Embed Size (px)
Citation preview

UNIVERSITE DE GENEVE Département de Biologie Moléculaire ECOLE POLYTECHNIQUE FEDERALE DE LAUSANNE Institute Suisse de Recherche Expérimentale sur le Cancer
FACULTE DES SCIENCES Professeur David Shore FACULTE DES SCIENCES DE LA VIE Professeur Joachim Lingner
Telomere Length Homeostasis in the Yeast
Saccharomyces cerevisiae
THESE
présentée à la Faculté des sciences de l’Université de Genève
pour obtenir la grade de Docteur ès sciences
par
Milica ARNERIĆ de
Split (Croatie)
Thèse n° 3939
Lausanne, 2007


Acknowledgments
My first big “thank You” goes to Joachim, who made me feel welcome and free during my time in his lab. He was a great support and a calming presence when I was panicking the most. Thanks to Susan and David – my PhD committee, for encouragement and belief in me – which still surprises me sometimes.
Since the last few years I found quite challenging in many aspect I felt privileged to have an understanding as well as lively and dynamic lab team to spend most of my days with. I love you ALL! Still, I want to mention a few past-present lab members with whom I shared more intensive moments:
Isabel honey - nothing was the same since you left. No one to teach me Swiss customs, no playing elephant, no one to save a woman in danger (me?)… We grew a lot together, and I hope for more to come!
Nele Gwendolin Ruth - we shared so much…laughs, tears, evil sense of humor, moods, problems, finally even kitchen, bathroom and the living room! And I know there is a lot of fun in front!
Gaël – you will always be my only lab husband! Thanks for taking all my moods so well; even though you left me - you are the one who was always there when needed!
Claus – I’ll never forget crazy nights in Thun! Even though we’ve never organized that Virginia Wolf party, when you are ready just call! I will wear my best Diesel jeans!
Sophie – for being kind and gentle as you are! Kathi – for almost convincing me that I am way younger then I am! Dr. Luke (ok - Brian!)– for unique humor and warm, enthusiastic nature!
Special thanks goes to my French-translators! They know who they are ;). …and finally two of my collaborators – Teresa for opening the door to the secret
life of STEX, and Mike – for being the best yeast postdoc I could have hoped for! You turned to be “all that” and even more, thank you for you patience and acceptance. You are full of surprises and I didn’t have enough yet! This time would be way harder without the knowledge that I’m loved and supported from the friends back home (including my mum ) and my dancing tribe, which keeps reminding me who I really am.


Table of Contents Résumé en français.......................................................................................................... 1
Références ................................................................................................................. 13
CHAPTER 1: Introduction............................................................................................... 17
1 Looking at chromosome ends - historical background.................................. 17
2 Telomere structure........................................................................................... 18
2.1 Telomeric DNA sequences ........................................................................ 18
2.2 Telomere Capping Structure ..................................................................... 20
2.3 Organization of the Subtelomeric Region ................................................ 23
3 Telomere Length Regulation ........................................................................... 24
3.1 The End-Replication Problem.................................................................... 24
3.2 Telomerase................................................................................................. 25
3.3 Cell Cycle Dependent Recruitment of Telomerase.................................. 28
3.4 Telomere Elongation is Coupled to DNA-replication................................ 31
3.5 A “Protein Counting Model” of Telomere Length Control........................ 33
3.6 DNA-damage Response Proteins in Telomere Length Regulation ......... 37
3.6.1 The Ku Heterodimer ........................................................................... 37
3.6.2 The MRX Complex............................................................................... 39
3.6.3 Tel1 and Mec1 Kinases...................................................................... 41
3.7 Other Regulators of Telomere Length ...................................................... 42
3.8 Recombination-Dependent Maintenance of Telomeres ......................... 43
4 Thesis Research Aims...................................................................................... 44
CHAPTER 2: Telomere Length Homeostasis Is Achieved via a Switch between
Telomerase-Extendible and -Nonextendible States ..................................................... 47
CHAPTER 3: Tel1 Kinase and Subtelomere-Bound Tbf1 Mediate Preferential
Elongation of Short Telomeres by Telomerase in Yeast............................................... 69
CHAPTER 4: Telomerase Repeat Addition Processivity is Increased at Critically Short
Telomeres in a Tel1-Dependent Manner in Saccharomyces cerevisiae .................... 85
CHAPTER 5: Discussion and Perspectives.................................................................103
References ....................................................................................................................111


1
Résumé en français
Les télomères chez la levure Saccharomyces cerevisiae Les génomes eucaryotes sont organisés en chromosomes linéaires. Les extrémités
de ces chromosomes linéaires sont à l’origine de deux problèmes pour la cellule. Le
premier problème concerne la machinerie de réplication de l’ADN qui n’est pas
capable de répliquer la totalité de l’ADN linéaire. Sans un mécanisme spécifique
pour résoudre ce problème, cela entrainerait le raccourcissement des
chromosomes, ainsi que la perte de séquences terminales à chaque cycle de
réplication et de division cellulaire. Le deuxième problème porte sur les extrémités
chromosomiques, qui, sans une protection spécifique, seraient reconnues comme
lésions double brin de l’ADN, et par conséquence soumis à des dégradations
exonucléolytiques ou des phénomènes de réparation des cassures double-brin par
recombinaison homologue ou par ligature non-homologue (NHEJ: «non-homologous
end-joining»). La réparation de type NHEJ est particulièrement dangereuse pour la
cellule, puisque ce processus entraîne la création de chromosomes dicentriques, qui
posent des problèmes considérables lors de la répartition des chromosomes
pendant l’anaphase de la mitose (cassure).
Les télomères sont des complexes nucléoprotéiques spécialisés qui ont
évolué à l’extrémité des chromosomes pour contribuer à la résolution de ces deux
problèmes: ils permettent leur réplication complète et les protègent contre
l’instabilité génétique. Les séquences ADN telomériques sont organisées de façon
similaire chez la plupart des eucaryotes; elles contiennent des plages de répétitions
courtes (Blackburn & Gall, 1978), mais dont la séquence et le nombre varient
considérablement selon les espèces. Chez la levure Saccharomyces cerevisiae, les
télomères sont composés d’environ 200-300 pb de la répétition irrégulière
5’- (TG)0–6TGGGTGT(G)1–2 -3’ (abrégée TG1-3) (Szostak & Blackburn, 1982; Shampay
et al, 1984; Forstemann & Lingner, 2001). Cette séquence dégénérée n’est pas une
caractéristique unique des télomères de S. cerevisiae car il a été démontré que la

Résumé en français
2
variation des répétitions télomériques est une caractéristique partagée par plusieurs
espèces fongiques (pour revue, voir (Lundblad, 2006)). Une autre caractéristique
propre aux télomères est la présence d’une extension simple brin 3’ sortante, riche
en guanine («G-tail»). Ces «G-tails» sont détectables tout au long du cycle cellulaire et
leur élimination est létale (Wellinger et al, 1993).
Les régions sous-télomériques de S. cerevisiae sont composées d’une
mosaïque de séquences à répétition modérée. Ces éléments répétés sont
regroupés en deux catégories: X et Y’ (Chan & Tye, 1983a; Chan & Tye, 1983b). Les
éléments sous-télomériques Y’ sont des séquences hautement conservées de 5.2 à
6.7 kb en taille, qui se retrouvent dans environ 1/2 à 2/3 des extrémités
chromosomiques, intégrés au milieu de courts blocs de séquences télomériques. En
revanche, l’élément X, dit «core element», se retrouve sur chaque extrémité
chromosomique. Il apparaît dans certains cas en combinaison avec d’autres petits
éléments sous-télomériques A, B, C et D, dénommés «subtelomeric repeats» (STR)
(Pryde et al, 1997). La taille de l’élément X est de 473 pb, et il contient des sites de
fixation pour le complexe qui reconnaît l’origine de réplication chez les levures. Les
éléments STR contiennent des blocs de répétitions télomériques de type Vertébrés
TTAGGG, qui sont reconnues par Tbf1, une protéine impliquée dans la régulation de
la taille des télomères (Brigati et al, 1993).
La télomérase
La taille des télomères est hétérogène, mais restreinte à un certain écart, spécifique
à chaque espèce. Afin de compenser la perte d’ADN provoquée par sa réplication
incomplète, la plupart des cellules eucaryotes utilisent une reverse transcriptase
spécifique, appelée télomérase, qui allonge spécifiquement les télomères (Greider &
Blackburn, 1985; Lingner et al, 1997b; Nakamura et al, 1997). Chez la levure S.
cerevisiae, cette ribonucleoprotéine est composée d’une sous-unité catalytique
(protéique) et d’une sous-unité ARN, codées par les gènes EST2 et TLC1,
respectivement (Singer & Gottschling, 1994; Lingner et al, 1997b). Ces deux sous-
unités sont suffisantes à l’activité télomérase in vitro, néanmoins, in vivo la
maintenance des télomères nécessite au moins trois protéines additionnelles: Est1
et Est3 (deux sous-unités protéiques de la télomérase) et une protéine Cdc13 qui se

Résumé en français
3
lie à l’extension simple brin 3’ sortante des télomères (Lendvay et al, 1996; Lingner
et al, 1997a). L’absence d’un seul de ces composants provoque un
raccourcissement des télomères qui s’accompagne d’une perte de viabilité
cellulaire; ce phénomène étant dénommé «sénescence» (Lundblad & Szostak,
1989).
L’élongation des télomères a lieu en phase S tardive (Marcand et al, 2000),
même si l’activité enzymatique de la télomérase peut être détectée dans des
extraits obtenus dans d’autres phases du cycle cellulaire (Diede & Gottschling,
1999). Deux explications sont possibles pour cette restriction de l’élongation à une
phase donnée du cycle cellulaire: le recrutement de la télomérase pourrait être
restreint à un moment particulier, ou l’activation de la télomérase ne se produit
qu’après sa fixation au télomère. Le recrutement de la télomérase aux télomères
nécessite une interaction avec la protéine Cdc13 qui se lie à l’ADN simple brin des
extrémités télomériques, ou avec l’hétérodimère Ku, qui se lie aux lésions d’ADN
double brin et reconnaît aussi les extrémités télomériques (Qi & Zakian, 2000;
Peterson et al, 2001). La fixation de la protéine Cdc13 aux télomères atteint un
maximum en phase S, coïncidant avec l’extension de l’extrémité simple brin riche en
guanine (Taggart et al, 2002). Le fait que Cdc13 interagit avec Est1 suggère que
Cdc13 serait responsable du recrutement de la télomérase en phase S (Evans &
Lundblad, 1999; Taggart et al, 2002). Cependant, la télomérase (Est2) est présente
au niveau des télomères pendant les phases G1 et S. Le recrutement de la
télomérase en phase G1 dépend ainsi de son interaction avec Ku, tandis que Ku et
Cdc13 contribuent au recrutement en phase S (Taggart et al, 2002; Fisher et al,
2004). En outre, Est1 est dégradée par l’activité protéosomale en phase G1, ce qui
empêche l’assemblage du complexe télomérique (Osterhage et al, 2006). La
télomérase agit exclusivement sur des extensions télomériques 3’ sortantes simple
brin, en catalysant leur élongation par reverse transcription de la matrice de sa
propre sous-unité ARN. Le brin d’ADN complémentaire est vraisemblablement
synthétisé par les ADN polymérases conventionnelles α et δ. D’ailleurs, la
coordination entre la télomérase et l’ADNpolymérase dédiée à la réplication du brin
retardé est nécessaire à la préservation des télomères (Carson & Hartwell, 1985;
Diede & Gottschling, 1999; Adams Martin et al, 2000).

Résumé en français
4
Le modèle «comptage protéique» du contrôle de taille des télomères
La protéine Rap1 est le composant majeur de la chromatine télomérique . Elle fixe
les répétitions télomériques avec une haute affinité (Conrad et al, 1990). La partie
C-terminale de Rap1 recrute deux protéines, Rif1 et Rif2 et ce complexe exerce une
influence négative sur la taille des télomères (Lustig et al, 1990; Hardy et al, 1992;
Wotton & Shore, 1997). La fixation de molécules additionnelles de Rap1 sur le
télomère induit un raccourcissement du télomère proportionnellement au nombre de
molécules Rap1 fixées (Marcand et al, 1997). Cette observation est la base
dumodèle dit du «comptage protéique», qui explique la régulation de la taille des
télomères (Marcand et al, 1997). Selon ce modèle, l’extension des télomères par la
télomérase est régulée par le nombre de complexes Rap1-Rif1 et -Rif2 fixés au
télomère. Quand le télomère atteint une certaine taille, le nombre croissant de
complexes Rap1 fixés produirait le signal qui empêche l’action de la télomérase. La
télomérase étant inhibée, le télomère est progressivement raccourci jusqu'à ce que
le nombre de complexes fixés ne décroisse en dessous d’un certain seuil. En
dessous de ce seuil, la télomérase pourrait à nouveau allonger le télomère.
Marcand et al. (1999) ont démontré chez S. cerevisiae que l’élongation
moyenne des télomères est restreinte à seulement quelques paires de bases qui
sont ajoutées à chaque génération, et que la répression de l’activité de la
télomérase se fait de manière progressive (Marcand et al, 1999). En revanche, la
vélocité du raccourcissement des télomères est constante (autour de 3 paires de
bases par génération) et ne dépend pas de la taille des télomères. Deux pistes sont
a priori envisageables pour expliquer que les télomères courts sont plus rapidement
allongés: soit la télomérase ajoute des répétitions plus longues sur les télomères
courts, soit les télomères courts sont plus fréquemment allongés. Il est aussi
envisageable que les deux mécanismes soit co-existants. Afin de discerner entre
ces possibilités, un système in vivo a été mis au point dans lequel l’élongation d’un
seul télomère peut être suivie au nucléotide près au cours d’un seul cycle cellulaire
chez S. cerevisiae. Cette approche expérimentale a été dénommée STEX («single
telomere extension assay»).

Résumé en français
5
Chapitre 2: L’homéostasie de la taille des télomères dépend de
l’équilibre entre deux états, extensible et non-extensible par la
télomérase.
Résultats et discussion
En utilisant l’approche STEX, nous avons déterminé la fréquence et l’efficacité de
l’élongation des télomères en fonction de leur taille (Chapitre 2) (Teixeira et al,
2004). Dans ces expériences, nous avons conjugué une souche de levure
télomérase-négative avec une souche télomérase-positive, toutes deux haploïdes et
de sexe différent. Les télomères raccourcis, en provenance du parent télomérase-
négatif, sont réallongés dans le zygote, grâce à la présence de la télomérase issue
de l’autre parent. L’élongation des télomères est ensuite détectée par amplification,
clonage et séquençage de la population d’un télomère spécifique du parent
télomérase-négatif. Puisque la télomérase de levure ajoute des répétitions
imparfaites, l’élongation des télomères est détectable par la divergence entre les
répétitions télomériques distales et l’ADN télomérique synthétisé par la réplication
semi-conservative qui ne présente aucune variation durant l’expansion clonale.
L’analyse des séquences télomériques a ainsi révélé que l’élongation des
télomères est un processus stochastique, qui ne se produit pas au niveau de chaque
télomère à chaque cycle cellulaire. Cependant les télomères courts ont une chance
plus élevée d’être allongés: la fréquence d’extension décroît de façon linéaire avec
la taille du télomère, de 42-46% des télomères de 100 nucléotides à 6-8% des
télomères de 300 nucléotides. Le nombre de répétitions qui peuvent être ajoutées
à untélomèreà chaque cycle cellulaire varie, mais le nombre de nucléotides ajoutés
n’est pas corrélé à la taille du télomère (pour les télomères de plus de 100 pb). En
dessous de ce seuil de 100 pb, la télomérase ajoute beaucoup plus de répétitions
télomériques. La fréquence de l’extension par la télomérase augmente dans les
mutants rif1Δ et rif2Δ. En conclusion, nos résultats soutiennent un modèle selon
lequel les télomères existent dans au moins deux états structuraux, et ces états
influencent directement la capacité de la télomérase à catalyser leur extension.
Selon ce modèle que nous proposons, il y a une forte probabilité qu’un
télomère court adopte une structure extensible par la télomérase. Après élongation,
cet équilibre bascule vers un état non-extensible. La probabilité que le télomère soit

Résumé en français
6
allongé décroît avec l’augmentation de sa taille. La taille moyenne des télomères
correspond alors à une taille dont la probabilité d’extension, multipliée par la taille
moyenne d’extension est égale à la vitesse de raccourcissement. Les protéines Rif1
et Rif2 ont été identifiées comme des médiateurs cruciaux de l’état des télomères.
Comment ces protéines arrivent-elles à induire un état non-extensible et comment
arrivent-elles à empêcher l’association productive entre télomérase et extrémité 3’
du télomère d’une manière dépendante de la taille? Les complexes Rap1-Rif1 et –
Rif2 pourraient effectuer cela en favorisant la formation d’une structure d’ADN
alternative, comme par exemple des quartets de G, connus comme inhibiteurs de
l’activité télomérase in vitro (Zahler et al, 1991). Effectivement, il a été démontré
que Rap1 favorise la formation des quartets de G in vitro (Giraldo & Rhodes, 1994).
Une autre structure inaccessible à la télomérase pourrait être la boucle t, décrit chez
les Vertébrés et les Protozoaires, et dans laquelle l’extrémité 3’ télomérique est
masquée par repliement et hybridation à la région télomérique proximale,
conduisant à la formation d’une boucle de déplacement («D-loop») (Griffith et al,
1999; Murti & Prescott, 1999; Munoz-Jordan et al, 2001). Ces structures repliés
(fold-back structures) ont été mises en évidence aussi chez S. cerevisiae, mais l’état
de l’extrémité 3’ simple brin dans ces structures n’est pas claire (Strahl-Bolsinger et
al, 1997; de Bruin et al, 2001). Néanmoins des mutations perturbant cette
structure ne causent pas de phénotype télomérique (Palladino et al, 1993). Il est
possible que dans ce cas la présence des protéines Rif empêche l’association d’un
facteur inconnu qui active la télomérase, mais pas l’accès de la télomérase en soi.
Les kinases Tel1 et Mec1
Bien que par définition, les télomères empêchent l’extrémité des chromosomes
d’activer la réponse aux dommages de l’ADN, un aspect surprenant du maintien des
télomères réside dans le fait que beaucoup de protéines de réparation de l’ADN et
de «point de contrôle», incluant l’hétérodimère Ku, le complexe MRX, Tel1 et Mec1
s’associent avec les télomères et contribuent à leur homéostasie (d'Adda di Fagagna
et al, 2004). Des levures privées de Tel1 présentent des télomères courts mais
stables alors que la délétion de MEC1 ne se traduit que par un raccourcissement
modéré des télomères (Greenwell et al, 1995; Ritchie et al, 1999). Les kinases Tel1

Résumé en français
7
et Mec1 semblent fonctionner de façon partiellement redondante pour le maintien
de la longueur des télomères puisque la délétion simultanée de TEL1 et MEC1
conduit à la sénescence cellulaire (Ritchie et al, 1999). En dépit du rôle critique de
Tel1 et Mec1 dans le maintien des télomères, les mécanismes moléculaires de
l’action de ces protéines restent très mal connus. Tel1 n’affecte pas directement
l’activité catalytique de la télomérase mais semble plutôt modifier la structure du
complexe télomérique (Chan et al, 2001; Tsukamoto et al, 2001). De façon
intéressante, la protéine Cdc13 qui se lie à la région simple brin des télomères a été
identifiée comme substrat de ces kinases (Tseng et al, 2006).
Tel1 semble fonctionner comme médiateur dans le mécanisme de
«comptage protéique» par Rap1(Ray & Runge, 1999). Il a en effet été observé dans
des levures tel1∆ que des complexes Rap1 additionnels adjacents aux répétitions
télomériques ne sont pas comptabilisés dans la longueur des télomères (Ray &
Runge, 1999). De plus, des répétitions télomériques humaines, qui ne sont pas
reconnues par Rap1, intégrées aux séquences télomériques de levure, sont
comptées dans ce fond génétique, ce qui n’est pas le cas dans un contexte sauvage
(Brevet et al, 2003). De façon surprenante, l’inactivation du mécanisme de
comptage par Rap1 par délétion de TEL1 a donc permis de révéler l’existence d’un
mécanisme additionnel cryptique de régulation de la longueur des télomères. Celui-
ci implique potentiellement deux protéines insulatrices essentielles des régions
sous-télomériques de la levure: Tbf1 et Reb1 (Bilaud et al, 1996; Fourel et al, 1999;
Koering et al, 2000). Tbf1 lie directement les répétitions télomériques in vitro, et
Tbf1 et Reb1 affectent la longueur des télomères quand ils sont ciblés sur les
télomères de levure dans le fond génétique tel1∆ (Brigati et al, 1993; Alexander &
Zakian, 2003; Berthiau et al, 2006).
Qui plus est, en absence de Tel1, Tbf1 et Reb1 se comportent comme
facteurs «d’anti-ancrage», empêchant la localisation des télomères au voisinage de
l’enveloppe nucléaire, suggérant l’existence d’un lien entre ancrage à la périphérie
nucléaire et régulation de la longueur des télomères (Gotta et al, 1996; Hediger et
al, 2006).

Résumé en français
8
Chapitre 3: La kinase Tel1 et la fraction sous-télomérique de Tbf1
assurent l’élongation préférentielle des télomères courts par la
télomérase de levure
Résultats et discussion
La télomérase assure l’homéostasie de la longueur des télomères et montre une
préférence pour les télomères inversement proportionnelle à leur longueur. Cette
régulation implique la liaison de Rap1 aux séquences répétées télomériques, qui
constitue un signal de rétrocontrôle négatif dépendant de la taille, ainsi que les
kinases Tel1 et Mec1 qui sont des régulateurs positifs de la longueur des télomères.
En analysant l’élongation des télomères de chromosomes natifs à une résolution
moléculaire (tests STEX décrits), je montre que dans des cellules tel1∆, la fréquence
d’élongation globale décroît considérablement, expliquant le phénotype associé de
télomère court. Dans ces cellules tel1∆, l’élongation d’un télomère artificiel privé de
région sous-télomérique n’augmente plus proportionnellement au raccourcissement
des télomères endogènes. A l’inverse, un télomère naturel comprenant la séquence
sous-télomérique montre toujours une élongation préférentielle des télomères
courts. Je démontre grâce à l’utilisation d’une autre version étiquetée de télomère
que la liaison de Gbd-Tbf1N mais pas celle de Gbd seul, sauve le phénotype de
délétion de TEL1 relatif à l’activation de la télomérase au niveau des télomères
courts. La liaison de la protéine sous-télomérique Tbf1 à un télomère artificiel dans
des cellules tel1∆ est donc suffisante pour restaurer l’action préférentielle de la
télomérase vis-à-vis des télomères courts. Par conséquent, Tbf1 pourrait
fonctionner parallèlement à Tel1, une protéine cruciale pour l’activation de la
télomérase au niveau des télomères courts, sous le contrôle de séquences répétées
TG.
A ce jour, les mécanismes sous-jacents par lesquels Tel1 reconnaît les
télomères courts ne sont pas connus. Il est toutefois intéressant de noter que Tel1
peut phosphoryler Cdc13 in vitro (Tseng et al, 2006), suggérant que Tel1 pourrait
réguler l’étape de recrutement de la télomérase. La découverte récente que Tel1
est requise pour la pleine association de Est1 et Est2 à l’ADN télomérique est
cohérente avec cette hypothèse (Goudsouzian et al, 2006). De plus, il a été montré
très récemment que les télomères courts sont plus avidement liés par Est1, Est2 et

Résumé en français
9
Tel1 (Bianchi & Shore, 2007; Hector et al, 2007; Sabourin et al, 2007).
Curieusement, la fréquence d’extension d’un télomère naturel est toujours régulée
en fonction de sa longueur, même en absence de TEL1, ce qui révèle une différence
essentielle entre versions naturelle et étiquetée des télomères. En effet, bien que la
liaison de Tbf1 au voisinage des répétitions télomériques suffit à restaurer l’action
préférentielle de la télomérase au niveau des télomères courts, ce mécanisme
semble conditionné à la délétion de TEL1 puisque la modification du nombre de
répétitions 5’-TTAGGG-3’ au sein des télomères de levure n’affecte la longueur des
télomères que dans le fond génétique tel1∆ mais pas dans le contexte sauvage
(Brevet et al, 2003). Nous postulons que le mécanisme de «secours» dépendant de
Tbf1 pourrait s’avérer extrêmement important pour la réparation des télomères qui
auraient accidentellement perdu tout ou partie de leurs séquences répétées ainsi
que leurs sites de liaison à Rap1, suite à une cassure double-brin proche de la base
du télomère ou à l’effondrement de la fourche de réplication. En effet, il est bien
établi que l’ADN télomérique constitue un obstacle à la machinerie de réplication
semi-conservative (voir par exemple (Cooper et al, 1997; Ivessa et al, 2002; Crabbe
et al, 2004)), et la perte de l’intégralité du télomère est un évènement qui pourrait
survenir à une fréquence faible mais toutefois significative.
La processivité de la Télomérase
Afin d’efficacement allonger les télomères, la télomérase humaine doit ajouter
plusieurs répétitions télomériques lors d’un même cycle cellulaire puisqu’elle doit
compenser la perte moyenne de 50-200 nucléotides par cycle de réplication (Harley
et al, 1990; Huffman et al, 2000). In vitro, la télomérase humaine peut allonger une
amorce télomérique de manière processive (Greider, 1991), ce qui n’est pas le cas
de l’enzyme de levure (Lingner et al, 1997a). L’activité télomérase d’extraits de
levure est très inefficace en terme de translocation: l’enzyme reste en effet
stablement liée à l’amorce étendue, après l’addition d’une unique séquence
télomérique (Prescott & Blackburn, 1997a; Prescott & Blackburn, 1997b). La
signification physiologique de ce comportement in vitro n’est pas claire puisque les
télomères de levure ne perdant que ~3 nucléotides par cycle, l’ajout d’une seule
répétition devrait être suffisante pour maintenir la longueur des télomères (Lundblad

Résumé en français
10
& Szostak, 1989). En effet, l’élongation des télomères est limitée à quelques paires
de bases par génération du télomère moyen et ce taux diminue progressivement
avec l’allongement des télomères (Marcand et al, 1999). Cependant, l’analyse in
vivo présentée au Chapitre 2 suggère que la télomérase de levure peut ajouter >100
nt dans un même cycle cellulaire, suggérant qu’in vivo, l’enzyme est soit processive,
soit très rapidement échangée.
Chapitre 4: La processivité de la télomérase est augmentée de façon
dépendante de Tel1 au niveau des télomères courts critiques chez
Saccharomyces cerevisiae
Résultats et discussion
Grâce à la co-expression de deux sous-unités ARN comportant des séquences
télomériques répétées distinctes et en examinant les extensions télomériques après
un cycle cellulaire, nous avons déterminé que la télomérase de S. cerevisiae peut se
dissocier et se réassocier à un télomère donné durant un cycle cellulaire. Nous
avons aussi confirmé que la télomérase n’est pas processive en termes d’addition
de séquences répétées. Toutefois, cette processivité est significativement
augmentée au niveau des télomères très courts. Nous proposons que cette
augmentation de la processivité d’addition de répétitions de la télomérase aux
télomères courts est mise à profit pour rapidement allonger des télomères
présentant une longueur inférieure à un seuil critique.
L’augmentation de processivité de la télomérase aux télomères courts est un
processus dépendant de Tel1. Je mentionnais déjà au Chapitre 3 que dans des
cellules tel1Δ, les séquences ajoutées au niveau des télomères courts n’étaient pas
plus longues que celles ajoutées aux télomères plus longs (en moyenne 42 nt pour
les télomères <100 bp contre 44 nt pour les télomères plus longs, au télomère
natif). Ceci semble révéler un rôle dual de Tel1: d’une part comme senseur de la
longueur télomérique via le mécanisme de comptage protéique dépendant de Rap1
et d’autre part comme stimulateur de l’addition de séquences répétées au niveau
des extrémités télomériques courtes. La surexpression de Tel1 conduit à
l’élongation des télomères (Hector et al, 2007; Sabourin et al, 2007), il serait donc

Résumé en français
11
intéressant de savoir laquelle de ces deux fonctions de Tel1 est responsable de ce
phénotype. Les télomères sont-ils dans ce contexte plus fréquemment allongés ou
bien la processivité de la télomérase est-elle augmentée, y compris au niveau de
télomères plus longs? De même, le rôle de l’activité kinase de Tel1 sur la
processivité de la télomérase n’est pas établi mais si cette activité s’avérait être
requise, on peut se demander quel substrat de Tel1 serait impliqué dans ce
processus. A ce jour, le seul substrat télomérique avéré de Tel1 est Cdc13 (Tseng et
al, 2006) mais des travaux récents suggèrent que Rif1 pourrait aussi être
phosphorylé par Tel1, une modification dont la fonction reste cependant totalement
mystérieuse (Smolka et al, 2007)


13
Références
Adams Martin A, Dionne I, Wellinger RJ, Holm C (2000) The function of DNA polymerase alpha at telomeric G tails is important for telomere homeostasis. Mol Cell Biol 20: 786-796.
Alexander MK, Zakian VA (2003) Rap1p telomere association is not required for mitotic stability of a C(3)TA(2) telomere in yeast. Embo J 22: 1688-1696.
Berthiau AS, Yankulov K, Bah A, Revardel E, Luciano P, Wellinger RJ, Geli V, Gilson E (2006) Subtelomeric proteins negatively regulate telomere elongation in budding yeast. Embo J 25: 846-856.
Bianchi A, Shore D (2007) Increased association of telomerase with short telomeres in yeast. Genes Dev 21: 1726-1730.
Bilaud T, Koering CE, Binet-Brasselet E, Ancelin K, Pollice A, Gasser SM, Gilson E (1996) The telobox, a Myb-related telomeric DNA binding motif found in proteins from yeast, plants and human. Nucleic Acids Res 24: 1294-1303.
Blackburn EH, Gall JG (1978) A tandemly repeated sequence at the termini of the extrachromosomal ribosomal RNA genes in Tetrahymena. J Mol Biol 120: 33-53.
Brevet V, Berthiau AS, Civitelli L, Donini P, Schramke V, Geli V, Ascenzioni F, Gilson E (2003) The number of vertebrate repeats can be regulated at yeast telomeres by Rap1-independent mechanisms. Embo J 22: 1697-1706.
Brigati C, Kurtz S, Balderes D, Vidali G, Shore D (1993) An essential yeast gene encoding a TTAGGG repeat-binding protein. Mol Cell Biol 13: 1306-1314.
Carson MJ, Hartwell L (1985) CDC17: an essential gene that prevents telomere elongation in yeast. Cell 42: 249-257.
Chan CS, Tye BK (1983a) A family of Saccharomyces cerevisiae repetitive autonomously replicating sequences that have very similar genomic environments. J Mol Biol 168: 505-523.
Chan CS, Tye BK (1983b) Organization of DNA sequences and replication origins at yeast telomeres. Cell 33: 563-573.
Chan SW, Chang J, Prescott J, Blackburn EH (2001) Altering telomere structure allows telomerase to act in yeast lacking ATM kinases. Curr Biol 11: 1240-1250.
Conrad MN, Wright JH, Wolf AJ, Zakian VA (1990) RAP1 protein interacts with yeast telomeres in vivo: overproduction alters telomere structure and decreases chromosome stability. Cell 63: 739-750.
Cooper JP, Nimmo ER, Allshire RC, Cech TR (1997) Regulation of telomere length and function by a Myb-domain protein in fission yeast. Nature 385: 744-747.
Crabbe L, Verdun RE, Haggblom CI, Karlseder J (2004) Defective telomere lagging strand synthesis in cells lacking WRN helicase activity. Science 306: 1951-1953.
d'Adda di Fagagna F, Teo SH, Jackson SP (2004) Functional links between telomeres and proteins of the DNA-damage response. Genes Dev 18: 1781-1799.
de Bruin D, Zaman Z, Liberatore RA, Ptashne M (2001) Telomere looping permits gene activation by a downstream UAS in yeast. Nature 409: 109-113.
Diede SJ, Gottschling DE (1999) Telomerase-mediated telomere addition in vivo requires DNA primase and DNA polymerases alpha and delta. Cell 99: 723-733.
Evans SK, Lundblad V (1999) Est1 and Cdc13 as comediators of telomerase access. Science 286: 117-120.
Fisher TS, Taggart AK, Zakian VA (2004) Cell cycle-dependent regulation of yeast telomerase by Ku. Nat Struct Mol Biol 11: 1198-1205.
Forstemann K, Lingner J (2001) Molecular basis for telomere repeat divergence in budding yeast. Mol Cell Biol 21: 7277-7286.
Fourel G, Revardel E, Koering CE, Gilson E (1999) Cohabitation of insulators and silencing elements in yeast subtelomeric regions. Embo J 18: 2522-2537.
Giraldo R, Rhodes D (1994) The yeast telomere-binding protein RAP1 binds to and promotes the formation of DNA quadruplexes in telomeric DNA. Embo J 13: 2411-2420.
Gotta M, Laroche T, Formenton A, Maillet L, Scherthan H, Gasser SM (1996) The clustering of telomeres and colocalization with Rap1, Sir3, and Sir4 proteins in wild-type Saccharomyces cerevisiae. J Cell Biol 134: 1349-1363.

Résumé en français: Références
14
Goudsouzian LK, Tuzon CT, Zakian VA (2006) S. cerevisiae Tel1p and Mre11p are required for normal levels of Est1p and Est2p telomere association. Mol Cell 24: 603-610.
Greenwell PW, Kronmal SL, Porter SE, Gassenhuber J, Obermaier B, Petes TD (1995) TEL1, a gene involved in controlling telomere length in S. cerevisiae, is homologous to the human ataxia telangiectasia gene. Cell 82: 823-829.
Greider CW, Blackburn EH (1985) Identification of a specific telomere terminal transferase activity in Tetrahymena extracts. Cell 43: 405-413.
Greider CW (1991) Telomerase is processive. Mol Cell Biol 11: 4572-4580. Griffith JD, Comeau L, Rosenfield S, Stansel RM, Bianchi A, Moss H, de Lange T (1999) Mammalian
telomeres end in a large duplex loop. Cell 97: 503-514. Hardy CF, Sussel L, Shore D (1992) A RAP1-interacting protein involved in transcriptional silencing
and telomere length regulation. Genes Dev 6: 801-814. Harley CB, Futcher AB, Greider CW (1990) Telomeres shorten during ageing of human fibroblasts.
Nature 345: 458-460. Hector RE, Shtofman RL, Ray A, Chen BR, Nyun T, Berkner KL, Runge KW (2007) Tel1p preferentially
associates with short telomeres to stimulate their elongation. Mol Cell 27: 851-858. Hediger F, Berthiau AS, van Houwe G, Gilson E, Gasser SM (2006) Subtelomeric factors antagonize
telomere anchoring and Tel1-independent telomere length regulation. Embo J 25: 857-867. Huffman KE, Levene SD, Tesmer VM, Shay JW, Wright WE (2000) Telomere shortening is proportional
to the size of the G-rich telomeric 3'-overhang. J Biol Chem 275: 19719-19722. Ivessa AS, Zhou JQ, Schulz VP, Monson EK, Zakian VA (2002) Saccharomyces Rrm3p, a 5' to 3' DNA
helicase that promotes replication fork progression through telomeric and subtelomeric DNA. Genes Dev 16: 1383-1396.
Koering CE, Fourel G, Binet-Brasselet E, Laroche T, Klein F, Gilson E (2000) Identification of high affinity Tbf1p-binding sites within the budding yeast genome. Nucleic Acids Res 28: 2519-2526.
Lendvay TS, Morris DK, Sah J, Balasubramanian B, Lundblad V (1996) Senescence mutants of Saccharomyces cerevisiae with a defect in telomere replication identify three additional EST genes. Genetics 144: 1399-1412.
Lingner J, Cech TR, Hughes TR, Lundblad V (1997a) Three Ever Shorter Telomere (EST) genes are dispensable for in vitro yeast telomerase activity. Proc Natl Acad Sci U S A 94: 11190-11195.
Lingner J, Hughes TR, Shevchenko A, Mann M, Lundblad V, Cech TR (1997b) Reverse transcriptase motifs in the catalytic subunit of telomerase. Science 276: 561-567.
Lundblad V, Szostak JW (1989) A mutant with a defect in telomere elongation leads to senescence in yeast. Cell 57: 633-643.
Lundblad V (2006) Budding Yeast Telomeres. In: de Lange T, Lundblad V, Blackburn E (eds), Telomeres. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, pp. 345-386.
Lustig AJ, Kurtz S, Shore D (1990) Involvement of the silencer and UAS binding protein RAP1 in regulation of telomere length. Science 250: 549-553.
Marcand S, Gilson E, Shore D (1997) A protein-counting mechanism for telomere length regulation in yeast. Science 275: 986-990.
Marcand S, Brevet V, Gilson E (1999) Progressive cis-inhibition of telomerase upon telomere elongation. Embo J 18: 3509-3519.
Marcand S, Brevet V, Mann C, Gilson E (2000) Cell cycle restriction of telomere elongation. Curr Biol 10: 487-490.
Munoz-Jordan JL, Cross GA, de Lange T, Griffith JD (2001) t-loops at trypanosome telomeres. Embo J 20: 579-588.
Murti KG, Prescott DM (1999) Telomeres of polytene chromosomes in a ciliated protozoan terminate in duplex DNA loops. Proc Natl Acad Sci U S A 96: 14436-14439.
Nakamura TM, Morin GB, Chapman KB, Weinrich SL, Andrews WH, Lingner J, Harley CB, Cech TR (1997) Telomerase catalytic subunit homologs from fission yeast and human. Science 277: 955-959.
Osterhage JL, Talley JM, Friedman KL (2006) Proteasome-dependent degradation of Est1p regulates the cell cycle-restricted assembly of telomerase in Saccharomyces cerevisiae. Nat Struct Mol Biol 13: 720-728.
Palladino F, Laroche T, Gilson E, Axelrod A, Pillus L, Gasser SM (1993) SIR3 and SIR4 proteins are required for the positioning and integrity of yeast telomeres. Cell 75: 543-555.
Peterson SE, Stellwagen AE, Diede SJ, Singer MS, Haimberger ZW, Johnson CO, Tzoneva M, Gottschling DE (2001) The function of a stem-loop in telomerase RNA is linked to the DNA repair protein Ku. Nat Genet 27: 64-67.

Résumé en français: Références
15
Prescott J, Blackburn EH (1997a) Functionally interacting telomerase RNAs in the yeast telomerase complex. Genes Dev 11: 2790-2800.
Prescott J, Blackburn EH (1997b) Telomerase RNA mutations in Saccharomyces cerevisiae alter telomerase action and reveal nonprocessivity in vivo and in vitro. Genes Dev 11: 528-540.
Pryde FE, Gorham HC, Louis EJ (1997) Chromosome ends: all the same under their caps. Curr Opin Genet Dev 7: 822-828.
Qi H, Zakian VA (2000) The Saccharomyces telomere-binding protein Cdc13p interacts with both the catalytic subunit of DNA polymerase alpha and the telomerase-associated est1 protein. Genes Dev 14: 1777-1788.
Ray A, Runge KW (1999) Varying the number of telomere-bound proteins does not alter telomere length in tel1Delta cells. Proc Natl Acad Sci U S A 96: 15044-15049.
Ritchie KB, Mallory JC, Petes TD (1999) Interactions of TLC1 (which encodes the RNA subunit of telomerase), TEL1, and MEC1 in regulating telomere length in the yeast Saccharomyces cerevisiae. Mol Cell Biol 19: 6065-6075.
Sabourin M, Tuzon CT, Zakian VA (2007) Telomerase and Tel1p Preferentially Associate with Short Telomeres in S. cerevisiae. Mol Cell.
Shampay J, Szostak JW, Blackburn EH (1984) DNA sequences of telomeres maintained in yeast. Nature 310: 154-157.
Singer MS, Gottschling DE (1994) TLC1: template RNA component of Saccharomyces cerevisiae telomerase. Science 266: 404-409.
Smolka MB, Albuquerque CP, Chen SH, Zhou H (2007) Proteome-wide identification of in vivo targets of DNA damage checkpoint kinases. Proc Natl Acad Sci U S A 104: 10364-10369.
Strahl-Bolsinger S, Hecht A, Luo K, Grunstein M (1997) SIR2 and SIR4 interactions differ in core and extended telomeric heterochromatin in yeast. Genes Dev 11: 83-93.
Szostak JW, Blackburn EH (1982) Cloning yeast telomeres on linear plasmid vectors. Cell 29: 245-255.
Taggart AK, Teng SC, Zakian VA (2002) Est1p as a cell cycle-regulated activator of telomere-bound telomerase. Science 297: 1023-1026.
Teixeira MT, Arneric M, Sperisen P, Lingner J (2004) Telomere length homeostasis is achieved via a switch between telomerase- extendible and -nonextendible states. Cell 117: 323-335.
Tseng SF, Lin JJ, Teng SC (2006) The telomerase-recruitment domain of the telomere binding protein Cdc13 is regulated by Mec1p/Tel1p-dependent phosphorylation. Nucleic Acids Res 34: 6327-6336.
Tsukamoto Y, Taggart AK, Zakian VA (2001) The role of the Mre11-Rad50-Xrs2 complex in telomerase- mediated lengthening of Saccharomyces cerevisiae telomeres. Curr Biol 11: 1328-1335.
Wellinger RJ, Wolf AJ, Zakian VA (1993) Saccharomyces telomeres acquire single-strand TG1-3 tails late in S phase. Cell 72: 51-60.
Wotton D, Shore D (1997) A novel Rap1p-interacting factor, Rif2p, cooperates with Rif1p to regulate telomere length in Saccharomyces cerevisiae. Genes Dev 11: 748-760.
Zahler AM, Williamson JR, Cech TR, Prescott DM (1991) Inhibition of telomerase by G-quartet DNA structures. Nature 350: 718-720.


17
Chapter 1: Introduction
Eukaryotic genomes are organized into multiple linear chromosomes. The ends of
linear DNA create two main problems for cells to solve. First, the semi-conservative
DNA-replication machinery is not able to replicate linear DNA to completion. Without
a specific mechanism to solve this problem, this would lead to chromosome
shortening and loss of sequences at chromosome ends with each round of DNA
replication and cell division. Second, uncapped chromosome ends would be
recognized as accidentally occurring double strand breaks (DSBs) that would be
subjected to exonucleolytic degradation, recombination, and end-to-end joining.
End-to-end fusions particularly would have fatal consequences for the cell, since di-
centric chromosomes would be created, which would cause problems during cell
division.
In order to cope with these problems, the ends of linear chromosomes are
organized into essential protein-DNA complexes – telomeres. Telomeres allow a way
to complete chromosome-end synthesis by the activity of a reverse transcriptase
called telomerase, which synthesizes new telomeric DNA. Telomeres also have
structural properties that protect them from recognition as DSBs. In the following
sections, I will describe telomere organization and the mechanisms of their
preservation and protection, mainly focusing on the lessons learned from budding
yeast, Saccharomyces cerevisiae. Furthermore, since the main focus of my research
was to understand the regulation of telomere length homeostasis, I specifically
highlight factors and processes that affect telomere length regulation.
1 Looking at chromosome ends - historical background
Telomere biology started in the 1930s, primarily by the cytological work of Barbara
McClintock and Hermann Muller (reviewed in (Blackburn, 2006)). In the 1920s,
Muller mutagenized fruit flies with X-rays and was analyzing products of the X-ray-

Introduction
18
induced broken chromosomes: inversions, translocations, and deficiencies.
Surprisingly, rearrangements including terminal regions of chromosomes were
strikingly under-represented, suggesting the presence of protective structures at
natural chromosome ends, preventing their end-to-end joining with other
mutagenized, broken DNA pieces. In his 1938 paper, Muller named those natural,
protected “free ends” the “telomeres” (Muller, 1938).
Muller’s contemporary, Barbara McClintock, developed microscopic
visualization methods by which she could distinguish special “knobs”, or
heterochromatic regions, including some at the chromosome ends, as landmarks
along chromosome arms distinguishing the individual chromosomes of maize cells.
Monitoring X-ray-induced chromosome abnormalities in mutagenized maize, she as
well found that no piece of broken chromosome was joined to an intact natural
chromosome-end. Thus, by 1931, McClintock had already recognized that the end
of an intact chromosome has distinct functional properties, functionally
distinguishing it from a broken DNA end (McClintock, 1931). In her 1939 paper, she
suggested that the new telomeres could be formed by an active process (McClintock,
1939). She noticed that a broken end can lose its tendency to fuse with another
broken end, in other words it can “heal” and stay stable, presumably by adopting the
structure of a normal telomere. The “healing” of a broken end was noticed only in
young plant embryos, an observation which already then suggested that there was
some specific factor – as we know today the reverse transcriptase telomerase -
allowing telomere synthesis in developing organisms. So already in the 1930s,
individual work from two remarkable geneticists led to the same conclusion:
telomeres are special functional structures, protecting the chromosome-ends from
being processed as DNA-damage.
2 Telomere structure
2.1 Telomeric DNA sequences
The first telomeric DNA sequence determined belongs to the telomeres of rDNA
minichromosomes in the somatic nucleus of the ciliated protozoan Tetrahymena
thermophila. These linear rDNA molecules contain 20-70 tandem repeats of 5’-
TTGGGG-3’ sequence on their ends (Blackburn & Gall, 1978). Telomeric sequences

Introduction
19
of most eukaryotes are organized in a similar manner, even though the actual
sequence varies between species. The number of telomeric repeats also
significantly differs between species and the length of telomeres is inherited (Kipling
& Cooke, 1990; Starling et al, 1990; Zhu et al, 1998).
When S. cerevisiae telomeres were subcloned and sequenced a few years
after Tetrayhmena by Szostak and Blackburn, it came as a surprise that they consist
of 200-300 bp of irregular tandem repeats with a consensus sequence of 5’-(TG)0–
6TGGGTGT(G)1–2-3’ (Szostak & Blackburn, 1982; Shampay et al, 1984; Forstemann
& Lingner, 2001) (see as well Section 3.2.). Even though irregular repeats are more
the exception, there are several other fungal species, including the evolutionarily
distant fission yeast, where telomeric sequence degeneracy is observed (reviewed in
(Lundblad, 2006)). Detailed analysis of clonally derived telomeres revealed a
centromere proximal region of 40-100 bp that rarely displays sequence variation,
while at the extreme termini sequences were highly dynamic (Wang & Zakian, 1990;
Forstemann et al, 2000). The degenerate nature of yeast telomeric repeats turned
out to be the feature that can be exploited experimentally to elucidate the
mechanism that leed to telomere length homeostasis. A set of experiments
described later in my thesis exploited the divergence of newly added yeast telomeric
repeats to determine the basis of telomere length homeostasis.
Mammals and all other vertebrates studied to date have telomeres made up
of tandem TTAGGG repeats (Moyzis et al, 1988; Meyne et al, 1990). The longest
telomeres in vertebrates are found in rats and some mouse strains, with more than
150 kb of telomeric sequence (Makarov et al, 1993). Telomeres in human cell lines
vary from 2 to 30 kb, but they are typically around 10 kb in a newborn human, and
gradually shorten with successive cell divisions (Cooke & Smith, 1986; de Lange et
al, 1990; Harley et al, 1990).
Another unique telomeric feature is the presence of a 3’ single-stranded G-
rich overhang (G-tail). The mechanism by which the overhang is generated is still
unclear, but there are protein complexes known that contribute to its formation. The
ribonucleoprotein (RNP) telomerase, which elongates telomeres by templating the
synthesis of new G-strand repeats, is not necessary for G-tail formation, but Mre11-
Rad50-Xrs2 (MRX) complex activity may be responsible for the C-rich strand
resection, as overhangs are reduced in mre11 mutants (Dionne & Wellinger, 1996;

Introduction
20
Larrivee et al, 2004) (properties and mechanism of telomerase are described in
Section 3.2). It has also been found in S. cerevisiae that the length of the overhang
is cell-cycle dependent. Shorter G-tails (12-14 bases) are detectable throughout the
whole cell-cycle, while they are longer than 30 bases in late S-phase (Wellinger et al,
1993; Larrivee et al, 2004). In humans, the 3’ single-stranded protrusion is roughly
30-110 nucleotides long (Chai et al, 2006a). Downregulation of the Mre11-Rad50-
Nbs1 (MRN) complex as well reduces the G-tail length, but interestingly only in
telomerase-positive cells (Chai et al, 2006b). This can be explained by a model in
which the MRN complex is involved in telomerase recruitment, thus not necessarily
having a direct role in C-strand resection. G-tails are suggested to be involved in the
protection of chromosome ends, either by the formation of protective telomeric t-
loops or/and by binding specific single stranded DNA (ss DNA) binding proteins
(mechanisms described in next Section 2.2) (Griffith et al, 1999; Murti & Prescott,
1999).
2.2 Telomere Capping Structure
Even though there are many proteins involved in telomere biology, there is only a
small number of specialized telomere-binding proteins currently known to protect
chromosome ends from degradation and from end-to-end joining events. Yeast
double stranded DNA (dsDNA) telomeric repeats are bound by Rap1
(repressor/activator site binding protein), a major component of telomeric chromatin
(Conrad et al, 1990; Lustig et al, 1990). Rap1 is a large protein of 827 amino acid
residues (Shore & Nasmyth, 1987) that contains two distinct but structurally similar
Myb-like DNA-binding domains between residues 361 and 596 (Henry et al, 1990;
Konig et al, 1996). A structured linker between the domains and a long C-terminal
tail contribute to the binding specificity (Konig et al, 1996). Rap1 also has a BRCT
(BRCA1 C-terminal) domain in the N-terminal region, which is well conserved among
Rap1-related proteins in other organisms.
Through a C-terminal domain Rap1 recruits two proteins, Rif1 and Rif2 (Rap1
interacting factor 1 and 2) (Hardy et al, 1992a; Hardy et al, 1992b; Kyrion et al,
1992; Wotton & Shore, 1997) (Fig. 1), which, together with Rap1, establish a protein
counting mechanism regulating telomere length (properties and mechanism of this

Introduction
21
complex are described in Section 3.5). Rap1 C-terminal domain binds also proteins
mainly involved in heterochromatin formation, Sir3 (silent information regulator 3)
and Sir4 (Moretti et al, 1994; Cockell et al, 1995). Sir2, a nicotinamide adenine
dinucleotide (NAD+)-dependent histone deacetylase, forms a complex with Sir3 and
Sir4 by interacting mainly with Sir4 (Strahl-Bolsinger et al, 1997). Sir3 and Sir4
interact not only with Rap1 but also with the N-termini of histones H3 and H4, which
are hypoacetylated by Sir2 (Hecht et al, 1995; Hoppe et al, 2002). Thus, the Sir2-
Sir3-Sir4 complex spreads over the nucleosomes proximal to the telomeres from the
nucleation sites with Rap1 at the telomeres.
The Cdc13 protein recognizes telomeric ss DNA and binds the G-overhang
with high affinity and sequence specificity (Lin & Zakian, 1996; Nugent et al, 1996).
The Cdc13 DNA-binding domain is thought to comprises a single OB
(oligonucleotide/oligosaccharide binding) fold augmented by an unusually large loop
for DNA recognition (Mitton-Fry et al, 2002), even though computational analysis
predicts the existence of multiple OB-folds (Theobald & Wuttke, 2004). Cdc13
performs its essential function at telomeres by protecting chromosome ends from
degradation, and loss of its function leads to extensive resection of the C-strand and
a Rad9-mediated cell cycle arrest (Weinert & Hartwell, 1993; Garvik et al, 1995;
Lydall & Weinert, 1995; Booth et al, 2001). Two essential Cdc13-associated
proteins, Stn1 and Ten1, also contribute to this capping activity, and the lethality of
the cdc13Δ strain can be rescued if Stn1 protein is artificially delivered to telomeres
by fusion with the DNA binding domain of Cdc13 (Grandin et al, 1997; Grandin et al,
2001; Pennock et al, 2001) (for the function of this complex in telomerase
recruitment and telomere replication see Session 3.3 and 3.4).
In humans, six proteins, TRF1, TRF2, hRap1, TIN2, TPP1, and POT1, form the
so-called shelterin complex, which is a constitutive component of human telomeres
(reviewed in (de Lange, 2005)). The related TRF1 and TRF2 proteins bind double-
stranded telomeric repeats as preformed homodimers (Bianchi et al, 1997),
whereas POT1 binds to the single-stranded telomeric 3′ overhang (Fig. 1). TIN2
connects, through protein-protein interactions, TRF1, TRF2, and TPP1 (Kim et al,
1999; Ye & de Lange, 2004; Ye et al, 2004a). TPP1 binds, in addition, POT1, thus
recruiting POT1 also to the double-stranded part of telomeres (Liu et al, 2004).
Human Rap1, the ortholog of yeast Rap1, associates with telomeres through

Introduction
22
interaction with TRF2 (Li et al, 2000; O'Connor et al, 2004). POT1 has also been
reported to interact directly with TRF2 (Loayza & De Lange, 2003; Yang et al, 2005).
Many examples are available in which perturbation of shelterin components disturbs
telomere length regulation, by interfering with telomere capping, telomerase action,
or possibly telomere heterochromatin states (reviewed in (Hug & Lingner, 2006)).
It is suggested that the G-tail loops back, invades duplex telomeric DNA
forming what is called a t-loop, which provides an elegant architectural solution for
telomere capping. The model for a t-loop came from in vitro observation that
telomeric repeat binding TRF2 can remodel linear telomeric DNA into large duplex
loops (Griffith et al, 1999). Furthermore, electron microscopy analysis of psoralen
cross-linked telomeric DNA purified from human and mouse cells revealed abundant
large loops with a size distribution consistent with their telomeric origin (Griffith et al,
1999). t-loops seem to be conserved feature of telomere structure. These
structures have also been found in the germline DNA (found in the micronucleus) of
the hypotrichous ciliate Oxytricha fallax (Murti & Prescott, 1999), in the flagellate
protozoan Trypanosoma brucei (Munoz-Jordan et al, 2001), and in Pisum sativum
(peas) (Cesare et al, 2003). Experimental evidence for fold-back structures exists in
S. cerevisiae, but the overhang status in these structures is unknown (Strahl-
Bolsinger et al, 1997; de Bruin et al, 2001). Very recently t-loops and homologous
recombination dependent t-circles were observed in yeast Kluyveromyces lactis
telomere mutant strain (Cesare et al, 2007). This strain, however, contains mutant
telomeres that are poorly bound by Rap1, resulting in a telomere uncapping
phenotype and significant elongation of the telomeric DNA. Thus, even though the
elongated telomeres allowed the isolation and examination of native yeast telomeric
DNA by electron microscopy, it is not shown that wild-type K. lactis telomeres form
such structures. t-loops are proposed to be precursors of t-circles formed in this
strain, and might be formed due to uncapping of the telomeres.
Sequestration of the 3’-OH terminus in a t-loop can mask the G-tail from a
variety of detrimental activities, but there is evidence that its binding by single-
stranded DNA (ssDNA) proteins is sufficient for protection. As mentioned above the
micronucleus of hypotrichous ciliates have t-loops, however their macronuclear
telomeres do not (Murti & Prescott, 1999). The macronucleus contains highly
amplified gene-sized DNA molecules at a remarkable copy number of 2 107 per

Introduction
23
nucleus and an average size of just a few kilobases. Rather than containing long
telomeres with t-loops, these DNA molecules are capped by ultrashort telomeres with
termini that are hidden inside a tenaciously bound protein complex (Horvath et al,
1998).
2.3 Organization of the Subtelomeric Region
The subtelomeric regions of S. cerevisiae are composed of a mosaic of moderately
repetitive sequences. Distribution of these repeated elements varies among
different chromosome ends, as well as between different parental strains. Repeat
elements are grouped into two categories, X and Y’ (Chan & Tye, 1983a; Chan & Tye,
1983b). Y’ subtelomeric elements are highly conserved sequences of 5.2 kb or 6.7
kb. They are located at ~half to 2/3 of chromosome ends, embedded within short
tracts of telomeric sequence. In contrast, all chromosome ends have a so-called
“core” X-element, which may be combined with an array of smaller subtelomeric
repeat (STR) A, B, C, and D elements (Pryde et al, 1997). This 473 bp long X-
element contains binding sites for the yeast origin recognition complex (ORC) as well
as the Abf1 transcriptional factor, while the STR elements contain multiple sites of
the vertebrate-like telomeric repeats TTAGGG which are recognized by Tbf1 (TTAGGG
repeat binding factor 1), a protein involved in telomere length regulation (see Section
3.5. and Chapter 3.). Subtelomeric regions are usually low in gene density and late
replicating (Raghuraman et al, 1997).

Introduction
24
In human cells a region made up of TTAGGG repeats, as well as variant
TTGGGG, TGAGGG, and TCAGGG units, is found proximal to telomeric repeat arrays
(Baird et al, 1995; Varley et al, 2002). Further inward, the human subtelomeric
regions are highly repetitive and undergo frequent deletions and translocations
(Brown et al, 1990; Wilkie et al, 1991; Riethman et al, 2004).
3 Telomere Length Regulation
3.1 The End-Replication Problem
The discovery of DNA polymerases and the resulting understanding of DNA
replication raised the question of how linear chromosome-ends are fully replicated.
DNA polymerases synthesize new DNA in the 5’ to 3’ direction, and in general they
extend from a short stretch of RNA (7-10 nucleotides), which primes DNA synthesis.
The RNA oligonucleotides are later degraded and replaced by DNA. However, this
mechanism cannot replace the most distal RNA primer on the 5’ end of the daughter
molecule used to start lagging strand synthesis (Fig. 2). In the subsequent round of
replication, this daughter molecule serves as a template for leading strand synthesis,
which will result in a loss of the G-overhang (Fig. 2d). The loss of telomeric sequence
becomes even more pronounced because the 5’ end of the parental strand must be
further resected to obtain a 3’ ssDNA overhang detected at the chromoseome ends
(Fig. 2e).
In order to counteract the DNA sequence loss due to incomplete replication
and resection, most eukaryotic cells employ an already mentioned specialized
reverse transcriptase called telomerase to extend their telomeres (Greider &
Blackburn, 1985; Lingner et al, 1997b; Nakamura et al, 1997). The mechanism of
telomerase action also offered an explanation for the repetitive nature of telomeric
sequences.

Introduction
25
3.2 Telomerase
The core of the budding yeast reverse transcriptase telomerase comprises the Est2
catalytic protein subunit and the TLC1 RNA component (Singer & Gottschling, 1994;
Lendvay et al, 1996; Lingner et al, 1997b) (Fig. 3a). Even though the core enzyme is
sufficient for telomerase activity in vitro, a screen for an “ever shorter telomeres”
(EST) phenotype in yeast uncovered four genes which exhibit this phenotype if
mutated (Lundblad & Szostak, 1989; Lendvay et al, 1996). The EST phenotype is

Introduction
26
characterized by two properties: continuous telomere shortening and a decline in cell
viability also referred to as senescence, essentially meaning that the genes
discovered are necessary for telomerase activity in vivo at the chromosome ends.
One of them is a mutant allele coding for an already mentioned ssDNA binding
protein Cdc13. The other three genes, EST1, EST2 and EST3, encode three protein
subunits of the S. cerevisiae telomerase.
Deleting any of the telomerase components results in the same senescence
phenotype, being equally defective in an in vivo telomere addition assay that
measures the extent of telomerase-mediated elongation at a de novo synthesized
telomere (Diede & Gottschling, 1999). Consistent with the phenotypic similarities, all
three protein subunits are found and function in the same complex (Hughes et al,
2000; Friedman et al, 2003). Still, telomeres in an est1Δ strain shorten somewhat
faster than in tlc1Δ, suggesting an independent role for Est1 in telomere capping
(Marcand et al, 1999).
TLC1 serves as a scaffold upon which the RNP assembles, with discrete stem-
loop structures that bind the RNP components Est1 and Est2 (Seto et al, 2002;
Dandjinou et al, 2004; Zappulla & Cech, 2004) (Fig. 3b). The association of Est3
with the RNP relies on its physical interaction with Est2 (Friedman et al, 2003). TLC1
contributes to telomerase recruitment to telomeres via the interaction with
conserved DNA end-binding protein Ku (Peterson et al, 2001; Stellwagen et al,
2003) (see Section 3.3). Immunoprecipitation of telomerase from yeast extracts
shows that most or all of the telomerase activity is associated with the complex of
seven Sm-proteins, which binding site is found near the 3' end of TLC1 (Seto et al,
1999). This complex is known to participate in maturation of small nuclear RNAs
(Jones & Guthrie, 1990), suggesting that the TLC1 transcript shares the similar
biogenesis pathway.
TLC1 also templates synthesis of new telomeric repeats. The sequence-
divergence pattern of yeast telomeres is due to the alignment of telomeres in
multiple registers along the 17-nucleotide-long region of TLC1 that serves as the
template for the addition of telomeric repeats (3’ ACACACACCCACACCAC 5’), possibly
combined with abortive synthesis at several points along the template (Prescott &
Blackburn, 1997b; Prescott & Blackburn, 1997a; Forstemann et al, 2000;
Forstemann & Lingner, 2001; Forstemann & Lingner, 2005). Other processes, such

Introduction
27
as nucleolytic degradation or incomplete semi-conservative DNA replication, may
also contribute to the degree of sequence diversity.
While in uni-cellular organisms, like yeast, telomerase is expressed
constitutively, human telomerase is expressed during early development and
remains fully active in specific-germ-line cells, but is undetectable in most somatic
cells except for proliferative cells of renewal tissue (e.g. bone marrow cells, basal
cells of epidermis, proliferative endometrium, and intestinal crypt cells). In all
proliferative cells in which telomerase is not expressed, progressive telomere
shortening is observed, and they ultimately enter a process called replicative
senescence – similarly as yeast cells in which telomerase is non-functional (Harley et
al, 1990).
The human telomerase core consists of the hTERT catalytic subunit and hTR
RNA component (Feng et al, 1995; Nakamura et al, 1997). Unlike the TERT
component, the TR is not well conserved across species – its length and sequence
are both diverged in different phylogenetic groups. However, in all telomerases the
template region is single stranded and connected with a pseudoknot domain
structure. Other yeast telomerase components also have homologues in human
cells. hEST1A, hEST1B, and hEST1C (also known as SMG6, SMG5, and SMG7) have
sequence homology with yeast Est1, and hEST1A and hEST1B both associate with
telomerase in cell extracts (Reichenbach et al, 2003; Snow et al, 2003).
In order to efficiently elongate telomeres, human telomerase has to add
multiple repeats in one cell cycle since it has to compensate for an average
shortening rate of 50-200 nucleotides per round of DNA replication (Harley et al,
1990; Huffman et al, 2000). In vitro human telomerase can elongate a telomeric
primer in a processive manner (Greider, 1991), which is not the case for the yeast
telomerase. Telomerase activity from S. cerevisiae extracts is very inefficient at
translocation: the enzyme remains stably bound with the extended primer used in
the assay after addition of a single telomeric repeat (Prescott & Blackburn, 1997b;
Prescott & Blackburn, 1997a). The functional significance of this in vitro behavior
has been unclear, since yeast telomerase has to compensate for the loss of only ~3
nucleotides per telomere in a single cell cycle. Thus, adding just one repeat would
theoretically be enough to preserve telomere length (Lundblad & Szostak, 1989).
Indeed, the average telomere elongation rate is only a few base pairs per generation

Introduction
28
and this rate decreases progressively with increasing telomere length (Marcand et al,
1999). However, in Chapters 2 and 4, the question of telomere elongation and
telomerase processivity in vivo is addressed in a more precise manner, at single
telomere, revealing telomerase turnover and processivity.
3.3 Cell Cycle Dependent Recruitment of Telomerase
Telomeres are extended by telomerase in late S phase (Marcand et al, 2000), even
though enzyme activity can be detected in extracts prepared during other phases of
the cell cycle (Diede & Gottschling, 1999). Regulation of telomerase recruitment or
activation at chromosome ends can explain this cell-cycle restriction. The only

Introduction
29
known components necessary for telomerase activity in vitro are Est2 and TLC1.
Est1 and Est3 are thought to be required for recruitment and/or activation of
telomerase at telomeres (Cohn & Blackburn, 1995; Lingner et al, 1997a).
Telomerase also interacts with the telomere binding components, Cdc13 and Ku,
although they are not stable constituents of the telomerase RNP in cellular extracts
(Qi & Zakian, 2000; Peterson et al, 2001). As already mentioned, Cdc13 is the
telomeric overhang binding protein. It seems to play two functionally discrete roles
at the chromosome ends. Mutant allele of CDC13, cdc13-2, leads to cellular
senescence, without affecting the end protection activity of Cdc13 (Lendvay et al,
1996; Nugent et al, 1996). However, in cdc13-5 strain, telomeres are increased by
more then 1000 bp, but there is significant amount of ss DNA present at the
telomeres, thus the essential end protection function of Cdc13 is impaired (Chandra
et al, 2001). This dual function can be explained by the Cdc13 property to recruit
both, telomerase and DNA-replication machinery, and coordinate their activity on the
chromosome ends (see Section 3.4). The Ku heterodimer (Yku70/Yku80) binds with
high affinity to ds DNA ends in a sequence independent fashion and is the main
mediator of DNA repair through the non-homologous end joining pathway (Downs &
Jackson, 2004). Paradoxically, despite the fact that telomeres are protected from
such end-to-end fusions, Ku is found on telomeres. Similarly to Cdc13, it exhibits
dual role in telomerase recruitment and telomere end-protection (mechanisms are
decribed later in this section and Section 3.6.1).
How are these proteins involved in cell cycle regulation of telomerase activity?
The presence of Cdc13 at telomeres peaks in S phase, concomitantly with the
appearance of longer G-rich overhangs (Taggart et al, 2002). Cdc13 interacts with
Est1 component of telomerase, suggesting that Cdc13 recruits telomerase to
telomeres in S phase (Evans & Lundblad, 1999; Pennock et al, 2001; Bianchi et al,
2004). Strains bearing either a cdc13-2 or an est1-60 mutant allele, both of which
disrupt the interaction of Cdc13 and telomerase, show a senescing phenotype, with
no effect on telomerase activity in vitro (Nugent et al, 1996; Lingner et al, 1997a;
Pennock et al, 2001). However, telomerase (Est2) is present at the telomeres in
both G1 and S phase; recruitment in G1 is dependent on its interaction with Ku and
interactions with both Ku and Cdc13 contributes to S phase recruitment (Taggart et
al, 2002; Fisher et al, 2004). Ku interacts with telomerase through a defined TLC1-

Introduction
30
loop (Fig. 3b) (Peterson et al, 2001; Stellwagen et al, 2003). Mutants in which the
interaction of Ku and telomerase is perturbed (yku80-135i and tlc-Δ48) have short
but stable telomeres with normal overhang structure (Porter et al, 1996; Boulton &
Jackson, 1998; Peterson et al, 2001; Stellwagen et al, 2003) (for a description
about other roles of Ku in telomere biology, see Section 3.6.1).
Interestingly, in yku70Δ mutants Est1 is absent from telomeres in G1 and its
association with telomeres in late S phase was strongly decreased (Fisher et al,
2004). Since the major contribution of Est1 recruitment is through its interactions
with TLC1 and indirectly with Ku, why is Est1 not present at telomeres in G1 as well?
The answer came from the recent observation that Est1 is actively degraded by
proteosomal activity in G1, thus Est1 is simply absent from the nucleus (Osterhage
et al, 2006). Est3 is also excluded from the telomerase complex during G1 phase.
Stabilization of Est1 in G1 promotes telomerase assembly with Est3, revealing a role
for Est1 in the recruitment of Est3 to the telomerase complex. Still, even if Est1 and
Est3 are assembled with the telomerase complex during G1 due to stabilization of
Est1, it is insufficient to elongate telomeres (Osterhage et al, 2006), suggesting that
Est1 is a necessary but not sufficient activator of telomerase, and that telomerase
may need further activation in late S phase. One possibility is that telomere addition
is tightly coupled to the activity of DNA polymerases α and δ, thus their activity
restricts telomere elongation to S phase (see Section 3.4).
In budding yeast, the regulation of the cell cycle is dominated by the periodic
expression of cyclins and the activity of the cyclin-dependent kinase Cdk1 (Cdc28).
In addition to regulating passage through the cell cycle, Cdk1 activity regulates many
processes in the cell that may not be directly involved in cell-cycle progression by
phosphorylating hundreds of different protein targets (Ubersax et al, 2003). It is
recently observed that the completion of S phase and cyclin-dependent kinase
activity is required for de novo telomere elongation and for the proper formation of G
strand overhangs at both de novo and native telomeres (Frank et al, 2006;
Vodenicharov & Wellinger, 2006). In addition to Cdk1 activity, elongation in G2/M
required the nuclease activity of Mre11, the C-terminal domain of Xrs2 (two
members of the MRX-complex) and Tel1 (Frank et al, 2006) (mechanisms of the
MRX-complex and Tel1 activity are described in Sections 3.6.2 and 3.6.3). One
possibility is that Cdk1 activity is required to activate a nuclease in late S phase to

Introduction
31
generate the long 3′ single-strand overhang or, rather than a nuclease being the
target, another possibility is that a structural protein at the telomere regulates the
access of a nuclease.
Human telomeres are also more accessible in S phase and they seem to lose
part of their protective structure at the end of S phase, becoming recognized as DNA
damage in G2 (Verdun et al, 2005; Jady et al, 2006). However, as in budding yeast,
measurements of telomerase in vitro activity did not reveal cell cycle-dependent
changes (Holt et al, 1997). So either the active complex is not assembled in vivo
throughout cell cycle or recruitment is limited. TPP1 protein, member of the
shelterin complex, interacts with POT1 and is suggested to recruit telomerase (Xin et
al, 2007), but possible cell-cycle regulation of this recruitment is still not addressed.
It associates with the telomerase in a TPP1-OB-fold-dependent manner, providing a
physical link between telomerase and the shelterin complex. Even though the
factors that are responsible to recruit telomerase to telomeres in S phase are not
determined, recent experiments following the localization of hTR by in situ
hybridization indicate S phase-specific assembly of human telomerase and its
subsequent association with telomeres (Jady et al, 2006; Tomlinson et al, 2006).
3.4 Telomere Elongation is Coupled to DNA-replication
Telomerase elongates only the telomeric G-rich strand. Data suggest that the
conventional DNA replication machinery is exploited to synthesize the C-strand. The
first observation pointing to a connection between replication and telomere length
control was that strains bearing temperature sensitive alleles of the catalytic subunit
of DNA polymerase α (POL1) have longer (>2 kb) and more heterogeneous
telomeres when grown at a semi-permissive temperature and exhibit an increase in
the length of the G-strand (Carson & Hartwell, 1985; Adams Martin et al, 2000). By
monitoring de novo telomere synthesis, it was observed that telomere elongation by
telomerase is completely abolished if the lagging-strand polymerase (DNA pol α or
pol δ) is inactive, pointing out that the recruitment of telomerase and the lagging-
strand polymerases are both critical for telomere preservation (Diede & Gottschling,
1999). Shortly after, supporting this model, the replication of a telomere was
suggested to facilitate its extension by telomerase (Marcand et al, 2000).

Introduction
32
It is unclear how these two processes are coordinated, but several lines of
evidence point to the role of the Cdc13 complex and its associated partners, Stn1
and Ten1. Many mutations in CDC13, STN1, or TEN1 result in the same telomeric
phenotype as observed for DNA pol α mutants (Grandin et al, 1997; Chandra et al,
2001; Grandin et al, 2001). More direct evidence of their possible functional
interaction is that both Cdc13 and Stn1 physically interact with subunits of the DNA
pol α/primase complex (Qi & Zakian, 2000; Grossi et al, 2004). In vivo disruption of
the Cdc13–Pol1 interaction results in elongated telomeres, as well as disruption of
the interaction between Stn1 and Pol12, another subunit of DNA pol α (Qi & Zakian,
2000; Grossi et al, 2004). This led to a model in which the Cdc13-Stn1-Ten1
complex would recruit both telomerase and the lagging-strand DNA-replication
machinery, ensuring coordinated synthesis of the G- and C-rich telomeric strands.
Interestingly, the Cdc13-complex shares structural features and shows limited
sequence homology with the heterotrimeric replication protein A (RPA), the major
ssDNA–binding complex in eukaryotic cells essential for multiple processes in DNA
metabolism including DNA replication, recombination, and DNA repair pathways
(reviewed in (Wold, 1997)). Since Cdc13, Stn1, and Ten1 are as well ssDNA-binding
proteins, but with specificity for telomeric DNA substrates, the question was raised
whether the Cdc13-complex could be viewed as a telomere-specific RPA (Gao et al,
2007). Still, RPA seems to have functions at telomeres which can not be
complemented by the presence of the Cdc13-complex (Schramke et al, 2004).
Chromatin immunoprecipitation (ChIP) experiments showed that the RPA component
Rfa2 associates with telomeres in S-phase (Schramke et al, 2004). In the same
work, the authors noticed strong telomere shortening in strains bearing a defective
rfa2Δ40 allele, which expresses Rfa2 lacking the N-terminus domain. In these
strains, telomere recruitment of Est1 was significantly reduced in S-phase, provoking
two possible explanations. Either RPA directly plays a role in Est1 recruitment or this
can be viewed as another example that telomerase action must be coordinated with
replication. Since rfa2Δ40 has no noticeable accumulation of single-stranded
overhangs, and the telomere shortening phenotype can be completely rescued by
expressing a Cdc13-Est1 fusion protein in this strain, the first model is the preferred
one.

Introduction
33
Nevertheless, there is no direct proof that a telomere has to replicate in order
to be elongated by telomerase. Very recent data from the Shore lab suggests that
this is actually the case, and propose that short telomeres are replicated earlier and
elongated more by telomerase then the longer ones (Bianchi & Shore, 2007a). The
authors take advantage of a system which generates a short telomere after the
excision, via recombination, of internal Rap1 binding sites (Marcand et al, 1999).
Internally inserted Rap1 sites cause in cis shortening of the terminal telomeric tract
since they are counted as a measure of telomere length (see Section 3.5). After
their excision this telomere is left with a short TG-tract which is elongated by
telomerase. Comparing the binding of DNA polymerase ε (Pol2) at short telomeres
and at normal-length telomeres showed that shortened telomeres replicate early in S
phase, unlike normal-length telomeres, due to the early firing of origins of DNA
replication in subtelomeric regions (Bianchi & Shore, 2007a). This correlated with
earlier loading of Cdc13 and Est1, as well as a slightly increased average elongation
rate of the short telomeres. These results connect replication timing with telomerase
activation, revealing an epigenetic effect of telomere length on the activity of nearby
subtelomeric replication origins.
3.5 A “Protein Counting Model” of Telomere Length Control
In yeast cells (as in all organisms studied to date) telomere length is kept within a
well-defined range, implying the existence of mechanisms that are able to sense the
length of the TG-tract and regulate the elongation or shortening rate. The main
“sensor” proteins in yeast consist of the Rap1 protein, which directly binds to the
telomeric double stranded region, and its interacting partners, Rif1 and Rif2. Rap1
is an essential protein with multiple roles in the cell, and was originally identified as
a repressor or activator of transcription (Shore & Nasmyth, 1987). Rap1 binds to the
promoter regions of about 5% of yeast genes and participates in the activation of
about 37% of all RNA polymerase II initiation event (Shore et al, 1987; Lieb et al,
2001). Thus it is no surprise that its essential role in cell viability is connected with
its role in gene expression. However, Rap1 is as well tightly involved in the
regulation of telomere length (Conrad et al, 1990; Lustig et al, 1990; Sussel &
Shore, 1991; Marcand et al, 1997b) and telomere position effect (TPE) (Kyrion et al,

Introduction
34
1993; Moretti et al, 1994). In vitro studies revealed that one Rap1 molecule binds
on average every 18 bp of double-stranded telomeric DNA (Longtine et al, 1989;
Gilson et al, 1993) and it was shown to be localized on chromosome ends in vivo as
well (Conrad et al, 1990; Klein et al, 1992). The role of Rap1 in telomere length
regulation clarified with the isolation of rap1t mutants (such as rap1-17) which
display very long telomeres (up to 3 kb) and carry a mutation leading to C-terminal
deletions of the protein (Kyrion et al, 1992). Such proteins can still bind telomeres,
but can not interact with Rif1 nor Rif2 (Hardy et al, 1992b). Indeed, deletion strains
of RIF1 have elongated telomeres (up to 1 kb) (Hardy et al, 1992b), while rif2Δ
strains have a less strong telomere elongation phenotype, with the effects being
striking in a double mutant, having telomeres indistinguishable from the rap1t
mutant-phenotype unable to bind any of these factors (Wotton & Shore, 1997)

Introduction
35
These data suggest a model in which Rif1 and Rif2 are negative regulators of
telomere length, with a partially overlapping function, when recruited to telomeres by
the Rap1 C-terminal domain. Consistent with this model, it was observed that
tethering the C-terminal domain adjacent to a telomeric tract causes shortening of
the telomeric tract (Marcand et al, 1997a). A similar shortening of telomeres was
induced if just Rif1 or Rif2 were artificially tethered internally to a telomeric tract
(Levy & Blackburn, 2004). These observations form the basis of the so-called
“counting model” for the regulation of telomere length, which proposes that the
number of bound Rap1-Rif1 and -Rif2 molecules at telomeres dictates telomere
length (Fig. 4). When telomeres reach a certain length, a growing number of bound
Rap1 complexes will produce a signal that blocks telomerase action, and telomeres
will shorten progressively due to the end-replication problem and telomere resection,
until the number of bound complexes will drop below the threshold value. The loss
of Rap1 with bound Rifs will switch telomeres to an active state in which they will
become accessible to telomerase activity (Fig. 4).
Research by Marcand et al. (1999) demonstrated that repression of
telomerase activity does not occur suddenly, but is a result of gradual inhibition upon
telomere elongation (Marcand et al, 1999). By contrast the rate of telomere
shortening is independent of telomere length, and is constant at the rate of about 3
bp/generation. Two possibilities can be envisioned to explain the observation that
the shorter telomeres are more rapidly elongated – either telomerase adds longer
repeats on the short telomeres, or short telomeres are more frequently elongated, or
both.
There is data available revealing the existence of Rap1-independent
mechanism for telomere length setting. Tel1, a member of the PI-3 (phosphatidyl
inositol-3) kinase-like kinase family, seems to be a mediator of the Rap1-dependent
counting mechanism (see Section 3.6.3) (Ray & Runge, 1999). It was observed that
in tel1∆ yeast cells, in contrast to wild type, additional Rap1-complexes added
adjacent to telomeric repeats are not counted as a signal for telomere length (Ray &
Runge, 1999). Furthermore, human telomeric repeats, which are not bound by
Rap1, inserted internally to native yeast telomeric tracts are counted in this deletion
background, whereas they are not counted in wild type cells (Brevet et al, 2003).
Surprisingly, disruption of the Rap1-counting mechanism by deletion of TEL1

Introduction
36
revealed the presence of an additional cryptic mechanism of telomere length
regulation, possibly depending on two essential insulator proteins present in the
yeast subtelomeric region: Tbf1 and Reb1 (Bilaud et al, 1996; Fourel et al, 1999;
Koering et al, 2000). Tbf1, a protein related to the mammalian telomere repeat-
binding factors, TRF1 and TRF2, binds directly to human-like telomeric repeats and
negatively regulates telomere elongation (Brigati et al, 1993; Alexander & Zakian,
2003). Tbf1 and Reb1 both cause telomere shortening when targeted to yeast
telomeres in the tel1∆ background, proportionally to the number of bound molecules
(Berthiau et al, 2006). Furthermore, telomeres are clustered in 4-8 foci, which
associate with the nuclear envelope (Gotta et al, 1996). In the absence of Tel1, Tbf1
and Reb1 act as anti-anchoring factors, preventing telomere localization at the
nuclear envelope, suggesting a link between telomere anchoring to the nuclear
periphery and telomere length regulation (Hediger et al, 2006).
Apart from its function in the protein counting mechanism, Rap1 plays a role
in telomere end protection. Rap1 loss from telomeres, by the means of induced
degradation of a degron tagged Rap1, causes frequent fusions between telomeres of
normal length (Pardo & Marcand, 2005). These fusions are dependent on factors
required for nonhomologous end joining (NHEJ) showing that Rap1 is essential to
suppress NHEJ between telomeres.
Can the shelterin complex on human telomeres control telomere elongation
in a similar manner as described for the yeast “counting” mechanism? Longer
telomeres load more shelterin and this may provide a length-sensing mechanism
(Smogorzewska et al, 2000). As well, the association of POT1 with shelterin in the
dsDNA telomere region may affect POT1 loading on the single-stranded telomeric
DNA where it may act as an inhibitor of telomerase activity (Loayza & De Lange,
2003; Liu et al, 2004; Ye et al, 2004b; Kelleher et al, 2005). Alternatively or in
addition, shelterin may inhibit telomerase by promoting the formation of the t-loop
structure, in which the 3′ overhang is inaccessible, being tucked into the double-
stranded part of the telomere (Griffith et al, 1999). Telomere length is also
negatively regulated by histone methyltransferases in the mouse, which may be
required to assemble telomeric heterochromatin, suggesting another possible
mechanism in regulating telomere accessibility (Garcia-Cao et al, 2004).

Introduction
37
3.6 DNA-damage Response Proteins in Telomere Length Regulation
The protein counting mechanism describes how to sense telomere length, but it is
unclear what are downstream transmitters of this signal; furthermore what are the
molecular mechanisms of telomerase regulation? As I already mentioned, Tel1 (the
orthologue of human ATM) is a downstream regulator of the Rap1-dependent
mechanism of telomere length regulation, which together with Mec1 (yeast
orthologue of human ATR), belongs to PI-3 kinase-like kinase family. Even though by
definition, telomeres prevent chromosome ends from activating the DNA damage
response, a surprising aspect of telomere maintenance is that many DNA repair and
checkpoint proteins, including the Ku heterodimer, MRX complex, Tel1 and Mec1,
associate with telomeres and contribute to telomere length homeostasis. (d'Adda di
Fagagna et al, 2004).
3.6.1 The Ku Heterodimer
Ku is a heterodimer made up of two subunits of approximately 70 and 80 kDa,
known as Ku70 and Ku80, respectively. It is of central importance to DNA repair in
eukaryotes, though, in addition, it has roles in a number of other fundamental
cellular processes such as telomere maintenance, transcription and apoptosis. Ku
binds with high affinity to DSBs with 3' or 5' overhangs, as well as blunt ones, and is
a key player in DNA repair through the non-homologous end joining pathway (NHEJ)
in both yeast and mammals. An early consequence of Ku – DSB binding might be to
maintain the DNA ends in proximity in some regulated manner prior to DNA repair to
prevent the ends drifting apart and to protect them from unwanted or excessive
nucleolytic attack. Since ends need to be further processed to be repaired and
ligated, Ku also allows recruitment of repair factors and manipulation of the broken
ends. This can be explained by an asymmetry of the Ku ring structure, which allows
the DNA to be held, while potentially allowing access to the DNA on the opposite side
(reviewed in (Downs & Jackson, 2004)).
At the telomeres, Ku may bind at the transition from double strand telomeric
repeats to single-stranded overhang, with very little variation throughout the cell-
cycle (Gravel et al, 1998; Fisher et al, 2004). However, Ku is, like Rap1, required for
establishment of telomeric heterochromatin, suggesting that it also resides on the
duplex portion of the telomere (Boulton & Jackson, 1998; Laroche et al, 1998;

Introduction
38
Nugent et al, 1998). It affects silencing of the telomere proximal genes through its
interaction with the Sir4 protein (Tsukamoto et al, 1997; Mishra & Shore, 1999; Roy
et al, 2004).
Ku’s role at the chromosome ends is expected to differ from that on DSBs,
since telomeres are protected from end-to-end fusions. The functions of Ku at
telomeres and DSBs can be separated in Ku mutants that affect one function but
not the other (Bertuch & Lundblad, 2003). Ku’s dual activities are also observed at
telomeres where it performs two functions: first, as already mentioned, it facilitates
telomerase recruitment (Section 3.3) (Porter et al, 1996; Boulton & Jackson, 1998;
Peterson et al, 2001; Stellwagen et al, 2003) and second, it prevents excessive
resection of the C-strand, thereby contributing to protection of chromosome termini
(Gravel et al, 1998; Polotnianka et al, 1998). In accordance with the suggested
roles of Ku, its deletion mutants have very short telomeres with long overhangs
(Gravel et al, 1998; Polotnianka et al, 1998). If these strains are shifted to the
restrictive temperature (37°C), Exo1 nuclease–dependent resection of the C-strand
can extend to subtelomeric regions (Maringele & Lydall, 2002; Bertuch & Lundblad,
2004). On the other hand, mutants in which the interaction of Ku and telomerase is
perturbed (yku80-135i and tlc-Δ48) have short but stable telomeres, with normal
overhang structure (Peterson et al, 2001; Stellwagen et al, 2003). Ku has also been
shown to help localize telomeres to the nuclear periphery (Hediger et al, 2002) and it
is required for the establishment of transcriptional silent telomeric chromatin
(Boulton & Jackson, 1998; Nugent et al, 1998). Furthermore, it has been proposed
that Ku regulates the replication timing of telomeres, since sub-telomeric origins of
replication are activated much earlier in mutants lacking Ku function (Cosgrove et al,
2002). This observation might be an indirect consequence of the short telomere
phenotype in kuΔ strains, as short telomeres induces earlier telomeric replication, or
their misslocalization from nuclear periphery (Bianchi & Shore, 2007a).
The Ku heterodimer is associated with the shelterin complex and mammalian
telomeres (Hsu et al, 1999; Song et al, 2000; d'Adda di Fagagna et al, 2001;
O'Connor et al, 2004). Since Ku is essential in human cells, its role in telomere
biology is more difficult to uncover. However, even partial reduction of Ku86 results
in a telomere deprotection phenotype that included loss of telomeric DNA and
increased frequency of chromosome end fusions (Jaco et al, 2004; Myung et al,

Introduction
39
2004). In mouse cells deletion of Ku is not lethal, though it increases the frequency
of telomeric fusions, without obvious loss of telomeric sequence (Bailey et al, 1999;
Hsu et al, 2000; Samper et al, 2000; d'Adda di Fagagna et al, 2001). Also, in mouse
cells Ku70 represses exchanges between sister telomeres - a form of homologous
recombination implicated in the alternative lengthening of telomeres (ALT) pathway
(Celli et al, 2006). Significantly, consequences of Ku loss on mammalian telomeres
are far less serious than perturbation of the shelterin complex, suggesting that
telomere protection by Ku is redundant with other mechanisms.
3.6.2 The MRX Complex
MRX (Mre11-Rad50-Xrs2) is an evolutionary conserved protein complex which acts
in the DSB response, in break repair by either homologous recombination (HR),
primarily between sister chromatids, or nonhomologous end-joining (NHEJ) (reviewed
in (Haber, 1998; Stracker et al, 2004)). The three MRX polypeptides have different
biochemical activities. Mre11 acts as an ATP-stimulated nuclease that is both a
single stranded endonuclease and a 3’ to 5’ exonuclease, even though MRX defect
in vivo results in impaired 5’ to 3’ resection. Rad50 demonstrates ATP-dependent
binding to DNA and contains a zinc-coordinating motif (Rad50 hook) that is critical
for MRX-dependent DNA repair and telomere maintenance (Raymond & Kleckner,
1993; Wiltzius et al, 2005). Xrs2 is important for targeting Mre11 and Rad50 to
DNA ends, and it also plays a critical role in recruiting Tel1 to damaged DNA (Nakada
et al, 2003; Trujillo et al, 2003). In mammalian cells, analogous interactions occur
and NBS1, the orthologue of Xrs2, recruits ATM to DSBs (Falck et al, 2005).
In yeast, deletion of any MRX gene results in shortened, but stable, telomeres
and slow growth (Kironmai & Muniyappa, 1997; Ritchie & Petes, 2000). There are
at least three potential roles for MRX in telomere maintenance that would explain
the short telomeres of mrxΔ mutants. Epistasis analysis demonstrated that the MRX
complex is required for telomerase-mediated telomere elongation and a possible
role of MRX is creating the substrate for telomerase (Nugent et al, 1998).
Telomerase requires a 3′ substrate and mrxΔ mutants are partially defective in
generating the 3′ overhang at telomeres, causing proportional shortening of the
overhang throughout cell cycle (Diede & Gottschling, 2001; Larrivee et al, 2004). A
second role for MRX is activation of the Tel1 kinase, which may, through its kinase

Introduction
40
activity, enhance the activity of telomerase at telomeres (Tsukamoto et al, 2001).
Recent data confirmed that recruitment of Est2 and Est1 is severely reduced in
mre11Δ and tel1Δ cells, association of Est2 being affected both in G1 and S phase
(Goudsouzian et al, 2006). However, MRX and Tel1 do not have identical telomere
functions, as MRX (but not Tel1) is required for WT-length G tails. This difference is
also reflected in the levels of telomere-bound Cdc13 in the two strains: tel1Δ
telomeres have WT levels of Cdc13 binding, whereas Cdc13 binding to mre11Δ
telomeres is 65% of WT levels (Goudsouzian et al, 2006).
A third role for the MRX complex is the capping of telomeres to protect them
from nuclease or other DNA repair activities that respond to DSBs elsewhere in the
genome and that, if active at telomeres, might shorten telomere length.
Interestingly, a capping role for MRX is supported by recent experiments in
Drosophila, an organism that does not use telomerase to maintain or cap telomeres.
In fruit flies, mutation of MRE11 or RAD50 causes an increase in telomere fusions.
This capping function occurs through a pathway involving ATM and the
heterochromatic proteins HP1 and HOAP (Bi et al, 2004; Ciapponi et al, 2004; Bi et
al, 2005). This range of MRX functions may also be explained by the hypothesis that
MRX is needed to load different regulator proteins onto the telomeres.
Mammalian Mre11 complex (MRN) is recruited to telomeres through shelterin
(primarily TRF2) interactions (Zhu et al, 2000; Cano et al, 2002). Mre11 and Rad50
are present at telomeres throughout the cell cycle, but Nbs1 is only detectable in S
phase (Zhu et al, 2000; Loayza & De Lange, 2003). Because the MRN complex is
essential in mammals, it has been difficult to establish its function on telomeres,
however a mutant allele of Nbs1 induces telomere loss in some settings, and Nbs1
contributes to telomerase-mediated telomere elongation (Ranganathan et al, 2001;
Bai & Murnane, 2003).
3.6.3 Tel1 and Mec1 Kinases
Tel1 (along with the NHEJ proteins) is among the first proteins to localize to a DSB in
order to initiate DNA repair, but is subsequently replaced by Mec1 (and the HR
proteins); this transition is dependent on the resection of one strand of the DSB
(reviewed in (Garber et al, 2005)). While its human homologue, ATM, is involved in
sensing DSBs generated by DNA damaging agents like ionizating radiation, the yeast

Introduction
41
checkpoint response mainly depends on Mec1. Tel1 has a more pronounced role in
telomere preserving processes. Indeed, TEL1 was one of the first genes identified
for its role in telomere length maintenance and the first PI-3 kinase-like kinase
recognized to participate in telomere length maintenance (Lustig & Petes, 1986).
Yeast cells lacking Tel1 harbor short but stable telomeres, while deletion of MEC1
has a mild telomere shortening effect (Greenwell et al, 1995; Ritchie et al, 1999).
The kinase functions of Tel1 and Mec1 appear to be partially redundant in telomere-
length maintenance since simultaneous deletion of both TEL1 and MEC1 results in
progressive loss of telomeric sequences and finally senescence (Ritchie et al, 1999).
Epistasis analysis showed that Tel1, but not Mec1, functions in the same pathway of
telomere length regulation as MRX (Ritchie & Petes, 2000).
What is the reason for short telomeres in tel1Δ strains? One possibility could
be that Tel1 regulates telomerase activity itself. It was found that telomerase activity
in vitro is not affected by lack of either Tel1 or Mec1. In addition, the senescent
phonotype of tel1Δ mec1Δ double mutants is bypassed by targeting a Cdc13-Est2 or
a Cdc13-Est1 protein fusion to chromosome ends, suggesting a role in telomerase
recruitment, not directly in its activation (Chan et al, 2001; Tsukamoto et al, 2001).
This hypothesis is supported by findings that strong perturbation of telomeric
structure in rif1Δ rif2Δ cells is as well able to prevent senescence in a tel1Δ mec1Δ
double mutant.
Recently data has been accumulating to support a role for Tel1 in telomerase
recruitment. As mentioned above, the Zakian lab confirmed that recruitment of Est2
and Est1 is severely reduced in mre11Δ and tel1Δ cells, while mec1Δ have just a
mild effect on the Est2 presence on the telomeres (Goudsouzian et al, 2006). This
observation is consistent with Mec1 having a less pronounced impact on telomere
length. Tel1 presumably modulates telomere length by phosphorylating a target
protein. Since Tel1 seems to be a downstream transducer of the Rap1-counting
mechanism, it can be envisioned that this phosphorylation can differentiate short
from long telomeres. What its relevant downstream substrate is for the protein
counting mechanism is still an open question. The single stranded binding protein
Cdc13p has recently been revealed as a substrate of these kinases (Tseng et al,
2006). Although the number of Cdc13 molecules bound on the telomere is
independent of telomere length (Sabourin et al, 2007), changing the Cdc13-Stn1-

Introduction
42
Ten1 conformation by phosphorylating Cdc13 would be consistent with a role of Tel1
in regulating the telomerase recruitment step by modifying the structure of the end-
complex. In addition, it has been recently shown that short telomeres are more
avidly bound by Est1, Est2 and Tel1, more directly defining Tel1 as a molecular
marker of short telomeres (Bianchi & Shore, 2007b; Hector et al, 2007; Sabourin et
al, 2007). Binding of Tel1 at short telomeres is independent of its kinase activity
and requires active MRX complex (Hector et al, 2007; Sabourin et al, 2007).
At mammalian telomeres, ATM activates a major telomere-damage signal if
telomeres get unprotected (Takai et al, 2003; Silverman et al, 2004). In the
absence of ATM, ATR (ataxia telangiectasia-Rad3 related) can respond to
dysfunctional telomeres (Herbig et al, 2004). As well, an accelerated rate of
telomere shortening was suggested for patients suffering ataxia telangiectasia,
which is caused by mutations in ATM (Metcalfe et al, 1996). A recent report shows
that ATM can function together with the MRN complex to control TRF1 binding to
telomeres (Wu et al, 2007). Previously it was established that TRF1 can be
phosphorylated by ATM in response to DNA damage (Kishi et al, 2001). Upon
phosphorylation, TRF1 dissociates from telomeres, leading to increased access of
telomerase to the ends of telomeres (Wu et al, 2007). This model proposes that the
MRN- and ATM-mediated control of TRF1 is evolutionarily conserved between yeast
and human.
3.7 Other Regulators of Telomere Length
Knowing the importance of telomere length regulation for cellular viability, it is not
surprising that there are many protein families involved in this process. Very
important, and still unmentioned, helicase family members that can unwind double-
stranded nucleic acids or remove proteins from ssDNA are regulators of telomere
length (Krejci et al, 2003; Veaute et al, 2003). One of them, the PIF1 helicase family
is conserved from yeast to humans (Bessler et al, 2001). In yeast cells, Pif1 inhibits
telomerase-mediated telomere lengthening and de novo telomere addition at double-
strand breaks (Schulz & Zakian, 1994; Zhou et al, 2000; Mangahas et al, 2001;
Myung et al, 2001). Pif1 helicase activity limits telomerase action both in vivo and in
vitro by displacing active telomerase from DNA ends (Boule et al, 2005; Boule &

Introduction
43
Zakian, 2007). In accordance with this proposed function, overexpression of Pif1
leads to modest telomere shortening while Pif1 deletion results in long telomeres
(Schulz & Zakian, 1994; Zhou et al, 2000). The human Pif1 as well inhibits
telomerase activity, and ectopic expression of wild-type Pif1 causes telomere
shortening in certain cell lines (Zhang et al, 2006).
There are hints about a role of Pif1 in telomere replication as well. It interacts
genetically with the Dna2 helicase/nuclease, which is required for Okazaki fragment
processing (Budd et al, 2006). It has been proposed that Dna2 and Pif1 may be
required for processing the last Okazaki fragment at the telomere. Interestingly, the
other yeast member of this helicase family, Rrm3, appears to promote the
progression of the replication fork through telomeres (Ivessa et al, 2002; Ivessa et al,
2003).
Additionally to the factors that are already intensively studied, there are many
others involved in telomere length homeostasis. Two genome-wide screens
monitoring telomere length in deletion strains were performed in S. cerevisiae to
identify genes and pathways that regulate telomere length (Askree et al, 2004;
Gatbonton et al, 2006). Over 250 genes were detected to affect telomere length,
many of them very diverse in their functions. Broadly, they can be separated into
telomerase-dependent and telomerase-independent pathways, including genes that
participate in DNA and RNA metabolism, chromatin modification, sister chromatid
cohesion, and vacuolar protein sorting.
3.8 Recombination-Dependent Maintenance of Telomeres
As described above, in most eukaryotes telomere length is preserved by telomerase.
When any of the genes essential for telomerase activity is absent, telomeres
progressively shorten, and eventually cells fail to divide and arrest. However, from
these senescing cells, survivors can arise and continue to divide. This was first
discovered in budding yeast, revealing an alternative pathway for telomere
maintenance that does not require telomerase (Lundblad & Blackburn, 1993). The
rare survivors recovered from an est1Δ strain displayed dramatic changes at their
chromosomal termini, due to global amplification of both telomeric or sub-telomeric
repeat sequences (named Type II and Type I survivors, respectively) (Teng & Zakian,

Introduction
44
1999). Type I survivors are characterized by tandem duplications of the sub-
telomeric Y’ elements and by short tracts of telomeric repeats, while Type II have
very long and heterogeneous stretches of TG repeats (Teng et al, 2000). Both types
of rearrangements arise as a result of recombination, as the appearance of survivors
depends on Rad52, a protein responsible for the majority of homologous
recombination events in yeast. Still, there is a difference in recombination pathways,
since Type I survivors depend on Rad51 (the yeast homologue of the bacterial strand
exchange protein RecA), and Type II survivors arise by a Rad50 dependent process
(Teng et al, 2000; Chen et al, 2001).
Even though type I survivors initially represent 90% of survivors, they have a
slower growth rate, so they are overgrown in a senescant population by type II. As
well, in a different mutant background certain type of survivors are prevealing. For
example type II recombination is mediated by the cell cycle checkpoint proteins Tel1
and Mec1, and telomere-telomere recombination is regulated by telomere binding
protein Cdc13 and the Ku complex (Tsai et al, 2002).
Some immortalized mammalian cell lines and tumors are as well able to
maintain their telomeres over many population doublings in the absence of
telomerase. These cells have extremely heterogeneous telomere length which is
preserved by a telomerase-independent mechanism called the ALT (alternative
lengthening of telomeres)-pathway (Bryan et al, 1995).
4 Thesis Research Aims
In the beginning of my thesis work, it was thought that short telomeres were
preferentially elongated by telomerase over longer ones. Telomere elongation was
shown to be restricted to a few base pairs per generation on average, and this rate
decreased gradually with increasing telomere length (Marcand et al, 1999).
However, it was not known how this is achieved or what is really happening at the
level of single telomeres. At least two possibilities could have been envisioned:
either telomerase adds longer repeats on the short telomeres, or short telomeres are
more frequently elongated, or both. Furthermore it was not known whether or not
every telomere is elongated in each cell cycle, or just a limited subset. To answer
these questions, an in vivo system was set up in the laboratory in which the

Introduction
45
elongation of single telomeres could be assayed at nucleotide resolution within one
cell cycle (Chapter 2). This experimental setup we named STEX for “single telomere
extension” assay.
In the continuation of my work presented in Chapter 3, I tried to decipher the
molecular mechanism involved in the preferential elongation of short telomeres; or
in other words, what proteins are necessary to distinguish short from longer wild-type
length telomeres?
And finally in Chapter 4, the question of telomerase repeat addition
processivity in vivo is addressed, showing once more that there are differences
between the mechanism of elongation of very short and “normal” length telomeres.


Chapter 2
Telomere Length Homeostasis Is Achieved via a Switch
between Telomerase-Extendible and -Nonextendible States
Article published in Cell, vol. 117, pp. 323-335, April 30th, 2004


Mechanism of Telomere Length Homeostasis
49
Short telomeres are preferentially elongated over longer ones. Telomere elongation
per average telomere is restricted to a few base pairs per generation in S. cerevisiae,
and this rate decreased gradually with increasing telomere length (Marcand et al,
1999). The following chapter answers how this regulation is achieved: are more
nucleotides added upon short telomeres than on longer ones, or are just the shortest
telomeres elongated in each cell cycle? M.T. Teixeira set up the “single telomere
extension” (STEX) assay in the laboratory at the time of my arrival and joining the
project. STEX enabled us to follow the extension events at a single chromosome end
in vivo, in one cell cycle, at nucleotide resolution.
I contributed to Fig. 3, Fig. 4, and Fig. 5 by performing a number of STEX
assays for the analysis of telomere VR sequence divergence events in wild-type and
rif1Δ cells, in parallel with M.T. Teixeira. I also did the construction of the strains and
STEX experiments assaying the elongation events at the native telomere IL in wild-
type cells, and at telomere VR in rif2Δ mutants (Fig. 1 c, g; Fig. 4 c; Fig. 5 b).


Cell, Vol. 117, 323–335, April 30, 2004, Copyright 2004 by Cell Press
Telomere Length Homeostasis Is Achievedvia a Switch between Telomerase-Extendible and -Nonextendible States
acts telomere shortening (reviewed in Kelleher et al.[2002]). It extends the 3� end of chromosomes by reversetranscribing in an iterative fashion the template regionof its tightly associated telomerase RNA moiety. Thecomplementary DNA strand is presumably synthesized
M. Teresa Teixeira,1,3 Milica Arneric,1
Peter Sperisen,2 and Joachim Lingner1,*1Swiss Institute for Experimental Cancer Researchand National Center of Competence in Research
“Frontiers in Genetics”by the conventional DNA polymerases � and � (Diede2 Swiss Institute for Bioinformaticsand Gottschling, 1999; Ray et al., 2002). TelomeraseCH-1066 Epalinges/s Lausanneexpression is required for unlimited proliferation of uni-Switzerlandcellular organisms such as yeast or protozoa as well asfor immortal cells in multicellular organisms such asgerm cells, some stem cells, and many cancer cells. InSummaryhumans, a decline in telomere length correlates with theonset of age-dependent mortality (Cawthon et al., 2003).Telomerase counteracts telomere erosion that stemsShort telomeres and reduced levels of telomerase havefrom incomplete chromosome end replication andbeen observed in individuals that carry a defective tel-nucleolytic processing. A precise understanding ofomerase RNA gene or a defective allele of the tel-telomere length homeostasis has been hampered byomerase-associated dyskerin protein (Mitchell et al.,the lack of assays that delineate the nonuniform telo-1999; Vulliamy et al., 2001). Both genetic disorders givemere extension events of single chromosome mole-rise to the bone marrow failure syndromes dyskeratosiscules. Here, we measure telomere elongation at nu-congenita and aplastic anemia, which may be causedcleotide resolution in Saccharomyces cerevisiae. Theby telomere exhaustion and a reduced replicative poten-number of nucleotides added to a telomere in a singletial of stem cells. On the other hand, telomere lengthcell cycle varies between a few to more than 100 nucle-stabilization and upregulation of telomerase expressionotides and is independent of telomere length. Tel-has fatal consequences during malignant transforma-omerase does not act on every telomere in each celltion of cells that are normally telomerase negative (Kimcycle, however. Instead, it exhibits an increasing prefer-et al., 1994).ence for telomeres as their lengths decline. Deletion of
In cells that express telomerase, the length of thethe telomeric proteins Rif1 or Rif2 gives rise to longerduplex telomeric repeat array is kept within a species-telomeres by increasing the frequency of elongationand cell type-specific narrow range. In humans, the aver-events. Thus, by taking a molecular snapshot of aage telomere length varies between 5 and 15 kb,single round of telomere replication, we demonstratewhereas in the yeast S. cerevisiae, telomere length isthat telomere length homeostasis is achieved via aaround 300 nucleotides. Telomere length homeostasisswitch between telomerase-extendible and -nonex-is the result of a balance between telomere shorteningtendible states.and telomere lengthening activities (see Smogorzewskaand de Lange [2004] for a recent review). As discussedIntroductionabove, a primary cause of telomere shortening is incom-plete DNA end replication. Nucleolytic processing ofThe ends of eukaryotic chromosomes, known as telo-telomere ends is presumably important for the genera-meres, are essential protein-DNA complexes that pro-tion of 3� overhangs, which are present on both chromo-tect chromosome ends from fusion and degradationsomal ends (Huffman et al., 2000; Makarov et al., 1997;
(see Ferreira et al. [2004] for a recent review). The DNAMcElligott and Wellinger, 1997; Wellinger et al., 1996;
component of telomeres typically comprises tandemWright et al., 1997). The telomere shortening rate in the
repeats of simple sequences that are rich in guanosine absence of telomerase is insensitive to telomere lengthresidues in the strand containing the 3� end, which pro- and corresponds to 50–200 nucleotides per round oftrudes to form a single strand overhang. Therefore, the DNA replication in human cells and three nucleotidesrecessed 5� end-containing strand, which is replicated per generation in S. cerevisiae (Harley et al., 1990; Huff-by the leading strand synthesis machinery, cannot func- man et al., 2000; Lundblad and Szostak, 1989; Marcandtion as a template for the synthesis of the 3� overhang et al., 1999). Telomere shortening can also occur in a(Lingner et al., 1995). Hence, in the absence of special more stochastic fashion through so-called telomerictelomere maintenance mechanisms, linear chromo- rapid deletions (TRD), which involve intrachromatid re-somes shorten progressively with every round of DNA combination events (Bucholc et al., 2001; Li and Lus-replication. In addition, nucleolytic processing of tel- tig, 1996).omere ends may contribute to shortening (Jacob et al., Telomere lengthening requires telomerase in most2003; Wellinger et al., 1996). Telomere length limits the species. However, under rare circumstances, recombi-replicative potential of many normal human somatic nation-mediated pathways may maintain telomeres incells (Bodnar et al., 1998). the absence of telomerase (Bryan et al., 1995; Dunham
The cellular reverse transcriptase telomerase counter- et al., 2000; Lundblad and Blackburn, 1993; Teng andZakian, 1999). This is observed at low frequency in yeastcells that escape the lethality caused by telomerase loss*Correspondence: [email protected]
3 Present address: Institut Pasteur, 75724 Paris Cedex 15, France. and in some in vitro-immortalized and cancer-derived

Cell324
human cell lines. The contribution of recombination to to a specific telomere induced its shortening in cis (An-telomere length maintenance in wt cells is unknown. celin et al., 2002). Evidence for regulation of telomerase
In S. cerevisiae, telomerase extends telomeres in late activity in cis had been provided even earlier in telomereS phase (Marcand et al., 2000), and this is thought to healing experiments in which newly seeded telomeresbe coupled to semiconservative DNA replication (Diede were smaller than the endogenous telomeres (Barnettand Gottschling, 1999). Numerous gene products have et al., 1993). Furthermore, in mice, crosses betweenbeen implicated in telomere maintenance. Mutations in strains with short and long telomeres result in a preferen-some of these genes abolish telomerase activity, leading tial elongation of short telomeres (Zhu et al., 1998; Hem-to progressive telomere shortening. Telomere replica- ann et al., 2001; Samper et al., 2001). Similarly, expres-tion factors that are essential in vivo include telomerase sion of limiting amounts of telomerase activity in humansubunits and proteins that bind to the telomeric 3� over- fibroblasts leads to preferential elongation of short telo-hang, where they mediate the access and activation of meres (Ouellette et al., 2000).telomerase (reviewed by Evans and Lundblad [2000]). At least two not mutually exclusive models could pro-ATM- and ATR-related protein kinases are also required vide a mechanistic basis for the protein-counting modelfor telomerase activity in vivo in fission and budding of telomere length control and the increased activity ofyeast (Naito et al., 1998; Ritchie et al., 1999), but their telomerase on shorter telomeres. First, the elongationcritical substrates are yet to be identified. Mutations in efficiency of telomerase, i.e., the number of nucleotidesother telomeric factors perturb but do not abolish the added to an individual telomere per elongation event,end replication machinery, leading to an altered steady- could be regulated as a function of telomere length.state telomere length. For example, some double- Thus, all telomeres would be available for telomerase-stranded telomeric DNA binding proteins are negative mediated extension, but the telomere structure wouldregulators of telomere length. Budding yeast Rap1p is regulate the catalytic activity of telomerase in a length-a major component of telomeric chromatin (reviewed by dependent manner, perhaps by influencing the pro-Shore [1997]) that binds the telomeric repeats with high cessivity or turnover of the enzyme. Second, the produc-affinity and negatively affects telomere length (Lustig et tive association of telomerase with telomere 3� endsal., 1990; Conrad et al., 1990). Through a C-terminal could be regulated by length-dependent changes in tel-domain, Rap1p recruits two proteins, Rif1p and Rif2p, omeric chromatin structure. A long telomere would havewhich contribute to telomere length control (Hardy et al., a lower probability to be in a telomerase-extendible state1992; Wotton and Shore, 1997). Targeting of additional than a short telomere. In this model, a telomere couldcopies of the Rap1p C terminus to a telomere induces shorten for several rounds of DNA replication withouttelomere shortening to an extent that is roughly propor- being elongated by telomerase, before its chromatintional to the number of targeted molecules (Marcand et structure would switch and become competent for tel-al., 1997; Ray and Runge, 1999b). This observation led to omerase-mediated elongation. Thus, the association,the “protein counting model” of telomere length control rather than the activity, of telomerase would be the regu-(Marcand et al., 1997), in which telomerase-mediated lated element.telomere extension is regulated by the number of telo- To distinguish between these two models, we devisedmere bound Rap1p molecules. Additional support for a a system to measure elongation of single telomeres incounting mechanism comes from work with telomerase vivo at nucleotide resolution. Using this system, we de-RNA template mutants in S. cerevisiae and the related termined the frequency and elongation efficiency of telo-yeast Kluveromyces lactis (Chan et al., 2001; McEachern meres as a function of their length. We find that telomereand Blackburn, 1995). Such mutations reduced the num- elongation is a stochastic process that does not occurber of telomeric binding sites for Rap1p and con- at all telomeres in every cell cycle. Short telomeres havecomitantly induced massive telomere elongation. The a higher probability of being extended than long telo-protein-counting model was also supported by mea-
meres. A variable number of telomeric repeats can besurement of the reelongation kinetics of an artificially
added in one round of replication to a single chromo-shortened telomere (Marcand et al., 1999). These experi-
some end, but the number of added nucleotides doesments indicated that the rate of telomere elongation isnot correlate with telomere length, unless the telomereinversely proportional to telomere length.is almost completely eroded. The frequency of tel-Double-stranded telomere binding proteins in fissionomerase-mediated extension increases in rif1-� andyeast and vertebrates also function as negative regula-rif2-� mutants. Taken together, our results provide di-tors of telomere length, similarly to S. cerevisiae Rap1p.rect evidence for a model in which telomeres switch inDisruption of the fission yeast taz1 gene gives rise to aa length-dependent manner between at least two struc-10-fold increase in telomere length (Cooper et al., 1997),tural states that dictate the extendibility by telomerase.and depletion of the orthologous human TRF1 from telo-
meres also leads to substantial telomere lengtheningResults(van Steensel and de Lange, 1997). Overexpression of
TRF1 induces telomere shortening without affectingAn Assay for Measuring Single Telomeretelomerase activity in vitro (Smogorzewska et al., 2000).Extension at Nucleotide Resolution In VivoSimilarly, the TRF1-interacting proteins TIN2 and tankyr-To study the mechanisms that control telomere accessase 1 regulate telomere length without affecting tel-and extension efficiency, we developed a system toomerase activity in vitro (Kim et al., 1999; Smith and demeasure single telomere elongation events in vivo atLange, 2000). These experiments suggest that tel-nucleotide resolution in S. cerevisiae (Figure 1). A tel-omerase is regulated by the telomeric substrate in cis.
Consistent with this notion, artificial tethering of TRF1 omerase-negative strain was created by deleting the

Mechanism of Telomere Length Homeostasis325
Figure 1. An Experimental System for Measuring Telomere Extension In Vivo at Nucleotide Resolution
(A) Introduction of telomerase activity in a controlled fashion to a telomerase-negative cell by functional complementation. Clonal telomerase-negative cells (recipients) are mated with telomerase-positive cells (donors) of the opposite mating type. Upon karyogamy, the newly formedzygotes enter their first S phase, and telomerase from the donor cell can elongate shortened telomeres inherited from the telomerase-negative recipients.(B) Specific amplification of telomere VR of the recipient strain by telomere PCR (Forstemann et al., 2000; see Experimental Procedures). DNAis extracted from the zygotes, and DNA ends are tailed with terminal transferase and dCTP (green). Telomere VR of the recipient contains aunique sequence in the subtelomeric region (purple). This allows specific amplification by PCR with a primer that anneals to the sequencetag and a primer that is complementary to the oligo-dC tail. Telomerase-mediated telomere DNA extension is indicated by the blue line.(C) Sequence alignment of telomere VR from a recipient clonal population before mating (P; upper alignment) and 3 hr after mating (M; loweralignment). Telomere extension events are detected by sequence divergence that arises among sister telomeres due to the action of yeasttelomerase, which adds imperfect telomeric repeats (blue letters). Most sister telomeres that stem from the telomerase-negative parent areidentical in sequence, because they are replicated faithfully by the semiconservative DNA replication machinery. They only differ in lengthdue to irregular shortening. One out of nine parental telomeres analyzed (P11) diverged in sequence in the telomerase-negative clonalpopulation. This is due to a telomerase-independent telomere rearrangement (green). N, ambiguous bases.

Cell326
telomerase RNA gene TLC1. An isolated tlc1-� colony After approximately 3.5 hr, the diploid cell number haddoubled, indicating that most diploids had undergonewas grown for approximately 30 generations. Active tel-
omerase was provided by mating the tlc1-� strain (re- mitosis and completed one cell cycle (Figure 2A). Thecombined analyses indicated that zygote formation andferred to as “recipient”) with a telomerase-positive strain
(referred to as “donor”) of the opposite mating type the subsequent S phase occurred with high synchronyand that, 3 hr after initial mixing, most zygotes had com-(Figure 1A). Thus, telomeres that had shortened in the
telomerase-negative parent become reextended in the pleted their first round of DNA replication.Telomere VR was amplified by telomere PCR (Figurezygote due to the presence of complementing tel-
omerase. Telomere elongation was detected by cloning 2D) and cloned from the parental tlc1-� strain and fromthe tlc1-�/TLC1 diploid 3, 5, and 7 hr postmating. Telo-and sequencing telomeres that originated from the re-
cipient. To this end, we used as recipient a strain harbor- mere sequence analysis (Figure 2E) indicated that duringthe time course experiment, the fraction of telomereing the ADE2 gene in the subtelomeric region of chromo-
some VR, which served as a sequence tag. Telomere extension events increased. After 3 hr, which corre-sponds to a time point when most zygotes had com-VR was amplified by telomere PCR as described (Fors-
temann et al., 2000, and Figure 1B). In this method, pleted one S phase, nine out of 25 (36%) sequencedtelomeres had been elongated. After 5 hr, the ratio ofgenomic DNA is isolated and tailed in vitro with dCTP
and terminal transferase. Telomere VR is amplified spe- extended to nonextended telomeres was roughly main-tained (nine out of 22 � 40%), whereas after 7 hr, orcifically with a primer that anneals with the ADE2 gene
at VR and a primer that is complementary to the oligo approximately three population doublings, 10 out of 11telomeres were elongated. This experiment demon-dC tail. PCR products were resolved on agarose gels,
cloned, and sequenced (see Experimental Procedures). strates that not all telomeres become extended by tel-omerase in every cell cycle, even if they are considerablyYeast telomerase adds imperfect 5�-(TG)0–6TGGGTG
TG(G)-3� repeats (Forstemann and Lingner, 2001). Thus, shorter than wt.telomere elongation is detectable via the divergence oftelomeric sequences in the telomere-distal region rela- Telomerase-Independent Lengtheningtive to the telomeric DNA synthesized by semiconserva- of Chromosome Endstive DNA replication, which does not vary during clonal Inspection of telomere sequences in tlc1-� parents (Fig-expansion. Thus, even if two telomere extension events ure 2E) revealed that, even in the absence of telomerase,occurred at the same nucleotide, they can be distin- one out of ten telomere 3� ends diverged in sequenceguished due to tract divergence. A representative exam- from their sisters. To quantify these telomerase-inde-ple is given in Figure 1C, in which telomere VR was pendent elongation events, we cloned and sequencedamplified, cloned, sequenced, and aligned in the tlc1-� the telomeres from haploid (Figures 3A–3C) and diploidparental strain after propagation for approximately 30 (Figures 3D and 3E) telomerase-negative strains thatgenerations without telomerase and from the tlc1-�/ lacked either TLC1 or EST1. EST1 encodes a telom-TLC1 zygote 3 hr after mating. The frequency of telo- erase-associated protein that is essential for telomerasemeres showing sequence divergence in the mating activity in vivo, but it is not required for in vitro activity.mixture was significantly increased compared to the Est1p has been implicated in recruitment and activationparental telomeres, allowing us to examine telomerase- of telomerase at chromosomal ends (Evans and Lund-dependent telomere extensions at nucleotide reso- blad, 1999; Pennock et al., 2001; Taggart et al., 2002).lution. Sequence analysis of VR telomeres revealed that tel-
omerase-independent elongation events occurred atsimilar rates in all four telomerase-negative strains.Telomerase Does Not Extend Every TelomereThirty generations of clonal expansion resulted in 6.6%in Every Cell Cycle(13 out of 197 at VR) telomeres showing this type ofTo correlate telomere elongation kinetics with zygoteevent. Involved telomeres were variable in size from 41formation and subsequent cell divisions, time courseto 226 nucleotides, and elongations varied from three toexperiments were performed (Figure 2). Mating was fol-179 nucleotides. Thus, these telomerase-independentlowed by plating on selective media to assess the pres-extension events occurred prior to achievement of criti-ence of auxotrophic markers (Figure 2A) by cytologicalcal telomere shortening. We suspect that the divergenceobservation (Figure 2B) and by fluorocytometric (FACS)is due to recombination events involving TG repeats ofanalysis of DNA content (Figure 2C). Mating efficiencydifferent chromosome ends.was quantified and optimized to nearly 100% by collect-
ing the telomerase-negative strain together with a 10-fold excess of the telomerase-positive strain on filter Variability of Telomere Extension Length
To analyze the relationship between telomere length andpapers (see Experimental Procedures). Microscopicanalysis and assessment of auxotrophic markers indi- telomerase activity in vivo, we quantified the telomere
elongation events that occurred after one round of DNAcated that almost all recipient cells had initiated cell-to-cell contact and zygote formation after approximately replication following telomerase reintroduction in tlc1-�/
TLC1 zygotes and in est1-�/EST1 zygotes (Figure 4).2 hr. At this time point, most cells were in G1, as deter-mined by FACS (Figure 2C). After 3 hr, approximately In order to achieve an extensive coverage of telomere
lengths, we performed the mating experiments with sev-9% of the cells contained a 4C DNA content (Figure 2C).Because of the 10-fold excess of wt over tlc1-� haploids, eral independent clones for each strain. Figures 4A and
4B show the results of independent experiments inthis was consistent with the notion that, at this timepoint, most zygotes had completed a single S phase. which the tagged telomere VR had a different length
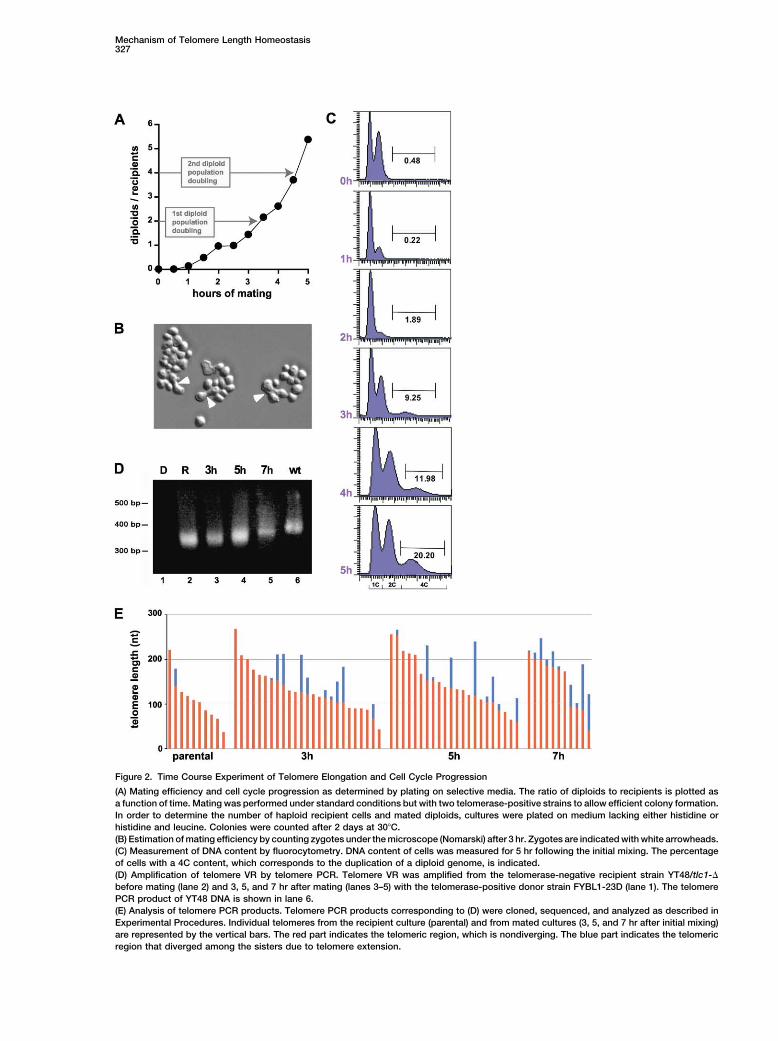
Mechanism of Telomere Length Homeostasis327
Figure 2. Time Course Experiment of Telomere Elongation and Cell Cycle Progression
(A) Mating efficiency and cell cycle progression as determined by plating on selective media. The ratio of diploids to recipients is plotted asa function of time. Mating was performed under standard conditions but with two telomerase-positive strains to allow efficient colony formation.In order to determine the number of haploid recipient cells and mated diploids, cultures were plated on medium lacking either histidine orhistidine and leucine. Colonies were counted after 2 days at 30�C.(B) Estimation of mating efficiency by counting zygotes under the microscope (Nomarski) after 3 hr. Zygotes are indicated with white arrowheads.(C) Measurement of DNA content by fluorocytometry. DNA content of cells was measured for 5 hr following the initial mixing. The percentageof cells with a 4C content, which corresponds to the duplication of a diploid genome, is indicated.(D) Amplification of telomere VR by telomere PCR. Telomere VR was amplified from the telomerase-negative recipient strain YT48/tlc1-�
before mating (lane 2) and 3, 5, and 7 hr after mating (lanes 3–5) with the telomerase-positive donor strain FYBL1-23D (lane 1). The telomerePCR product of YT48 DNA is shown in lane 6.(E) Analysis of telomere PCR products. Telomere PCR products corresponding to (D) were cloned, sequenced, and analyzed as described inExperimental Procedures. Individual telomeres from the recipient culture (parental) and from mated cultures (3, 5, and 7 hr after initial mixing)are represented by the vertical bars. The red part indicates the telomeric region, which is nondiverging. The blue part indicates the telomericregion that diverged among the sisters due to telomere extension.

Cell328
Figure 3. Telomerase-Independent TelomereExtension
Telomere VR (A, B, and D–G) and telomere1L (C) were PCR amplified and analyzed asin Figure 2 from clonal populations after 30cell doublings from YT48/tlc1-� cells (A),YT48/est1-� cells (B and C), diploid tlc1-�/tlc1-� (C), diploid est1-�/est1-� cells (D),est1-� rif1-� cells (F), and est1-� rif2-�
cells (G).
when reextended by the complementing telomerase in in est1-�/EST1 zygotes. The average extension lengthin tlc1-�/TLC1 cells was 45 nucleotides, with an inter-the zygote. To analyze elongation events at telomeres
of wt length, we analyzed a strain in which telomeres quartile range from 14 to 66 nucleotides (Table 1). Inest1-�/EST1 cells, the average extension length was 86were overelongated before telomerase removal and
mating (Figure 4B, left). To do this, we transformed the nucleotides, with an interquartile range from 14 to 80nucleotides. The longer average seen in est1-�/EST1recipient est1-� strain with plasmid pVL1120 expressing
a fusion protein between the DNA binding domain of zygotes stems from very long extension events of ex-tremely short telomeres (Figure 4B and Table 1). As theCdc13p and Est1p (Est1-DBDCdc13), a construct that com-
plements est1-� and leads to telomere overelongation mean telomeric repeat length in S. cerevisiae is 12–13nucleotides (Forstemann and Lingner, 2001), telomere(Evans and Lundblad, 1999). Upon culturing the est1-�/
pVL1120 strain for 50 generations under selection for extension length corresponded in the majority of casesto several telomeric repeats. Thus, the in vivo propertiesthe plasmid, the strain was plated on nonselective YPD
medium. Cells that had lost pVL1120 were telomerase of yeast telomerase differ markedly from its biochemicalproperties in vitro, where it performs only one elongationnegative and were mated as above with a telomerase-
positive wt strain for 3 hr. cycle and prevents further elongation by stably associat-ing with the telomeric primer (Cohn and Blackburn, 1995;After 3 hr of mating, most of the resulting zygotes
were in G2 following their first round of DNA replication. Prescott and Blackburn, 1997).Surprisingly, the number of added nucleotides is notStrikingly, the telomere extensions were very heteroge-
neous in size, ranging from four to 187 nucleotides in correlated significantly with telomere length (Figure 4D)(tlc1-�/TLC1: r � �0.29, ppearson � 0.15; � � �0.27,tlc1-�/TLC1 zygotes and from four to 417 nucleotides

Mechanism of Telomere Length Homeostasis329
Figure 4. Frequency and Length of TelomereExtension as a Function of Telomere Size
(A) Telomere elongation events in tlc1-�/TLC1 zygotes. YT48/tlc1-� cells were matedwith FYBL1-23D cells for 3 hr. The taggedtelomere at VR (tel VR::ADE2) was amplified,cloned, and sequenced. Two independentexperiments (M2111 and M2510) are de-picted on the left and right sides. Labeling isas in Figure 2.(B) Telomere VR elongation events in est1-�/EST1 zygotes. YT57/est1-� (left side, M0302_2)and YT48/est1-� (right side, M2811) cells weremated with FYBL1-23D as in (A).(C) Elongation events at the natural telomereIL in est1-�/EST1 zygotes. YT48/est1-� cellswere mated with MAY01 for 3 hr. MAY01 lacksthe subtelomeric region at telomere IL. Twoindependent experiments (M2511 and M0711)are depicted on the left and right sides.(D) Length of telomere extension as functionof telomere size. The sizes of diverging re-gions obtained in the experiments describedin (A), (B), and (C) are plotted as a functionof the nondiverging sequence, which corre-sponds to the length of the original telomere.Green dots indicate events from (A), blue dotsevents from (B), and pink dots from (C). Pinkcircles indicate events that could not be at-tributed without ambiguity to telomerase-mediated extension due to the high recombi-nation background for short telomeres atchromosome 1L in the parent (see Figure 3C).(E) Frequency of telomere extension as func-tion of telomere size. Sequences obtainedfrom (A), (B), and (C) were ordered accordingto nondiverging telomere size and pooled intosubgroups containing ten telomeres each.The frequency of elongation in each sub-group was calculated and plotted as a func-tion of telomere length. Data points were fit-ted to the following logistic regression model:[(frequency of elongation) � 0 1 (meanlength) �]. The p1 value for tlc1-�/TLC1
zygotes is 0.00447; for est1-�/EST1 (at the tagged telomere VR), 0.00018; and for est1-�/EST1 (at the natural telomere IL), 0.00592. The curvecorresponding to est1-�/EST1 (telomere 1L) is dashed for telomeres below 100 nt, because, in this region, the events could not be attributedwith high confidence to telomerase-mediated extension, due to the high recombination background for short telomeres at chromosome 1Lin the parent (see Figure 3C).
pspearman � 0.19; est1-�/EST1: r � �0.37, ppearson � 0.062; meres according to the length of their nondiverging telo-meric sequence and formed subgroups containing ten� � �0.37, pspearman � 0.059). However, extremely long
extensions of several hundred nucleotides were ob- telomeres each. The average telomere size of each sub-group was calculated, and the frequency of elongationserved with very short telomeres of less than 100 nucleo-
tides, indicating loss of normal telomerase control. in each subgroup was plotted as a function of telomerelength (Figure 4E). Although the data scattered substan-tially, the frequency of elongation correlated signifi-Telomerase Acts Preferentially
on Short Telomeres cantly with telomere size (tlc1-�/TLC1: r � �0.75,ppearson � 0.0125, � � �0.80, pspearman � 0.0082; est1-�/To determine whether the frequency of telomere elonga-
tion was regulated by telomere size, we sorted telo- EST1: r � �0.70, ppearson � 0.0057, � � �0.59, pspearman �
Table 1. Comparison of Telomere Elongation Lengths in tlc1-�/TLC1, est1-�/EST1, est1-�/EST1 rif1-�/rif1-�, and est1-�/EST1 rif2-�/rif2-�
Zygotes
Average (Interquartile Range)
Original telomere length �100 nt 200 nt alltlc1-�/TLC1 49 (22–67) 19 (13–22) 45 (14–66)est1-�/EST1 212 (79–378) 44 (15–70) 86 (14–80)est1-�/EST1 (tel IL) NA ND 67 (21–93)est1-�/EST1 rif1-�/rif1-� NA 59 (31–78) NAest1-�/EST1 rif2-�/rif2-� 36 (20–46) 56 (19–98) 68 (20–116)

Cell330
0.029). Furthermore, a significant departure from ran- tions (Wotton and Shore, 1997). To better understand themechanism of action of Rif1p, we mated a presenescentdomness was apparent when applying the �2 test forrif1-� est1-� strain with a rif1-� EST1 strain and ana-contingency tables (ptlc1 � 0.029; pest1 � 0.00071). Thelyzed telomere elongation as described above (Figuredata were fit to a generalized linear model using logistic5). After 30 generations of clonal expansion, telomeresregression (Figure 4E). At wt telomere length (300 nucle-in the parental est1-� rif1-� strain had been extendedotides), the frequency of telomere elongation per roundby telomerase-independent elongation at a frequencyof DNA replication was 0.078 for tlc1-�/TLC1 zygotesof 23% (25 out of 110) (Figure 3F). Most of these eventsand 0.067 for est1-�/EST1 zygotes. This frequency in-corresponded to telomeres that had shortened belowcreased steadily to 0.46 for tlc1-�/TLC1 zygotes and120 nucleotides. In the rif1-�/rif1-� est1-�/EST1 zygote,0.42 for est1-�/EST1 zygotes as telomeres shortenedtelomere elongation frequency increased significantlyto 100 nucleotides. These data indicate that a typical(p � 0.0084) by approximately 2-fold when comparedtelomere undergoes several rounds of semiconservativeto the est1-�/EST1 zygote, considering the telomeresDNA replications without being elongated by telom-that were longer than 120 nucleotides (Figure 5D). Thiserase. Therefore, telomeres switch between telomerase-indicates that Rif1p promotes formation of a telomereextendible and -nonextendible states. The equilibriumstructure that prevents the productive association ofbetween the two states is regulated by telomere length.telomerase. For telomeres that had shortened below120 nucleotides, the high recombination backgroundTelomere Length-Dependent Controldid not allow identification of telomerase-dependentof Extendibility at a Natural Telomereelongation events without ambiguity. However, compar-Natural yeast telomeres contain in their subtelomericison of the frequency and length of extension in theregion repetitive elements that are referred to as X andpresence and absence of telomerase (compare FiguresY�, whereas the tagged telomere VR lacks these ele-5A to Figure 3F) suggested that telomerase-mediatedments. To test if the above telomere elongation analysisextension events contributed substantially to the ob-of tagged VR could be extrapolated to natural telomeres,served elongation of short telomeres.we analyzed telomere 1L, which contains an X element
We also analyzed the extent of telomere elongationin its subtelomeric region. For this telomere, we had(Figure 5C) as a function of telomere length. As seenpreviously identified a unique subtelomeric sequencein est1-�/EST1 zygotes, the telomere extensions wereallowing specific amplification by telomere PCR (Teixe-heterogeneous in size, ranging from ten to 261 nucleo-ira et al., 2002). In order to amplify 1L from the recipienttides for the telomeres that had not shortened belowstrain only, telomere IL of the donor strain was truncated120 nucleotides. The average extension length was 83and replaced with an artificial telomere (tel IL::URA3; seenucleotides, with an interquartile range from 32 to 96.Supplemental Data at http://www.cell.com/cgi/content/In the absence of Rif1p, the frequency of elongation (r �full/117/3/323/DC1).�0.68, ppearson � 6.04e-6, � � �0.62, pspearman � 8.17e �Parental est1-� IL telomeres were sequenced after 305) but not the extent of elongation (r � �0.40, ppearson �generations of clonal expansion to determine telom-0.09, � � �0.23, pspearman � 0.37) correlated significantlyerase-independent events (Figure 3C). Divergence waswith telomere length (for telomeres that were longer thanobserved in four out of 39 sequenced telomeres. All four120 nucleotides). In telomeres with a nearly physiologi-diverging chromosome ends corresponded to telo-cal length of above 200 nucleotides, the rif1-� zygotesmeres that had shortened to near or below 100 nucleo-displayed a slightly longer extension length than RIF1tides (Figure 3C), suggesting that telomerase-indepen-controls (59 versus 44 nucleotides; see Table 1).
dent recombination may be regulated at this naturalThe effects of a rif2 deletion were analyzed as for
telomere in a length-dependent manner. These est1-�rif1 (Figure 5B and Table 1). The frequency of telomere
cells were mated with EST1 tel IL::URA3 donor cells, elongation increased similarly for rif2-� by approxi-and intact telomere IL was amplified as above (Figure mately 2-fold when compared to RIF2 cells, whereas4C). As for telomere VR, the frequency of telomerase- the extension length was not affected (Table 1). As fordependent elongation events increased with telomere rif1-�, the frequency (r � �0.83, ppearson � 0.04, � �shortening (r � �0.75, ppearson � 0.020, � � �0.82, �0.75, pspearman � 0.0083) but not the extent of telomerepspearman � 2.75e � 05) (Figure 4E). Furthermore, a signifi- elongation (r � �0.28, ppearson � 0.234, � � �0.26,cant departure from randomness was apparent when pspearman � 0.275) correlated significantly with telomereapplying the �2 test for contingency tables (pteI1L � 0.036). length. However, in contrast to rif1-� cells, the recombi-The extent of elongation, as for telomere VR, did not nation background in rif2-� cells was, with four out ofconvincingly correlate with telomere length (r � �0.45, 63 analyzed telomeres, low and comparable to RIF2ppearson � 0.058; � � �0.52, pspearman � 0.029) for telomeres cells (Figure 3G), allowing a more straightforward as-longer than 120 nucleotides (Figure 4D). sessment of telomerase-mediated events also for short
telomeres. In conclusion, the data indicate that in rif1-�Rif1p and Rif2p Regulate the Equilibrium and rif2-� cells, telomeres are longer because they be-between Extendible and Nonextendible come more frequently extended by telomerase. Thus,Telomeric States both Rif1p and Rif2p control the productive associationThe Rif1 and Rif2 proteins are recruited to telomeres of the telomere substrate with telomerase.via the Rap1p C-terminal domain and are thought tomediate the Rap1-counting mechanism of telomere Discussionlength control (Marcand et al., 1997). Simultaneous dele-tion of RIF1 and RIF2 has a synergistic effect on telomere Telomere length homeostasis is fundamental for the
maintenance and propagation of stable chromosomes.length, indicating that they have distinct regulatory func-

Mechanism of Telomere Length Homeostasis331
Figure 5. Frequency and Length of TelomereExtension as a Function of Telomere Size inrif1-� and rif2-� Cells
(A) Telomere elongation events in est1-�/EST1 rif1-�/rif1-� zygotes. YT56/est1-� cellswere grown and mated with YT55 cells for 3hr, and the tagged telomere at VR was ana-lyzed. Two independent experiments (M2012_1and M1604) are depicted on the left and rightsides. Labeling is as in Figure 2.(B) Same as (A) with est1-�/EST1 rif2-�/rif2-�zygotes.(C) Length of telomere extension as a functionof telomere size. The sizes of diverging re-gions obtained in the experiments describedin (A) (red) and (B) (orange) are plotted as afunction of the nondiverging sequence. Theextension events in est1-�/EST1 zygotes(blue) from Figure 4D are also indicated forcomparison.(D) Frequency of telomere extension as afunction of telomere size. Sequences from (A)and (B) were analyzed as in Figure 4D. Theplot corresponding to est1-�/EST1 zygotesis reported from Figure 4D (blue). The p1
value for est1-�/EST1 rif1-�/rif1-� zygotes is2.45�5 and for est1-�/EST1 rif2-�/rif2-� is0.0137. The curve corresponding to est1-�/EST1 rif1-�/rif1-� zygotes is dashed for telo-meres below 100 nt, because, in this region,the events could not be attributed withoutambiguity to telomerase-mediated exten-sion, due to the high recombination back-ground for short telomeres in the parent (seeFigure 3F).
Loss of a single telomere is a potentially lethal event (Figure 6). The nearly constant rate of telomere shorten-ing due to incomplete end replication, nucleolytic pro-eliciting chromosome end fusions and missegregation.cessing, and telomere rapid deletion events is balancedPrevious work has indicated that telomerase activityby telomerase- and recombination-mediated telomereis regulated in cis at individual telomeres through thelengthening. A short telomere has a high probability ofnumber of double-stranded telomere binding proteinsadopting the telomerase-extendible structure (Figure 6,(Ancelin et al., 2002; Marcand et al., 1997; Ray andlower). Upon elongation (Figure 6, upper), the equilib-Runge, 1999a). This implies that telomeric chromatinrium shifts to the nonextendible state, and the telo-controls either the enzymatic properties of telomerase,mere’s likelihood of being in the extendible state de-the accessibility of telomeric ends, or both (Marcand etcreases with length. The average telomere lengthal., 1999). To directly address and distinguish betweencorresponds to the size at which the probability of exten-these possibilities, we developed a method to measuresion multiplied by the average extension length equalstelomere elongation events at single molecules. Ourthe shortening rate. In est1�/EST1 zygotes, the probabil-analysis demonstrates for the first time that telomeraseity of extension at the wt telomere length of 300 nucleo-does not act on every telomere in every cell cycle. Wetides (0.067) multiplied by the average extension lengthdemonstrate the existence of at least two distinct tel-(44 nucleotides for telomeres 200 nucleotides; Table
omeric states: one that allows the productive associa-1) matches the measured shortening rate of 2.95 � 0.2
tion with telomerase and one that prohibits telomerase-nucleotides per generation (Marcand et al., 1999).
mediated telomere extension. We show that the frequency Our analysis of telomere sequence divergence in tel-of telomere extension increases steadily as a function of omerase-negative cells (Figure 3) also indicates that re-telomere length, from roughly 6%–8% at 300 nucleotide- combination is contributing to telomere maintenance.long telomeres to 42%–46% at 100 nucleotides. Thus, The contribution of this pathway to telomere mainte-the equilibrium between the two states is regulated by nance was detected in the telomerase-negative cellstelomere length. The existence of different telomeric after clonal expansion for approximately 30 generations.states that prevent or permit telomerase access has At this time point, approximately 6% of the telomeresbeen suggested, but direct experimental evidence for had undergone telomerase-independent elongation,their existence was lacking. corresponding to an estimated frequency of less than
0.3% per generation (Lea and Coulson, 1949). Thus,we estimate that the contribution of recombination to
Mechanism of Telomere Length Homeostasis telomere maintenance is 20- to 30-fold lower than thatThe telomere length dependent regulation of extendibil- of telomerase. The frequency of telomere recombination
strongly increased in the rif1-� background for shortity is sufficient to explain telomere length homeostasis

Cell332
Figure 6. Model for Telomere Length Homeostasis
Telomeres exist in an equilibrium between two states: one that is extendible by telomerase (left side) and one that is nonextendible (rightside). Long telomeres have a higher probability to be in the nonextendible state (upper part). As telomere length declines, the equilibriumbetween the two states shifts continuously to the extendible state (lower part). The average telomere length corresponds to the size at whichthe ratio of extendible over nonextendible states (probability of extension) multiplied by the average extension length equals the shorteningrate. Rif1p and Rif2p, which associate with telomere bound Rap1p, promote the nonextendible state, shifting the equilibrium to the right. Theymay promote conformational changes of the telomeric 3� end that prevent telomerase access or activation.
telomeres (Figure 3F). Interestingly, Rif1p was previously polymerase. Upon reaching the natural length of approx-imately 250 nucleotides, the poly(A) binding proteinidentified as a factor that inhibits type II recombination
(Teng et al., 2000), a pathway that involves homologous loses its stimulating activity, and poly(A) polymeraseswitches from a processive to a distributive mode ofrecombination, which can be observed in a small frac-
tion of yeast that escaped the lethality caused by tel- extension (Wahle, 1991).Yeast telomerase associates in vitro with telomericomerase loss. However, deletion of RIF2 did not en-
hance telomerase-independent elongation in our study, primers in a stable manner and is able to add only asingle telomeric repeat (Cohn and Blackburn, 1995;while it favored the occurrence of type II survivors even
more strongly than rif1-� (Teng et al., 2000). Prescott and Blackburn, 1997). Thus, our in vivo analysisindicates that additional factors must exist that eitherThe number of nucleotides added by telomerase var-
ied substantially between individual telomerase exten- promote a processive mode of elongation or that enablerapid turnover of the enzyme. The described assay sys-sion events and was not tightly regulated by telomere
length unless they were shorter than 100 nucleotides. tem provides a tool to identify processivity factors. Apotential candidate might be the Est1p protein, whichHowever, telomeres of this size are unlikely to occur in
wt yeast cells. The average length of the ADE2-tagged has been proposed to promote enzyme turnover or pro-cessivity (Singh and Lue, 2003). However, in our analy-VR is 291 nucleotides, with a standard deviation of �31
nucleotides (Forstemann et al., 2000). Thus, for the telo- sis, est1-� cells were indistinguishable from tlc1-� cells,being unable to add even a single telomeric repeat (Fig-mere size range of telomerase-positive cells, the extent
of the elongation by telomerase does not correlate with ure 4). We also determined telomere addition in est1-60cells, which express mutant Est1p with a single Lys →the length of the original telomere. We conclude that
under normal conditions, telomere length does not sig- Glu mutational change at residue 444 (data not shown).This mutation disrupts an electrostatic interaction withnificantly control telomerase processivity or turnover.
On the other hand, dramatic telomere shortening may Cdc13p, causing an ever-shorter telomeres phenotype(Pennock et al., 2001). In our analysis, this allele was, asremove telomere binding factors that block excessive
elongation by telomerase, or a distinct pathway for the est1-�, completely inactive in telomere addition. Thus,Est1p cannot act solely by promoting processivity orhealing of severely shortened telomeres may become
activated. turnover but is also involved in the initial elongation cycleconsistent with its function in telomerase recruitmentThe mechanism for length homeostasis described
here has interesting similarities and dissimilarities with (Evans and Lundblad, 1999; Pennock et al., 2001).another terminal nucleotide transferase, poly(A) poly-merase, which polyadenylates mRNA precursors by Rif1p and Rif2p Promote Formation
of the Nonextendible Telomeric Stateadding approximately 250 nucleotides to precleavedmRNA molecules. Similarly to telomeres, poly(A) tail ex- We identify the Rif1 and Rif2 proteins as critical media-
tors of the state of telomere extendibility. In both rif1 andtension is regulated in cis by the number of bound nu-clear poly(A) binding proteins (PABPN1) associated with rif2 mutant cells, the average telomere length increases,
because the equilibrium between the extendible andthe growing tail (Wahle, 1991). However, in contrast todouble-stranded telomere binding proteins, PABPN1 do nonextendible state is shifted (Figure 5). The extent of
telomere elongation did not increase significantly withnot regulate the initial binding of poly(A) polymerase butrather the processivity during extension. When the tail telomere shortening (Figure 5C).
How do Rif1p and Rif2p promote the nonextendibleis short, PABPN1 stimulates the processivity of poly(A)

Mechanism of Telomere Length Homeostasis333
Telomere Sequencing and Analysisstate and prevent the productive association of tel-Genomic DNA was extracted using the Wizard Genomic Kit (Pro-omerase with the telomere 3� end in a length-dependentmega). Total genomic DNA (30–100 ng) was used for telomere PCRmanner? The Rap1p-Rif1p and -Rif2 complexes mightas described (Forstemann et al., 2000). Briefly, genomic DNA was
mediate their effects by favoring the formation of alter- tailed with dCTP and terminal transferase. Telomeres were amplifiednative DNA structures such as G quartets, which are using a primer (5�-TGTCCGAATTGATCCCAGAG-3�) that hybridized
specifically to the ADE2 gene at the telomere of chromosome VRknown to prohibit telomerase extension in vitro (Zahlerof the recipient strains and with a primer (5�-CGGGATCCG18-3�)et al., 1991). Interestingly, Rap1p is known to promotecomplementary to the oligo-dC tail (see Figure 1B). For amplifyingG quartet formation in vitro (Giraldo et al., 1994). Anothertelomeres of chromosome 1L, a primer (5�-GCGGTACCAGGGTTAtelomerase-inaccessible DNA structure could be pro-GATTAGGGCTG-3�) specific for the subtelomeric sequence of this
vided through t loops, which have been described in chromosome was used. PCR products were separated on 2.5%vertebrates and protozoa, and in which the telomeric 3� preparative agarose gels, purified with a gel extraction kit (Qiagen),
and cloned into the pDrive cloning vector using the PCR Cloningoverhang is tucked away by forming a displacementPlus Kit (Qiagen). Plasmid DNA was prepared from overnight cul-loop with the telomere-proximal region (Griffith et al.,tures using the Millipore Montage Plasmid Miniprep96 Kit. Plasmid1999; Munoz-Jordan et al., 2001; Murti and Prescott,DNA was sequenced using M13 forward or reverse primers and the1999). Experimental evidence for fold-back structuresSequiTherm EXCEL II DNA Sequencing Kit (Epicentre Technologies,
also exists in S. cerevisiae, but the status of the 3� end Madison, WI) on a LiCor DNA Sequencer. Sequences were aligned,in these structures is unknown (de Bruin et al., 2001; and sequence divergence was detected using the contig assembler
of Sequencher (Gene Codes Corporation) or by analyzing the se-Strahl-Bolsinger et al., 1997). If this fold-back structurequencing gel image directly.were to prevent telomerase access, one would predict
that mutations in Sir proteins, which have been shownStatistical Analysisto disrupt this structure, should lead to a dramatic in-Package R v1.8.1 (Ihaka and Gentleman, 1996) was used for stan-crease in telomere length. However, telomere length indard parametric and nonparametric tests. Telomeres were orderedsir mutants is slightly shorter than in wt cells (Palladino according to their nondiverging size, and subgroups containing 10
et al., 1993). It is also possible that Rif1p and Rif2p telomeres were formed. The average telomere size of each subgroupstimulate the telomere association of Cdc13p com- was calculated and the frequency of elongation determined. The
relationship between frequency of initiation and mean length ofplexes, which may not only act as positive but also astelomere of each subgroup was modeled using logistic regression.negative regulators of telomere length (see, e.g., Chan-Several diagnostics were applied to test for lack of fit, outliers, anddra et al., [2001] and Grandin et al. [2001]). A similarleverage points.mechanism has been proposed for human cells in which
TRF1 may transmit its telomerase inhibition via the sin-Acknowledgments
gle-strand end binding protein POT1 (Loayza and DeLange, 2003), while, under other conditions, POT1 may We thank K. Forstemann, J. Promisel Cooper, and the referees forstimulate telomerase activity (Colgin et al., 2003). helpful comments on the manuscript; K. Forstemann and S. Marcand
for discussion; S. van der Straten for DNA sequencing; E. Schwob,In summary, our data demonstrate that telomereV. Lundblad, and S. Marcand for plasmids; and B. Dujon for yeastlength homeostasis is mediated through the telomerestrains. M.T.T. was the recipient of a postdoctoral fellowship fromsubstrate, which is subject to a structural switch thatthe Human Frontier Science Program. This work was supported by
regulates its productive association with telomerase. a Human Frontier Science Program grant and the Swiss NationalThe described experimental system provides a means Science Foundation.of deciphering which telomere-length regulators affectthe equilibrium of the two telomeric states and ultimately Received: July 23, 2003will allow definition of their molecular natures. Revised: February 17, 2004
Accepted: March 3, 2004Experimental Procedures Published: April 29, 2004
Mating of Yeast Cells ReferencesTelomerase-negative isolated transformants were inoculated in 50ml of synthetic medium lacking histidine and grown overnight to a Ancelin, K., Brunori, M., Bauwens, S., Koering, C.E., Brun, C., Ricoul,density of 108 cells/ml, as determined by cell counting. 108 recipient M., Pommier, J.P., Sabatier, L., and Gilson, E. (2002). Targeting assaycells were mixed with 109 exponentially growing telomerase-positive to study the cis functions of human telomeric proteins: evidencedonor cells. The mixtures were immediately concentrated by filtering for inhibition of telomerase by TRF1 and for activation of telomereon four membranes with a diameter of 47 mm each and a pore size degradation by TRF2. Mol. Cell. Biol. 22, 3474–3487.of 0.8 �m (type AA from Millipore). Membranes were placed on
Barnett, M.A., Buckle, V.J., Evans, E.P., Porter, A.C., Rout, D., Smith,prewarmed YPD plates at 30�C. After 3 hr, cells were resuspendedA.G., and Brown, W.R. (1993). Telomere directed fragmentation ofby vigorous vortexing in 30 ml of synthetic medium lacking histidinemammalian chromosomes. Nucleic Acids Res. 21, 27–36.and leucine. Ten milliliters of this suspension was diluted to 40 ml
in the same selective medium and incubated at 30�C under agitation Bodnar, A.G., Ouellette, M., Frolkis, M., Holt, S.E., Chiu, C.-P., Morin,to allow diploids to grow for an additional 2 to 4 hr (5 and 7 hr after G.B., Harley, C.B., Shay, J.W., Lichtsteinter, S., and Wright, W.E.initial mixing, respectively). At each time point (0, 3, 5, and 7 hr after (1998). Extension of life-span by introduction of telomerase intoinitial mixing), aliquots of the suspensions were analyzed. Genomic normal human cells. Science 279, 349–352.DNA was prepared from at least 5 � 107 of each of the parental cells Bryan, T.M., Englezou, A., Gupta, J., Bacchetti, S., and Reddel,and approximately 109 cells from mating mixtures. Mating efficiency R.R. (1995). Telomere elongation in immortal human cells withoutwas estimated by inspection of the cell mixtures under the micro- detectable telomerase activity. EMBO J. 14, 4240–4248.scope at the 3 hr time point. A spot assay on synthetic medium
Bucholc, M., Park, Y., and Lustig, A.J. (2001). Intrachromatid exci-lacking either leucine, histine, or both was performed to quantifysion of telomeric DNA as a mechanism for telomere size control inmating efficiency and to quantify the initial proportion of recipientSaccharomyces cerevisiae. Mol. Cell. Biol. 21, 6559–6573.cells and donor cells. DNA content was measured by fluorocytome-
try as described (Henchoz et al., 1997). Cawthon, R.M., Smith, K.R., O’Brien, E., Sivatchenko, A., and Kerber,

Cell334
R.A. (2003). Association between telomere length in blood and mor- omeric G strand overhangs involves both G and C strand cleavage.Mol. Cell 11, 1021–1032.tality in people aged 60 years or older. Lancet 361, 393–395.
Kelleher, C., Teixeira, M.T., Forstemann, K., and Lingner, J. (2002).Chan, S., Chang, J., Prescott, J., and Blackburn, E. (2001). AlteringTelomerase: biochemical considerations for enzyme and substrate.telomere structure allows telomerase to act in yeasts lacking ATMTrends Biochem. Sci. 27, 572–579.kinases. Curr. Biol. 11, 1240–1250.
Kim, N.W., Piatyszek, M.A., Prowse, K.R., Harley, C.B., West, M.D.,Chandra, A., Hughes, T.R., Nugent, C.I., and Lundblad, V. (2001).Ho, P.L., Coviello, G.M., Wright, W.E., Weinrich, S.L., and Shay,Cdc13 both positively and negatively regulates telomere replication.J.W. (1994). Specific association of human telomerase activity withGenes Dev. 15, 404–414.immortal cells and cancer. Science 266, 2011–2015.Cohn, M., and Blackburn, E.H. (1995). Telomerase in yeast. ScienceKim, S.H., Kaminker, P., and Campisi, J. (1999). TIN2, a new regulator269, 396–400.of telomere length in human cells. Nat. Genet. 23, 405–412.
Colgin, L.M., Baran, K., Baumann, P., Cech, T.R., and Reddel, R.R.Lea, D.E., and Coulson, C.A. (1949). The distribution of the numbers(2003). Human POT1 facilitates telomere elongation by telomerase.of mutants in bacterial populations. J. Genet. 49, 264–285.Curr. Biol. 13, 942–946.Li, B., and Lustig, A.J. (1996). A novel mechanism for telomere sizeConrad, M., Wright, J., Wolf, J., and Zakian, V. (1990). RAP1 proteincontrol in Saccharomyces cerevisiae. Genes Dev. 10, 1310–1326.interacts with yeast telomeres in vivo: overproduction alters tel-Lingner, J., Cooper, J.P., and Cech, T.R. (1995). Telomerase andomere structure and decreases chromosome stability. Cell 63,DNA end replication: no longer a lagging strand problem? Science739–750.269, 1533–1534.Cooper, J.P., Nimmo, E.R., Allshire, R.C., and Cech, T.R. (1997).Loayza, D., and De Lange, T. (2003). POT1 as a terminal transducerRegulation of telomere length and function by a Myb-domain proteinof TRF1 telomere length control. Nature 424, 1013–1018.in fission yeast. Nature 385, 744–747.Lundblad, V., and Blackburn, E.H. (1993). An alternative pathwayde Bruin, D., Zaman, Z., Liberatore, R.A., and Ptashne, M. (2001).for yeast telomere maintenance rescues est1-senescence. CellTelomere looping permits gene activation by a downstream UAS in73, 347–360.yeast. Nature 409, 109–113.Lundblad, V., and Szostak, J.W. (1989). A mutant with a defect inDiede, S.J., and Gottschling, D.E. (1999). Telomerase-mediated tel-telomere elongation leads to senescence in yeast. Cell 57, 633–643.omere addition in vivo requires DNA primase and DNA polymerasesLustig, A., Kurtz, S., and Shore, D. (1990). Involvement of the silenceralpha and delta. Cell 99, 723–733.and UAS binding protein Rap1 in regulation of telomere length.Dunham, M.A., Neumann, A.A., Fasching, C.L., and Reddel, R.R.Science 250, 549–553.(2000). Telomere maintenance by recombination in human cells. Nat.Ihaka, R., and Gentleman, R. (1996). R: A language for data analysisGenet. 26, 447–450.and graphics. J. Comp. Graph. Stat. 5, 299–314.Evans, S.K., and Lundblad, V. (1999). Est1 and Cdc13 as comedia-Makarov, V.L., Hirose, Y., and Langmore, J.P. (1997). Long G tails attors of telomerase access. Science 286, 117–120.both ends of human chromosomes suggest a C strand degradation
Evans, S.K., and Lundblad, V. (2000). Positive and negative regula-mechanism for telomere shortening. Cell 88, 657–666.
tion of telomerase access to the telomere. J. Cell Sci. 113, 3357–Marcand, S., Gilson, E., and Shore, D. (1997). A protein-counting3364.mechanism for telomere length regulation in yeast. Science 275,
Ferreira, M.G., Miller, K.M., and Cooper, J.P. (2004). Indecent expo- 986–990.sure: when telomeres become uncapped. Mol. Cell 13, 7–18.
Marcand, S., Brevet, V., and Gilson, E. (1999). Progressive cis-inhibi-Forstemann, K., and Lingner, J. (2001). Molecular basis for telomere tion of telomerase upon telomere elongation. EMBO J. 18, 3509–repeat divergence in budding yeast. Mol. Cell. Biol. 21, 7277–7286. 3519.Forstemann, K., Hoss, M., and Lingner, J. (2000). Telomerase- Marcand, S., Brevet, V., Mann, C., and Gilson, E. (2000). Cell cycledependent repeat divergence at the 3� ends of yeast telomeres. restriction of telomere elongation. Curr. Biol. 10, 487–490.Nucleic Acids Res. 28, 2690–2694.
McEachern, M.J., and Blackburn, E.H. (1995). Runaway telomereGiraldo, R., Suzuki, M., Chapman, L., and Rhodes, D. (1994). Promo- elongation caused by telomerase RNA gene mutations. Naturetion of parallel DNA quadruplexes by a yeast telomere binding pro- 376, 403–409.tein: a circular dichroism study. Proc. Natl. Acad. Sci. USA 91, 7658–
McElligott, R., and Wellinger, R.J. (1997). The terminal DNA structure7662.
of mammalian chromosomes. EMBO J. 16, 3705–3714.Grandin, N., Damon, C., and Charbonneau, M. (2001). Ten1 functions Mitchell, J.R., Wood, E., and Collins, K. (1999). A telomerase compo-in telomere end protection and length regulation in association with nent is defective in the human disease dyskeratosis congenita. Na-Stn1 and Cdc13. EMBO J. 20, 1173–1183. ture 402, 551–555.Griffith, J.D., Comeau, L., Rosenfield, S., Stansel, R.M., Bianchi, A., Munoz-Jordan, J.L., Cross, G.A., de Lange, T., and Griffith, J.D.Moss, H., and de Lange, T. (1999). Mammalian telomeres end in a (2001). t-loops at trypanosome telomeres. EMBO J. 20, 579–588.large duplex loop. Cell 97, 503–514.
Murti, K.G., and Prescott, D.M. (1999). Telomeres of polytene chro-Hardy, C.F., Sussel, L., and Shore, D. (1992). A RAP1-interacting mosomes in a ciliated protozoan terminate in duplex DNA loops.protein involved in transcriptional silencing and telomere length reg- Proc. Natl. Acad. Sci. USA 96, 14436–14439.ulation. Genes Dev. 6, 801–814.
Naito, T., Matsuura, A., and Ishikawa, F. (1998). Circular chromo-Harley, C.B., Futcher, A.B., and Greider, C.W. (1990). Telomeres some formation in a fission yeast mutant defective in two ATMshorten during ageing of human fibroblasts. Nature 345, 458–460. homologues. Nat. Genet. 20, 203–206.Hemann, M.T., Strong, M.A., Hao, L.Y., and Greider, C.W. (2001). Ouellette, M.M., Liao, M., Herbert, B.S., Johnson, M., Holt, S.E., Liss,The shortest telomere, not average telomere length, is critical for H.S., Shay, J.W., and Wright, W.E. (2000). Subsenescent telomerecell viability and chromosome stability. Cell 107, 67–77. lengths in fibroblasts immortalized by limiting amounts of tel-
omerase. J. Biol. Chem. 275, 10072–10076.Henchoz, S., Chi, Y., Catarin, B., Herskowitz, I., Deshaies, R.J., andPeter, M. (1997). Phosphorylation- and ubiquitin-dependent degra- Palladino, F., Laroche, T., Gilson, E., Axelrod, A., Pillus, L., anddation of the cyclin-dependent kinase inhibitor Far1p in budding Gasser, S.M. (1993). SIR3 and SIR4 proteins are required for theyeast. Genes Dev. 11, 3046–3060. positioning and integrity of yeast telomeres. Cell 75, 543–555.
Huffman, K.E., Levene, S.D., Tesmer, V.M., Shay, J.W., and Wright, Pennock, E., Buckley, K., and Lundblad, V. (2001). Cdc13 deliversW.E. (2000). Telomere shortening is proportional to the size of the separate complexes to the telomere for end protection and replica-G-rich telomeric 3�-overhang. J. Biol. Chem. 275, 19719–19722. tion. Cell 104, 387–396.
Prescott, J., and Blackburn, E.H. (1997). Telomerase RNA mutationsJacob, N.K., Kirk, K.E., and Price, C.M. (2003). Generation of tel-

Mechanism of Telomere Length Homeostasis335
in Saccharomyces cerevisiae alter telomerase action and reveal Inhibition of telomerase by G-quartet DNA structures. Nature 350,718–720.nonprocessivity in vivo and in vitro. Genes Dev. 11, 528–540.
Zhu, L.X., Hathcock, K.S., Hande, P., Lansdorp, P.M., Seldin, M.F.,Ray, A., and Runge, K.W. (1999a). Varying the number of telomere-and Hodes, R.J. (1998). Telomere length regulation in mice is linkedbound proteins does not alter telomere length in tel1� cells. Proc.to a novel chromosome locus. Proc. Natl. Acad. Sci. USA 95, 8648–Natl. Acad. Sci. USA 96, 15044–15049.8653.Ray, A., and Runge, K.W. (1999b). The yeast telomere length count-
ing machinery is sensitive to sequences at the telomere-nontelo-mere junction. Mol. Cell. Biol. 19, 31–45.
Ray, S., Karamysheva, Z., Wang, L., Shippen, D.E., and Price, C.M.(2002). Interactions between telomerase and primase physically linkthe telomere and chromosome replication machinery. Mol. Cell. Biol.22, 5859–5868.
Ritchie, K.B., Mallory, J.C., and Petes, T.D. (1999). Interactions ofTLC1 (which encodes the RNA subunit of telomerase), TEL1, andMEC1 in regulating telomere length in the yeast Saccharomycescerevisiae. Mol. Cell. Biol. 19, 6065–6075.
Samper, E., Flores, J.M., and Blasco, M.A. (2001). Restoration oftelomerase activity rescues chromosomal instability and prematureaging in Terc�/� mice with short telomeres. EMBO Rep. 2, 800–807.
Shore, D. (1997). Telomere length regulation: getting the measureof chromosome ends. Biol. Chem. 378, 591–597.
Singh, S.M., and Lue, N.F. (2003). Ever shorter telomere 1 (EST1)-dependent reverse transcription by Candida telomerase in vitro:evidence in support of an activating function. Proc. Natl. Acad. Sci.USA 100, 5718–5723.
Smith, S., and de Lange, T. (2000). Tankyrase promotes telomereelongation in human cells. Curr. Biol. 10, 1299–1302.
Smogorzewska, A., and de Lange, T. (2004). Regulation of tel-omerase by telomeric proteins. Annu. Rev. Biochem. 73, 177–208.
Smogorzewska, A., van Steensel, B., Bianchi, A., Oelmann, S.,Schaefer, M.R., Schnapp, G., and de Lange, T. (2000). Control ofhuman telomere length by TRF1 and TRF2. Mol. Cell. Biol. 20, 1659–1668.
Strahl-Bolsinger, S., Hecht, A., Luo, K., and Grunstein, M. (1997).SIR2 and SIR4 interactions differ in core and extended telomericheterochromatin in yeast. Genes Dev. 11, 83–93.
Taggart, A.K., Teng, S.C., and Zakian, V.A. (2002). Est1p as a cellcycle-regulated activator of telomere-bound telomerase. Science297, 1023–1026.
Teixeira, M.T., Forstemann, K., Gasser, S.M., and Lingner, J. (2002).Intracellular trafficking of yeast telomerase components. EMBORep. 3, 652–659.
Teng, S.C., and Zakian, V.A. (1999). Telomere-telomere recombina-tion is an efficient bypass pathway for telomere maintenance inSaccharomyces cerevisiae. Mol. Cell. Biol. 19, 8083–8093.
Teng, S., Chang, J., McCowan, B., and Zakian, V.A. (2000). Tel-omerase-independent lengthening of yeast telomeres occurs by anabrupt Rad50p-dependent, Rif-inhibited recombinational process.Mol. Cell 6, 947–952.
van Steensel, B., and de Lange, T. (1997). Control of telomere lengthby the human telomeric protein TRF1. Nature 385, 740–743.
Vulliamy, T., Marrone, A., Goldman, F., Dearlove, A., Bessler, M.,Mason, P.J., and Dokal, I. (2001). The RNA component of telomeraseis mutated in autosomal dominant dyskeratosis congenita. Nature413, 432–435.
Wahle, E. (1991). A novel poly(A)-binding protein acts as a specificityfactor in the second phase of messenger RNA polyadenylation. Cell66, 759–768.
Wellinger, R.J., Ethier, K., Labrecque, P., and Zakian, V.A. (1996).Evidence for a new step in telomere maintenance. Cell 85, 423–433.
Wotton, D., and Shore, D. (1997). Novel Rap1p-interacting factor,Rif2p, cooperates with Rif1p to regulate telomere length in Sacchar-omyces cerevisiae. Genes Dev. 11, 748–760.
Wright, W.E., Tesmer, V.M., Huffman, K.E., Levene, S.D., and Shay,J.W. (1997). Normal human chromosomes have long G-rich tel-omeric overhangs at one end. Genes Dev. 11, 2810–2821.
Zahler, A.M., Williamson, J.R., Cech, T.R., and Prescott, D.M. (1991).


Mechanism of Telomere Length Homeostasis
Supplemental Data for:
Teixeira et al., Cell 117, pp. 323–335
Yeast Strains and Plasmids
Yeast strains are listed in Supplemental Table S1. YT48 was generated by
transformation of GA426 with Stu I-cut plasmid 422 containing the URA3 gene
(Lengronne et al., 2001). Telomerase-deficient cells (termed ‘recipients’) were
constructed in YT48 using PCRgenerated disruption cassettes according to Longtine
et al., (1998), by replacing TLC1 or EST1 with the HIS5 marker (Teixeira et al., 2002)
to obtain YT48/tlc1-Δ and YT48/est1-Δ, respectively. YT57 was obtained by
transformation of YT48/est1-Δ with plasmid pVL1120 (CEN TRP1 Est1-DBDCdc13 )
(Evans and Lundblad, 1999). FYBL1-23D (Fairhead et al., 1996), and MAY01 were
used as telomerase-positive strains for mating (termed ‘donors’). MAY01 was
derived from FYBL1-23D by replacing the subtelomeric region at chromosome 1L
with the URA3 marker as follows. A 1582 bp DNA fragment corresponding to the
Cterminus of open reading frame SEO1 was amplified from genomic DNA using
primers SEO1F (5’-ACATGCATGCGAGACTAGTAGCATAGTGAAC-3’) and SEO1R1 (5’-
CCCAAGCTTGTTCGCGG-AACGAAAACTGTGCG-3’). The PCR product was cut with SphI
and HindIII and subcloned into the corresponding sites of plasmid pVII-L_URA3-TEL
(Gottschling et al., 1990), giving pI-L_URA3-TEL. Strain FYBL1-23D was transformed
with a linear SphI/EcoRI- restriction fragment from pI-L_URA3-TEL containing the C-
terminus of the SEO1 gene, theURA3-marker and a telomeric seeding sequence.
Transformants were selected as URA3 positive colonies and integration at 1L was
confirmed by PCR analysis. YT56 and YT55 rif1-Δ strains were obtained by replacing
RIF1 with a PCR-product containing the TRP1 marker (Longtine et al., 1998)
obtained with primers RIF1F1 (5’-ACGTTATTACAGG-
ATTGCCATTGCAAAATCGTTTTTGTGGTCAATTTGCACGGATCCCCGGGTTAATTAA-3’) and
RIF1R1 (5’-GAGCTCGTTTAAAC-3’). For construction of the rif2-Δ strains MAY02 and
MAY03, the disruption cassettes were obtained by PCR with primers RIF2F1 (5’-
ATGGAGCATGTAGATTCCGATTTTGCACCTATAAGGAGATCGAAAAAGGTCGGATCCCCGGG-
TTAATTAA-3’) and RIF2R1 (5’-GCCATCTCTTTGTATTGTTCGAACTCTTTCAAAAGACCTT-
GGTAATTTATGAATTCGAGCTCGTTTAAAC-3’).

Mechanism of Telomere Length Homeostasis
References
Evans, S.K., and Lundblad, V. (1999). Est1 and Cdc13 as comediators of telomerase access. Science 286, 117–120.
Fairhead, C., Llorente, B., Denis, F., Soler, M., and Dujon, B. (1996). New vectors for combinatorial deletions in yeast chromosomes and for gap-repair cloning using ‘split-marker’ recombination. Yeast 12, 1439–1457.
Gottschling, D.E., Aparicio, O.M., Billington, B.L., and Zakian, V. (1990). Position effect at S. cerevisiae telomeres: reversible repression of Pol II transcription. Cell 16, 751–762.
Lengronne, A., Pasero, P., Bensimon, A., and Schwob, E. (2001). Monitoring S phase progression globally and locally using BruU incorporation in TK(+) yeast strains. Nucleic Acids Res. 29, 1433–1442.
Longtine, M.S., McKenzie, A., III, Demarini, D.J., Shah, N.G., Wach, A., Brachat, A., Philippsen, P., and Pringle, J.R. (1998). Additional modules for versatile and economical PCR-based gene deletion and modification in Saccharomyces cerevisiae. Yeast 14, 953–961.
Teixeira, M.T., Forstemann, K., Gasser, S.M., and Lingner, J. (2002). Intracellular trafficking of yeast telomerase components. EMBO Rep. 3, 652–659.

Mechanism of Telomere Length Homeostasis
Strain Genotype Source
GA426 MATa ura3-52 trp1 leu2 his3-11 ade2 can1-∆ DIA5-1 Lingner laboratory collection
YT48 MATa ura3-52 trp1 leu2 his3-11 ade2 can1-∆ DIA5-1 URA3::TKx7 This work
YT57 MATa ura3-52 trp1 leu2 his3-11 ade2 can1-∆ est1∆::HIS5(S.p.) DIA5-1 URA3::TKx7 + pVL1120 (CEN TRP1 Est1-DBDCdc13) This work
YT56 MATa ura3-52 trp1 leu2 his3-11 ade2 can1-∆ DIA5-1 URA3::TKx7 rif1∆::TRP1 This work
MAY02 MATa ura3-52 trp1 leu2 his3-11 ade2 can1-∆ DIA5-1 URA3::TKx7 rif2∆::TRP1 This work
FYBL1-23D MATα ura3-∆851 trp1∆63 his3∆200 Fairhead et al., 1996
MAY01 MATα ura3-∆851 trp1∆63 his3∆200 SEO1::URA3 tel1L∆ This work
YT55 MATα ura3-∆851 trp1∆63 his3∆200 rif1∆::TRP1 This work
MAY03 MATα ura3-∆851 trp1∆63 his3∆200 rif2∆::TRP1 This work
Supplemental Table S1. Saccharomyces cerevisiae Strains


Chapter 3
Tel1 Kinase and Subtelomere-Bound Tbf1 Mediate Preferential
Elongation of Short Telomeres by Telomerase in Yeast
Article published in EMBO Reports, vol. 8. pp. 1080-1085, November 8th, 2007


Tel1 Activates Telomerase at Short Telomeres
71
It was established in the Chapter 2 that the shortest telomeres are preferentially
elongated in each cell cycle. Rap1-Rif1 and -Rif2 complexes associate with the
double stranded telomeric repeats of budding yeast and create a negative feedback
loop, called “counting mechanism”, that regulates telomerase in a length dependent
manner (Marcand et al, 1997a; Wotton & Shore, 1997). How this regulatory signal
is transmitted to telomerase is not clear, but it is dependent on the presence of the
PI-3 kinase-like kinase Tel1 (Ray & Runge, 1999). The kinase function of Tel1 and
the other member of the same protein family, Mec1, seem to be partially redundant
and essential for the telomere preservation, since simultaneous deletion of both
leads to cellular senescence (Ritchie et al, 1999).
To gain more precise insight into the mechanisms by which Tel1 and Mec1
regulate telomere length, I performed STEX analysis in tel1Δ and mec1Δ cells, as
described in the following chapter. As well, I describe the role of a backup counting
mechanism of telomere length control, which depends on yeast subtelomeric
regions, or more precisely, the insulator protein Tbf1.


Tel1 kinase and subtelomere-bound Tbf1 mediatepreferential elongation of short telomeres bytelomerase in yeastMilica Arneric & Joachim Lingner+
Swiss Institute for Experimental Cancer Research (ISREC), Ecole Polytechnique Federale de Lausanne (EPFL) and NCCR Program
‘Frontiers in Genetics’, Epalinges s/Lausanne, Switzerland
Telomerase enables telomere length homeostasis, exhibitingincreasing preference for telomeres as their lengths decline. Thisregulation involves telomere repeat-bound Rap1, which providesa length-dependent negative feedback mechanism, and the Tel1and Mec1 kinases, which are positive regulators of telomerelength. By analysing telomere elongation of wild-type chromo-some ends at single-molecule resolution, we show that in tel1Dcells the overall frequency of elongation decreases considerably,explaining their short telomere phenotype. At an artificialtelomere lacking a subtelomeric region, telomere elongation nolonger increases with telomere shortening in tel1D cells. Bycontrast, a natural telomere, containing subtelomeric sequence,retains a preference for the elongation of short telomeres.Tethering of the subtelomere binding protein Tbf1 to the artificialtelomere in tel1D cells restored preferential telomerase action atshort telomeres; thus, Tbf1 might function in parallel to Tel1,which has a crucial role in a TG-repeat-controlled pathway forthe activation of telomerase at short telomeres.Keywords: Tel1; Mec1; Tbf1; STEX; telomerase;telomere length homoeostasisEMBO reports (2007) 8, 1080–1085. doi:10.1038/sj.embor.7401082
INTRODUCTIONBy measuring telomere elongation at single chromosome endmolecules at nucleotide resolution, we established that telomerasedoes not extend every telomere in every cell cycle and that itshows an increasing preference for telomeres as their lengthsdecrease (Teixeira et al, 2004). Rap1–Rif1–Rif2 complexesassemble with the double-stranded region of the telomere andcreate a negative feedback loop that regulates telomerase in a
length-dependent manner (Kyrion et al, 1992; Krauskopf &Blackburn, 1996; Marcand et al, 1997; Wotton & Shore, 1997;Teixeira et al, 2004), but how this regulatory signal is transmittedto telomerase is not yet known. At the telomeric 30 end,telomerase is recruited during the S phase of the cell cyclethrough a direct interaction between the telomerase-associatedEst1 and Cdc13, which binds to the telomeric 30 overhang (Evans& Lundblad, 1999; Pennock et al, 2001; Fisher et al, 2004).
The two yeast members of the phosphoinositide-3-kinase-related family—Tel1 and Mec1 (Saccharomyces cerevisiae homo-logues of human ataxia telangiectasia mutated (ATM) and ATMand Rad3-related (ATR), respectively)—are also found at telo-meres and are necessary for the correct regulation of telomerelength. Yeast cells lacking Tel1 have short but stable telomeres,whereas deletion of MEC1 yields only a mild telomere shorteningphenotype (Greenwell et al, 1995; Ritchie et al, 1999). The kinasefunctions of Tel1 and Mec1 seem to be partly redundant intelomere length maintenance, as simultaneous deletion ofboth TEL1 and MEC1 results in a progressive loss of telomericsequences, leading to cellular senescence (Ritchie et al, 1999). Inwild-type cells, ectopic tethering of additional Rap1 moleculesadjacent to telomere repeats results in telomere shortening,whereas tel1D cells are not responsive to increased localconcentrations of Rap1 (Marcand et al, 1997; Ray & Runge, 1999).
Despite the crucial roles of Tel1 and Mec1 in telomeremaintenance, their mechanisms of action remain unknown. Tel1does not directly affect telomerase catalytic activity, but seemsto mainly modify the structure of the telomere end complex(Chan et al, 2001; Tsukamoto et al, 2001). Interestingly, thesingle-stranded telomere-binding protein Cdc13 was identifiedas a substrate of these kinases (Tseng et al, 2006). Furthermore, veryrecently, Est2, Est1 and Tel1 have been reported to associatepreferentially with short telomeres (Bianchi & Shore, 2007; Sabourinet al, 2007), and the association of Est2 and Est1 with telomeres isseverely impaired in tel1D cells (Goudsouzian et al, 2006).
To gain deeper insight into the roles of Tel1 and Mec1 in telo-mere maintenance and length regulation, we analysed telomereelongation of single chromosome end molecules at nucleotideresolution in vivo, in yeast strains deleted for TEL1, MEC1 or both.
Received 10 April 2007; revised 2 August 2007; accepted 20 August 2007;published online 5 October 2007
+Corresponding author. Tel: þ 41 21 692 5912; Fax: þ 41 21 652 6933;E-mail: [email protected]
Swiss Institute for Experimental Cancer Research (ISREC), Ecole PolytechniqueFederale de Lausanne (EPFL) and NCCR Program ‘Frontiers in Genetics’,Epalinges s/Lausanne CH-1066, Switzerland
EMBO reports VOL 8 | NO 11 | 2007 &2007 EUROPEAN MOLECULAR BIOLOGY ORGANIZATION
scientificreportscientific report
1080

RESULTSExtension analysis of single telomeresTo study the roles of Tel1 and Mec1 in telomere elongationin S. cerevisiae, we monitored single telomere extension eventsat nucleotide resolution at two different chromosome endsin vivo as described (Teixeira et al, 2004; Fig 1A; Methods). Werefer to this method hereafter as STEX, for Single TelomereEXtension assay. Telomerase-negative strains—referred to as‘recipient’—were created from haploid tel1D or mec1D sml1Dyeast cells by deleting one of the essential components necessaryfor telomerase activity in vivo, EST1 or TLC1. EST1 encodes atelomerase protein subunit that is involved in telomeraserecruitment (Lundblad & Szostak, 1989) and TLC1 encodes thetelomerase RNA moiety (Singer & Gottschling, 1994). All mec1Dstrains were also sml1D, to suppress lethality of the MEC1 deletion(Zhao et al, 2001). For simplicity, hereafter we use mec1D torepresent mec1D sml1D double mutants. As the steady-statetelomere length in tel1D cells is short (170 bp at telomere VR (rightarm of chromosome V) in our strain background), we had toensure that recipient telomeres were not lost before mating.Therefore, telomeres were pre-elongated in the recipient byexpressing a fusion protein between the DNA-binding domainof Cdc13 and Est1 (Est1-DBDCdc13)—a construct that comple-ments est1D (and est1D tel1D, this study) and leads to telomereoverelongation (Evans & Lundblad, 1999). Before matings, cellsthat had lost the Est1-DBDCdc13 plasmid (pVL1120) were selectedto obtain telomerase-negative cells. Functional telomerase wasprovided by mating the telomerase-negative recipients withtelomerase-positive donor strains of the opposite mating typethat were also deleted for TEL1, MEC1 or both. Thus, telomeresthat had shortened in the telomerase-negative parents becamere-extended in the zygote owing to the presence of complement-ing telomerase (Fig 1A). DNA was isolated from the matingwhen most zygotes had completed a single S phase, and telomereelongation was detected by cloning and sequencing two differenttelomeres originating from the recipient strain. Telomere VR wasamplified by telomere PCR (Forstemann et al, 2000). Nativetelomere IL (left arm of chromosome I) from the recipients wasamplified using a primer complementary to a sequence that wasdeleted in the donor strains (Teixeira et al, 2002, 2004). PCR-amplified telomeres were cloned and sequenced. As yeasttelomerase adds imperfect 50-(TG)0–6TGGGTGTG(G)0–1-30 repeats(Forstemann & Lingner, 2001), telomere elongation is detectableon sequence alignment by the divergence of telomeric sequencesin the telomere-distal region. As expected, telomere sequencedetermination at VR and IL in the tel1D est1D and mec1D tlc1Drecipient strains on clonal expansion before mating revealeddifferences in size between the telomeres emerging from thedifferent sisters, whereas sequence divergence was rare. Attelomere VR, 1 out of 35 sequences (2.8%) diverged in tel1Dest1D and 3 out of 45 sequences (6.6%) diverged in mec1Dtlc1D cells (supplementary Fig S1A online); at telomere IL, 4 outof 54 sequences (7.4%) diverged in tel1D est1D and 2 out of34 sequences (5.8%) diverged in mec1D tlc1D cells (supplemen-tary Fig S1B online). These results are similar to the 6.6%of telomerase-independent events at VR seen previously inwild-type cells (Teixeira et al, 2004). Thus, TEL1 or MEC1deletion does not significantly affect telomerase-independentmodes of telomere elongation.
Tel1 mediates the recognition of short telomeresTelomerase-mediated elongation of telomeres in a tel1D back-ground was analysed in four independent STEX assays (Fig 1D).Among the 133 sequences analysed, we detected 12 events inwhich sequences diverged (9%). This frequency is considerablylower than that observed in wild-type cells for the correspondinglength range, in which the frequency increased from approxi-mately 20% at a telomere length of 200 nucleotides to 45% at 100nucleotides (Fig 1C,F; data pooled from Teixeira et al, 2004).Thus, the lower frequency of extension in tel1D cells provides anexplanation for their short telomere phenotype. Furthermore, inmarked contrast to wild-type cells, the frequency of elongation atVR did not increase for short telomeres in tel1D cells. Indeed,not a single telomere extension event was detected for theanalysed telomeres that were shorter than 100 nucleotides(Fig 1D). The apparent lack of correlation between frequencyof telomere elongation and telomere length was confirmed bystatistical analysis. Sequences were pooled into groups of 15and the frequency of elongation was plotted against the meantelomere length for each subgroup (Fig 1F). In wild-type cells shorttelomeres were preferentially elongated (Fig 1F, grey curve;Teixeira et al, 2004), whereas no correlation was establishedbetween divergence frequency and telomere length in thetel1D background (tel1D/tel1D est1D/EST1: R¼ 0.34, Ppearson
¼ 0.3749246; P¼ 0.29, Pspearman¼ 0.4366237). Thus, Tel1 isnecessary for preferential elongation of short telomeres at VR bytelomerase. The ADE2-tagged telomere VR lacks subtelomericelements, and telomere length sensing is thought to rely solelyon TG repeats and the Rap1-counting mechanism. Thus, ouranalysis suggests that Rap1-mediated telomerase control relieson the presence of Tel1, at least during the first zygotic cell cycleanalysed in this experiment. However, the lack of correlationbetween telomere elongation frequency and length observedhere does not provide an explanation for the residual telomerelength regulation at VR, which maintains a short but stable sizerange in tel1D cells.
In mec1D cells, we analysed sequence divergence in mec1D/mec1D tlc1D/TLC1 diploid cells from five independent STEXassays (Fig 1E). The overall frequency of elongation in the ana-lysed size range was 26%. In contrast to tel1D cells, mec1D cellsextended very short telomeres at high frequency. However, theinverse correlation between the frequency of telomere elongationand telomere length was also perturbed in mec1D cells, as thedata could not be fitted to the logistic regression model (seethe supplementary information online; mec1D/mec1D tlc1D/TLC1: R¼�0.53, Ppearson¼ 0.1407485; P¼�0.42, Pspearman
¼ 0.2499173; Fig 1F). Thus, Mec1 and Tel1 are both requiredfor optimal regulation of telomerase at chromosome VR, with Tel1having crucial role.
Role of a subtelomeric region for length sensingTo monitor elongation events at a natural telomere, we performedSTEX for telomere IL in the same tel1D genomic DNA samples asabove. To analyse telomere IL in the mec1D background,however, a new set of matings were performed using a donorstrain that lacked the subtelomeric region at telomere IL. Thetelomere elongation analyses at IL are shown in Fig 2B–D.Markedly, at this native telomere, and in contrast to the taggedtelomere VR, short telomeres were preferentially extended by
Tel1 activates telomerase at short telomeres
M. Arneric & J. Lingner
&2007 EUROPEAN MOLECULAR BIOLOGY ORGANIZATION EMBO reports
scientificreport
1081

telomerase in tel1D cells (Fig 2C). The sequences were pooled ingroups of 15 and the frequency of elongation was plotted againstmean telomere length for each subgroup (Fig 2E). Although theelongation frequencies were lower than in the wild-type cells attelomere IL (grey line in Fig 2E; Teixeira et al, 2004), the inversecorrelation between the frequency of elongation and telomere lengthwas significant (tel1D/tel1D est1D/EST1: R¼�0.64, Ppearson
¼ 0.03462063; P¼�0.75, Pspearman¼ 0.01115861). Thus, despite
the TEL1 deletion and a presumed inactivation of the Rap1-mediatedcounting mechanism, telomere length was still sensed at telomereIL and transmitted to the machinery that regulates telomerase. Thus,our analysis of telomere elongation at telomere IL suggests theexistence of a cryptic telomere length-sensing mechanism that seemsto rely on chromosomal regions other than the TG repeats.
For STEX on telomere IL in mec1D/mec1D tlc1D/TLC1 zygotes,three independent matings were analysed (Fig 2D). The overall
B
Telo
mer
e le
ngth
(nt)
100
200
300
0
400
Telo
mer
e le
ngth
(nt)
100
200
300
0
1.0
0.2
0.4
0.6
0.0
0.8
100 200 3000
Freq
uenc
y of
elo
ngat
ion
400
wt
• •
•
••
•
•
••
Telomere length (nt)
FRecipienttelomerase-negative
cell
Donortelomerase-positive
cell
Mating
Replicationtelomerase acts
A
STEX
Telomere PCRClone and sequence
telomeresAnalysis of sequence
divergence
Telo
mer
e le
ngth
(nt)
100
200
300
0
400
500
ADE2 (TG1–3)nTel ADE2::VR
Oligo
C
E
D
tel1/∆/tel1∆ est1∆/EST1
mec1∆/mec1∆ tlc1∆/TLC1
est1∆/EST1
tel1∆/tel1∆ est1∆/EST1
mec1∆/mec1∆ tlc1∆/TLC1
Tel1 activates telomerase at short telomeres
M. Arneric & J. Lingner
EMBO reports VOL 8 | NO 11 | 2007 &2007 EUROPEAN MOLECULAR BIOLOGY ORGANIZATION
scientificreport
1082

Fig 1 | Single telomere extension analysis at tagged telomere VR in tel1D and mec1D cells. (A) Methodology of the STEX assay; see text for details.
(B) Schematic of tagged telomere VR. The vector-derived sequence is between the ADE2 gene and the telomeric tract ((TG1–3)n). (C) Telomere VR sequence
analysis of est1D/EST1 zygotes. Recipient est1D cells were mated with EST1 cells for 3 h. Telomere VR was PCR amplified, cloned, sequenced and analysed for
sequence divergence as described in the Methods. Individual telomeres are represented by vertical bars. The red bars indicate the telomeric region that is non-
diverging; the blue bars indicate the telomeric region in which the sequence diverges from the sisters. Data are pooled from experiments published in Teixeira
et al (2004). (D) Telomere VR sequence analysis of tel1D/tel1D est1D/EST1 zygotes. Recipient tel1D est1D cells were mated with tel1D EST1 cells for 3 h and
the sequences of the tagged telomere VR were analysed. Results from four individual matings are pooled. (E) Telomere VR sequence analysis of mec1D/mec1Dtlc1D/TLC1 zygotes. Recipient mec1D tlc1D cells were mated with mec1D TLC1 cells for 3 h and the sequences of the tagged telomere VR were analysed.
Results from five individual matings are pooled. (F) Frequency of telomere VR extension as a function of telomere length. Sequences obtained from (C) and
(D) were ordered according to non-diverging telomere size (as shown in the graphs in (C) and (D)) and pooled into subgroups each containing 15 telomeres.
The frequency of elongation in each subgroup was calculated and plotted as a function of telomere length. The grey curve (wild type; wt) describing diverging
events at telomere VR in an est1-D/EST1 zygote (otherwise wild-type cells) was pre-established (Teixeira et al, 2004). STEX, single telomere extension analysis.
b
A
B
C
1.0
0.2
0.4
0.6
0.0
0.8
100 200 3000
Freq
uenc
y of
elo
ngat
ion
400
wt
Telomere length (nt)
E Telomere IL
Telo
mer
e le
ngth
(nt)
100
200
300
0
Telo
mer
e le
ngth
(nt)
100
200
300
0
Telo
mer
e le
ngth
(nt)
100
200
300
0
D
XC XR (TG1–3)nTel IL
Oligo
XC=X element core sequence XR=X element combinatorial repeats
est1∆/EST1
tel1∆/tel1∆ est1∆/EST1
tel1∆/tel1∆ est1∆/EST1
mec1∆/mec1∆ tlc1∆/TLC1
mec1∆/mec1∆ tlc1∆/TLC1
Fig 2 | Single telomere extension analysis at native telomere IL in tel1D and mec1D cells. (A) Schematic of native telomere IL (Tel IL). The telomeric
X element combinatorial repeat region contains repeats of the D, C, B and A types, as well as Tbf1-binding sites. (B) Telomere IL sequence analysis
of est1D/EST1 zygotes. Data are pooled from experiments published in Teixeira et al (2004). Methods and labelling are as described in Fig 1.
(C) Telomere IL sequence analysis of tel1D/tel1D est1D/EST1 zygotes. tel1D est1D cells were mated with tel1D EST1 cells lacking the subtelomeric
region of telomere IL. Results from four individual matings are pooled. The DNA samples analysed here for telomere IL are the same as in Fig 1C for
telomere VR. (D) Telomere IL sequence analysis of mec1D/mec1D tlc1D/TLC1 zygotes. Results from three individual matings are pooled. (E) Frequency
of telomere IL extension as a function of telomere length. Analysis was performed as in Fig 1E. STEX, single telomere extension analysis; wt, wild type.
Tel1 activates telomerase at short telomeres
M. Arneric & J. Lingner
&2007 EUROPEAN MOLECULAR BIOLOGY ORGANIZATION EMBO reports
scientificreport
1083

frequency of elongation (24% for all analysed telomeres) was similarto that observed at telomere VR, although the mating efficiency in theIL experiments was lower (70%). In addition, a significant inversecorrelation between divergence events and telomere length wasobserved at telomere IL (mec1D/mec1D tlc1D/TLC1: R¼�0.84,Ppearson¼ 0.009121891; P¼�0.75, Pspearman¼ 0.03675595; Fig 2E).Notably, elongation frequency was lower at longer telomeres inmec1D than in wild type, which cannot be solely explained by lowermating efficiency. This lower overall extension frequency couldexplain the shorter telomere phenotype of mec1D cells.
Tbf1 tethering provides a backup counting mechanismA Rap1-independent telomere length-sensing mechanism, whichrelies on the presence of subtelomeric DNA bound by Tbf1,was shown in tel1D cells (Koering et al, 2000; Alexander & Zakian,2003; Brevet et al, 2003; Berthiau et al, 2006). The ADE2-tagged VRtelomere lacks subtelomeric elements and therefore is presumably notbound by Tbf1. To test whether tethering of Tbf1 to a subtelomere-lesschromosome end could restore preferential elongation of shorttelomeres in tel1D cells, we used plasmids from the Gilson laboratorythat express the amino terminus of Tbf1 fused to the Gal4-DNA-binding domain (Gbd-Tbf1N). Effects of expression and telomerictethering of Gbd-Tbf1N fusion protein or Gbd alone were tested bySTEX analysis of a tagged telomere VIIL containing six subtelomericGbd-binding sites (Fig 3A). Consistent with the analysis of tagged VR,telomerase did not preferentially elongate short tagged VIIL in tel1Dcells expressing the Gbd domain (Fig 3B). However, on expression ofGbd-Tbf1N in tel1D cells, short tagged VIIL telomeres were againpreferentially elongated over long telomeres (Fig 3C). This analysistherefore shows that Gbd-Tbf1N provides a Tel1-independentmechanism to activate telomerase at short telomeres.
Telomerase activity requires Tel1 or Mec1Strains deleted simultaneously for TEL1 and MEC1 show an evershorter-telomere phenotype and senesce (Ritchie et al, 1999). It wasproposed that the presence of telomerase could delay senescence intel1D mec1D strains, suggesting residual telomerase activity attelomeres in the absence of both kinases (Chan et al, 2001). Weanalysed three independent tel1D mec1D mutant spores aftersporulating heterozygous mutants, and checked for telomeresequence divergence at telomere VR after 29 generations of clonalexpansion. The results are pooled in supplementary Fig S2 online. Of53 telomeres 7 showed divergence (13.2%). This frequency is onlyslightly higher than the recombinational background seen in tel1Dest1D or mec1D tlc1D cells (see above). A slightly elevatedrecombination frequency in mec1D tel1D cells is consistent with aprevious report addressing mitotic recombination and telomeric fusionevents (Craven et al, 2002). Hence, these results show that telomerasecannot extend telomeres in the absence of Mec1 and Tel1 kinases.
DISCUSSIONThe mechanisms by which telomerase is preferentially activated atshort telomeres have been poorly defined. In this study, we haveshown that Tel1 is a crucial mediator of this control. Indeed, at anartificial telomere lacking subtelomeric repeats, telomerase wasunable to distinguish short from long telomeres in the absenceof the Tel1 kinase. Thus, it seems that Tel1 recognizes shorttelomeres and mediates their preferential elongation by telo-merase. Mec1 has a less pronounced but still important role in the
regulation of telomerase activation; the underlying mechanismsare not yet known. Interestingly, Tel1 can phosphorylate Cdc13in vitro (Tseng et al, 2006); thus, it is conceivable that Tel1 regu-lates the telomerase recruitment step. Consistent with this idea isthe recent finding that Tel1 is required for normal levels of Est1and Est2 association with telomeric DNA (Goudsouzian et al,2006). In addition, it has been recently shown that short telomeresare more avidly bound by Est1, Est2 and Tel1 (Bianchi & Shore,2007; Sabourin et al, 2007).
Curiously, the extension frequency at a natural telomere wasstill regulated in a length-dependent manner even in the absenceof TEL1. This shows a crucial difference between the ADE2-taggedtelomere VR and the natural telomere IL. We have shown usingthe URA3-tagged telomere VIIL that tethering of Gbd-Tbf1N,but not Gbd alone, can rescue the TEL1-deletion phenotype, withregard to the activation of telomerase at short telomeres.Interestingly, the Tbf1-mediated mechanism seems active only
B
Telo
mer
e le
ngth
(nt)
100
200
300
0
Telo
mer
e le
ngth
(nt)
100
200
300
0
ADH4 URA3
Oligo (TG1–3)n
6 × UASG
Tel URA3::VIIL
tel1∆ /tel1∆ est1∆ /EST1 pGbd
tel1∆ /tel1∆ est1∆ /EST1 pGbd-Tbf1N
A
C
P=0.0002
<100 nt >100 nt
<100 nt >100 nt
P=0.3638
Fig 3 | Tethering of the Tbf1 amino-terminal domain to the subtelomere
restores preferential elongation of short telomeres on tagged telomere
VIIL in tel1D cells. (A) Schematic of tagged telomere VIIL. Six UASGs
are present between the URA3 reporter gene and the telomeric tract
((TG1–3)n). (B) Telomere VIIL sequence analysis of tel1D/tel1Dest1D/EST1 diploids expressing Gbd alone. tel1D est1D pGbd cells were
grown and mated with tel1D EST1 cells. Diploids were propagated for
five generations after initial mating, and then telomeres were analysed
for sequence divergence. Results from three individual matings are
pooled. (C) Telomere VIIL sequence analysis of tel1D/tel1D est1D/EST1
diploids expressing Gbd-Tbf1N fusion. tel1D est1D pGbd-Tbf1N cells
were grown and mated with tel1D EST1 cells. Diploids were propagated
for five generations after initial mating, and then telomeres were
analysed for sequence divergence. Results from three individual matings
are pooled. Gbd, Gal4-DNA-binding domain.
Tel1 activates telomerase at short telomeres
M. Arneric & J. Lingner
EMBO reports VOL 8 | NO 11 | 2007 &2007 EUROPEAN MOLECULAR BIOLOGY ORGANIZATION
scientificreport
1084

upon TEL1 deletion, as changing the number of 50-TTAGGG-30
repeats internally to yeast telomeric repeats influenced telomerelength in tel1D but not in wild-type cells (Brevet et al, 2003). Weestimate that the Tbf1-mediated backup mechanism mightbe important for the efficient healing of telomeres that have acci-dentally lost most or all of the telomeric repeats and Rap1-bindingsites, either owing to a double-strand break near the base of thetelomere or replication fork collapse. Indeed, it is well establishedthat telomeric DNA provides a stumbling block for the semi-conservative DNA replication machinery (for example, Cooperet al, 1997; Ivessa et al, 2002; Crabbe et al, 2004), and entiretelomere tract loss might occur at a low but significant frequency.
METHODSSingle telomere extension analysis (STEX). Yeast cells weremated as described previously (Teixeira et al, 2004). Matingefficiency was measured using a spot dilution assay on syntheticmedia to select for growth of recipient, donor or mated cells, orby plating a mixture of approximately 200 recipient or mated cellson plates selective for recipients, and then replica plating themon double selective media, specific for mated cells. To determinethe effects of Gbd-Tbf1N tethering to the subtelomere in tel1Dcells, mated cells were grown in medium selective for diploids forfive cell- cycles after initial mating, before sequence divergencewas analysed at the tagged telomere VIIL (Berthiau et al, 2006).These yeast strains mated with a lower efficiency (5–15%), but theselection of diploids for five generations allowed the analysisof cultures in which diploids prevailed, although preference forelongation of short versus long telomeres could still be distin-guished. See the supplementary information online for furtherdetails and a description of yeast strains.Supplementary information is available at EMBO reports online(http://www.emboreports.org).
ACKNOWLEDGEMENTSWe thank Eric Gilson, Vicki Lundblad and Susan Gasser for plasmids andstrains; Teresa Teixeira, Brian Luke and Michael Chang for technicalcontributions; Marc Gasser for computational expertise; and JuliaPromisel Cooper and Michael Chang for reading the manuscript. Thiswork was supported by the Swiss National Science Foundation, the SwissCancer League, the Human Frontier Science Program and the EuropeanUnion 6th Framework Programme.
REFERENCESAlexander MK, Zakian VA (2003) Rap1p telomere association is not
required for mitotic stability of a C3TA2 telomere in yeast. EMBO J 22:1688–1696
Berthiau AS, Yankulov K, Bah A, Revardel E, Luciano P, Wellinger RJ, Geli V,Gilson E (2006) Subtelomeric proteins negatively regulate telomereelongation in budding yeast. EMBO J 25: 846–856
Bianchi A, Shore D (2007) Increased association of telomerase with shorttelomeres in yeast. Genes Dev 21: 1726–1730
Brevet V, Berthiau AS, Civitelli L, Donini P, Schramke V, Geli V, Ascenzioni F,Gilson E (2003) The number of vertebrate repeats can be regulated at yeasttelomeres by Rap1-independent mechanisms. EMBO J 22: 1697–1706
Chan SW, Chang J, Prescott J, Blackburn EH (2001) Altering telomerestructure allows telomerase to act in yeast lacking ATM kinases. Curr Biol11: 1240–1250
Cooper JP, Nimmo ER, Allshire RC, Cech TR (1997) Regulation of telomerelength and function by a Myb-domain protein in fission yeast. Nature385: 744–747
Crabbe L, Verdun RE, Haggblom CI, Karlseder J (2004) Defective telomerelagging strand synthesis in cells lacking WRN helicase activity.Science 306: 1951–1953
Craven RJ, Greenwell PW, Dominska M, Petes TD (2002) Regulation ofgenome stability by TEL1 and MEC1, yeast homologs of the mammalianATM and ATR genes. Genetics 161: 493–507
Evans SK, Lundblad V (1999) Est1 and Cdc13 as comediators of telomeraseaccess. Science 286: 117–120
Fisher TS, Taggart AK, Zakian VA (2004) Cell cycle-dependent regulationof yeast telomerase by Ku. Nat Struct Mol Biol 11: 1198–1205
Forstemann K, Lingner J (2001) Molecular basis for telomere repeatdivergence in budding yeast. Mol Cell Biol 21: 7277–7286
Forstemann K, Hoss M, Lingner J (2000) Telomerase-dependent repeatdivergence at the 30 ends of yeast telomeres. Nucleic Acids Res 28:2690–2694
Goudsouzian LK, Tuzon CT, Zakian VA (2006) S. cerevisiae Tel1p andMre11p are required for normal levels of Est1p and Est2p telomereassociation. Mol Cell 24: 603–610
Greenwell PW, Kronmal SL, Porter SE, Gassenhuber J, Obermaier B,Petes TD (1995) TEL1, a gene involved in controlling telomere lengthin S. cerevisiae, is homologous to the human ataxia telangiectasia gene.Cell 82: 823–829
Ivessa AS, Zhou JQ, Schulz VP, Monson EK, Zakian VA (2002)Saccharomyces Rrm3p, a 50 to 30 DNA helicase that promotes replicationfork progression through telomeric and subtelomeric DNA. Genes Dev16: 1383–1396
Koering CE, Fourel G, Binet-Brasselet E, Laroche T, Klein F, Gilson E (2000)Identification of high affinity Tbf1p-binding sites within the budding yeastgenome. Nucleic Acids Res 28: 2519–2526
Krauskopf A, Blackburn EH (1996) Control of telomere growth by interactionsof RAP1 with the most distal telomeric repeats. Nature 383: 354–357
Kyrion G, Boakye KA, Lustig AJ (1992) C-terminal truncation of RAP1 resultsin the deregulation of telomere size, stability, and function inSaccharomyces cerevisiae. Mol Cell Biol 12: 5159–5173
Lundblad V, Szostak JW (1989) A mutant with a defect in telomere elongationleads to senescence in yeast. Cell 57: 633–643
Marcand S, Gilson E, Shore D (1997) A protein-counting mechanism fortelomere length regulation in yeast. Science 275: 986–990
Pennock E, Buckley K, Lundblad V (2001) Cdc13 delivers separate complexesto the telomere for end protection and replication. Cell 104: 387–396
Ray A, Runge KW (1999) Varying the number of telomere-bound proteinsdoes not alter telomere length in tel1D cells. Proc Natl Acad Sci USA 96:15044–15049
Ritchie KB, Mallory JC, Petes TD (1999) Interactions of TLC1 (which encodes theRNA subunit of telomerase), TEL1, and MEC1 in regulating telomere lengthin the yeast Saccharomyces cerevisiae. Mol Cell Biol 19: 6065–6075
Sabourin M, Tuzon CT, Zakian VA (2007) Telomerase and Tel1ppreferentially associate with short telomeres in S. cerevisiae. Mol Cell 27:550–561
Singer MS, Gottschling DE (1994) TLC1: template RNA component ofSaccharomyces cerevisiae telomerase. Science 266: 404–409
Teixeira MT, Forstemann K, Gasser SM, Lingner J (2002) Intracellulartrafficking of yeast telomerase components. EMBO Rep 3: 652–659
Teixeira MT, Arneric M, Sperisen P, Lingner J (2004) Telomere lengthhomeostasis is achieved via a switch between telomerase-extendibleand -nonextendible states. Cell 117: 323–335
Tseng SF, Lin JJ, Teng SC (2006) The telomerase-recruitment domain of thetelomere binding protein Cdc13 is regulated by Mec1p/Tel1p-dependentphosphorylation. Nucleic Acids Res 34: 6327–6336
Tsukamoto Y, Taggart AK, Zakian VA (2001) The role of the Mre11–Rad50–Xrs2 complex in telomerase-mediated lengthening of Saccharomycescerevisiae telomeres. Curr Biol 11: 1328–1335
Wotton D, Shore D (1997) A novel Rap1p-interacting factor, Rif2p,cooperates with Rif1p to regulate telomere length in Saccharomycescerevisiae. Genes Dev 11: 748–760
Zhao X, Chabes A, Domkin V, Thelander L, Rothstein R (2001) Theribonucleotide reductase inhibitor Sml1 is a new target of the Mec1/Rad53 kinase cascade during growth and in response to DNA damage.EMBO J 20: 3544–3553
Tel1 activates telomerase at short telomeres
M. Arneric & J. Lingner
&2007 EUROPEAN MOLECULAR BIOLOGY ORGANIZATION EMBO reports
scientificreport
1085

Tel1 Activates Telomerase at Short Telomeres
Supplemental Data for:
Arnerić and Lingner,
Experimental Procedures
Telomere Sequencing and Analysis
Genomic DNA was extracted using the Wizard® genomic kit (Promega). Up to
100ng of total genomic DNA was used for telomere-PCR as described (Forstemann
et al, 2000). The oligonucleotide sequence to amplify tagged telomere VIIL (5’-
GCATGTATACTAAACTCACAAATT-AGAGC-3’) was kindly provided by T.M. Teixeira
(CNRS Ecole Normale Supérieure de Lyon, Lyon, France). Oligonucleotides specific
for tagged telomere VR and native telomere IL, cloning procedure, and mini-prep
plasmid preparation were as previously described (Teixeira et al, 2004). Plasmid
DNA was sequenced using M13 forward or reverse primers (Fasteris, Geneva,
Switzerland). Sequences were aligned and sequence divergence was detected using
the contig assembler of Sequencher (Gene Codes Corporation).
Statistical Analysis
Package R v1.8.1 (Ihaka & Gentleman, 1996) was used for standard
parametric and nonparametric tests. Telomeres were ordered according to their non-
diverging size and subgroups containing 15 telomeres were formed. The average
telomere size of each sub-group was calculated and the frequency of elongation
determined. The relationship between [frequency of initiation] and [mean length of
telomere of each sub-group] was fitted to the following logistic regression model:
[(frequency of elongation) ~ β0 + β1(mean length) +ε]. Several diagnostic analyses
were applied to test for lack of fit, outliers and leverage points.
Yeast Strains and Plasmids
Yeast strains are listed in Supplemental Table 1. Deletions of TEL1, MEC1 and SML1
were generated using PCR generated disruption cassettes according to (Longtine et
al, 1998). GA1415 was a gift from the S. Gasser laboratory (FMI, Basel,Switzerland).
EST1 or TLC1 were disrupted as described (Teixeira et al, 2002) to obtain

Tel1 Activates Telomerase at Short Telomeres
MAY06/est1∆ and MAY18/tlc1∆, respectively. MAY06, MAY46 and MAY47 contain
the EST1 deletion covered with plasmid pVL1120 (CEN TRP1 Est1-DBDCdc13) (Evans
& Lundblad, 1999). MAY46 and MAY47 are derivatives from W303 strain with
telomere VIIL tagged (see scheme Fig.3 and (Berthiau et al, 2006)), and express Gbd
(from plasmid pGbd) and Gbd-Tbf1N (from pGbd-Tbf1N) respectively. Original strain
and plasmids are gifts from E. Gilson laboratory (CNRS Ecole Normale Supérieure de
Lyon, Lyon, France). MAY07, GA1415, MAY37 and MAY50 were used as telomerase-
positive strains for mating (termed ‘donors’). In MAY07 and MAY37 the subtelomeric
region at chromosome IL was replaced with the URA3 marker as described (Teixeira
et al, 2004).
Supplemental References
Berthiau AS, Yankulov K, Bah A, Revardel E, Luciano P, Wellinger RJ, Geli V, Gilson E (2006) Subtelomeric proteins negatively regulate telomere elongation in budding yeast. Embo J 25: 846-856.
Evans SK, Lundblad V (1999) Est1 and Cdc13 as comediators of telomerase access. Science 286: 117-120.
Forstemann K, Hoss M, Lingner J (2000) Telomerase-dependent repeat divergence at the 3' ends of yeast telomeres. Nucleic Acids Res 28: 2690-2694.
Ihaka R, Gentleman R (1996) R: A Language for data analysis and graphics. J of Comp Graph Statistics 5: 299-314.
Longtine MS, McKenzie A, 3rd, Demarini DJ, Shah NG, Wach A, Brachat A, Philippsen P, Pringle JR (1998) Additional modules for versatile and economical PCR-based gene deletion and modification in Saccharomyces cerevisiae. Yeast 14: 953-961.
Teixeira MT, Forstemann K, Gasser SM, Lingner J (2002) Intracellular trafficking of yeast telomerase components. EMBO Rep 3: 652-659.
Teixeira MT, Arneric M, Sperisen P, Lingner J (2004) Telomere length homeostasis is achieved via a switch between telomerase- extendible and -nonextendible states. Cell 117: 323-335.

Tel1 Activates Telomerase at Short Telomeres
Supplemental Table 1. Saccharomyces cerevisiae Strains
Strain name Strain genotype Source
MAY06 MATa ura3-52 trp1-1 leu2 his3-11,15 ade2-1 can1-100 URA3::TKx7 tel1∆::KANMX6 est1∆::HIS3MX6 ADE2-(DIA5-1)telVR + pVL1120 (CEN TRP1 Est1-DBDCdc13)
this work
MAY21 MATa ura3-52 trp1-1 leu2 his3-11,15 ade2-1 can1-100 tel1∆::HISMX6 ADE2-(DIA5-1)telVR this work
MAY18 MATa ura3-52 trp1-1 leu2 his3-11,15 ade2-1 can1-100 sml1∆::KANMX6 mec1∆::TRP1 ADE2-(DIA5-1)telVR this work
MAY46 MATa ura3-52 trp1-1 leu2 his3-11,15 ade2-1 can1-100 tel1∆::LEU2 est1∆::NATMX6 URA3-(6xUASG)telVIIL + pGbd pVL1120 (CEN TRP1 Est1-DBDCdc13)
this work
MAY47 MATa ura3-52 trp1-1 leu2 his3-11,15 ade2-1 can1-100 tel1∆::LEU2 est1∆::NATMX6 URA3-(6xUASG)telVIIL + pGbd-Tbf1N pVL1120 (CEN TRP1 Est1-DBDCdc13)
this work
MAY07 MATα ura3-∆851 trp1∆63 his3-∆200 tel1∆::TRP1 SEO1-URA3 tel1L∆ this work
GA1415 MATα ura3-52 trp1 leu2 his3-11 ade2 can1∆ sml1∆::KANMX6 mec1∆::TRP1
Gasser strain collection
MAY37 MATα ura3-∆851 trp1∆63 his3-∆200 sml1∆::KANMX6 mec1∆::TRP1 SEO1-URA3 tel1L∆ this work
MAY50 MATα his3-∆1 leu2-∆0 ura3-∆0 tel1∆::NATMX6 this work
MAY23 diploid GA1415xMAY21 this work

Tel1 Activates Telomerase at Short Telomeres

Tel1 Activates Telomerase at Short Telomeres


Chapter 4
Telomerase Repeat Addition Processivity is Increased at
Critically Short Telomeres in a Tel1-Dependent Manner in
Saccharomyces cerevisiae
Article published in Genes & Development, vol. 21. pp. 2485-2494, October 1st, 2007


Regulation of Telomerase Processivity
87
Budding yeast telomerase appears to be nonprocessive in vitro (Cohn & Blackburn,
1995; Lingner et al, 1997b). However, in vivo analysis of telomere elongation
presented in Chapter 2 showed that telomerase is able to add much more than just
one telomeric repeat on a single telomere in one cell cycle. Thus, in vivo, the enzyme
is either processive or it turns over very rapidly. To measure telomerase processivity
in vivo, we took advantage of a modified STEX system in which telomere elongation
is monitored in the presence of both a wild-type TLC1 and a mutant template tlc1tm,
a setup named the two-template single-telomere extension (2T-STEX) assay.
I contributed to research in Chapter 4 by performing 2T-STEX analysis in a
tel1Δ background, establishing that telomerase processivity, which is observed at
short telomeres in vivo is Tel1-dependent.


Telomerase repeat addition processivityis increased at critically short telomeresin a Tel1-dependent manner inSaccharomyces cerevisiaeMichael Chang, Milica Arneric, and Joachim Lingner1
Swiss Institute for Experimental Cancer Research (ISREC), CH-1066 Epalinges, Switzerland; École Polytechnique Fédéralede Lausanne (EPFL), CH-1015 Lausanne, Switzerland; National Centre of Competence in Research (NCCR) “Frontiersin Genetics,” CH-1211 Geneva, Switzerland
Telomerase is the ribonucleoprotein enzyme that elongates telomeres to counteract telomere shortening.The core enzyme consists of a reverse transcriptase protein subunit and an RNA subunit. The RNA subunitcontains a short region that is used as a template by the reverse transcriptase to add short, tandem, G-richrepeats to the 3� ends of telomeres. By coexpressing two RNA subunits that differ in the telomeric repeatsequence specified and examining the telomere extensions after one cell cycle, we determined thatSaccharomyces cerevisiae telomerase can dissociate and reassociate from a given telomere during one cellcycle. We also confirmed that telomerase is nonprocessive in terms of telomeric repeat addition. However,repeat addition processivity is significantly increased at extremely short telomeres, a process that is dependenton the ATM-ortholog Tel1. We propose that this enhancement of telomerase processivity at short telomeresserves to rapidly elongate critically short telomeres.
[Keywords: Telomerase; processivity; Saccharomyces cerevisiae; TLC1; Tel1]
Supplemental material is available at http://www.genesdev.org.
Received June 29, 2007; revised version accepted August 10, 2007.
Telomeres, the physical ends of eukaryotic chromo-somes, protect chromosome ends from end fusions anddegradation (Ferreira et al. 2004). Telomere length ismaintained by a dynamic process of lengthening andshortening (Hug and Lingner 2006). Shortening can occuras a result of nucleolytic degradation and incompleteDNA replication. Lengthening is primarily accom-plished by the action of a specialized reverse transcrip-tase called telomerase, whose core consists of a catalyticprotein subunit and an RNA subunit (Greider and Black-burn 1985, 1987, 1989; Lingner et al. 1997). Telomeraseextends telomeres by repeated reverse transcription of ashort sequence, using the RNA subunit as a template, tothe 3� ends of telomeres (Greider and Blackburn 1989; Yuet al. 1990; Singer and Gottschling 1994). Lagging strandsynthesis then presumably fills in the complementary 5�strand. Cells lacking telomerase have telomeres thatshorten with each cell division, which can ultimatelylead to replicative senescence (Lundblad and Szostak1989; Harley et al. 1990; Yu et al. 1990).
Telomerase complexes isolated from ciliated protozoaand humans are processive in vitro, capable of elongatinga given telomeric primer by hundreds of nucleotides
through multiple rounds of telomeric repeat addition(Morin 1989; Greider 1991). In contrast, in other organ-isms such as mouse, Chinese hamster, and various fungi,telomerase appears to be less or nonprocessive in vitro(Prowse et al. 1993; Cohn and Blackburn 1995; Lingner etal. 1997; Lue and Peng 1997, 1998; Fulton and Blackburn1998). The budding yeast Saccharomyces cerevisiaetelomerase enzyme remains stably bound to the telo-mere substrate after a single round of telomere elonga-tion in vitro (Cohn and Blackburn 1995; Prescott andBlackburn 1997a). However, in vivo analysis suggeststhat yeast telomerase can add >100 nucleotides (nt) in asingle cell cycle (Teixeira et al. 2004). Thus, in vivo, theenzyme is either processive or it turns over very rapidly.
In vivo analysis in S. cerevisiae has also shown thattelomerase does not act on every telomere in each cellcycle but instead exhibits an increasing preference fortelomeres as their lengths decline, suggesting that telo-meres switch between nonextendible and extendiblestates (Teixeira et al. 2004). Although the number ofnucleotides added per extension event does not signifi-cantly correlate with telomere length, telomere exten-sion length is strongly increased at very short telomeres(Teixeira et al. 2004). However, it was not determinedwhether or not these long extension events were due toprocessive telomerase action.
1Corresponding author.E-MAIL [email protected]; FAX 41-21-652-69-33.Article is online at http://www.genesdev.org/cgi/doi/10.1101/gad.1588807.
GENES & DEVELOPMENT 21:2485–2494 © 2007 by Cold Spring Harbor Laboratory Press ISSN 0890-9369/07; www.genesdev.org 2485

To analyze telomerase repeat addition processivity invivo, and to determine whether telomere length affectsprocessivity, we coexpressed a mutant tlc1 that specifiesaltered telomeric sequences together with wild-typeTLC1 and analyzed telomere elongation events after onecell cycle. This analysis indicated that telomerase canuse both RNA templates to extend a telomere during onecell cycle, and that the telomerase complex can dissoci-ate and reassociate with telomeres during one round oftelomere elongation. We found that telomerase does gen-erally act nonprocessively in vivo, consistent with con-clusions drawn from a previous study (Prescott andBlackburn 1997b). However, we found that the repeataddition processivity was significantly enhanced at ex-tremely short telomeres. We propose that this enhance-ment of processivity, which is dependent on the ATMortholog Tel1, allows cells to rapidly elongate criticallyshort telomeres that can arise as a result of a double-strand break caused by DNA damage or DNA replicationfork collapse in the telomeric tracts.
Results
Characterization of a tlc1 template mutant
To study telomerase action in cells expressing two dif-ferent telomerase RNA subunits, we needed a mutanttlc1 that incorporated telomeric repeats that were dis-tinguishable from wild type, yet would still be able tocomplement a deletion of TLC1 and elongate telomeresto similar levels as wild type. We previously screened anS. cerevisiae tlc1 library, in which 10 of the 16 templat-ing nucleotides were randomized, for complementation
of a TLC1 deletion (Forstemann et al. 2003). Out of anestimated 60,000 templates screened, only 40 differentmutant templates were able to rescue the senescencephenotype of tlc1� (Forstemann et al. 2003). One of thetemplate mutations was isolated three times indepen-dently and phenotypically most closely resembled wildtype (Forstemann et al. 2003). We called this templatemutant tlc1-tm. The template region of tlc1-tm is 3�-CACCUAAACCACACAC-5�, whereas it is 3�-CACACACCCACACCAC-5� for wild-type TLC1.
Wild-type TLC1 was replaced at its endogenous locusby tlc1-tm. After propagating this strain for >100 genera-tions to allow mutant telomeric sequences to be incor-porated into telomeres, we performed telomere PCR toamplify telomere VI-R sequences (Fig. 1A). The lengthsof the tlc1-tm telomeres were ∼80 base pairs (bp) shorterthan those of the isogenic wild-type strain. Despite themodestly shorter telomeres, telomere length homeosta-sis was maintained, confirming what we reported previ-ously for this mutant when it was expressed from a plas-mid in a different genetic background (Forstemann et al.2003).
The amplified VI-R telomeres from the tlc1-tm mu-tant were cloned and sequenced. Analysis of these se-quences revealed that telomerase using this templateRNA added degenerate repeats with a consensus se-quence of [(TG)0–4TGG]nATTTGG, where n indicatesthat the [(TG)0–4TGG] motif can be repeated for a vari-able number of times before the incorporation of anATTTGG sequence (Fig. 1B; Supplementary Fig. S1).Mutant sequences could easily be differentiated fromwild-type sequences because of the frequent usage of theGG dinucleotide. Wild-type telomeres, with a consensus
Figure 1. Characterizing the tlc1-tm mutant. (A) Telomere lengths of the indicated strains were determined by telomere PCR oftelomere VI-R. The size of the PCR product is the sum of the telomere length, the distance of the subtelomeric primer to the beginningto the telomeric tracts, and the size of the primers used in the telomere PCR (59 bp). Telomere lengths were calculated to be 353 bpand 271 bp for TLC1 and tlc1-tm strains, respectively. The calculated telomere lengths are an average of telomere lengths from twodifferent isolates of each strain. (B) Telomere PCR products from A amplified from the tlc1-tm mutant were cloned and sequenced.Three representative sequences are shown. The complete set of sequences is shown in Supplementary Figure 1. Mutant divergentregions of each sequence (in red) were used to derive a mutant consensus sequence of [(TG)0–4TGG)]nATTTGG. Bolded sequenceshighlight occurrences of GGG trinucleotide incorporation by the mutant telomerase.
Chang et al.
2486 GENES & DEVELOPMENT

sequence of (TG)0–6TGGGTGTG(G)0–1 (Forstemann andLingner 2001), very rarely contain two consecutive GGdinucleotide sequences without an intervening GGG tri-nucleotide. GGG trinucleotides are rarely found in mu-tant repeat sequences, whereas they are present in everywild-type repeat incorporated. Furthermore, the occa-sional incorporation by the mutant telomerase ofATTTGG is a clear indication of mutant telomeric se-quence.
Since the tlc1-tm template region is 3�-CACCUAAACCACACAC-5�, the mutant telomerase should not beable to incorporate GGG trinucleotides according to ca-nonical Watson-Crick base-pairing. The rare occurrencesof GGG trinucleotides in mutant sequences (Fig. 1B;Supplementary Fig. S1) are most likely due to the U resi-due in the template being able to base-pair with Gnucleotides (Crick 1966; Varani and McClain 2000). Thispotentially poses a problem for differentiating mutantfrom wild-type sequences. However, some of the GGGtrinucleotide occurrences in the mutant sequences werestill easily identifiable as mutant (e.g., in cases where thesequence was GGGTTT or GGGTGG, neither of whichoccurs in wild-type telomeric sequences). The remainingGGG trinucleotide occurrences were very rare (twotimes in 1373 nt of mutant telomeric sequence, or 217mutant repeats, from a tlc1-tm mutant). Hence the tlc1-tm mutant added telomeric repeats that were easily dis-tinguishable from wild-type repeats and therefore suit-able for use in our coexpression assays.
Two-template single-telomere extension assay(2T-STEX)
Although much work has been done to characterizetelomerase activity, it is not known whether a telomere,if extended, is extended by only one telomerase complexonce per cell cycle, or whether telomerase complexescan dissociate and reassociate from telomeres while ex-tending a given telomere. To answer this question, weused a modified version of the STEX assay that we de-veloped and described previously (Teixeira et al. 2004;Arneric and Lingner 2007). In the STEX assay, a telo-merase-negative strain (referred to as the “recipient”)was created by deleting EST1, which encodes an essen-tial subunit of yeast telomerase (Lundblad and Szostak1989). Telomerase was added by mating this strain to atelomerase-positive “donor” strain that contains wild-type EST1. Thus, telomeres that had shortened in therecipient strain are extended in the zygote. Telomereelongation was detected by cloning and sequencing oftelomeres derived from the recipient parental strain afterbeing amplified by telomere PCR, which is made pos-sible because the recipient chromosome V-R telomerewas tagged with the ADE2 gene in the subtelomeric re-gion (Forstemann et al. 2000; Teixeira et al. 2004). Sinceyeast telomerase adds imperfect repeats, telomere elon-gation can be detected by the divergence of telomericsequences in the telomere-distal region relative to thetelomeric DNA synthesized by semiconservative DNA
replication (Wang and Zakian 1990; Forstemann et al.2000; Forstemann and Lingner 2001).
We modified the STEX assay by replacing wild-typeTLC1 with tlc1-tm at its endogenous locus in the donorstrain. We named this modified assay 2T-STEX (Fig. 2A).This strain was mated to a recipient strain that expresseswild-type TLC1. Telomere extensions were analyzed 3 hafter mating (Fig. 2B; Supplementary Fig. S2A), the timerequired for most cells to mate and complete one S phase(Teixeira et al. 2004). Of 21 telomere elongation events,11 contained only wild-type sequence and three con-tained only mutant sequence. Seven contained a mixtureof wild-type and mutant sequences.
We also performed the reciprocal experiment wherethe donor strain expressed wild-type TLC1 while the re-cipient expressed the tlc1-tm mutant (SupplementaryFig. S2B). The results were very similar. Of 23 telomereelongation events, 11 contained only wild-type se-quence, four contained only mutant sequence, and eightcontained a mixture of both wild-type and mutant se-quences. In both experiments, the short telomeres werepreferentially elongated and long extension events oc-curred at very short telomeres (data not shown) as ex-pected (Teixeira et al. 2004). Together, these results in-dicate that telomerase can use both templates to extendthe same telomere during one cell cycle.
We also performed a 2T-STEX assay using an est1�telomerase-negative donor strain to control for telomer-ase-independent divergence events (Fig. 2C). Instead oflimiting cells to one cell cycle, they were allowed topropagate for ∼30 generations after mating. Three of 38(7.9%) telomeres sequenced showed telomerase-inde-pendent divergence events. This corresponds to an ex-tremely low frequency of divergence after one cell cycle,similar to what we reported previously (Teixeira et al.2004). Thus, the sequence divergence events we ob-served in our 2T-STEX assays were due to telomerase-mediated telomere elongation events.
Yeast telomerase is generally nonprocessive for repeataddition except at extremely short telomeres
The average extension length for S. cerevisiae telomer-ase during one cell cycle is 44 nt for telomeres >200 bp inlength (Teixeira et al. 2004). Since the mean telomericrepeat length is 11 nt (Forstemann and Lingner 2001),yeast telomerase in vivo can extend telomeres by aboutfour repeats, which is in contrast with its in vitro bio-chemical properties, where it can only add one repeatbefore stopping (Cohn and Blackburn 1995; Prescott andBlackburn 1997b). Therefore, either yeast telomerase hassome in vivo processivity or it is able to dissociate fromand reassociate to telomeres within a single cell cycle.Our 2T-STEX results suggest that the latter model ispossible (Fig. 2A; Supplementary Fig. S2). However, thisdoes not necessary imply that telomerase is completelynonprocessive in vivo.
To determine whether telomerase has any repeat ad-dition processivity in vivo, we considered the scenariowhere telomerase is completely nonprocessive—i.e., if it
Regulation of telomerase processivity
GENES & DEVELOPMENT 2487

stops after each repeat added. We counted the total num-ber of repeats added in all extensions from our 2T-STEXassays and determined the fraction of wild-type repeatsused (207/295 = 0.702). This fraction did not change sig-nificantly as a function of telomere length (data notshown). Given a 70.2% incorporation of wild-type re-peats, the expected average number of consecutive wild-type repeats per stretch or run of wild-type repeatsshould be 3.3, assuming that telomerase is completelynonprocessive in vivo (see Materials and Methods for themathematical equation used to determine this value). Ifyeast telomerase is processive, the observed averagenumber of wild-type repeats per run should be higherthan 3.3. However, the observed average for telomeres>125 bp in length was 3.2, a value that did not changesignificantly as a function of telomere lengths >125 bp,indicating that yeast telomerase is indeed nonprocessivein vivo. Strikingly, repeat addition processivity was sig-nificantly increased at telomeres <125 bp in length, asthe observed average number of wild-type repeats per run
was 7.6 (Fig. 3). This effect was also seen for telomeraseusing the mutant tlc1-tm, where the average number ofmutant repeats per run of mutant repeats increased from2.1 to 5.9 (Fig. 3). Thus, the increased extension length atvery short telomeres (Teixeira et al. 2004) can be attrib-uted to the increased telomerase repeat addition proces-sivity (Fig. 3).
Telomeres <125 bp in length do arise in vivo
In the absence of telomerase, S. cerevisiae telomeresshorten at a rate of 3 bp per generation (Marcand et al.1999). Thus, starting with a wild-type-length telomere of300 bp in length, it would take ∼58 generations [(300–125)/3 = 58.3] to shorten to a length of 125 bp if telom-erase did not act in the intervening time. The frequencyof telomere extension by telomerase steadily increasesfrom ∼7% at 300-bp-long telomeres to ∼39% at 125-bp-long telomeres (Teixeira et al. 2004). Consequently, theprobability of a telomere shortening from 300 bp to 125
Figure 2. 2T-STEX analysis. (A) 2T-STEX methodology (see text for details). (B) Example of aligned sequences from a 2T-STEX assayin which the donor strain carried the tlc1-tm allele. Telomere extension events were detected by sequence divergence. Sequences inblue are repeats added by telomerase with wild-type TLC1, while sequences in red are repeats added by telomerase with the mutanttlc1-tm. (C) Analysis of a 2T-STEX assay in which both the recipient and the donor strains were est1�. Unlike a standard 2T-STEXassay, the diploids were allowed to undergo ∼30 cell doublings after mating before sequence analysis. Each telomere sequence isrepresented by a vertical bar where the red and blue parts indicate the nondiverging and diverging regions, respectively. This analysisshows that sequence divergence in the absence of telomerase is very low and not perturbing the 2T-STEX analysis.
Chang et al.
2488 GENES & DEVELOPMENT

bp without telomerase acting on it in the interveningtime is 2.6 × 10−7. Therefore, it is highly improbable thata wild-type-length telomere could shorten to 125 bp inlength via normal telomere erosion mechanisms such asincomplete DNA replication or nucleolytic degradation.
To detect whether or not telomeres <125 bp in lengthactually occur in vivo, we sequenced telomeres fromtelomerase-positive cells after ∼40 generations of clonalexpansion. While all yeast telomeric repeats are degen-erate, only the distal region of telomeres have sequencesthat are divergent (Wang and Zakian 1990; Forstemannet al. 2000). This sequence divergence is dependent ontelomerase activity (Forstemann et al. 2000). The averagepoint of divergence in a wild-type strain is 214 bp intothe telomeric tract, counting away from the centromere(Fig. 4). A rad52� strain was included in this experimentto eliminate any possibility of recombination-dependentdivergence events. The rad52� strain had an averagepoint of divergence of 253 bp into the telomeric tract.The slightly greater point of divergence in the rad52�strain can be attributed to the slightly longer telomeresseen in this strain (27 wild-type telomeres sequencedwith an average length of 291 bp compared with 17rad52� telomeres sequenced with an average length of348 bp). Although the average points of divergence inboth strains were much >125 bp, both strains containedtelomere sequences in which the points of divergencewere before 125 bp into the telomeric tract (three of 27sequences for wild type; seven of 17 sequences for
rad52�). Taken together, this indicates that telomeres<125 bp in length do occur in vivo and most likely do notarise from normal telomere shortening processes.
Tel1 is needed for the enhanced repeat additionprocessivity at extremely short telomeres
The phosphoinositide 3-kinase-related kinases Tel1 andMec1 (S. cerevisiae orthologs of human ATM and ATR,respectively) are required for maintaining telomerelength homeostasis, as simultaneous deletion of bothTEL1 and MEC1 results in progressive shortening of telo-meres leading to cellular senescence (Ritchie et al. 1999).Cells lacking MEC1 have slightly shortened telomeres,while cells lacking TEL1 have very short but stable telo-meres (Greenwell et al. 1995; Ritchie et al. 1999), sug-gesting that Tel1 has the major role in telomere lengthregulation. To test if Tel1 is important for makingtelomerase become more processive at telomeres <125bp in length, we performed the 2T-STEX assay in a tel1�background (Fig. 5). The percentage of wild-type repeatsused was 62.5% (25 of 40). For this percentage, the ex-pected average number of wild-type repeats per run ofwild-type repeats should be 2.7 if telomerase was com-pletely nonprocessive. At telomeres <125 bp in length,the observed average number of wild-type repeats per runwas 2.6, indicating that in cells lacking TEL1, telomeraseis nonprocessive even at telomeres <125 bp in length.Accordingly, we found that the increase in telomere ex-tension lengths at telomeres <125 bp in length was notobserved in a tel1� background (Fig. 5). Regular “one-
Figure 4. Telomeres <125 bp in length do occur in vivo. ARAD52/rad52� heterozygous diploid was sporulated and tetradwas dissected. Wild-type or rad52� isolates derived from thesame tetrad were allowed to propagate for ∼40 generations.Chromosome VI-R telomeres were amplified by telomere PCR,cloned, and sequenced. Each telomere sequence is representedby a vertical bar where the red and blue parts indicate the non-diverging and diverging regions, respectively.
Figure 3. Telomere extension length (top panel) correlateswith repeat addition processivity (bottom panel). The averagelengths of the telomere extensions from the 2T-STEX assayswere determined for telomeres greater than and less than 125 bpin length. Repeat addition processivity is shown as the averagenumber of wild-type repeats added per run of wild-type repeats(blue bars) or the average number of mutant repeats added perrun of mutant repeats (red bars) for telomeres greater than andless than 125 bp in length. The P-values associated with two-tailed unpaired t-tests are shown.
Regulation of telomerase processivity
GENES & DEVELOPMENT 2489

template” STEX assays (in which both the donor andrecipient cells carry wild-type copies of TLC1) performedin a tel1� background also lacked long extension eventsat short telomeres (Arneric and Lingner 2007). Thechange in repeat addition processivity at short telomeresas measured by the average number of wild-type repeatsper run (∼2.9-fold; 7.6 repeats for wild type vs. 2.6 repeatsfor te1l�) mirrored the change in the average telomereextension length (threefold; 102 nt for wild type vs. 34 ntfor tel1�) (Fig. 5). Thus, the inability of telomerase to addlong telomeric extensions on extremely short telomeresin tel1� mutants is a result of its inability to act proces-sively at these telomeres.
Discussion
In this study, we showed that S. cerevisiae telomerasecomplexes can dissociate and reassociate to extend agiven telomere during one cell cycle. We demonstratedthat telomerase acts nonprocessively in repeat additionexcept at extremely short telomeres (i.e., telomeres <125bp in length). We determined that, although these telo-meres do not likely arise from normal telomere erosionevents such as incomplete DNA replication or nucleo-lytic degradation, these telomeres do occur in vivo. Fur-thermore, we found that the enhancement of repeat ad-dition processivity at extremely short telomeres is me-diated by Tel1.
Telomerase can dissociate and reassociate to extenda given telomere during one cell cycle
Previous work (Singer and Gottschling 1994; Prescottand Blackburn 1997b; Henning et al. 1998; Lin et al.
2004) and the work reported here clearly demonstratethat mutations in the templating region of TLC1 can becopied into telomeric DNA in vivo. By coexpressing atlc1 template mutant along with wild-type TLC1 andanalyzing telomere extension events after one cell cycle,we found that some extensions contained both wild-typeand mutant repeats, indicating that both RNA templateswere used to extend these telomeres. This suggests thattelomerase complexes can dissociate and reassociate toextend a given telomere during one cell cycle.
Est2, the reverse transcriptase protein subunit oftelomerase (Lingner et al. 1997; Counter et al. 1997), isfound at telomeres throughout the cell cycle, whereasEst1 only associates with telomeres in late S and G2phases, when telomeres are lengthened by telomerase(Taggart et al. 2002). Since our data show that telomerasecan dissociate and reassociate from telomeric 3� ends,the localization of Est2 to telomeres is likely dynamic. Inother words, telomerase core complexes are unlikely tobe sitting statically at telomeres waiting for Est1 to ar-rive and activate them in late S phase. Indeed, the obser-vation from our 2T-STEX assays that some telomere ex-tension events began with the incorporation of mutantsequence in an assay using a wild-type TLC1 recipient orwild-type sequence in an assay using a mutant tlc1-tmrecipient support the argument that telomerase com-plexes can and do dissociate and reassociate from telo-meres, both before they act on telomeres and while telo-meres are being extended.
As it has been shown that yeast telomerase is likely adimer or multimer (Prescott and Blackburn 1997a,b), it isformally possible that the telomere extensions contain-ing a mixture of wild-type and mutant sequences are theresult dimeric or multimeric telomerase complexes,with each telomere extension event carried out by onlyone telomerase complex that is capable of using differentRNA templates present in the complex. We are not infavor of this model because the extensions containing amixture of wild-type and mutant sequences do not revealany discernable pattern that would be indicative of aregular intracomplex template switching mechanism(Fig. 2; Supplementary Fig. S2).
Telomerase is nonprocessive except at extremely shorttelomeres
Previous work by Prescott and Blackburn (1997b) alsoanalyzed telomerase processivity by coexpressing wild-type TLC1 and various template-mutated tlc1. Sequenc-ing telomeres from the TLC1/tlc1 heterozygous diploidsafter extensive clonal expansion, Prescott and Blackburn(1997b) found that the mutant repeats were not presentas long stretches; instead, they occurred most often ei-ther singly or in pairs. Prescott and Blackburn (1997b)suggested that the most likely interpretation for this pat-tern is that the mutant enzymes acted nonprocessively.Furthermore, some of mutant repeats were separated bysingle wild-type repeats, indicating that the wild-typetelomerase was likely similarly nonprocessive. How-ever, since their study did not analyze STEX events after
Figure 5. Tel1 is needed for enhanced repeat addition proces-sivity at critically short telomeres. (Top panel) The averagelengths of the telomere extensions from the 2T-STEX and tel1�
2T-STEX assays were determined for telomeres <125 bp inlength. (Bottom panel) Repeat addition processivity is shown asthe average number of wild-type repeats added per run of wild-type repeats for telomeres <125 bp in length. The P-values as-sociated with two-tailed unpaired t-tests are shown.
Chang et al.
2490 GENES & DEVELOPMENT

one cell cycle, Prescott and Blackburn (1997b) did notexclude the possibility that the mixed wild-type and mu-tant repeat patterns were the result of processive elon-gation by telomerase, with successive bursts of elonga-tion separated by degradation of newly added telomericrepeats, generated over many cell cycles. Since our studyanalyzes extension events during one cell cycle, we con-clusively showed that telomerase does indeed act non-processively in vivo at telomeres that are >125 bp inlength.
The Prescott and Blackburn (1997b) study was also notable to assess telomerase processivity as a function oftelomere length. Our work demonstrated that whiletelomerase acts nonprocessively on telomeres >125 bp inlength, it possesses significant repeat addition processiv-ity when acting on telomeres <125 bp in length. We pro-pose that this enhancement of processivity is a mecha-nism that cells use to rapidly elongate critically shorttelomeres. We showed previously that the frequency oftelomere extension decreases steadily as a function oftelomere length. Thus, cells have two ways to respond tocritically short telomeres—an increase in frequency ofextension coupled with an increase in telomerase pro-cessivity. While the increase in repeat addition proces-sivity is only approximately two- to threefold, thiswould correspond to extension lengths of ∼88–132 nt,given that the average extension length during one cellcycle is 44 nt for telomeres >200 bp in length (Teixeira etal. 2004), which is quite significant considering thatwild-type yeast telomeres are only ∼300 bp in length.
Although both the frequency of extension and telo-merase processivity change with telomere length, therelationship of each to telomere length is different.While the extension frequency steadily increases as telo-meres get shorter, repeat addition processivity is bimo-dal with respect to telomere length. Telomerase is gen-erally nonprocessive at all telomeres greater than ∼125bp in length but processive on telomeres <125 bp inlength. This agrees with the bimodal distribution seenfor telomere length extension, where much longer exten-sions are observed at very short telomeres (Teixeira et al.2004). Telomerase repeat addition processivity, unliketelomere extension frequency, does not steadily increasewith decreasing telomere length. Hence, although an in-crease in both extension frequency and telomerase pro-cessivity occurs at critically short telomeres, the in-crease in processivity may occur specifically to rapidlyelongate these telomeres.
One potential caveat of this study is that it assumesthat there is an excess of telomerase of both templatesequences available at every telomere. If this assumptionis incorrect, there may not be free competition of the twotelomerases in our 2T-STEX assays to extend a giventelomere. Consequently, the same telomerase wouldhave an advantage to rebind and extend the same telo-mere. As a result, the calculated processivity, as mea-sured by the average number of wild-type repeats per runof wild-type repeats, could be viewed as only a maxi-mum estimate. Indeed, it has been reported that telo-mere length homeostasis requires that telomerase levels
are limiting in cultured human cells (Cristofari and Ling-ner 2006), and that telomerase is present in low abun-dance in yeast (Mozdy and Cech 2006). Our calculatedprocessivity for telomerase is extremely low at telo-meres >125 bp in length, so even this “maximum” esti-mate suggests a nonprocessive enzyme. At a telomere<125 bp in length, it is formally possible that one telom-erase enzyme undergoes several cycles of single repeatadditions separated by periods where the enzyme has“dissociated” from the telomeric 3� end, but stayingwithin its vicinity, yielding a pattern of repeats that re-sembles processive repeat addition. However, since theconcept of repeat addition processivity is unique to thetelomerase enzyme, and given that the molecularmechanisms behind processive repeat addition by telo-merase from any organism has not yet been elucidated, itis not possible to discount this mode of action as proces-sive repeat addition, especially considering that in anymodel for processive repeat addition, after the extensionof the telomeric 3� end, the new 3� end must be “disso-ciated” from the active site of the telomerase enzyme inorder for it to be translocated into a position where theenzyme can once again extend it. Repeat addition pro-cessivity is the ability of one telomerase enzyme to syn-thesize more than one repeat at one telomere, an abilitythat we have shown in this study that yeast telomerasepossesses at extremely short telomeres.
Critically short telomeres do not occur due to normaltelomere erosion
As discussed in the Results section, it is highly improb-able for a yeast telomere to shorten to 125 bp or less inlength through telomere erosion caused by incompleteDNA replication or nucleolytic degradation. However,we showed that these extremely short telomeres do oc-cur in vivo. Such telomeres likely arise from double-strand DNA breaks caused by DNA damage or the col-lapse of DNA replication forks. Indeed, it is well estab-lished that telomeric repeats are difficult to replicate andresult in increased replication fork pausing events (Coo-per et al. 1997; Ivessa et al. 2002; Crabbe et al. 2004;Makovets et al. 2004; Fouche et al. 2006). If a replicationfork were to collapse while traversing the telomerictracts, there would be no replication origin distal to thesite of fork collapse to generate a fork to finish replica-tion of the telomere resulting in a truncated telomere. Aprocessive telomerase may then be essential to rapidlyelongate a critically shortened truncated telomere.
The function of Tel1 in telomere length homeostasis
Cells lacking TEL1 have very short, but stable, telomeres(Greenwell et al. 1995; Ritchie et al. 1999). Tel1 is able toin vitro phosphorylate Cdc13 (Tseng et al. 2006), asingle-stranded telomere-binding protein involved inboth telomere protection and telomerase recruitment(Garvik et al. 1995; Lin and Zakian 1996; Nugent et al.1996; Chandra et al. 2001; Pennock et al. 2001). Further-
Regulation of telomerase processivity
GENES & DEVELOPMENT 2491

more, Tel1 has been shown recently to specifically asso-ciate with short telomeres (Bianchi and Shore 2007; Sa-bourin et al. 2007) and is needed for the preferential bind-ing of Est1 and Est2 to short telomeres (Goudsouzian etal. 2006; Sabourin et al. 2007). Thus, Tel1 appears to beimportant for the recruitment of telomerase to telo-meres. Consistent with this view, we recently demon-strated that cells lacking TEL1 have a greatly decreasedfrequency of telomerase-mediated telomere elongation(Arneric and Lingner 2007).
In addition, at an artificial telomere lacking subtelo-meric repeats, telomerase is unable to mediate the pref-erential elongation of short telomeres by telomerase in astrain lacking TEL1 (Arneric and Lingner 2007). In thisstudy, we found that Tel1 is also important for increas-ing telomerase processivity at critically short telomeres.Thus, Tel1 is a major regulator in telomere length ho-meostasis, affecting multiple aspects of telomerase ac-tivity including telomerase recruitment, telomere lengthdetermination, and telomerase repeat addition processiv-ity. Since occurrences of telomeres that are <125 bp inlength should be relatively infrequent, the reduced fre-quency of elongation, rather than the reduced processiv-ity, is likely the primary cause of the shortened telo-meres of tel1� cells.
Interestingly, the frequency of telomere extension at anatural telomere is still regulated in a length-dependentmanner even in the absence of Tel1 (Arneric and Lingner2007). This can be attributed to a redundant mechanismfor telomerase regulation that is dependent on the sub-telomeric region. Curiously, although our tel1� 2T-STEX assayed extension events at an artificial telomerelacking subtelomeric repeats, the extension length de-fect at short telomeres in tel1� mutants is also present ata natural telomere (Arneric and Lingner 2007), suggest-ing that repeat addition processivity is not affected bythe presence or absence of a subtelomeric region. Hence,this redundant mechanism for telomerase regulationseems to only affect the frequency of telomere extension,not extension length and telomerase processivity.
It will be of considerable interest to know if these mul-
tiple functions of Tel1 are separable through the phos-phorylation of different substrates. Although Cdc13 isthe only known substrate for the Tel1 kinase so far(Tseng et al. 2006), it is likely that other substrates existand phosphorylation of these different substrates maymediate the different aspects of Tel1 function. SinceTel1 has been found recently to be enriched at shorttelomeres (Bianchi and Shore 2007; Sabourin et al. 2007),it is likely that the phosphorylation of these substratesoccur specifically at short telomeres. Some of these sub-strates may have been identified by a recent proteomicstudy to identify Mec1 and Tel1 targets (Smolka et al.2007). In particular, the telomeric protein Rif1 wasamong the substrates identified (Smolka et al. 2007).Phosphorylation of Rif1 by Tel1 may mediate some ofthe functions of Tel1 at telomeres. Clearly, much workstill needs to be done to elucidate the mechanisms be-hind telomere length homeostasis and the role of Tel1 inthese processes.
Materials and methods
Yeast strains and media
Yeast strains used in this study are listed in Table 1. Standardyeast media and growth conditions were used (Sherman 1991).Mating of yeast cells was performed essentially as described(Teixeira et al. 2004).
Telomere sequencing and analysis
Telomere V-R PCR, cloning, and sequence analysis were per-formed as described (Forstemann et al. 2000; Teixeira et al.2004). For amplifying telomeres of chromosome VI-R, a primerspecific for the subtelomeric sequence of this chromosome wasused (5�-ACGTGTGCGTACGCCATATCAATATGC-3�). Se-quencing was performed by Fasteris SA.
Expected average number of repeats per run
Assuming that telomerase is completely nonprocessive at add-ing repeats, the expected average number of wild-type repeats
Table 1. Yeast strains used in this study
Strain name Genotype Source
BY4741 MATa his3�1 leu2�0 ura3�0 met15�0 Brachmann et al. 1998
YBL39 MAT� his3�1 leu2�0 ura3�0Brian Luke (ISREC,
Epalanges, Switzerland)MCY400 MATa TLC1�kanMX4 leu2�0 ura3�0 met15� This studyMCY407 MATa tlc1-tm�kanMX4 leu2�0 ura3�0 met15� This studyMCY416 MAT� tlc1-tm�kanMX4 leu2�0 ura3�0 This studyMCY418 MATa est1��HIS3 URA3�TKx7 DIA5-1 ade2 can1� his3-11 leu2-3,112 ura3-1 trp1-1
kar1�15This study
MCY430 MATa tlc1-tm�kanMX4 est1��HIS3 URA3�TKx7 DIA5-1 ade2 can1� his3-11leu2-3,112 ura3-1 trp1-1 kar1�15
This study
MCY444 his3�1 leu2�0 ura3�0 This studyMCY445 rad52��kanMX4 his3�1 leu2�0 ura3�0 This studyMCY456 MAT� tel1��natMX4 tlc1-tm�kanR tel1L�URA3 This studyMAY06 MATa tel1��kanMX4 est1��HIS3 URA3�TKx7 DIA5-1 ade2 can1� his3-11 leu2-3,112
ura3-1 trp1-1 kar1�15This study
Chang et al.
2492 GENES & DEVELOPMENT

per run of wild-type repeats from the 2T-STEX events was de-termined using the following equation:
�n=0
�
�n + 1� pn
�n=0
�
pn
,
where p is the probability of telomerase adding a wild-type re-peat, which, for a nonprocessive telomerase, would be equiva-lent to the fraction of added repeats that have wild-type se-quence.
Acknowledgments
We thank Alessandro Bianchi for providing the telomere VI-Rprimer sequence for telomere PCR and Brian Luke for construc-tive comments on the manuscript. M.C. was supported by anEMBO Long-term Fellowship and a Human Frontiers ScienceProgram Long-term Fellowship Award. This work was sup-ported by the Swiss National Science Foundation, the SwissCancer League, the Human Frontier Science Program, and theEU Sixth Framework Programme.
References
Arneric, M. and Lingner, J. 2007. Tel1p kinase and subtelomerebound Tbf1p mediate preferential elongation of short telo-meres by telomerase in yeast. EMBO Rep. (in press).
Bianchi, A. and Shore, D. 2007. Increased association of telom-erase with short telomeres in yeast. Genes & Dev. 21: 1726–1730.
Brachmann, C.B., Davies, A., Cost, G.J., Caputo, E., Li, J., Hi-eter, P., and Boeke, J.D. 1998. Designer deletion strains de-rived from Saccharomyces cerevisiae S288C: A useful set ofstrains and plasmids for PCR-mediated gene disruption andother applications. Yeast 14: 115–132.
Chandra, A., Hughes, T.R., Nugent, C.I., and Lundblad, V. 2001.Cdc13 both positively and negatively regulates telomere rep-lication. Genes & Dev. 15: 404–414.
Cohn, M. and Blackburn, E.H. 1995. Telomerase in yeast. Sci-ence 269: 396–400.
Cooper, J.P., Nimmo, E.R., Allshire, R.C., and Cech, T.R. 1997.Regulation of telomere length and function by a Myb-do-main protein in fission yeast. Nature 385: 744–747.
Counter, C.M., Meyerson, M., Eaton, E.N., and Weinberg, R.A.1997. The catalytic subunit of yeast telomerase. Proc. Natl.Acad. Sci. 94: 9202–9207.
Crabbe, L., Verdun, R.E., Haggblom, C.I., and Karlseder, J. 2004.Defective telomere lagging strand synthesis in cells lackingWRN helicase activity. Science 306: 1951–1953.
Crick, F.H. 1966. Codon–anticodon pairing: The wobble hy-pothesis. J. Mol. Biol. 19: 548–555.
Cristofari, G. and Lingner, J. 2006. Telomere length homeosta-sis requires that telomerase levels are limiting. EMBO J. 25:565–574.
Ferreira, M.G., Miller, K.M., and Cooper, J.P. 2004. Indecentexposure: When telomeres become uncapped. Mol. Cell 13:7–18.
Forstemann, K. and Lingner, J. 2001. Molecular basis for telo-mere repeat divergence in budding yeast. Mol. Cell. Biol. 21:7277–7286.
Forstemann, K., Hoss, M., and Lingner, J. 2000. Telomerase-dependent repeat divergence at the 3� ends of yeast telo-meres. Nucleic Acids Res. 28: 2690–2694.
Forstemann, K., Zaug, A.J., Cech, T.R., and Lingner, J. 2003.Yeast telomerase is specialized for C/A-rich RNA templates.Nucleic Acids Res. 31: 1646–1655.
Fouche, N., Ozgur, S., Roy, D., and Griffith, J.D. 2006. Replica-tion fork regression in repetitive DNAs. Nucleic Acids Res.34: 6044–6050.
Fulton, T.B. and Blackburn, E.H. 1998. Identification ofKluyveromyces lactis telomerase: Discontinuous synthesisalong the 30-nucleotide-long templating domain. Mol. Cell.Biol. 18: 4961–4970.
Garvik, B., Carson, M., and Hartwell, L. 1995. Single-strandedDNA arising at telomeres in cdc13 mutants may constitutea specific signal for the RAD9 checkpoint. Mol. Cell. Biol.15: 6128–6138.
Goudsouzian, L.K., Tuzon, C.T., and Zakian, V.A. 2006. S. cere-visiae Tel1p and Mre11p are required for normal levels ofEst1p and Est2p telomere association. Mol. Cell 24: 603–610.
Greenwell, P.W., Kronmal, S.L., Porter, S.E., Gassenhuber, J.,Obermaier, B., and Petes, T.D. 1995. TEL1, a gene involvedin controlling telomere length in S. cerevisiae, is homolo-gous to the human ataxia telangiectasia gene. Cell 82: 823–829.
Greider, C.W. 1991. Telomerase is processive. Mol. Cell. Biol.11: 4572–4580.
Greider, C.W. and Blackburn, E.H. 1985. Identification of a spe-cific telomere terminal transferase activity in Tetrahymenaextracts. Cell 43: 405–413.
Greider, C.W. and Blackburn, E.H. 1987. The telomere terminaltransferase of Tetrahymena is a ribonucleoprotein enzymewith two kinds of primer specificity. Cell 51: 887–898.
Greider, C.W. and Blackburn, E.H. 1989. A telomeric sequencein the RNA of Tetrahymena telomerase required for telo-mere repeat synthesis. Nature 337: 331–337.
Harley, C.B., Futcher, A.B., and Greider, C.W. 1990. Telomeresshorten during ageing of human fibroblasts. Nature 345:458–460.
Henning, K.A., Moskowitz, N., Ashlock, M.A., and Liu, P.P.1998. Humanizing the yeast telomerase template. Proc.Natl. Acad. Sci. 95: 5667–5671.
Hug, N. and Lingner, J. 2006. Telomere length homeostasis.Chromosoma 115: 413–425.
Ivessa, A.S., Zhou, J.Q., Schulz, V.P., Monson, E.K., and Zakian,V.A. 2002. Saccharomyces Rrm3p, a 5� to 3� DNA helicasethat promotes replication fork progression through telo-meric and subtelomeric DNA. Genes & Dev. 16: 1383–1396.
Lin, J.J. and Zakian, V.A. 1996. The Saccharomyces CDC13protein is a single-strand TG1-3 telomeric DNA-binding pro-tein in vitro that affects telomere behavior in vivo. Proc.Natl. Acad. Sci. 93: 13760–13765.
Lin, J., Smith, D.L., and Blackburn, E.H. 2004. Mutant telomeresequences lead to impaired chromosome separation and aunique checkpoint response. Mol. Biol. Cell 15: 1623–1634.
Lingner, J., Hughes, T.R., Shevchenko, A., Mann, M., Lundblad,V., and Cech, T.R. 1997. Reverse transcriptase motifs in thecatalytic subunit of telomerase. Science 276: 561–567.
Lue, N.F. and Peng, Y. 1997. Identification and characterizationof a telomerase activity from Schizosaccharomyces pombe.Nucleic Acids Res. 25: 4331–4337.
Lue, N.F. and Peng, Y. 1998. Negative regulation of yeast telo-merase activity through an interaction with an upstreamregion of the DNA primer. Nucleic Acids Res. 26: 1487–1494.
Lundblad, V. and Szostak, J.W. 1989. A mutant with a defect intelomere elongation leads to senescence in yeast. Cell 57:633–643.
Makovets, S., Herskowitz, I., and Blackburn, E.H. 2004.Anatomy and dynamics of DNA replication fork movement
Regulation of telomerase processivity
GENES & DEVELOPMENT 2493

in yeast telomeric regions. Mol. Cell. Biol. 24: 4019–4031.Marcand, S., Brevet, V., and Gilson, E. 1999. Progressive cis-
inhibition of telomerase upon telomere elongation. EMBO J.18: 3509–3519.
Morin, G.B. 1989. The human telomere terminal transferaseenzyme is a ribonucleoprotein that synthesizes TTAGGGrepeats. Cell 59: 521–529.
Mozdy, A.D. and Cech, T.R. 2006. Low abundance of telomer-ase in yeast: Implications for telomerase haploinsufficiency.RNA 12: 1721–1737.
Nugent, C.I., Hughes, T.R., Lue, N.F., and Lundblad, V. 1996.Cdc13p: A single-strand telomeric DNA-binding proteinwith a dual role in yeast telomere maintenance. Science 274:249–252.
Pennock, E., Buckley, K., and Lundblad, V. 2001. Cdc13 deliversseparate complexes to the telomere for end protection andreplication. Cell 104: 387–396.
Prescott, J. and Blackburn, E.H. 1997a. Functionally interactingtelomerase RNAs in the yeast telomerase complex. Genes &Dev. 11: 2790–2800.
Prescott, J. and Blackburn, E.H. 1997b. Telomerase RNA muta-tions in Saccharomyces cerevisiae alter telomerase actionand reveal nonprocessivity in vivo and in vitro. Genes &Dev. 11: 528–540.
Prowse, K.R., Avilion, A.A., and Greider, C.W. 1993. Identifica-tion of a nonprocessive telomerase activity from mousecells. Proc. Natl. Acad. Sci. 90: 1493–1497.
Ritchie, K.B., Mallory, J.C., and Petes, T.D. 1999. Interactions ofTLC1 (which encodes the RNA subunit of telomerase),TEL1, and MEC1 in regulating telomere length in the yeastSaccharomyces cerevisiae. Mol. Cell. Biol. 19: 6065–6075.
Sabourin, M., Tuzon, C.T., and Zakian, V.A. 2007. Telomeraseand Tel1p preferentially associate with short telomeres in S.cerevisiae. Mol. Cell 27: 550–561.
Sherman, F. 1991. Getting started with yeast. Methods Enzy-mol. 194: 3–21.
Singer, M.S. and Gottschling, D.E. 1994. TLC1: Template RNAcomponent of Saccharomyces cerevisiae telomerase. Sci-ence 266: 404–409.
Smolka, M.B., Albuquerque, C.P., Chen, S.H., and Zhou, H.2007. Proteome-wide identification of in vivo targets ofDNA damage checkpoint kinases. Proc. Natl. Acad. Sci. 104:10364–10369.
Taggart, A.K., Teng, S.C., and Zakian, V.A. 2002. Est1p as a cellcycle-regulated activator of telomere-bound telomerase. Sci-ence 297: 1023–1026.
Teixeira, M.T., Arneric, M., Sperisen, P., and Lingner, J. 2004.Telomere length homeostasis is achieved via a switch be-tween telomerase- extendible and -nonextendible states.Cell 117: 323–335.
Tseng, S.F., Lin, J.J., and Teng, S.C. 2006. The telomerase-re-cruitment domain of the telomere binding protein Cdc13 isregulated by Mec1p/Tel1p-dependent phosphorylation.Nucleic Acids Res. 34: 6327–6336.
Varani, G. and McClain, W.H. 2000. The G × U wobble basepair. A fundamental building block of RNA structure crucialto RNA function in diverse biological systems. EMBO Rep.1: 18–23.
Wang, S.S. and Zakian, V.A. 1990. Sequencing of Saccharomy-ces telomeres cloned using T4 DNA polymerase reveals twodomains. Mol. Cell. Biol. 10: 4415–4419.
Yu, G.L., Bradley, J.D., Attardi, L.D., and Blackburn, E.H. 1990.In vivo alteration of telomere sequences and senescencecaused by mutated Tetrahymena telomerase RNAs. Nature344: 126–132.
Chang et al.
2494 GENES & DEVELOPMENT

Regulation of Telomerase Processivity
Tel443 TGGTGTGTGT GTGTGGGTGT GGTGTGTGTG TGGGTGTGGG TGTGGGTGTG GGTGTGGATT TGGTGTGTGG TGTGGTGTGG TGTGGTG-3’
Tel433 TGTGGGTGTG TGGGTGTGGT GTGTGTGTGG GTGTGGGTGT GGGTGTGGGT GTGGGTGTGG TGTGGTGTGG GTGTGGTGTG TGTGTGGGTG TGGTGTGTGG TGTGGTGTGT GGTGTGGTGG >
Tel437 TGTGGGTGTG TGGGTGTGGT GTGTGTGTGT GGGTGTGGGT GTGGTGTGGT GTGGTGTGTG GTGTGTGTGG TGTGGTGTGG TGTGTGGTGT GGATTTGGTG TGTGGTGTGG TGTGGTGGTG >
Tel438 TGTGGGTGTG TGGGTGTGGT GTGTGTGTGT GGGTGTGGGT GTGGGTGTGG GTGTGGTGTG GTGTGGGTGT GGTGTGTGTG TGGGTGTGGT GTGTGGTGTG GTGTGTGGTG TGGTGTGTGT >
Tel431 TGTGGGTGTG TGGGTGTGGT GTGTGTGTGT GGGTGTGGGT GTGGGTGTGG GTGTGGGTGT GGTGTGGTGT GGGTGTGGAT TTGGTGTGTG GTGTGTGTGT GGTGTGGTGT GTGGTGTGGT >
Tel429 TGTGGGTGTG TGGGTGTGGT GTGTGTGTGT GGGTGTGGGT GTGGGTGTGG GTGTGGGTGT GGTGTGGTGT GGGTGTGGTG GTGTGTGGTG TGGTGGTGTG GTGTGGTGGT GTGGTGTGGT >
Tel435 TGTGGGTGTG TGGGTGTGGT GTGTGTGTGT GGGTATGGGT GTGGGTGTGG GTGTGGGTGT GGTGTGGTGT GGGTGTGGTG TGGTGTGTGG TGTGTGTAGT GTGTGGTGGG TTTGGTGTGT >
Tel439 TGTGGGTGTG TGGGTGTGGT GTGTGTGTGT GGGTGTGGGT GTGGGTGTGG GTGTGGGTGT GGTGTGGTGT GGGTGTGGTG TGGTGTGGTG TGGTGTGTGG TGTGGTGTGT GGTGTGGTGT >
Tel442 TGTGGGTGTG TGGGTGTGGT GTGTGTGTGT GGGTGCGGGT GTGGGTGTGG GTGTGGGTGT GGTGTGGTGT GGGTGTGGTG TGGTGTGGTG TGTGGATTTG GTGTGGTGTG TGTGGTGGTG >
Tel434 TGTGGGTGTG TGGGTGTGGT GTGTGTGTGT GGGTGTGGGT GTGGGTGTGG GTGTGGGTGT GGTGTGGTGT GGGTGTGGTG TGTGTGTGGG TGTGGATTTG GTGTGTGGTG TTTGGTGTGT >
Tel436 TGTGGGTGTG TGGGTGTGGT GTGTGTGTGT GGGTGTGGGT GTGGGTGTGG GTGTGGGTGT GGTGTGGTGT GGGTGTGGTG TGTGTGTGGG TGTGGTGTGT GGTGTGGTGT GTGGTGTGTG >
Tel432 TGTGGGTGTG TGGGTGTGGT GTGTGTGTGT GGGTGTGGGT GTGGGTGTGG GTGTGGGTGT GGTGTGGTGT GGGTGTGGTG TGTGTGTGGG TGTGGTGTGT GGTGTGGTGT GTGGTGTGGT >
Tel441 TGTGGGTGTG TGGGTGTGGT GTGTGTGTGT GGGTGTGGGT GTGGGTGTGG GTGTGGGTGT GGTGTGGTGT GGGTGTGGTG TGTGTGTGGG TGTGGTGTGT GGTGTGGTGT GTGGTGTGGT >
Tel444 TGTGGGTGTG TGGGTGTGGT GTGTGTGTGT GGGTGTGGGT GTGGGTGTGG GTGTGGGTGT GGTGTGGTGT GGGTGTGGTG TGTGTGTGGG TGTGGTGTGT GGTGTGGTGT GTGGTGTGGT >
Tel440 TGTGGGTGTG TGGGTGTGGT GTGTG
Tel433 TGTGGTGGTG TGTGGTGTGT-3’
Tel437 TGGTGTGGTG TGGTGTGGTG TGTGGTGTGG TGGTGTGGTG TGGTGTGGTG TGTGTGGTGT GGGTGGT-3’
Tel438 GGTGTGGATT TGGTGTG-3’
Tel431 GTGGTGTGGT GTGGT-3’
Tel429 GTGGTGTG-3’
Tel435 GGTGTGTGGG TGGTGT-3’
Tel439 GGTGTGGTGT GTGGTGTGGT GTGTGGTGTG TGTGGTGTGG TGTGGTGTGG TGTGGTGTGT GGTGTGGTGT GGTGTGGTGT GGTGTGGTGT GGTGTGGTGC GGTGTGTGTG TGGGGGGTTT >
Tel442 TGGTGGATTT GGTGTGTGGT GTGTGGTGTG TGGTGTTGGT GTGGATTTTG GTGTGGGGTG GTGTGGTGTG TGTGTGGTGT GGTGTTTGGT GTGTGGTGTG TGGATTTGGT GTGGTGTGGT >
Tel434 GGTGTGGTGT GGTGTGGTGT GGTGT-3’
Tel436 TGGTGTGGTG TGGTGTGTGG TGTGGTGTGT GTGGTGTGGT GTGTGTGTGG TGTGTGTGGT GTGGTGTGTG GTGTGTGGTG TTTTGGTGTG TGGTGGTGTG GTGTGTGGTG TGGTGGTGGT >
Tel432 GTGTGGTGTG GTGTGTGG-3’
Tel441 GTGGTGTGGT GTGGTGTGGA TTTGGTGTGT GGTGTGGTGT GGTGT-3’
Tel444 GTGGTGTGGT GTGGTGTGGT GTTGGTGTGT GGTGGTGGTG TGGTGTGGTG TGTGGT-3’
Tel439 TTTTGGTGTG TGGGTGTGGT GTGTGGTGTG GTGTGGTGTG TGGTGGTGTG GTGTGTGGAT TTGGTGTGGA TTTGGTGTGG TGTGTGGTGT GGTGGTGTGG TGTGGTGTGT GTGGTGTGGT >
Tel442 GTGGTGTG-3’
Tel436 GTGGTGTGTG GTGTGGTGTG GTGTGGGTTT GGTGTGGTGT GGTGTGGTGT GGTGGTGTGG TGTGGTGTGG TGGTGTGGTG-3’
Tel439 GTGGTGTGGT GTGTGG-3’
Supp
lem
enta
ry F
igur
e S1
. Tel
omer
ic s
eque
nces
from
a tl
c1-tm
mut
ant (
som
e of
whi
ch a
re s
how
n in
Fig
ure
1B
). O
nly
the
mut
ant d
iver
gent
regi
ons
of e
ach
sequ
ence
(in
red)
wer
e us
ed to
det
erm
ine
the
mut
ant c
onse
nsus
seq
uenc
e. S
eque
nces
in b
lue
are
dive
rgen
t but
con
tain
wild
-type
repe
ats
due
to te
lom
eric
re
com
bina
tion
even
ts. B
olde
d se
quen
ces
high
light
rare
occ
urre
nces
of G
GG
trin
ucle
otid
e in
corp
orat
ion
by th
e m
utan
t tel
omer
ase.
Bol
ded
and
unde
rline
d se
quen
ces
show
the
two
occu
rren
ces
of G
GG
trin
ucle
otid
es th
at c
anno
t be
easi
ly id
entif
ied
as m
utan
t.

Regulation of Telomerase Processivity
Supplementary Figure S2A. Aligned sequences from a 2T-STEX assay in which the donor strain carried the tlc1-tm allele. Telomere extension events were detected by sequence divergence. Sequences in blue are repeats added by telomerase with wild-type TLC1 while sequences in red are repeats added by telomerase with the mutant tlc1-tm
Tel147 TGTGGGTGTG GTGTGTGTGG GTGTGGTGTG GGTGTGGTGT GTGGTGTGGT GTGGTGTGGT GTGGGTGT-3’
Tel156 TGTGGGTGTG GTGTGTGTGG GTGTGGTGTG GGTGTGGTGT GTGTGGGTTT GGTGTGGTGT GGTGTGGTGG TGTGGTGTGT GTGGGTGTGG TGT-3’
Tel152 TGTGGGTGTG GTGTGTGTGG GTGTGGTGTG GGTGTGGTGT GTGTGGGTGT GGTGTGTGGG TGTGGGTGTG GTGGGTGTGG TGTGTGTGGG TGTGGTGTGT GTGGGTGTGG TGTGGGT-3’
Tel159 TGTGGGTGTG GTGTGTGTGG GTGTGGTGTG GGTGTGGTGT GTGTGGGTGT GGT-3’
Tel163 TGTGGGTGTG GTGTGTGTGG GTGTGGTGTG GGTGT-3’
Tel281 GTGGGTGTGG TGGGTGTGGT GTGTGTGGGT GTGGTGTGTG TGGGTGTGGT GTGGGTGTGG TGTGTGTGGT GTGT-3’
Tel274 GTGGGTGTGG TGTGTGGGTG TGTGGGTGTG GCGTGTGGGT GTGGTGTGTG TGGGTGTGGT GTGTGGGTGT GGGTGTGGTG GGTGTGGTGT GTGTGGGTGT GGTGTGTG-3’
Tel277 GTGGGTGTGG GTGTGTGGGT GTGTGGGTGT GTGGATGTGG GTGTGGTGTG GGTGTGGTGT GGGTGTGGTG GCAGGGGGAA GGGTGTGGTG TGGGTGTGGT GTGTGGGTGT >
Tel278 GTGGGTGTGG GTGTGTGGGT GTGTGGGTGT GTGGATGTGG GTGTGGTGTG GGTGTGGTGT GGGTGTGGTG GCAGGGGGAA GGGTGTGGTG TGGGTGTGGT GTGTGGGTGT >
Tel282 GTGGGTGTGG GTGTGTGGGT GTGTGGGTGT GTGGATGTGG GTGTGGTGTG GGTGTGGTGT GGGTGTGGTG GCAGGGGGAA GGGTGTGGTG TGGGTGTGGT GTGTGGGTGT >
Tel277 GGGTGTGGTG GGTGTGGTGT GTGTGGGTGT GGTGTGTGTG GGTGTGGTGT GGGTTT-3’
Tel278 GTGGGTGTGG TGTGTGGGTG TGGTGTGTGT GGGTGTGGTG TGTGGGTGTG GGTGTGGTGG GTGTGGTGTG TGTGGGTGTG GTGTGTGTGG GTGTGGTGTG GGTGTGGTGT GTGT-3’
Tel282 GTGGGTGTGG TGTGTGGGTG TGGTGTGTGT GGGTGTGGTG TGTGGGTGTG GGTGTGGTGG GTGTGGTGTG TGTGGGTGTG GTGTGTGTGG GTGTGGTGTG G-3’
Tel375 GTGGGTGTGG GTGTGGTGGG TGTGGTGTGT GTGGGTGTGG TGTGTGTGG-3’
Tel361 GTGGGTGTGG GTGTGTGGGT GTGTGGGTGT GTGGATGTGG GTGTGGTGTG TGGTGTGGTG TGGTGTGGTG TGTGGTGTGT GTGGTGTGGT GTGTGTGGAT TTGGTGTGTG GTGTGGTGTG >
Tel369 GTGGGTGTGG GTGTGTGGGT GTGTGGGTGT GTGGATGTGG GTGTGGTGTG GGTGTGGTGT GGGTGTGGTG GCAGGGGGAA GGGTGTGGTG TGGGTGTGGT GTGTGGGTGT GTGGGTGTGG >
Tel343 GTGGGTGTGG GTGTGTGGGT GTGTGGGTGT GTGGATGTGG GTGTGGTGTG GGTGTGGTGT GGGTGTGGTG GCAGGGGGAA GGGTGTGGTG TGGGTGTGGT GTGTGGGTGT GTGGGTGTGG >
Tel357 GTGGGTGTGG GTGTGTGGGT GTGTGGGTGT GTGGATGTGG GTGTGGTGTG GGTGTGGTGT GGGTGTGGTG GCAGGGGGAA GGGTGTGGTG TGGGTGTGGT GTGTGGGTGT GTGGGTGTGG >
Tel361 TGGTGTGTGG TGTGGTGTGT GGGGGTGTGG TGTGGTGTGG TGTGGTGTGT GGTGTGTGTG TGGTGTGTGT GGTGTGGTGT GGTGTGTG-3’
Tel369 TGTGTGGGTG TGGTGTGTGT GGGTGTGGTA TGTGGGTGTG GGTGTGGTGT GTGTGGGTGT GGTGTGTGTG GGTGT-3’
Tel343 TGTGTGGGTG TGGTGTGTGT GGGTGTGGTG TGTGGGTGTG GGTGTGGTGG GTGTGGTGTG TGTGGGTGTG GTGTGTGTGG GTGTGGTGTG GGTGTGGTGT GTGTGGGTGT GGTGGTGT-3’
Tel357 TGTGTGGGTG TGGTGTGTGT GGGTGTGGTG TGTGGGTGTG GGTGTGGTGG GTGTGGTGTG TGTGGGTGTG GTGTGTGTGG GTGTGGTGTG GGTGTGGTGT GTGTGGGTGT GG-3’
Tel455 GTGGATGTGG GTGTGGTGTG TGGGTGTGTG G-3’
Tel453 GTGGATGTGG GTGTGGTGTG GTGTGTGTGG GTGTGGTGTG TGGGTGTGGG TGT-3’
Tel499 GTGGATGTGG GTGTGGTGTG GGTGTGGTGT GGGTGTGGTG GCAGGGGGAA GGGTGTGGTG TGGGTGTGTG GGTGTGGTGT GTGGGTGTGG TGTGTGTGTG GGTGTGGTGT GGTGT-3’
Tel481 GTGGATGTGG GTGTGGTGTG GGTGTGGTGT GGGTGTGGTG GCAGGGGGAA GGGTGTGGTG TGGGTGTGGT GTGGGTGTGT GGTGTGTGGT GTGGTGTGGG TGTGGGTGTG GATTTGGTGC >
Tel458 GTGGATGTGG GTGTGGTGTG GGTGTGGTGT GGGTGTGGTG GCAGGGGGAA GGGTGTGGTG TGGGTGTGGT GTGTGGGTGT GTGGGTGTGG TGTGTGGGTG TGGTGTGTGG GGTGTGGTGG >
Tel498 GTGGATGTGG GTGTGGTGTG GGTGTGGTGT GGGTGTGGTG GCAGGGGGAA GGGTGTGGTG TGGGTGTGGT GTGTGGGTGT GTGGGTGTGG TGTGTGGGTG TGGTGTGTGT GGGTGTGGGT >
Tel473 GTGGATGTGG GTGTGGTGTG GGTGTGGTGT GGGTGTGGTG GCAGGGGGAA GGGTGTGGTG TGGGTGTGGT GTGTGGGTGT GTGGGTGTGG TGTGTGGGTG TGGTGTGTGT GGGTGTGGGT >
Tel484 GTGNATGTGG NTGTGGNGTG GNTGTGGNGT GGNTGTGGTG GCAGGGGGAA GGGTGTGGTG TGGGTGTGGT GTGTGGGTGT GTGGGTGTGG TGTGTGGGTG TGGTGTGTGT GGGTGTGGTG >
Tel496 GTGGATGTGG GTGTGGTGTG GGTGTGGTGT GGGTGTGGTG GCAGGGGGAA GGGTGTGGTG TGGGTGTGGT GTGTGGGTGT GTGGGTGTGG TGTGTGGGTG TGGTGTGTGT GGGTGTGGTG >
Tel493 GTGGATGTGG GTGTGGTGTG GGTGTGGTGT GGGTGTGGTG GCAGGGGGAA GGGTGTGGTG TGGGTGTGGT GTGTGGGTGT GTGGGTGTGG TGTGTGGGTG TGGTGTGTGT GGGTGTGGTG >
Tel460 GTGGATGTGG GTGTGGTGTG GGTGTGGTGT GGGTGTGGTG GCAGGGGGAA GGGTGTGGTG TGGGTGTGGT GTGTGGGTGT GTGGGTGTGG TGTGTGGGTG TGGTGTGTGT GGGTGTGGTG >
Tel497 GTGGATGTGG GTGTGGTGTG GGTGTGGTGT GGGTGTGGTG GCAGGGGGAA GGGTGTGGTG TGGGTGTGGT GTGTGGGTGT GTGGGTGTGG TGTGTGGGTG TGGTGTGTGT GGGTGTGGTG >
Tel457 GTGGATGTGG GTGTGGTGTG GGTGTGGTGT GGGTGTGGTG GCAGGGGGAA GGGTGTGGTG TGGGTGTGGT GTGTGGGTGT GTGGGTGTGG TGTGTGGGTG TGGTGTGTGT GGGTGTGGTG >
Tel464 GTGGATGTGG GTGTGGTGTG GGTGTGGTGT GGGTGTGGTG GCAGGGGGAA GGGTGTGGTG TGGGTGTGGT GTGTGGGTGT GTGGGTGTGG TGTGTGGGTG TGGTGTGTGT GGGTGTGGTG >
Tel481 GGTGTGGTGT GTGGGTTTTT-3’
Tel458 GTGTGTG-3’
Tel498 GTGGTGTGTG TGGGTGTGGG TT-3’
Tel473 GTGGTGTGTG GGTGTGGGCG TGGGTGTGGG TGTGGTGTGT GGGTGTGGTG T-3’
Tel484 GGTGTGGTTG GGTGTGGTGT GGGTGTGGTG TGTGTG-3’
Tel496 TGGGTGTGGT GTGGTGTGTG GTGTGCGGGT GTGGTGTGGT GTGTGTGGGT GT-3’
Tel493 TGTGGGTGTG GTGCGGTGTG GTGT-3’
Tel460 TGTGGGTGTG GTGTGTGTGG GTGTGGT-3’
Tel497 TGTGGGTGTG GGTGTGGTGG GTGTGGTGTG TGGGTGTGGG TGTGGTG-3’
Tel457 TGTGGGTGTG GGTGTGGTGG GTGTGGTGTG TGTGGGTGGT GTGGTG-3’
Tel464 TGTGGGTGTG GGTGTGGTGG GTGTGGTGTG TGTGGGTGTG GTGTGTGTGG GTGT-3’

Regulation of Telomerase Processivity
Supplementary Figure S2B. Aligned sequences from a 2T-STEX assay in which the recipient strain carried the tlc1-tm allele. Telomere extension events were detected by sequence divergence. Sequences in blue are repeats added by telomerase with wild-type TLC1 while sequences in red are repeats added by telomerase with the mutant tlc1-tm.
Tel852 GTGTGGGTGT GTGGGTGTGG GTGTGTGGGT GTGGGTGTGG TGTGTGGGTG TGGTGTGGGT GTGGTGTGTG GGTGTGGGTG TGTGGGTGTG GGTGTGGGTG >
Tel853 GTGTGTGGGT GTGGTGTGTG TGGGTGTGGT GTGTGGGTGT GGGTGTGGTG GGTGTGGTGT GTGGGTGTGG TGTGTGTGGG TGTGGTGT-3’
Tel860 GTGTGTGGGT GTGGGTGTGG GTGTGTGGGT GTGTGGGTGT GTGGATGTGG GTGTGGTGTG GGTGTGGTGG GTGTGGTGTG TGTGGGTGTG GTGTGTGTGG >
Tel861 GTGTGTGGGT GTGGGTGTGG GTGTGTGGGT GTGTGGGTGT GTGGATGTGG GTGTGGTGTG GGTGTGGTGT GTGGGCGTGT GGGTGTGGTG TGTGGGTGTG >
Tel879 GTGCGTGGGT GTGGGTGTGG GTGTGTGGGT GTGTGGGTGT GTGGATGTGG GTGTGGTGTG GGTGTGGTGT GGGTGTGGTG GCAGGGGGAA GGGTGTGGTG >
Tel900 GTGTGTGGGT GTGGGTGTGG GTGTGTGGGT GTGTGGGTGT GTGGATGTGG GTGTGGTGTG GGTGTGGTGT GGGTGTGGTG GCAGGGGGAA GGGTGTGGTG >
Tel877 GTGTGTGGGT GTGGGTGTGG GTGTGTGGGT GTGTGGGTGT GTGGATGTGG GTGTGGTGTG GGTGTGGTGT GGGTGTGGTG GCAGGGGGAA GGGTGTGGTG >
Tel842 GTGTGTGGGT GTGGGTGTGG GTGTGTGGGT GTGTGGGTGT GTGGATGTGG GTGTGGTGTG GGTGTGGTGA GGGTGTGGTG GCAGGGGGAA GGGTGTGGTG >
Tel857 GTGTGTGGGT GTGGGTGTGG GTGTGTGGGT GTGTGGGTGT GTGGATGTGG GTGTGGTGTG GGTGTGGTGT GGGTGTGGTT GCAGGGGGAA GGGTGTGGTG >
Tel862 GTGTGTGGGT GTGGGTGTGG GTGTGTGGGT GTGTGGGTGT GTGGATGTGG GTGTGGTGTG GGTGTGGTGT GGGTGTGGTG GCAGGGGGAA GGGTGTGGTG >
Tel863 GTGTGTGGGT GTGGGTGTGG GTGTGTGGGT GTGTGGGTGT GTGGATGTGG GTGTGGTGTG GGTGTGGTGT GGGTGTGGTG GCAGGGGGAA GGGTGTGGTG >
Tel899 GTGTGTGGGT GTGGGTGTGG GTGTGTGGGT GTGTGGGTGT GTGGATGTGG GTGTGGTGTG GGTGTGGTGT GGGTGTGGTG GCAGGGGGAA GGGTGTGGTG >
Tel833 GTGTGTGGGT GTGGGTGTGG GTGTGTGGGT GTGTGGGTGT ATGGATGTGG GTGTGGTGTG GGTGTGGTGT GGGTGTGGTG GCAGGGGGAA GGGTGTGGTG >
Tel840 GTGTGTGGGT GTGGGTGTGG GTGTGTGGGT GTGTGGGTGT GTG:ATGTGG GTGTGGTGTG GGTGTGGTGT GGGTGTGGTG GCAGGGGGAA GGGTGTGGTG >
Tel852 TGGTGTGTGT GTGTGGGTGC GGTGTGGTGT GTGTGGGTGT GGGTGTGGTG TGGGTGTGTG GGTGTGGTGT GGGTGTGGTG TGTGGGTGTG GGTGTGGGTG >
Tel860 GTGTGGTGTG GGTGTGGTGT GTGTGGGTGT GTGGGTGTGG TGTGTGGTGT GGTGTGGTGT GGTGTGTGTG GTGTGGTGTG GTGTGT-3’
Tel861 GTGTGTGTGG GTGTGGTGTG TGGGTGTGGG TGTGGTGGGT GTGGTGTGTG TGGGTGTGGT GTGTGTGGGT GTGGTGTGGG TGTGGTGTGT GTGGGTGTGG >
Tel879 TGGGTGTGGT GTGCGGGTGT GGGTGCGGGT GTGGTGTGGG TGTGGTGTGT G-3’
Tel900 TGGGTGTGGT GTGTGGGTGT GGGTGTGGTG TGTGGGTGTG GTGTGTGTGG GTGTGGTGTG TGGGTGTGGG TGTGGTGGGT GTGGTGTGTG TGGGTGTGGT >
Tel877 TGGGTGTGGT GTGTGGGTGT GTGGGTGTGG TGTGTGGGTG TGGTGTGTGT GGGTGTGGTG TGTGTGGGTG TGGGTGTGGT GTGGGTGTGG TGTGCGTGGG >
Tel842 TGGGTGTGGT GTGTGGGTGT GTGGGTGTGG TGTGTGGGTG TGGTGTGTGT GGGTGTGGTG TGTGGGTGTG GGTGTGGTGG GTGTGGTGTG TGTGTGTGTG >
Tel857 TGGGTGTGGT GTGTGGGTGT GTGGGTGTGG TGTGTGGGTG TGGTGTGTGT GGGTGTGGTG TGTGGGTGTG GGTGTGGTGG GTGTGGTGTG TGTGGGTGTG >
Tel862 TGGGTGTGGT GTGTGGGTGT GTGGGTGTGG TGTGTGGGTG TGGTGTGTGT GGGTGTGGTG TGTGGGTGTG GGTGTGGTGG GTGTGGTGTG TGTGGGTGTG >
Tel863 TGGGTGTGGT GTGTGGGTGT GTGGGTGTGG TGTGTGGGTG TGGTGTGTGT GGGTGTGGTG TGTGGGTGTG GGTGTGGTGG GTGTGGTGTG TGTGGGTGTG >
Tel899 TGGGTGTGGT GTGTGGGTGT GTGGGTGTGG TGTGTGGGTG TGGTGTGTGT GGGTGTGGTG TGTGGGTGTG GGTGTGGTGG GTGTGGTGTG TGTGGGTGTG >
Tel833 TGGGTGTGGT GTGTGGGTGT GTGGGTGTGG TGTGTGGGTG TGGTGTGTGT GGGTGTGGTG TGTGGGTGTG GGTGTGGTGG GTGTGGTGTG TGTGGGTGTG >
Tel840 TGGGTGTGGT GTGTGGGTGT GTGGGTGTGG TGTGTGGGTG TGGTGTGTGT GGGTGTGGTG TGTGGGTGTG GGTGTGGTGG GTGTGGTGTG TGTGGGTGTG >
Tel852 TGGTGTGTGG GTGTGTGTGG GTGTGGGTGT GGGTGTGGTG TGTGTGGGTG TGGTGTGTGG GTGTGGGTGT GTGTGGGTGT GGGTGT-3’
Tel861 TGTGGGTGTG GTGTGTGTGG GTGTGTGGGT GTGGTGTGTG GTGTG-3’
Tel900 GTGTGTGGGT GTGGTGTGGG TGTGGTGTGT GTGGGTGTGG TGTGGGTGTG GTGTGTGTGG GTGTGTGGGT GTGGTGTGTG GTGTGGTGTG GTGTGGTGTG >
Tel877 TGTGGTGTGT GTGGGTGTGG TGTGTGGGTG TGGT-3’
Tel842 GTGTGTG-3’
Tel857 GTGTGGGTGT GGTGTGTGTG GGTGTGGTGT GGGTGTGGTG TGTGTGGGTG TGTG-3’
Tel862 GTGTGTGTGG GTGTGGTGTG GGTGTAGTGT GTGGGTGTGG GTGTGGGTGT GGTGTGTGGG TGTGGTGTGT GGGTGTGTGG GTGTGGTGTG GGTGTGGGTG >
Tel863 GTGTGTGTGG GTGTGGTGTG GGTGTGGTGT GTGTGGGTGT GGTGTGGGTG TGGTGTGTGT GGGTGTGTGG TGTGTGTGGT GTGT-3’
Tel899 GTGTGTGTGG GTGTGGTGTG GGTGTGGTGT GTGTGGGTGT GGTGTGGGTG TGGTGTGTGT GGGTGTGTGG GTGTGGTGTG TGGTGTGGTG TGTGGTGTGT >
Tel833 GTGTGTGTGG GTGTGGTGTG GGTGTGGTGT GTGTGGGTGT GGTGTGGGTG TGGTGTGTGT GGGTGTGTGG GTGTGGTGTG TGGTGTGGTG TGGTGTGGTG >
Tel840 GTGTGTGTGG GTGTGGTGTG GGTGTGGTGT GTGTGGGTGT GGTGTGGGTG TGGTGTGTGT GGGTGTGTGG GTGTGGTGTG TGGTGTGGTG TGGTGTGGTG >
Tel900 TGTGGTGTGG TGTGGTGTGG TGT-3’
Tel862 TGGTGT-3’
Tel899 GTGGTGTGT-3’
Tel833 TGTGTGGTGT GGTGTGGTGT GTGGTGTGGT GTGGTGTGGT GTGGTGTGG-3’
Tel840 TGTGTGGTGT GGTGTGGTGT GTGGTGTGGT GTG-3’


103
Chapter 5: Discussion and Perspectives
Extendible versus Non-extendible Telomeric States
Telomere length homeostasis is crucial for the maintenance and propagation of
stable chromosomes. Data preceding the research presented in my thesis already
indicated that S. cerevisiae telomerase activity is regulated in cis at individual
telomeres through the “counting” mechanism which is able to sense the number of
Rap1-Rif1 and -Rif2 complexes bound to the double-stranded telomere tract
(Marcand et al, 1997a; Ray & Runge, 1999). This suggested that the state of the
telomeric chromatin can affect either the enzymatic activity or the recruitment of
telomerase at the chromosome ends. Furthermore, on average, short telomeres are
more elongated that longer ones (Marcand et al, 1999). But it was not clear whether
all telomeres are extended in each round of DNA replication and just the number of
added nucleotides would differ (i.e. telomerase would add more nucleotides at the
shorter telomeres than at the longer ones), or telomerase would act just on the
shortest telomeres, and the longer ones would acquire some kind of protected
structure. Of course, a combination of both mechanisms can lead to an even more
controlled lengthening of the shortest telomeres.
In order to monitor what is really happening on a single telomere in vivo
during one cell cycle, the STEX (single telomere extension) assay was established in
the laboratory (described in Chapter 2) (Teixeira et al, 2004). Data presented in
Chapter 2 confirmed the model in which just a subset of telomeres is subject to
elongation in a single cell cycle i.e. the shorter a telomeres is, the more likely it will
get elongated. While the frequency of telomere extension decreased steadily with
the increase of telomere length, the number of added nucleotides per single
telomere remained constant and was not following the logistic regression model.
However, on the short telomeres, <100 bp in length, very long extension events were
often observed.

Discussion and Perspectives
104
What is the nature of short, extendible telomeres which differentiates them
from the longer, less-extendible ones? On the most basic level, it is the absence of
Rif1 and Rif2 proteins bound on the telomeric tract (Levy & Blackburn, 2004).
Indeed, in both rif1Δ and rif2Δ cells, the telomeres were more extendible at all
telomere length, but a balance between extendible and nonextendible states was
preserved (Chapter 2, Fig. 5). Recent data indicates that the level of Rif2 is reduced
(but not lost, ~50%) on short telomeres. However the binding of Rif1 stays
unchanged upon induced telomere shortening (Sabourin et al, 2007). In order to
explain this result one can only assume that the density of telomere-bound Rif1
would increase with the decreasing telomere length. However, there are other
differences between Rif1 and Rif2 in telomere length regulation. Targeting a single
Rif2 molecule to a single telomere in a rap1ΔC strain prevents the hyper-elongation
that is usually seen in this background, whereas Rif1 is not effective at inhibiting
telomerase by this assay (Levy & Blackburn, 2004). This is an additionally
interesting notion, since just one molecule was enough to partially restore telomere
length control, suggesting the possibility that Rifs can nucleate the formation of a
higher-order structure by initiating the recruitment of additional Rif and/or other
proteins, a mechanism reminiscent of the establishment of silenced
heterochromatin initiated by yeast Sir proteins (Rusche et al, 2003). It is discussed
in more detail in Chapter 2 what the non-extendible heterochromatin structure may
correspond to. However, at least in human cells, this non-extendible structure can
still be extended, meaning that long telomeres do not permanently adopt a closed
conformation. Simultaneous over-expression of both hTR and hTERT led to a
dramatic increase of telomerase activity and its association with telomeres
(Cristofari & Lingner, 2006). This was sufficient to elongate the whole population of
telomeres in primary or cancer cells, far beyond their physiological length. This
brings yet another factor into the telomere homeostasis equation – at least in
human cells, it is achieved with limiting amounts of telomerase. Thus in addition to
switching between non-extendible and extendible states as a function of telomere
length, one can imagine the additional equilibrium between extendible and
extending states as a function of telomerase concentration. These results predict
that long telomeres are not permanently locked in a non-extendible state, but that
the switch between these two telomeric states is rather dynamic, particulary after

Discussion and Perspectives
105
the passage of the DNA-replication machinery. If the telomerase concentration in
this point is not limiting even the long telomeres may be bound and extended by
telomerase. A haploinsuficiency effect was reported in heterozygous mTR+/- and
mTERT+/- mice (Hathcock et al, 2002; Liu et al, 2002) and in TLC1/tlc1Δ
heterozygote yeast cells, where telomeres were noticed to be shorter (Mozdy & Cech,
2006). As described in Chapter 2 of my thesis, however, there is no difference in
elongation frequencies in EST1/est1Δ versus TLC1/tlc1Δ cells.
It remains an open question as to whether or not it would be enough just to
“open” telomeres and make them more accessible for telomerase to template
synthesis of new telomeric repeats, or whether some other molecular downstream
regulators are needed to mark short telomeres for preferential elongation.
Tel1 as a Molecular Marker of Short Telomeres
Tel1 kinase seemed to be an obvious choice as a downstream mediator for the
counting mechanism of telomere length control, since additional Rap1 molecules
tethered adjacent to a telomeric tract were not counted in tel1Δ cells (Ray & Runge,
1999). Furthermore, in tel1Δ cells the association of Est2 and Est1 with telomeres
is significantly reduced (Goudsouzian et al, 2006). In Chapter 3, I showed that the
Tel1 kinase mediates preferential elongation of short telomeres. At the artificial
telomere which lacked subtelomeric elements, short telomeres were not
preferentially elongated in the absence of Tel1. Parallel research showed by ChIP
that Est1, Est2, and Tel1 are all enriched at short telomeres (Bianchi & Shore,
2007b; Hector et al, 2007; Sabourin et al, 2007), and that preferential localization
of Est1 and Est2 on short telomeres depends on Tel1 (Sabourin et al, 2007).
However, localization of Est1 and Est2 in tel1Δ cells were egzamined just on the
artificial, tagged telomere. In my research I as well explored elongation of native
short telomeres, and discovered that they were still prefferentialy elongated even in
the absence of Tel1.

Discussion and Perspectives
106
Tbf1 Can Mark Short Telomeres for Preferential Elongation
The extension frequency at a natural telomere (telomere IL) was still regulated in a
length dependent manner even in the absence of Tel1 (Chapter 3, Fig. 2). The most
obvious difference between this native telomere and the artificial one used in the
previous analysis was the presence of the natural subtelomeric region, which is
absent from the tagged telomere. One can predict that there is an element in the
subtelomere that triggers the Tel1-independent counting mechanism and enables
preferential recognition of short telomeres by telomerase. Two essential insulator
proteins bound to the subtelomeric region, Tbf1 and Reb1, were known to be
negative regulators of telomere length (Fourel et al, 1999; Koering et al, 2000;
Alexander & Zakian, 2003; Berthiau et al, 2006). Strikingly, when I tethered six
copies of Gbd-Tbf1N adjacent to the telomeric tract of the artificial telomere, the
correlation between telomerase-dependent lengthening and telomere length was
restored, even in the absence of Tel1. Thus, even though Tbf1 is a negative
regulator of telomere length, it can restore the recognition of short telomeres by
telomerase (Chapter 3, Fig. 3). Maybe the effect on the telomere length
homeostasis and the ability to restore preferential elongation of the short telomeres
are separated processes and are not achieved by the same mechanism.
Interestingly, tethering Tbf1 (or Reb1 and VP16) can cause delocalization of the
telomeres away from the nuclear periphery (Hediger et al, 2006). The authors
suggest that the delocalization is a cause for the noticed shortening of such
telomeres in tel1Δ cells. Can it be that this delocalization is somehow needed for
the alternative counting mechanism to expose short telomeres and make them more
available for telomerase action? In a more general scheme, the correlation between
telomere elongation and tethering of telomeres to nuclear periphery is not clear.
Perhaps Tbf1 helps the recruitment and/or the activation of telomerase at the short
telomeres, and perhaps this would cause the delocalization of telomeres. It would
be interesting first to establish if active lengthening of telomeres disturbs telomere
localization in any way, like in the situation when telomerase activity is tethered to
telomeres by expressing the Cdc13-Est1 or -Est2 fusion, to conclude more about
possible meaning of Tbf1-dependent delocalization of telomeres. Since tethering of
VP16 as well can delocalize telomeres, it would be interesting to see if this, or
tethering of some other transcriptional activator, would restore the counting

Discussion and Perspectives
107
mechanism in the absence of Tel1, i.e. is this property unique to Tbf1/Reb1, or is it a
more general phenomenon due to the change in the chromatin state?
Another open question is how Tbf1 bound in the subtelomeric region can
“count” the length of the telomeric tract. Perhaps it is nucleating binding of the
protein that actually senses the length of the telomeric tract. Such a mechanism
was suggested for Rif2 (as discussed above), when targeting a single Rif2 molecule
to the telomere in a rap1ΔC strain was sufficient to prevent the hyper-elongation of
that particular telomere (Levy & Blackburn, 2004). On the other hand, it is important
to note that on the artificial telomere in the absence of Tel1, when none of the
“counting” mechanisms are active, the telomere elongation rate for all telomere-
lengths analysed was very low. This suggests that for the establishment of proper
telomere elongation there must be an active positive factor present on short
telomeres, rather then only opening telomeres from the non-extendible structure due
to their shotening. To favor elongation of short telomeres bound Tbf1 protein must
somehow act as positive regulator, perhaps by recruiting a protein that can activate
telomerase.
I showed that artificially tethered Tbf1 can restore preferred extension of
short telomeres. Which part of the natural subtelomeric sequence is necessary and
sufficient to recruit Tbf1? Since Tbf1 can in vivo bind (TTAGGG)n sequence inserted
in the yeast genome (Koering et al, 2000) and in vitro double stranded DNA probes
containing (TTAGGG)n sequence and a yeast telomeric junction sequence that
contains two copies of the TTAGGG separated by 5 bp (Liu & Tye, 1991; Brigati et al,
1993), I added such a sequence to a tagged telomere internally to the TG-tract. This,
however, was not sufficient to re-activate telomerase at short telomeres (data not
shown). It would be interesting to see which part of the X-element would be
sufficient to restore proper counting of telomere length in tel1Δ cells. Considering
the difficulties and cost associated with the STEX assay, one should approach this
experiment with an elaborate hypothesis.
The recent observation that short telomeres are replicated earlier than the
longer ones due to the activation of cryptic origins of replication present in the X-
element suggests an mechanism by which preferential elongation of the short
telomeres is achieved, or a possibility for an alternative counting mechanism
(Bianchi & Shore, 2007a). By this model short telomeres, which replicate earlier,

Discussion and Perspectives
108
would be excesible to the elongation by telomerase for a longer time then the wild-
type telomeres. Supporting this hypothesis preferential elongation of short and
earlier replicated telomeres was observed. So even though I did confirm that Tbf1
can mediate preferential elongation of short telomeres, maybe earlier replication can
be an alternative or parallel possibility. By adding a functional or mutated origin of
replication from the X-element, one could test this hypothesis.
S. cerevisiae Telomerase is Processive in vivo at Critically Short Telomeres
In contrast to many other organisms examined, budding yeast telomerase appears to
be less or nonprocessive in vitro (Cohn & Blackburn, 1995; Lingner et al, 1997b).
However, in vivo analysis suggests that yeast telomerase can add >100 nt in a single
cell cycle (Chapter 2). Thus, in vivo, the enzyme is either processive or it turns over
very rapidly. Even though the number of added nucleotides did not steadily increase
with decline in the telomere length, very long repeat addition events were very often
noticed at short telomeres (<100 bp) (Chapter 2). In Chapter 4, a modified version
of STEX analysis is described, called the two-template single-telomere extension (2T-
STEX) assay. Monitoring telomere elongation in the presence of both a wild-type
TLC1 and a mutant template tlc1tm revealed that telomerase generally acts
nonprocessively in vivo, and can dissociate and re-associate with telomeres during
one round of telomere elongation. However, the repeat addition processivity was
significantly enhanced at extremely short telomeres in a Tel1-dependent manner. As
mentioned in Chapter 3, I noticed that the repeats added to short telomeres in tel1Δ
cells were not longer than the ones added on the longer telomeres (on average 42 nt
on telomeres <100 bp versus 44 nt on the longer ones, at the native telomere IL).
These results would point to a dual role for Tel1 – on one hand it helps the Rap1-
dependent protein counting mechanism to properly sense telomere length. On the
other hand it enables enhancment of telomere repeat addition processivity on short
telomeric ends. Overexpression of Tel1 leads to telomere elongation (Hector et al,
2007; Sabourin et al, 2007). It would be interesting to see which of its functions is
responsible for this phenotype, i.e. are telomeres more frequently elongated or is
telomerase processivity enhanced even on long telomeres? It is also unknown if
Tel1 kinase activity is needed for telomerase processivity, and if yes, phosphorylation

Discussion and Perspectives
109
of which Tel1 substrate is critical. So far, the only known telomeric substrate is
Cdc13 (Tseng et al, 2006), though recent analysis suggests Rif1 as a possible
substrate of Tel1 (Smolka et al, 2007). The possible function of this modification is
still unknown.
There is certainly the possibility that other telomere-length regulating proteins
regulate telomerase processivity as well. One of the best candidates seems to be
the Pif1 helicase, which negatively regulates telomere length, and which can
promote telomerase turnover in vitro (Boule et al, 2005; Boule & Zakian, 2007).
Results obtained by using the STEX assay and monitoring events on single
telomeres helped us understand the basic mechanisms for telomere length
regulation. It was demonstrated for the first time that just a subset of telomeres is
elongated in each cell cycle and that elongation occurred at the shortest telomeres
in the population. This result was the basis for further investigation presented in my
thesis and also performed by other laboratories. The finding that the Tel1 kinase is
critical for the elongation of short telomeres at multiple levels (frequency of
elongation and telomerase processivity) opens new questions for the roles of
posttranslational modifications in telomere length regulation. Even though a few
telomeric proteins (Cdc13, Rap1) are suggested to be Tel1-substrates, the function
of these phosphorylation events has not yet been determined. It is also possible that
the main halmark of short telomeres is not the modification of a single telomeric
protein, but a more general change in chromatin structure. A large number of
proteins that affect telomere length were identified in genome-wide screens. The
precise function of many of these proteins remains unknown. The STEX analysis
would provide more precise insight into their role for telomere length homeostasis.
.


111
References
Adams Martin A, Dionne I, Wellinger RJ, Holm C (2000) The function of DNA polymerase alpha at telomeric G tails is important for telomere homeostasis. Mol Cell Biol 20: 786-796.
Alexander MK, Zakian VA (2003) Rap1p telomere association is not required for mitotic stability of a C(3)TA(2) telomere in yeast. Embo J 22: 1688-1696.
Askree SH, Yehuda T, Smolikov S, Gurevich R, Hawk J, Coker C, Krauskopf A, Kupiec M, McEachern MJ (2004) A genome-wide screen for Saccharomyces cerevisiae deletion mutants that affect telomere length. Proc Natl Acad Sci U S A 101: 8658-8663.
Bai Y, Murnane JP (2003) Telomere instability in a human tumor cell line expressing NBS1 with mutations at sites phosphorylated by ATM. Mol Cancer Res 1: 1058-1069.
Bailey SM, Meyne J, Chen DJ, Kurimasa A, Li GC, Lehnert BE, Goodwin EH (1999) DNA double-strand break repair proteins are required to cap the ends of mammalian chromosomes. Proc Natl Acad Sci U S A 96: 14899-14904.
Baird DM, Jeffreys AJ, Royle NJ (1995) Mechanisms underlying telomere repeat turnover, revealed by hypervariable variant repeat distribution patterns in the human Xp/Yp telomere. Embo J 14: 5433-5443.
Berthiau AS, Yankulov K, Bah A, Revardel E, Luciano P, Wellinger RJ, Geli V, Gilson E (2006) Subtelomeric proteins negatively regulate telomere elongation in budding yeast. Embo J 25: 846-856.
Bertuch AA, Lundblad V (2003) The Ku heterodimer performs separable activities at double-strand breaks and chromosome termini. Mol Cell Biol 23: 8202-8215.
Bertuch AA, Lundblad V (2004) EXO1 contributes to telomere maintenance in both telomerase-proficient and telomerase-deficient Saccharomyces cerevisiae. Genetics 166: 1651-1659.
Bessler JB, Torredagger JZ, Zakian VA (2001) The Pif1p subfamily of helicases: region-specific DNA helicases? Trends Cell Biol 11: 60-65.
Bi X, Wei SC, Rong YS (2004) Telomere protection without a telomerase; the role of ATM and Mre11 in Drosophila telomere maintenance. Curr Biol 14: 1348-1353.
Bi X, Srikanta D, Fanti L, Pimpinelli S, Badugu R, Kellum R, Rong YS (2005) Drosophila ATM and ATR checkpoint kinases control partially redundant pathways for telomere maintenance. Proc Natl Acad Sci U S A 102: 15167-15172.
Bianchi A, Smith S, Chong L, Elias P, de Lange T (1997) TRF1 is a dimer and bends telomeric DNA. Embo J 16: 1785-1794.
Bianchi A, Negrini S, Shore D (2004) Delivery of yeast telomerase to a DNA break depends on the recruitment functions of Cdc13 and Est1. Mol Cell 16: 139-146.
Bianchi A, Shore D (2007a) Early replication of short telomeres in budding yeast. Cell 128: 1051-1062.
Bianchi A, Shore D (2007b) Increased association of telomerase with short telomeres in yeast. Genes Dev 21: 1726-1730.
Bilaud T, Koering CE, Binet-Brasselet E, Ancelin K, Pollice A, Gasser SM, Gilson E (1996) The telobox, a Myb-related telomeric DNA binding motif found in proteins from yeast, plants and human. Nucleic Acids Res 24: 1294-1303.
Blackburn E (2006) A History of Telomere Biology. In: de Lange T, Lundblad V, Blackburn E (eds), Telomeres. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, pp. 1-19.
Blackburn EH, Gall JG (1978) A tandemly repeated sequence at the termini of the extrachromosomal ribosomal RNA genes in Tetrahymena. J Mol Biol 120: 33-53.
Booth C, Griffith E, Brady G, Lydall D (2001) Quantitative amplification of single-stranded DNA (QAOS) demonstrates that cdc13-1 mutants generate ssDNA in a telomere to centromere direction. Nucleic Acids Res 29: 4414-4422.
Boule JB, Vega LR, Zakian VA (2005) The yeast Pif1p helicase removes telomerase from telomeric DNA. Nature 438: 57-61.

References
112
Boule JB, Zakian VA (2007) The yeast Pif1p DNA helicase preferentially unwinds RNA DNA substrates. Nucleic Acids Res 35: 5809-5818.
Boulton SJ, Jackson SP (1998) Components of the Ku-dependent non-homologous end-joining pathway are involved in telomeric length maintenance and telomeric silencing. Embo J 17: 1819-1828.
Brevet V, Berthiau AS, Civitelli L, Donini P, Schramke V, Geli V, Ascenzioni F, Gilson E (2003) The number of vertebrate repeats can be regulated at yeast telomeres by Rap1-independent mechanisms. Embo J 22: 1697-1706.
Brigati C, Kurtz S, Balderes D, Vidali G, Shore D (1993) An essential yeast gene encoding a TTAGGG repeat-binding protein. Mol Cell Biol 13: 1306-1314.
Brown WR, MacKinnon PJ, Villasante A, Spurr N, Buckle VJ, Dobson MJ (1990) Structure and polymorphism of human telomere-associated DNA. Cell 63: 119-132.
Bryan TM, Englezou A, Gupta J, Bacchetti S, Reddel RR (1995) Telomere elongation in immortal human cells without detectable telomerase activity. Embo J 14: 4240-4248.
Budd ME, Reis CC, Smith S, Myung K, Campbell JL (2006) Evidence suggesting that Pif1 helicase functions in DNA replication with the Dna2 helicase/nuclease and DNA polymerase delta. Mol Cell Biol 26: 2490-2500.
Cano MI, Blake JJ, Blackburn EH, Agabian N (2002) A Trypanosoma brucei protein complex that binds G-overhangs and co-purifies with telomerase activity. J Biol Chem 277: 896-906.
Carson MJ, Hartwell L (1985) CDC17: an essential gene that prevents telomere elongation in yeast. Cell 42: 249-257.
Celli GB, Denchi EL, de Lange T (2006) Ku70 stimulates fusion of dysfunctional telomeres yet protects chromosome ends from homologous recombination. Nat Cell Biol 8: 885-890.
Cesare AJ, Quinney N, Willcox S, Subramanian D, Griffith JD (2003) Telomere looping in P. sativum (common garden pea). Plant J 36: 271-279.
Cesare AJ, Groff-Vindman C, Compton SA, McEachern MJ, Griffith JD (2007) T-loops and homologous recombination dependent t-circles in a Kluyveromyces lactis telomere mutant strain. Mol Cell Biol.
Chai W, Du Q, Shay JW, Wright WE (2006a) Human telomeres have different overhang sizes at leading versus lagging strands. Mol Cell 21: 427-435.
Chai W, Sfeir AJ, Hoshiyama H, Shay JW, Wright WE (2006b) The involvement of the Mre11/Rad50/Nbs1 complex in the generation of G-overhangs at human telomeres. EMBO Rep 7: 225-230.
Chan CS, Tye BK (1983a) A family of Saccharomyces cerevisiae repetitive autonomously replicating sequences that have very similar genomic environments. J Mol Biol 168: 505-523.
Chan CS, Tye BK (1983b) Organization of DNA sequences and replication origins at yeast telomeres. Cell 33: 563-573.
Chan SW, Chang J, Prescott J, Blackburn EH (2001) Altering telomere structure allows telomerase to act in yeast lacking ATM kinases. Curr Biol 11: 1240-1250.
Chandra A, Hughes TR, Nugent CI, Lundblad V (2001) Cdc13 both positively and negatively regulates telomere replication. Genes Dev 15: 404-414.
Chen Q, Ijpma A, Greider CW (2001) Two survivor pathways that allow growth in the absence of telomerase are generated by distinct telomere recombination events. Mol Cell Biol 21: 1819-1827.
Ciapponi L, Cenci G, Ducau J, Flores C, Johnson-Schlitz D, Gorski MM, Engels WR, Gatti M (2004) The Drosophila Mre11/Rad50 complex is required to prevent both telomeric fusion and chromosome breakage. Curr Biol 14: 1360-1366.
Cockell M, Palladino F, Laroche T, Kyrion G, Liu C, Lustig AJ, Gasser SM (1995) The carboxy termini of Sir4 and Rap1 affect Sir3 localization: evidence for a multicomponent complex required for yeast telomeric silencing. J Cell Biol 129: 909-924.
Cohn M, Blackburn EH (1995) Telomerase in yeast. Science 269: 396-400. Conrad MN, Wright JH, Wolf AJ, Zakian VA (1990) RAP1 protein interacts with yeast telomeres in vivo:
overproduction alters telomere structure and decreases chromosome stability. Cell 63: 739-750. Cooke HJ, Smith BA (1986) Variability at the telomeres of the human X/Y pseudoautosomal region.
Cold Spring Harb Symp Quant Biol 51 Pt 1: 213-219. Cosgrove AJ, Nieduszynski CA, Donaldson AD (2002) Ku complex controls the replication time of DNA
in telomere regions. Genes Dev 16: 2485-2490. Cristofari G, Lingner J (2006) Telomere length homeostasis requires that telomerase levels are
limiting. EMBO J 25: 565-574.

Referencess
113
d'Adda di Fagagna F, Hande MP, Tong WM, Roth D, Lansdorp PM, Wang ZQ, Jackson SP (2001) Effects of DNA nonhomologous end-joining factors on telomere length and chromosomal stability in mammalian cells. Curr Biol 11: 1192-1196.
d'Adda di Fagagna F, Teo SH, Jackson SP (2004) Functional links between telomeres and proteins of the DNA-damage response. Genes Dev 18: 1781-1799.
Dandjinou AT, Levesque N, Larose S, Lucier JF, Abou Elela S, Wellinger RJ (2004) A phylogenetically based secondary structure for the yeast telomerase RNA. Curr Biol 14: 1148-1158.
de Bruin D, Zaman Z, Liberatore RA, Ptashne M (2001) Telomere looping permits gene activation by a downstream UAS in yeast. Nature 409: 109-113.
de Lange T, Shiue L, Myers RM, Cox DR, Naylor SL, Killery AM, Varmus HE (1990) Structure and variability of human chromosome ends. Mol Cell Biol 10: 518-527.
de Lange T (2005) Shelterin: the protein complex that shapes and safeguards human telomeres. Genes Dev 19: 2100-2110.
Diede SJ, Gottschling DE (1999) Telomerase-mediated telomere addition in vivo requires DNA primase and DNA polymerases alpha and delta. Cell 99: 723-733.
Diede SJ, Gottschling DE (2001) Exonuclease activity is required for sequence addition and Cdc13p loading at a de novo telomere. Curr Biol 11: 1336-1340.
Dionne I, Wellinger RJ (1996) Cell cycle-regulated generation of single-stranded G-rich DNA in the absence of telomerase. Proc Natl Acad Sci U S A 93: 13902-13907.
Downs JA, Jackson SP (2004) A means to a DNA end: the many roles of Ku. Nat Rev Mol Cell Biol 5: 367-378.
Evans SK, Lundblad V (1999) Est1 and Cdc13 as comediators of telomerase access. Science 286: 117-120.
Falck J, Coates J, Jackson SP (2005) Conserved modes of recruitment of ATM, ATR and DNA-PKcs to sites of DNA damage. Nature 434: 605-611.
Feng J, Funk WD, Wang SS, Weinrich SL, Avilion AA, Chiu CP, Adams RR, Chang E, Allsopp RC, Yu J, et al. (1995) The RNA component of human telomerase. Science 269: 1236-1241.
Fisher TS, Taggart AK, Zakian VA (2004) Cell cycle-dependent regulation of yeast telomerase by Ku. Nat Struct Mol Biol 11: 1198-1205.
Forstemann K, Hoss M, Lingner J (2000) Telomerase-dependent repeat divergence at the 3' ends of yeast telomeres. Nucleic Acids Res 28: 2690-2694.
Forstemann K, Lingner J (2001) Molecular basis for telomere repeat divergence in budding yeast. Mol Cell Biol 21: 7277-7286.
Forstemann K, Lingner J (2005) Telomerase limits the extent of base pairing between template RNA and telomeric DNA. EMBO Rep 6: 361-366.
Fourel G, Revardel E, Koering CE, Gilson E (1999) Cohabitation of insulators and silencing elements in yeast subtelomeric regions. Embo J 18: 2522-2537.
Frank CJ, Hyde M, Greider CW (2006) Regulation of telomere elongation by the cyclin-dependent kinase CDK1. Mol Cell 24: 423-432.
Friedman KL, Heit JJ, Long DM, Cech TR (2003) N-terminal domain of yeast telomerase reverse transcriptase: recruitment of Est3p to the telomerase complex. Mol Biol Cell 14: 1-13.
Gao H, Cervantes RB, Mandell EK, Otero JH, Lundblad V (2007) RPA-like proteins mediate yeast telomere function. Nat Struct Mol Biol 14: 208-214.
Garber PM, Vidanes GM, Toczyski DP (2005) Damage in transition. Trends Biochem Sci 30: 63-66. Garcia-Cao M, O'Sullivan R, Peters AH, Jenuwein T, Blasco MA (2004) Epigenetic regulation of
telomere length in mammalian cells by the Suv39h1 and Suv39h2 histone methyltransferases. Nat Genet 36: 94-99.
Garvik B, Carson M, Hartwell L (1995) Single-stranded DNA arising at telomeres in cdc13 mutants may constitute a specific signal for the RAD9 checkpoint. Mol Cell Biol 15: 6128-6138.
Gatbonton T, Imbesi M, Nelson M, Akey JM, Ruderfer DM, Kruglyak L, Simon JA, Bedalov A (2006) Telomere length as a quantitative trait: genome-wide survey and genetic mapping of telomere length-control genes in yeast. PLoS Genet 2: e35.
Gilson E, Roberge M, Giraldo R, Rhodes D, Gasser SM (1993) Distortion of the DNA double helix by RAP1 at silencers and multiple telomeric binding sites. J Mol Biol 231: 293-310.
Gotta M, Laroche T, Formenton A, Maillet L, Scherthan H, Gasser SM (1996) The clustering of telomeres and colocalization with Rap1, Sir3, and Sir4 proteins in wild-type Saccharomyces cerevisiae. J Cell Biol 134: 1349-1363.
Goudsouzian LK, Tuzon CT, Zakian VA (2006) S. cerevisiae Tel1p and Mre11p are required for normal levels of Est1p and Est2p telomere association. Mol Cell 24: 603-610.

References
114
Grandin N, Reed SI, Charbonneau M (1997) Stn1, a new Saccharomyces cerevisiae protein, is implicated in telomere size regulation in association with Cdc13. Genes Dev 11: 512-527.
Grandin N, Damon C, Charbonneau M (2001) Ten1 functions in telomere end protection and length regulation in association with Stn1 and Cdc13. Embo J 20: 1173-1183.
Gravel S, Larrivee M, Labrecque P, Wellinger RJ (1998) Yeast Ku as a regulator of chromosomal DNA end structure. Science 280: 741-744.
Greenwell PW, Kronmal SL, Porter SE, Gassenhuber J, Obermaier B, Petes TD (1995) TEL1, a gene involved in controlling telomere length in S. cerevisiae, is homologous to the human ataxia telangiectasia gene. Cell 82: 823-829.
Greider CW, Blackburn EH (1985) Identification of a specific telomere terminal transferase activity in Tetrahymena extracts. Cell 43: 405-413.
Greider CW (1991) Telomerase is processive. Mol Cell Biol 11: 4572-4580. Griffith JD, Comeau L, Rosenfield S, Stansel RM, Bianchi A, Moss H, de Lange T (1999) Mammalian
telomeres end in a large duplex loop. Cell 97: 503-514. Grossi S, Puglisi A, Dmitriev PV, Lopes M, Shore D (2004) Pol12, the B subunit of DNA polymerase
alpha, functions in both telomere capping and length regulation. Genes Dev 18: 992-1006. Haber JE (1998) The many interfaces of Mre11. Cell 95: 583-586. Hardy CF, Balderes D, Shore D (1992a) Dissection of a carboxy-terminal region of the yeast regulatory
protein RAP1 with effects on both transcriptional activation and silencing. Mol Cell Biol 12: 1209-1217.
Hardy CF, Sussel L, Shore D (1992b) A RAP1-interacting protein involved in transcriptional silencing and telomere length regulation. Genes Dev 6: 801-814.
Harley CB, Futcher AB, Greider CW (1990) Telomeres shorten during ageing of human fibroblasts. Nature 345: 458-460.
Hathcock KS, Hemann MT, Opperman KK, Strong MA, Greider CW, Hodes RJ (2002) Haploinsufficiency of mTR results in defects in telomere elongation. Proc Natl Acad Sci U S A 99: 3591-3596.
Hecht A, Laroche T, Strahl-Bolsinger S, Gasser SM, Grunstein M (1995) Histone H3 and H4 N-termini interact with SIR3 and SIR4 proteins: a molecular model for the formation of heterochromatin in yeast. Cell 80: 583-592.
Hector RE, Shtofman RL, Ray A, Chen BR, Nyun T, Berkner KL, Runge KW (2007) Tel1p preferentially associates with short telomeres to stimulate their elongation. Mol Cell 27: 851-858.
Hediger F, Neumann FR, Van Houwe G, Dubrana K, Gasser SM (2002) Live imaging of telomeres: yKu and Sir proteins define redundant telomere-anchoring pathways in yeast. Curr Biol 12: 2076-2089.
Hediger F, Berthiau AS, van Houwe G, Gilson E, Gasser SM (2006) Subtelomeric factors antagonize telomere anchoring and Tel1-independent telomere length regulation. Embo J 25: 857-867.
Henry YA, Chambers A, Tsang JS, Kingsman AJ, Kingsman SM (1990) Characterisation of the DNA binding domain of the yeast RAP1 protein. Nucleic Acids Res 18: 2617-2623.
Herbig U, Jobling WA, Chen BP, Chen DJ, Sedivy JM (2004) Telomere shortening triggers senescence of human cells through a pathway involving ATM, p53, and p21(CIP1), but not p16(INK4a). Mol Cell 14: 501-513.
Holt SE, Aisner DL, Shay JW, Wright WE (1997) Lack of cell cycle regulation of telomerase activity in human cells. Proc Natl Acad Sci U S A 94: 10687-10692.
Hoppe GJ, Tanny JC, Rudner AD, Gerber SA, Danaie S, Gygi SP, Moazed D (2002) Steps in assembly of silent chromatin in yeast: Sir3-independent binding of a Sir2/Sir4 complex to silencers and role for Sir2-dependent deacetylation. Mol Cell Biol 22: 4167-4180.
Horvath MP, Schweiker VL, Bevilacqua JM, Ruggles JA, Schultz SC (1998) Crystal structure of the Oxytricha nova telomere end binding protein complexed with single strand DNA. Cell 95: 963-974.
Hsu HL, Gilley D, Blackburn EH, Chen DJ (1999) Ku is associated with the telomere in mammals. Proc Natl Acad Sci U S A 96: 12454-12458.
Hsu HL, Gilley D, Galande SA, Hande MP, Allen B, Kim SH, Li GC, Campisi J, Kohwi-Shigematsu T, Chen DJ (2000) Ku acts in a unique way at the mammalian telomere to prevent end joining. Genes Dev 14: 2807-2812.
Huffman KE, Levene SD, Tesmer VM, Shay JW, Wright WE (2000) Telomere shortening is proportional to the size of the G-rich telomeric 3'-overhang. J Biol Chem 275: 19719-19722.
Hug N, Lingner J (2006) Telomere length homeostasis. Chromosoma 115: 413-425.

Referencess
115
Hughes TR, Evans SK, Weilbaecher RG, Lundblad V (2000) The Est3 protein is a subunit of yeast telomerase. Curr Biol 10: 809-812.
Ivessa AS, Zhou JQ, Schulz VP, Monson EK, Zakian VA (2002) Saccharomyces Rrm3p, a 5' to 3' DNA helicase that promotes replication fork progression through telomeric and subtelomeric DNA. Genes Dev 16: 1383-1396.
Ivessa AS, Lenzmeier BA, Bessler JB, Goudsouzian LK, Schnakenberg SL, Zakian VA (2003) The Saccharomyces cerevisiae helicase Rrm3p facilitates replication past nonhistone protein-DNA complexes. Mol Cell 12: 1525-1536.
Jaco I, Munoz P, Blasco MA (2004) Role of human Ku86 in telomere length maintenance and telomere capping. Cancer Res 64: 7271-7278.
Jady BE, Richard P, Bertrand E, Kiss T (2006) Cell cycle-dependent recruitment of telomerase RNA and Cajal bodies to human telomeres. Mol Biol Cell 17: 944-954.
Jones MH, Guthrie C (1990) Unexpected flexibility in an evolutionarily conserved protein-RNA interaction: genetic analysis of the Sm binding site. Embo J 9: 2555-2561.
Kelleher C, Kurth I, Lingner J (2005) Human protection of telomeres 1 (POT1) is a negative regulator of telomerase activity in vitro. Mol Cell Biol 25: 808-818.
Kim SH, Kaminker P, Campisi J (1999) TIN2, a new regulator of telomere length in human cells. Nat Genet 23: 405-412.
Kipling D, Cooke HJ (1990) Hypervariable ultra-long telomeres in mice. Nature 347: 400-402. Kironmai KM, Muniyappa K (1997) Alteration of telomeric sequences and senescence caused by
mutations in RAD50 of Saccharomyces cerevisiae. Genes Cells 2: 443-455. Kishi S, Zhou XZ, Ziv Y, Khoo C, Hill DE, Shiloh Y, Lu KP (2001) Telomeric protein Pin2/TRF1 as an
important ATM target in response to double strand DNA breaks. J Biol Chem 276: 29282-29291. Klein F, Laroche T, Cardenas ME, Hofmann JF, Schweizer D, Gasser SM (1992) Localization of RAP1
and topoisomerase II in nuclei and meiotic chromosomes of yeast. J Cell Biol 117: 935-948. Koering CE, Fourel G, Binet-Brasselet E, Laroche T, Klein F, Gilson E (2000) Identification of high
affinity Tbf1p-binding sites within the budding yeast genome. Nucleic Acids Res 28: 2519-2526. Konig P, Giraldo R, Chapman L, Rhodes D (1996) The crystal structure of the DNA-binding domain of
yeast RAP1 in complex with telomeric DNA. Cell 85: 125-136. Krejci L, Van Komen S, Li Y, Villemain J, Reddy MS, Klein H, Ellenberger T, Sung P (2003) DNA
helicase Srs2 disrupts the Rad51 presynaptic filament. Nature 423: 305-309. Kyrion G, Boakye KA, Lustig AJ (1992) C-terminal truncation of RAP1 results in the deregulation of
telomere size, stability, and function in Saccharomyces cerevisiae. Mol Cell Biol 12: 5159-5173. Kyrion G, Liu K, Liu C, Lustig AJ (1993) RAP1 and telomere structure regulate telomere position
effects in Saccharomyces cerevisiae. Genes Dev 7: 1146-1159. Laroche T, Martin SG, Gotta M, Gorham HC, Pryde FE, Louis EJ, Gasser SM (1998) Mutation of yeast
Ku genes disrupts the subnuclear organization of telomeres. Curr Biol 8: 653-656. Larrivee M, LeBel C, Wellinger RJ (2004) The generation of proper constitutive G-tails on yeast
telomeres is dependent on the MRX complex. Genes Dev 18: 1391-1396. Lendvay TS, Morris DK, Sah J, Balasubramanian B, Lundblad V (1996) Senescence mutants of
Saccharomyces cerevisiae with a defect in telomere replication identify three additional EST genes. Genetics 144: 1399-1412.
Levy DL, Blackburn EH (2004) Counting of Rif1p and Rif2p on Saccharomyces cerevisiae telomeres regulates telomere length. Mol Cell Biol 24: 10857-10867.
Li B, Oestreich S, de Lange T (2000) Identification of human Rap1: implications for telomere evolution. Cell 101: 471-483.
Lieb JD, Liu X, Botstein D, Brown PO (2001) Promoter-specific binding of Rap1 revealed by genome-wide maps of protein-DNA association. Nat Genet 28: 327-334.
Lin JJ, Zakian VA (1996) The Saccharomyces CDC13 protein is a single-strand TG1-3 telomeric DNA-binding protein in vitro that affects telomere behavior in vivo. Proc Natl Acad Sci U S A 93: 13760-13765.
Lingner J, Cech TR, Hughes TR, Lundblad V (1997a) Three Ever Shorter Telomere (EST) genes are dispensable for in vitro yeast telomerase activity. Proc Natl Acad Sci U S A 94: 11190-11195.
Lingner J, Hughes TR, Shevchenko A, Mann M, Lundblad V, Cech TR (1997b) Reverse transcriptase motifs in the catalytic subunit of telomerase. Science 276: 561-567.
Liu D, Safari A, O'Connor MS, Chan DW, Laegeler A, Qin J, Songyang Z (2004) PTOP interacts with POT1 and regulates its localization to telomeres. Nat Cell Biol 6: 673-680.
Liu Y, Kha H, Ungrin M, Robinson MO, Harrington L (2002) Preferential maintenance of critically short telomeres in mammalian cells heterozygous for mTert. Proc Natl Acad Sci U S A 99: 3597-3602.

References
116
Liu ZP, Tye BK (1991) A yeast protein that binds to vertebrate telomeres and conserved yeast telomeric junctions. Genes Dev 5: 49-59.
Loayza D, De Lange T (2003) POT1 as a terminal transducer of TRF1 telomere length control. Nature 423: 1013-1018.
Longtine MS, Wilson NM, Petracek ME, Berman J (1989) A yeast telomere binding activity binds to two related telomere sequence motifs and is indistinguishable from RAP1. Curr Genet 16: 225-239.
Lundblad V, Szostak JW (1989) A mutant with a defect in telomere elongation leads to senescence in yeast. Cell 57: 633-643.
Lundblad V, Blackburn EH (1993) An alternative pathway for yeast telomere maintenance rescues est1- senescence. Cell 73: 347-360.
Lundblad V (2006) Budding Yeast Telomeres. In: de Lange T, Lundblad V, Blackburn E (eds), Telomeres. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, pp. 345-386.
Lustig AJ, Petes TD (1986) Identification of yeast mutants with altered telomere structure. Proc Natl Acad Sci U S A 83: 1398-1402.
Lustig AJ, Kurtz S, Shore D (1990) Involvement of the silencer and UAS binding protein RAP1 in regulation of telomere length. Science 250: 549-553.
Lydall D, Weinert T (1995) Yeast checkpoint genes in DNA damage processing: implications for repair and arrest. Science 270: 1488-1491.
Makarov VL, Lejnine S, Bedoyan J, Langmore JP (1993) Nucleosomal organization of telomere-specific chromatin in rat. Cell 73: 775-787.
Mangahas JL, Alexander MK, Sandell LL, Zakian VA (2001) Repair of chromosome ends after telomere loss in Saccharomyces. Mol Biol Cell 12: 4078-4089.
Marcand S, Gilson E, Shore D (1997a) A protein-counting mechanism for telomere length regulation in yeast. Science 275: 986-990.
Marcand S, Wotton D, Gilson E, Shore D (1997b) Rap1p and telomere length regulation in yeast. Ciba Found Symp 211: 76-93; discussion 93-103.
Marcand S, Brevet V, Gilson E (1999) Progressive cis-inhibition of telomerase upon telomere elongation. Embo J 18: 3509-3519.
Marcand S, Brevet V, Mann C, Gilson E (2000) Cell cycle restriction of telomere elongation. Curr Biol 10: 487-490.
Maringele L, Lydall D (2002) EXO1-dependent single-stranded DNA at telomeres activates subsets of DNA damage and spindle checkpoint pathways in budding yeast yku70Delta mutants. Genes Dev 16: 1919-1933.
McClintock B (1931) Cytological observation of deficiencies involving known genes, translocations and an inversion in Zea mays. Mo Agric Exp Res Stn Res Bull 163: 4-30.
McClintock B (1939) The Behavior in Successive Nuclear Divisions of a Chromosome Broken at Meiosis. Proc Natl Acad Sci U S A 25: 405-416.
Metcalfe JA, Parkhill J, Campbell L, Stacey M, Biggs P, Byrd PJ, Taylor AM (1996) Accelerated telomere shortening in ataxia telangiectasia. Nat Genet 13: 350-353.
Meyne J, Baker RJ, Hobart HH, Hsu TC, Ryder OA, Ward OG, Wiley JE, Wurster-Hill DH, Yates TL, Moyzis RK (1990) Distribution of non-telomeric sites of the (TTAGGG)n telomeric sequence in vertebrate chromosomes. Chromosoma 99: 3-10.
Mishra K, Shore D (1999) Yeast Ku protein plays a direct role in telomeric silencing and counteracts inhibition by rif proteins. Curr Biol 9: 1123-1126.
Mitton-Fry RM, Anderson EM, Hughes TR, Lundblad V, Wuttke DS (2002) Conserved structure for single-stranded telomeric DNA recognition. Science 296: 145-147.
Moretti P, Freeman K, Coodly L, Shore D (1994) Evidence that a complex of SIR proteins interacts with the silencer and telomere-binding protein RAP1. Genes Dev 8: 2257-2269.
Moyzis RK, Buckingham JM, Cram LS, Dani M, Deaven LL, Jones MD, Meyne J, Ratliff RL, Wu JR (1988) A highly conserved repetitive DNA sequence, (TTAGGG)n, present at the telomeres of human chromosomes. Proc Natl Acad Sci U S A 85: 6622-6626.
Mozdy AD, Cech TR (2006) Low abundance of telomerase in yeast: implications for telomerase haploinsufficiency. Rna 12: 1721-1737.
Muller HJ (1938) The remaking of chromosomes. The Collecting Net 8: 182-195. Munoz-Jordan JL, Cross GA, de Lange T, Griffith JD (2001) t-loops at trypanosome telomeres. Embo J
20: 579-588. Murti KG, Prescott DM (1999) Telomeres of polytene chromosomes in a ciliated protozoan terminate
in duplex DNA loops. Proc Natl Acad Sci U S A 96: 14436-14439.

Referencess
117
Myung K, Chen C, Kolodner RD (2001) Multiple pathways cooperate in the suppression of genome instability in Saccharomyces cerevisiae. Nature 411: 1073-1076.
Myung K, Ghosh G, Fattah FJ, Li G, Kim H, Dutia A, Pak E, Smith S, Hendrickson EA (2004) Regulation of telomere length and suppression of genomic instability in human somatic cells by Ku86. Mol Cell Biol 24: 5050-5059.
Nakada D, Matsumoto K, Sugimoto K (2003) ATM-related Tel1 associates with double-strand breaks through an Xrs2-dependent mechanism. Genes Dev 17: 1957-1962.
Nakamura TM, Morin GB, Chapman KB, Weinrich SL, Andrews WH, Lingner J, Harley CB, Cech TR (1997) Telomerase catalytic subunit homologs from fission yeast and human. Science 277: 955-959.
Nugent CI, Hughes TR, Lue NF, Lundblad V (1996) Cdc13p: a single-strand telomeric DNA-binding protein with a dual role in yeast telomere maintenance. Science 274: 249-252.
Nugent CI, Bosco G, Ross LO, Evans SK, Salinger AP, Moore JK, Haber JE, Lundblad V (1998) Telomere maintenance is dependent on activities required for end repair of double-strand breaks. Curr Biol 8: 657-660.
O'Connor MS, Safari A, Liu D, Qin J, Songyang Z (2004) The human Rap1 protein complex and modulation of telomere length. J Biol Chem 279: 28585-28591.
Osterhage JL, Talley JM, Friedman KL (2006) Proteasome-dependent degradation of Est1p regulates the cell cycle-restricted assembly of telomerase in Saccharomyces cerevisiae. Nat Struct Mol Biol 13: 720-728.
Pardo B, Marcand S (2005) Rap1 prevents telomere fusions by nonhomologous end joining. Embo J 24: 3117-3127.
Pennock E, Buckley K, Lundblad V (2001) Cdc13 delivers separate complexes to the telomere for end protection and replication. Cell 104: 387-396.
Peterson SE, Stellwagen AE, Diede SJ, Singer MS, Haimberger ZW, Johnson CO, Tzoneva M, Gottschling DE (2001) The function of a stem-loop in telomerase RNA is linked to the DNA repair protein Ku. Nat Genet 27: 64-67.
Polotnianka RM, Li J, Lustig AJ (1998) The yeast Ku heterodimer is essential for protection of the telomere against nucleolytic and recombinational activities. Curr Biol 8: 831-834.
Porter SE, Greenwell PW, Ritchie KB, Petes TD (1996) The DNA-binding protein Hdf1p (a putative Ku homologue) is required for maintaining normal telomere length in Saccharomyces cerevisiae. Nucleic Acids Res 24: 582-585.
Prescott J, Blackburn EH (1997a) Telomerase RNA mutations in Saccharomyces cerevisiae alter telomerase action and reveal nonprocessivity in vivo and in vitro. Genes Dev 11: 528-540.
Prescott J, Blackburn EH (1997b) Functionally interacting telomerase RNAs in the yeast telomerase complex. Genes Dev 11: 2790-2800.
Pryde FE, Gorham HC, Louis EJ (1997) Chromosome ends: all the same under their caps. Curr Opin Genet Dev 7: 822-828.
Qi H, Zakian VA (2000) The Saccharomyces telomere-binding protein Cdc13p interacts with both the catalytic subunit of DNA polymerase alpha and the telomerase-associated est1 protein. Genes Dev 14: 1777-1788.
Raghuraman MK, Brewer BJ, Fangman WL (1997) Cell cycle-dependent establishment of a late replication program. Science 276: 806-809.
Ranganathan V, Heine WF, Ciccone DN, Rudolph KL, Wu X, Chang S, Hai H, Ahearn IM, Livingston DM, Resnick I, Rosen F, Seemanova E, Jarolim P, DePinho RA, Weaver DT (2001) Rescue of a telomere length defect of Nijmegen breakage syndrome cells requires NBS and telomerase catalytic subunit. Curr Biol 11: 962-966.
Ray A, Runge KW (1999) Varying the number of telomere-bound proteins does not alter telomere length in tel1Delta cells. Proc Natl Acad Sci U S A 96: 15044-15049.
Raymond WE, Kleckner N (1993) RAD50 protein of S.cerevisiae exhibits ATP-dependent DNA binding. Nucleic Acids Res 21: 3851-3856.
Reichenbach P, Hoss M, Azzalin CM, Nabholz M, Bucher P, Lingner J (2003) A human homolog of yeast Est1 associates with telomerase and uncaps chromosome ends when overexpressed. Curr Biol 13: 568-574.
Riethman H, Ambrosini A, Castaneda C, Finklestein J, Hu XL, Mudunuri U, Paul S, Wei J (2004) Mapping and initial analysis of human subtelomeric sequence assemblies. Genome Res 14: 18-28.

References
118
Ritchie KB, Mallory JC, Petes TD (1999) Interactions of TLC1 (which encodes the RNA subunit of telomerase), TEL1, and MEC1 in regulating telomere length in the yeast Saccharomyces cerevisiae. Mol Cell Biol 19: 6065-6075.
Ritchie KB, Petes TD (2000) The Mre11p/Rad50p/Xrs2p complex and the Tel1p function in a single pathway for telomere maintenance in yeast. Genetics 155: 475-479.
Roy R, Meier B, McAinsh AD, Feldmann HM, Jackson SP (2004) Separation-of-function mutants of yeast Ku80 reveal a Yku80p-Sir4p interaction involved in telomeric silencing. J Biol Chem 279: 86-94.
Rusche LN, Kirchmaier AL, Rine J (2003) The establishment, inheritance, and function of silenced chromatin in Saccharomyces cerevisiae. Annu Rev Biochem 72: 481-516.
Sabourin M, Tuzon CT, Zakian VA (2007) Telomerase and Tel1p Preferentially Associate with Short Telomeres in S. cerevisiae. Mol Cell.
Samper E, Goytisolo FA, Slijepcevic P, van Buul PP, Blasco MA (2000) Mammalian Ku86 protein prevents telomeric fusions independently of the length of TTAGGG repeats and the G-strand overhang. EMBO Rep 1: 244-252.
Schramke V, Luciano P, Brevet V, Guillot S, Corda Y, Longhese MP, Gilson E, Geli V (2004) RPA regulates telomerase action by providing Est1p access to chromosome ends. Nat Genet 36: 46-54.
Schulz VP, Zakian VA (1994) The saccharomyces PIF1 DNA helicase inhibits telomere elongation and de novo telomere formation. Cell 76: 145-155.
Seto AG, Zaug AJ, Sobel SG, Wolin SL, Cech TR (1999) Saccharomyces cerevisiae telomerase is an Sm small nuclear ribonucleoprotein particle. Nature 401: 177-180.
Seto AG, Livengood AJ, Tzfati Y, Blackburn EH, Cech TR (2002) A bulged stem tethers Est1p to telomerase RNA in budding yeast. Genes Dev 16: 2800-2812.
Shampay J, Szostak JW, Blackburn EH (1984) DNA sequences of telomeres maintained in yeast. Nature 310: 154-157.
Shore D, Nasmyth K (1987) Purification and cloning of a DNA binding protein from yeast that binds to both silencer and activator elements. Cell 51: 721-732.
Shore D, Stillman DJ, Brand AH, Nasmyth KA (1987) Identification of silencer binding proteins from yeast: possible roles in SIR control and DNA replication. Embo J 6: 461-467.
Silverman J, Takai H, Buonomo SB, Eisenhaber F, de Lange T (2004) Human Rif1, ortholog of a yeast telomeric protein, is regulated by ATM and 53BP1 and functions in the S-phase checkpoint. Genes Dev 18: 2108-2119.
Singer MS, Gottschling DE (1994) TLC1: template RNA component of Saccharomyces cerevisiae telomerase. Science 266: 404-409.
Smogorzewska A, van Steensel B, Bianchi A, Oelmann S, Schaefer MR, Schnapp G, de Lange T (2000) Control of human telomere length by TRF1 and TRF2. Mol Cell Biol 20: 1659-1668.
Smolka MB, Albuquerque CP, Chen SH, Zhou H (2007) Proteome-wide identification of in vivo targets of DNA damage checkpoint kinases. Proc Natl Acad Sci U S A 104: 10364-10369.
Snow BE, Erdmann N, Cruickshank J, Goldman H, Gill RM, Robinson MO, Harrington L (2003) Functional conservation of the telomerase protein Est1p in humans. Curr Biol 13: 698-704.
Song K, Jung D, Jung Y, Lee SG, Lee I (2000) Interaction of human Ku70 with TRF2. FEBS Lett 481: 81-85.
Starling JA, Maule J, Hastie ND, Allshire RC (1990) Extensive telomere repeat arrays in mouse are hypervariable. Nucleic Acids Res 18: 6881-6888.
Stellwagen AE, Haimberger ZW, Veatch JR, Gottschling DE (2003) Ku interacts with telomerase RNA to promote telomere addition at native and broken chromosome ends. Genes Dev 17: 2384-2395.
Stracker TH, Theunissen JW, Morales M, Petrini JH (2004) The Mre11 complex and the metabolism of chromosome breaks: the importance of communicating and holding things together. DNA Repair (Amst) 3: 845-854.
Strahl-Bolsinger S, Hecht A, Luo K, Grunstein M (1997) SIR2 and SIR4 interactions differ in core and extended telomeric heterochromatin in yeast. Genes Dev 11: 83-93.
Sussel L, Shore D (1991) Separation of transcriptional activation and silencing functions of the RAP1-encoded repressor/activator protein 1: isolation of viable mutants affecting both silencing and telomere length. Proc Natl Acad Sci U S A 88: 7749-7753.
Szostak JW, Blackburn EH (1982) Cloning yeast telomeres on linear plasmid vectors. Cell 29: 245-255.
Taggart AK, Teng SC, Zakian VA (2002) Est1p as a cell cycle-regulated activator of telomere-bound telomerase. Science 297: 1023-1026.

Referencess
119
Takai H, Smogorzewska A, de Lange T (2003) DNA damage foci at dysfunctional telomeres. Curr Biol 13: 1549-1556.
Teixeira MT, Arneric M, Sperisen P, Lingner J (2004) Telomere length homeostasis is achieved via a switch between telomerase- extendible and -nonextendible states. Cell 117: 323-335.
Teng SC, Zakian VA (1999) Telomere-telomere recombination is an efficient bypass pathway for telomere maintenance in Saccharomyces cerevisiae. Mol Cell Biol 19: 8083-8093.
Teng SC, Chang J, McCowan B, Zakian VA (2000) Telomerase-independent lengthening of yeast telomeres occurs by an abrupt Rad50p-dependent, Rif-inhibited recombinational process. Mol Cell 6: 947-952.
Theobald DL, Wuttke DS (2004) Prediction of multiple tandem OB-fold domains in telomere end-binding proteins Pot1 and Cdc13. Structure 12: 1877-1879.
Tomlinson RL, Ziegler TD, Supakorndej T, Terns RM, Terns MP (2006) Cell cycle-regulated trafficking of human telomerase to telomeres. Mol Biol Cell 17: 955-965.
Trujillo KM, Roh DH, Chen L, Van Komen S, Tomkinson A, Sung P (2003) Yeast xrs2 binds DNA and helps target rad50 and mre11 to DNA ends. J Biol Chem 278: 48957-48964.
Tsai YL, Tseng SF, Chang SH, Lin CC, Teng SC (2002) Involvement of replicative polymerases, Tel1p, Mec1p, Cdc13p, and the Ku complex in telomere-telomere recombination. Mol Cell Biol 22: 5679-5687.
Tseng SF, Lin JJ, Teng SC (2006) The telomerase-recruitment domain of the telomere binding protein Cdc13 is regulated by Mec1p/Tel1p-dependent phosphorylation. Nucleic Acids Res 34: 6327-6336.
Tsukamoto Y, Kato J, Ikeda H (1997) Silencing factors participate in DNA repair and recombination in Saccharomyces cerevisiae. Nature 388: 900-903.
Tsukamoto Y, Taggart AK, Zakian VA (2001) The role of the Mre11-Rad50-Xrs2 complex in telomerase- mediated lengthening of Saccharomyces cerevisiae telomeres. Curr Biol 11: 1328-1335.
Ubersax JA, Woodbury EL, Quang PN, Paraz M, Blethrow JD, Shah K, Shokat KM, Morgan DO (2003) Targets of the cyclin-dependent kinase Cdk1. Nature 425: 859-864.
Varley H, Pickett HA, Foxon JL, Reddel RR, Royle NJ (2002) Molecular characterization of inter-telomere and intra-telomere mutations in human ALT cells. Nat Genet 30: 301-305.
Veaute X, Jeusset J, Soustelle C, Kowalczykowski SC, Le Cam E, Fabre F (2003) The Srs2 helicase prevents recombination by disrupting Rad51 nucleoprotein filaments. Nature 423: 309-312.
Verdun RE, Crabbe L, Haggblom C, Karlseder J (2005) Functional human telomeres are recognized as DNA damage in G2 of the cell cycle. Mol Cell 20: 551-561.
Vodenicharov MD, Wellinger RJ (2006) DNA degradation at unprotected telomeres in yeast is regulated by the CDK1 (Cdc28/Clb) cell-cycle kinase. Mol Cell 24: 127-137.
Wang SS, Zakian VA (1990) Sequencing of Saccharomyces telomeres cloned using T4 DNA polymerase reveals two domains. Mol Cell Biol 10: 4415-4419.
Weinert TA, Hartwell LH (1993) Cell cycle arrest of cdc mutants and specificity of the RAD9 checkpoint. Genetics 134: 63-80.
Wellinger RJ, Wolf AJ, Zakian VA (1993) Saccharomyces telomeres acquire single-strand TG1-3 tails late in S phase. Cell 72: 51-60.
Wilkie AO, Higgs DR, Rack KA, Buckle VJ, Spurr NK, Fischel-Ghodsian N, Ceccherini I, Brown WR, Harris PC (1991) Stable length polymorphism of up to 260 kb at the tip of the short arm of human chromosome 16. Cell 64: 595-606.
Wiltzius JJ, Hohl M, Fleming JC, Petrini JH (2005) The Rad50 hook domain is a critical determinant of Mre11 complex functions. Nat Struct Mol Biol 12: 403-407.
Wold MS (1997) Replication protein A: a heterotrimeric, single-stranded DNA-binding protein required for eukaryotic DNA metabolism. Annu Rev Biochem 66: 61-92.
Wotton D, Shore D (1997) A novel Rap1p-interacting factor, Rif2p, cooperates with Rif1p to regulate telomere length in Saccharomyces cerevisiae. Genes Dev 11: 748-760.
Wu Y, Xiao S, Zhu XD (2007) MRE11-RAD50-NBS1 and ATM function as co-mediators of TRF1 in telomere length control. Nat Struct Mol Biol 14: 832-840.
Xin H, Liu D, Wan M, Safari A, Kim H, Sun W, O'Connor MS, Songyang Z (2007) TPP1 is a homologue of ciliate TEBP-beta and interacts with POT1 to recruit telomerase. Nature 445: 559-562.
Yang Q, Zheng YL, Harris CC (2005) POT1 and TRF2 cooperate to maintain telomeric integrity. Mol Cell Biol 25: 1070-1080.
Ye JZ, de Lange T (2004) TIN2 is a tankyrase 1 PARP modulator in the TRF1 telomere length control complex. Nat Genet 36: 618-623.

References
120
Ye JZ, Donigian JR, van Overbeek M, Loayza D, Luo Y, Krutchinsky AN, Chait BT, de Lange T (2004a) TIN2 binds TRF1 and TRF2 simultaneously and stabilizes the TRF2 complex on telomeres. J Biol Chem 279: 47264-47271.
Ye JZ, Hockemeyer D, Krutchinsky AN, Loayza D, Hooper SM, Chait BT, de Lange T (2004b) POT1-interacting protein PIP1: a telomere length regulator that recruits POT1 to the TIN2/TRF1 complex. Genes Dev 18: 1649-1654.
Zappulla DC, Cech TR (2004) Yeast telomerase RNA: a flexible scaffold for protein subunits. Proc Natl Acad Sci U S A 101: 10024-10029.
Zhang DH, Zhou B, Huang Y, Xu LX, Zhou JQ (2006) The human Pif1 helicase, a potential Escherichia coli RecD homologue, inhibits telomerase activity. Nucleic Acids Res 34: 1393-1404.
Zhou J, Monson EK, Teng SC, Schulz VP, Zakian VA (2000) Pif1p helicase, a catalytic inhibitor of telomerase in yeast. Science 289: 771-774.
Zhu L, Hathcock KS, Hande P, Lansdorp PM, Seldin MF, Hodes RJ (1998) Telomere length regulation in mice is linked to a novel chromosome locus. Proc Natl Acad Sci U S A 95: 8648-8653.
Zhu XD, Kuster B, Mann M, Petrini JH, de Lange T (2000) Cell-cycle-regulated association of RAD50/MRE11/NBS1 with TRF2 and human telomeres. Nat Genet 25: 347-352.