Embed Size (px)
Citation preview

BIBLIOTECAFaculdade de Ciências Farmacêuticas
Universidade de São Paulo
UNIVERSIDADE DE SÃO PAULOFACULDADE DE CIÊNCIAS FARMACÊUTICAS
Programa de Pós-Graduação em FarmáciaÁrea de Análises Clínicas
Alterações na resposta imune inata e adaptativa induzidas porEscherichia calí enterainvasora em modelo murino
Juliana Mata Khalil Amhaz
Tese para obtenção do grau deDOUTOR
Orientador:Prafa. Tit. Marina Baquerizo MartinezCo-orientador:Praf. Dr. Sandro Rogério de Almeida
São Paulo2007
.JB~92

DEDALUS - Acervo - CQ
1I11I111111111111 11111 111111111111111 1111111111 11111 11111 1111 I11I
30100013020
Ficha CatalográficaElaborada pela Divisão de Biblioteca e
Documentação do Conjunto das Químicas da USP.
Amhaz, luliana Mata KhalilAS15a Alterações na resposta imune inata e adaptativa induzidas
por Escherichia coli enteroinvasora em modelo murino I JulianaMota Khalil Amhaz. -- São Paulo, 2006.
118p.
Tese (doutorado) - Faculdade de Ciências Farmacêuticasda Universidade de São Paulo. Departamento de AnálisesClínicas e Toxicológicas.
Orientador: Martinez, Marina BaquerizoCo-orientador: Almeida, Sandro Rogério de
1. Microbiologia médica 2. Microorganismo patogênico3. Relação hospedeiro-parasita I. T. lI. Martinez. MarinaBaquerizo, orientador. III. Almeida, Sandro Rogério de,co-orientador.
616.01 COD

Juliana Mota Khalil Amhaz
Alterações na resposta imune inata e adaptativa induzidas porEscherichia cali enteroinvasora em modelo murino
Comissão Julgadorada
Tese para obtenção do grau de Doutor
Prafa. Tit. Marina Baquerizo Martinezorientador/presidente
~ d, Dt- a- CV\tlô ~
U~~a-~~~2°. examinador
~ GJ4ry~J~3°. examinador
Md R-và... ~ ~~~ \Jv~4°. examinador
São Paulo, ~ ok ~e cJjXF':, .

·o6!weeO~WJ!new'le!Ueoo't;/
·o~:5eJ!wpeeJowewo::>
'4l!P3eopt;?V\1's!edsnewso't;/

Agradecimentos
À Prafa. Tit. Marina Baquerizo Martinez, por ter me recebido em seulaboratório, e por estes dez anos de orientação.
Ao Praf. Dr. Sandra Rogério de Almeida, pela valiosa co-orientação epaciência nas correções finais da tese.
Ao Praf. Tit. Momtchilo Russo, por ter aceitado colaborar com este trabalho,pelas contribuições fundamentais e pela amizade.
Ao John, por todo o auxílio científico e por ter se tornado nestes anos o meumelhor amigo.
Ao Lucas, pelo auxílio valioso na execução dos experimentos e pelaamizade.
À Andyara, pela amizade e por me encorajar na reta final.
À Patrícia, Nilton e Marina, pelos bons momentos compartilhados.
À Estela, Antônio, Cristina, Priscila, Silvia e Vanessa, meus antigos colegasde laboratório, que estiveram presentes nos estágios iniciais dodesenvolvimento deste trabalho.
À Agda e a Carla, por terem guiado meus primeiras passos no laboratório.
Aos Professores Eisa Mamizuka, Primavera Borelli, Ana Campa e AntônioAltair, por permitirem a utilização de seu laboratório e pelo agradávelconvívio de todos estes anos.
Aos colegas do laboratório de Micologia, em especial ao Eliver e a Glória,pelo auxílio e amizade.
Aos colegas do laboratório de Parasitologia pelos empréstimos e convívioagradável.
À Karina, Esther e Eliane, do laboratório do Praf. Momtchilo, pelo auxílio nosensaios de FACS, NO e citocinas.
Ao Praf. Dr. Gustavo Amarante Mendes e sua aluna Jaqueline, pelo auxíliono ensaio de viabilidade dos macrófagos.
Ao Prof. Carlos Alberto Moreira Filho e a Patrícia Renovato Tobo, peloauxílio no ensaio de PCR em tempo real.

À Silvânia, Renata, Flávia e Fátima, do biotério do conjunto das químicas.
À Regina, do biotério do Instituto de Ciências Biomédicas.
Às secretárias do departamento de Análises Clínicas Ana, Sueli, Dora eEdna.
Às "meninas" da limpeza Dorlei, Cláudia, Carmem e Rose.
Ao Jorge e a Elaine, da secretaria de pós-graduação.
Ao José Edgar, pelo auxílio na análise densitométrica dos ensaios de RTPCR.
À minha família, em especial aos meus tios Bene, Vicente e Eliza, por toda atorcida.
À Tânia, Lívia, Alexandre, Lígia e André, pela amizade de todos estes anos.
À Andrezza, mais que uma prima, uma grande amiga.
À FAPESP, pela bolsa de doutorado, pelo financiamento do projeto, e porinvestir no desenvolvimento da ciência no país.
Aos camundongos que foram sacrificados para a realização deste trabalho.

RESUMO
Durante uma infecção, uma complexa seqüência de eventos é inkiadaapós a invasão do hospedeiro por microrganismos patogênicos. Escherichíacolí enteroinvasora (EIEC), assim como Shigella, causa disenteria através dainvasão da mucosa do cólon, levando à destruição tecidual e inflamação. Paraque ocorra um processo infeccioso, porém, são necessários inóculos de 102
Shigella e 106 EIEC. Foram avaliados aspectos da resposta inflamatóriadesencadeada pela infecção por EIEC em modelo murino, comparativamentea Shigella. A infecção de macrófagos J774 por EIEC resultou em fagocitosebacteriana, comprometimento da viabilidade do macrófago e produção decitocinas. Macrófagos de camundongos C57BU6 infectados com EIECproduziram NO, que parece ser importante no controle da infecção. Foiobservado que camundongos INOS nocaute apresentaram maior produção decitocinas pró-inflamatórias e maior letalidade após infecção do que osselvagens. EIEC induziu a migração de granulócitos e monócitos para operitônio, e a secreção de citocinas por estas células. Houve proliferação delinfócitos em resposta aos antígenos solúveis de EIEC, mas não foi detectadaprodução de citocinas por estes linfócitos.Comparativamente a Shigella, EIECescapou mais lentamente do macrófago, induziu menor produção de citocinaspró-inflamatórias e NO, e menor ativação dos linfócitos T. Estes dadossugerem o desafio com EIEC desencadeia uma resposta menos severa nohospedeiro do que Shigella, o que explicaria a forma mais branda de disenteriae resolução mais rápida do processo infeccioso causado por EIEC.

ABSTRACT
During an infection, a complex sequence of events in iniciated afterinvasion of the host by pathogenic microorganisms. EnteroinvasiveEscherichia coli (EIEC) and Shigella cause dysentery by means of invading thecolonic mucosa, leading to tissue destruction and inflammation. In arder for aninfectious process to occur, inocula of 102 Shigella are necessary incontrast toe 106 EIEC. The infection of J774 macrophages by EIEC resulted inphagocytosis of the bacterium, a hindering of the viability of the macrophageand in the production of cytokines. Macrophages obtained from C57BU6 miceinfected with EIEC produced NO, which seems to be important for the control ifinfection. We observed that in iNOS knockout mice, both the production of proinflammatory cytokines and lethality were higher than that observed in wild-typemice. EIEC induced the migration of granulocytes and monocytes to theperitoneum as well as the secretion of cytokines by these cells. We observed aproliferation of Iymphocytes in response to inoculation with soluble EIECantigens, however, in this case, the production of cytokines was not detected.Compared to Shigella, EIEC was slower in escaping from the macrophage, andinduced a shyer production of pro-inflammatory cytokines and NO, as well aspromoted a smaller activation of T Iymphocytes. These data suggest that whenchallenged with EIEC, the host produces a less severe response than thatelicited by Shigella, which might explain why the infectious process with EIECproduces a milder form of dysentery with a quicker resolution.

SUMÁRIO
1. Introdução
1.1 - O sistema mononuclear fagocítico
1.2 - Ativação de macrófagos
1.2.1 - Reconhecimento do patógeno e fagocitose
1.2.2 - Os receptores "TolI-like"
1.2.3 - Produção de espécies reativas de nitrogênio
1.3 - Macrófago: ponte entre a imunidade inata e a adaptativa
1.3.1 - B7-1 e B7-2
1.3.2 - Geração da resposta adaptativa
1.4 - O macrófago e os enteropatógenos
1.5 - Disenteria bacilar
1.6 - Shígella spp.
1.6.1 - Geração da resposta imune na infecção por Shígella
1.6.2 - Participação dos linfócitos T na patogênese da shigelose
1.7 - Escherichía colí enteroinvasora
2. Objetivos
2.1 - Objetivo geral
2.2 - Objetivos Específicos
3. Material e Métodos
3.1 - Amostras Bacterianas
3.2 - Padronização do inóculo bacteriano
3.3 - Cultura celular
Página
01
02
04
05
07
08
11
12
13
14
17
18
19
22
23
26
26
26
27
27
27
28

3.4 - Obtenção dos macrófagos de camundongo 28
3.5 - Extração de RNA e síntese de DNA complementar 29
3.6 - PCR em tempo real 29
3.7 - RT-PCR semiquantitativo 30
3.8 - "Enzyme-Iynked immunosorbent assay" (ELISA) para quantificação 32das citocinas
3.9 - Interação entre EIEC e macrófagos J774 (ensaios "in vitro') 33
3.9.1 - Sobrevivência bacteriana no interior dos macrófagos- 33
3.9.2 - Escape bacteriano após fagocitose e determinação da viabilidade 34dos macrófagos
3.9.3 - Produção de citocinas por macrófagos infectados 35
3.10- Interação entre EIEC e macrófagos de camundongos C57BU6 e 36C57BU6 iNOS KO (ensaios "in vitro")
3.10.1 - Produção de óxido nítrico (NO) 36
3.10.2 - Transcrição dos genes das citocinas, da enzima iNOS e do TLR4 36
3.10.3 - Produção de TNF-a e IL-10 37
3.11 - Interação entre EIEC e camundongos C57BU6 e C57BU6 iNOS KO 37"in vivo"
3.11.1 - Determinação do influxo celular para o peritônio após infecção 37
3.11.2 - Análise da transcrição dos genes das citocinas, da enzima iNOS e 39do TLR4
3.11.3 - Produção de TNF-a e IL-10 39
3.11.4 - Determinação da mortalidade de camundongos C57BU6 e 39C57BU6 iNOS KO após infecção
3.12 - Resposta de linfócitos à infecção por EIEC 40
3.12.1 - Ensaio de linfoproliferação 40
3.12.1.1 - Obtenção de linfócitos 40

3.12.1.2 - Obtenção de Antígenos Solúveis 40
3.12.1.3 - Linfoproliferação 41
3.12.2 - Produção de IFN-y, IL-10 e IL-4 42
3.12.3 - Determinação das subpopulações de linfócitos 42
4. Resultados 44
4.1 - Interação entre EIEC e macrófagos J774 (ensaios "in vitro") 44
4.1.1 - Determinação do número de bactérias viáveis no interior de 44macrófagos J774 após fagocitose
4.1.2 - Determinação do escape bacteriano após fagocitose 45
4.1.3 - Determinação da viabilidade dos macrófagos infectados 46
4.1.4 - Cinética da transcrição dos genes de TNF-a, IL-1 e IL-10 por PCR 47em tempo real
4.1.5 - Determinação da produção de TNF-a, IL-1 e IL-10 por ELISA 50
4.2 - Interação entre EIEC e macrófagos de camundongos C57BU6 e 52C57BU6 iNOS KO (ensaios "in vitro")
4.2.1 - Determinação da produção de NO pelo método de Griess 52
4.2.2- Análise da transcrição relativa dos genes das citocinas, da enzima 53iNOS e do TLR4 por RT-PCR semiquantitativo
4.2.3 - Determinação da produção de TNF-a e IL-10 por ELISA 60
4.3 - Interação entre EIEC e camundongos C57BU6 e C57BU6 iNOS KO 63"in vivo"
4.3.1 - Determinação do influxo celular para o peritônio após infecção 63
4.3.2 - Análise da transcrição relativa dos genes das citocinas, iNOS 68eTLR4 por RT-PCR semiquantitativo
4.3.3 - Determinação da produção de IL-1, IL-10 e IFN-y por ELISA 77
4.3.4 - Determinação da mortalidade de camundongos C57BU6 e 82C57BU6 iNOS KO após infecção

4.4 - Resposta de linfócitos à infecção por EIEC
4.4.1 - Linfoproliferação
4.4.2 - Produção de IFN-y, IL-10 e IL-4
4.4.3 - Determinação das subpopulações de linfócitos
5. Discussão
6. Conclusões
7. Referências Bibliográficas
83
83
88
89
91
111
113

LISTA DE FIGURAS
Figura 1 - Representação esquemática da patogênese da infecção porShigella
Figura 4.1 - Unidades Formadoras de Colônia de Shigella flexneriM9üT e EIEC FBC124/13 em lisados de macrófagosJ774 após diferentes intervalos de infecção.
Figura 4.2 - Unidades Formadoras de Colônia de Shigella flexneriM9üT e EIEC FBC124/13 no sobrenadante de culturasde macrófagos J774 após diferentes intervalos deinfecção.
Figura 4.3 - Determinação de alterações na viabilidade dosmacrófagos J774 em diferentes intervalos de infecçãopor Shigella flexneri M9üT e EIEC FBC124/13.
Figura 4.4 - Transcrição relativa do gene de TNF-y avaliada por PCRem tempo real.
Figura 4.5 - Transcrição relativa do gene de IL-1 avaliada por PCR emtempo real.
Figura 4.6 - Transcrição relativa do gene de IL-1ü avaliada por PCRem tempo real.
Figura 4.7 - Determinação, por ELISA de captura, da produção deTNF-y, IL-1 e IL-1ü por macrófagos J774 não infectadose infectados por sete horas com Shigella flexneri M9üT eEIEC FBC124/13.
Figura 4.8 - Determinação da produção de NO por macrófagosinfectados com Shigella M9üT e EIEC FBC124/13 pelométodo de Griess.
Figura 4.9 - Transcrição relativa do gene de TNF-a por macrófagos decamundongos C57BU6 e C57BU6 iNOS KO após 4horas de infecção, avaliada por RT-PCRsemiquantitativo.
Figura 4.10 -Transcrição relativa do gene de IL-1ü por macrófagos decamundongos C57BU6 e C57BU6 iNOS KO após 4horas de infecção, avaliada por RT-PCRsemiquantitativo.
Página
20
45
46
47
48
49
50
51
53
55
56

Figura 4.11 - Transcrição relativa do gene de IL-1 por macrófagos de 57camundongos C57BU6 e C57BU6 iNOS KO após 4horas de infecção, avaliada por RT-PCRsemiquantitativo.
Figura 4.12 - Transcrição relativa do gene de IL-18 por macrófagos de 58camundongos C57BU6 e C57BU6 iNOS KO após 4horas de infecção, avaliada por RT-PCRsemiquantitativo.
Figura 4.13 - Transcrição relativa do gene da TLR4 por macrófagos de 59camundongos C57BU6 e C57BU6 iNOS KO após 4horas de infecção, avaliada por RT-PCRsemiquantitativo.
Figura 4.14 - Transcrição relativa do gene de TNF por macrófagos de 60camundongos C57BU6 e C57BU6 iNOS KO após 4horas de infecção, avaliada por RT-PCRsemiquantitativo.
Figura 4.15 - Determinação, por ELISA de captura, da produção de 61TNF-a por macrófagos de camundongos C57BU6 eC57BU6 iNOS após 4 horas de infecção.
Figura 4.16 - Determinação, por ELISA de captura, da produção de 62IL-10 por macrófagos de camundongos C57BU6 eC57BU6 iNOS após 4 horas de infecção.
Figura 4.17 - Número de células em camundongos C57BU6 não 64infectados e após 12, 24 e 48 horas de infecção comShigella M90T e EIEC FBC124/13 determinado porcitometria de fluxo.
Figura 4.18 - Análise da transcrição de MHCII por macrófagos em 65peritônio de camundongos não infectados (linha preta)e após 12h (linha verde), 24h (linha rosa) e 48h (linhaazul) de infecção com Shigella flexneri M90T e EIECFBC124/13.
Figura 4.19 - Análise da transcrição de CD80 por macrófagos em 66peritônio de camundongos não infectados (linhavermelha) e após 12h (linha verde), 24h (linha rosa) e48h (linha azul) de infecção com Shigella flexneri M90Te EIEC FBC124/13.
Figura 4.20 - Análise da transcrição de CD86 por macrófagos em 67peritônio de camundongos não infectados (linhavermelha) e após 12h (linha verde), 24h (linha rosa) e

48h (linha azul) de infecção com Shigella f1exneri M9DTe EIEC FBC124/13.
Figura 4.21 - Análise da transcrição de F4/8D por macrófagos em 68peritônio de camundongos não infectados (linhavermelha) e após 12h (linha verde), 24h (linha rosa) e48h (linha azul) de infecção com Shigella f1exneri M9DTe EIEC FBC124/13.
Figura 4.22 - Transcrição relativa do gene de TNF-a por células 69peritoniais de camundongos C57BU6 e C57BU6 iNOSKO após 4 e 48 horas de infecção, avaliada por RT-PCR semiquantitativo.
Figura 4.23 - Transcrição relativa do gene de IL-1D por células 71peritoniais de camundongos C57BU6 e C57BU6 iNOSKO após 4 e 48 horas de infecção, avaliada por RT-PCR semiquantitativo.
Figura 4.24 - Transcrição relativa do gene de IL-1 por células 72peritoniais de camundongos C57BU6 e C57BU6 iNOSKO após 4 e 48 horas de infecção, avaliada por RT-PCR semiquantitativo.
Figura 4.25 - Transcrição relativa do gene de IL-18 por células 73peritoniais de camundongos C57BU6 e C57BU6 iNOSKO após 4 e 48 horas de infecção, avaliada por RT-PCR semiquantitativo.
Figura 4.26 - Transcrição relativa do gene de IFN-y por células 74peritoniais de camundongos C57BU6 e C57BU6 iNOSKO após 4 e 48 horas de infecção, avaliada por RT-PCR semiquantitativo .
Figura 4.27 - Transcrição relativa do gene de TLR4 por células 75peritoniais de camundongos C57BU6 e C57BU6 iNOSKO após 4 e 48 horas de infecção, avaliada por RT-PCR semiquantitativo.
Figura 4.28 - Transcrição relativa do gene de IP-1D por células 76peritoniais de camundongos C57BLl6 e C57BU6 iNOSKO após 4 e 48 horas de infecção, avaliada por RT-PCR semiquantitativo.
Figura 4.29 - Transcrição relativa do gene da iNOS por células 77peritoniais de camundongos C57BU6 após 4 e 48horas de infecção, avaliada por RT-PCRsemiquantitativo.

Figura 4.30 - Determinação, por ELISA de captura, da produção de 79TNF-a por células peritoniais de camundongosC57BU6 e C57BU6 iNOS KO após 4 e 48 horas deinfecção.
Figura 4.31 - Determinação, por ELISA de captura, da produção de IL- 8010 por células peritoniais de camundongos C57BU6 eC57BU6 iNOS KO após 4 e 48 horas de infecção.
Figura 4.32 - Determinação, por ELISA de captura, da produção de 82IFN-y por células peritoniais de camundongos C57BU6e C57BU6 iNOS KO após 4 e 48 horas de infecção.
Figura 4.33 - SDS-PAGE dos antígenos secretados por EIEC 84FBC124/13 e Shigella M90T após 5 horas de incubaçãoem contato com células Hep-2.
Figura 4.34 - índice de proliferação de linfócitos de camundongos 85imunizados com Shigella flexneri M90T e EIEC FBC124/13.
Figura 4.35 - índice de proliferação de linfócitos de camundongos 88imunizados com Shigella flexneri M90T, EIECFBC124/13 e E.coJi K12 por sete dias e posteriormenteestimulados com 100 ~g de antígenos solúveis deM90T, EIEC e K12.
Figura 4.36 - Produção, de IL-10 e IFN-y por linfócitos de 89camundongos Balb/C imunizados com Shigella flexneriM90T e EIEC FBC 124/13 e posteriormenteestimulados por 72h com antígenos solúveis deShigella e EIEC, respectivamente.
Figura 4.37 - Citometria de fluxo de células dos Iinfonodos de 90camundongos infectados com Shigella M90T e EIECFBC24/13.

1 -INTRODUÇÃO
Durante uma infecção, uma complexa seqüência de eventos é iniciada
após a invasão do hospedeiro por microrganismos patogênicos (MEDZHITOV
& JANEWAY, 2000).
Para estabelecer e manter uma infecção, os patógenos microbianos têm
desenvolvido uma série de estratégias para invadir o hospedeiro, evitar ou
resistir à resposta imune, e colonizar sítios específicos do organismo
(COSSART & SANSONETTI, 2004; SANSONETTI & DI SANTO, 2007).
Por outro lado, o sistema imune inato é responsável por detectar os
primeiros sinais da infecção e mantê-Ia sob controle até que a imunidade
adaptativa instale uma resposta eficiente. As células componentes da
imunidade inata detectam as infecções bacterianas por meio de receptores
que se ligam a estruturas comumente expressas em microrganismos, mas não
nas células do hospedeiro. Esta ligação desencadeia uma cascata de
sinalização que leva, em última instância, à expressão de genes pró
inflamatórios e secreção de uma série de mediadores (MEDZHITOV &
JANEWAY, 2000; GRASSL & FINLAY, 2007).
Neste contexto, as interações entre microrganismos e macrófagos
desempenham um papel crucial na patogênese de muitas infecções. Os
macrófagos são células fundamentais da resposta imune inata e adaptativa,
funcionando como "sentinelas" que guardam os pontos potenciais de infecção.
Os patógenos encontram os macrófagos assim que ganham acesso aos
tecidos do hospedeiro e o resultado deste encontro determina se o hospedeiro

2
vai produzir uma resposta imune eficiente ou se o patógeno vai estabelecer
com sucesso a infecção (NAVARRE & ZYCHLlNSKY, 2000).
1.1 - O sistema mononuclear fagocítico
O sistema mononuclear fagocítico é definido como uma linhagem celular
derivada de células progenitoras da medula óssea. É constituído por
monoblastos e promonócitos da medula óssea, monócitos do sangue e
macrófagos residentes. Estes últimos encontram-se amplamente distribuídos
em diferentes tecidos, exibindo uma grande heterogeneidade fenotípica e
funcional (PAULNOCK, 2000; HUME, 2006).
O macrófago origina-se do monócito sanguíneo, cuja célula precursora
na medula é o monoblasto. Após migração para sítios extravasculares o
monócito sofre um processo de maturação, sendo então denominado
macrófago residente. O processo de diferenciação resulta tanto em perda
como em aquisição de características fenotípicas e funcionais. (PAULNOCK,
2000; GORDON, 2002).
Os macrófagos podem se apresentar como células fixas em tecidos ou
como macrófagos livres. Alguns macrófagos tissulares recebem denominações
específicas, como células de Kupffer (fígado), microglia (sistema nervoso
central), células de Langerhans (pele). Macrófagos livres estão presentes em
uma série de localizações anatômicas, como espaço alveolar e cavidade
peritonial (PAULNOCK, 2000; GORDON, 2002).
A migração do monócito para os tecidos é mediada por moléculas
expressas na sua superfície e de células endoteliais. Estas moléculas,

3
denominadas moléculas de adesão, são agrupadas em três grandes famílias:
A superfamília das imunoglobulinas, das selectinas e das integrinas. A L
selectina é expressa em leucócitos enquanto a P e a E selectinas são
expressas no endotélio. As ~2 integrinas são expressas na superfície dos
leucócitos e se ligam aos seus receptores da família das imunoglobulinas (as
"intracellular adhesion molecules", ICAM-1, 2 e 3), expressas na superfície de
células endoteliais. Em processos inflamatórios, um número maior de
monócitos migra através do endotélio vascular a fim de alcançar o sítio de
infecção (PANES e coL, 1999; MARSHALL & HASKARD, 2002).
Os fagócitos mononucleares participam de numerosos processos
homeostáticos e imunológicos. Sua atividade endocítica é responsável pela
remoção de células apoptóticas e debris celulares, e pela fagocitose de
microrganismos e apresentação de antígenos aos linfócitos para geração da
resposta imune celular e humoral (GORDON, 1998; UNDERHILL e coL, 1999).
Além disso, estas células possuem uma grande atividade secretora. Algumas
moléculas são secretadas constitutivamente, mas a maioria delas é produzida
apenas após ativação por produtos bacterianos, por exemplo. Entre as
moléculas secretadas estão os mediadores lipídicos, enzimas, citocinas, e
receptores de citocinas (PAULNOCK, 2000).
Citocinas são proteínas que desempenham funções fisiológicas e
inflamatórias. IL-3 e os CSF ("colony stimulating factor") , por exemplo, atuam
na hemopoiese, promovendo a proliferação e maturação de células miéloides,
granulócitos e monócitos (HUME, 2002). Outras citocinas são essenciais na
indução e resolução de processos inflamatórios. IL-1, IL-5, IL-18, TNF-a, IL12

'"
4
e IFN-y são exemplos de citocinas pró-inflamatórias que participam da ativação
de macrófagos e linfócitos T e estimulam o influxo celular. TGF-13 e IL-10,
citocinas anti-inflamatórias, têm como função principal limitar a magnitude da
infecção, inibindo a produção de citocinas pró-inflamatórias por macrófagos e
linfócitos (GORDON, 1998; PAULNOCK, 2000; GORDON, 2002).
1.2 - Ativação de macrófagos
O surgimento de anormalidades no tecido, como traumas, infecções ou
células neoplásicas ocasiona um rápido recrutamento de macrófagos. Os
macrófagos recrutados exibem diferenças fenotípicas dos residentes que
dependem da natureza e localização do estímulo. O termo genérico
"macrófago ativado" é comumente usado para descrever este processo,
(GORDON, 2002; HUME, 2006).
Macrófagos ativados apresentam aumento na capacidade microbicida e
tumoricida, resultante do aumento na produção de espécies reativas de
nitrogênio (RNI) e oxigênio (ROI). Apresentam, também, aumento na
capacidade de apresentar antígenos para as células T, pelo aumento na
expressão de moléculas do complexo principal de histocompatibilidade (MHC),
e moléculas co-estimulatórias, principalmente 87-1 e 87-2 (MEDZHITOV &
JANEWAY, 1997; NASCIMENTO e coL, 2003).
O processo de ativação celular está relacionado a um aumento na
expressão, ou ainda, à supressão de genes que codificam proteínas
essenciais para o desempenho de funções como fagocitose, processamento e
apresentação de antígenos, inativação e destruição de microrganismos,

5
ativação e proliferação dos próprios macrófagos e outros tipos celulares. Uma
análise do transcriptoma de macrófagos murinos mostrou que, em resposta a
estímulos como o LPS, há uma grande mudança no perfil de expressão de
genes. Os genes que são expressos em altos níveis no estado basal são
fortemente reprimidos pela ativação com LPS, enquanto a expressão de outros
genes é fortemente ativada (HUME, 2002). Entre os genes cuja expressão é
ativada por LPS, podemos citar genes que codificam citocinas (IL-1, TNFa, IL
6, IL-12, IL-10, MIP, IP-10), fatores de transcrição, enzimas, (hidrolases,
colagenases), enzimas relacionadas à produção de óxido nítrico e
prostaglandinas (iNOs, arginase, cicloxigenases), moléculas relacionadas à
apresentação de antígeno (MHCII, 87.1, 87.2), entre outros.
1.2.1 - Reconhecimento do patógeno e fagocitose
Entre as estratégias para internalizar partículas desenvolvidas pelas
células ao longo da evolução das espécies está a fagocitose, mecanismo de
captura de partículas grandes dependente de actina. Em organismos
unicelulares a fagocitose é utilizada para aquisição de nutrientes, enquanto
que em metazoários é desempenhada principalmente por células fagocíticas
especializadas, como macrófagos, células dendríticas e neutrófilos (ADEREM
& UNDERHILL, 1999; ADEREM, 2002). Entretanto, outros tipos celulares,
como células epiteliais e fibroblastos, também apresentam a capacidade de
internalizar partículas, tanto em cultura quanto "in vivo". Estas células,
denominadas fagócitos não-profissionais, não possuem receptores de
imunoglobulinas e complemento, e também não secretam toda a gama de

6
citocinas secretadas pelos fagócitos especializados. Sua capacidade
fagocítica, portanto, é limitada (RABINOVICH, 1995). A fagocitose por células
do epitélio intestinal é importante na patogênese de algumas infecções, como
a shigelose (item 1.6).
O reconhecimento de componentes alterados do hospedeiro ou
estranhos é mediado por receptores de membrana do macrófago. A ligação do
componente ao receptor gera sinais intracelulares que são responsáveis pelo
rearranjo e polimerização da actina e coordenam as forças tracionais que
internalizam as partículas (ADEREM & UNDERHILL, 1999; ADEREM, 2002).
O reconhecimento de patógenos consiste em uma séria dificuldade para
a imunidade inata, devido à enorme variabilidade e alta taxa de mutação
característica dos microrganismos. Em contrapartida, o sistema imune inato
desenvolveu receptores que têm como alvos estruturas moleculares que (i)
são comuns a grandes grupos de patógenos, (ii) são produzidos por
patógenos, não pelo hospedeiro e (iii) são essenciais para a sobrevivência ou
patogenicidade do microrganismo. Estas estruturas são denominadas padrões
moleculares associados a patógenos (PAMPs) (MEDZHITOV & JANEWAY,
1997; MEDZHITOV & JANEWAY, 2000).
Os PAMPs são reconhecidos pelos receptores denominados receptores
de reconhecimento de padrões (PRRs). Membros de várias famílias de
proteínas funcionam como PRRs. As mais proeminentes são os domínios de
Lectina tipo C, os receptores "scavenger" e os domínios ricos em repetições de
leucina (LRR). Os PRRs são expressos em células que são a primeira linha de
defesa do organismo, como células epiteliais, e também em células

7
apresentadoras de antígeno, como macrófagos e células dendríticas
(ADEREM & UNDERHILL, 1999; MEDZHITOV & JANEWAY, 2000).
Os PRRs estão envolvidos na opsonização de vírus e bactérias para
fagocitose ou ativação de complemento; na fagocitose de patógenos por
APCs; ou na ativação de vias de sinalização que ativam fatores transcricionais,
como NF-K8 e AP-1, responsáveis pela indução da expressão de mediadores
inflamatórios, entre eles citocinas, moléculas de adesão e enzimas como a
cicloxigenase 2 (COX-2) (GORDON, 1998; MUKHOPADHYAY e coL, 2004).
1.2.2 - Os receptores "TolI-like"
A identificação dos receptores To11 foi um grande avanço na
compreensão sobre o reconhecimento microbiano pelo sistema imune inato.
Seu envolvimento na imunidade inata foi inicialmente descrito em Drosophila,
e, posteriormente foram identificados homólogos em humanos, denominados
receptores ''TolI-like" (TLR). Tanto as proteínas TolI de humanos quanto de
Drosophila são receptores transmembrânicos que contém um domínio LRR
(Leucine-rich repeat) extracelular e um domínio citoplasmático (responsável
pela sinalização intracelular) que é homólogo ao domínio citoplasmático do
receptor da IL-1 (IL-1R), e, portanto, é chamado de domínio ToII/IL-1R (TIR)
(MEDZHITOV &JANEWAY, 2000).
Foi demonstrado que Drosophilas que apresentavam mutações em
genes de diferentes proteínas TolI mostraram ser suscetíveis a diferentes tipos
de patógenos, como fungos e bactérias Gram-negativas. Da mesma maneira,
existem dez membros da família dos TLRs em humanos e 11 em roedores já

8
identificados, que permitem às células do sistema imune inato reconhecer
diversas estruturas de microrganismos (TAKEDA e coL, 2003).
O primeiro receptor "toll-like" identificado foi o TLR4. Foi demonstrado
que duas linhagens de camundongos hipo-responsivas ao LPS, C3H/HeJ e
C57BL1O/ScCr, apresentavam mutações no gene Tlr4, confirmando que este
receptor é fundamental no reconhecimento do LPS. Além do LPS, o TLR4
reconhece outros ligantes, como o Taxol, um produto vegetal, e algumas
proteínas de choque tóxico (HSP60 e HSP70) (PERERA e coL, 2001; TAKEDA
e coL, 2003).
A expressão dos TLRs varia de acordo com o tipo celular. Os monócitos
e macrófagos possuem mRNA para todos os TLRs, com exceção de TLR3. Os
TLRs são expressos também em células epiteliais. A expressão de TLR4 por
células intestinais é relativamente baixa, e provavelmente está relacionada à
ausência de uma resposta inflamatória severa no intestino, apesar da
constante exposição ao LPS (TAKEDA e coL, 2003).
1.2.3 - Produção de espécies reativas de nitrogênio
Os RNI são compostos pelo radical óxido nítrico (NO), que em meio
aquoso reage formando outros radicais (N02), ânions(N02-, N03-), óxidos
(N20 3) e peróxidos (ONOO}
O NO é gerado em uma reação na qual uma molécula de L-arginina é
oxidada produzindo uma molécula de L-citrulina e uma de NO. A enzima que
catalisa esta reação é a NO sintase, que existe em três isoformas: endotelial
(eNOS, também conhecida como NOS3), neuronal (nNOS, também conhecida

9
como NOS1) e induzível (iNOS) (RESTA-LENERT & BARRET, 2002). Grosso
modo, a expressão de eNOS e nNOS é constitutiva, e a produção de NO por
estas isoformas é pequena e responsável pelas funções fisiológicas no
indivíduo saudável. A iNOS, no entanto, produz grandes quantidades de NO
após estímulo (daí o nome induzível), geralmente de natureza inflamatória e
infecciosa.
A expressão da iNOS ocorre em vários tipos celulares e é estimulada
por várias citocinas, como IL-113, TNF-a., IFN-y e componentes microbianos,
como o LPS.
O NO produzido pela iNOS desempenha um importante papel em
processos patológicos, sendo um dos responsáveis pelo efeito microbicida e
tumoricida de macrófagos ativados. Este efeito ocorre pela inibição de enzimas
das células-alvo, como GAPDH, ribonucleotídeo redutase, complexos I e IV da
cadeia respiratória, além de modificação do DNA (MACMICKING e coL, 1997,
CONNELLY e coL, 2001).
O NO produzido por macrófagos mostrou ter efeito citostático ou
citotóxico sobre uma série de patógenos, sejam eles bactérias, fungos ou
parasitas. Foi demonstrado que camundongos com uma deleção no gene da
iNOS apresentaram um aumento na suscetibilidade a infecções por
Mycobacterium tuberculosis, Listeria monocitogenes, Toxoplasma gondii e
Leishmania major (WAY & GOLDBERG, 1998; CHAKRAVORTIY e coL,
2002). Estudos envolvendo camundongos infectados com M. tuberculosis
mostraram uma correlação entre expressão da iNOS e subseqüente resolução
da infecção, enquanto a falha na expressão desta enzima foi associada à

10
progressão da infecção (SHILOH & NATHAN, 2000). A importância do NO na
eliminação de SalmoneJla por macrófagos murinos também está documentada
(BABU e coL, 2006).
O NO também é importante na fisiopatologia do processo inflamatório,
pois estimula a síntese de citocinas, como IL-6 e TNF-a (MACMICKING e coL,
1997; NASCIMENTO e coL, 2002). Na patogênese da infecção por L.major, foi
evidenciado o papel da iNOS como indutor da produção de IFN-y. No princípio
da infecção há um aumento na produção de IFN-a, que induz um pequeno
aumento na expressão da iNOS, necessário à maturação das células NK.
Estas células produzem IFN-y, que estimula o incremento da expressão de
iNOS, levando à eliminação do parasita (SHILOH & NATHAN, 2000).
A produção de RNI pelos macrófagos reflete não só no aumento da sua
capacidade microbicida, mas também afeta a suscetibilidade destas células a
apoptose. Foi observado que pequenas concentrações de RNI produzidos por
macrófagos ativados retardam a apoptose por inibição dos sítios ativos das
caspases, proteínas que participam de um dos mecanismos de apoptose.
Porém, em altas concentrações, os RNI podem reagir com uma série de alvos
moleculares, levando à morte dos macrófagos por apoptose ou necrose
(SHILOH & NATHAN, 2000).
Além de seus efeitos fisiológicos e pró-inflamatórios, o NO em altas
concentrações também tem efeito imunossupressor. Em macrófagos murinos,
a regulação da expressão da iNOS é controlada pelo fator transcricional NF
KB. O NO produzido em resposta a estímulos bacterianos, como o LPS,
estimula a ativação do NF-KB, resultando em aumento da produção de

11
mediadores inflamatórios responsáveis pelo recrutamento e ativação de
células no local da infecção. A produção de NO por estas células leva à
inibição da ativação do NF-K8, diminuindo assim a expressão dos genes das
citocinas, moléculas de adesão, e da própria iNOS. Este mecanismo está em
acordo com a necessidade do hospedeiro em produzir uma resposta rápida à
infecção, porém esta resposta deve ser suprimida a tempo de minimizar o
dano ao próprio hospedeiro (CONNELLY e col., 2001).
1.3 - Macrófago: ponte entre a imunidade inata e a adaptativa
Além da sua função efetora, os macrófagos também participam da
coordenação da resposta imune, através da produção de citocinas e
quimiocinas, e apresentação de antígenos às células T.
A ativação dos linfócitos T é descrita como resultado de várias
interações entre células apresentadoras de antígenos e células T. O MHC
proporciona interações moleculares entre o peptídeo processado e o receptor
da célula T (TCR) (UNDERHILL e col., 1999, PAULNOCK, 2000).
As APCs, após fagocitose e processamento enzimático, expõem o
peptídeo bacteriano associado ao MHCII, proporcionando assim o
reconhecimento antigênico pelos linfócitos TCD4T
• Os linfócitos TCDS+
reconhecem os antígenos sintetizados pela própria célula hospedeira, como
nos casos de infecções virais, associados ao MHCI (PAULNOCK, 2000).
Entretanto, só a ligação do MHC ao TCR não é suficiente para estimular
o linfócito. É necessário um segundo sinal, fornecido tanto por fatores solúveis,
como a IL-2, quanto pelas interações entre as moléculas coestimulatórias,

12
expressas na superfície de linfócitos e APCs. Uma das principais interações
coestimulatórias descritas envolve a molécula C028 e a família B7 de ligantes
(MEOZHITOV & JANEWAY, 1997; ALBA SOTO E COL., 2003).
1.3.1 - 87-1 e 87-2
As glicoproteínas B7-1 (C080) e B7-2 (C086) pertencem à superfamília
das imunoglobulinas e estão presentes em células B, macrófagos e células
dendríticas. Elas se ligam a C028, cuja expressão na superfície das células T
é constitutiva, mas aumenta após ativação; e CTLA-4, expresso apenas após
ativação da célula T (FREEMAN e coL, 1993; LENSCHOW E COL., 1996).
As interações de C080 e C086 com C028 são essenciais para iniciar a
produção de citocinas, promover a expansão e diferenciação dos clones
específicos de células T, e determinar a polarização da resposta. Além disso, a
sinalização via C028 parece estar envolvida na prevenção da apoptose e na
manutenção dos níveis de proliferação (FREEMAN e coL, 1993; LENSCHOW
E COL., 1996; BOULOUGOURIS e coL, 1998).
Por outro lado, a ligação de CTLA-4 com os membros da família B7
parece ter efeitos de modulação negativa da ativação dos linfócitos T. A
ativação linfocitária resultante da interação MHCfTCR e C028/B7 resulta em
aumento da expressão de CTLA-4 que, ao se ligar a C080 e C086, transmite
sinais inativadores aos linfócitos T, num mecanismo auto-regulador
(LENSCHOW E COL., 1996; BOULOUGOURIS e coL, 1998).

13
A regulação da expressão de C080 e C086 é controlada por interações
célula-célula e também pela presença de citocinas no microambiente. A
produção de IFN-y está relacionada a um aumento na expressão destas
moléculas, enquanto a IL-10, por outro lado, inibe a expressão de C080 e
C086 (LENSCHOW E COL., 1996;).
1.3.2 - Geração da resposta adaptativa
O reconhecimento do MHCII pelo TCR gera respostas donais aos
patógenos, influenciadas por sinais recebidos pelas interações das moléculas
coestimulatórias. Na presença do sinal, ocorre a ativação, caracterizada pela
expansão clonal dos linfócitos T específicos ao antígeno, e aumento na
secreção de citocinas e expressão de receptores para citocinas. Na ausência,
não ocorre resposta e a célula T se torna tolerante ou anérgica (MEOZHITOV
& JANEWAY, 1997; UNOERHILL e coL, 1999).
As células efetoras da imunidade inata, como os macrófagos,
reconhecem os patógenos de uma maneira não-clonal pela ligação dos PRRs
aos PAMs, e assim induzem respostas efetoras apropriadas, discriminando o
self do non-self e também diferenciando os diversos tipos de patógenos. Os
linfócitos "naive", por outro lado, são pluripotentes, e não são especializados
em nenhum tipo de resposta em particular antes da ativação. Estas células vão
se tornar células efetoras distintas, dependendo dos sinais recebidos durante a
ativação, e estes sinais provêm das células efetoras do sistema imune inato
(MEOZHITOV &JANEWAY, 1997; MEOZHITOV & JANEWAY, 2000).

14
As citocinas que são produzidas nos primeiros estágios da resposta
inflamatória, pelas células T ativadas determinam o padrão da resposta
adaptativa. Citocinas do tipo 1 (IFN-y e IL-12) induzem a diferenciação de
células T em células Th1, enquanto as citocinas do tipo 2 (IL-10, IL-4),
induzem a via de diferenciação do tipo Th2. Portanto, a polarização da
resposta inflamatória depende da concentração das citocinas no
microambiente, das moléculas acessórias envolvidas na interação da APC
com o linfócito T e da concentração e natureza do antígeno (MEDZHITOV &
JANEWAY, 1997; PAULNOCK, 2000).
1.4 - O macrófago e os enteropatógenos
A mucosa intestinal possui uma série de folículos linfóides, isolados ou
em agregados (placas de Peyer) , responsáveis pelo reconhecimento de
microrganismos como comensais ou patógenos e também pela indução da
resposta imunológica. Para que os microrganismos estimulem a resposta
imune, eles devem ser transportados através da barreira epitelial até estes
folículos Iinfóides. O transporte é possível graças à existência das células M no
epitélio associado ao folículo (FAE). Este tipo celular especializado não possui
microvilosidades na sua superfície, mas sim domínios por onde ocorre a
endocitose de partículas e microrganismos. A membrana basolateral destas
células é extremamente invaginada, formando um bolso que contém linfócitos
T, linfócitos B, macrófagos e células dendríticas. Esta estrutura permite que as
superfícies apicais e basolaterais fiquem muito próximas, diminuindo a
distância que as vesículas endocíticas precisam percorrer para atravessar a

15
barreira epitelial. Assim, os antígenos endocitados são rapidamente
transportados até o interior do bolso, onde são fagocitados e apresentados aos
linfócitos B, resultando na produção de anticorpos IgA, que auxiliam na
regulação da microbiota endógena (NEUTRA e col.,1996; SANSONETII,
2004).
A colonização do trato gastrointestinal é benéfica para o hospedeiro,
pois a microbiota exerce uma função protetora, competindo por nutrientes e
receptores com os enteropatógenos; sintetiza vitaminas; e influencia no
desenvolvimento e maturação do sistema imune da mucosa.
A interação entre a microbiota e o sistema imune do hospedeiro é
complexa e baseia-se em um sistema de apresentação microbiana que resulta
em um estado de "inflamação fisiológica", caracterizado por tolerância às
bactérias comensais e mediado pela ação da IgA e do muco (SANSONETII,
2004; SANSONETII & DI SANTO, 2007).
Uma série de enteropatógenos utiliza as células M como rota de invasão
da mucosa intestinal. Ao se translocarem através destas células, no entanto,
são fagocitados por macrófagos residentes. Conseqüentemente, necessitam
de mecanismos de escape e sobrevivência para que a infecção seja
estabelecida com sucesso (NEUTRA e col., 1996; MARTINO e coI. , 2005).
A apoptose é um tipo de morte celular que ocorre em processos
fisiológicos, mas também em estados patológicos. Um dos mecanismos pelos
quais a célula entra em apoptose é pela ativação de uma cascata de proteases
intracelulares, denominadas caspases. Estas enzimas c1ivam muitas proteínas
celulares e ativam proteoliticamente outras caspases, levando a uma

16
amplificação irreversível do processo apoptótico (NAVARRE & ZYCHLlNSKY,
2000; NONAKA e coL, 2003).
Uma série de microrganismos utiliza a apoptose das células fagocíticas
como estratégia de sobrevivência. Macrófagos infectados por algumas
bactérias intracelulares facultativas, como Salmonella, Shigella, Yersinia e
algumas categorias de Escherichia coli sofrem apoptose (ZYCHLlNSKI e coL,
1996; HERSH e coL, 1998; FERNANOEZ-PRAOA e coL, 1998 MARTINO e coL,
2005).
Y. enterocolitica e Y. pseudotuberculosis penetram no epitélio intestinal
via células M, e, ao encontrar os macrófagos residentes, secretam proteínas
através de um sistema de secreção do tipo três (SSTT). Estas proteínas
inativam o citoesqueleto de actina dos macrófagos, impedindo a fagocitose da
bactéria, e inibem a produção de TNF-a, essencial para o controle da infecção.
Por outro lado, as bactérias que foram fagocitadas escapam do macrófago
induzindo sua apoptose (OONNEMBERG, 2000; SANSONETTI & DI SANTO,
2007).
Salmonella, no entanto, apresenta outro padrão de interação, também
baseado na secreção de efetores por um SSTT. Após ser fagocitada pelos
macrófagos residentes, esta bactéria tanto pode causar apoptose quanto
remodelar o vacúolo fagocítico, sobrevivendo e multiplicando-se em seu
interior (OONNEMBERG, 2000; SANSONETTI & DI SANTO, 2007).

17
1.5 - Disenteria bacilar
A disenteria bacilar é uma doença inflamatória intestinal aguda que tem
como agentes principais Shigella sp. e Escherichia coli enteroinvasora (EIEC).
Estas bactérias invadem o epitélio do cólon de humanos, causando a
inflamação aguda que caracteriza a doença (SANSONETTI, 2001; PARSOT,
2005). As manifestações clínicas mais comuns são febre, cólicas intestinais e
emissão de fezes com sangue e mucopurulentas. A análise de biópsias do
cólon de pacientes com shigelose mostra um infiltrado de células inflamatórias
na camada epitelial, edema tissular e áreas destruídas do epitélio intestinal
(MARTINO e coL, 2005).
A disenteria bacilar é endêmica através do mundo, mas
aproximadamente 99% dos 160 milhões de casos anuais ocorrem em países
subdesenvolvidos, onde as condições sanitárias e sócio-econômicas são
desfavoráveis. O número de mortes anuais por disenteria bacilar é de
aproximadamente um milhão, ocorrendo principalmente em crianças com
idades entre um e cinco anos (KOTLOFF e coL, 1999; SANSONETTI, 2001).
Recentemente, foi divulgado um estudo sobre shigelose realizado em 6
países asiáticos: China, Vietnã, Tailândia, Bangladesh, Paquistão e Indonésia.
Os dados mostram que a incidência da disenteria bacilar permanece alta (40
indivíduos em 1000, por ano, em todas as faixas etárias, e 254 em 1000, por
ano, no grupo de crianças com menos de cinco anos). Porém, em desacordo
com estudos divulgados anteriormente, as taxas de mortalidade foram
menores. Contribuem para este fato as melhoras nas condições alimentares, o

18
acesso mais rápido aos primeiros cuidados, e a maior utilização de
antibióticos. (VON SEIDLEIN e coL, 2006, SANSONETII, 2006).
1.6 - Shigella spp.
O gênero Shigella é constituído de quatro espécies: S. boydii, S.
dysenteriae, S. flexneri e S. sonnei. A maioria dos estudos sobre
patogenicidade destes microrganismos foi realizada em S. flexneri, mas os
resultados podem ser aplicados às outras espécies e a EIEC. (PARSOT,
2005).
Uma das principais características da patogênese das infecções por
Shigella e EIEC é a capacidade de invasão, ou seja, a capacidade de induzir
sua própria fagocitose por células que normalmente não são fagocíticas, como
as células do epitélio intestinal (COSSART & SANSONETII, 2004). O
responsável por este fenótipo invasor é o plasmídio de virulência (PV-220 Kb).
Células sem o PV são avirulentas e não penetram em células epiteliais
(SANSONETII et ai., 1982a; SANSONETII, 2001; PARSOT, 2005).
Uma região de aproximadamente 30 kb do PV é essencial para a
entrada da bactéria na célula. Esta região alberga componentes de um SSTT,
translocons, efetores e ativadores de transcrição. Os genes do locus mxi-spa
são responsáveis pela síntese de elementos do SSTI, que é ativado através
do contato da bactéria com as células epiteliais. Este contato promove a
inserção do translocon (formado pelas Ipas B e C) na membrana da célula
eucariótica e secreção dos efetores para o interior da célula do hospedeiro.
Entre estes efetores estão os antígenos plasmidiais de invasão IpaA, IpaB ,

19
IpaC e IpaO , essenciais na internalização da bactéria e escape do fagossomo
(COSSART & SANSONETII, 2004; PARSOT, 2005).
O processo de invasão é complexo e ocorre em várias etapas. Shigella
inicialmente adere à superfície da célula epitelial, provavelmente pela ligação
da IpaB com o receptor C044. Esse contato é seguido da ativação do SSTI,
que promove a entrada dos efetores bacterianos no citoplasma da célula.
Ocorre, então, a reorganização dos filamentos de actina no sítio de ligação da
bactéria com a célula, levando à formação de um "bolso macropinocítico". O
evento seguinte é o fechamento deste bolso,_resultado da despolimerização da
actina (OONNENBERG, 2000; COSSART & SANSONETII, 2004). Uma vez
dentro da célula, o microrganismo escapa do vacúolo e ganha acesso ao
citoplasma, onde ocorre a multiplicação. O movimento bacteriano leva à
formação de protrusões, que penetram nas células adjacentes. O ciclo da
infecção é então completado pela lise da dupla membrana formada pela
protrusão, a fim de liberar a bactéria no citoplasma da célula vizinha. (PARSOT
& SANSONETII, 1995; COSSART & SANSONETII, 2004).
1.6.1 - Geração da resposta imune na infecção por Shigella
Ensaios envolvendo monocamadas celulares demonstraram que S.
f1exneri invade células epiteliais apenas por seu pólo basolateral, sendo
incapaz de fazê-lo pelo pólo apical. Para promover a infecção, portanto, a
bactéria necessitaria de outra rota de invasão. Utilizando-se alça ligada de
coelho, observou-se que cepas de Shigella translocam-se através das células
M, sendo esta então sua via preferencial de entrada no epitélio intestinal

20
(COSSART & SANSONETII, 2004; SANSONETII & DI SANTO, 2007). A
fagocitose da bactéria pelos macrófagos residentes resulta em apoptose como
resultado da ligação da IpaB com a caspase-1. Além disso, ocorre liberação de
IL-1 e IL-18 como resultado da c1ivagem de seus precursores pela caspase-1
(figura1) (ZYCHLlNSKI et aI., 1996; GUICHON et aI., 2001; SANSONETII,
2005; MARTINO e col., 2005; SANSONETII, 2007).
Translocation through M cel!
M cel! Epithelial cells
Paracellular entry
- Macrophage/Dendritic cel!apoptosis ~""=-'"'1i
- Initiation of inflammation
IL-8 (<.-.;~~acrOPhageslLymPhocytes~\..--' IpaBCA +-.~ 'Dendritic cells type 111 secretion
Polymorpho~uclear '" IL-1~ / O OLeukocyte -
Figura 1 - Representação esquemática da patogênese da infecção por
Shigella (COSSART & SANSONETTI, 2004).
Diversos estudos evidenciam o papel de uma série de citocinas no
estabelecimento, manutenção e resolução da infecção por Shigella.

21
A síntese de IL-1 tem papel fundamental na disseminação bacteriana e
na forte resposta inflamatória característica da infecção por Shigella. IL-1
juntamente com IL-8 são responsáveis pelo recrutamento de células do
sistema imune inato, como monócitos e principalmente neutrófilos
polimorfonucleares (PMN). A migração destas células para o local da infecção
causa desestabilização do epitélio intestinal, permitindo assim a passagem de
um maior número de microrganismos que irão invadir os enterócitos, levando à
destruição tecidual. (DONNENBERG, 2000; SANSONETII, 2001; MARTINO e
col., 2005).
Além da IL-1, a ligação da IpaB com a çaspase-1 também é responsável
pela clivagem e liberação de IL-18. Utilizando camundongos nocauteados no
gene da caspase 1 (casp-rf) SANSONETII e col. (2000) demonstraram que
IL-1J3 é responsável pela intensidade da inflamação aguda, enquanto IL-18 é
necessária para o controle da infecção. Esta citocina contribuiria para a
erradicação da bactéria, provavelmente pelo estímulo à síntese de IFN-y, que
ativa uma resposta do tipo Th1.
RAQIB e col. (1995) observaram a produção de IL-1a, IL-1J3, TNF-a, IL
6, IL-4, IL-8, IL-10, IFN-y e TGF-J3 em biópsia retaI de pacientes durante o
estágio agudo de shigelose. Além disso, foi possível observar um número
muito maior de células produzindo TNF-a, IL-6 e IFN-y na colite severa em
relação à moderada.
WAY e col. (1998) avaliaram a suscetibilidade para infecção pulmonar
por Shigella em diferentes linhagens de camundongos com deleções em
aspectos específicos do sistema imune. Foi observado um aumento de cinco

22
vezes na magnitude da susceptibilidade à infecção por Shigella em
camundongos nocauteados no gene do IFN-y (GKO -f) em relação aos
selvagens. Além disso, as manifestações inflamatórias foram bem mais
severas nos GKO -r, o que permite concluir que a citocina IFN-y tem grande
importância na proteção da infecção por este microrganismo. Foi verificado
também que animais com deficiência em células NK apresentaram uma
susceptibilidade maior à infecção, e os autores sugerem que estas células
provavelmente são as fontes do IFN-y produzido durante a infecção.
O papel do IFN-y no controle da infecção por S. flexneri já foi
demonstrado 0JVAY e coL, 1998; LE-BARILLEC e coL, 2005). Um dos
mediadores da ação desta citocina é o óxido nítrico. Porém, não foram
observadas diferenças na dose letal de S. flexneri entre camundogos tratados
e não-tratados com aminoguanidina (inibidor da iNOS), e também não houve
diferença na dose letal entre camundongos selvagens e nocauteados no gene
da iNOS (NOS2-f). Estes resultados permitem concluir que o "killing" de S.
flexneri em células infectadas in vitro e a resistência dos camundongos são
processos independentes de NO (WAY & GOLDBERG, 1998).
1.6.2 - Participação dos linfócitos T na patogênese da shigelose
Durante muito tempo acreditou-se que somente a resposta humoral
estava envolvida na defesa contra Shigella. Posteriormente, foi demonstrada a
participação dos linfócitos T na resposta inflamatória a este microrganismo.
Uma das primeiras observações foi a de que indivíduos com shigelose
apresentaram um aumento no número de células TCD4+ e TCD8+ (AZIM e

23
coI. ,1996). ISLAM e col. (1995) demonstraram que indivíduos com shigelose
apresentaram um aumento no número de linfócitos, bem como aumento da
expressão de certas moléculas de superfície. Foi observado, por exemplo, um
aumento inicial da expressão de CD25 (considerado um marcador inicial de
ativação de células T), seguido de um aumento de HLA-DR, marcador de
ativação tardia, o que confirma o envolvimento de uma resposta T específica
na infecção por Shígella.
Em estudo conduzido por LE-BARILLEC e col. (2005) observou-se que
camundongos RagOyO (deficientes em linfócitos B, Te NK) foram incapazes de
controlar a infecção, enquanto camundongos reconstituídos com linfócitos T a13
e camundongos RagO (deficientes em linfócitos B e T, mas possuidores de
linfócitos NK) controlaram a infecção de maneira similar aos camundongos
selvagens. O controle da infecção mostrou ser dependente da produção de
IFN-y, e esta produção ocorreu de maneira redundante, ou seja, foi produzida
independentemente por linfócitos T a13 e NK. Estes resultados indicam
claramente que os linfócitos desempenham um papel fundamental na proteção
de camundongos na infecção pulmonar primária por Shígella f1exnerí.
1.7 - Escherichia coli enteroinvasora
Na década de 60 observou-se que várias amostras de E. colí
compartilhavam com bactérias do gênero Shígella a capacidade de invadir e se
proliferar no interior de células epiteliais, e de induzir ceratonjuntivite ao serem
inoculadas em olho de cobaia, ensaio conhecido como teste de Serény
(SERÉNY, 1963; TRABULSI e coI. , 1965). Em vários casos, a disenteria

24
causada por essas amostras foi semelhante à disenteria provocada por
Shigella spp. (TRABULSI e coL, 1965), observações que foram confirmadas
posteriormente por DuPONT e coL (1971), em ensaios com voluntários. Essa
nova classe de enteropatógeno foi denominada de E colí enteroinvasora
(EIEC).
A primeira descrição de EIEC foi feita por EWING e GRAWATII (1947).
Uma amostra de Ecoli foi isolada a partir de fezes de soldados americanos
com diarréia, durante a segunda guerra mundial, e estudos posteriores
realizados por SERÉNY (1963) demonstraram se tratar de uma EIEC. No
Brasil, a primeira descrição foi feita por TRABULSI e coL (1965), que isolaram
das fezes de um paciente com enterite aguda uma amostra de Ecolí capaz de
produzir ceratoconjuntivite em cobaia.
As cepas de EIEC possuem características bioquímicas e genéticas
praticamente idêntica às de Shigella. Além disso, o mecanismo de invasão e
os sintomas da doença são semelhantes. Estudos com voluntários humanos
mostraram, contudo, que a dose infectante de EIEC é muito maior do que a de
Shigella. De fato, para que ocorra um processo infeccioso é necessário um
inóculo de 10 a 100 microrganismos para Shigella, enquanto para EIEC o
inóculo é de 106 (DuPONT e coL, 1971).
Uma possibilidade para explicar essas diferenças seria uma capacidade
maior de invasão por amostras de Shigella. Em testes de Serény realizados
em nosso laboratório pudemos observar que o desenvolvimento de
ceratoconjuntivite em cobaias inoculadas com Shigella ocorre após 48 horas,
enquanto que em cobaias inoculadas com EIEC são necessários de quatro a

25
cinco dias. No entanto, demonstrou-se que não há diferenças importantes
entre os PV das duas espécies (fORMAL e coL, 1983; LAN e coL, 2001). Além
disso, GIBOTII e coL (2004) observaram que os genes ipaA, ipaB, ipaC e ipaD
não possuem alterações que possam explicar as diferenças na dose infectante
e na severidade de sintomas da disenteria causada por EIEC e Shigella.
Devido à maior incidência, os estudos ficaram praticamente restritos a
amostras de Shigella. Além do processo de invasão, pouco se sabe acerca
das alterações provocadas no hospedeiro na infecção por EIEC. Nossa
proposta durante o presente trabalho é a de buscar uma maior compreensão
sobre a interação entre EIEC e o hospedeiro. Para tanto, foi escolhido o
modelo murino de infecção, onde foram avaliadas: (i) a fagocitose, escape
bacteriano e morte dos macrófagos; (ii) a ativação dos macrófagos, através da
produção de citocinas e NO; (ii) a ativação linfocitária, através da medida da
proliferação e produção de citocinas; e (iv) o influxo celular e produção de
citocinas no peritônio de animais após infecção. Esperamos, com os
resultados obtidos, ter contribuído para elucidar algumas diferenças de
patogenicidade entre Shigella e EIEC, e os resultados desse estudo com
certeza são uma ferramenta para a compreensão do mecanismo patogênico
de EIEC.

26
2 - OBJETIVOS
2.1 - Objetivo geral
Avaliação das alterações induzidas por EIEC no hospedeiro durante o
curso da inflamação, utilizando-se de modelos murinos in vivo e in vitro.
2.2 - Objetivos Específicos
• Determinar a cinética de fagocitose, morte celular e produção de citocinas
por macrófagos da linhagem J774 infectados com EIEC;
• Avaliar a produção de óxido nítrico e citocinas por macrófagos peritoniais
de camundongo, após infecção com EIEC;
• Caracterizar o influxo celular e a produção de citocinas pelas células
peritoniais de camundongos infectados com EIEC;
• Avaliar a proliferação e produção de citocinas por linfócitos T em resposta
ao estímulo com antígenos de EIEC;
• Observar diferenças na resposta do hospedeiro após desafio com Shigella
e EIEC que possam explicar o maior inóculo requerido para causar infecção e
os sintomas mais brandos da infecção por EIEC.

27
3 - MATERIAIS E MÉTODOS
3.1 - Amostras Bacterianas
Foram utilizadas Shigella f1exneri M90T 5, cedida pelo Or. Philippe
Sansonetti, do Instituto Pasteur, e Escherichia coli enteroinvasora do sorogrupo
0124:H- (EIEC FBC 124/13), isolada de fezes diarréicas. As cepas foram
submetidas a teste de Serény (SERÉNY, 1963) para confirmação de sua
capacidade invasora. Nos ensaios de análise da transcrição dos genes de
citocinas por RT-PCR também foi utilizada Ecoli K12 (OH5a).
3.2 - Padronização do inóculo bacteriano
Para todos os ensaios de infecção descritos a seguir, as amostras
invasoras (Shigella e EIEC) foram semeadas em ágar vermelho congo (TSA +
0,2% de corante vermelho congo) e incubadas à 37°C por 48h, enquanto a
amostra não invasora (Ecoli K12) foi semeada em ágar MacConkey e
incubada à 37°C por 24h. A coloração vermelha das colônias de EIEC e
Shigella crescidas em ágar vermelho congo é resultado da precipitação do
corante pelas proteínas secretadas pelas bactérias. Esta propriedade é
utilizada para distinguir entre amostras invasoras (cujas proteínas se coram
com o vermelho congo ou colônias vc+) e não invasoras (incapazes de se corar
com o vermelho congo ou colônias vc} O fenótipo de precipitação do vermelho
congo atua como um marcador de virulência (MAURELLI e coL, 1984). Foram
selecionadas colônias vermelho-congo positivo e colônias de Ecoli K12
provenientes do ágar MacConkey para semeadura em caldo TSB. As amostras
foram incubadas em agitador (220 rpm, 37°C) por aproximadamente quatro

28
horas para obtenção da fase logarítmica, centrifugadas (1500 rpm, por cinco
minutos) e ressuspensas em solução fisiológica. A concentração bacteriana na
suspensão foi ajustada em espectrofotômetro, sendo de 108 células por
mililitro. A cada ensaio realizado, o número de bactérias do inóculo foi
confirmado através da semeadura de diluições da suspensão em ágar
MacConkey.
3.3 - Cultura celular
Macrófagos murinos da linhagem J774 1.6 foram cultivados em meio
RPMI (Cultilab), suplementado com 5% de Soro Fetal Bovino (R5) e penicilina
estreptomicina (0,1 g de estreptomicina e 100.000 UI de penicilina por litro). A
cultura celular foi mantida em estufa a 37°C, com atmosfera de 5% de CO2 .
3.4 - Obtenção dos macrófagos de camundongo
Para obtenção dos macrófagos utilizados nos ensaios "in vitro"
camundongos C57BU6 e C57BU6 nocauteados no gene da NO sintase
induzível (iNOS KO) foram sacrificados e submetidos a lavado peritonial com
PBS estéril gelado. A suspensão celular foi transferida para um tubo Falcon e o
número de células foi determinado através de contagem em Câmara de
Neubauer com o corante vital azul de tripan (0,1% em PBS), na proporção de
1:1. A seguir, a suspensão foi centrifugada por cinco minutos a 1.500 rpm. O
sobrenadante foi descartado e os macrófagos ressuspensos em meio R5 com
penicilina - estreptomicina. Alíquotas contendo 3x105 células foram distribuídas
em placa de 96 orifícios, que foi incubada por uma hora a 37°C, na presença

29
de 5% de CO2, possibilitando a aderência dos macrófagos. O meio de cultura
foi retirado, eliminando-se assim as células não aderentes, os orifícios foram
lavados com PBS, e foi adicionado R5 sem antibióticos. Estas células foram
utilizadas nos ensaios de produção de óxido nítrico e análise da transcrição
dos genes de citocinas por RT-PCR.
3.5 - Extração de RNA e síntese de DNA complementar
A extração do RNA total foi feita utilizando-se o RNeasy Mini Kit
(Quiagen), de acordo com as recomendações do fabricante. As amostras
foram submetidas à eletroforese em gel de agarose a 1,5% para análise da(
integridade do RNA. O RNA foi quantificado em espectrofotômetro a 260 e 280
nm e diluído para uma concentração final de 10ng/J.!L para a síntese de DNA
complementar (cDNA). Para a síntese de cDNA foi utilizado o kit Supercript
First-Strand Synthesis System for RT-PCR (Invitrogen). Como iniciador utilizou-
se oligo (Dt), que se hibridiza com a cauda de poly(A) encontrada na maioria
do RNA de eucariotos.
3.6 - peR em tempo real
O cDNA obtido conforme o item anterior foi amplificado por PCR em
tempo real, utilizando-se o sistema TaqMan. Este sistema baseia-se na
utilização de uma sonda (TaqMan probe) duplamente marcada, com um
fluorocromo "reporter" e um "quencher". A sonda é desenhada de modo a se
anelar com a sequência alvo, entre os iniciadores "forward" e "reverse". Na
forma inativada, a fluorescência do "reporter" é absorvida pelo "quencher".

30BIBLIOTECA
Faculdade de Ciências FarmacêuticasUniversidade de São Paulo
Durante a fase de extensão da PCR, a sonda é degradada pela atividade de
exonuclease da Taq DNA polimerase, separando os fluorocromos "quencher" e
"reporter". A quantidade de fluorescência liberada pelo "reporter" é diretamente
proporcional à quantidade de DNA gerada em cada ciclo da PCR
(OVERBERGH e coI. , 2003). A reação foi realizada em um equipamento ABI
Prism 7000 Sequence Detection System versão 1.6 (Applied Biosystems). Os
parâmetros da reação foram: 50°C /2 min, 95°C /10 min, 50 ciclos de 95° C/15
seg e 60°C /1 mino Foram adicionados 3 ~g of cDNA a 12.5 ~I de 2x TaqMan
PCR Master mix (Applied Biosystems), 100 nM de sonda fluorescente
específica (Applied Biosystems) e água livre de nucleases em um volume final
de 251..1L. Em cada reação realizada, foi utilizada a-actina como controle
endógeno. Os primers e sondas utlizados estão descritos na tabela 1.
Os dados foram normatizados para os níveis de a-actina e analisados
pelo método da curva padrão. Foi utilizado o método da comparação relativa
para quantificar os níveis de transcrição dos genes das citocinas.
Gene Primer 5' (5'-3') Sonda (5'FAM) Primer 3' (5'-3')
TNF-a CACAAGATGCTGGGACAGTGA ATGCACCACCATCAAGG CAGTGAATTCGGAAAGCCCATTTG
IL-1 GCAGGATTTTCTAGGTGGTCAGTTA CCTCTAGAGCACCATGCTAC ~GCTTCGTGGCTGTGGAAA
IL-10 CCTGTGTTTAAGCTGTTTCCATTGG CCCCTCCCATCATATAATATA AGCTTCAATTGCTTCCCAAGGA
a-actina CCCTGAAGTACCCCATTGAACATG CCAGATCTTCTCCATGTCGC ACGCAGCTCATTGTAGAAGGT
Tabela 3.1 - Primers e sondas utilizados no real-time PCR
3.7 - RT-peR semiquantitativo
A análise da transcrição dos genes das citocinas por RT-PCR
semiquantitativo foi avaliado em ensaios "in vitro" e "in vivo". O cDNA obtido de

31
acordo com o item 3.5 foi amplificado por reação de PCR. Os reagentes foram
utilizados nas seguintes concentrações: 1,5 mM de MgCh, 150 !-lM de DNTPs,
O,4!-lM de cada iniciador, 1,5 U de Taq DNA polimerase, tampão da enzima 1x
concentrado, 1 !-lI de cDNA e água miliQ estéril num volume final de 20!-lL.
As condições da PCR foram as seguintes: Etapa incial de desnaturação
por 5 minutos a 94°C, 30 ciclos com desnaturação por 45 segundos a 94°C,
anelamento por 45 segundos em temperatura adequada a cada par de
iniciadores e extensão por 1 minuto a 72°C, seguida de extensão final por 7
minutos a 72°C.
Os iniciadores utilizados estão descritos na tabela 2. Como controle
interno foi utilizado o gene da u-actina. Os produtos de PCR foram analisados
em gel de agarose, a 1,5% em tampão TBE. Os produtos da PCR foram
submetidos à separação eletroforética por 45 minutos a 100 volts. Em seguida,
o gel foi corado em solução contendo brometo de etídeo (O,5IJg/ml) e as
bandas visualizadas sob luz ultravioleta. As imagens foram adquiridas através
de um equipamento de fotodocumentação (Doc Print DP-001-FDC, Vilber
Lourmat, França). As imagens das bandas foram submetidas à análise
densitométrica com software apropriado (Scion Image). O nível de expressão
do gene de cada citocina foi calculado como a razão da intensidade da banda
do gene sobre a intensidade da banda da u- actina.

32
IIniciador ISeqüência
TNF-a F - 5'- CCTTIATTGTCTACTCCTCAG - 3'R - 5'- GGAAAGGTCTGAAGGTAGG - 3'
IL-1 F - 5'- CTCAAGAAGGAGAAGACT - 3'R - 5'- GATGGTTGGTGTIAGTIAC - 3'
IL-10 F - 5'-TACCTCAGTICCCATIC -3'R - 5'- GAAAGGACACCATAGC -3'
IL-18 F - 5'- ACTGTACAACCGCAGTAATACGG - 3'R - 5'- AGTGAACATIACAGATTTATCCC - 3'
INOs F - 5'- CATGGCTIGCCCCTGGAAGTTICTCT - 3'R - 5'- GCAGCATCCCCTCTGATGGTGCCATC - 3'
TLR4 F- 5'- GACCTCAGCTICAATGGTGC - 3'R - 5'-TATCAGAAATGCTACAGTGGATACC - 3'
IFN-y F- 5'- CTGTGATIGCGGGGTIG - 3'R - 5'- TTTCTITCAGGGACAGCCTG - 3'
IP-10 F- 5'- CGCGGATCCTCGCACCTCCACATAGCTTAC - 3'R - 5'- CGCGGATCCTGAGCAGAGATGTCTGAATC- 3'
a-actina F - 5'-TCCTICGTIGCCGGTCCACA - 3'R - 5'- CGTCTCCGGAGTCCATCACA - 3'
Tabela 3.2 - Iniciadores usados no RT-PCR semiquantitativo.
3.8 - "Enzyme-Iynked immunosorbent assay" (ELISA) para quantificação
das citocinas
A concentração das citocinas no sobrenadante de macrófagos foi
determinada por ELISA de captura (PEPROTECH), de acordo com as
recomendações do fabricante, a fim de confirmar que a transcrição dos genes
resultou em produção de proteína. Em resumo, placas de microtitulação de 96
poços foram sensibilizadas com o anticorpo primário diluído em PBS e
incubadas de um dia para o outro. Após lavagem com PBS acrescido de tween
20 a 0,05% as placas foram incubadas por uma hora com PBS com 1% de
albumina de soro bovino (BSA, SIGMA), para promover o bloqueio dos sítios
livres do plástico. As citocinas recombinantes utilizadas para realizar a curva
padrão foram diluídas em PBS contendo 1% de BSA. Após lavagem, a curva

33
padrão e as amostras foram incubadas por duas horas. As placas foram
lavadas e incubadas por duas horas com os anticorpos secundários
conjugados à biotina diluídos em PBS. Após nova lavagem, as placas foram
incubadas com conjugado estreptoavidina-peroxidase por 30 minutos. As
placas foram novamente lavadas e a revelação foi realizada com 0
fenilenodiamina (1 mg/mL) diluída em tampão citrato de sódio (0,1 M, pH 4,5)
por 15 a 30 minutos. Para o bloqueio da reação foi utilizado ácido sulfúrico 4N,
e a leitura das densidades ópticas foi realizada em leitor automático, em
comprimento de onda de 450 nm. As concentrações de cada citocina nas
amostras foram calculadas mediante equação de regressão linear com base na
curva padrão obtida com diluições das citocinas recombinantes. Todas as
incubações foram realizadas em temperatura ambiente.
3.9 - Interação entre EIEC e macrófagos J774 (ensaios "in vitro'1
3.9.1 - Sobrevivência bacteriana no interior dos macrófagos
Este ensaio foi realizado de acordo com a metodologia descrita por
FERNANDEZ-PRADA e col. (1997). Cerca de 24 horas antes do teste de
fagocitose, os macrófagos foram retirados dos frascos de cultura com a ajuda
de um espalhador Ccel! scraper"). A suspensão celular foi transferida para um
tubo Falcon e o número de células viáveis foi determinado através de
contagem em Câmara de Neubauer com o corante vital azul de tripan (0,1%
em PBS), na proporção de 1:1. A seguir, a suspensão foi centrifugada por
cinco minutos a 1.500 rpm. O sobrenadante foi descartado e os macrófagos

34
ressuspensos em meio R5 com penicilina - estreptomicina. As células foram
distribuídas em placas de cultura de 24 orifícios numa concentração de 106
células por orifício. Após 24 horas de incubação a 37°C em atmosfera úmida
com 5% de CO2 as placas foram centrifugadas (100 x g, por cinco minutos); o
meio foi retirado e os macrófagos lavados com PBS. Foi adicionado R5 sem
antibióticos e as monocamadas celulares foram infectadas com 107 bactérias.
As placas foram novamente centrifugadas para promover o contato bacteriano
com os macrófagos, e a seguir incubadas por zero, 30, 60 e 90 minutos. A
seguir, os macrófagos foram novamente lavados e incubados em meio R5
contendo gentamicina (50llg/ml), por 50 minutos, para matar bactérias
extracelulares e prevenir a reinfecção. O tempo zero foi determinado da
seguinte maneira: A placa foi centrifugada e os macrófagos foram
imediatamente lavados e incubados com gentamicina. A seguir, as células
foram novamente lavadas e lisadas com Triton X-100 a 10%. Diluições dos
lisados foram semeadas em ágar TSA e MacConkey (incubados a 37°C por
18-24 horas), para determinação das UFC.
3.9.2 - Escape bacteriano após fagocitose e determinação da viabilidade
dos macrófagos
Para avaliar o escape bacteriano após fagocitose, e se este escape era
acompanhado de perda de viabilidade dos macrófagos, foram feitas algumas
modificações no ensaio anterior. Os macrófagos foram infectados com Shigella
e EIEC na proporção 10 bactérias por macrófago por uma hora, e, a seguir,
incubados em meio R5 contendo gentamicina (50llg/ml), por 50 minutos. As

35
células foram então lavadas e incubadas em R5 sem antibióticos por 30, 60 e
120 minutos, e seis e oito horas. A seguir, alíquotas do sobrenadante foram
diluídas e semeadas em ágar TSA e MacConkey para determinação das UFC.
O meio foi retirado e os macrófagos foram ressuspensos em 300 J..l1 de tampão
HFS (0,1% de citrato de sódio, 0,1% de triton X - 100 e 50 J..lg/ml de iodeto de
propídio). A perda do DNA fragmentado dos núcleos apoptóticos foi
quantificada através de citometria de fluxo (FACScalibur, Beckton Dickinson).
Como controle negativo foram utilizadas monocamadas celulares não
infectadas, para determinação das UFC e viabilidade dos macrófagos.
3.9.3 - Produção de citocinas por macrófagos infectados
A transcrição dos genes de TNF-a, IL-1 e IL-10 por macrófagos J774 foi
analisada após infecção com Shigella M90T e EIEC FBC124/13. Macrófagos
J774 foram cultivados em placas de 96 orifícios na concentração de 105
células por orifício, por 24 horas, a 37°C, na presença de 5% de COzo As
células foram infectadas com suspensões de Shigella e EIEC contendo 105
bactérias, por uma hora. O meio foi substituído por RPMI acrescido de 5% de
soro fetal bovino e 50J..lg/ml de gentamicina. Após zero, duas e seis horas de
incubação o meio foi retirado e as células recolhidas para a extração do RNA,
realizada de acordo com o item 3.5. Após a síntese do cDNA (item 3.5), a
transcição dos genes das citocinas foi avaliada por PCR em tempo real (item
3.6). A transcrição basal dos genes das citocinas foi determinada através da
extração de RNA de macrófagos não infectados. As concentrações TNF-a, IL
1 e IL-10 foram determinadas no sobrenadante de monocamadas celulares

36
não-infectadas e infectadas (tempo de 7 horas), por ELISA de captura (item
3.8).
3.10 - Interação entre EIEC e macrófagos de camundongos C57BLl6 e
C57BLl6 iNOS KO (ensaios "in vitro'1
3.10.1 - Produção de óxido nítrico (NO)
Os macrófagos obtidos conforme o item 3.4 foram infectados com
suspensões bacterianas de Shigella ou EIEC contendo 104 e 105 bactérias e
incubados por 1 hora a 37°C, na presença de 5% de CO2 . Em seguida o meio
de cultura foi substituído por R5 com gentamicina (50/-lg/ml). Após 24 horas de
incubação, foram transferidos 50/-lL do sobrenadante de cada orifício para
outras placas, e a dosagem de N02- foi realizada pelo método de Griess. (Ding
e coL, 1988). Resumidamente, foi adicionado aos 50/-lL do sobrenadante 50/-lL
do reativo de Griess. A absorbância foi determinada em leitor de ELISA com
filtro de 550nm, contra branco constituído de meio de cultura e reativo de
Griess v/v. Os resultados foram expressos em /-lmoles de N02-, com base em
uma curva padrão feita com concentrações conhecidas de nitrato de sódio em
meio de cultura (5,10, 30 e 60 /-lmoles de N02- ). O limite de detecção do
método é de 5 /-lmoles de N02-.
3.10.2 - Transcrição dos genes das citocinas, da enzima iNOS e do TLR4
Os macrófagos de camundongos C57BU6 e C57BU6 iNOS KO obtidos
conforme o item 3.4 foram infectados com suspensões bacterianas de

37
Shigella, EIEC FBC 124/13 e E.coli K12 contendo 105 bactérias e incubados
por uma hora a 37°C, na presença de 5% de CO2 . Em seguida o meio de
cultura foi substituído por RPMI com 5% de soro fetal bovino e gentamicina
(50J.!g/ml). Após três horas de incubação, num total de quatro horas de
infecção, o meio foi retirado e as células recolhidas para a extração do RNA
(item 3.5). Também foi extraído RNA de macrófagos tratados por 4h com
10ng/ml de LPS de Escherichia coli (SIGMA) O cONA obtido conforme o item
3.5 foi utilizado para determinação da transcrição dos genes das citocinas IL-1,
TNF-a, IL-10 e IL-18, da enzima iNOS e do receptor transmebrânico "toll like
receptor 4" (TLR4) (item 3.7).
3.10.3 - Produção de TNF-a e IL-10
Para confirmação de que a transcrição gênica resultou em produção de
proteína, os sobrenadantes das culturas dos macrófagos não infectados e
infectados por quatro horas, (cuja transcrição gênica foi avaliada por RT-PCR
no item 3.10.2) foram recolhidos para determinação da concentração de TNF-a
e IL-10, por ELISA de captura (item 3.8).
3.11 - Interação entre EIEC e camundongos C57BLl6 e C57BLl6 iNOS KO
"in vivo"
3.11.1 - Determinação do influxo celular para o peritônio após infecção
Lotes de dois camundongos C57BLl6 foram inoculados
intraperitonialmente com suspensões de EIEC FBC124/13 ou Shigella M90T

38
contendo 107 bactérias. Após 12, 24 e 48 horas de infecção os animais foram
sacrificados e submetidos a lavado peritonial com meio RPMI gelado. O
número de células foi determinado através de contagem em Câmara de
Neubauer com azul de tripan e alíquotas contendo 106 células foram
transferidas para placas de 96 orifícios com fundo em U. Foi feita uma lavagem
com "Staining Buffer" (1 % de soro fetal bovino e 0,05% de azida em PBS). A
seguir foram adicionados os marcadores numa diluição de 1:100 em "Staining
Buffer". Após 30 minutos de incubação em gelo as amostras foram lavadas
com "Staining Buffer", centrifugadas (1300 rpm, cinco minutos), ressuspensas
em "Staining Buffer" e analisadas em citômetro de fluxo (FACScalibur; Becton
Oickinson). Todo o procedimento foi realizado em banho de gelo.
Os seguintes anticorpos monoclonais, com os respectivos fluorocromos
foram utilizados: anti-C04 (Cychrome) e anti-C08 (PE), marcadores de
linfócitos T, anti-B220 (Cychrome), marcador de linfócitos B, anti-F4/80
(FITC), marcador de macrófagos, anti-RB6 (PE), marcador de granulócitos, e
anti-MHCII (FITC), anti-C080 (FITC) e anti-C086 (FITC), moléculas
expressas na superfície de macrófagos. As células peritoniais receberam as
seguintes marcações: tripla marcação de C04 com C08 e F4/80; dupla
marcação de RB6 com F4/80, dupla marcação de F4/80 com B220, e dupla
marcação de F4/80 com MHCII, de F4/80 com C080 e de F4/80 com C086.

39
3.11.2 - Análise da transcrição dos genes das citocinas, da enzima iNOS e
do TLR4
Camundongos C57BU6 e C57BU6 iNOS Kü foram inoculados
intraperitonialmente com 107 bactérias. Após quatro e 48 horas de infecção os
animais foram sacrificados e submetidos a lavado peritoníal com meio RPMI
gelado. O número de células foi determinado através de contagem em Câmara
de Neubauer com azul de tripan. A extração do RNA das células totais do
peritônio e a obtenção do cDNA foram realizadas de acordo com o item 3.5 e o
RT-PCR semiquantitativo foi realizado de acordo com o item 3.7.
3.11.3 - Produção de TNF-a e IL-10
Para confirmação de que a expressão gênica resultou em produção de
proteína, a concentração de TNF-a e IL-10 foi determinada no lavado peritonial
dos animais não infectados, e após quatro e 48 horas de infecção, por ELISA
de captura (item 3.8).
3.11.4 - Determinação da mortalidade de camundongos C57BLl6 e
C57BLl6 iNOS KO após infecção
Camundongos C57BU6 e C57BU6 iNOS Kü foram inoculados
intraperitonialmente com suspensões de Shigella M90T, EIEC FBC124/13 e
E.coli K12 contendo 107 e 108 bactérias (item 3.2), a fim de determinar se a
ausência de produção de NO ocasionaria alterações na mortalidade dos
animais.

40
3.12 - Resposta de linfócitos à infecção por EIEC
3.12.1 - Ensaio de linfoproliferação
3.12.1.1 - Obtenção de linfócitos
Lotes de três camundongos Balb/C foram inoculados com suspensões
bacterianas de Shigella e EIEC na concentração de 106 bactérias por coxim
plantar. Sete dias após a inoculação os camundongos foram sacrificados e
seus linfonodos inguinais e poplíteos extraídos. Os Iinfonodos foram
macerados em placa, para dispersão das células, com meio RPMI. As células
foram centrifugadas (1500 rpm, por cinco minutos) e ressuspensas em RPMI.
Estas células foram utilizadas nos ensaios de linfoproliferação e determinação
de população de linfócitos.
3.12.1.2 - Obtenção de Antígenos Solúveis
Como agente indutor de proliferação linfocitária foram utilizados
antígenos solúveis, entre eles os Antígenos de Invasão Plasmidial (Ipas). Estes
antígenos foram obtidos pelo estímulo do contato bacteriano com células
epiteliais.
Células Hep-2 foram cultivadas em meio MEM, suplementado com 10%
de Soro Fetal Bovino e penicilina - estreptomicina, em garrafas de 75cm2. Após
obtenção da confluência celular o meio foi substituído por MEM com 10% de
soro fetal bovino sem antibióticos. Foram adicionados dois ml de suspensão
bacteriana de Shigella ou EIEC na fase logarítmica de crescimento. Após cinco

41
horas de incubação o meio foi recolhido e centrifugado (8000 rpm, 10 minutos).
Foram adicionados fenilmetilsulfonil fluoreto (PMSF; Sigma Aldrich) numa
concentração de 50 Ilg/ml, e EDTA (Sigma Aldrich) numa concentração de 0,5
11M, a fim de se inibir a atividade de proteases. O sobrenadante foi filtrado (filtro
Millipore, 0.22 IJm), e concentrado 10 vezes (Ultrafree 15, Millipore). A
concentração protéica nas amostras foi determinada pelo método de Bradford.
Para confirmação da presença de Ipas na amostra, foi realizada eletroforese
em gel de poliacrilamida (SDS-PAGE) a 10%, seguida de reação de
Westernblot. Utilizou -se anticorpos monoclonais anti - Ipa B, anti - Ipa C e anti
- Ipa O (título: 1/100) e conjugado antimouse IgG marcado com peroxidase
(Sigma), num título de 1:1000.
3.12.1.3 - Linfoproliferação
As células dos linfonodos foram centrifugadas (1500 rpm, 10 minutos),
ressuspensas em RPMI suplementado com 2,5% de soro humano e
distribuídas em placas de 96 orifícios, na concentração de 3x105 células por
orifício. Para obtenção de uma curva de proliferação adicionou-se aos orifícios
as seguintes concentrações de antígeno: 10, 20, 30, 40, 50, 60, 70, 80, 90 e
100 Ilg. Os linfócitos de animais inoculados com Shigella foram estimulados
com antígenos de Shigella e os inoculados com EIEC receberam antígenos de
EIEC. A placa foi mantida em estufa umidificada a 37 ac, em atmosfera de 5%
de CO2 , por 72 horas. Os controles positivos foram incubados na presença de
41lg de Concanavalina A (ConA, SIGMA).

42
Cerca de 16 horas antes de coletar as células foi adicionado à cultura
1!J.Ci por poço de metil-3H-timidina. As células foram coletadas com o auxílio de
um coletor automático (Cell Harvester, LKB Wallac 1295-001), e a
incorporação da timidina mensurada através da emissão de radiações beta
(Liquid Scintillation Counter, 1205 betaplate, Wallac).
Após determinação da curva de proliferação, foi realizado novo ensaio
de linfoproliferação. Os animais foram inoculados com Shigella, EIEC e E.coli
K12 para a obtenção de linfócitos (conforme descrito no item 3.12.1.1). Os
linfócitos foram então estimulados com os antígenos solúveis bacterianos na
concentração de 100 !J.g. Os linfócitos de animais inoculados com Shigella
foram estimulados com antígenos de Shigella, EIEC e K12, o mesmo
acontecendo com os linfócitos de animais inoculados com EIEC e K12. Os
controles positivos foram incubados na presença de 4!J.g fitohemaglutinina
(SIGMA).
3.12.2 - Produção de IFN-y, IL-10 e IL-4
Em ensaio paralelo à linfoproliferação, os sobrenadantes das células
dos linfonodos estimuladas com os antígenos solúveis bacterianos foram
recolhidos após 72 horas de incubação para determinação das concentrações
de IFN-y, IL-10 e IL-4 por ELISA de captura (item 3.8).
3.12.3 - Determinação das subpopulações de linfócitos
As células dos linfonodos foram centrifugadas (1500 rpm, 10 minutos) e
o sedimento ressuspenso em PBS Dulbeco com 5% de Soro Fetal Bovino.

43
Alíquotas contendo 5 x 105 células foram transferidas para tubos eppendorf
onde foram adicionados os marcadores, numa diluição de 1:100, incubando-se
em seguida por 30 minutos em gelo. A suspensão foi centrifugada (1000rpm,
dois minutos), e o sedimento ressuspenso em PBS Oulbeco com 5% de Soro
Fetal Bovino. As amostras foram analisadas em citômetro de fluxo
(FACScalibur Becton Oickinson).
As células receberam dupla marcação: anticorpos monoclonais anti-C03
(PE) e Anti-C04 (FITC), ou anti-C03 (PE) e anti-C08 (FITC).

44
4 - RESULTADOS
4.1 -Interação entre EIEC e macrófagos J774 (ensaios "in vitro"
4.1.1 - Determinação do número de bactérias viáveis no interior de
macrófagos J774 após fagocitose
Para comparar a viabilidade de EIEC e Shigella no interior dos
macrófagos após fagocitose foi feita a determinação das UFC em diferentes
intervalos de tempo. A gentamicina não penetra no macrófago, assim,
apenas as bactérias não fagocitadas são mortas pela ação do antibiótico.
Pode-se observar na figura 4.1. que não há diferenças na captura
inicial das duas bactérias pelos macrófagos (p=0,5). Após 30 minutos de
infecção, porém, há um aumento significativo de UFC de EIEC no interior
dos macrófagos, se comparado a Shigella (p=0,007). Nos tempos
subseqüentes há um decréscimo no número de UFC obtidas dos
macrófagos infectados com ambas as bactérias, sugerindo o escape
bacteriano do macrófago. Observa-se, porém, que o número de UFC de
EIEC recuperada do macrófago é sempre maior que o número de UFC de
Shigella (p=0,036 após uma hora de infecção e p=0,06 após 90 minutos).

12000 l
I10000 -
8000 -!
EU 6000u.=>
4000
2000 ~
O
O
*
0,5
Tempo (horas)
*
*
1,5
.M90T
• FBC124/13
45
FigA.1- Unidades Formadoras de Colônia de Shigella flexneri M90T e EIEC FBC124/13 em
lisados de macrófagos J774 após diferentes intervalos de infecção. Os dados representam
as médias e os desvios-padrão de infecções em triplicata e são representativos de dois
experimentos independentes. * - Intervalos de infecção onde a diferença no número de
UFC de Shigella e EIEC é estatisticamente significativa (p<O,05 segundo o teste t não
pareado).
4.1.2 - Determinação do escape bacteriano após fagocitose
Os resultados obtidos no experimento de escape bacteriano após
fagocitose em macrófagos J774 são mostrados na figura 4.2. De acordo
com o gráfico, o número de UFC de Shigella M90T no sobrenadante é maior
que o número de UFC de EIEC em todos os intervalos de infecção. Em 30
minutos (p=0,0013) e uma hora (p=O,006) de incubação em meio sem
gentamicina o número de UFC de Shigella no sobrenadante é praticamente
quatro vezes maior que o número de UFC de EIEC. Em duas horas o
número de UFC de Shigella é dez vezes maior que o número de UFC de
EIEC (p<0,0001) e, em seis horas, não houve diferenças significativas entre
o número de UFC das duas bactérias.

50000
45000
40000
35000 -
30000 -EÜ 25000u..::::> 20000
15000
10000 -
5000 -
O -
0.5
*
*
2
Tempo (horas)
6
.M90T
• FBC124/13
46
Fig.4.2- Unidades Formadoras de Colônia de Shigella f1exneri M90T e EIEC FBC124/13 no
sobrenadante de culturas de macrófagos J774 após diferentes intervalos de infecção. Os
dados representam médias e desvios padrão de infecções em duplicata e são
representativos de dois experimentos independentes. * - Intervalos de infecção onde a
diferença no número de UFC de Shigella e EIEC é estatisticamente significativa (p<0,05
segundo o teste t não pareado).
4.1.3 - Determinação da viabilidade dos macrófagos infectados
Para determinar se a infecção ocasionaria alterações na viabilidade
dos macrófagos foi utilizado o tampão HFS, que avalia o nível de
fragmentação do DNA celular. O número de macrófagos não viáveis em
monocamadas de células não-infectadas foi ao redor de 5% (dados não
mostrados). Em macrófagos infectados observou-se que o número de
macrófagos não viáveis é maior em monocamadas infectadas com ShigelJa
do que naquelas infectadas com EIEC (figura 4.3). Em duas horas de
incubação em meio sem gentamicina a porcentagem de macrófagos não
viáveis na infecção por Shigella é aproximadamente duas vezes maior do
que na infecção por EIEC (p=O,006). Em quatro horas a porcentagem de

47
macrófagos não viáveis é de 85% em monocamadas infectadas por
Shigel/a, e de 42% em monocamadas infectadas por EIEC (p=O,0003). No
tempo de seis horas observa-se 93% de macrófagos não viáveis na infecção
por Shigel/a, contra 75% de macrófagos não viáveis na infecção por EIEC
(p=O,02). Após oito horas de incubação em meio sem gentamkina o número
de macrófagos não viáveis é semelhante na infecção por Shigel/a (94%) e
.M90T
• FBC124/13
---,
864
*
2
*
1.5
EIEC (98%).
120
CIl
"Qj 100>
'l1l.;:I
o 801l1lc:CIlo 60Cl~'0L-U 40l1l
EQ)
"O 20~o
o -j----,--
0,5
Tempo (horas)
Fig. 4.3- Determinação de alterações na viabilidade dos macrófagos J774 em diferentes
intervalos de infecção por Shigella flexneri M90T e EI EC FBC124/13. Os dados
representam médias e desvios padrão de infecções em triplicata e são representativos de
três experimentos independentes. * - Intervalos onde a diferença na porcentagem de
macrófagos não-viáveis após infecção por Shigella e EIEC é estatisticamente significativa
(p<0,05 segundo o teste t não pareado).
4.1.4 - Cinética da transcrição dos genes de TNF-a, IL-1 e IL-10 por PCR
em tempo real
A transcrição dos genes das citocinas TNF-a,( IL-1 e IL-10 por
macrófagos J774 infectados e não infectados foi quantificada por PCR em
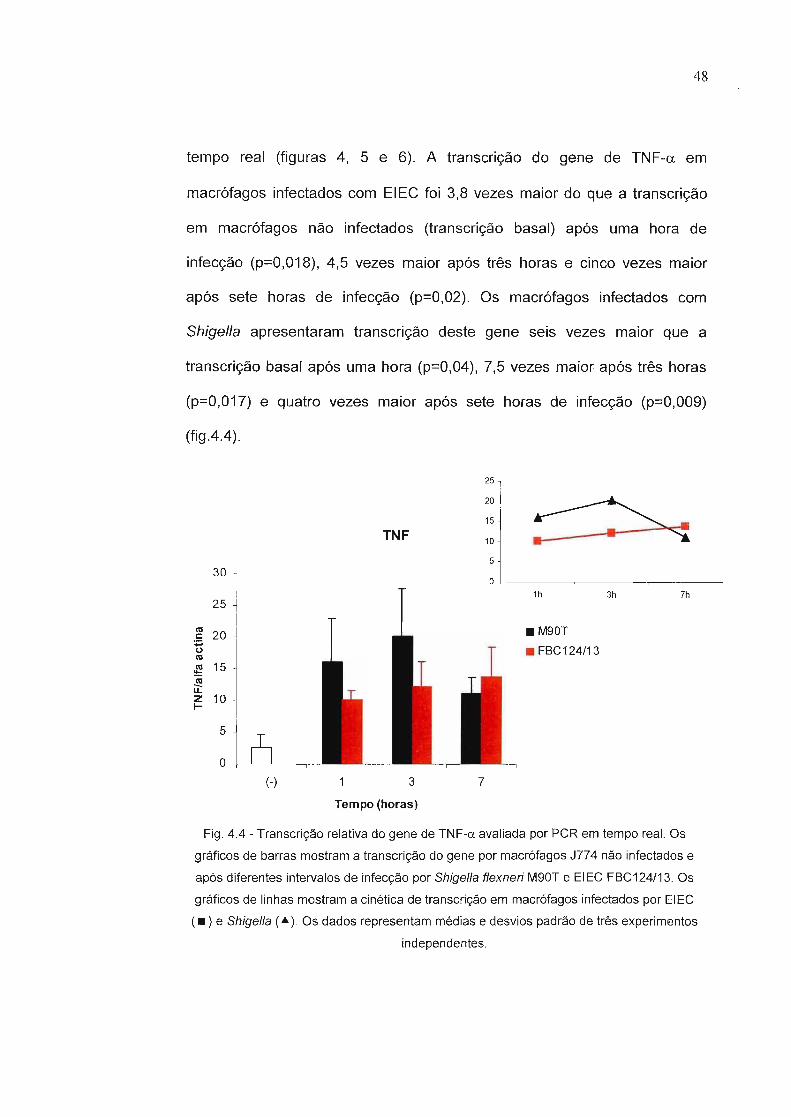
48
tempo real (figuras 4, 5 e 6). A transcrição do gene de TNF-a em
macrófagos infectados com EIEC foi 3,8 vezes maior do que a transcrição
em macrófagos não infectados (transcrição basal) após uma hora de
infecção (p=O,018), 4,5 vezes maior após três horas e cinco vezes maior
após sete horas de infecção (p=O,02). Os macrófagos infectados com
Shigella apresentaram transcrição deste gene seis vezes maior que a
transcrição basal após uma hora (p=O,04), 7,5 vezes maior após três horas
(p=O,017) e quatro vezes maior após sete horas de infecção (p=O,009)
(figAA).
:]---~--~-~7h3h
~~1h
.M90T
• FBC124/13
25 1
20 I15
10TNF
30 l
II
25 1I
:g 20 J:;:;oCll
~ 15 ~
~ Iz 10 .•I- I
I
:~(-) 1 3 7
Tempo (horas)
Fig. 4.4 - Transcrição relativa do gene de TNF-a avaliada por PCR em tempo real. Os
gráficos de barras mostram a transcrição do gene por macrófagos J774 não infectados e
após diferentes intervalos de infecção por Shigella f1exneri M9üT e EIEC FBC124/13. Os
gráficos de linhas mostram a cinética de transcrição em macrófagos infectados por EIEC
( • ) e Shigella ( ... ). Os dados representam médias e desvios padrão de três experimentos
independentes.

49
A transcrição do gene de IL-1 em macrófagos infectados com EIEC
foi três vezes maior do que a transcrição basal após uma hora de infecção,
10 vezes maior após três horas (p=0,012), e 12,6 vezes maior após sete
horas (p=0,014). Na infecção por Shigella observamos um aumento de 3,8
vezes na transcrição deste gene em relação à transcrição basal após uma
hora de infecção, 12 vezes após três horas (p=0,004) e oito vezes após sete
horas (p=0,009) (figA.5).
7h3h1h
.M90T
• FBC124/13
---,
0.35
0.3
0.25
0.2
0.15
0.1
0.05
oJ---~---~---~
0,1
0,05
° I I I
IL-1
-:::; 0,15-
0,45
0,4 -
0,35
.~ 0,3...g 0,25J!lii 0,2
(-) 3 7
Tempo (horas)
Fig. 4.5 - Transcrição relativa do gene de IL-1 avaliada por PCR em tempo real. Os gráficos
de barras mostram a transcrição do gene por macrófagos J774 não infectados e após
diferentes intervalos de infecção por Shigella f1exneri M9üT e EIEC FBC124/13. Os gráficos
de linhas mostram a cinética de transcrição em macrófagos infectados por EI EC (. ) e
Shigella ("'). Os dados representam médias e desvios padrão de três experimentos
independentes.
o aumento na transcrição do gene de IL-10 em relação à transcrição
basal só foi significativo após três horas de infecção por EIEC (p=0,018).

'i0
Apesar de este aumento ocorrer também na infecção por Shigella, ele não
foi considerado estatisticamente significativo (fig. 4.6).
Os macrófagos infectados por Shigella apresentaram maior transcrição dos
genes de TNF e IL-1 após uma e três horas de infecção, enquanto que após
sete horas de infecção a transcrição dos genes destas duas citocinas foi
maior na infecção por EIEC. Entretanto, não houve diferenças significativas
entre os macrófagos infectados por Shigella ou EIEC.
---,
0,12 1
I0,08 1
IL-100,04
°.M90T 1h 3h 7h
• FBC124/13
0.2
10,16 -I
Cllc::g 0,12-Cll
~Cll
Õ 0,08....~
0,04
o(-) 3 7
Tempo (horas)
Fig. 4.6 - Transcrição relativa do gene de IL-10 avaliada por PCR em tempo real. Os
gráficos de barras mostram a transcrição do gene por macrófagos J774 não infectados e
após diferentes intervalos de infecção por Shigella flexneri M90T e EIEC FBC124/13. Os
gráficos de linhas mostram a cinética de transcrição em macrófagos infectados por EIEC
( • ) e Shigella ( "'). Os dados representam médias e desvios padrão de três experimentos
independentes.
4.1.5 - Determinação da produção de TNF-a, IL-1 e IL-10 por ELISA
Os sobrenadantes das culturas de macrófagos não infectados e
infectados por sete horas com Shigella e EIEC (cuja transcrição dos genes

51
foi avaliada no ensaio anterior) foram utilizados no ELISA de captura para
determinação da concentração de TNF-a, IL-1 e IL-10. Os resultados
obtidos confirmaram que a transcrição do RNA mensageiro resultou em
produção das três citocinas (figA.7). Houve um aumento de 13 vezes na
produção de TNF-a por macrófagos infectados por EIEC (p=O,001) e de 15
vezes em macrófagos infectados por Shigella (p=O,007) em relação à
produção basal. A produção de IL-1 por macrófagos infectados por EIEC foi
1,2 vez maior do que a produção basal. Em macrófagos infectados por
Shigella esta produção foi 3,8 vezes maior. Não foi detectada produção
basal de IL-10. A produção desta citocina foi significativa em macrófagos
infectados por EIEC (p=O,007) e Shigella (p=O,004). Assim como no ensaio
de PCR em tempo real, não houve diferenças significativas na produção de
citocinas por macrófagos infectados por Shigella ou EIEC.
TNF IL-1
200 l 300 l250 ~
160200
_ 120.§ 150E
Õl Clo- 80
o-
100
40 50
o o(-) flfeOT FBC124/13 (-) flfeOT FBC124/13
IL-10
30°1250
I200
.§ 150Clo-
100
50
o(- ) flfeOT FBC124/13

52
Fig. 4.7- Determinação, por ELISA de captura, da produção de TNF-a, IL-1 e IL-1ü por
macrófagos J774 não infectados e infectados por sete horas com Shigella f1exneri M9üT e
EIEC FBC124/13. Os dados representam médias e desvios padrão de duplicatas e são
representativos de dois experimentos independentes.
4.2 - Interação entre EIEC e macrófagos de camundongos C57BLl6 e
C57BLl6 iNOS KO (ensaios Itin vitra '1
4.2.1- Determinação da produção de NO pelo método de Griess
A oxidação do NO a N02- em meio aquoso permite a detecção da
produção de NO pelo reativo de Griess. Observou-se que macrófagos de
camundongos C57BU6 nocauteados no gene da NO sintase induzível
(iNOS KO) não produziram quantidades detectáveis de NO (figA.8).
Macrófagos de camundongos C57BU6 selvagens infectados com 105 EIEC
produziram significativamente mais NO do que os não infectados (p=0,009),
enquanto macrófagos infectados com 104 (p=0,0003) e 105 (p<0,0001)
Shigella produziram significativamente mais NO que os não infectados. Os
macrófagos infectados com Shigella produziram significativamente mais NO
do que aqueles infectados com EIEC nos dois inóculos testados (p= 0,0006
quando o inóculo foi de 104, e p= 0,0003 quando o inóculo foi de 105
).

53
*
o C57BL16 iNOs KO
.C57BL16
(105)(104
)(105)(104
)
30 -.
25
20
i'2. 15bz
10
5
O
(-)
... v v
Shigella M90T EIEC FBC124/13
Fig. 4.8 - Determinação da produção de NO por macrófagos infectados com Shigella M90T
e EIEC FBC124/13 pelo método de Griess.Os dados representam médias de triplicatas e
são representativos de dois experimentos independentes. * - Macrófagos infectados por
Shigella produzem significativamente mais NO que aqueles infectados por EIEC (p<O,OS
segundo o teste t não pareado).
4.2.2 - Análise da transcrição relativa dos genes das citocinas, da
enzima iNOS e do TLR4 por RT-peR semiquantitativo
Neste ensaio, a transcrição relativa de genes de citocinas por
macrófagos de camundongos C57BU6-e C57BU6 iNOS KO infectados por 4
horas com EIEC, Shigella, E.coli K12 (proporção de uma bactéria por
macrófago) e estimulados com LPS (10 ng/ml) foi avaliada por RT-PCR

54
semiquantitativo. Foi utilizado um software (Scion Image) para análise
densitométrica dos produtos de RT-PCR (figuras 4.9 a 4.15).
Pode-se observar que em camundongos selvagens o estímulo com
LPS resultou em aumento significativo da transcrição dos genes de TNF
(p=0,01), IL-1 (p=0,008), IL-18 (p=0,005), TLR4 (0,003) e IP-10 (p=0,05), em
relação aos macrófagos não infectados (transcrição basal).
A transcrição do gene de TNF foi significativa na infecção por EIEC
(p=0,0004) e Shigella (p=0,008), em relação à transcrição basal (fig. 4.9),
em macrófagos de camundongos selvagens. Macrófagos infectados com
EIEC apresentaram maior transcrição deste gene do que macrófagos
infectados com Shigella (p=0,008).
Em macrófagos de camundongos nocauteados no gene da NO
sintase induzível (iNOS KO), o aumento na transcrição do gene do TNF em
relação à transcrição basal é evidente em valores absolutos. Porém, estas
diferenças não foram consideradas estatisticamente significativas, devido ao
elevado valor do desvio padrão da transcrição basal (fig. 4.9).
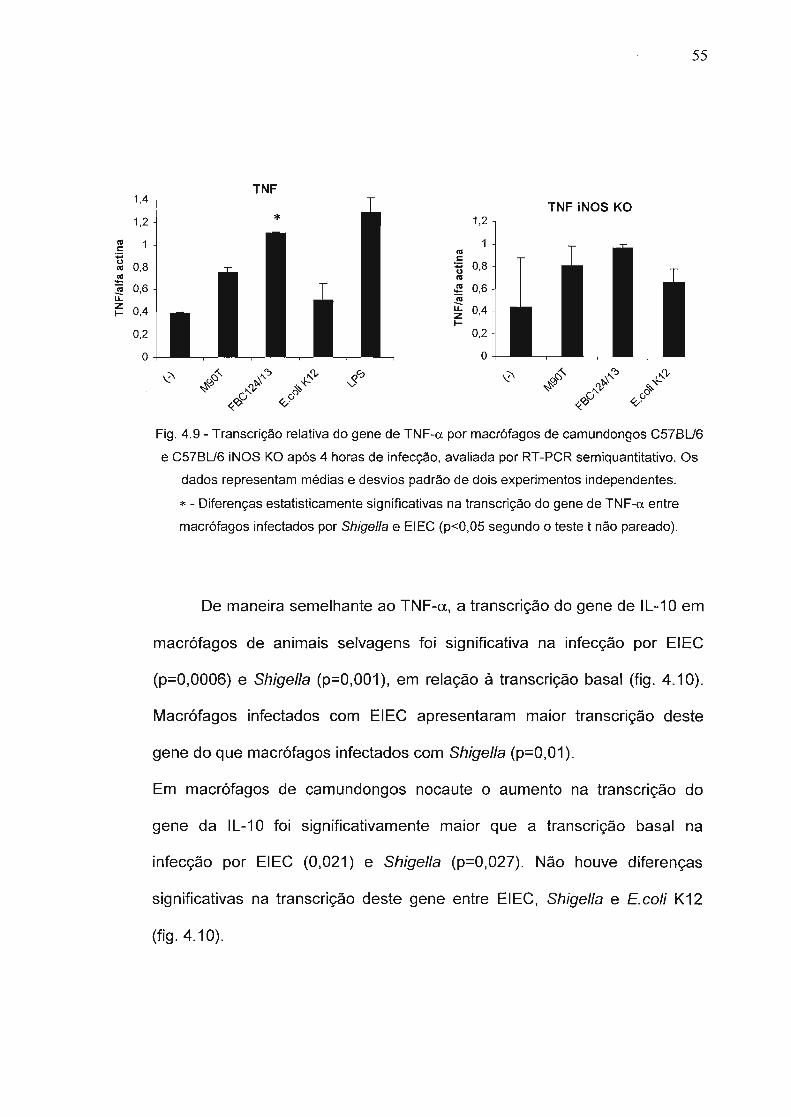
55
TNF iNOS KO1,4 1
1,2
CIIC~oCII 08~ ,
~ 0,6LI.zI- 0,4
0,2
O
\:,
TNF
s- ,,~
$3 ~0"
«<o
.+,-'l,~
<v'?
~0v
1,2
CIIC:g 0,8CII
~ 0,6~~ 0,4I-
0,2
O
\:, ~$'(:) ';),,,~
0"rf«<o
.+,-'l,~
<v'?
Fig. 4.9 - Transcrição relativa do gene de TNF-a por macrófagos de camundongos C57BU6
e C57BL/6 iNOS KO após 4 horas de infecção, avaliada por RT-PCR semiquantitativo. Os
dados representam médias e desvios padrão de dois experimentos independentes.
* - Diferenças estatisticamente significativas na transcrição do gene de TNF-a entre
macrófagos infectados por Shigella e EIEC (p<O,05 segundo o teste t não pareado).
De maneira semelhante ao TNF-a, a transcrição do gene de IL-10 em
macrófagos de animais selvagens foi significativa na infecção por EIEC
(p=0,0006) e Shigella (p=0,001), em relação à transcrição basal (fig. 4.10).
Macrófagos infectados com EIEC apresentaram maior transcrição deste
gene do que macrófagos infectados com Shigella (p=0,01).
Em macrófagos de camundongos nocaute o aumento na transcrição do
gene da IL-10 foi significativamente maior que a transcrição basal na
infecção por EIEC (0,021) e Shigella (p=0,027). Não houve diferenças
significativas na transcrição deste gene entre EIEC, Shigella e E.coli K12
(fig.4.10).

56
1,4 lIL-10
1,2 ~*
CllI • • .- IL-10 iNOS KO
c
1,2
:;:;o: 0,8
Cll
-~ 0,6
c~ 0,8
~ 0,4
Cll
~ 0,6~
0,2
~ 0,4
°
:::!0,2
°f..~OjÇ)o ~"O:> . +-'], -$0 0 RJrS- ').,,0:> +-'],
0"rv ~ ~ "tf ~«<Q <v~ «'<)0 <v~
Fig. 4.10 - Transcrição relativa do gene de IL-10 por macrófagos de camundongos CS7BLl6
e CS7BLl6 iNOS KO após 4 horas de infecção, avaliada por RT-PCR semiquantitativo. Os
dados representam médias e desvios padrão de dois experimentos independentes.
* - Diferenças estatisticamente sign ificativas na transcrição do gene de IL-10 entre
macrófagos infectados por Shigel/a e EIEC (p<O,OS segundo o teste t não pareado).
o aumento da transcrição do gene da IL-1 em relação à transcrição
basal foi significativo na infecção por EIEC (p=O,OOS), Shigella (p=O,02S) e
E.coli K12 (p=O,007). No entanto, não houve diferenças significativas na
transcrição deste gene entre EIEC, Shigella e E.coli K12 (fig. 4.11). Em
camundongos nocaute não houve diferenças significativas na transcrição do
gene de IL-1 na infecção pelas três bactérias, em relação à transcrição
basal.

57
IL-1 IL-1 iNOS KO
.+,'1~
<v9
~"fl:Jv"rv
«<Q
fo-.$'\:)
1,6
1,4
1,2l'CI.:ül'CI
:! 0,8~"';' 0,6~
0,4
0,2
O
~,«0v.+,'1
~<v9
'}.,,,fl:J
V"rf«<Q
fo-.$'\:)
1,6 1
I1,4
1,2l'CIc::p.,l'CI
:! 0,8ni;;: 0,6~
0,4
0,2
O
0
Fig. 4.11 - Transcrição relativa do gene de IL-1 por macrófagos de camundongos C57BU6 e
C57BU6 iNOS KO após 4 horas de infecção, avaliada por RT-PCR semiquantitativo. Os
dados representam médias e desvios padrão de dois experimentos independentes.
o aumento na transcrição do gene da IL-18 em relação à transcrição
basal foi significativo em macrófagos de camundongos selvagens na
infecção por EIEC (p=O,014) e Shigella (p=O,028). Não houve diferenças
significativas entre a transcrição do gene desta citocina entre macrófagos
infectados com EIEC, Shigella e E.coli K12 (fig. 4.12).
Curiosamente, em macrófagos de camundongos nocaute foi
observada uma inibição da transcrição do gene da IL-18 após infecção por
EIEC (p=O,027) e E.coli K12 (p=O,028) nos dois ensaios realizados. (fig.
4.12). A inibição da transcrição deste gene na infecção por Shigella foi
considerada pouco significativa (p=O,09), devido ao elevado desvio padrão.
Não foram observadas diferenças significativas na inibição da transcrição de
IL-18 entre EIEC, Shigella e E.coli K12.

58
IL-18 iNOS KO1,4 .,
1,2 l IL-181,2
nscns :;::;cu~ 0,8 : 0,8ns -.1! 0,6 ~ 0,6::§ 0Cl
~ 0,4 ...~
~ 0,4
0,2 0,2
° °0 s- ~,,~ +,1- ~0 0 s- ,,~ .+,1-$3 ~ ~ v $3 ~ ~0" 0° 0" <v':J««) <v. ««)
Fig. 4.12 - Transcrição relativa do gene de IL-18 por macrófagos de camundongos C57BU6
e C57BU6 iNOS KO após 4 horas de infecção, avaliada por RT-PCR semiquantitativo. Os
dados representam médias e desvios padrão de dois experimentos independentes.
Ao contrário do observado na transcrição gênica das citocinas, a
transcrição do receptor transmembrânico TLR4 após infecção, em relação à
transcrição basal, foi significativo apenas em macrófagos infectados por
E.coli K12 (p=O,03) (fig. 4.13). Em macrófagos de camundongos nocaute
não houve aumento significativo na transcrição deste gene após infecção
por Shigella, EIEC e E.coli K12, devido aos elevados valores de desvio
padrão (figA.13).

59
TLR4 iNOS KO
1,6I TLR41,4 ~ 1,4
la 1,2 1,2t::
:;:;1
la<.> t::la :;:;
~ 08<.>: 0,8
la ' -~ 0,6 ~ 0,6..J a::I- 0,4 ~ 0,4
0,2 0,2
O O
0 S- ~,,"J . ~'1- «0 0$! ,,'I) ~v
~<Q0 <vy
f...$'<;) ~,,"J
0"rf~«)
. ~'1~
<v':J
Fig. 4.13 - Transcrição relativa do gene de TLR4 por macrófagos de camundongos C57BU6
e C57BU6 iNOS KO após 4 horas de infecção, avaliada por RT-PCR semiquantitativo. Os
dados representam médias e desvios padrão de dois experimentos independentes.
o aumento na transcrição do gene da enzima iNOS em relação à
transcrição basal foi significativo na infecção por Shigella (p=O,01) e E.coli
K12 (p=O,04). A transcrição do gene da iNOS em macrófagos infectados por
Shigella foi maior do que em EIEC (p=O,02)(fig. 4.14).

60
*
~C:>vo
"' 1,2c::=<J
"'~ 0,8..!!!til
OZ
1,6 l
o
0,4
B,§. ~,,":J . +,-'"~ ,,'1: (t
«<Qv <v5i
Fig. 4.14 - Transcrição relativa do gene da iNOS por macrófagos de camundongos CS7BU6
e CS7BU6 iNOS KO após 4 horas de infecção, avaliada por RT-peR semiquantitativo. Os
dados representam médias e desvios padrão de dois experimentos independentes.
* - Diferenças estatisticamente significativas na transcrição do gene de iNOS entre
macrófagos infectados por Shigella e EIEC (p<O,OS segundo o teste t não pareado).
4.2.3 - Determinação da produção de TNF-a e IL-10 por ELISA
Os sobrenadantes das culturas de macrófagos não infectados e
infectados por quatro horas com Shigella, EIEC e E.coli K12, e estimulados
com LPS (cuja transcrição dos genes foi avaliada no ensaio anterior) foram
utilizados no ELISA de captura para determinação da concentração de TNF-
a e IL-10. De maneira semelhante ao ocorrido nos ensaios com macrófagos
J774, confirmou-se que a transcrição dos genes resultou em produção de
proteína (figuras 4.15 e 4.16).
Em relação aos macrófagos de camundongos selvagens não-
infectados (produção basal da citocina), a produção de TNF-a foi
significativamente maior em macrófagos infectados por Shigella (p=0,001),
EIEC (p=0,002), E.coli K12 (0,003) e estimulados com LPS (p=0,0001).
Macrófagos infectados com EIEC produziram mais TNF-a que

61
aqueles infectados com Shigella (p=O,043); estes resultados vão de
encontro aos obtidos no ensaio de transcrição gênica. Os macrófagos de
camudongos iNOS nocaute infectados por Shigella (p=O,001), EIEC
(p=O,0002) e E.coli K12 (p=O,001) produziram significativamente mais TNF
comparando-se aos macrófagos não-infectados, e a diferença na produção
desta citocina entre macrófagos infectados por Shigella e EIEC também foi
significativa (p=O,002) (fig. 16).
4000 l TNF
3500
3000 " ... - TNFiNOSKO2500
_ 2500 IlIIIIiiIII 11 ~ 11 2000-§, 2000
Q.
1500 J • • • • E 1500
~ 10001000
500500
o o
0 S- ,,"3 .+,'" ~C:J 0 S- ,,"3 .+,'".$S :f' v .$S :f' ~
0" ~ 0" <vy«.<Q <vy «.<Q
Fig. 4.15 - Determinação, por ELISA de captura, da produção de TNF-a por macrófagos de
camundongos C57BU6 e C57BU6 iNOS KO após 4 horas de infecção. Os dados
representam médias e desvios padrão de duplicatas e são representativos de dois
experimentos independentes. * - Diferenças estatisticamente significativas na produção de
TNF-a entre macrófagos infectados por Shigella e EIEC (p<O,05 segundo o teste t não
pareado).

62
A produção de IL-10 foi maior na infecção por Shigella (p<O,0001),
EIEC (p<O,0001), e E.coli K12 (p=O,0014), em relação à produção basal
desta citocina. Macrófagos infectados com EIEC produziram mais IL-10 que
aqueles infectados com Shigella (p=O,0016). Os macrófagos de
camundongos iNOS nocaute produziram significativamente mais IL-10
quando infectados com Shigella (p=O,005), EIEC (p=O,001) e E.coli K12
(p=O,001) comparando-se aos macrófagos não-infectados (fig.17). Neste
caso, também houve diferença significativa na produção de IL-10 entre
macrófagos infectados por EIEC e Shigella (p=O,02).
IL-10 iNOS KO
IL-10*
.~'l(J>
xl~,,":J
0"rv~<Q
~$\;)
9000
8000
7000
6000
5000
4000
3000
2000
1000
o0«C:Jv. ~'l
(J>xl
*
~,,":J
0"rv~<Q
~~C!)\;)o
3000
1000
500
o
3500
2500
E 2000
~ 1500
Fig. 4.16 - DeterminaçãO, por ELISA de captura, da produção de IL-10 por macrófagos de
camundongos C57BLl6 e C57BLl6 iNOS KO após 4 horas de infecção. Os dados
representam médias e desvios padrão de duplicatas e são representativos de dois
experimentos independentes. * - Diferenças estatisticamente significativas na produção de
IL-10 entre macrófagos infectados por Shigella e EIEC (p<O,05 segundo o teste t não
pareado).

63
4.3 - Interação entre EIEC e camundongos C57BU6 e C57BU6 iNOS KO
"in vivo"
4.3.1 - Determinação do influxo celular para o peritônio após infecção
A análise dos resultados obtidos na citometria de fluxo pode ser
observada na figura 4.17. A infecção intraperitonial com ShigeJla e EIEC
resulta em influxo celular, uma vez que o número de células aumentou da
ordem de 106 para 107. Os granulócitos são a população predominante em
todos os tempos de infecção por ShigeJla e EIEC. Camundongos infectados
por ShigeJla, porém, permanecem com o número destas células aumentado
mesmo após 48 horas de infecção, enquanto que, em na infecção por EIEC,
observa-se um decréscimo a partir de 24 horas.
Observa-se também o influxo de macrófagos para o peritônio após
infecção com as duas bactérias. No peritônio de camundongos infectados com
Shigella o número de macrófagos praticamente não se altera após 12 e 24
horas de infecção, no entanto, após 48 horas, este número é três vezes maior
do que em camundongos não-infectados. A infecção por EIEC estimulou um
padrão semelhante de influxo de macrófagos; no entanto, após 48 horas o
número de macrófagos na infecção por Shigella é aproximadamente 1,3 vez
maior que na infecção por EIEC.
O número de linfócitos no peritônio de animais infectados com Shigella
e EIEC diminuiu em relação aos não-infectados. Na infecção por Shigella esta
diminuição é observada após 12 e 24 horas de infecção; após 48 horas o

64
número de linfócitos é semelhante ao número encontrado em camundongos
não-infectados. Na infecção por EIEC a diminuição ocorre apenas após 12
horas de infecção; nos tempos subseqüentes os valores são restaurados aos
valores basais.
EIEC FBC124/13_ Macrófagos
_ Neulrófilos
_ Linfócitos
24h 48h
2000,
o 1600~
'".!!!1200:>
:c;;".."C 800e..E
400.:>Z
o12 24 48 (-) 12h
Shigella flexneri M90T2000,
f 1600I
'".!!!1200:>
:c;;".."C 800·e..E
.:> 400Z
o(-)
Tempo de infecção (horas) Tempo de infecção (horas)
FigA.17 - Número de células em camundongos C57BL/6 não infectados e após 12, 24 e 48
horas de infecção com Shigella M9üT e EIEC FBC124/13, determinado por citometria de fluxo.
Os dados são representativos de dois experimentos independentes.
Foi realizada também marcação com anti-MHCII, anti-C080 e anti-
C086, moléculas que formam o complexo de ligação ao linfócito T, e são um
indicativo do estado de ativação dos macrófagos. Além disso, foi analisada
isoladamente a transcrição do marcador de macrófagos F4/80 (figuras de 4.18
a 4.21).
A infecção por Shigella e EIEC induz a modulação positiva da
transcrição de MHCII na superfície dos macrófagos peritoniais. No entanto,
macrófagos de camundongos infectados com Shigella apresentaram maior
expressão desta molécula do que os macrófagos de camundongos infectados
com EIEC, em todos os tempos de infecção (figA.18)

65
M1
11102Fl1-M1C 11
FBC124/13M90T~l
o1 '"MHCII __ M90T
-- FBC124/1~1 I MI I ov
250 l200 ::::====---:: q
I
oM
~
1~ j D, ~ll.~<J~
100 -
50· =' ~ IIL~ü"··'~I.. ... 1 1"1. I ;:>
O -I o' r '-"'~"ILW ~\
lJO 10 ' 1~2 103 lIi4 o11)012 24 48 Fl1-M1C 11
Terrpo de infecção (horas)
Fig. 4.18 - Análise da expressão de MHCII por macrófagos em peritônio de camundongos não
infectados (linha preta) e após 12h (linha verde), 24h (linha rosa) e 48h (linha azul) de infecção
com Shigella f1exneri M9üT e EIEC FBC124/13. Os gráficos e histogramas são
representativos de dois experimentos independentes.
Também se observa aumento na expressão da molécula co-
estimulatória C080 (87-1) em todos os tempos de infecção por Shigella e
EIEC, em relação aos controles negativos. Após 12 horas de infecção, a
expressão desta molécula é maior em macrófagos de camundongos infectados
com EIEC, e, nos tempos subseqüentes, maior na infecção por Shigella
(figA.19).

- ,.
-..1.:_ dadf. c; (.;i'" ;.\..,~ 66
coso M90T FBC124/13
_M90T :B ] <:>500, I c
_FBC124/13
~<:>
400 '"M1 ] I M1
~ C>v
11l 300
A.l/lic: IA IA
.!l1 cc>'O
cc>Q) 5"" """::;: 200 <> o
,,(1IM~~'~,\<>
C> C>N N
': je'j 1!ljfjJ '\ V! lAll'L I e
C>
,ÓOI "" lÓ' 1~2 ' , " '1~3 I I 1~4 C>
12 24 481(;0 . ... "io1 . .. ';'02 .... ";'03 ";Ô4
FL2-CD80 FL2-CD80
Tempo de infecção (horas)
Fig. 4.19 .. Análise da expressão de C080 por macrófagos em peritônio de camundongos não
infectados (linha vermelha) e após 12h (linha verde), 24h (linha rosa) e 48h (linha azul) de
infecção com Shigella flexneri M90T e EIEC FBC124/13. Os gráficos e histogramas são
representativos de dois experimentos independentes.
Curiosamente, houve diminuição da expressão da molécula co-
estimulatória CD86 (87-2) em macrófagos de camundongos infectados com as
duas bactérias em relação aos não infectados. Esta modulação negativa foi
maior em camundongos infectados com EIEC em todos os tempos de infecção
(fig. 4.20).

67
M90T FBC124/13
104
M1
10 1 - - - -;02 - -i03
FL2-CD86
oM
...,N
oN
IA;:...,:3~
oV
~
...,
o104 11)0
M1
101 ... '1()2 - - -i03FL2-CD86
oN
li') -.nr-.!lJIW.T\l\I .....
~
IAC...,:3~
OV
g, i
__M90T
___ FBC124/13 :!3
CD86
~
Tempo de infecção (horas)
12 24 48
O+1-----.-----------,-----,
40
160
120
lO<:
;3 80~
Fig. 4.20 - Análise da expressão de CD86 por macrófagos em peritônio de camundongos não
infectados (linha vermelha) e após 12h (linha verde), 24h (linha rosa) e 48h (linha azul) de
infecção com Shigella flexneri M90T e EIEC FBC124/13. Os gráficos e histogramas são
representativos de dois experimentos independentes.
A expressão de F4/80 também decresce após 12 horas de infecção
pelas duas bactérias, em relação aos não infectados. Na infecção por Shigella,
os valores da expressão de F4/80 continuam baixos nos tempos
subseqüentes, enquanto na infecção por EIEC este vaiare é praticamente
restaurado ao nível basal após 48 horas de infecção (fig. 4.21).

68
M90T FBC124/13F4/80
-+-tv'9üT o o~ ~
__ F8C124/13
500o oo:>
/lro
400 -
o o300 -
",.oM1 "'u::>E E::>
o ::><.)0 o
200 - v
\<">0v
100 - o I
'" \ o\ N
I0- ~.....
o12 24 48
100
101 10
2 104 o
100
101
102
Terrpo de infecção (horas) FL1-F4180FL 1-F4180
Fig. 4.21 - Análise da expressão de F4/8ü por macrófagos em peritônio de camundongos não
infectados (linha vermelha) e após 12h (linha verde), 24h (linha rosa) e 48h (linha azul) de
infecção com Shigella flexneri M9üT e EIEC FBC124/13. Os gráficos e histogramas são
representativos de dois experimentos independentes.
4.3.2 - Análise da transcrição relativa dos genes das citocinas, iNOS
eTLR4 por RT-PCR semiquantitativo
Neste ensaio foi avaliada a transcrição relativa de genes das citocinas
TNF-a, IL-10, IL-1, IL-18 e IFN-y, da quimiocina IP-10, do TLR4 e da enzima
iNOS por células peritoniais totais de camundongos C57BLl6 e C57BLl6
iNOS KO infectados por 4 e 48 horas com EIEC, Shigella, E.coli K12 (107
bactérias) por RT-PCR semiquantitativo. Foi realizada análise densitométrica
com software apropriado (Scion Image) (fig. 4.22 a 4.30).
A transcrição do gene do TNF-a após 4 horas de infecção foi
significativamente maior em células peritoniais de animais selvagens
infectados com Shigella (p=0,03), EIEC (p=0,003) e E.coli K12 (p=0,052),
em relação à transcrição basal (camundongos não infectados) (fig. 4.22). A
transcrição do gene do TNF-a foi maior por células de camundongos
infectados com Shigella do que com EIEC (p=0,02) e maior na infecção por

69
ShigeIJa do que na infecção por Ecoli K12 (p=O,02). Após 48 horas de
infecção, apenas o aumento na transcrição deste gene por camundongos
infectados com ShigeIJa foi significativo (p=O,001). Camundongos infectados
com ShigeIJa apresentaram maior transcrição deste gene do que aqueles
infectados com Ecoli K12 (p=O,002).
Em camundongos nocauteados no gene da iNOS também houve
aumento significativo da transcrição do gene de TNF-a após 4 horas de
infecção por ShigeIJa (p=O,003), EIEC (p=O,004) e Ecoli K12 (p=O,005);
porém, após 48 horas, este aumento foi significativo apenas após infecção
por EIEC (p=O,02), provavelmente devido aos altos valores de desvio padrão
obtidos na transcrição do gene de TNF-a em animais infectados com
ShigeIJa e Ecoli K12 (fig. 4.22).
*1,6
1,4
<11 1,2c:13 1lO
J! 08lii 'Li: 0,6zI- 0,4
, 0,2
° -\ À ~ ~ ~ <0'(:- c§' c§''- ~ ~ ~ bI bI bI
p'>\:l ~" ~ ~ ,,~ ,,'J,~ ,,'I) ~C8:f +:
~0 0"« «<Q
TNF iNOS KO1,2
~ 1~ 0,8
~ 0,6lO
Li: 0,4
~ 0,2
0 ' i
\ À ~ ~ ~ '0'(:- c§' c§''- ~ ~ ~ bI bI bI
p'>\:l ~" ~ ~ ,::-"~ ,,'J,~ ,,'I) $' l' +:
~0 0"« «<Q
Fig. 4.22 - Transcrição relativa do gene de TNF-a por células peritoniais de camundongos
C57BU6 e C57BU6 iNOS KO após 4 e 48 horas de infecção, avaliada por RT-PCR
semiquantitativo. Os dados representam médias e desvios padrão de dois experimentos
independentes. * - Diferenças estatisticamente significativas na transcrição do gene de
TNF-a entre macrófagos infectados por Shigella e EIEC (p<O,05 segundo o teste t não
pareado).

70
o aumento na transcrição do gene de IL-10 por células de
camundongos selvagens após 4 horas foi significativo na infecção por EIEC
(p=O,008), Shigella (p=O,01) e EcoJi K12 (p=O,027) em relação à transcrição
basal (fig. 4.23). EIEC induziu uma maior transcrição deste gene do que
Shigella (p=O,014) e EcoJi K12 (p=O,007), e Shigella estimulou maior
transcrição deste gene que EcoJi K12 (p=O,03). Curiosamente, após 48
horas de infecção, a bactéria que mais induziu a transcrição de gene de IL
10 em relação à transcrição basal foi Shigella (p=O,018). A transcrição deste
gene foi maior em camundongos infectados com Shigella do que naqueles
infectados com EcoJi K12 (p=O,OOS).
Em células de camundongos iNOS nocauteados, o aumento na
transcrição do gene de IL-10 foi significativa na infecção por Shigella
(p=O,002), EIEC (p=O,007) e EcoJi K12 (p=O,018). Porém, nestes
camundongos o aumento na transcrição é semelhante em camundongos
infectados com Shigella e EIEC, e maior em camundongos infectados com
Shigella do que naqueles infectados com EcoJi K12 (p=O,OOS), e também
maior em camundongos infectados com EIEC do que na infecção por EcoJi
K12 (p=O,017). Após 48 horas de infecção o aumento na transcrição do
gene de IL-10 foi significativo apenas na infecção por EIEC, (p=O,011) em
relação à transcrição basal, e não há diferenças entre a transcrição deste
gene nas células peritonias de camundongos infectados com Shigella, EIEC
e EcoJi K12 (fig.4.23).

71
1,8* IL-10 IL-10 iNOS KO
1,61,6
1,41,4
11l
~ 1,215 1,2u
~11l 1 1~ 11l~ 0,8 ~ 0,8o 11l:::; 0,6 ~ 0,6
0,4 ~ 0,40,2 0,2
O O
o ~~ ~~ ~ b.?J:~ t;..ciJ:' $' 1(' ~ ~~ ~ t$>~ $' t$>~$J~ ~" +''"], _-RJrS- .~p +''"], $JrS-~" +''"], PJrS- ~,,~ +''"],
0" y ,,'V 0" ~ ,,'V«<Q «<Qu «<Q ~u
Fig. 4.23 - Transcrição relativa do gene de IL-10 por células peritoniais de ~mundongosC57BU6 e C57BLl6 iNOS KO após 4 e 48 horas de infecção, avaliada por RT-PCR
semiquantitativo. Os dados representam médias e desvios padrão de dois experimentos
independentes. * - Diferenças estatisticamente significativas na transcrição do gene de IL
10 entre macrófagos infectados por Shigella e EIEC (p<O,05 segundo o teste t não
pareado).
Em camundongos selvagens a transcrição do gene de IL-1 foi
significativamente maior que a transcrição basal na infecção por Shigella
(p=O,003) e EIEC (p=O,006) (fig. 4.24). A transcrição do gene de IL-1 foi
maior em camundongos infectados com Shigella do que naqueles infectados
com EIEC (p=O,04) e E.coli K12 (p=O,02). As células peritoniais de
camundongos infectados por 48 horas com Shigella foram as únicas a
apresentar aumento significativo da transcrição do gene de IL-1 em relação
à transcrição basal (p=O,003), e Shigella induziu maior transcrição deste
gene do que E.coli K12 (p=O,006). Entre os camundongos nocaute o
aumento na transcrição do gene de IL-1 em relação à transcrição basal foi
maior na infecção por EIEC (p=O,002) e Shigella (p=O,005). EIEC induziu
maior transcrição deste gene que Shigella (p=O,01). Após 48 horas não

72
houve diferenças significativas no aumento da transcrição deste gene em
células de camundongos infectados comparando-se à transcrição basal;
também não houve diferenças entre a transcrição do gene de IL-1 na
infecção por Shigella, EIEC e Ecoli K12 (fig. 4.24).
IL-1 iNOS KO
1,4
1,2
.~ 113til 0,8~~ 0,6~
~ 0,4
0,2
o
IL-1*
1,6 1I
1,4
<li 1,2cTI 1<li
~ 0,8~~ 0,6~
0,4
0,2
o 1-
*
,>, ~ ~ ~ co"'" c§' c§''-~ ~ ~ ~ ~ ~
2)Ç) ~" +' sS- '},.,,~ ,,'1-~ ,,'V $ l' +:
~0 0"« «<Q
,>, ~ ~ ~ co"'" c§' 0.."",'- ~ ~ ~ ~ ~ b-V
$JÇ) #" +' 2)rS- ~"~ *-"'1-0" ~ r~'V
«<Q ~v«
Fig. 4.24 - Transcrição relativa do gene de IL-1 por células peritoniais de camundongos
CS7BU6 e CS7BU6 iNOS KO após 4 e 48 horas de infecção, avaliada por RT-PCR
semiquantitativo. Os dados representam médias e desvios padrão de dois experimentos
independentes. * - Diferenças estatisticamente significativas na transcrição do gene de IL-1
entre macrófagos infectados por Shigella e EIEC (p<O,OS segundo o teste t não pareado).
o aumento na transcrição do gene da IL-18 em relação à transcrição
basal foi significativo após quatro horas de infecção por Shigella (p=O,021) e
pouco significativo na infecção por Ecoli K12 (p=O,051) (fig. 4.25). Shigella
induziu maior transcrição deste gene que Ecoli K12 (p=O,04). Após 48 horas
de infecção não houve aumento significativo na transcrição do gene de IL-18
na infecção por EIEC, Shigella e Ecoli K12 em relação à transcrição basal.
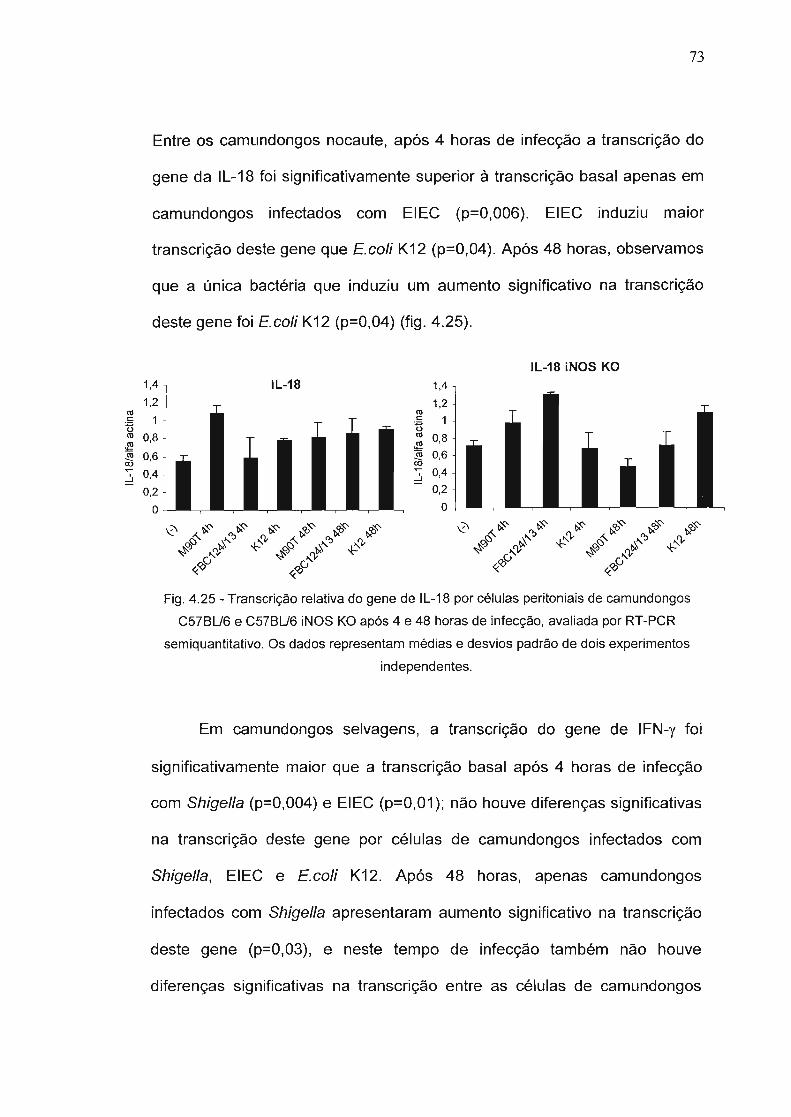
73
Entre os camundongos nocaute, após 4 horas de infecção a transcrição do
gene da IL-18 foi significativamente superior à transcrição basal apenas em
camundongos infectados com EIEC (p=O,006). EIEC induziu maior
transcrição deste gene que Ecoli K12 (p=O,04). Após 48 horas, observamos
que a única bactéria que induziu um aumento significativo na transcrição
deste gene foi Ecoli K12 (p=O,04) (fig. 4.25).
IL-18 iNOS KO
o ~ ~ ~ 'b'<::- c§' <§'~ ,,':> ,,'I- ~bl ,:>b/ bl
-$3 ~ ~ RJ<:> ~" ~'I-0" ~ ,,'I:«<?) ~0«
1,4 1I
1,2'"É 1o: 0,8
~ 0,6co:::; 0,4- 0,2
°o ~ ~ ~ 'b'<::- c§' 0..'<::-~ ':> ~ bl bl ~"V
RJ<:> ~" ~ ~ ,,':> ,,'I-~ ,,'I: -$3 ~ ~
~0 0"« «<?)
1,4 l IL-181,2 ~
'".!: 113: 0,8"=~ 0,6
:::; 0,4
0,2
°
Fig. 4.25 - Transcrição relativa do gene de IL-18 por células peritoniais de camundongos
C57BU6 e C57BU6 iNOS KO após 4 e 48 horas de infecção, avaliada por RT-PCR
semiquantitativo. Os dados representam médias e desvios padrão de dois experimentos
independentes.
Em camundongos selvagens, a transcrição do gene de IFN-y foi
significativamente maior que a transcrição basal após 4 horas de infecção
com Shigella (p=O,004) e EIEC (p=O,01); não houve diferenças significativas
na transcrição deste gene por células de camundongos infectados com
Shigella, EIEC e Ecoli K12. Após 48 horas, apenas camundongos
infectados com Shigella apresentaram aumento significativo na transcrição
deste gene (p=O,03), e neste tempo de infecção também não houve
diferenças significativas na transcrição entre as células de camundongos

74
infectados com as três bactérias (fig.4.26). Na infecção de camundongos
nocaute, o aumento da transcrição do gene de IFN-y em relação à
transcrição basal só foi significativo em células de camundongos infectados
por 4 horas com EIEC (0,006). O aumento na transcrição não foi significativo
após 48 horas de infecção com nenhuma bactéria, provavelmente devido
aos altos valores dos desvios padrão (fig.4.26).
1,4IFN-y
1,6IFN- Y iNOS KO
1,2
1,4
coc 1
ê 1,2
·ü~ 0,8
n 1co
':li 06
ª 0,8
- '~ 0,4
coZ 0,6
-Li.
0,2
- 0,4
o0,2
o~, ~ ~ ~ 'b'<:- <§' <§'
~ ,,~ ,,'l- :<...1>< ~I>< bI..fJ ~ +: Rl<::l ~" +-'l-
0" ~ ,,'V~«) ~0
~
o ~ ~ ~ 'b'<:- <§' ~:<... ~ ~ bI I>< ~v
Rl<::l ~" +- ~ '::.."~ ,,'l-~ ,,'V ..fJ l' +:
~0 0"~ ~«)
Fig. 4.26 - Transcrição relativa do gene de IFN-y por células peritoniais de camundongos
C57BU6 e C57BU6 iNOS KO após 4 e 48 horas de infecção, avaliada por RT-PCR
semiquantitativo. Os dados representam médias e desvios padrão de dois experimentos
independentes.
A transcrição do gene do receptor transmembrânico TLR4 em
camundongos selvagens foi maior em macrófagos infectados por Shigella
(p=0,03) e Ecoli K12 (p=O,OS) após 4 horas de infecção, em relação à
transcrição basa!. Shigella induziu maior transcrição deste gene que EIEC
(p=O,OS). Após 48 horas apenas Ecoli K12 (p=O,OS) induziu aumento
significativo da transcrição deste gene. Ecoli K12 induziu maior transcrição
do gene do TLR4 que EIEC (p=0,007) (fig. 4.27). Camundongos nocaute só
apresentaram aumento significativo na transcrição do gene de TLR4 após

75
48 horas de infecção por Ecoli K12 (p=O,02) em relação à transcrição basal
(fig. 4.27).
1,4
1,2
.~ 1Õ(1J 08(1J ,
~ 06~ ,
~ 0,4
I- 0,2
o
TLR4TLR4 iNOS KO
1,2
~ 1
~ 0,8
'* 0,6
~ 0,4
~ 0,2o
\À ~ ~ ~ co~ ~ &''- ~ ~ ~ ~ ~ b<V
RJ<::l ~.... +' sS' ....~ ....'1-~ ....'V $' #- ~
~0 0....« ««)
\À ~ ~ ~ co~ >--c§' ~'- ~ ~ ~ ~ '" ~
RJ<::l ~.... +' sS' ~....~ ....'1-~ ....'V $' ~ ~
~0 0....« ««)
Fig. 4.27 - Transcrição relativa do gene de TLR4 por células peritoniais de camundongos
C57BU6 e C57BU6 iNOS KO após 4 e 48 horas de infecção, avaliada por RT-PCR
semiquantitativo. Os dados representam médias e desvios padrão de dois experimentos
independentes. * - Diferenças estatisticamente significativas na transcrição do gene do
TLR4 entre macrófagos infectados por Shigella e EIEC (p<O,05 segundo o teste t não
pareado).
Após 4 horas de infecção a transcrição do gene da quimiocina IP-10
foi significativamente maior do que a transcrição basal na infecção por
Shigella (p=O,OOOS), EIEC (p=O,002) e Ecoli K12 (p=O,007), e, após 48
horas, o aumento na transcrição foi significativo apenas na infecção por
Shigella (p=O,OOS) e Ecoli K12 (p=O,04). Em camundongos nocauteados no

76
gene da iNOS houve aumento significativo da transcrição do gene de IP-10
após 4 horas de infecção por Shigella (p=O,003), EIEC (p=O,01) e E.coli K12
(p=O,009); após 48 horas, este aumento foi significativo na infecção por
EIEC (p=O,OS) e E.coli K12 (p=O,003) (figA.28).
2
1,8
1,6
~ 1,4
:g 12co '
J!! 1ro~ 0,8
~ 0,6
0,4
0,2
° I -
IP-10 2
1,8
1,6
~ 1,4
~ 1,2
J!! 1ro~ 0,8
~ 0,6
0,4
0,2
°I -
IP-10 iNOS KO
o ~~ ~~ ~ ~~ ':).~ ':).~RlÇ) ~" +,'1.- s} ",,,~ ,,'1.-~ ,,'V ~Oj rf ~
~v 0"« «(()
~, ~~ ~~ ~ ~~ $' $'RlÇ) ~" +,'1.- s} ,,~ ,,'1.-~ ,,'V $':f ~
~v 0"« «(()
Fig. 4.28 - Transcrição relativa do gene de IP-10 por células peritoniais de camundongos
C57BU6 e C57BU6 iNOS KO após 4 e 48 horas de infecção, avaliada por RT-PCR
semiquantitativo. Os dados representam médias e desvios padrão de dois experimentos
independentes.
o aumento na transcrição do gene da enzima iNOS em relação à
transcrição basal foi significativo após 4 horas de infecção por Shigella
(p=O,003) e EIEC (p=O,01). Shigella induziu maior transcrição deste gene
que EIEC (p=O,02). Após 48 horas de infecção houve aumento significativo
na transcrição do gene da iNOS apenas em camundongos infectados com

77
Shigella (p=O,02). Neste tempo de infecção não houve diferenças
significativas no aumento da transcrição deste gene entre as células
peritonias de camundongos infectados com as três bactérias (fig 4.29).
1,6 11,4
:g 1,2
U 1tU
~ 0,8tU
~ 0,6
~ 0,4
0,2
° I ..
iNOS
o ~ ~ ~ ~ ~ ~~ ~ ~ ~ ~ b<V
Rl<:::l ~..... ~ ~ .....~ ....."),~ ~ $' #- ~
~0 0.....~ ~<Q
Fig. 4.29 - Transcrição relativa do gene da iNOS por células peritoniais de camundongos
C57BU6 após 4 e 48 horas de infecção, avaliada por RT-PCR semiquantitativo. Os dados
representam médias e desvios padrão de dois experimentos independentes. * - Diferenças
estatisticamente significativas na transcrição do gene do iNOS entre macrófagos infectados
por Shigella e EIEC (p<O,05 segundo o teste t não pareado).
4.3.3 - Determinação da produção de TNF-a, IL-10 e IFN-y por ELISA
Os sobrenadantes dos lavados peritoniais de camundongos C57BU6
e C57BU6 iNOS nocaute, não infectados e infectados por 4 e 48 horas com
Shigella, EIEC e E.coli K12 (cuja transcrição dos genes das células foi

78
avaliada no ensaio anterior) foram utilizados no ELISA de captura para
determinação da concentração de TNF-a, IL-10 e IFN-y (figA.30 a 4.32).
Em relação às células peritoniais de camundongos selvagens não
infectados (produção basal da citocina), a produção de TNF-a foi
significativa após 4 horas de infecção por .Shigella (p=O,03) e EIEC
(p=O,0007). A produção de TNF-a foi maior em células de animais
infectados com Shigella do que em células infectadas com Ecoli K12
(p=O,04), e também maior na infecção por EIEC do que na infecção por
Ecoli K12 (p=O,002). A diferença na produção de TNF-a por camundongos
infectados por EIEC e Shigella foi pouco significativa (p=O,OS2),
provavelmente devido ao alto desvio padrão obtido nas dosagens de TNF-a
na infecção por Shigella. Após 48 horas de infecção houve aumento
significativo da produção de TNF por células de camundongos infectados
por Shigella (p-O,OOOS), EIEC (p=O,0009) e Ecoli K12 (p=O,03). Houve maior
produção de TNF por camundongos infectados com Shigella em relação
àqueles infectados com EIEC (p=O,002) e Ecoli K12 (p=O,008).
Os camudongos iNOS nocaute infectados por 4 horas com Shigella
(p=O,003), EIEC (p=O,003) e Ecoli K12 (p=O,04S) produziram
significativamente mais TNF-a comparando-se aos não-infectados, e a
diferença na produção desta citocina entre camundongos infectados com as
três bactérias não foi significativa. Após 48 horas de infecção houve
aumento significativo de produção de TNF-a na infecção por Shigella
(p=O,009) e EIEC (p=O,003). A produção desta citocina foi significativamente
maior na infecção por Shigella do que na infecção por EIEC (p=O,OS).

79
o ~ ~ ~ n.~ c§' c§'~ ~ ~ b<V ~ ~
$'<) #" '$:.+' 9Jç} ~,,~ .*-"'),0" (,0 ~ ,,'V ~
«<?) x,,' «<QV x,,'ií
TNF TNFiNOS KO
2400, ** 2000
1600
~ 1200a.
800
400
O
~ n.~ c§' ~~ b<V ~ ~+' ~ ,,~ ~
(§> -~<) cf' :s:,~x,,'ií ~. 0" <,o
«<?) x,,0
~ ~~ ~
$'<) #"0"
«<?)
2400 l
2oo0~I
1600
~ 1200a.
800
400
O
~,
Fig. 4.30 - Determinação, por ELISA de captura, da produção de TNF-a por células
peritoniais de camundongos C57BU6 e C57BLl6 iNOS KO após 4 e 48 horas de infecção.
Os dados representam médias e desvios padrão de duplicatas e são representativos de
dois experimentos independentes. * - Diferenças estatisticamente significativas na produção
de TNF-a entre macrófagos infectados por Shigella e EIEC (p<O,OS segundo o teste t não
pareado).
A produção de IL-10 foi maior após 4 horas de infecção por Shigella
(p=O,008), EIEC (p=O,005), e Ecoli K12 (p=O,001), em relação à produção
basal desta citocina. Houve maior produção de IL-10 em células de
camundongos infectados com Shigella do que naquelas infectadas com
Ecoli K12 (p=O,02); a produção desta citocina também foi maior na infecção
por EIEC do que por Ecoli K12 (p=O,0003). Após 48 horas de infecção a
produção de IL-10 foi significativamente superior à produção basal apenas
na infecção por Shigella (p=O,006). Camundongos infectados com Shigella
produziram mais IL-10 do que aqueles infectados com EIEC (p=O,005) e

80
Ecoli K12 (p=O,006) (fig. 4.31).
A produção de IL-10 após 4 horas de infecção em camundongos
nocaute foi significativamente superior à produção basal na infecção por
Shigella (p=O,002), EIEC (p=O,04) e Ecoli K12 (p=O,02). A produção de IL-
10 foi maior em camundongos infectados com Shigella do que naqueles
infectados por EIEC (p=O,001) e Ecoli K12 (p=O,001). Após 48 horas de
infecção não houve produção significativa de IL-10 em camundongos
infectados por qualquer uma das três bactérias (fig. 4.31).
6000, IL-10
IL-10 iNOS KO
7000
5000 *I *6000
5000
1 3000
E 4000
c-C»
2000
c. 3000
1000
2000
o
1000
oo ~~ ":J~ ~~ ~-<::- ~-<::- ';;.<§' \:.' ~ ~ ~ '1>"'<::- '0"'<::- '1>"'<::-
RI '::." .L" ~ ,,":J ,,'].o ~ ,,":J ,,'l- bI ":JbI n bI~Oj ~ :s,.' R>~ ~ .+ RlÇ) ~ .~ ~ ';:.." .LV
0" cP ~ ú"rv ~ ~ ,,')) ~ .~ ~ ~"«<Q <v. «<Q <v'? ~v <y'P ~. 0" cP« «<Q <y.
Fig. 4.31 - Determinação, por ELISA de captura, da produção de IL-10 por células
peritoniais de camundongos CS7BLl6 e CS7BLl6 iNOS KO após 4 e 48 horas de infecção.
Os dados representam médias e desvios padrão de duplicatas e são representativos de
dois experimentos independentes. * - Diferenças estatisticamente significativas na produção
de IL-10 entre macrófagos infectados por Shigella e EIEC (p<O,OS segundo o teste t não
pareado).
As células de camundongos selvagens infectados por 4 horas com
Shigella (p=O,007), EIEC (p=O,004) e Ecoli K12 (p=O,03) apresentaram
aumento significativo na produção de IFN-y em relação à produção basal

81
(fig. 4.32). Não houve diferenças na produção desta citocina entre
camundongos infectados com Shigella e EIEC, porém a diferença foi
significativa entre camundongos infectados com Shigella e Ecoli K12
(p=O,02), e EIEC e Ecoli K12 (p=O,009). Após 48 horas de infecção houve
aumento significativo da produção de IFN-y por células de camundongos
infectados por Shigella (p=IO,03) e EIEC (p=O,01). As células peritonias de
camundongos infectados com Shigella e EIEC produziram quantias
semelhantes de IFN-y, mas camundongos infectados com Shigella
produziram mais IFN-y do que aqueles infectados com Ecoli K12 (p=O,04).
Os camudongos iNOS nocaute infectados por 4 horas com Shigella
(p<O,0001), EIEC (p=O,03) e Ecoli K12 (p=O,04) produziram
significativamente mais IFN-y comparando-se aos não-infectados. Observou
se maior produção de IFN-y, na infecção por EIEC, se comparada à infecção
por Shigella (p=O,04) e Ecoli K12 (p=O,04). As células de camundongos
infectados com Shigella e Ecoli K12 produziram quantias semelhantes de
IFN-y. Após 48 horas de infecção houve aumento significativo de produção
de IFN-y apenas na infecção por Ecoli K12 (p=O,03) em relação à produção
basal; camundongos infectados por esta bactéria produziram mais IFN-y do
que os infectados por Shigella (p=O,002), e EIEC (p=O,002).

82
4500 1
4000
3500
IFN-y 30001600
1400 E 2500-..
1200 ~ 20001000
~ 800 1500O-
600 1000400
200500
O O
IFN-y iNOS KO
*
Fig. 4.32 - Determinação, por ELISA de captura, da produção de IFN-y por células
peritoniais de camundongos CS7BU6 e CS7BU6 iNOS KO após 4 e 48 horas de infecção.
Os dados representam médias e desvios padrão de duplicatas e são representativos de
dois experimentos independentes. * - Diferenças estatisticamente significativas na produção
de IFN-y entre macrófagos infectados por Shigella e EIEC (p<O,OS segundo o teste t não
pareado).
4.3.4 - Determinação da mortalidade de camundongos C57BU6 e C57BU6
iNOS KO após infecção
Lotes de quatro camundongos C57BU6 e C57BU6 iNOS KO foram
inoculados intraperitonialmente com suspensões de EIEC FBC124/13 e
Shigella M90T contendo 107 e 108 bactérias. Os resultados são mostrados
na tabela 1. Observa-se que a infecção por E.coli K12 não foi letal para os
camundongos em nenhuma das duas concentrações testadas. O inóculo de
108 foi letal para 100% dos animais selvagens e nocautes inoculados com

83
Shigella e EIEC, após 48 horas de infecção. O inóculo de 107 não foi letal
para os animais selvagens inoculados com EIEC e Shigella, porém foi letal
para 25% dos camundongos nocaute infectados com Shigella, e para 75%
dos camundongos nocaute infectados com EIEC.
C57BU6 C57BU6 iNOS KO
Shigella M90T (1 Of) 0% 25%
Shigella M90T (1 O!l) 100% 100%
EIEC FBC 124/13 (10 f) 0% 75%
EIEC FBC 124/13 (10!l) 100% 100%
E.coli K12 (10 f) 0% 0%
E.coli K12 (10!l) 0% 0%
Tabela 4.1 - Porcentagem de mortalidade em camundongos C57BL/6 e
C57BL/6 iNOS KO infectados com inóculos de 107 e 108 Shigella f1exneri
M90T, EIEC FBC124/13 e E.coli K12 por 48 horas.
4.4 - Resposta de linfócitos à infecção por EIEC
4.4.1 - Linfoproliferação
Neste ensaio foi avaliada a proliferação da população celular dos
linfonodos de animais previamente imunizados no coxim plantar com EIEC e
Shigella, e posteriormente estimulados com os antígenos solúveis secretados
por estas bactérias (fig 4.33) Para tanto, inicialmente foi obtida uma curva de
proliferação, onde foram testadas as concentrações antigênicas de 10 a 100

84
I-Ig (fig.23). Observou-se que, em relação aos controles negativos (linfócitos
de animais imunizados que não foram posteriormente estimulados com os
antígenos bacterianos) houve um aumento na proliferação de linfócitos de
animais imunizados com Shigella e EIEC e posteriormente estimulados com
os antígenos.
2 3
67 ...
43'" •
30 ...
20 ...
Fig. 4.33 - SOS-page dos antígenos secretados por EIEC FBC124/13 e Shigel/a M90T apóscinco horas de incubação em contato com células Hep-2. 1- Padrão de peso molecular; 2
Antígenos secretados por Shigel/a M90T (100 fl9); 3- Antígenos secretados por EIECFBC124/13 (100 fl9).
Entre os linfócitos de animais que foram imunizados com Shigella e
posteriormente estimulados com antígenos de Shigella este aumento foi
significativo quando os linfócitos foram estimulados com 20l-lg de antígenos
(p=0,019); 30l-lg (p=0,0038); 40l-lg (p=O,0005), 50l-lg (p<0,0001), 60l-lg
(p=O,0002), e 70l-lg, 80l-lg, 90l-lg e 100l-lg de antígenos (p=0,0001 para todos).
Entre os linfócitos de animais que foram imunizados com EIEC e
posteriormente estimulados com antígenos de EIEC este aumento foi
significativo quando os linfócitos foram estimulados com 10l-lg de antígenos
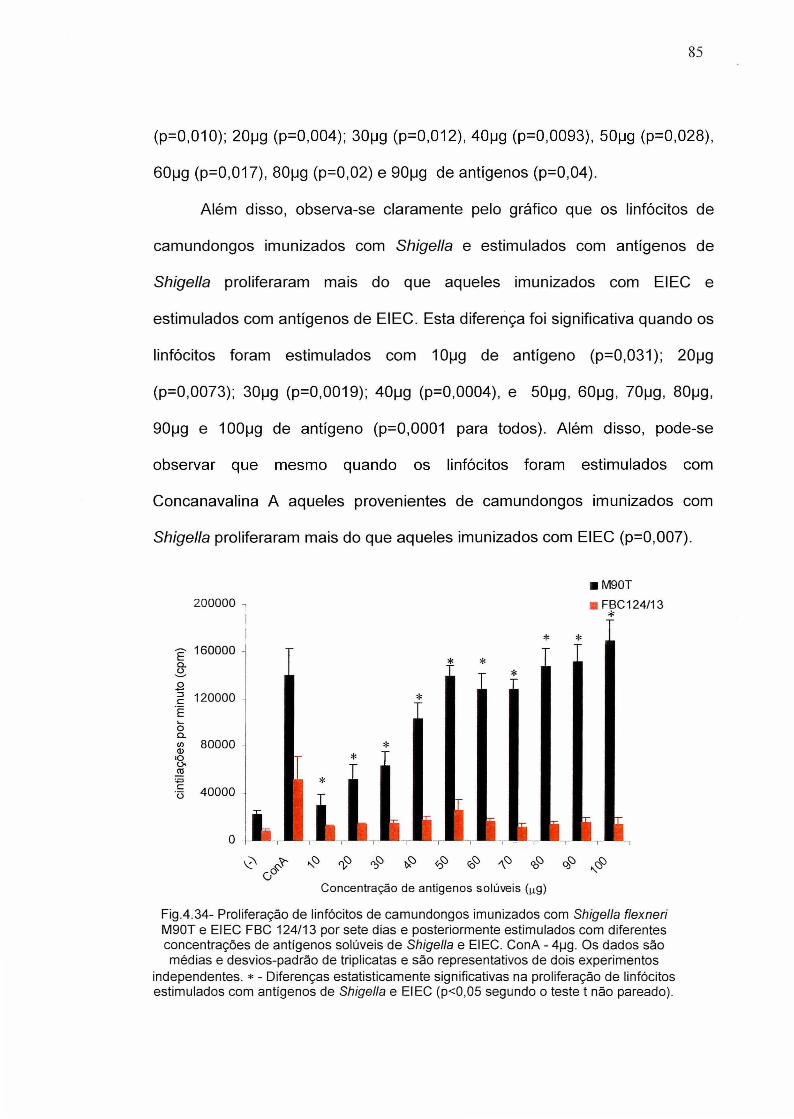
85
(p=O,010); 20jJg (p=O,004); 30jJg (p=O,012), 40jJg (p=O,0093), 50jJg (p=O,028),
60jJg (p=O,017), 80jJg (p=O,02) e 90jJg de antígenos (p=O,04).
Além disso, observa-se claramente pelo gráfico que os linfócitos de
camundongos imunizados com Shigella e estimulados com antígenos de
Shigella proliferaram mais do que aqueles imunizados com EIEC e
estimulados com antígenos de EIEC. Esta diferença foi significativa quando os
linfócitos foram estimulados com 10jJg de antígeno (p=O,031); 20jJg
(p=O,0073); 30jJg (p=O,0019); 40jJg (p=O,0004), e 50jJg, 60jJg, 70jJg, 80jJg,
90jJg e 100jJg de antígeno (p=O,0001 para todos). Além disso, pode-se
observar que mesmo quando os linfócitos foram estimulados com
Concanavalina A aqueles provenientes de camundongos imunizados com
Shigella proliferaram mais do que aqueles imunizados com EIEC (p=O,007).
200000 -,
II * *
• M90T
• FBC124/13
*
Ê 160000a.~
.8~ 120000·E *
* **
80000
l **
* I40000
o -r----,----,----,----,----,----,----,----,-...".-,-
\:.' f;:-'?- "Ç) '1"Ç) fl:JÇ) b<Ç) ~Ç) roÇ) "Ç) coÇ) q,Ç) Ç)Ç)
cP "Concentração de antígenos solúveis (I-Ig)
L-
oa.Cf)Q)
100ro
E·13
FigA.34- Proliferação de linfócitos de camundongos imunizados com Shigella fiexneriM90T e EIEC FBC 124/13 por sete dias e posteriormente estimulados com diferentesconcentrações de antígenos solúveis de Shigella e EIEC. CanA - 41Jg. Os dados sãomédias e desvios-padrão de triplicatas e são representativos de dois experimentos
independentes. * - Diferenças estatisticamente significativas na proliferação de linfócitosestimulados com antígenos de Shigella e EIEC (p<O,05 segundo o teste t não pareado).

86
Avaliou-se posteriormente a capacidade de proliferação linfocitária após
estímulo com antígenos secretados por uma bactéria diferente da que o
animal foi imunizado. Os linfócitos de animais inoculados com Shigella foram
estimulados com antígenos de Shigella, EIEC e Ecoli K12, o mesmo
acontecendo com os linfócitos de animais inoculados com EIEC e Ecoli K12
(fig.24).
Entre os linfócitos de camundongos imunizados com Shigella, o maior
índice de proliferação ocorreu após estímulo com antígenos de Shigella
(p<O,0001), seguida do estímulo com antígenos de Ecoli K12 (p=O,002). O
menor índice de proliferação ocorreu após estimulo com antígenos de EIEC
(p=O,0004), comparando-se aos controles negativos. As diferenças entre os
índices de proliferação foram significativas entre linfócitos estimulados com
antígenos de Shigella e EIEC (p=O,0016) e Shigella e Ecoli K12 (p=O,0196), e
pouco significativas entre linfócitos estimulados com antígenos de EIEC e
Ecoli K12 (p=O,065).
Entre os linfócitos de animais imunizados com EIEC, observou-se
comportamento semelhante. O maior índice de proliferação ocorreu após
estímulo com antígenos de Shigella (p=O,0004), seguido do estímulo com
antígenos de Ecoli K12 (p=O,001) e EIEC (p=O,001). Neste caso, as
diferenças entre os índices de proliferação foram significativas entre linfócitos
estimulados com antígenos de Shigella e EIEC (p=O,0006) e EIEC e Ecoli
K12 (p=O,00180). Não houve diferenças significativas entre os índices de
Iinfoproliferação induzidos pelos antígenos de Shigella e Ecoli K12.

87
Finalmente, os linfócitos de camundongos imunizados com antígenos
de Ecoli K12 apresentaram maior índice de proliferação quando estimulados
com antígenos de Ecoli K12 (p<O,0001), seguido dos estimulados com
antígenos de Shigella (p=O,0001), e EIEC (p=O,001). Novamente não houve
diferenças significativas entre os índices de linfoproliferação induzidos pelos
antígenos de Shigella e Ecoli K12. Porém, a linfoproliferação induzida pelos
antígenos de Shigella (p=O,0004) e Ecoli K12 (p=O,0003) foram
significativamente maiores do que a induzida pelos antígenos de EIEC.
Neste ensaio foi utilizada a fitohemaglutinina como controle positivo, e
os linfócitos dos camundongos imunizados com as três bactérias proliferaram
de maneira semelhante após este estímulo (a diferença entre estes índices de
proliferação não foi significativa).

88
Shigella M90T E1EC FBC 124/13
~, * ~l~000 6000050000 50000
E 40000 í E 40000
~~ _. I ~:~j í. - •~\ :A~ [):,ç} ~,,":> ~'), ~\ ;<::-4.,,0 _~ç} ~,,":> "~'"
«~ ~ ,,'I: ~ « ~. ,(I) (j>ú cP ~ú ,-,Y
~<Q <v" ~ v
antígenos solúveis (100 I1g) antígenos solúveis (100 I1g)
E.coli K1270000~000
50000
~=j li2000010000
O
"~'),(j>
<vY
~~~ ~,,":>0,(1)
~<Q
I...*
_~o
«~.,~\
antigenos solúveis (100 Ilg)
Fig. 4.35 - Proliferação de linfócitos de camundongos imunizados com Shigella flexneri
M90T, EIEC FBC124/13 e E.coli K12 por sete dias e posteriormente estimulados com
100 /lg de antígenos solúveis de M90T, EIEC e K12. Fitohemaglutinina - 41Jg. Os dados
representam média e desvio padrão de triplicatas e são representativos de um
experimento. * - Diferenças estatisticamente significativas na proliferação de linfócitos
estimulados com antígenos de Shigella e EIEC (p<0,05 segundo o teste t não pareado).
4.4.2 - Produção de IFN-y, IL-10 e IL-4
o resultados podem ser vistos na figura 9. O ELISA de captura
detectou produção de IFN-y e IL-10 apenas no sobrenadante de linfócitos de
camundongos imunizados com Shigel/a e estimulados por 72 horas com 100
J.lg de antígenos solúveis de Shigella. Os linfócitos de animais imunizados com
EIEC não produziram estas citocinas "nem mesmo quando estimulados com
100 J.lg de antígenos solúveis de EIEC. Não foi detectada produção de IL-4,
por linfócitos de camundongos imunizados com Shigel/a e EIEC, tanto nos

89
controles negativos quanto nos linfócitos estimulados por 72 horas com os
antígenos (dados não mostrados).
1FN-r
'\~~"O:>
,,<f«<Qv
o~"O:>
0"rv«q)
,\rV'~
$f:::>
1200
1000
250] IL-10800
200E 600C,
150 . Do
E
~100j I400
20050
OI O\(, rv'<:' 0:>0 '\~ \('S' ~'\
~" "O:> S'$3 ~~f:::> ,,'V ",f- $30«q) ~v«
Fig. 4.36 - Produção, de IL-1ü e IFN-y por linfócitos de camundongos Balb/C imunizados com
Shigella f1exneri M9üT e EIEC FBC 124/13 e posteriormente estimulados por 72h com
antígenos solúveis de Shigella e EIEC, respectivamente. Os dados representam médias e
desvios padrão de duplicatas e são representativos de um experimento.
4.2.4 - Determinação das subpopulações de linfócitos
As células dos linfonodos de animais imunizados com Shigella e EIEC
foram marcadas com anticorpos anti-CD3 e anti-CD4, ou anti-CD3 e anti-CD8.
Os resultados obtidos através de citometria de fluxo podem ser observados
na figura 8. Pode -se observar através dos gráficos que tanto em linfonodos
de animais imunizados com Shigella quanto com EIEC a população linfocitária
predominante é a de TCD4+ (histogramas A e C). A porcentagem de linfócitos
TC8+ é menor do que a de TCD4+ nos linfonodos de animais inoculados com
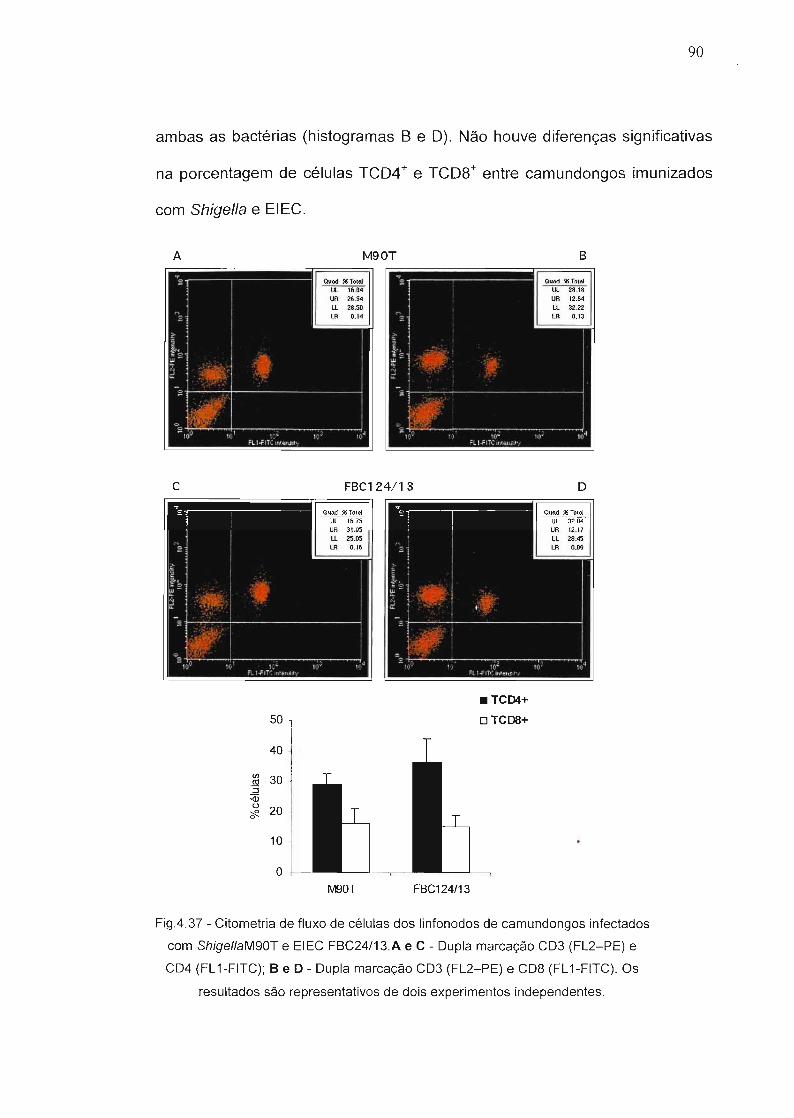
90
ambas as bactérias (histogramas B e O). Não houve diferenças significativas
na porcentagem de células TC04+ e TCOS+ entre camundongos imunizados
com Shigella e EIEC.
A M90T B
c FBC124/13 D
50 1
I40 -
12 30::J
:a>~ 20o
10
orv190T F8C124/13
• TCD4+
o TC08+
Fig.4.37 - Citometria de fluxo de células dos linfonodos de camundongos infectados
com Shigel/aM9üT e EIEC FBC24/13.A e C - Dupla marcação CD3 (FL2-PE) e
CD4 (FL1-FITC); B e D - Dupla marcação CD3 (FL2-PE) e CD8 (FL1-FITC). Os
resultados são representativos de dois experimentos independentes.

91
5 - DISCUSSÃO
Shigella sp e Escherichia coli enteroinvasora são os agentes
etiológicos da disenteria bacilar, doença intestinal aguda com incidência de
aproximadamente 160 milhões de casos anuais, 99% deles em países
subdesenvolvidos (KOTLOFF e coL, 1999; SANSONETTI, 2001).
EIEC e Shigella apresentam o mesmo perfil bioquímico, o mesmo
plasmídeo de virulência e causam síndromes semelhantes, porém, a dose
infectante de EIEC é muito maior. Os estudos sobre a relação entre EIEC e
o hospedeiro são praticamente inexistentes; nossa proposta, portanto, foi a
de elucidar alguns aspectos desta interação. Foram avaliados parâmetros da
resposta imune inata, como fagocitose e escape bacteriano de macrófagos,
influxo celular para o peritônio após infecção e produção de citocinas e óxido
nítrico por macrófagos infectados. Além disso, a resposta imune adaptativa
foi avaliada através da proliferação e produção de citocinas por linfócitos
ativados. Estes estudos foram realizados utilizando-se modelos murinos, in
vitro e in vivo, e comparativamente a Shigella.
Os primeiros estudos foram conduzidos em macrófagos murinos da
linhagem J774. Este modelo de infecção foi utilizado em uma série de
estudos envolvendo apoptose e produção de citocinas na infecção por
Shigella e outros patógenos, como Mycobacterium tuberculosis, Salmonella
typhimurium e Escherichia colí enteroagregativa (ZYCHLlNSKI e coL, 1992;
FERNANDEZ-PRADA e coL, 1998; MUSSO e coL, 2001; SERAFíN-LOPEZ
e coL, 2004; KALUPAHANA e coL, 2005).

92
A fagocitose foi avaliada pela determinação do número de bactérias
viáveis intracelulares após infecção (fig.4.1). Não foram observadas
diferenças na captura inicial das duas bactérias, uma vez que no tempo zero
(as monocamadas foram infectadas, centrifugadas e em seguida lavadas e
incubadas com gentamicina) o número de UFC de Shigella e EIEC no
interior do macrófago foi praticamente o mesmo. Após 30 minutos e uma
hora de infecção, contudo, há um número significativamente maior de UFC
de EIEC no interior dos macrófagos em relação a Shigella. Estes resultados
são similares aos obtidos por FERNANDEZ-PRADA e colaboradores (1997),
que observaram um maior número de UFC de amostras avirulentas de
Shigella flexneri em relação à amostra virulenta.
Para determinar se as diferenças no número de UFC no interior dos
macrófagos eram resultado de sua ação microbicida, ou do escape
bacteriano, foi feita também a determinação do número de bactérias viáveis
no sobrenadante após infecção (fig.4.2). Os dados obtidos mostram que
EIEC e Shigella escapam do macrófago, e corroboram os resultados
anteriores, uma vez que o número de UFC de Shigella no sobrenadante foi
significativamente maior que o número de UFC de EIEC após 30, 60 e 120
minutos de incubação em meio sem gentamicina.
Foi demonstrado que Shigella causa morte dos macrófagos. Os
estudos conduzidos com monócitos humanos são controversos, uma vez
que alguns autores afirmam que esta morte ocorre por oncose
(FERNANDEZ-PRADA e coL, 1998), enquanto outros afirmam ocorrer por

93
apoptose (HATHAWAY e coL, 2002). Porém, em macrófagos murinos, a
indução de apoptose na infecção por Shigella flexneri está bem
documentada (ZYCHLlNSKI, 1992; FERNANDEZ-PRADA, 1998;
SANSONETII,2000).
Observou-se em nosso estudo que a infecção com EIEC e Shigella
alterou a viabilidade dos macrófagos, o que provavelmente ocorreu em
decorrência do escape bacteriano (fig. 4.3). A porcentagem de células não
viáveis foi dependente do tempo de infecção. EIEC apresentou
significativamente menor capacidade de alteração da viabilidade que
Shigella, após 2, 4 e 6 horas de infecção. Nos tempos de 2 e 4 horas,
especialmente, nota-se que há o dobro de macrófagos não viáveis na
infecção por Shigella, confirmando mais uma vez o maior potencial
patogênico desta bactéria.
A ativação de macrófagos é essencial na resposta inflamatória, tanto por
sua atividade microbicida quanto pelo estímulo da secreção de citocinas e
outros mediadores que orquestram a resposta imune (PARSOT &
SANSONETII, 1995; FERNANDEZ-PRADA, 1997; SANSONETII, 2001).
Em nosso estudo foi caracterizada a cinética de transcrição dos genes
das citocinas TNF-a, IL-1 e IL-10 por macrófagos J774 infectados com EIEC e
Shigella. Houve um aumento significativo da transcrição do mRNA da citocina
TNF-a, após uma, três e sete horas de infecção por Shigella, e após uma e
sete horas de infecção por EIEC, em relação à transcrição basal (fig. 4.4). A
transcrição do mRNA de IL-1 foi significativa após três e sete horas de infecção
por Shigella e EIEC (fig. 4.5). Por fim, somente macrófagos infectados com

94
EIEC mostraram aumento significativo na transcrição do mRNA de IL-10 em
relação à transcrição basal (fig. 4.6). Não foram encontradas diferenças
significativas, no entanto, na transcrição do mRNA das três citocinas entre
macrófagos infectados com Shigella e EI EC. A produção destas citocinas foi
confirmada por ELISA.
A rápida produção in vitro de TNF-a e IL-1 por macrófagos em resposta
ao estímulo bacteriano está de acordo com a função destas citocinas na fase
aguda da resposta imune. A síntese de TNF-a está associada ao influxo celular
e também ao aumento das funções microbicidas do próprio macrófago, num
processo de ativação autócrina. Além disso, D'HAUTEVILLE e col. (2002)
observaram que amostras de S. flexneri que apresentavam mutações no
lipídeo A induziam menor produção de TNF-a do que as amostras selvagens,
resultando em inflamação menos severa e preservação da barreira intestinal. A
produção de IL-1 estimula o recrutamento de leucócitos e induz a produção de
outras citocinas (SERAFiN-LOPEZ e col., 2004). O papel da IL-1 no processo
de migração celular relacionado à ruptura da barreira intestinal na shigelose
está bem estabelecido (SANSONETTI, 2000). A produção de IL-10
provavelmente representa uma tentativa da célula em evitar uma resposta
imune descontrolada (DEMUTH e col., 1996).
Os resultados obtidos nos ensaios in vitro, analisados em conjunto,
demonstraram que o encontro de EIEC com macrófagos J774 resulta em
fagocitose; a bactéria escapa do macrófago induzindo apoptose, e este
processo é acompanhado de síntese de citocinas TNF-a, IL-1 e IL-10.

95
Ao comparar a interação dos macrófagos com EIEC e Shigella pode-se
sugerir que esta última apresenta menor capacidade de escape e indução de
apoptose do que Shigella, apesar da fagocitose ocorrer de maneira
semelhante. EIEC, portanto, ficaria por mais tempo no interior do macrófago,
estando sujeita à ação microbicida da célula. Não foram observadas
diferenças, no entanto, no estímulo à síntese de citocinas, tanto nos ensaios
de PCR em tempo real quanto no ELISA.
No modelo de infecção que utiliza macrófagos J774, portanto, as
diferenças patogênicas entre EIEC e Shigella podem ser explicadas pela
maior permanência de EIEC no macrófago, mas não por diferenças na
produção de citocinas. Este resultado é surpreendente, pois se deveria
esperar que a bactéria mais patogênica estimulasse uma maior produção de
citocinas, ou, pelo menos, uma maior produção de IL-1, uma vez que a
ligação da IpaB com a caspase-1 resulta em ativação da cascata apoptótica e
também na clivagem do precursor de IL-1 em sua forma ativa (ARONDEL e
col., 1999; SANSONETTI & DI SANTO, 2007). Porém, é importante
considerar que os macrófagos J774 apresentam diferenças fisiológicas de
macrófagos provenientes de animais. Além disso, em um processo infeccioso,
a produção de citocinas é regulada por múltiplos fatores além da presença do
patógeno.
Os estudos com linhagens celulares são bastante úteis na avaliação
da relação entre o microrganismo e o hospedeiro. Este modelo apresenta
algumas vantagens em relação a estudos com animais, como a relativa
facilidade de manipulação e a garantia de obtenção de um número

96
abundante de células. No entanto, apresentam limitações, por se tratar de
células modificadas e também porque a interação entre o patógeno e o
hospedeiro ocorre em um ambiente fechado, sem a influência do
microambiente. Assim, nos propusemos a avaliar outro modelo de infecção,
estudando a interação entre EIEC e macrófagos de camundongos C57BU6.
Inicialmente foi avaliada a produção de NO por macrófagos peritoniais
de camundongos C57BU6 e C57BU6 nocauteados no gene da iNOS.
Observou-se menor produção de NO por células infectadas com EIEC,
comparativamente a Shigella, e a produção foi dependente do inóculo
bacteriano (fig. 4.8).
Uma vez que houve diferenças na produção de NO na infecção pelas
duas bactérias, nos propusemos a avaliar o papel do NO na resposta do
hospedeiro à infecção por EIEC. Para tanto, inicialmente foi comparada a
transcrição dos genes das citocinas IL-1, TNF-a, IL-10, IL-18, da enzima iNOs
e do receptor transmembrânico TLR4 por macrófagos de camundongos
C57BU6 selvagens e nocauteados no gene da iNOS, após 4 horas de infecção
por Shigella EIEC e E. colí K12. Nosso objetivo foi o de avaliar se a diferença
na produção de NO estaria associada a diferenças na produção de citocinas.
E.coli K12, uma Escherichia coli que não apresenta plasmídeo de invasão, foi
acrescentada ao ensaio para avaliar se a ativação transcricional do macrófago
seria dependente apenas do LPS ou também da secreção dos antígenos de
invasão pelo SSTI. Em nossos ensaios também foi utilizado LPS como
controle positivo da transcrição, uma vez que PERERA e coI. (2001)

97
descreveram aumento na transcrição de genes como os de TNF-a e iNOS por
macrófagos de camundongos C57BU6 estimulados com LPS.
Observou-se um aumento significativo da transcrição dos genes de
TNF-a, IL-10, IL-1 e IL-18 em macrófagos de camundongos C57BU6
selvagens infectados com EIEC, enquanto os camundongos infectados com
Shigella apresentaram aumento significativo na transcrição dos mesmos
genes, além de iNOS (figs. 4.9 a 4.14). Macrófagos infectados com Ecoli
K12 apresentaram aumento apenas na transcrição dos genes de IL-1 e iNOS.
Estes resultados estão em acordo com os obtidos nos ensaios com
macrófagos J774, mostrando que a infecção bacteriana induz a ativação da
transcrição de citocinas pelos macrófagos. No entanto, foi observado que
EIEC induz uma transcrição significativamente maior de TNF-a e IL-10,
comparativamente a Shigella, e que esta última induz maior ativação da
transcrição da enzima iNOS do que EIEC. Demonstra-se, assim, que a maior
produção de NO por macrófagos infectados com Shigella é decorrente de
uma maior transcrição da enzima NO sintase induzível.
Um resultado interessante é a transcrição significativa do gene do
TLR4 por macrófagos infectados com Ecoli K12, o que não ocorreu na
infecção por Shigella e EIEC. Este resultado sinaliza que a ativação
transcricional dos macrófagos pela Ecoli K12 ocorre primordialmente pela
ação do LPS, enquanto que as bactérias invasoras ativam a transcrição dos
genes das citocinas também por outras vias, provavelmente relacionadas aos
antígenos de invasão (SAMANDARI e col., 2000).

98
Os ensaios conduzidos com macrófagos de camundongos iNOS
nocaute não apresentaram resultados satisfatórios, uma vez que a análise
estatística foi prejudicada pelas diferenças entre os dois experimentos
independentes. Apesar disso, pode-se sugerir que a produção elevada de IL
10 pelos macrófagos dos camundongos nocaute após infecção, observada no
ensaio de ELISA de captura, inibiu a produção das citocinas pró-inflamatórias
(fig. 4.16). Foi demonstrado que a produção de IL-10 por monócitos humanos
inibiu a transcrição de genes de citocinas pró-inflamatóiias pelos próprios
monócitos, num mecanismo de regulação autócrina (DE WAAL MALEFYT e
coL, 1991).
Estes resultados, analisados em conjunto, nos permitem sugerir que
em macrófagos de camundongos C57BU6 EIEC estimula uma maior
produção de TNF-a e IL-10 (dados corroborados pelos resultados obtidos no
ensaio de ELISA). Este resultado novamente nos surpreendeu porque,
embora haja diferenças no estímulo à síntese de citocinas pelos macrófagos
infectados com as duas bactérias, a bactéria que mais estimulou a síntese de
TNF-a e IL-1D foi justamente a que provoca um quadro mais brando de
disenteria. Este dado poderia ser sugestivo de que embora a produção de
TNF-a seja necessária para a patogênese da infecção, a produção de IL-1D
estaria relacionada a uma maior resposta anti-inflamatória do hospedeiro na
infecção por EIEC, comparativamente a Shigella;
Num modelo in vitra, que analisa isoladamente a relação entre
macrófagos e patógenos, a produção de citocinas poderia ou não estar
relacionada com a maior patogenicidade da bactéria. Ao analisar a transcrição

99
de genes de citocinas por macrófagos derivados de medula óssea infectados
com diferentes genotipos de M. tuberculosis, CHACON-SALlNAS e col.
(2005) observaram que o genotipo mais virulento foi o que induziu maior
transcrição das citocinas pró-inflamatórias, enquanto a transcrição de IL-10 foi
pequena. DEMUTH e col. (1996) observaram que o aumento na transcrição
dos genes de IL-1, TNF-a e IL-10 em macrófagos derivados de medula óssea
foi semelhante na infecção pela espécie virulenta L. monocytogenes e pela
avirulenta L. innocua. Estudos conduzidos com monócitos humanos
mostraram que a infecção com uma cepa de Shigella flexneri M90T selvagem
inibiu a transcrição dos genes de IL-1 e TNF-a, enquanto a infecção com uma
cepa de M90T sem plasmídeo de invasão induziu a transcrição destes genes
(HATHAWAYe col., 2002).
Sabe-se, que, in vivo, a interação de alguns enteropatógenos com os
macrófagos residentes da lâmina própria é fundamental para o
estabelecimento da infecção. Yersinia, por exemplo, pode inibir a fagocitose,
ou ser fagocitada e induzir a apoptose do macrófago com concomitante
bloqueio da produção de TNF-a, garantindo sua sobrevivência; Salmonella
pode promover apoptose do macrófago ou modificar o vacúolo fagocítico de
modo a propiciar a sobrevivência no seu interior; Shigella por outro lado, induz
apoptose do macrófago com liberação de citocinas pró-inflamatórias, ativando
assim a resposta imune.
Na infecção por Shigella, a elicitação da resposta imune resulta no
rompimento da barreira, facilitando a invasão nas áreas adjacentes ao ponto
inicial de translocação (SANSONETTI, 2004). Por outro lado, as células

[; I ~: i.. I O r r: c AFaculdade de Ciências Farmacêutici]~
Universidade de São Pauln
100
recém-migradas, como neutrófilos polimorfonucleares e monócitos, podem
eliminar a bactéria. Numa tentativa de entender a importância das diferentes
linhagens celulares na resposta à infecção e avaliar possíveis diferenças
entre o padrão de resposta gerado por EIEC e Shigella foram iniciados
estudos de indução de resposta in vivo.
Inicialmente determinou-se, por citometria de fluxo, a população celular
presente no peritônio de camundongos C57BU6 não infectados e infectados
por 12, 24 e 48 horas com Shigella e EIEC (fig.4.17).
Em animais não infectados, observou-se que a população
predominante é a de macrófagos (os macrófagos peritoniais residentes),~
~ seguida de linfócitos, provavelmente linfócitos B1 (YANG e coL, 2007). Em%~
animais não infectados praticamente não foram encontrados neutrófilos nesta
cavidade (cerca de 2% da população total).
Houve um aumento de uma ordem de grandeza (100 para 10') no
número de células da cavidade peritoniaL O influxo foi predominantemente de
neutrófilos, na infecção pelas duas bactérias. A migração de neutrófilos para o
peritônio em resposta à infecção por outros enteropatógenos já foi
documentada (YANG e coL, 2002).
Neutrófilos polimorfonucleares são células citotóxicas que têm papel
importante na eliminação bacteriana. Os patógenos fagocitados são
eliminados pela ação de enzimas proteolíticas e espécies reativas de oxigênio
(BRINKMANN e coL, 2004). Foi demonstrado que a elastase produzida pelos
neutrófilos degrada antígenos como as Ipas produzidas por Shigella; além
disso, os neutrófilos aprisionam a bactéria em estruturas filamentosas

101
extracelulares ricas em moléculas antimicrobianas, conhecidas como
"neutrophil extracellular traps" (NETs) (WEINNRAUCH e coL, 2002;
BRINKMANN e coL, 2004). Estas células, portanto, não apenas contribuem
para a ruptura da barreira intestinal, mas também participam da resolução da
infecção. Na infecção por Shigella, o número de neutrófilos permaneceu alto,
mesmo após 48 horas de infecção, porém, em animais infectados com EIEC,
o número destas células começa a diminuir a partir de 24 horas de infecção.
A população de linfócitos sofreu uma redução em seu número após
infecção por Shigella e EIEC, sugerindo sua morte por apoptose.
ZICHLlNSKY e coL (1996) demonstraram a ocorrência de apoptose de
linfócitos T e B na infecção por Shigella in vivo. A partir de 12 horas, na
infecção por EIEC, e 24 horas, na infecção por Shigella, o número de linfócitos
começa a se aproximar dos níveis basais, o que sugere o recrutamento de
células B e T para o foco da infecção.
O número de macrófagos praticamente não se altera nos primeiros
tempos de infecção, porém a partir de 24 horas de infecção observa-se um
aumento no número destas células.
No que diz respeito à expressão de moléculas de ativação por
macrófagos, foi possível observar que a infecção por EIEC e Shigella induziu
um aumento na expressão de MHCII e C080 (figs. 4.18 e 4.19), e uma
diminuição da expressão de C086 (fig. 4.20). Enquanto a modulação positiva
de MHCII e C080 foi maior na infecção por Shigella, a modulação negativa de
C086 foi maior na infecção por EIEC.

102
o MHCII e as moléculas co-estimulatórias participam da apresentação
do antígeno ao linfócito T são um indicativo do estado de ativação dos
macrófagos.
Macrófagos peritoniais de camundongos infectados com Neisseria
meningifidis apresentaram modulação positiva da expressão de C080 e C086,
o mesmo ocorrendo com macrófagos J774 infectados com Salmonella
Typhimurium (Mukhopadhyay e coL, 2004; Kalupahana e col., 2005). ALBA
SOTO e coL, (2003), no entanto, mostraram que macrófagos e células
dendríticas de camundongos infectados com Tripanosoma cruzi apresentaram
modulação negativa da expressão de C086. A ligação das moléculas co
estimulatórias C080 e C086 a seus ligantes C028 e CTLA-4, expressos em
linfócitos, ocorre em uma série de processos infecciosos (LENSCHOW e coL,
1996; ALBA SOTO e coL, 2003). FREEMAN e coL (1993) demonstraram que a
expressão de C086 ocorre mais precocemente do que a expressão de C080.
Os autores sugerem que a ligação de C086 a C028/CTLA-4 influencia se o
linfócito T vai desenvolver uma resposta eficiente ou vai se tornar anérgico.
Sua modulação negativa, portanto, pode contribuir para supressão da resposta
imune. (ALBA SOTO e coL, 2003).
Nota-se também uma diminuição da expressão de F4/80 (figA.21). Este
marcador é especifico para macrófagos e já foi demonstrado que macrófagos
residentes do peritônio expressam níveis elevados de F4/80 (GOROON, 1998;
HAMERMAN, 2001). A diminuição da expressão em 12 horas com aumento nos
tempos subseqüentes representaria um aumento de monócitos recém-migrados
em resposta ao estímulo inflamatório. Portanto, a partir dos dados obtidos,

103
pode-se inferir que os macrófagos residentes são mortos após a infecção
bacteriana, e tem início uma migração de monócitos.
Os resultados obtidos na caracterização das células envolvidas na
infecção por Shigella e EIEC demonstram que estas duas bactérias, em um
primeiro momento, estimulam de maneira semelhante o influxo celular. Os
macrófagos, no entanto, parecem estar menos ativados em camundongos
infectados com EIEC, se compararmos a modulação da transcrição do MHCII e
moléculas co-estimulatórias. Observa-se também que na infecção por EIEC o
número de granulócitos começa a diminuir a partir de 24 horas, o número de
linfócitos é restaurado ao valor basal após 24 horas e, após 48 horas, o número
de macrófagos e a transcrição de F4/80 também estão mais próximos dos
valores basais do que na infecção por Shigella. Estes dados sugerem que a
resolução da infecção ocorre mais precocemente em animais infectados com
EIEC.
O padrão de citocinas produzidas pelas células componentes da
imunidade inata influencia o perfil da resposta imune adaptativa subseqüente
(MEDZHITOV & JANEWAY, 2000). Uma série de citocinas está envolvida na
patogênese da infecção por Shigella. Indivíduos portadores de shigelose
apresentaram produção de IL-1a, IL-1p, TNF-a, IL-6, IL-4, IL-8, IL-10, IFN-y e
TGF-p (RAQIB e coL, 1995). O papel de certas citocinas, como IL-1, IFN-y e IL
18 no processo inflamatório desencadeado por Shigella está bem
documentado. A IL-1, como já foi dito, contribui para a colonização bacteriana,
graças ao estímulo ao influxo celular. O IFN-y, por sua vez, é fundamental no
controle da infecção, como foi demonstrado elegantemente por LE-BARILLEC

104
e col. (2005). A IL-18 seria necessária para gerar uma resposta inflamatória
capaz de erradicar o patógeno, uma vez que camundongos nocauteados no
gene desta citocina não controlam o crescimento bacteriano, e,
conseqüentemente, a inflamação. (SANSONETTI e coI. , 2000). A IL-18,
juntamente com a IL-12, induz a produção de IFN-y, ativando uma resposta do
tipo Th 1. Com o propósito de estabelecer o perfil de citocinas ligado ao
desenvolvimento da imunidade a EIEC, foi feita a determinação da transcrição
dos genes das citocinas IL-1, TNF-u, IL-10, IL-18, IFN-y, IP-10, da enzima iNOs
e do receptor transmembrânico TLR4 por células peritoniais totais de
camundongos não infectados e infectados com EIEC, Shigella e E.coli K12 por
4 e 48 horas (figs. 4.22 a 4.29). Avaliou-se, desta maneira, a produção in vivo
de citocinas pelas células residentes do peritônio, logo após o desafio com os
patógenos (tempo de 4 horas), e também pelas células que migraram para o
peritônio em resposta ao estímulo inflamatório (tempo de 48 horas).
Assim como nos ensaios in vitro, observou-se que a infecção bacteriana
resultou em ativação da transcrição, no hospedeiro, de genes relacionados à
resposta inflamatória.
Após 4 horas de infecção, as células p~ritoniais de camundongos
selvagens infectados com Shigella apresentaram transcrição significativamente
maior dos genes das citocinas pró-inflamatórias TNF-u e IL-1, da enzima iNOS
e do receptor TLR4 que aqueles infectados com EIEC. EIEC, no entanto,
induziu a maior transcrição da citocina anti-inflamatória IL-10. A produção de
IFN-y foi maior na infecção pelas bactérias invasoras, em relação à E.coli K12;
a transcrição do gene da quimiocina IP-10 foi semelhante na infecção pelas

105
três bactérias. Após 48 horas de infecção, no entanto, o que se observa é que
apenas células de camundongos infectados com Shigella apresentaram
produção significativa das citocinas TNF-a, IL-1, IL-10, IFN-y, IP-10 e iNOS.
De maneira semelhante ao ocorrido in vitro, os ensaios realizados com
camundongos nocauteados foram prejudicados pelos elevados valores de
desvio padrão, especialmente no tempo de infecção de 48 horas. No tempo de
infecção de 4 horas, no entanto, observou-se que a infecção por EIEC induziu
um aumento na transcrição do gene de IL-1 significativamente maior que
Shigella; que apenas camundongos infectados com EIEC apresentaram
aumento significativo na transcrição de IL-18 e IFN-y; e que EIEC e Shigella
induziram de maneira semelhante a transcrição do gene de IL-10. Nos estudos
in vivo, portanto, a ausência da produção de NO interferiu no perfil
transcricional de camundongos infectados com EIEC, que produziram menos
IL-10, mais IL-18, IFN-y e IL-1, enquanto camundongos infectados com Shigella
produziram mais IL-10, e menos IL-18, IFN-y e IL-1.
A quimiocina IP-10 é produzida por macrófagos e regula o tráfego de
macrófagos, linfócitos e células NK para os sítios de inflamação (HAMERMAN
& ADEREM, 2001; PERERA e coL, 2001). A transcrição do gene da IP-10 foi
semelhante em macrófagos infectados com as três bactérias, sugerindo que
macrófagos secretam esta quimiocina em resposta a um estímulo bacteriano,
assim como documentado por HAMERMAN & ADEREM (2001), seja esta
bactéria invasora ou não. Não houve diferença na transcrição do gene da IP-10
entre camundongos selvagens e nocaute.

106
o ensaio de determinação da dose letal demonstrou que, enquanto o
inóculo de 10' bactérias não provocou a morte de nenhum animal selvagem
infectado com Shigella ou EIEG, em camundongos nocaute este inóculo foi
letal para 75% dos animais infectados com EIEG, e para 25% dos animais
inoculados com Shigella (tabela 4.1).
Foi demonstrado por WAY & GOLDBERG (1998) que o controle da
infecção por Shigella ocorre independente da produção de NO. Em seu estudo,
estes autores observaram que, embora tenha havido aumento de NO após
desafio intranasal com S.f1exneri, não houve diferenças na letalidade entre
animais selvagens e iNOS nocaute.
Estes resultados analisados em conjunto sugerem a hipótese de que as
diferenças na produção de citocinas entre camundongos selvagens e nocaute
estariam relacionadas à eficiência da resposta imune em controlar a infecção.
Macrófagos infectados com EIEG produzem pequenas quantidades de NO,
mas, ao que parece, mesmo esta pequena quantidade exerce efeito protetor,
uma vez que na ausência da produção de NO há um aumento da letalidade
desta bactéria. Na ausência de NO observou-se um aumento da produção de
citocinas pró-inflamatórias por esta bactéria, o que provavelmente estaria
relacionado à exacerbação da resposta inflamatória com conseqüente morte do
hospedeiro. A ausência de NO, no entanto, parece não influir na letalidade de
Shigella, e o aumento da produção de IL-10 por camundongos nocauteados
poderia estar relacionado a um efeito anti-inflamatório, protegendo o
hospedeiro.

107
A ativação de linfócitos é uma etapa importante da resposta inflamatória,
pois linfócitos ativados produzem citocinas, proliferam-se em clones específicos
e estimulam a ativação de macrófagos. Diversos autores demonstraram o
envolvimento dos linfócitos T na resposta do hospedeiro a Shigella (ISLAM e
coI. , 1995; AZIM e col., 1996;. ISLAM e col.; 2000; LE-BARILLEC e col.;2005).
No estudo da linfoproliferação foi utilizado o sobrenadante de cultura de
células Hep-2 incubadas com bactérias como possível antígeno indutor (fig.
4.33). Os resultados obtidos mostram que os antígenos destas duas bactérias
estimulam diferentemente a proliferação linfocitária (fig. 4.34). Houve maior
proliferação das células estimuladas com os antígenos de Shigella para todas
as concentrações testadas, enquanto que EIEC estimulou muito pouco a
linfoproliferação.
Em seguida, foi avaliada a capacídade de proliferação linfocitária após estímulo
com antígenos secretados por uma bactéria diferente da que o animal foi
imunizado (fig. 4.35). Observou-se que a imunização com Shigella foi a mais
eficiente em estimular o desenvolvimento de uma resposta T específica, uma
vez que a posterior estimulação com antígenos solúveis de Shigella resultou
em aumento significativo da proliferação. A imunização com EIEC foi a mais
ineficiente em estimular a geração de memória imunológica nos linfócitos T,
pois, embora o estímulo com antígenos de EIEC tenha resultado em
proliferação significativa, ela foi menor do que a proliferação induzida pelos
antígenos de Shigella e E.coli K12.
Além disso, a imunização resultou em reação cruzada a antígenos
compartilhados pelas três bactérias, já que linfócitos de animais imunizados por

108
uma bactéria também proliferaram significativamente em resposta ao estímulo
com antígenos solúveis de outras bactérias. Linfócitos de animais imunizados
com EIEC, surpreendentemente, proliferaram mais em resposta a antígenos
solúveis de Shigella e E.coli K12 do que em resposta a antígenos da própria
EIEC. Este dado sugere que, além da imunização com EIEC conferir uma
resposta T específica de menor magnitude, existem diferenças entre os
antígenos secretados por estas três bactérias.
A maior capacidade de Shigella em estimular a ativação dos linfócitos T
também pôde ser demonstrada pela maior produção de IFN-y e IL-10 por
linfócitos de animais imunizados com Shigella e posteriormente estimulados
com antígenos de Shigella. Não foi detectada produção de citocinas por
linfócitos de animais imunizados com EIEC e posteriormente estimulados com
antígenos de EIEC (fig. 4.36).
A produção de IFN-y e IL-10 em resposta a estímulo com Ipas por
células mononucleares de voluntários previamente infectados com Shigella
dysenteriae foi demonstrada elegantemente por SAMANDARI e col. (2000),
que sugerem a indução de uma resposta imune específica, adquirida e
predominantemente Th1 por esta bactéria.
Os resultados obtidos neste estudo trazem contribuições relevantes
acerca da caracterização da resposta inflamatória que a infecção por EIEC
desencadeia no hospedeiro. Foi demonstrado que o encontro de EIEC com
macrófagos murinos resulta em (i) fagocitose, (ii) escape bacteriano, e (iii)
alterações na viabilidade do macrófago. Este aumento de macrófagos não
viáveis pode estar relacionado à ocorrência de apoptose. Ocorre síntese de

109
citocinas pró-inflamatórias, e também da citocina anti-inflamatória IL-10. EIEC
induz a migração de granulócitos e monócitos para o peritônio, e as células
peritoniais respondem à infecção secretando citocinas pró-inflamatórias, e
também IL-10. A infecção por EIEC induz ativação dos macrófagos,
evidenciada pela produção de citocinas, pelo aumento na expressão de MHCII
e CD80, e pela produção de NO. O NO parece ser importante no controle da
bactéria, uma vez que camundongos nocauteados apresentam maior produção
de citocinas pró-inflamatórias e maior letalidade após administração de um
inóculo que não mata camundongos selvagens. Finalmente, a infecção por
EIEC parece ativar fracamente a resposta adaptativa, uma vez que a
proliferação de linfócitos em resposta aos antígenos solúveis de EIEC foi
pequena, e não foi detectada produção de citocinas por estes linfócitos. A
modulação negativa da expressão de CD86 pode contribuir para que não
ocorra ativação eficiente do linfócito T.
No que concerne às diferenças patogênicas entre EIEC e Shigella,
podemos destacar que o escape de EIEC do macrófago se dá mais
lentamente que o escape de Shigella. Além disso, na infecção por Shigella
parece haver um maior estímulo dos mecanismos imunes inatos, como
produção de NO. Neste ponto, a maior produção de TNF-a e IL-10
observadas nos ensaios ex vivo poderiam estar relacionadas com um uma
eliminação mais eficiente da bactéria pelo macrófago. Nos ensaios in vivo, foi
observada uma maior produção de citocinas pró-inflamatórias na infecção por
Shigella, enquanto EIEC estimulou uma maior produção de IL-10. O NO
parece ter um papel importante no controle da infecção por EIEC, embora a

110
eliminação de Shigella pareça ocorrer independente desta via. O maior
número de neutrófilos e macrófagos no peritônio e a maior produção de
citocinas, observados após 48 horas de infecção por Shigella, sugerem que o
hospedeiro controla mais rapidamente a infecção por EIEC. Shigella também
mostrou ser mais eficiente na geração da resposta adaptativa, uma vez que
os linfócitos de animais previamente imunizados com esta bactéria mostraram
proliferação e produção significativa de citocinas após estímulo com antígenos
de Shigella.
O tipo e a magnitude da resposta imune elicitada por um microrganismo
vivo depende de inúmeros fatores, como as moléculas de superfície e os
produtos por ele secretados. Nossos resultados demonstram que bactérias
que possuem LPS, mas não são invasoras, induzem uma menor produção de
citocinas do que aquelas que, além do LPS, secretam antígenos de invasão.
Estes antígenos, portanto, não estão apenas relacionados à invasão
bacteriana, mas também à ativação dos mecanismos inatos de defesa.
Finalmente, é importante ressaltar que Shigella e EIEC são patógenos
entéricos. Portanto, por mais que o modelo de infecção peritonial tenha se
mostrado valioso no estudo da relação de EIEC com as células de defesa do
hospedeiro, e que tenha permitido a observação de diferenças patogênicas
entre as duas bactérias, certamente alguns aspectos importantes deixaram de
ser abordados.

111
5 - Conclusões
• Macrófagos J774 e macrófagos peritoniais de camundongos C57BU6 são
ativados após infecção com EIEC; os macrófagos J774 fagocitam a bactéria e
secretam citocinas, e a infecção resulta em decréscimo da viabilidade dos
macrófagos. Os macrófagos de camundongos C57BU6 produzem NO e
aumentam a transcrição dos genes de citocinas pró-inflamatórias (TNF-a, IL-1,
IL-18) e anti-inflamatória (IL-10).
• A infecção por EIEC induz influxo celular e produção de citocinas pelas
células peritoniais. A infecção também gera uma resposta T específca.
• A ausência de NO resulta em aumento da letalidade de EIEC. Este dado
pode estar relacionado ao aumento de citocinas pró-inflamatórias observado
em camundongos nocaute, ou à necessidade de NO para o controle da
infecção por EIEC.
• In vitra, a infecção por Shigella resulta em maior número de macrófagos
não viáveis em menor tempo do que a infecção por EIEC. In vivo, há maior
produção de citocinas inflamatórias e, após 48 horas de infecção, a
produção de citocinas e o número de granulócitos são menores na infecção
por EIEC do que na infecção por Shigella. Além disso, a indução de uma
resposta T específica foi menos eficiente na infecção por EIEC. Estes
dados sugerem que o hospedeiro desenvolve uma resposta menos severa

112
em resposta à infecção por EIEC, e que esta bactéria é eliminada mais
rapidamente. Os resultados obtidos ajudam a explicar a necessidade de
um inoculo de 106 EIEC para causar infecção, em contraste com o inóculo
de Shigella, que é de 10 a 100 bactérias.
• Existem diferenças na infecção conduzida in vitro e in vivo no que
concerne à produção de citocinas, evidenciando a importância do
microambiente na fisiopatologia da infecção.
• A ativação da produção de citocinas é menor quando o hospedeiro é
desafiado com uma bactéria Gram-negativa, cujo único fator de virulência é
o LPS, em comparação a bactérias dotadas de plasmídeo de invasão. Os
antígenos de invasão, portanto, não apenas permitem a entrada de
Shigella e EIEC na célula epitelial, mas também interferem nos
mecanismos de defesa.

113
7. Referências Bibliográficas
ADEREM, A; UNDERHILL, D. M. Mechanisms of Phagocytosis inMacrophages. Annual Reviews in Immunology, 17:593-623,1999.
ADEREM, A How to Eat Something Bigger than Your Head. Cell, 110:5-8,2002.
ALBA SOTO, C. A; MIRKIN, G. A; SOLANA, M. E., GONZÁLEZ CAPPA, S.M. Trypanosoma cruzi Infection Modulates in vivo Expression of MajorHistocompatibility Complex Class 11 Molecules on Antigen-Presenting Cellsand T-Cell Stimulatory Activity of Dendritic Cells in a Strain-DependentManner. Infection and Immunity, 71 (3): 1194-1199, 2003.
ARONDEL, J.; SINGER, M.; MATSUKAWA, A; ZYCHLlNSKY, A;SANSONETII, P. Increased Interleukin-1 (IL-1) and Imbalance between IL-1and IL-1 Receptor Antagonist during Acute Inflammation in ExperimentalShigellosis. Infection and Immunity, 67:6056-6066, 1999.
AZIM, T.; SARKER, M. S.; HAMADANI, J.; KHANUM, N.; HALDER, R. C.;SALAM, M. A; ALBERT, M. J. Alterations in Lymphocite Phenotype andfunction in Children with Shigellosis Who Develop Complications. Clin. Diag.Lab. Immun, 3:191-196,1996.
BABU, U. S.; GAINES, D. W.; L1LLEHOJ, H.; RAYBOURNE, R. B.Diferencial Reactive Oxygen and Nitrogen Production and Clearance ofSalmonella Serovars by Chicken and Mouse Macrophages. Developmentand Comparative Immunology, 30:942-953,2006.
BOULOUGOURIS, G., MCLEOD, J. O., PATEL, Y.I., ELLWOOD, C.N.,WALKER, L. S. K., SANSOM, D. M. Positive and Negative Regulation ofHuman T Cell Activation Mediated by the CTLA-4/CD-28 ligand CD80. J.Immunol., 161 :3919-3924, 1998.
BRINKMANN, V., REICHARD, U., GOOSMANN, C., FAULER, B.,UHLEMANN, Y., WEISS, D. S., WEINRAUCH, Y., ZYCHLlNSKI, ANeutrophil Extracellular Traps Kill Bacteria. Science, 303: 1532-1535, 2002.
CHAKRAVORTIY, O., HANSEN-WESTER, 1., HENSEL, M. SalmonellaPathogenicity Island 2 Mediates Protection of Intracellular Salmonella fromReactive Nitrogen Intermediates. J. Exp. Med., 9: 1155-1166, 2002.
CONNELLY, L.; PALACIOS-CALLENDER, M.; AMEIXA, C.; MONCADA, S.;HOBBS, AJ. Biphasic regulation of NFKB activity underlies the pro- and antiinflammatory actions of nitric oxide. J. Immunol., 166:3873-3881, 2001.

114
COSSART, P.; SANSONETTI, P. J. Bacterial invasion: the paradigm ofenteroinvasive pathogens. Science, 304:242-248, 2004.
DEMUTH, A, GOEBEL, W., BEUSCHER, H. U., KUHN, M. DifferentialRegulation of Cytokine and Cytokine Receptor mRNA Expression uponInfection of Bone Marrow-Derived Macrophages with Listeriamonocytogenes. Infection and Immunity, 64: 3475-3483, 1996.
DE WAAL MALEFYT, R; ABRAMS, J.; BENNETT, B.; FIGDOR, C.G.; DEVRIES, J.E. Interleukin 10(IL-10) inhibits cytokine synthesis by humanmonocytes: an autoregulatory role of IL-1 Oproduced by monocytes. J ExpMed., 174(5):1209-1220, 1991.
D'HAUTEVILLE, H.; KHAN, S.; MASKELL, D. J.; KUSSAK, A; WEINTRAUB,A; MATHISON, J.; ULEVITCH, R J.; WUSCHER, N.; PARSOT, C.;SANSONETTI, P. J. Two msbB Genes Encoding Maximal Acylation of LipidA Are Required for Invasive Shigella f1exneri to Mediate InflammatoryRupture and Destruction of the Intestinal Epithelium. J. Immunol., 168:52405251,2002.
DING, A H.; NATHAN C. F.; STUEHR D. J. Release of reactive nitrogenintermediates and reactive oxygen intermediates from mouse peritonialmacrophages. Comparison of activating cytokines and evidence forindependent production. Joumal of Immunology, 141 :2407-2412, 1988.
DONNENBERG, M. S. Pathogenic strategies of enteric bacteria. Nature,406:768-774, 2000.
DUPONT, H. L.; FORMAL, S. B.; HORNICK, R B.; SNYDER, M. J.;L1BONATI, J. P.; SHEHAN, D. G.; LABREC, E. H.; KALAS, J. P.Phatogenesis of Escherichia coli diarrhea. N. Engl. J. Med., 285: 1-9, 1971.
EWING, W.H.; GRAVATTI, J.L. Shigella types encountered in themediterranean area. J. Bacteriol., 53:191-195, 1974.
FERNANDEZ, M.I.; THUIZAT, A; PEDRON, T.; NEUTRA, M.; PHALlPON,A; SANSONETTI, P.J. A newborn mouse model for the study of intestinalpathogenesis of shigellosis. Cellular Microbiology, 5(7):481-491,2003.
FERNANDEZ-PRADA, C.; TALL, B. T.; ELLlOTT, S. E.; HOOVER, D. L.;NATARO, J. P.; VENKATESAN, M. M. Hemolysin- positiveenteroaggregative and cell-detaching Escherichia coli strains cause oncosisof human monocyte-derived macrophages and apoptosis of murine J774cells. Infection and Immunity, 66: 3918-3924, 1998.
FREEMAN, G. J.; BORRIELLO, F.; HODES, R J. Murine B7-2, anAlternative CTLA-4 Counter-receptor that Costimulates T Cell Proliferationand Interleukin-2 Production. J. Exp. Med., 178:2185-2192, 1993.

115
FORMAL, S.8.; HALE, T.L.; SANSONETII, P.J. Invasive enteric pathogens.Rev. Infect. Ois., 5: 702-707, 1983.
GIBOTII, A; TANAKA, T.L.; OLlVEIRO, V.R; TADDEI C.R; MARTINEZ,M.B. - Molecular Characterization of Enteroinvasive Escherichia coli ipagenes by PCR- RFLP analysis. B.J Microbiol., 2004, no prelo
GORDON, S. The role of macrophage in immune regulation. Ressearch inImmunology, 149:685-688, 1998.
GORDON, S. Alternative activation of macrophages. Nature Reviews, 3:2335,2002.
GRASSL, G. A; FINLAY, B. B. Shigella rewrites host transcriptionalresponses. Nature Immunology, 8:15-16,2007.
GUICHON, A; HERSH, D.; SMITH, M. R; ZYCHLlNSKY, A Structurefunction analysis of the Shigella virulence factor Ipa B. J. bacteriol.,183:1269-1276,2001.
HAMERMAN, J. A; ADEREM, A. Functional transitions in macrophages duringin vivo infection with Mycobacterium bovis bacillus Calmette-Guérin. J.Immunol., 167(4):2227-33,2001.
HATHAWAY, L. J.; GRIFFIN, G. E.; SANSONETII, P. J.; EDGEWORTH, J. D.Human Monocytes kill Shigella flexneri but Then Die by Apoptosis AssociatedWith Supression of Proinflammatory Cytokine Production. Infection andImmunity, 70(7): 3833-3842, 2002.
HUME, D. A The mononuclear phagocyte system. Joumal of LeukocyteBiology, 72:621-627, 2002.
HUME, D. A; ROSS, I. L.; HIMES, S. R; SASMONO R T.; WELLS, C. A;RAVASI, T. The mononuclear phagocyte system revisited. Current Opinion inImmunology, 18:49-53, 2006.
ISLAM, D.; BARDHAN, P.K.; L1NDBERG, AA; CHRISTENSSON, B.Shigella Infection Induces Cellular Activation of T and B Cells and DistinctSpecies-Related Changes in Peripheral Blood Lymphocyte Subsets duringthe course ofthe Disease. Infection and Immunity, 63: 2941-2949,1995.
KOTLOFF, K.L.; WINICKOFF, J.P.; IVANOFF, 8.; CLEMENS, J.D.;SWERDLOW, D.L.; SANSONETTI, P.J.; ADAK, G.K.; LEVINE, M.M. Globalburden of Shigella infections: implications for vaccine development andimplementation of control strategies. Buli. World Hea/lh Organization 77:651-666, 1999.

116
LAN, R; LUMB, 8.; RYAN, D.; REEVES, P.R Molecular evolution of largevirulence plasmid in Shigella clones and enteroinvasive Escherichia coli.Infection and Immunity, 69: 6303-6309, 2001.
LE-BARILLEC, K.; MAGALHAES, J. G.; CORCUFF, E.; THUIZAT, A;SANSONETII, P.J.; PHALlPON, A; DISANTO, J.P. roles for T and NK cellsin the immune response to Shigella flexneri. J. immunol., 175:1735-1740.
LENSCHOW, D.J.; WALUNAS, TL.; BLUESTONE, J.A CD28/B7 system ofT cell costimulation. Annu. Rev. Immunol., 14:233:258, 1996.
MACMICKING, J.; XIE, O. Q.W.; NATHAN, C. Nitric oxide and macrophagefiunction. Annu. Rev. Immunol., 15:323-50, 1997.
MANDIC-MULEC, 1., WEISS, J., ZYCHLlNSKI, A Shigella flexneri isTrapped in Polymorphonuclear Leukocyte Vacuoles and Efficienty Killed.Infection and Immunity, 65: 110-115, 1997.
MARSHALL, D.; HASKARD, D. O. Clinicai Overview of Leukocyte Adhesionand Migration: Where Are We Now? Semanars in Imunnology, 14:133-140,2002.
MARTINO, M.C.; ROSSI, G.; MARTINI, 1.; TATIOLl, 1.; CHIAVOLlNI, D.;PHALlLPON, A; SANSONETII, P.J., BERNARDINI, M.L. Mucosallymphoidinfiltrate dominates colonic phatological changes in murine experimentalshigellosis. J. Inf. Dis., 192:136-148,2005.
MAURELLI, AT, BLACKMON, B., CURTIS 111, R Loss of Pigmentation inShigella flexneri 2a is Correlated with the Loss of Virulence-associatedPlasmid. Infect. Immun, 43: 397-401, 1984.
MEDZHITOV, R; JANEWAY, C. Innate immune: impact on the adaptativeimmune response. Current Opinion in Immunology, 9:4-9, 1997.
MEDZHITOV, R; JANEWAY, C. Innate Immune Recognition: Mechanismsand Pathways. Immunological Reviews, 173:89-97,2000.
MUKHOPADHYAY, S.; PEISER, L.; GORDON, S. Activation of MurineMacrophages by Neisseria meningitidis and IFN-y in vitro: Distinct Roles ofClass A Scavenger and Toll-like pattem recognition receptors in selectivemodulation of surface phenotype. Joumal of Leukocyte Biology, 76:577584,2004.
MUSSO, T; BADOLATO, R; RAVARINO, D.; STORNELLO, S.;PANZANELLI, P.; MERLlNO, C.; SAVOIA, D.; CAVALLO, R; PONZI, AN.;ZUCCA, M. Interaction of Barlonella henselae with the murine macrophage

117
cellline J774: infection and proinflammatory response. Infect Immun.,(10):5974-80,2001.
NASCIMENTO, F.RF; CALlCH, V.L.G.; RODRíGUEZ, D.; RUSSO, M. Dualrole for nitric oxide in paracoccidioidomicosis: essential for resistance, butoverproduction associated with susceptibility. J Immunol., 68(9):4593-600,2002.
NASCIMENTO, F. R F.; RODRíGUEZ, D.; GOMES, E.; FERNVIK E. C.;RUSSO, M. A Method for Multiple Sequential Analyses of MacrophageFunctions Using a Small Single Cell Sample. Brazilian Joumal of Medicai andBiological Research, 36: 1221-1226, 2003.
NAVARRE, W. W.; ZYCHLlNSKY, A Pathogen-induced apoptosis ofmacrophages: a Common end for Different Pathogenic strategies. CellularMicrobiology, 2(4):265-273,2000.
NEUTRA, M.R; FREY, A; KRAEHENBUHL, J.P. Epithelial M cells:gateways for mucosal infection and immunization. Cell, 86:345-348, 1996.
NONAKA, T; KUWABARA, T; MIMURO, H.; KUWAE, A.; IMAJOH-OHMI, S.Shigella-induced necrosis and apoptosis of U937 cells and J774macrophages. Microbiology, 149: 2513 - 2527,2003.
OVERBERGH, L.; GIULlETII, A.; VALCKX, D.; DECALLONNE, B.;BOUILLON, R; MATHIEU, C. The Use of Real-Time Reverse TranscriptasePCR for the Quantification of Cytokine Gene Expression. JoumalofBiomolecular Techniques,14:33-43, 2003.
PANES, J.; PERRY, M.; GRANGER, D. N. Leukocyte-Endothelial CellAdhesion: Avenues for Therapeutic Intervention. British Joumal ofPharmacology, 126:537-550, 1999.
PARSOT, C.; SANSONETII, P.J. - Invasion and pathogenesis of Shigellainfections. In: Miller, V.L - Bacterial Invasiveness. Curo Trop. Microbiol.Immunol. Springer, New York. 25-42, 1995.
PARSOT, C. - Shigella spp. and enteroinvasive Escherichia colipathogenicity factors. FEMS Microbiology Letters, 252: 11-18, 2005.
PAULNOCK, D.M. Macrophages A praticai approach. 1si edition., OxfordUniversity Press, 2000.PARSOT, C. Shigella spp. and enteroinvasive Escherichia coli pathogenicityfactors. FEMS Microbiology Letters, 252:11-18, 2005.
PERERA, P.; MAYADAS, T N.; TAKEUCHI, O.; AKIRA, S.; ZAKSZILBERMAN, M.; GOYERT, S. M.; VOGEL, S. N. CD11b/CD18 acts in

118
concert with CD14 and toll-like receptor (TLR) 4 to elicit fulllipopo/ysaccharide and taxol-inducible gene expression. The Joumal ofImmunology, 166:574-581,2001.
RAQIB, R.; L1NDBERG, AA; WRETLlND, 8.; BARDHAN, P.K.;ANDERSSON, U.; ANDERSSON, J. Persistence of local cytokine productionin shigellosis in acute and convalescent stages. Infection and Immunity,63:289-296, 1995.
RESTA-LENERT, S.; BARRET, K. E. Enteroinvasive bacteria alter barrier andtransport properties of human intestinal epithelium role of iNOS and COX-2Gastroenterology, 122:1070-1 087,2002.SAMANDARI, T.; KOTLOFF, K. L.; LOSONSKY, G. A; PICKING, W. O.;SANSONETII, P. J.; LEVINE, M. M.; SZTEIN, M. 8. Production of IFN-y and IL10 to Shigella Invasins by Mononuclear Cells from Volunteers Orally Inoculatedwith a Shiga-Toxin Deleted Shigel/a dysenteriae Type 1 Strain. The Joumal ofImmunology, 164:2221-2232,2000.
SANSONETII, P.J.; D'HAUTEVILLE, H.; FORMAL, S.B.; TOUCAS, M.Plasmid - mediated invasiness of "Shigel/a-like" Escherichia coli. Annu.Microbiol. (lnst. Pasteur). v.132, p.351-355, 1982a.
SANSONETII, P. J.; PHALlPON, A.; ARONDEL, J.; THIRUMALAI, K.;BANERJEE, S.; AKIRA, S.; TAKEDA, K.; ZYCHLlNSKI, A Caspase-1activation of IL-113 and 11-18 are essential for Shigella f1exneri- inducedinflammation. Immunity, 12:581-590,2000.
SANSONETII, P. J. Rupture, invasion and inflammatory destruction of theintestinal barrier by Shigella, making sense of prokaryote-eucaryote cross-talk.FEMS Microbiology reviews, 25:3-14,2001.
SANSONETII, P. J. War and peace at mucosal surfaces. Nat. Rev. Immunol.,4:953-964, 2004.
SANSONETII, P. J. Shigellosis: An Old Disease in New Clothes? PIoSMedicine, 3:51465-1466,2006.
SANSONETII, P. J.; DI SANTO, J. P. Debugging how bacteria manipulatethe immune response. Imunity, 26:149-161,2007.
SERAFIN-LOPEZ, J.; CHACON-SALlNAS, R.; MUNOZ-CRUZ, S.; ENCISOMORENO, J.A; ESTRADA-PARRA, S.A; ESTRADA-GARCIA, I. The effect ofiron on the expression of cytokines in macrophages infected withMycobacterium tuberculosis. Scand J Immunol. (4):329-37,2004.
SERENY, B. Biochemical reactions and virulence of E. coli 0124, K27 (B17).Acta Microbiol. Acad. Sei. Hung., 10: 11-18, 1963.

119
SHILOH, M. U.; NATHAN, C. F. Reactive Nitrogen Intermediates and thePathogenesis of Salmonella and mycobacteria. Current Opinion inMicrobiology, 3:35-42, 2000.
TAKEOA, K., KAISHO, T., AKIRA, S. TolI-like receptors. Annu. Rev.Immunol., 21 :335-376,2003.
TRABULSI, L.R.; ZULlANI, M.E.; SERRANO, J.A On two newenterobacteria pathogenic to the guinea-pig eye (cultures 185T-64 and 193T64). Rev Inst Med Trop Sao Paulo. (4):241-6, 1965.
UNOERHILL, O. M.; BASSETII, M.; RUOENSKY, A; ADEREM, A Oynamicinteractions of macrophages with T cells during antigen presentation. JoumalofExperimental Medicine, 190(12): 1909-1914, 1999.
VON SEIOLEIN, L., KIM, O. R, ALI, M., LEE, H., WANG, X. Y. A MulticentreStudy of Shigella Oiarrhoea in Six Asian Countries: Oisease Burden, ClinicaiManifestations, and Microbiology. PIoS Med 3:e353.DOI: 10.1371/journal.pmed.0030353, 2006.
WAY, S.S.; BOREZUK, AC.; OOMINITZ, R; GOLOBERG, M.B. An essentialrole for y -interferon in innate resistance to Shigella f1exneri infection. Infect.Immun., 66: 1342-1348, 1998.
WAY, S.S.; GOLOBERG, M.B. Clearance of Shigella f1exneri infection occursthrough a nitric oxide-independent mechanism. Infection and Imunity,66(6):3012-3016, 1998.
WEINRAUCH, Y., ORJAN, O., SHAPIRO, S.O., WEISS, J., ZYCHLlNSKI, ANeutrophil elastase targets virulence factors of enterobacteria. Nature,417:91-94,2002.
YANG, K.K; OORNER, B. G.; MERKEL, U.; RYFFEL, B.; SCHÜTI, C.;GOLENBOCK, O.; FREEMAN, M. W.; JACK, R S. Neutrophillnflux inResponse to a Peritonial Infection with Salmonella Is Oelayed in LPS-BindingProtein or CO-14 Oeficient Mice. The Joumal of Immunology, 169:4475-4480,2002.
YANG, Y., TUNG, J. W., GHOSN, E. E. B., HERZENBERG, L. A,HERZENBERG, L. A Oivision and differentiation of Natural Antibody Producingcells in mouse spleen. PNAS, 104:4542-4546,2007.
ZYCHLlNSKI, A; PREVOST, M. C.; SANSONETII, P. J. Shigella f1exneriinduces apoptosis in infected macrophages. Nature, 358: 167-168, 1992.
ZYCHLlNSKI, A; THIRUMALAI, K.; ARONOEL, J.; CANTEY, J. R;ALlPRANTIS, A O., M.; SANSONETTI, P. J. In vivo apoptosis in Shigellaflexneri infections. Infection and Imunity, 64:5357-5365, 1996.

UNIVERSIDADE DE SÃO PAULOFaculdade de Ciências Farmacêuticas
Comissão de Ética em Experimentação Animal
orCEEA n009
CERTIFICADO
Certificamos que o Projeto "Estudo da resposta celular induzida por
Escherichia coli Enteroinvasora (EIEC) no Processo Inflamatório"
(Protocolo nO}O), sob a responsabilidade do(a) Sr(a). Juliana Mota
Khalil Amhaz e do orientador(a) Marina Baquerizo Martinez, está de
acordo com os Princípios Éticos na Experimentação Animal adotado
pelo Colégio Brasileiro de Experimentação Animal (COBEA) e foi
aprovado pela Comissão de Ética em Experimentação Animal
(CEEA) desta Faculdade, em 02/06/2003.
São Paulo, 03 de junho de 2003.
i.. J -,' :} -." i,"')' '/ /'• .' - ., I .. ' ,,-.,,-~; .............. \. '..'. 'J .'_ \ v' '-- \. \. "_
Pro! Assoe. Elfriede Marianne BacchiCoordenadora da CEEA
Av. Prof. L1neu Prestes, 580 - Bloco 13 A - Cidade Universitária - CEP 05508-900 - São Paulo - SPFone: (11) 309136n I Fax: (11) 3031-8986 - e-mail: [email protected]





























