Embed Size (px)
Citation preview

P.S.Z.N. I: Marine Ecology, 16 (2): 133-144 (1995) 0 1995 Blackwell Wissenschafts-Verlag, Berlin ISSN 0173-9565
Accepted: June 29, 1994
Temperature Effects on Photosynthesis and Depth Distribution of the Seagrass Cymodocea nodosa (UCRIA) ASCHERSON in a Mediterranean Coastal Lagoon: the Mar Menor (SE Spain) J. TERRADOS’* & J. D. ROS’
‘Depto. de Ecologia y Medio Ambiente, Universidad de Murcia, Campus Univ. de Espinardo, E-30100 Murcia, Spain.
’Dept. d’Ecologia, Universitat de Barcelona, Diagonal, 645, E-08028 Barcelona, Spain.
With 5 figures and 2 tables
Key words: Seagrasses, Cymodocea, net photosynthesis, dark respiration. temperature, depth distribution.
Abstract. Seasonal variation of temperature effects on photosynthesis and respiration of Cymodorea nodosa (UCRIA) ASCHERSON was investigated in a 1 m depth meadow in the Mar Menor lagoon (SE Spain). Light-saturated net photosynthesis (P,J and dark respiration (R) rates were linearly correlated with water temperature between 10 and 30°C. and 10 and 3 5 T , respectively, during the year. Water temperature had a similar effect on R rates in August, November, and February, but increased P,,, rates in August vs. those in February, thus promoting a shift of the maximum P,,,/R ratio values from 15-C in February to 30‘C in August. P-I curves had higher P,,, values in May and August than in November or February, while I, values remained relatively constant throughout the year. Whole-plant carbon balances suggested that C. nodosa might not be able to survive in the central basin of the lagoon and that the spatial distribution of this seagrass in the Mar Menor might be controlled by light availability.
Problem
Water temperature is a major factor controlling seagrass photosynthesis ( BUL- THUIS, 1987; HILLMAN et al., 1990). In general, the photosynthesis-irradiance (P-I) relationship shows an initial light-dependent portion and a subsequent tem- perature-dependent portion (RAMUS, 198 1). Within normal physiological limits, the initial slope, c1, of the P-I curve is not affected by temperature, but, as the upper temperature limit is approached, the photosynthetic capacity is reduced (BULTHUIS,
*Present address and for correspondence: Centro de Estudios Avanzados de Blanes, C.S.I.C., Cami de Santa Birbara, sjn, E-17300 Blanes, Gerona, Spain.
U. S. Copyright Clearance Center Code Statement: 01 73-9565/95/1602-0133$11 .oo/o

134 TERRADOS & Ros
1987). Light-saturated net photosynthesis, P,,,,, and dark respiration, R, rates increase with temperature, whereas u usually remains constant. Thus, com- pensation irradiance, I,, and saturation irradiance, Ik, must also increase with temperature, and their changes during the year mainly reflect the seasonal changes of P,,, and R (BULTHUIS, 1987; HILLMAN et al., 1990). Plant carbon balances and, therefore, growth and temporal distribution of seagrasses may be driven by the changing influence of water temperature on photosynthesis and respiration during the year (MARSH et al., 1986; ZIMMERMAN et al., 1989; PEREZ & ROMERO, 1992). The ability of the plants to develop thermal acclimation may also play a role (EVANS et al., 1986).
Meadows of the chlorophyte Caulerpa prolifera (FORSSKAL) LAMOUROUX and the seagrass Cymodocea nodosa (UCRIA) ASCHERSON cover more than 85 YO of the bottom of the Mar Menor lagoon: C. nodosa is restricted to depths less than 4 m, while C. prolifera is distributed throughout the lagoon (TERRADOS, 1991). C. nodosa shows a clear unimodal growth cycle, with maximum vegetative development reached during the summer (TERRADOS & Ros, 1992a). Water temperature changes from 12°C in January-February to 30°C in July-August and might be a major factor controlling the seasonality of C. nodosa growth. Together with light avail- ability, the responses of C. nodosa photosynthesis and respiration to seasonal water temperature variation might also help to explain the spatial distribution of this seagrass in the lagoon.
Material and Methods
1. Collection site and plant material
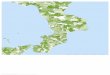
The Mar Menor is a 135 km' coastal lagoon located on the SE Mediterranean coast of Spain (Fig. I ) . Its average depth is 4 m, with a maximum of 7 m at some places. Water salinity varies between 42% and 47%. depending on area and season. Water temperature at the sampling site ranges from 10 to 12°C in January-February to 30 to 33°C in July-August, with a daily variation of 5°C (TERRADOS, 1991). Maximum photon flux densities, PFD (PAR), just above macrophyte meadows vary between 900 and 1700 p n o l * m - * . s - ' at the collection site, and between 160 and 600 p m o l . m - 2 * s - ' in the central lagoon basin. Nitrate and orthophosphate water column concentrations are below 1 pM and 0.3 pM, respectively (TERRADOS, 1991). Additional information regarding environmental characteristics and biotic communities may be found in Ros (1987).
Cyrnodocea nodoso shoots were collected at 1 m depth in November 1989 and February, May and August 1990 at an on-sand meadow in the Encafiizada de la Torre (Fig. I ) . Plant material was transferred to the laboratory within 4 h and maintained there in aerated seawater, indirect sunlight (< 150 pmol. m - ' * s -I), and at the same temperature and light-dark photoperiod as in situ conditions. Experiments were performed within 4 days of collection.
2. Experiments
The influence of water temperature on dark respiration rates (R) was determined by incubations (six replicates) at 10, 15, 20. 25. 30, and 35'C in November, February, May, and August. The influence of water temperature on light-saturated net photosynthesis rates (PmaX) was established by incubations (three replicates) a t the above temperatures in February and August (saturating irradiance, 665 p n o l . m * s - I ) . To determine the seasonal variation of the influence of water temperature on photo- synthetic charactenstics (1'. l k , P,,,,, a), photosynthesis-irradiance (P-I) curves (six replicates) were

Temperature effects on C . nodosa photosynthesis 135
Fig. I , Location of the Mar Menor lagoon on the Spanish Mediterranean coast, and of the collection site at the Encailizada de la Torre (solid square). Solid circle indicates the central basin site at which daily light cycles were measured; the 4 m depth isobath and the spatial distribution of C. nodosa meadows (inverted-T symbols) in the lagoon are also represented.
obtained between 0 and 665 p n 0 1 . m ~ ~ s - ' at 10, 20, and 30°C in November, February, May, and August.
On the evening prior to the experiments, whole shoots of Cymodocea nodosa were cut from the rhizome at the level of the second internode of the respective orthotropic rhizome; the senescent leaves (yellow or brown in colour and/or broken) were discarded and the rest were gently cleaned ofepiphytes. The shoots were then held overnight at the experimental temperature in order to obtain stable photo- synthesis and respiration (LITTLER. 1979; KNWP & BATE, 1988). The mean shoot weight/bottle volume ratio of the entire set of incubations done was 0.1 107 g dw*l - ' (n = 495, S.E. = 0.0044). Although these values are higher than recommended by LITTLER (1979). there was no negative correlation between photosynthetic rates and ratio values, which would be indicative of self-shading.
Incubation chambers were 280 ml glass bottles submerged in an aquarium. Incubation temperature was controlled with submersible standard laboratory heaters and coolers. Light was provided laterally from one side by two 500 W halogen lamps. Stirring inside the chambers was effected by placing a multisite magnetic stirrer just below the aquarium. Photon flux density (PAR) at the incubation chambers was regulated by changing the distance between the aquarium and the lamps, and by inserting

136 TERRADOS & Ros
neutral density screens between them. Dark respiration rates were measured in the same system, but covering the bottles with aluminium sheets.
Net photosynthesis and dark respiration rates were estimated by measuring the variations of dissolved oxygen concentration inside the incubation chambers for 2-h periods. Previous experiments showed that this was the optimum incubation time (TERRADOS, unpublished data). Oxygen concentration was measured using a Clark-type oxygen electrode (Orbisphere, 27141). Photon flux density (PAR) was measured using a cosine-corrected underwater quantum sensor (LiCor, LI-192SA). Cymodorea notfosa has a lacunar system (BORNET, 1864). SORRELL & DROMCi001.E ( I 986) contended that an oxygen-storage error is always present when measuring photosynthesis of aquatic macrophytes with a lacunar system. but ROBERTS & MORIARTY (1987) showed that there was a linear relationship between the oxygen discharged from the lacunae and the oxygen released into the water column. Stirring of the water inside the chamber (ROBERTS & MORIARTY, 1987), 2-h incubation periods (KEMP P I uI., 1990) and high volumes of the chamber (280 ml) relative to the lacunar airspace (SORRELL & DROMGOOLE, 1986) should minimize this error. After incubation, plant material was rinsed with distilled water and its dry weight determined after drying at 105 'C for 24 h.
3. Calculations
The influence of temperature on P,,,,, and R was described by linear least squares regression analysis ol' the results using different models (linear, potential, exponential). The best fit, i e . , that with the highest Pearson correlation coefficient. was chosen. Critical values for significance levels of the correlation coefficients of ROHLF & SOKAL (1969) were used. Significance of differences between the obtained regression lines (slope, Y-intercept) was tested at the 0.05 level (MARTIN-ANDRES & LUNA DEL CASTILLO, 1989).
P-I curve results were fitted to the models
p "el = a~&z '~~p, , ,8 .e~l -R ( 1 )
PDc, = [f',,,AI- L)l/KPnVdx/a) + (1- L)1 (2)
(modified from NELSOU & SIEGKIST, 1987, and from PEREZ & ROMERO, 1992, respectively) using the non-linear fitting program Enzfitter (LEATHERBARROW, 1982). Best fit was chosen using the fidelity index of JASSBY & PLATT (1976). CHALKER (1980) proposed an hyperbolic tangent model to describe light saturation curves of photosynthesis and, in a comparative study, JASSBY & PI.ATT (1976) showed that model to be the better descriptor of phytoplanktonic photosynthesis. However, this model might not work as well for macrophytes, as NELSON & SIEGRIST (1987) have shown. The CHALKER (1980) model gave no better fits (fidelity index, JASSBY & PLATT, 1976) than the models used here. FOURQURFAN & ZIBMAN ( I 991 ) found no major differences among rectangular hyperbola, exponential, and hyperbolic tangent models in the description of Thalussiu irsrudinum P-I curves.
Diurnal light curves at shallow and deep lagoon sites were constructed for selected days during 1990 using averaged PFD data obtained every 15 min from dawn to dusk with a cosine-corrected underwater quantum sensor (LiCor, Ll-lY2SA) connected to a datalogger Li-1000. The leaf carbon balance for those days was calculated by multiplying in s i /u diurnal PFD curves by corresponding points on the P- I curve at 15-min intervals. Oxygen measurements were converted to carbon values by multiplying by 0.3, assuming a photosynthetic quotient of 1.25 (MCROY & MCMILLAN, 1977; DBNNISON, 1987).
Root-rhizome respiration rates are also required to calculate whole-plant carbon balances. The root- rhizome/leaf biomass ratio and seasonal growth pattern of Cymodocea nodosa at the shallow site are thoroughly known (TERRADOS & Ros, 1992a). Assuming that absolute respiration rates are positively correlated with specific growth rates (FALKOWSKY P I a/.. 1985). root and rhizome respiration rates were estimated as:
leaf resoiration rate x annual rhizome P/B ratio annual leaf P/B ratio
with annual P/B ratios being the quotient between annual production and mean year biomass of leaves and rhizomes, respectively. Whole-plant carbon balances have been calculated by multiplying the root-

Temperature effects on C. nodosa photosynthesis 137
rhizomeileaf biomass ratio by estimated root-rhizome respiration rates, and adding the result to leaf carbon balances.
4. Starch content of tissues
The starch content of Cymodocea nodosa leaves, rhizomes, and roots at the shallow site was determined monthly from October 1987 to November 1988. Samples of leaves, rhizomes, and roots were taken at four different points of the meadow patch, pooled together, and frozen in sifu with dry ice. Prior to analysis, plant material was dried at 105°C for 24 h and ground to fine powder. Ground material (0.150 g) was washed repeatedly ( 4 6 times) with 80% ethanol to remove all trace of soluble sugars and extracted with 30% perchloric acid (4°C) to solubilize starch (JENSEN, 1962). The starch concentration in the extract was measured by a spectrophotometric anthrone assay (JENSEN. 1962).
Results
The dark respiration rate was linearly correlated (P < 0.01) with water temperature between 10 and 35°C (Fig. 2a) in November, February, and August; in May, there was no relationship between water temperature and R. The effect of temperature on R was similar (P > 0.05) in November (regression coefficient, b = 0.0600), February (b = 0.0745), and August (b = 0.0617), although dark respiration values were greater (P < 0.05) in February and August than in November.
P,,, was linearly correlated (P < 0.05) with water temperature between 10 and 30°C in February and August (Fig. 2b); at 35°C the photosynthetic rate decreased. Rates were higher ( f e x p = 3.6166, P < 0.01) in August than in February, and the influence of temperature on P,,, was greater ( f e x p = 10.6790, P < 0.01) in August (b = 0.4224) than in February (b = 0.0677). The P,,,/R ratio showed a shift of its maximum from 15°C in February to 30°C in August (Fig. 2c).
P-I model 1, based on a two-parameter-exponential-type kinetics, gave better fits to the results obtained in February at 20"C, May at 10, 20, and 30°C and August at 10°C. P-I model 2, based on a MICHAELIS-MENTEN-type kinetics, gave better fits to the rest of the results (Table 1). I, values ranged between 7 and 116 pmol-m-2-s- ' , while Ik values ranged between 22 and 401 pmol-m-'*s- ' (Table 2). The P,,, rate ranged from 0.87 to 14.01 mg 0 2 * g (dw)-'.h-', and R ranged
Table 1. Fidelity index values (JASSBY & PLATT, 1976) of fits to exponential ( I ) and MICHAELIS-MENTEN type (2) P-1 models (see text)
temperature November February May August
10°C model 1 0.053 0.192 0.523 0.029 model 2 0.029 0.055 0.542 0.056
model 1 0.070 0.094 0.385 0.419 model 2 0.002 0.124 0.656 0.262
model 1 0.097 0.091 0.303 0.606 model 2 0.062 0.020 0.641 0.340
20°C
30°C

138 TERRADOS & Ros
3.0
2.5
2.0 f ';r
@ ON
tk U Q
0 1.0
0.5
0.0
8
6
Tr
g a E 0" 4 h
2
0
6
5
4
3
2
1
0
E
d
8 E
. h
- - - I 1 I I
B
l J.7-0-u
C
I I I I I I I
0 5 10 15 20 25 30 35 "c Fig. 2. a: Effect of temperature on dark respiration rates of Cymodorea nodosa in November (triangles, dash-dot line). February (circles, dashed line), May (diamonds), and August (squares, solid line). b: Effect of temperature on Pmax rates in February (circles) and August (squares). c: P,,./R ratio as a function of temperature between 10 and 35°C in February (circles) and August (squares). Vertical bars represent the standard error of the mean.
from 0.29 to 2.24 mg O,.g (dw)-' - h- ' . Photosynthetic efficiency at non-saturating irrddiances (a) ranged between 0.005 and 0.063 mg 0 2 . g (dw)-' * h- ' (Table 2). The influence of temperature on P-I curves was small in November and February; in May and August, however, the P-I curves showed higher P,,, values at 20 and 30°C than at 10°C. The highest P,,, value was obtained in August at 30°C (Fig. 3).
The starch content of Cymodocea nodosa leaves varied between 14.3 and 30.7 mg-g (dw)-' and showed no seasonal trend (Fig. 4). When compared with leaves,

Temperature effects on C. nodosa photosynthesis 139
the starch content of rhizomes was lower from December to March (1 1.8-14.1 mg-g (dw)-') and higher from May to August (30.149.9 mg-g (dw)-'). From March to June, the starch content of roots (9.8-15.5 mg-g (dw)-') was lower than that of leaves.
The constructed carbon balances suggest a net gain of carbon for the whole plant at the shallow site from March to September (Fig. 5); in October and January, photosynthesis did not supply enough carbon to maintain the plant. Except for June (13.7 mg C - g (dw)-' -day-'), the whole-plant carbon balance was always negative in the central basin.
Discussion
The estimates of Cymodocea nodosa photosynthetic parameters obtained in this study are similar to those found by DREW (1978) in Malta and PEREZ & ROMERO (1992) in the Ebro delta (NE Spain). The light compensation and saturation irradiances of C. nodosa in the Mar Menor are also within the range of values known for other seagrasses (EVANS et al., 1986; LIBES, 1986; MARSH et al., 1986; PIRC, 1986; DENNISON, 1987; HILLMAN et a/., 1990).
The similarity both between the dark respiration values in February and August and the effect of temperature on them (Fig. 2) suggests that water temperature has a constant influence on Cymodocea nodosa respiration throughout the year. Light- saturated net photosynthesis rates, however, were higher in August than in Febru- ary, and water temperature had a larger influence on them (Fig. 2). These results do not conform to those obtained by PEREZ & ROMERO (1992), who found C.
Table 2. P-I relationship parameters of Cymodocea nodosa at 10, 20, and 30°C during the year. Values in brackets represent the standard error of the estimated parameter. Units: a, in mg O , * g (dw)-' * h - ' -pmol-' - m 2 * s ' ; P,,, and R, in mg O,*g (dw)-' * h-'; I, and It . in p m o l * m - 2 * s - ' .
November February May
10'C a 0.039 0.0 19 0.009 (0.003) Pmax 0.87 (0.10) 1.32 (0.32) 2.40 (0.53) 1, 7.3 (6.6) 28.2 ( 1 I .O) 0.07 II, 22.3 (21.1) 67.9 (36.4) 25.9 R 0.29 0.55 0.01 (0.02)
U 0.025 0.020 (0.003) 0.025 (0.005) 20°C
P m a I .94 (0.04) 2.29 (0.26) 5.79 (0.65) I, 20.0 (0.9) 32.1 22.9 Ik 77.8 (4.5) 111.9 23 1.2 R 0.50 0.66 (0.14) 0.57 (0.35)
a 0.005 0.014 0.05 I (0.009) P m d x 2.20 (1.02) I .89 (0.26) 7.97 (0.69) 1, 1 16.4 (22.1) 63.6 ( I I .6) 43.3 1, 400.9 (196.5) 135.1 (35.2) 154.5 R 0.64 0.89 2.24 (0.52)
30°C
August
0.010 (0.002) I .68 (0.22)
50.6 165.9
0.51 (0.18)
0.046 7.59 ( I .94)
18.1 (8.2) 166.5 (86.7)
0.82
0.063 14.02 (2.06) 26.0 (3.6)
223.1 (68.2) 1.63

140 TEKRADOS & Ros
,,o , I , , . . . . . 0 ........... ....
~
0 100 200 300 400 500 600 0 100 200 300 400 500 600
photon flux density , prnol rn-2 s-' Fig. 3. P-I relationship ofC)modncea nodosci at different temperatures (IO"C, circles, dotted line; 20'-C, triangles, dashed line; 30"C, squares, solid line) in November, February, May, and August. Vertical bars represent the standard error of the mean.
nodosa respiration was more sensitive to temperature than photosynthesis. The shift of the maximum value of the P,,,/R ratio from 15°C in February to 30°C in August (Fig. 2) suggests that photosynthetic production is higher in August than in February. The high dark respiration values found in May and their lack in correlation with water temperature were anomalous and unexpected; they could be the result of an as yet undetermined stress caused to the plants during collection and handling.
The increased P,,, rate with increasing water temperature from February to August implies that Ik values must also increase, as observed (Table 2). On the other hand, I, values remained relatively constant during the year, which is con- sistent with the small variation in the observed dark respiration rates (Fig. 2, Table 2).
Cymodocea nodosa shows a clear unimodal growth cycle in the Mar Menor (TERRADOS & Ros, 1992a): the highest leaf and rhizome growth rates occur from May to July, leaf growth is minimal from November to February and almost stops in December to January, and rhizome growth is halted from October to February. P-I curves obtained in this study (Fig. 3) and the daily carbon balances calculated for whole plants at shallow sites (Fig. 5) are consistent with this temporal pattern of vegetative development.

Temperature effects on C
60
tiodosa photosynthesis
1 141
-
N D J F M A M J J A S O N 1988
Fig. 4. Seasonal variation of starch content of Cymodocea nodosa leaves (circles, dash-dot-dot line). rhizomes (squares, solid line), and roots (triangles, dashed line) at the Encafiizada de la Torre (shallow) site.
The starch content of Cyrnodocea nodosa rhizomes was higher from May to August than during the rest of the year. These data suggest that photosynthesis during this period might supply enough carbon to meet growth needs, and that the excess carbon is stored in the rhizome. Starch reserves would then be consumed in autumn to maintain leaf growth. In winter, net photosynthesis, leaf growth rates,
50
'z 40 e.
30 0
20
U r
a,
ISI
ti 5 '0 2
ii B 0
-10
-20 U
plant, shallow
I I I I 4 I I I I I
J F M A M J J A S O N D J Fig. 5 . Daily carbon balances constructed for leaves (circles) and whole-plant (squares) a t the Encari- izada de la Torre, shallow (solid line) and central basin, deeper (dashed line) sites.

142 TEKKADOS & Ros
and starch content of rhizomes were the lowest. The high rhizome starch content in December 1987 (Fig. 4) could be the result of interannual variability in the timing between leaf and rhizome growth seasons. PIRC (1989) detected no starch storage in C. nociosu rhizomes at Ischia, Naples. However, this author found that the content of soluble carbohydrates, mainly glucose, fructose, and 1 -chiro-inositol in all plant parts, was correlated with photosynthetic activity, and suggested that sucrose maintained plant metabolism from February to May.
P-I relationships and diurnal light curves may be used to predict growth responses to changes in light regime, seasonal growth patterns. and maximum depth limit for seagrass survival (DENNISON & ALBERTE, 1985; DENNISON. 1987). Several studies have shown the major role of light on seagrass survival and growth (BACKMAN & BARILOTTI, 1976; DENNISON & ALBERTE, 1982, 1985; GIESEN ('1 ul., 1990). Macrophytic vegetation in the deeper central basin of the Mar Menor is composed of a monospecific Cuulrrpu proliferu meadow: only scattered, isolated Cyniodoceu notioosu plants are present, always above 4 m depth (TERRADOS, 1991). True C. nodosu meadows are restricted to shallow sites. Because there are no plants at the deeper site, the whole-plant carbon balances of C. nociosu there were calculated by extrapolation of the results obtained from shallow plants. This might flaw the light-limitation hypothesis presented here if plants at the deeper site were able to acclimatize to lower light levels, but there are no plants there to test that. DENNISON & ALBERTE (1982, 1985) have shown that in situ manipulations of light availability along a depth gradient had a larger effect on Zostrru mariner growth than photosynthesis, and that photosynthetic acclimatization could not cope with net reductions in light availability.
Although the respiration rates of roots and rhizomes were not measured in this study, their estimated value and the model outputs for whole-plant carbon balances are consistent with growth seasonality and spatial distribution of C. nodosci in the Mar Menor lagoon. Whole-plant carbon balances indicate that only in June, when light availability is at its annual maximum, was there enough light for C. nodom to survive in the central basin. Short days and a low incident light angle in winter, along with increased attenuation of light in the water column in summer, may explain this situation. Carbon balances calculated for Caulerpa prolijkru in a similar way indicated that, even at the deeper central basin, this alga maintained a photosynthetic surplus throughout the year; it is therefore unlikely that its spatial distribution in the lagoon is limited by light availability (TERRADOS & Ros, 1992b).
The mean vertical attenuation coefficient for downwelling irradiance (K,) in the Mar Menor is 0.35, but K D may reach up to 0.57 during August and September due to phytoplankton growth ( J . GILABERT, pers. comm.). DUARTE (1991) suggests a mean empirical relationship for seagrasses of Z, = 1.86/KD, where Z, (light compensation depth) is equivalent to the depth at which light intensity is about 1 1 % of surface irradiance. Assuming the validity of this last equation and knowing the seasonal variation of KD in the Mar Menor, Z, values for Cymodoceu nodosu would oscillate between 3.3 and 5.5 m in August and September, depths which roughly delimitate C. riodosa spatial distribution in the lagoon (TERRADOS, 199 1).
These findings are consistent with the hypothesis that Cymodoceu nodosu dis- tribution in the Mar Menor lagoon is controlled by light availability. Although c. nodosu P,,, rates increased with temperature more than R rates, the whole-plant carbon balance was negative in summer because of reduced light availability at

Temperature effects on C. nodosu photosynthesis 143
deeper lagoon sites. Similar relationships have been found in lower Chesapeake Bay, Virginia (WETZEL & PENHALE, 1983; EVANS et al., 1986), where light avail- ability and light-temperature interactions are the main environmental controls of seasonal growth dynamics in Zostera marina and Ruppia maritima.
Summary
Both leaf respiration and P,,, rates of Cymodocea nodosa in the Mar Menor increased with increasing temperature between 1 O T , 35 "C and 30"C, respectively, but there were differences in the response of the two processes. Temperature had a similar influence on leaf respiration throughout the year; in contrast, the effect of temperature on P,,, rates was greater in August than in February, shifting the maximum P,,,/R ratio values from 15°C in February to 30°C in August. The main effect of temperature on the P-I relationship was an increase in the P,,, rates in May and August vs. those in November and February. Light availability is a major factor controlling Cymodocea nodosa spatial distribution in the Mar Menor lagoon.
Acknowledgements
This research was supported by a MEC-PFPI fellowship to J . TERRADOS. Reviewer comments greatly improved an earlier version of the manuscript.
References
BACKMAN, T. W. & D. C. BARILOTTI. 1986: Irradiance reduction: effects on standing crops of the
BORNET. E., 1864: Recherches sur le Phucugrostis major CAVOL. Ann. Sci. Nat., 5, Ser. Bot., 1: 5--51. BULTHUIS, D. A., 1987: Effects of temperature on photosynthesis and growth of seagrasses. Aquat.
Bot., 27: 2 7 4 0 . CHALKER, B. E., 1980: Modelling Light Saturation Curves for Photosynthesis: An Exponential Func-
tion. J . theor. Biol., 84: 205-215. DENNISON, W. C., 1987: Effects of light on seagrass photosynthesis, growth and depth distribution.
Aquat. Bot., 27: 15-26. - - & R. S. ALBERTE, 1982: Photosynthetic responses of Zosrera marina L. (Eelgrass) to in situ manipu-
lations of light intensity. Oecologia, 5 5 137-144. - - & R. S. ALBERTE, 1985: Role of the daily light period in the depth distribution of Zosreru murinu
(eelgrass). Mar. Ecol. Prog. Ser., 2 5 51-61. DREW, E. A,, 1978: Factors affecting photosynthesis and its seasonal variation in the seagrasses
Cymodocea nodosa (UCRIA) ASCHERSON and Posidonia oreanica (L.) DELlLE in the Mediterranean. J. Exp. Mar. Biol. Ecol., 31: 173-194.
eelgrass Zosreru marinu in a coastal lagoon. Mar. Biol., 34: 3 3 4 0 .
D U A R ~ , C. M., 1991: Seagrass depth limits. Aquat. Bot., 40: 363-377. EVANS, A. S., K. L. WEBB & P. A. PENHALE, 1986: Photosynthetic temperature acclimation in two
FALKOWSKY, P. G., 2. DUBINSKY & K. WYMAN, 1985: Growth-irradiance relationships in phyto-
FOURQUREAN, J. W. & J. C. ZIEMAN, 1991: Photosynthesis, respiration and whole plant carbon budget
GIESEN, W. B. J. T.. M. M. V A N KATWIJK & C. DEN HARTOG, 1990: Eelgrass condition and turbidity
HILLMAN, K., D. 1.. WALKER, A. W. D. LARKUM & A. J. MCCOMB, 1990: Productivity and nutrient
coexisting seagrasses, Zosrera marina L. and Ruppia maririma L. Aquat. Bot., 2 4 185-197.
plankton. Limnol. Oceanogr., 30: 31 1-321.
of the seagrass Thalassiu resrudinum. Mar. Ecol. Prog. Ser., 6 9 161-170.
in the Dutch Wadden Sea. Aquat. Bot., 37: 71-85.

1 44 TERRADOS & Ros
limitation. In: A. W. D. LARKUM. A. J . MCCOMB & S. A. SHEPHERD (Eds.), Biology of Seagrasses. Elsevier. Amsterdam: 635-685.
JASSHY. A. D. & T. PLATT. 1976: Mathematical formulation of the relationship between photosynthesis and light for phytoplankton. Limnol. Oceanogr.. 21: 54&547.
JENSBN. W. A,, 1962: Botanical Histochemistry. WH Freeman and Company. San Francisco; 408 pp. KEMP. M., L. MURRAY & C. P. McRov 1990: Primary productivity. In: R. C. PHILLIPS & C. P. McRou
KNWP, W. T. & G. C. Bn.rli. 1988: The effect of wounding on the photosynthetic rates of three subtidal
LEATIIERBARROW. R.. 1982: Enrfitter: User’s manual. Elsevier, Amsterdam: 91 pp. LIHES, M., 1986: Productivity-irradiance relationships of Posidunicr ocwrnicu and its epiphytes. Aquat.
Bot.. 2 6 285 306. LITTLER, M. M., 1979: The erects of bottle volume, thallus weight, oxygen saturation levels and water
movement on apparent photosynthetic rates in marine algae. Aquat. Bot.. 7: 21-34. MARSH, J . A,. W. C. DI~NNISON & R. S. ALHERTE, 1986: Effects of temperature on photosynthesis and
respiration in eelgrass (Zo.vrwu triurinu L.). J . Exp. Mar. Biol. Ecol., 101: 257-267. MARTIN ANDRES, A. & J. de D., LUNA DEL CAsTIi.Lo. 1989: Bioestadistica para las Ciencias de la Salud.
Ediciones Norma. Madrid; 614 pp. McRov. C. P. & C. MC.MILLAN. 1977: Production ecology and physiology of seagrasses. In: C. P.
MCROY & C. HELFFERICII (Eds.), Seagrass ecosystems: a scientific perspective. Marcel Dekker, Inc., New York: 53-87.
NI:I.SON, S. G. & A. W. SIEGRIST. 1987: Comparison of mathematical expressions describing light- saturation curves for photosynthesis by tropical marine macroalgae. Bull. Mar. Sci., 41: 6 17-62?,
PEREZ. M. & J . ROMERO. 1992: Photosynthetic response to light and temperature of the seagrass C’~, tnor /u tw ~ I ~ J C / O . S U and the prediction of its seasonality. Aquat. Bot., 43: 51-62.
PIKC. H. , 1986: Seasonal aspects of photosynthesis in Posiduniu oceanica: influence ofdepth. temperature and light intensity. Aquat. Bot., 2 6 203-212.
~ . 1989: Seasonal changes in soluble carbohydrates, starch and energy content in Mediterranetun seagrasses. P.S.Z.N. I : Marine Ecology, 1 0 97-105.
RAMUS, J . , 1981: The capture and transduction of light energy. In: C. S. LOHHAN & M. J. WYNNI: (Eds.). The Biology of Seaweeds. Blackwell Scientific Publications, Oxford: 458492.
ROHEKTS. D. G. & D. J . W. MOHIARTY, 1987: Lacuna1 gas discharge as a measure of productivity in the seagrasses Zo.yrrru cupricorrii, Cymoc/oceu srrrukulu and $vrinyodiurn isoef(/di,liurn. Aquat. Bol.. 28:
RoiiLl, F. J . & R. R. SolcAi., 1969: Statistical Tables. W. H. Freeman and Company. San Francisco;
Ros. .I. D.. 1987: El Mar Menor: fisiografia y biota acuiitica de una laguna hipersalina y su evolucion reciente. In: Bases cientificas para la proteccibn de 10s humedales en Espaila. Real Academia de Ciencias Exactas, Fisicas y Naturales. Madrid: 237- 256.
SORRELL, B. K. & F. I . DROMGOOLI~, 1986: Errors in measurements of aquatic macrophyte gas exchange due to oxygen storage in internal airspaces. Aquat. Bot., 24: 103-1 14.
TI~RRAI)OS, J . . 1991: Crecimiento y produccibn de las praderas de macrofitos del Mar Menor, Murcia. Ph.D. thesis. Univ. of Murcia, Murcia; 229 pp.
~ - & J . D. Ros, 1992a: Growth and primary production of C:)niock~icc~~ noc/osu (UCRIA) ASctII~RSON in a Mediterranean coastal lagoon: the Mar Menor (SE Spain). Aquat. Bot., 4 3 63-75.
- ~ & J . D. Ros. 1992b: The influence of temperature on seasonal variation of C ~ u l r r p u pro/r/iwc (FORSSKAL.) LAMOLIROKJX photosynthesis and respiration. J . Exp. Mar. Biol. Ecol.. 162: 199- 212.
WETZEL, R. L. & P. A. PiiNiiA1.E. 1982: Production ecology of seagrass communities in thc lower Chesapeake Bay. Mar. Technol. SOC. J . , 17: 22-31.
ZIMMERMAN. R. c.. R. D. SMITH & R. s. ALHERTE, 1989: Thermal acclimation and whole-plant carbon balance in Zosrcw nrurinu L. (eelgrass). J . Exp. Mar. Biol. Ecol.. 130: 93-109.
(Eds.), Seagrass research methods. UNESCO. Paris: 153-159.
rhodophytes. Bot. Mar., 31: 149~-153.
143- 160.
253 pp.