa de las
restantes a una distancia de similitud de 70 3 mientras que las cepas A43 y 006
muestran una similitud de 87 6
GØnero Debmyomyces
Las cepas de este gØnero se separan en dos fenones a partir de una distancia de
similitud de 544 En el primer fenón se reœnen las cepas A75 B12 Y C49 las dos
primeras presentan una similitudde 74 4 mientras que la cepa C49 se separa de ellas
53

a partir de una distancia de similitud de 69 2 En el otro fenón se agrupan las cepas
B13 B17 C12 y C22 todas idØnticas
9 Características generales de las amplificaciones al azar deADN polimórfico RAPDs
Los cinco oligonucleótidos que se utilizaron para este ensayo generaron patrones
específicos para cada cepa Al revisar cada una de las amplificaciones es evidente que
existen claras diferencias entre las cepas de cada uno de los gØneros así en todos los
casos las cepas del gØnero Rl1odotorula siempre presentan un patrón completamente
distinto a los patrones que se generaron con las cepas de Debmyomyces Las bandas
de mayor tamaæo que se generaron de las cepas de Debmyomyces siempre fueron
mayores que las bandas de mayor tamaæo en el gØnero Rl1odotomla
Una característica importante de esta tØcIÙca es que ongma patrones
electroforØticos mÆs claros y en los que es mÆs fÆcil de distinguir las diferencias que
existen entre especies y cepas que los patrones que se generaron mediante el uso de
enzimas de restricción
Con los oligonucleótidos 1 y 2 fue con los que se generaron mayor nœmero de
bandas dentro de un gel 14 en ambos casos corresponde a la amplificación con estos
oligonucleótidos en cepas de Debmyomyces Con el oligonuc1eótido 3 ÚIÙcamente no
amplificó la cepa C 12 del gØnero Debaryomyces mientras que con los demÆs fue mÆs
de una cepa la que no amplificó AdemÆs el patrón electroforØtico mÆs claro que se
obtuvo fue el que se generó al amplificar con el oligonucleótido 3
Por œltimo cabe mencionar que aœn y cuando muchas de las cepas presentan
patrones idØnticos se presentan variaciones en cuanto a la intensidad de las bandas
10 AnÆlisis de los patrones electroforØticos generados por RAPDs
Oligonucleótido 1 secuencia TCACGGTGCA
Al amplificar el ADN genómico en las cepas A75 B17 C12 C22 y C49 utilizando
este oligonucleótido la banda de mayor tamaæo que se generó para las cepas de
Debaryomyces vanrijˇi vanrijii fue de 2 6 Kb mientras que en las especies del gØnero
54

Rhodotorula la banda de mayor tamaæo fue de 2 15 Kb en las cepas 002 y 006 Por otro
lado la banda de menor tamaæo que se observa en el gel corresponde a 0 56 Kb y la
comparten las cepas A18 A43 002 006 A75 B17 C12 C22 Y C49 Como se observa
en la figura 11 los patrones electroforØticos de las cepas de los gØneros estudiados son
completamente diferentes
Rhodotomla Debaryomyæs
1 2 3 4 5 6 7 8 9 10 11 12 1 3
Kb
23139 426 56
2 322 03
111
0 90
0 69
0 000 400 32
Figura 11 Patrón electroforØtico de las cepas de los gØneros Rhodotorula y
DebaJyomyces mediante la amplificación del oligonucleótido 1 por PCR Carriles
1 cepa A18 2 cepa A18c 3 cepa A23 4 cepa A43 5 cepa 002 6 cepa 006 7
A75 8 cepa B12 9 cepa B13 10 cepa B17 11 cepa C12 12 cepa C22 13 cepaC49
Al observar el patrón electroforØtico de las cepas del gØnero Rhodotorula se nota
que en las cepas A18c y A23 no hubo productos de amplificación mientras que el resto
de las cepas de este gØnero presentan un patrón muy similar entre ellas con la
salvedad de que la intensidad de las bandas varían entre las cepas
Con respecto a las cepas del gØnero Debaryomyces se observa que las cepas
B12 y B13 no amplificaron este oligonuc1eótido Las cepas A75 B17 C12 C22 Y C49
presentan un patrón idØntico aunque existen variaciones en cuanto a la concentración
de las bandas
55

Oligonucleótido GTCGCCGAC 2
Al observar los patrones electroforØtícos generados mediante la amplificación de
ADN genómico utilizando este oligonucleótído en las cepas de Rhodotorula y
Debaryomyces Figura 12 se notan marcadas diferencias La banda de mayor tamaæo
que se obtuvo corresponde a las cepas del gØnero Debaryomyces y corresponde a
6 19 Kb Por su parte en las cepas del gØnero Rhodotorula la banda de mayor tamaæo
fue de 2 051 Kb Dentro del gel la banda de menor tamaæo molecular fue de 0 47 Kb Y
se generó en las cepas A75 B12 B13 B17 y C49 Dfb vanrijii vanrijiJ En las cepas del
gØnero Rhodotorula se identificó un menor nœmero debandas en relación al nœmero de
bandas identificadas en las cepas del gØnero Debaryomyces
Rhodotom1a Debaryomyces
1 2 3 4 5 6 7 8 9 10 11 12 13Kb
2 32
2 m
111
090
0 69
0 00
Figura 12 Patrón electroforØtico de las cepas de los gØneros Rhodotomla y
Debaryomyces mediante la amplificación del oligonuc1eótido 2 por PCR Carriles1 cepa A18 2 cepa A18c 3 cepa A23 4 cepa A43 5 cepa 002 6 cepa 006 7
A75 8 cepaB12 9 cepa B13 10 cepaB17 11 cepaC12 12 cepaC22 13 cepa
C49
En cuanto al patrón electroforØtico generado a partir del ADN del gØnero
RhodotoruJa se observa que œnicamente en las cepas A18 A23 002 y 006 existen
productos de amplificación con este oligonucleótído Las cepas A23 002 y 006
presentan patrones electroforØticos idØnticos mientras que la cepa A18 presenta un
patrón muy similar aunque con un menor nœmero de bandas
56

Las cepas del gØnero Debaryomyces generaron patrones electroforØticos
idØnticos y solamente las cepas C12 y C22 no amplificaron con este oligonucleótido
Como en los casos anteriores la concentración de las bandas presentó variaciones
entre las cepas
Oligonucleótido GTAGACGAGC 3
Los patrones generados mediante la amplificación usando como templete este
oligonucleótido presentan diferencias entre los gØneros RhodotoruJa y Debaryomyces
como se puede observar en la figura 13 La banda de mayor tamaæo que se obtuvo
corresponde a un peso de 187 Kb y como en los casos anteriores es producto de la
amplificación de las cepas del gØnero Debaryomyces Este gØnero presentó un menor
nœmero de bandas que el gØnero RhodotoruJa
Rhodotomla Deharyomyæs
1 2 3 4 5 6 1 8 9 10 11 12 13 Kb
6 56
2 322m
1110 90
uu
0 89Ol
0 400 32
Figura 13 Patrón electroforØtico de las cepas de los gØneros Rhodotomla y
Debaryomyces mediante la amplificación del oligonucleótido 3 por PCR Carriles
1 cepa A18 2 cepa A18c 3 cepa A23 4 cepa A43 5 cepa 002 6 cepa 006 7
A75 8 cepaB12 9 cepaB13 lO cepaB17 11 cepaC12 12 cepaC22 13 cepa
C49
57

Los patrones electroforØticos de las cepas del gØnero RhodotoruJa muestran una
gran similitud pues comparten bandas del mismo tamaæo sin embargo la cepa A23
presento un mayor nœmero de bandas que el resto
De las cepas del gØnero Debaryomyces en la œnica en que no se obtuvo
producto de amplificación fue la cepa C 12 mientras que el resto de las cepas presentan
patrones idØnticos
Oligonucleótido AAACGTCGGG 4
De la misma manera que en los casos anteriores se observan marcadas
diferencias entre las cepas del gØnero RhodotoruJa y las del gØnero Debaryomyces
Como se observa en la figura 14 la banda de mayor tamaæo corresponde a los
productos de amplificación en las cepas del gØnero Debaryomyces con un tamaæo de
2 6 Kb mientras que en el gØnero Rhodotorula la banda de mayor tamaæo es de 2 05
Kb La banda de menor tamaæo fue de 0 37 Kb Debaryomyces
Rhodotom1a Deb8IYomyces
1 2 3 4 5 6 7 8 9 la 11 12 13 Kb
23139 476 00
3 332 m
111
0 90
069Ol
0 400 32
Figura 14 Patrón electroforØtico de las cepas de los gØneros Rhodotorula y
Debaryomyces mediante la amplificación del oligonuc1eótido 4 por PCR Carriles
1 cepa A18 2 cepa A18c 3 cepa A23 4 cepa A43 5 cepa 002 6 cepa 006 7
A75 8 cepa B12 9 cepa B13 10 cepa B17 11 cepa C12 12 cepa C22 13 cepaC49
58

Las cepas A18 A23 Y 002 son las œnicas del gØnero Rhodotorula que generaron
productos de amplificación sus patrones electroforØticos son idØnticos Por su parte
todas las cepas del gØnero Debaryomyces presentaron bandas de productos de
amplificación con la excepción de la B 1 7 Los patrones obtenidos son idØnticos entre sí
y solo presentan variaciones en la concentración de las bandas
Oligonucleótido ACTGACTGCC 5
La figura 15 muestra el patrón electroforØtico obtenido en las cepas de los
gØneros Rhodotorula y Debaryomyces Como se puede observar existe gran
diferencia entre los patrones obtenidos de un gØnero y del otro En este caso las cepas
del gØnero RllOdotorula presentan la banda de mayor tamaæo en el gel 2 72 Kb
mientras que en el gØnero Debaryomyces la banda de mayor tamaæo fue de 2 3 Kb
Dentro del gel la banda de menor tamaæo corresponde a 0 71 Kb Y es producto de la
amplificación con este oligonucleótido en las cepas del gØnero Rhodotorula
Rhodotom1a Debaryomyces
1 2 3 4 5 6 1 8 9 10 11 12 13
Kb
33139 476 00
2 322 m
1110 90
0 69
0 50
0 400 32
Figura 15 Patrón electroforØtico de las cepas de los gØneros Rhodotorula y
Debaryomyces mediante la amplificación del oligonucleótido 5 por PCR Carriles1 cepa A18 2 cepa A18c 3 cepa A23 4 cepa A43 5 cepa 002 6 cepa 006 7
A75 8 cepa Bl2 9 cepa B13 10 cepa B17 11 cepa C12 12 cepa C22 13 cepaC49
59

De las seIS cepas del gØnero RhodotoruJa que se analizaron solamente 4
generaron productos de amplificación A18c A43 002 Y 006 En cuanto a la
concentración de las bandas se observan variaciones entre las cepas A18c y A43
lIÙentras que las cepas 002 y 006 son similares entre sí Por otro lado la œnica cepa del
gØnero Debaryomyces que no amplificó este oligonucleótido fue la A75 el resto de las
cepas presentaron patrones idØnticos entre sí
Cabe mencionar aquí que con este oligonucleótido las cepas del gØnero
Debaryomyces presentaron menor variación en cuanto a la concentración de las bandas
en sus patrones electroforØticos que con los oligonucleótidos anteriores
60

The field of systematics has been in considerable turmoil as various
investigators developed different methods ofc1assification and argued their merits
1 guarantee you that no one method or view has all the good pointsW M Fitch 1984
V DISCDSION
Es relativamente poco lo que se conoce sobre la diversidad de los
llÙcroorganismos que constituyen los ecosistemas naturales y artificiales organismos
que son potencialmente œtiles en aplicaciones biotecnológicas Nuestro conocimiento de
la biodiversidad microbial se encuentra limitado debido al obstÆculo tradicional de los
microbiólogos el no poder cultivar la vasta mayoría 99 de los microorganismos
que se presentan en la naturaleza usando las tØcnicas estÆndar Los resultados que se
han obtenido mediante tØcnicas de biología molecular sobresaltan el hecho de que la
mayoría de los microorganismos no han sido aœn descritos Hugenholtz y Pace 1996
Actualmente se conocen WlOS 60 gØneros y 600 especies de levaduras Olsen
1990 sin embargo este nœmero solamente representa una fracción de las especies
existentes Roberts y Wildman 1995 Una clave para entender la diversidad microbial
es un sistema de clasificación confiable en el cual se puedan basar la identificaciones
Arnann et al 1994 Cada vez es mÆs aceptado que la taxonomía y los estudios
filogenØticos particularmente pueden brindar gran apoyo a otras Æreas de la biologíaPara generar predicciones experimentalmente probables acerca de la evolución de
muchos sistemas biológicos a partir de estudios bioquímicos y fisiológicos se pueden
usar hipótesis filogenØticas bien fundamentadas El punto mÆs importante en este
argumento es que la filogenia deberÆ de ser lo mÆs precisa posible puesto que las
imprecisiones dirigirÆn a conclusiones totalmente erróneas Quicke 1993
Existe poca literatura sobre las relaciones filogenØticas en las levaduras quizÆsdebido a que resolver el orden de las divergencias evolutivas es dificil tanto como lo es
para cualquier sucesión rÆpida de eventos evolutivos que ocurrieron hace mucho
tiempo
Las pruebas fisiológicas son indispensables para la caracterización taxonómica
de las levaduras Sin embargo al evaluar tanto la rentabilidad como la sensibilidad de
esta metodología podemos observar lo siguiente
61

Esta tØcnica es œtil para identificar levaduras a nivel gØnero lo que queda
corroborado por el hecho de que los porcentajes de participación de cada gØnero
dentro de los tres grupos sea tan alto Esto es especialmente cierto para grupos como
Rhodotorula donde las 6 cepas pertenecientes a tres especies diferentes se
integraron en el grupo 1 Lo mismo ocurre para las cepas de los gØneros
Leueosporidium Aureobasidium y Kluyveromyces sin embargo en estos gØneros
habrÆ que tomar esto con las debidas reservas puesto que en ambos casos solamente
se estudiaron a 2 cepas
La dispersión de las cepas de la especie Deb vanrijii yarrowi y de las 4 cepas
de Piehia phi10gaea se deben a las variaciones que se obtuvieron en las respuestas a las
pruebas por cepas de una misma especie De hecho HernÆndez Saavedra 1990 hace
mención sobre el polimorfismo que presentaron algunas de las especies de su estudio
poniendo especial atención a las características macroscópicas donde se presentaron
todo tipo de variaciones que son propias de cada especie De este modo las
variaciones reportadas en las respuestas a diferentes pruebas de asimilación de fuentes
de carbono y de nitrógeno en las cepas de Deb vanrijii yarrowi dieron como resultado
que se dispersaran en los tres grupos
Pese a que teóricamente las relaciones filogenØticas entre las cepas de Deb
vanrijii vanrijii son estrechas no quedan integradas en un sólo fenón al someterlas a las
pruebas bioquímicas y fisiológicas junto con las cepas de RhodotoruJa 3 especies lo
que demuestra que existe cierta variación en la respuesta a las pruebas
No se debe descartar la posible correlación entre la variación infraespecíficas de
las cepas y los sitios de donde se obtuvieron estas muestras HernÆndez Saavedra
1990 indica la existencia de un gran nœmero de microambientes en la zona de
muestreo debidos al encuentro de dos frentes oceÆnicos el tØmplado cÆlido con aguas
de la región Californiana y el tropical con aguas de la región PanÆmica lo que provoca
condiciones especiales en cuanto a mezclas de agua y disponibilidad de nutrimentos
De este modo el polimorfismo de las cepas colectadas podría deberse a los diferentes
microambientes especialmente si se consideran las muestras que fueron tomadas a
diferentes profundidades Así por ejemplo la distribución de especies del gØnero
62

Sporobolomyces encontradas en esta zona no siguen un patrón definido Los resultados
sugieren que existe una correlación entre la presencia de este gØnero y la salinidad y
oxígeno disuelto en el ocØano HernÆndez Saavedra et al 1992 Por otro lado en un
estudio mÆs reciente se sugiere una correlación R 0 75 entre la presencia de Candida
y la concentración de oxígeno disuelto y temperatura HernÆndez Saavedra et al
1995 Sin embargo como puntualizan los autores estos no son los œnicos factores que
se combinan para permitir la presencia y desarrollo de este gØnero la cantidad y tipo
de nutrientes tienen una fuerte influencia sobre las poblaciones y distribución de las
levaduras
ReslÙta evidente que esta tØcnica se puede considerar como un procedimiento
conveniente y prÆctico para corroborar resultados preliminares sobre la identificación
de muestras de levaduras sin embargo como apunta Olsen 1990 no como un
indicador bÆsico para la para la taxonomía de levaduras AdemÆs como se discute mÆs
adelante se debe contemplar el hecho de que actualmente se cuentan con herramientas
mÆs poderosas mÆs económicas y mÆs rÆpidas en las que se deberÆ apoyar la
taxonomía de levaduras
Esta es una tØcrnca que reqmere de mucho tiempo para obtener resultados
aproximadamente 7 días y en la que se consumen gran cantidad de reactivos lo que
la hace cara
En referencia a los patrones electroforØticos de proteínas se puede observar
lo siguiente
Si bien con esta metodología se generan mÆs grupos que con las pruebas
convencionales cada uno de ellos presenta fenones mucho mÆs compactos Por
ejemplo considerando el mismo caso que con las pruebas morfológicas y fisiológicas
aœn y cuando las cepas de Deb vanrijii vanrijii se dispersan dentro de tres grupos es
evidente que se mantiene cierta cohesión dentro de esta especie Sin embargo en la
variedad yarrowi las cepas analizadas se dispersan mucho menos que con las pruebas
tradicionales Lo mismo ocurre para las cepas de Deb melissophi1us y P pllllogaea que
63

se distribuyen en un sólo grupo Idealmente en un contexto genØtico las bandas de los
patrones electroforØticos de proteínas se interpretaran como alelos de un locus
Vaughan Martini el al 1987 El patrón de bandeo de las proteínas solubles de un
microorganismo es un reflejo fiel de una gran parte de su genóma van Vuuren y van
der Meer 1987 y por lo tanto se deben esperar menores variaciones en los
resultados manteniendo constantes las condiciones del cultivo que con las pruebas de
identificación tradicionales
Un hecho que se debe enfatizar y que ya se ha discutido por Lewontin y Hubby
1966 es que el mØtodo de la separación electroforØtica detecta œnicamente algunas
de las diferencias entre proteínas Muchas substituciones de arninoÆcidos pueden
ocurrir en una proteína sin generar una diferencia detectable en la carga neta De modo
que si bien esta tØcnica ofrece resultados objetivos rÆpidos y reproducibles se debe
considerar que actualmente existen tØcnicas de biología molecular que son mucho mÆs
sensibles
Considerando lo anterior esta tØcnica es œtil para la identificación de levaduras a
nivel especie Inclusive esta es una tØcnica sumamente œtil para agrupar e identificar
cepas de levaduras estrechamente relacionadas resultados que concuerdan con los
obtenidos por otros autores Yamazaki y Komagata 1981 Yamada el aJ 1986 van
Vuuren y van der Meer 1987 Yamada y Nakagawa 1988 Olsen 1990 Vancanneyt el
al 1991 Querol elaJ 1992 Boekhout elal 1993
El anÆlisis de los resultados de las cepas de Rhodotorula y Debaryomycesmuestra un dendograma muy similar al obtenido mediante las pruebas tradicionales de
identificación sin embargo como los patrones electroforØticos de proteínas presentan
menor variación dentro de la misma especie todas las cepas de Deb vanrijil vanrijil
quedan integradas dentro de un sólo fenón
De igual forma que como ocurrió con los resultados obtenidos mediante la
taxonoITÚa tradicional no se debe descartar el hecho de que las pequeæas variaciones
infraespecíficas puedan tener alguna correlación con el origen ecológico de los
aislamientos lo que concuerda con los resultados presentados por Vancanneyt el al
1991
64

La ocurrencia universal de un extenso polimorfismo proteico en las poblaciones
ha dirigido a algunos investigadores a creer que esta variación podría ser
fisiológicamente importante y consecuentemente encontrarse bajo control selectivo
otros la consideran sin ningœn efecto fenotípico y por lo tanto selectivamente neutral
Selander 1976 A pesar de Østo los datos derivados de esta tØcnica proveen una
percepción mÆs profunda del proceso evolutivo y de la taxonomía Kurtzman y Phaff
1987
El anÆlisis numØrico de los patrones electroforØticas de proteínas permite un
agrupamiento rÆpido y adecuado de un gran nœmero de cepas lo que concuerda con
los resultados presentados por Vancarmeyt etal 1991
EstÆ tØcnica como lo han reportado varios autores Vancarmeyt et al 1991
Kersters y De Ley 1975 van Vuuren y van der Meer 1987 Yamazaki y Komagata
1981 tiene la ventaja de ser bastante simple rÆpida y relativamente barata ademÆs de
generar en un sólo experimento un gran nœmero de características por cepa Por otro
lado esta metodología ofrece resultados altamente reproducibles
Los resultados que se generaron mediante RFLP restriction fragment length
polymorphisms permiten discutir los siguientes puntos
EstÆ tØcnica ha sido reportada como œtil para la diferenciación de cepas de
microorgarøsmos por varios autores Lee et al 1985 Hartan y Horgen 1989 Laaser et
aJ 1989 McCormick etal 1990 Olsen 1990 VÆzquez etal 1991 1993 Querol etal
1992 Boekhout et al 1993 Kurtzman 1994 Sin embargo la mayoría de ellos han
realizado sus trabajos específicamente con ADN mitocondrial ADNmt o ribosomal
rADN Kreitman 1991 por ejemplo enfatiza las siguientes ventajas de utilizar ADNmt
en lugar de ADN nuclear a es altamente polimórfico b su tasa de evolución es muy
alta y c es haploide Aunado a esto se encuentra el hecho de que el ADNmt no
recombina lo que significa que para la construcción de arboles filogenØticos se pueden
generar a partir de algoritmo s estÆndar Todos estos factores hacen que el ADNmt sea
65

mÆs sensible en la diferenciación de cepas de levaduras que utilizando el ADN
genómico Lee etaJ 1985
En este trabajo se utilizó ADN genómico nuclear y mitocondrial lo que provocó
que al realizar las digestiones con las enzimas de restricción Alu 1 y Kpn 1 se generaran
un gran nœmero de bandas que se observaron como una mancha o barrido Si bien
con el resto de las enzimas utilizadas se observan patrones de restricción definidos en
todos ellos existe este barrido de modo que la identificación de algunas de las bandas
fue dificil de determinar en muchos casos y en otros imposible Existen dos formas
posibles de evitar estas manchas 1 la utilización de enzimas de restricción que
reconozcan secuencias de un mayor nœmero de pares de bases Not 8 pb Sil 1 13
pb lo que probablemente mejoraría ligeramente las imÆgenes obtenidas o 2 el uso
de mtADN en lugar del ADN genórnico
Pese a lo anteriormente expuesto los patrones que se generaron ofrecen
información œtil Con el uso de las enzimas Bam HI y Hinf11as especies de Rhodotorula y
Debaryomyces quedaron claramente separadas En todos los casos se observa que el
grupo del gØnero Debaryomyces es mucho mÆs compacto lo que nos indica que entre
las cepas que se estudiaron existe una mayor identidad que con las cepas del gØnero
Rhodotorula
Se debe de considerar que las variaciones que se presentan se pueden deber
tambiØn a que este tØcnica es tan sensible que detecta diferencias no sólo a nivel
especie sino a nivel cepa Esto concuerda con el trabajo de Lee et aJ l985 en el que
concluyen que esta tØcnica es œtil no sólo para diferenciar una especie de
Saccharomyces de otra sino para diferenciar cepas de la misma especie remarcando
que el uso de esta deberÆ tener un nœmero de aplicaciones en microbiología de
fermentaciones Resultados similares obtuvieron Hartan y Horgen 1989 al trabajar con
el Oornyceto Achlya
En lo referente a la rentabilidad de esta tØcnica se pudo comprobar que es
rÆpida relativamente barata y rinde resultados suficientemente buenos ademÆs de ser
œtil para diferenciar cepas de una misma especie
66

El anÆlisis de los resultados obtenidos mediante las amplificaciones al azar de
ADNpolimórfico RAPD permite discutir lo siguiente
Las tØcnicas de identificación de levaduras basadas en el ADN tienen la ventajade ser independientes de la expresión de los genes Ness et al 1993 Ya se ha
mencionado que el uso de los patrones de restricción de ADN y los cariotipos por
electroforØsis han probado ser herramientas œtiles en la taxonomía de levaduras
Baleiras Couto et al 1995 Esta metodología ya se ha reportado como una
herramienta rÆpida para identificar organismos
Recientemente se ha descrito el uso de RAPD para discriminar entre
Saccharomyces cerevisiae Zygosaccharomyces bailii y Z rouxii Baleiras Couto et al
1994 Así mismo el alcance de esta tØcnica ha facilitado la diferenciación de cepas de
Listena monocytogenespertenecientes al mismo serovar Estos estudios y otros indican
que esta metodología puede ser una herramienta eficaz en la discriminación entre
cepas que de otro modo serían indistinguible s mediante pruebas serológicas o
bioquímicas Martin Kearley etaJ 1994
La elección de los oligonucleótidos es importante con respecto al poderdiscrirninatorio de este mØtodo Trabajos previos indican que en la tØcnica de RAPD el
nivel de diferenciación ya sea interespecífico o intraespecífico depende en buena
medida en los oligonucleótidos utilizados Baleiras Couto et al 1996 Los primers
utilizados en este trabajo pueden ser utilizados exitosamente para la identificación de
cepas de levaduras marinas utilizando ADN genómico resultados que concuerdan con
un estudio realizado por Mahenthiralingam etaJ 1996 sobre la identificación de cepas
de Pseudomonas aureoginosa en pacientes con fibrosis quística en el que afirma que
primers de 10 nucleótidos pueden ser utilizados con Øxito para la identificación de este
organismo
Aœn y cuando se utilizó el ADN genómico con esta tØcnica se obtienen imÆgenesclaras en las que las diferencias entre especies y o cepas son muy evidentes resultados
que son contrarios a los obtenidos por Abed 1995 quien reportó que el uso de estÆ
67

tØcnica con ADN genómico de cepas de Mycobacterium tuberculosis generó patrones
que contenían un gran nœmero de bandas lo cual lo mzo dificil de analizar
El anÆlisis de las cepas mediante estÆ metodología resultó ser muy rÆpido y
relativamente sencillo Es una tØcnica que resulta muy œtil y como menciona Kurtzman
1994 promete una amplia aplicación
Se puede decir que esta tØcnica es muy rentable si se toman en cuenta factores
como rÆpidez y poder de resolución ya que brinda muy buenos resultados en poco
tiempo Por otro lado se debe de tener en mente que esta metodología es
relativamente costosa debido a que requiere equipo especializado Sin embargo se
debe considerar que esta es una herramienta altamente eficaz para la diferenciación de
levaduras hasta el nivel infraespecífico de cepa
68

All biologists are involved in identification
but the taxonomist is especially concemed
since he produces or alters the classifications of organismsas well as providing the tools by which his fellow scientist
can identify their specimenWalters 1975
It is crucial in what follows that fue reader
keep a certain simple distinction clearly in mind
Criticising the logic of an argumentis quite different from
criticising that argument s conclusion
Sober 1988
VI CONCLUSIONES
l Hasta ahora el mØtodo mÆs popular de extracción de ADN de levaduras ha
sido producir protoplastos mediante la lísis enzimÆtica de la pared celular Oshiro etal
1987 Este procedimiento sin embargo presenta algunas limitaciones en ciertas
especies como en el caso de los gØneros Rhodotorula y Rhodosporidium que poseen
paredes celulares resistentes a los procedimientos de producción de protoplastos
HernÆndez Saavedra D 1991 Vishnuvardan y Joseph 1995 De los resultados
obtenidos en este estudio se concluye que el empleo de la tØcnica de extracción de
ADN que se utilizó es rÆpida y eficiente que el ADN obtenido no presentó evidencias
de degradación considerÆndose de buena calidad para su anÆlisis mediante el uso de
tØcnicas de biología molecular
2 Las pruebas convencionales de identificación de levaduras pruebas
bioquímicas y fisiológicas se deben considerar como procedimientos prÆcticos y
convenientes en los diagnósticos de rutina de los laboratorios para la separación de axa
mÆs que como indicadores bÆsicos de la taxonoIIÚa de levaduras Sin embargo
tØcnicas como la electroforØsis de proteínas totales el anÆlisis de los patrones de
restricción de ADN y RAPDs se deberÆn usar cada vez con mayor frecuencia en la
identificación y clasificación de levaduras
3 La identificación de levaduras mediante la electroforØsis de proteínas resulta
œtil para reconocer niveles taxonómicos a nivel genØrico como es el caso en los
gØneros Piehia Kluyveromyees Leueosporidium y Aureobasidium Sin embargo
69

algunos autores recomiendan que la determinación de relaciones inter e intragenØricas
se confirmen mediante otras tØcnicas
4 Los resultados obtenidos mediante RFLP permiten concluir que utilizando
ADN genómico el uso de las enzimas Bam HI y HinfI son herramientas œtiles para
diferenciar levaduras hasta el nivel gØnero Si bien en este trabajo no se utilizó ADN
mitocondrial se sabe que tratado con enzimas de restricción puede dar resultados mÆs
finos en cuanto a la diferenciación de levaduras por lo que se recomienda su uso para
diferenciar levaduras a nivel cepa
5 Los resultados que se obtuvieron mediante el uso de RAPD muestran que esta
tØcnica es œtil no solo para diferenciar levaduras a nivel gØnero puesto que con todos
los oligonucleótidos los patrones obtenidos en los gØneros Rhodotorula y
Debaryomyces fueron siempre diferentes sino que es una herramienta poderosa para
diferenciar levaduras a nivel especie y en algunos casos hasta nivel cepa
6 El oligonucleótido 3 resultó œtil para diferenciar ambos gØneros se obtuvo una
buena diferenciación de algunas de las especies de Rhodotorula y se observó la
diferenciación de algunas cepas de Debaryomyces vanrijjj vanrijii De igual manera el
oligonucleótido 5 proporcionó una buena resolución para diferenciar ambos gØneros
generando patrones especialmente buenos del gØnero Rhodotorula
7 La tØcnica de RAPD ha probado ser una herramienta poderosa para la
identificación de levaduras y dependiendo del oligonucleótido utilizado la información
que se obtiene es œtil para diferenciar levaduras hasta nivel cepa en forma rÆpida Los
resultados obtenidos con esta metodología son consistentes con los obtenidos por otros
autores Baleiras Couto et al 1994 Messner et al 1994 Baleiras Couto et al 1995
Paffetti et al 1995 Baleiras Couto etal 1996 ademÆs de ser exactos reproducibles y
objetivos
8 Los distintos tipos de electroforØsis aquí empleados son muy œtiles para
caracterizar levaduras marinas Son tØcnicas que ofrecen la ventaja de ser mÆs rÆpidas y
mÆs sensibles que las utilizadas tradicionalmente y los resultados que proporcionan son
70

reproducibles y objetivos Por estas razones se recomienda su uso como herramienta
complementaria en la identificación y clasificación de levaduras marinas
9 Estas tØcnicas permitirÆn la creación de bases de datos que ayudarÆn a una
rÆpida y eficaz identificación de diferentes levaduras satisfaciendo así las necesidades
de los investigadores en campos tan diferentes como la identificación clínica y las
diferentes industrias biotecnológicas
10 La capacidad de identificar moleclÙarmente cepas y especies de levaduras
tendrÆ un profundo efecto en el anÆlisis de las poblaciones naturales Con un mejorentendimiento de la diversidad microbial podremos hacer uso de un mayor nœmero de
compuestos en Æreas de la salud y alimentación y conoceremos los posibles efectos de
las perturbaciones provocadas por las actividades humanas en la estructura de las
comunidades de estos microorganismos11 Los estudios que incorporen tanto datos moleclÙares como morfológicos
proveerÆn mucho mejores descripciones e interpretaciones de la diversidad biológicaque aquellos que se basen exclusivamente en alguno de estos
71

r
VIII BIBLIOGRAF1A
Abed Y Davin Regli A Bollet C y de Micco p 1995 Efficient Discrirninationi
of Mycobacterium tuberculosis Strains by 16S 23S Spacer Region Based Random
Amplified Polymorfic DNA Analysis Journal of Clinical Microbiology 33 1418 1420
Ainsworth G C 1973 En The flmgi Vol Va Ainsworth G C Sparrow F K Y
Sussman A S eds Acadernic Press NewYork pp 1 7
Amann R Ludwig W y Schleifer K H 1994 Identification of Uncultured
Bacteria A Challenging Task for Molecular Taxonornists American Society of
Microbiology News 60 360 365
Bak A L Y Stenderup A 1969 Deoxyribonuc1eic Acid Homology in Yeasts
Genetic Re1atedness Witbin the Genus Candida Journal of General Microbio10gy 59
21 30
Baleiras Couto M M van der Vossen J M B M Hofstra H y Huis in t
Veld J H J 1994 RAPD Analysis A Rapid Technique for Differentiation of SpoilageYeasts lnternational Journal ofFood Microbio10gy 24 249 260
Baleiras Couto M M Vogels J T W E Hofstra H Huis in t Veld J H J Y
van der Vossen J M B M 1995 Random Amplified Polymorpbic DNA and
Restriction Enzyrne Analysis of PCR Amplified rDNA in Taxonorny Two Identification
Techniques for Food Borne Yeasts Journal ofApplied Bacterio10gy 79 525 535
Baleiras Couto M M Eijsma B Hofstra H Huis in t Veld J H J Y van der
Vossen J M B M 1996 Evaluation of Molecular Typing Techniques to Assign
72

Genetic Diversity Among Saccharomyces cerevisiae strains Applled and Environmental
Microbiology 62 41 46
Boekhout T Fonseca A Sampaio J P Y Golubev W l 1993 Classification of
Heterobasidiomycetous Yeasts Characteristics ans Affiliation of Genera to Higher Taxa
ofHeterobasidiomycetes CanadianJournal ofMicrobilogy 39 276 290
Botstein D Y Fink G R 1988 Yeast An Experimental Organism for Modern
Biology Science 240 1439 1443
Bradford M M 1976 A Rapid and Sensitive Method for Quantitation of Microgram
Quantities of Protein Utilizing the PrincipIe of Protein Dye Binding Analytical
Biochemistry 72 248 254
Davenport R R 1980 An Introduction to Yeasts and Yeast like Organisms En
Biology and Activities ofYeasts Skinner F A Passmore S M y Davenport R R eds
Academic Press New York pp 1 23
Fell J W Hunter l L Y Tallman A S 1973 Marine Basidiomycetous Yeasts
Rhodosporidium spp n with Tetrapollar Multip1e Allellc Bipolar Mating Systems
CanadianJournal of Microbiology 19 643 657
Fitch W M 1984 Cladistic and Other Methods Problerns Pitfalls and Potentials En
Cladistics Perspectives on the Reconstruction of Evolutionary History Duncan T y
Stuessy F eds Columbia University Press NewYork pp 221 252
Gould S J 1985 Taxonorny ofDeath Nature 313 505 506
73

BemÆndez Saavedra N Y 1990 Levaduras Marinas Aisladas en la Costa Occidental
de Baja California Sur MØxico Tesis de Licenciatura UNAM ENEP Iztacala pp 1 107
HemÆndez Saavedra N Y HemÆndez Saavedra D y Ochoa J L 1992
Distribution of Sporobolomyces KIuyver et van Niel Genus on the Western Coast of
Baja California Sur MØxico Systematic and Applied Microbiology 15 319 322
HemÆndez Saavedra N Y HemÆndez Saavedra D y Ochoa J L 1995
Factors Meeting the Distribution ofthe Genus Candida Berkhout Along the West Coast
ofBaja California Sur MØxico Systematic and Applied Microbiology 18 109 112
BemÆndez Saavedra D 1991 Composición Química de la Pared Celular de Distintos
GØneros de Levaduras Marinas Tesis de Maestría Universidad de Guanajuato Facultad
de QlÚmica Instituto de Investigación en Biología Experimental pp 1 86
Hillis D M Y Moritz C 1990 Molecular Systematics Hillis D M Y Moritz C
eds Sinauer Associates Sunderland Massachusetts pp 1 588
Hoffman C S y Winston F 1987 A Ten minute Preparation from Yeast Efficiently
Releases Autonomous Plasmids for Transformation in EscheÚchia coli Gene 57
267 272
Horton J S Y Horgen P A 1989 Polymorphisms in the Nuclear DNA of Achlya
Species Some Taxonornic Implications Canadian Journal of Microbiology 35
1146 1155
Hugenholtz P Y Pace N R 1996 Identifying Microbial Diversity in the Natural
Environment a Mo1ecular Phy10genetic Approach Trends in Biotechno1ogy 14
190 197
74

lwu M M 1996 Irnplernenting the Biodiversity Treaty How to Make International
Co operative Agreernents Work Trends in Biotechnology 14 78 83
Kersters K Y De Ley J 1975 ldentification and Grouping of Bacteria by Numerical
Analysis of Their Electrophoretic Protein Patterns Journal of General Microbiology 87
333 342
Kohlmeyer E Y Kohlemeyer J 1979 Marine Mycology The Higher Fungi
Acadernic Press New York pp 556 606
Kreger vanRij N J W 1969 En The Yeasts Rose A H Y Harrison J S eds Vol I
pp 5 78 AcadeIIÚc Press London y New York
Kreger van Rij N J W 1984 Classification of the ascosporogenous yeasts En The
Yeasts A Taxonomic Study Tercera Edición Elsevier Biornedical Press Amsterdam pp
25 32
Kreitman M 1991 Variation at the DNA LeveSornething for Everyone En
Molecular Techniques in Taxonorny Hewitt et aJ eds NATO ASI series Vol H57
Springer Verlag Berlín Heidelberg pp 15 31
Kurtzman C P y Phaff H J 1987 Molecular Taxonorny ofYeast En The Yeasts
Vol 1 Rose A H Y Harrison J S eds Academic Press New York pp 63 94
Kurtzman C P 1994 Molecular Taxonorny ofthe Yeasts Yeast 10 1727 1740
Laemmli U K 1970 Cleavage of Structural Proteins During the Assembly of the Head
ofBacteriophage T4 Nature 227 680 685
75

Laaser G Möller E ahnke K D Bahnweg G Prillinger H y Prell H
H 1989 Ribosomal DNA Restriction Fragment Analysis as a Taxonomic Too1 in
Separating Physio1ogical1y Similar Basidiomycetous Yeasts Systematic and Applied
Microbio1ogy 11 170 175
Lee S Y Knudsen F B Y Poyton R O 1985 Differentiation of Brewery Yeast
Strains by Restriction Endonuclease Analysis of Their Mithocondrial DNA J Inst Brew
91 169 173
Lewontin R C Y Hubby J L 1966 A Mo1ecular Approach to the Stdudy of Geruc
Heterozygosity in Natural Populations n Amount of Variation and Degree of
Heterozygosity in Natural Populations of DrosophiJa pseudoobscura Genetics 54
595 609
Mahenthiralingam E Campbell M E Foster J Lam J S Y Speert D P
1996 Random Amplified Polymorphic DNA Typing of Pseudomonas aureoginosa
Isolates Recovered from Patients with Cystic Fibrosis Journal of Cliruca Microbio1ogy
34 1129 1135
Mahrous M Lott T J Meyer S A Sawant A D Y Aheam D G 1990
Electrophoretic Karyotyping of Atypical Candida albicans Journal of Clinical
Microbio1ogy 28 876 881
Martínez J 1989 3er Curso Internacional de Tópicos sobre Taxonomía GenØtica y
Conservación de las Levaduras y su Aplicación en Biotecno10gía CINVESTAV IPN
MØxico pp 5 24
Martin Kearley J Gow J A PØloquin M y Greer C W 1994 Numerical
Ana1ysis and the Application of Random Amplified Po1ymorphic DNA Polymerase Chain
76

Reaction to the Differentiation of Vjbrio Strains from a Seasonally Co1d Ocean Canadian
Journal of Microbio1ogy 40 446 455
McConnick W A Stevenson R M W y Maclnnes J l 1990 Restriction
Endonuc1ease Fingerprinting Analysis of Canadian Iso1ates of Aeromonas salmonicida
CanadianJournal ofMicrobio1ogy 36 24 32
Messner R Príllinger H Altmann r Lopandic K Wimmer K Molnar
O y Weigang F 1994 Molecular Characterization and Application of Random
Amplified Polymorphic DNA Ana1ysis of Mrakia and SteJigmatomyces Species
International ournal ofSystematic and Bacterio1ogy 44 694 703
Ness r Lavallee F Dubourdieu D 1ügle M y Dulau L 1993 Identification
of Yeast Strains Using Po1ymerase Chain Reaction Journal of the Science of Food and
Agriculture 62 89 94
0lsen l 1990 Chemotaxonomy ofYeasts Acta Odonto1ogica Scandinava 48 19 25
Oshiro S Katsura N Kitada K y Gunge N 1987 A Simple Method for
Extraction of DNA from Fungi and Yeasts with Anhydrous Hydrogen Fluoride FEBS
Letters 220 383 386
Paffetti D Barberío C Casalone E Cavalierí D y Fani R 1995 DNA
Fingerprinting by Random Amplifíed Po1ymorphic DNA and Restriction Fragment Length
Po1ymorphism is Useful for Yeast Typing Research on Microbio1ogy 146 587 594
Phaff H J y Príce c W 1977 Strength and Weakness of Traditional Criteria in the
Systematics of Yeast as revealed by Nuclear Genome Comparison En Single Cell
77

Protein Safety for Animal and Human Feeding Paglialilllga S y Scrimshaw N S eds
Pergamon Press London pp 1 12
Ouerol A Barrio E y Ramón D 1992 A Comparative Study ofDifferent Methods
ofYeast Strain Characterization Systematic and Applied Microbio10gy 15 439 446
Quicke D L J 1993 PrincipIes and Techniques of Contemporary Taxonomy Blackie
Academic Professionals London
Rheiheimer G 1974 Aquatic MicrobioIogy John Wiley and Sons New York pp
13 14 59 74
Roberts l N Y Wildman H G 1995 The Diverse Potential ofYeast Biotechnology
13 1246
Rose M D Y Broach J R 1991 Cloning genes by Complementation in Yeast En
Guide to Yeast Genetics and Molecular Biology Guthrie C y Fink G R eds
Methods in Enzymo1ogy vol 194 Academic Press Inc pp 218 219
Sarnbrook J Fritsch E F Y Maniatis T 1989 Molecular Cloning A Laboratory
Manual Segunda Edición Cold Spring Harbor Laboratory Press pp 6 7
Selander R K 1976 En Molecular Evolution Ayala F J ed Sinauer Associates
Sunderland Massachusetts pp 21 45
Sober E 1988 Reconstructing the Pasí Parsimony Evolution and Inference The MIT
Press Cambridge Massachusetts
Sokal R R 1966 Numerical Taxonomy Scientific American 215 106 116
78

Storck R 1Uexopoulos J y Phaff H J 1969 Nucleotide Composition of
Deoxyribonucleic Acid of Some Species of Cryptococcus Rhodotorula and
Sporobolomyces Journal ofBacterio1ogy 98 1069 1072
Uden N van y Fell J W 1968 Marine Yeasts En Advances in Microbiology ofThe
Sea Drop M R Y Ferguson Wood E l eds Academic Press London y New York
pp 167 201
Vancanneyt M Pot B Hennebert G y Kersters K 1991 Differentiation of
Yeasts Species Based on E1ectrophoretic Who1e Cell Protein Patterns Systematic and
Applied Microbio1ogy 14 23 32
van der Walt J P 1970 Criteria and Methods Used in Classification En The Yeasts A
Taxonomic Study Segunda Edición E1sevier Biomedical Press Amsterdam pp 34 113
van Vuuren H J J y van der Meer L 1987 Fingerprinting of Yeasts by Protein
Electrophoresis American Journal ofEnology and Viticulture 38 49 53
Vaughan Martini 11 Sidenberg D G Y Lachance M 11 1987 Analysis of a
Hybryd Between 13 Kluyveromyces marxianus uyveromyces thermotolerance by
Physiological Profile Comparison Isoenzyme Electrophoresis DNA Reassociation and
Restriction Mapping of Ribosomal DNA Canadian Journal of Microbio1ogy 33
971 978
VÆzquez J 11 Beckley 11 Sobel J D Y Zervos M J 1991 Comparison of
Restriction Enzyme Analysis and Pulse Fie1d Gradient Gel E1ectrophoresis as Typing
Systems for Candida albicans Journal of Clínical Microbiology 29 962 967
79

VÆzquez J A Beckley A Donobedian S Sobel J D Y Zervos M J 1993
Comparison of Restriction Enzyme Analysis versus Pulse Field Gradient Gel
Electrophoresis as a Typing Systems for ToruJopsis glabrata and Candida Species Other
than e albicans Journal ofClinical Microbio1ogy 31 2021 2030
Vishnuvardan D Y Joseph R 1995 A Simple Method for Isolation of
High Mo1ecular Weight DNA from RhodotoruJa gracilis World Journal of Microbiology
and Biotechno1ogy 11 291 293
Walters S M 1975 Traditional Methods for Bio1ogical Identiification En Biological
Identification with Computers Systematics Association Volumen especial 7 Pankhurst
ed Academic Press Londres pp 3 8
Warburg O Y Christian W 1942 Biochemistry 310 348
Winge O Y Roberts C 1949 Inheritance ofEnzymatic Characters in Yeast and the
Phenomen of Long Term Adaptation En Comptes Rendus des Travaux de Laboratoire
Carlesberg SŁrie Physiologique 24 263 315
Yamada Y Y Nakagawa Y 1988 An E1ectrophoretic Comparison of Enzymes in
Strains of the Q9 equipped Species in the Genus Sporobolomyces Kluyver et van Nie1
Sporobo1omycetaceae Journal of General and Applied Microbio1ogy 34 289 295
Yamada Y Watanabe M Akita M y Banno l 1986 The Electrophoretic
Comparison of Enzymes in Strains of Species in the Anamorphic Yeast Genera
Sterigmatomyces and Fellomyces and m the te1eomorphic yeast genus
Sterigmatosporidium Journal of General and Applied Microbiology 32 157 163
80

Yamasato K Gota S Ohwada K Okuno D Araki H e lizuka H 1974
Yeast from the Pacific Ocean Journal of General and Applied Microbiology 20
289 307
Yamazaki M Y Komagata K 1981 Taxonomic Signíficance of Electrophoretic
Comparison of Enzymes in the Genera Rhodotorula and RhodospoJidiwn lnternational
Journal ofSystematic Bacteriology 31 361 381
81

IX APENDICES
ApØndice l Medios de cultivo
Medio MI líquido Glucosa 2 0
Peptona 1 0
Extracto de levadura 0 5
Agua de mar filtrada
pH 4 5 ajustado con HCI 0 1N
Medio Ml agar Medio MI líquido 2 0 de agar
pH 4 5 ajustado con HCI 0 1N
Medio YFD líquido Glucosa 2 0
Peptona 2 0
Extracto de levadura 1 0
Agua destilada
pH 5 6 ajustado con HCI O IN
Medio YFD agar Medio YFD agar 2 0 de agar
pH 5 6 ajustado con HCl 0 1N
82

ApØndice 2 Determinación deprotefnaspor elmØtodo deBradford 1976
PreparaC˝ón del reactivo de Bradford Se disuelven 100 mg de Azul Brillante de
Coomassie G 250 en 50 rnL de etanol al 95 A esta solución se le agregan 100 rnL de
Æcido fosfórico al 85 Aforar a 1 litro con agua destilada
Curva estÆndar Se prepara una solución stock de albœmina sØrica bovina a una
concentración de 100 Ilg rnL Hacer las siguientes diluciones O 5 lO 15 20 25 Ilg de
proteína m1 A 100 IlL de cada dilución se le adiciona 1 m1 del reactivo de Bradford y
se mezcla DespuØs de 5 min de incubación a temperatura ambiente se procede a leer
en el espectrofotómetro a 595 nm en cuvetas de 1 rnL La concentración de proteína se
grafica contra la absorbancia correspondiente para obtener la curva estÆndar que se
utilizara para determinar la concentración de proteína de las muestras
Microensayo de cuantificación de protefnas De cada muestra de proteínas se
hace una dilución 1 100 tomÆndose 10 fJL del extracto y 990 IlL de agua Se toman 100
jJL de esta dilución y se vacían en tubos de ensayo de 11 x 125 rnm A estos se les
agrega 1 rnL del reactivo de Bradford y se agitan DespuØs de 5 min se procede a su
lectura en el espectrofotómetro a 595 nm en cuvetas de 1 rnL
83

ApØndice 3 Reactivos requeridos para la electroforØsis de proteínas en geles
desnaturalizantes SDS PAGE basado en el mØtodo de Laemm1i 1970 Tomado de
Miniprotean II Electrophoresis Gell Instruction Manual Bio Rad
Soluciones stock
A AcrilamidaBisacrilamida 30
Acrilamida 29 2 grs
Bisacrilarnida 0 8 grs
Ajustar a 100 mL con agua destilada Filtrar y almacenar a 40C
Antes de usar desgasificar al vacío por 15 minutos
B 1 5 M Tris HCpH 8 8
Tris base 18 15 grs
Disolver en 50 mL de agua destilada Ajustar el pH a 8 8 con HCl1 N
Aforar a 100 mL Y almacenar a 40C
e 0 5 M Tris HCl pH 6 8
Tris base 6 grs
Disolver en 60 mL de agua destilada Ajustar el pH a 6 8 con HCIl N
Aforar a 100 mL Y almacenar a 40C
D 10 SDS
Disolver 10 gr de SDS en agua destilada bajo agitación suave y llevar a 100 ruL
E Buffer de corrimiento pH 8 3 5x
Tris base 45 grs
Glicina 216 grs
SDS 15 grs
Aforar a 3litros con agua destilada Almacenar a 40C
Diluir 300 roL del buffer 5x con 1200 roL de agua destilada
84

F 10 Persulfato de amonio PSA
Disolver 0 1 grs de PSA en 1 rnL de agua destilada Agitar
Preparar unos momentos antes de usarse
G Buffer de carga
Agua destilada 4 0mL
0 5 M Tris HC1 pH 6 8 10 roL
Glicerol 80mL
10 SDS 16mL
2 ß mercaptoetanol0 4
rnL 0 05Azuldebromofenol 0 2
roL Diluir lamuestra por lo menos 14conel buffer de
carga Calentar a950C por4
minutos Preparación del gelde
corrimiento I II I IIII 111 1 11 1 1 1 1 1 1111111111111111111111111111 mi i i ii i i i i i i 6 6
rnL 111 fllljm i i i8
mi IflImIII˛í il ˛ I5
rnL IIllli i j i j i iO 2
roL III III i ii iii i iO 2
mL IIII i i i i i i i i j j i i i i i i8
IlLPreparación del gelde
empaquetamiento i i i i i i i i i i i i i iI il I Iimljj ifI 11 i i i i i i i i ii i ii i i 3 4
mi 11 I I i ii ii O 83
roL IflI fllltllíjllj ltl iO 63
roL 111 ii I i i i i i ii iO 05
mLI III I i 1 1 1 150
IlL 111I1 il i i I j I i i I 5j
tL85

H Solución para teæir Azul de Coomassie R 250
Azul de Coomassie R 250 0 1 grs
Metanol 40 rnL
Acido acØtico 10 m1
Agua destilada 50 rnL
Filtrar en filtros Whatman No 1
Teæir por 30 ITÙnutos
1 Solución para desteæir
Metanol
Acido acØtico
Agua destilada
Desteæir durante 1 3 horas
40rnL
lOrnL
50rnL
86

ApØndice 4 TØcnicas para la extracción de Æcidos nuc1Øicos de levaduras
Preparación de esferoplastospor elmØtodo deRose y Broach 1991
l Crecer las cepas hasta la fase logarítmica 2 x 108 cØlulas rnL en 150 rnL de
medio YPD líquido
2 Centrifugar a 2 060 x g rma por 15 min a 50C Retirar el sobrenadante Lavar
los paquetes celulares con 30 m1 de 50 rnM EDTA pH 9 5 Centrifugar nuevamente a
2 060 x g rmaJ por 5 min a 50C Retirar el sobrenadante Resuspender las cØlulas en 30
mL de 50 rnM Tris HCl pH 9 5 2 2 ß mercaptoetanol durante 15 mina
temperatura
ambiente 3 Centrifugar las muestras a 2 060 x g rmax por 5 mm a 50C retirar
el sobrenadanteyresuspender las cØlulas en 3 rnL de una solución 1 M sorbitol 1
rnM EDTA pH 8 5Yagregar 50 IJg rnL de Zymoliasa 100T Incubar a 370C bajo
agitación suave Los esferoplastos sehabrÆn formado una vez que al revisaralmicroscopio
una gota del medio las cØlulas aparezcan negras aprox 60 120 min Centrifugar a 2 060
x g rmax por 5 min a
50C Extracción deADN de esferoplastos delevadura medianteel mØtodo deHoffman
y Winston
1987 1 Agregar 3 rnL de buffer 2 Tritón 100x 1 SDS 100 rnM NaCl 10
rnM Tris HCl 1 rnM EDTA pH 8 0al paquete de esferoplastos Adicionalmente agregar
5 m1 de una mezclafenol cloroformo alcohol isoarrœlico 25 24 1 y 9 rnL de perlas
de vidrio estØriles Agitar envortex durante 10 minen pulsos de 1min por 6min en
hielo 2 Centrifugar lasmuestras a 43 600 x g rmax durante 20 min a 50C Recuperar
el sobrenadante Dividir las muestras en tubos Eppendorf de 1 7 rnL de modo que
cada tubo contenga 650 IJLYagregar 1 3 rnL deetanol absoluto preenfriado a20oC
Reposar por el resto de la noche a
20oC87

3 Centrifugar las muestras a 15 900 x g Ja
durante 20 min a 50C Retirar el
sobrenadante Lavar los pellets con 250 fJL de etanol 70 mediante inversión del tubo
un par de veces Secar al aire
4 Resuspender las muestras en 100 fJL de agua destilada Almacenar a 20oC
Eliminación deARN
l Agregar a cada muestra 35 fJL de una solución 100x deRNAsa Incubar a 370C
durante 60 mino
2 Transcurrido este tiempo adicionar 500 L de la mezcla
fenolcloroformo alcohol isoamílico 25 24 1 Agitar en vortex y centrifugar a 15 900 x g
rmaJ durante 5 mino Recuperar la fase superior acuosa y agregar 600 fJL de la mezcla
anterior Agitar en vortex y centrifugar a 15 900 x g rmax por 5 mino Recuperar el
sobrenadante Agregar 400 fJL de cloroformo Agitar en vortex y centrifugar a 15 900 x
grma
durante 10 mino Recuperar el sobrenadante
3 Dividir la muestra en tubos Eppendorf de 17 roL Agregar 600 1L de etanol
absoluto Enfriar a 800C durante 15 mino Centrifugar las muestras a 15 900 x g CrmaJdurante 15 min a 50C Retirar el sobrenadante
4 Lavar el pellet por inversión del tubo con etanol al 70 dos veces Secar las
muestras al aire Disolver las muestras en 400 fJL de agua destilada estØril Almacenar a
290C Verificar pureza mediante electroforØsis y cuantificar a 260 nm
88

ApØndice S Soluciones para electroforØsis de Æcidos nuc1eicos
Buffer TBE lOx Tomado de Sambrook et al 1989
EDTA 0 5M pH 8 0
EDTA 36 54 grs
Disolver en 200 mL de agua destilada
Ajustar a pH 8 0 con NaOH 10 N
Aforar a 250 rnL
Buffer TBE 10x
Tris base 108 grs
Acido bórico 55 grs
Disolver en 600 mL de agua destilada
Agregar 40 mL de 0 5 M EDTA pH 8 0
Aforar a 1 litro
Buffer de carga para muestras deADN
Disolver 400 rng de sacarosa en 1 mL de agua destilada
Agregar 0 25 rng de azul de bromofenol
89





























