Embed Size (px)
Citation preview

JOURNAL OF BACTERIOLOGY, Jan. 1971, p. 349-354Copyright e 1971 American Society for Microbiology
Vol. 105, No. IPrinted in U.S.A.
Taxonomic Implications of Spore Fine Structurein Clostridium bifermentans
L. J. RODE AND LOUIS DS. SMITH
Department ofMicrobiology, University of Texas, Austin, Texas 78712, and A naerobe Laboratory, VirginiaPolytechnic Institute, Blacksburg, Virginia 24060.
Received for publication I October 1970
Thirty-five strains of Clostridium bifermentans were, in most part, culturallyhomogeneous by conventional taxonomic criteria but were heterogeneous with re-
spect to spore fine structure. Fourteen of the strains produced spores with appen-
dages, distributed among four distinct ultrastructural types. No consistent correla-tion existed between spore type and other variable properties of these strains. It isproposed, therefore, that these spore appendage-type strains be considered as "vari-eties" of C. bifermentans and that they should not be designated as new species.
Gross spore features which are readily detect-able with the light microscope have historicallyplayed an important and proper role in speciationwithin the Bacillus and Clostridium genera. Dis-tinctive attributes such as spore shape, spore size,and location in the sporangium are consideredrelevant in taxonomy of the Bacillaceae (Bergey'sManual, 7th ed.)
Spore morphological diversity which extendsbeyond such general gross features is now recog-nized, though not generally utilized, for taxo-nomic purposes. The elegant, early electronmicroscope study of Franklin and Bradley (1)using carbon replicas documented such diversityfor some Bacillus species, and additional detailhas been provided through use of the freeze-etchtechnique (3).
Additional, even more pronounced, spore mor-phological diversity became evident with the re-port by Krasil'nikov, Duda, and Sokolov (5) ofelaborate protrusions (appendages) on spores ofClostridium isolates. Two ensuing developmentsin this immediate area appear to have importanttaxonomic implications: (i) the creation of newClostridium species with spore ultrastructure(i.e., spore appendage status), the apparent soleor primary criterion for speciation (6) and (ii) thediscovery that multiple spore appendage typesoccur within presently established species such asC. botulinum (2) and C. bifermentans (2, 8, 14).Thus, what role, if any, should properly be as-signed to spore ultrastructure features, such asthe possession of appendages, in the taxonomy ofsporeforming bacteria?
Since at least five distinctive spore morpholog-ical types were already known for C. bifermen-tans (8, 14), we have, in the present study, chosen
this species of known diversity for more intensivestudy. Thirty-five strains from diverse sources,including those for which electron microscopestudies had already been performed (2, 8, 14),have been characterized by both conventionaltaxonomic methods and by electron microscopeappraisal of spore fine structure. Our objectivehas been to provide experimental rationale forproper use of the details of spore ultrastructure inthe taxonomy of this species.
MATERIALS AND METHODS
C. bifermentans strains. Thirty-five strains of C.bifermentans were studied. Strain designations, strainidentifications, and strain sources are listed in Table 1.
Electron microscope appraisal of spores. The organ-isms were grown to sporulation at 30 C on the surface of2% agar plates of Brain Heart Infusion (Difco) medium,supplemented with 0.5 g per liter of sodium thioglyco-late, in desiccators with wet oats to provide an anaerobicenvironment (10).
For carbon replicas, free spores from growth plateswere washed six times with demineralized water, placedon mica squares, shadowed with platinum, and coatedwith carbon. After flotation onto water and dissolutionof the cellular matter with 0.5% sodium hypochlorite,the replica films were washed and transferred to coppergrids for examination.
For thin sections, specimens were used directly fromgrowth plates. Preliminary fixation was for 1 hr at 4 Cin 0.5% glutaraldehyde (I 1) and was followed by routineKellenberger osmium fixation at 4 C for 17 hr (4). Thespecimens were embedded in a plastic mixture of do-decenyl succinic anhydride, Araldite 6005, and Epon 812.Sections, cut on a Porter-Blum MT-2 microtome (IvanSorvall, Inc., Norwalk, Conn.) fitted with a diamondknife, were stained for 5 to 60 sec with Reynolds leadcitrate ( 19).An Hitachi HS-7S electron microscope with double
349
on May 28, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

RODE AND SMITH
TABLE 1. Clostridium bifermentans straindesignations, identification, and source
Strain Other or SSanj rio strain ]Source
ATCC 638CDC 258CDC 352
McClung collec-tion 134
McClung collec-tion 135
ATCC 638
SDH 9cSDH lACSDH 2ACSDH 3AcSDH 4AcFDA lcMcClung collec-
tion 431cFDA U llcFDA Ul5cFDA U-27cFDA U-49c
TRS 288Bd
TRS 244Bd
TRS 247Bd
NCIB 6800dNCIB 506dNCIB 1341dNCIB 2929dNCIB 6928dTRS 275Bd
Sterility test of tuberculinHuman infectionHuman infectionCanned tomatoes
Lagoon mudIsolated from Beerenscollection 544
Isolated from Beerenscollection 544
Dust on hardware partsSterility test of tuberculinMarine sedimentMarine sedimentStomach ulcer of goat
Sediment
SedimentSedimentSedimentSedimentHuman infectionVacuum-packedsmoked fish
Vacuum-packedsmoked fish
Vacuum-packedsmoked fish
Vacuum-packedsmoked fish
a Accession numbers of the Anaerobe Laboratory,Virginia Polytechnic Institute, Blacksburg, Va., whichidentify the C. bifermentans strains for the presentwork.
bAbsence of designation indicates that other or priordesignation does not exist or is not known. ATCC,American Type Culture Collection; CDC, Center forDisease Control; SDH, Texas State Department ofHealth; FDA, Food and Drug Administration; TRS,Torry Research Station; NCIB, National Collection ofIndustrial Bacteria.
c Strain designations used in two prior spore appen-dage studies (8, 14).
d The spore appendage status of these strains hasbeen investigated (2).
condenser and 50-kv accelerating voltage was used forspecimen examination. Micrographs were taken onKodak contrast process Ortho film. Spore dimensionswere determined by measurements performed on printsof suitable magnification.
Conventional taxonomic appraisal of strains. With afew minor exceptions, the media and methods used forstudying these strains were those of Moore, Cato, andHoldeman (7), and Smith and Holdeman (13).
RESULTSConventional taxonomic appraisal: constant
features. The 35 strains were identical in the fol-lowing respects: All strains were gram-positiverods with central to subterminal spores (lightmicroscopy), liquefied gelatin, digested casein inmilk, digested cooked-meat medium with theproduction of ammonia, formed indole, fer-mented glucose, and produced lecithinase on eggyolk agar. No strain fermented sorbose, galac-tose, xylose, arabinose, rhamnose, sucrose, lac-tose, trehalose, cellobiose, raffinose, dextrin,starch, glycogen, inulin, cellulose, erythritol,adonitol, mannitol, dulcitol, salicin, esculin,amygdalin, or inositol. No strain reduced nitrate,produced acetylmethylcarbinol, produced ureaseor lipase, formed pigment, or produced toxin de-tectable in mice. The diaminopimelic acid in thevegetative cell wall was of the DL form. Fermen-tation products from a peptone, yeast extractglucose medium included acetic, propionic, isobu-tyric, isovaleric, and isocaproic acids, with orwithout small amounts of butyric or heptanoicacids.
Conventional taxonomic appraisal; variable fea-tures. Variability among the 35 strains was ob-served with respect to the following: fermentationof fructose, mannose, ribose, maltose, glycerol,and sorbitol; the production of H2S; motility; cellwall sugars. These data are shown in Table 2.
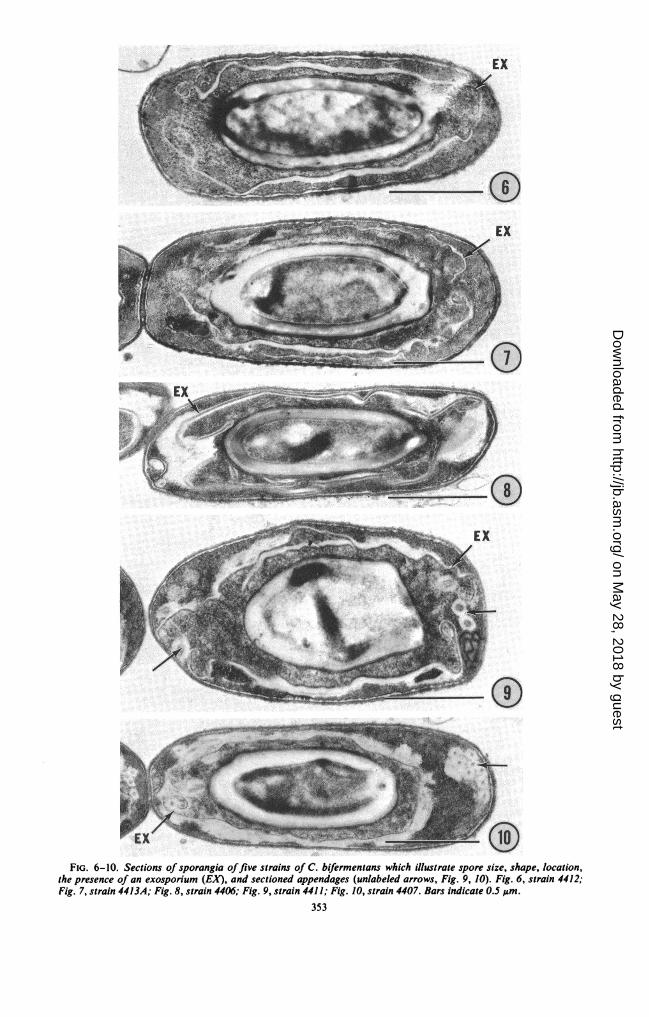
Electron microscopy; constant spore features.The spores of all 35 strains possessed an exo-sporium which was detected both in carbon rep-licas of free spores (Fig. 1-5) and in sections ofsporangia (Fig. 6-10). The spores were locatedcentrally to subterminally in sporangia (Fig. 6-10) and were oval to elongated (Fig. I-10). In nocase was a terminal spore observed. Measure-ments of spore size carried out on electron micro-graphs indicated that spore size, both length andwidth, was variable over a fairly narrow range;however, size variability between strains did notexceed size variability of spore populations withinstrains.
Electron microscopy; variable spore features.Significant ultrastructural variability amongspores of different strains was limited to thespore appendage aspect (Fig. 1-5; Table 2). Onthis basis, five spore morphological types were
20122035203624502658
2659
27082825A
2825-1
282928953036B31314131A4406440744084409441044114412
4413A44144415441644384669A
4670
4671
4672A467346744675A4676A4677A
350 J. BACTERIOL.
on May 28, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

VOL. 105, 1971 SPORE FINE STRUCTURE IN C. BIFERMENTANS
TABLE. 2. Variable properties of thirty-five strains of Clostridium bifermentans
T Fermentation of
Strain Type of spore H,S Vegetative cellno. appendage Fruc- Man- Ri- Mal- Gly- Sor- pro- Motil- wall sugars
tose nose bose tose cerol bitol duc- itytion
4413A Pinlike (Fig. 2)a4416 Pinlike
Smooth, tubular (Fig. 3)bSmooth, tubularSmooth, tubular
Hirsute, tubular (Fig. 4)a
Hirsute, tubularHirsute, tubularHirsute, tubularHirsute, tubular
Featherlike (Fig. 5)b
FeatherlikeFeatherlikeFeatherlike
None (Fig. 1)NoneNoneNoneNoneNoneNone
NoneNone
NoneNoneNoneNoneNoneNoneNone
NoneNoneNoneNoneNone
++ -- +
+
_
_ + _ + + - + +
+ + - - - - + +
_- - + + +
_- - + + + +
_ + _ - + + + +
_+ + - + +
_- - + + + +
_-
- + + +
+ -
_-
+ _
+ + +
+ + - - - + - +
+ _- - + + +
+ +
+ - - + - +
+ +
_+ + - - - + +
+ _- - + +
_-
+ + - - - - + +
+ + - - - - + +
+ + + - + + + +
+ + - - +- + + +
+ + - - + - + +
Glucose
Glucose
Glucose, rhamnose,mannose
Glucose, rhamnose,mannose
Glucose, rhamnose,galactose
Glucose, rhamnose,mannose
a For ultrastructure characterization see Yolton et al. (14).b For ultrastructure characterization see Pope et al. (8).
observed: (i) spores lacking appendages (Fig. 1; are distinctive and readily distinguishable. Expe-21 strains); (ii) spores with pinlike appendages rience to date indicates that spore appendage(Fig. 2; 2 strains); (iii) spores with smooth tu- status is a constant strain property. Multiplebular appendages (Fig. 3; 3 strains); (iv) spores appendage types within single strains have notwith hirsute tubular appendages (Fig. 4; 5 strains); been observed. No "new" spore appendage types(v) spores with featherlike appendages (Fig. 5; 4 were detected in the present study.strains). These five spore types, which correspond Lack of correlation between conventional taxo-in ultrastructure and in descriptive terminology nomic variables and spore appendage status. Thoseto the five spore types described in previous features which were found to be variable for thestudies of 12 strains of C. bifermentans (8, 14), 35 strains are summarized in Table 2. No corre-
351
440624502658
4411
4669A467046714677A
4407
440844094410
4412201220352036265927082825A
2825-12829
28953036B31314131A441444154438
4672A467346744675A4676A
on May 28, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

t-E-es^r
I0. , . . , . . . .-. :.s. :. ............ :. .ss, ... - Z S- z *, # - ^ s * -lyw . ,.,'-'; .......... ';' .,'. '' ........ ; i........... ...... _^ . - - . . _
.. P , _, .w ^: ^
,' '' ._ 'ZL ', *' .,- t
s<- ; j-r'_ w- ;l '-|>i-z2C; =
,*-.e
,i tacrs z \- s l o .S . . f." '_
..fS rS
.:^. t.> *-.. . : . , # :
;..v^
' *;_. . .';., '';. P^!'.W .. ,, :
.. o_,,l
4Fo: .. :.
., .. _ge
,2e,f,0
ofL;e' --/o-Js
_fif - etS .- EX*-I .-M. .:
- .f * KI.).
b-:01~ ~ ~ L
FIG. 1-5. Replicas of the free spores offive strains of C. bifermentans showing exosporium (EX) and appen-
dages (unlabeled arrows). Fig. 1, strain 4412, spore lacks appendages; Fig. 2, strain 4413A, spore has pinlike ap-
pendages; Fig. 3, strain 4406, spore has smooth tubular appendages; Fig. 4, strain 4411, spore has hirsute tubularappendages; Fig. 5, strain 4407, spore has featherlike appendages. Bars indicate 0.5 im.
352
0
i..- .'l.
I -.7.
on May 28, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
/1
EX
....
v0FIG. 6-10. Sections of sporangia offive strains of C. bifermentans which illustrate spore size, shape, location,
the presence of an exosporium (EX), and sectioned appendages (unlabeled arrows, Fig. 9, 10). Fig. 6, strain 4412;Fig. 7, strain 4413A; Fig. 8, strain 4406; Fig. 9, strain 4411; Fig. 10, strain 4407. Bars indicate 0.S ,um.
353
9=1.. 7;;- 1--
on May 28, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

RODE AND SMITH
lation seems apparent between spore appendagestatus and other strain properties.
DISCUSSIONThe 35 Clostridium strains, appraised by con-
ventional taxonomic methods, exhibited markedhomogeneity; variable properties were few anddid not serve to exclude any of the strains fromthe C. bifermentans species. This homogeneityextended to the gross spore features of the 35strains: uniformity of spore size, spore shape,exosporium status, and location of spores withinsporangia. These features are consistent with cur-rent characterization of the species (Bergey'sManual, 7th ed.).On the ultrastructural level, marked spore dif-
ferences were encountered among the strains.These differences were limited, however, to thespore appendage aspect; 21 strains lacked appen-dages, whereas 14 strains possessed appendages(Table 2), and these were distributed among fourultrastructurally distinct appendage types (Table2, Fig. 1-5).Two extreme approaches to the taxonomic
problem posed by the spore appendage variabilitycome to mind. The first of these is to ignorespore appendage status in the taxonomy of C.bifermentans and possibly of other clostridia. Asecond approach, used by the investigators atMoscow State University (6), is to create newClostridium species with spore appendage statusas the basis for speciation. This approach makesspore fine structure paramount in speciation, ig-nores all other natural areas of relatedness, andrisks the possibility that very similar or identicalspore appendage types may be found in severalotherwise distinct species.A more workable approach at this time is to
accommodate this new knowledge of spore finestructure within the framework of existing taxo-nomic schemes. This finds support in our observa-tion that variation in cultural characteristics doesnot correlate with spore appendage morphology.Although spore appendage morphology is highlyreliable as a strain characteristic, we see nothingto be gained by an attempt to subdivide a speciesas generally accepted and as uniform, on thewhole, as C. bifermentans.
Inherent in this approach is a designation ofthe several spore appendage types as "varieties"of C. bifermentans. This can be done by the ap-plication of appropriate nomenclature. For the
present, we prefer to retain the simple descriptiveterminology we have used in this report (Table 2,Fig. 1-5) and in earlier reports (8 and 14).
Since the conclusion of our experiments, anadditional spore appendage type of C. bifermen-tans has been described by Samsonoff, Hashi-moto, and Conti (12). Although additional diver-sity is not unanticipated, the basic problem oftaxonomy remains the same.
ACKNOWLEDGMENTS
We are grateful for the technical assistance of Robin N. Huettel andPeggy K. Johnson. We thank Leodocia Pope for advice, helpful discus-sions, and critical reading of the manuscript and Cecil S. Cummins fordetermining cell wall composition.
This investigation was supported by Public Health Service grants Al-07582 from the National Institute of Allergy and Infectious Diseases andGM 14604 from the National Institute of General Medical Sciences, andby research grant GB-17677 from the National Science Foundation.
LITERATURE CITED
1. Franklin, J. G., and D. E. Bradley. 1957. A further study of the sporesof the genus Bacillus in an electron microscope using carbon repli-cas, and some preliminary observations on Clostridiurn welchii. J.Appl. Bacteriol. 20:467-472.
2. Hodgkiss, W., Z. J. Ordal, and D. C. Cann. 1967. The morphologyand ultrastructure of the spore and exosporium of some Clostridiumspecies. J. Gen. Microbiol. 47:213-225.
3. Holt, S. C., and E. R. Leadbetter. 1969. Comparative ultrastructureof selected aerobic spore-forming bacteria: a freeze-etching study.Bacteriol. Rev. 33:346-378.
4. Kellenberger, E., A. R. Ryter, and J. C. Sechaud. 1958. Electron mi-croscope study of DNA-containing plasms. II. Vegetatative andmature phage DNA as compared with normal bacterial nucleoids indifferent physiological states. J. Biophys. Biochem. Cytol. 4:671678.
5. Krasil'nikov, N. A., V. 1. Duda, and A. A. Sokolov. 1963. Outwardgrowth on spores of anaerobic bacteria of the genus Clostridium.Proc. Acad. Sci. USSR Microbiol. Sect. 152:735-736.
6. Krasil'nikov, N. A., V. 1. Duda, and A. A. Sokolov. 1964. Protrusionson the surface of spores of anaerobic bacteria of the genus Closiri-dium. Microbiology 33:454-458.
7. Moore, W. E. C., Elizabeth P. Cato, and Lillian V. Holdeman. 1966.Fermentation patterns of some Clostridium species. Int. J. Syst.Bacteriol. 16:384-415.
8. Pope, L., D. P. Yolton, and L. J. Rode. 1967. Appendages of Clostri-dium bifermentans spores. J. Bacteriol. 94:1206-1215.
9. Reynols, E. S. 1963. The use of lead citrate at high pH as an elec-tron-opaque stain in electron microscopy. J. Cell Biol. 17:208 212.
10. Rode, L. J., M. A. Crawford, and M. G. Williams. 1967. Clostridiumspores with ribbon-like appendages. J. Bacteriol. 95:1160 1173.
11. Sabatini, D. D., F. Miller, and R. J. Barnett. 1964. Aldehvde tixationfor morphological and enzyme histochemical studies with theelectron microscope. J. Histochem. Cytochem. 12:57 71.
12. Samsonoff, W. A., T. Hashimoto, and S. F. Conti. 1970. Ultrastruc-tural changes associated with germination and outgrowth of anappendage-bearing clostridial spore. J. Bacteriol. 101:1()38 1(045.
13. Smith, L. DS., and Lillian V. Holdeman. 1968. The pathogenic an-aerobic bacteria. C. C. Thomas Co., Fort Lauderdale.
14. Yolton, D. P., L. Pope, M. G. Williams, and L. J. Rode. 1968. Fur-ther electron microscope characterization of spore appendages ofClostridium bifermentans. J. Bacteriol. 95:231-238.
354 J. BAICTERIOI..
on May 28, 2018 by guest
http://jb.asm.org/
Dow
nloaded from





























