Embed Size (px)
Citation preview

ANRV311-BB36-01 ARI 1 April 2007 9:12
Synthetic Gene Circuits:Design with DirectedEvolutionEric L. Haseltine and Frances H. ArnoldDivision of Chemistry and Chemical Engineering 210-41, California Instituteof Technology, Pasadena, California 91125; email: [email protected]
Annu. Rev. Biophys. Biomol. Struct. 2007. 36:1–19
First published online as a Review in Advance onDecember 13, 2006
The Annual Review of Biophysics and BiomolecularStructure is online at biophys.annualreviews.org
This article’s doi:10.1146/annurev.biophys.36.040306.132600
Copyright c© 2007 by Annual Reviews.All rights reserved
1056-8700/07/0609-0001$20.00
Key Words
mathematical modeling, gene regulation, synthetic biology
AbstractSynthetic circuits offer great promise for generating insights intonature’s underlying design principles or forward engineering novelbiotechnology applications. However, construction of these circuitsis not straightforward. Synthetic circuits generally consist of com-ponents optimized to function in their natural context, not in thecontext of the synthetic circuit. Combining mathematical modelingwith directed evolution offers one promising means for addressingthis problem. Modeling identifies mutational targets and limits theevolutionary search space for directed evolution, which alters circuitperformance without the need for detailed biophysical information.This review examines strategies for integrating modeling and di-rected evolution and discusses the utility and limitations of availablemethods.
1
Ann
u. R
ev. B
ioph
ys. B
iom
ol. S
truc
t. 20
07.3
6:1-
19. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
05/
08/0
7. F
or p
erso
nal u
se o
nly.

ANRV311-BB36-01 ARI 1 April 2007 9:12
Contents
INTRODUCTION. . . . . . . . . . . . . . . . . 2PROPOSED STRATEGIES FOR
MAKING FUNCTIONALSYNTHETIC CIRCUITS . . . . . . . 3“Plug and Play” Strategy . . . . . . . . . . 3“Design, Then Mutate” Strategy . . 4Combining the “Plug and Play”
and “Design, Then Mutate”Strategies . . . . . . . . . . . . . . . . . . . . . 5
USING DIRECTED EVOLUTIONTO PERTURB CIRCUITS . . . . . . 6Altering the Performance of a
Circuit . . . . . . . . . . . . . . . . . . . . . . . . 8Engineering Novel Regulation . . . . 10
THE ROLE OF MATHEMATICALMODELING . . . . . . . . . . . . . . . . . . . . 11Overview of Deterministic
Methods for IdentifyingMutational Targets . . . . . . . . . . . . 12
Overview of Stochastic Methodsfor Identifying MutationalTargets . . . . . . . . . . . . . . . . . . . . . . . 14
CONCLUSIONS AND FUTUREDIRECTIONS . . . . . . . . . . . . . . . . . . 15
INTRODUCTION
The synthetic gene circuits discipline can bedescribed succinctly as novel regulation ofpre-existing or engineered cellular functions.Consequently, synthetic gene circuits can beviewed as a subset of synthetic biology, the for-ward engineering of biological systems. Theemergence of synthetic gene circuits as an en-gineering goal has re-emphasized the impor-tance of accounting for the continuous natureof gene regulation. In this manner, it com-plements views prevalent, for example, in en-gineering of metabolic pathways where generegulation is either ignored or treated as all-or-none in an effort to construct genome-wide models that assimilate large numbers ofmeasurements (53). Synthetic gene circuitshave also focused attention on the fact thatregulation can be used to elicit specific re-
sponses to specific environmental stimuli (i.e.,pH, temperature, exogenous signals, or sig-nals from other cells). In this regard, they cancoordinate specific functions to internal or ex-ternal cues and execute precise instructions ina temporal or spatial fashion.
Synthetic gene circuits offer the promiseof generating insights into nature’s underly-ing design principles (38, 51), reconstructingand better understanding naturally occurringfunctions (56), and forward engineering novelbiotechnology applications (45). Three exem-plary contributions respectively demonstratethese promises:
� Pedraza & van Oudenaarden (49) inves-tigated how noise propagates through asynthetic cascade of transcriptional re-pressors on the single-cell level. Theydiscovered three sources for noise ingene expression: its own intrinsic fluc-tuations, transmitted noise from up-stream genes, and global noise affectingall genes.
� Suel et al. (59) elucidated an ingeniousnoise-based mechanism that Bacillussubtilis uses to spontaneously and tem-porarily induce competence. They val-idated their model of this novel mecha-nism by constructing a synthetic circuitthat permitted induction to, but not exitfrom, the competent state.
� Farmer & Liao (27) used syntheticcircuits to couple expression of rate-controlling enzymes in lycopene syn-thesis to the presence of a metabolicprecursor of lycopene. This regulationincreased both lycopene production andthe overall growth rate, emphasizing theimportance of dynamic control in opti-mizing gene expression.
The dream of gene circuit designers isthat well-characterized components can beeasily assembled to achieve particular goals.This dream reflects the field’s strong engi-neering influence. In traditional engineeringdisciplines, the concepts of standardization,decoupling, and abstraction have streamlined
2 Haseltine · Arnold
Ann
u. R
ev. B
ioph
ys. B
iom
ol. S
truc
t. 20
07.3
6:1-
19. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
05/
08/0
7. F
or p
erso
nal u
se o
nly.

ANRV311-BB36-01 ARI 1 April 2007 9:12
the process that transforms ideas into physicalrealities (4, 25). Standardization ensures thatindividual working components can be usedtogether or exchanged as needed. Decouplingrepresents a “divide and conquer” strategy inwhich complex problems are dissected intosimpler subsystems. Abstraction pares downinformation so that attention is focused solelyon those details relevant to the problem athand. Applying these concepts should enablesynthetic gene circuits to be conceived, de-signed, and implemented in a fashion simi-lar to how an electrical engineer might de-sign a computer or how a chemical engineermight construct a process flow sheet and ul-timately a chemical plant. As a result, circuitsconstructed from standardized synthetic com-ponents are conceptually superior to naturallyoccurring circuits encoding the same func-tions because the latter are context dependent;that is, nature has optimized them in the con-text of both their natural function and cellu-lar environment. In addition, synthetic genecircuits offer the possibility of implementingnew functions and regulation not available tonatural circuits.
The reality, however, is that the functionof synthetic components and their assembliesalso exhibits this context dependence. A genecircuit does not function outside the cellularcontext and may behave differently when thecontext changes. In fact, it can behave differ-ently even in the same nominal context be-cause of intrinsic and extrinsic noise. Muta-tion can routinely and suddenly alter circuitbehavior in unexpected ways. In addition, cir-cuits encounter variability owing to cell death,undefined and changing extracellular envi-ronments, and interactions with the cellularcontext. How these phenomena affect circuitfunction is not well understood and hindersthe characterization of even the simplest com-ponents (4). Also, most tasks for which circuitsare designed require precise, reproducible be-havior for an entire population of cells, neces-sitating coordination at the population level.Simply stated, details matter, and we don’t un-derstand the details.
In light of context dependence, what arethe strategies one can use to take a syntheticgene circuit from an initial design to an effec-tive physical implementation? Once the initialdesign is implemented, relatively few strate-gies exist for altering the behavior of thesecircuits. Directed evolution guided by math-ematical modeling is a powerful method foraltering circuit performance. Here, we reviewthe utility and limitations of directed evolu-tion to alter circuit performance and constructnovel regulatory components. We then con-sider how mathematical modeling and sys-tems theory tools (e.g., sensitivity analysis)can suggest specific manipulations, therebytargeting the directed evolution. Finally, wepresent conclusions and discuss future direc-tions and opportunities for growth in thisfield.
PROPOSED STRATEGIES FORMAKING FUNCTIONALSYNTHETIC CIRCUITS
Approaches for generating a conceptualdesign of a synthetic circuit include a mod-ular approach, as championed by the engi-neering community (4, 25, 65), and evolu-tionary design (29). We define a conceptualdesign as specifying how individual compo-nents (e.g., sensors, regulatory factors, out-puts) are connected to accomplish the desiredcircuit function. The next step is constructingthe functional circuit. Surprisingly, there existrelatively few strategies for this process giventhe ever-increasing number of published syn-thetic circuits, suggesting that genetic circuitconstruction is currently more of an art formthan a well-established engineering discipline.In this section, we review the “plug and play”strategy, the “design, then mutate” strategy,and a hybrid of the two.
“Plug and Play” Strategy
Using the engineering principle of decou-pling, circuits can be divided into simpler sub-problems consisting of different components.
www.annualreviews.org • Synthetic Gene Circuits 3
Ann
u. R
ev. B
ioph
ys. B
iom
ol. S
truc
t. 20
07.3
6:1-
19. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
05/
08/0
7. F
or p
erso
nal u
se o
nly.

ANRV311-BB36-01 ARI 1 April 2007 9:12
One could then envision exchanging thesecomponents from a well-characterized libraryto effectively alter the circuit performance.Consider, for example, the toggle switch de-sign of Gardner et al. (33), a mutually in-hibitory network consisting of two repressors.A functional toggle switch requires propermatching of the repressor characteristics toensure that the region of bistability falls withinthe operating regime so that transient pertur-bations can toggle the switch from one oper-ating point to the other. In an ideal world, onecould simply choose two such repressors froma well-characterized library (for example, theMIT Parts Registry, http://parts.mit.edu/)and construct a functional toggle switch. Mostcircuit constructs to date have been imple-mented using this approach. However, the dif-ficulties of this approach are not always ap-parent from the published literature, whichdoes not include the vast majority of the manyfailures.
Several practical considerations currentlylimit the utility of this approach. First, designconsiderations often restrict the pool fromwhich one can select components. In the tog-gle switch example, one may desire to main-tain the same switching effectors (thermal andchemical) that Gardner et al. (33) chose. Sec-ond, the function of each component is con-text dependent; different components may ex-hibit different interactions and/or sensitivitiesto inputs when taken out of their naturalcontexts. Finally, current libraries of well-characterized components are sparse. For ex-ample, as of the submission of this review,the MIT Parts Registry currently reports onlyaround 40 repressors.
“Design, Then Mutate” Strategy
An alternative to “plug and play” is “design,then mutate.” Here, one a priori selects thecomponents that comprise the designed cir-cuit. Rather than swapping out different com-ponents, directed evolution is used to manip-ulate the behavior of these components, andthe desired phenotype is obtained via screen-
ing or selection. That directed evolution is anappropriate tool for tuning circuits was recog-nized early in the emergence of the syntheticgene circuit discipline (44), and the utility ofthis approach has been demonstrated experi-mentally via random mutagenesis and screen-ing for functional mutants (66). However, im-plementation of this strategy must be donethoughtfully because the evolutionary searchis restricted to the number of mutants thatcan be screened or selected at each generation,usually 103 to 108. Genetic circuits are com-posed of multiple genes. The number of waysto mutate these genes grows rapidly with thesize of the system, and the probability that arandom mutation will confer the desired func-tionality is correspondingly small.
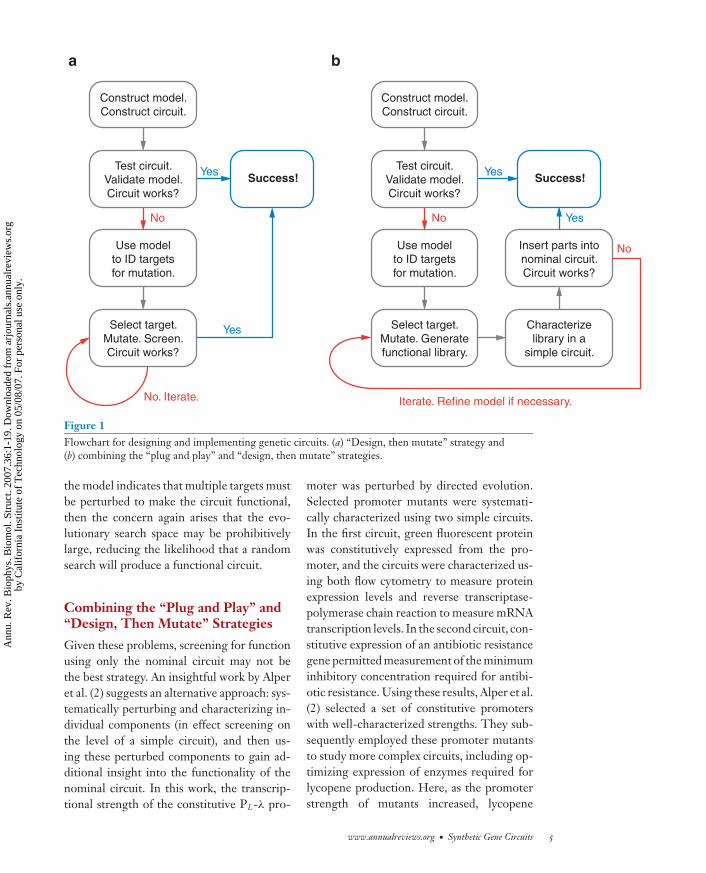
Feng et al. (28) proposed to overcome thisproblem with mathematical modeling of thegene circuit. A flowchart of this strategy ispresented in Figure 1a. Here, global sensi-tivity analysis interrogates a model of the cir-cuit to identify targets for mutation. A mu-tational target is then selected and modifiedusing directed evolution, screening for thedesired overall circuit behavior. Identifying atarget restricts the evolutionary search spaceand increases the likelihood that a functionalcircuit will be screened (provided the righttarget is identified). However, there are sev-eral conceptual problems with this strategy.First, what happens when a single iterationstill yields a nonfunctional circuit? One mustthen decide whether to try the next mutationaltarget or attempt to figure out what wentwrong. One possible explanation is that themathematical model is incorrect and that theidentified mutational target does not alterthe circuit performance in the desired way. Inthis case, trusting the model to provide a sec-ond mutational target is not justified, and theabove procedure has not provided much ad-ditional data with which to refine the model.A second possibility is that the directed evolu-tion has not correctly or sufficiently perturbedthe mutational target. Here, debugging of thedirected evolution may be hindered by thecomplexity of the genetic circuit. Finally, if
4 Haseltine · Arnold
Ann
u. R
ev. B
ioph
ys. B
iom
ol. S
truc
t. 20
07.3
6:1-
19. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
05/
08/0
7. F
or p
erso
nal u
se o
nly.
ANRV311-BB36-01 ARI 1 April 2007 9:12
Yes
Yes
No
No. Iterate. Iterate. Refine model if necessary.
Success!
Construct model.Construct circuit.
Test circuit.Validate model.Circuit works?
Use modelto ID targetsfor mutation.
Select target.Mutate. Screen.Circuit works?
Yes
YesNo
No
Success!
Construct model.Construct circuit.
Test circuit.Validate model.Circuit works?
Use modelto ID targetsfor mutation.
Insert parts intonominal circuit.Circuit works?
Select target.Mutate. Generatefunctional library.
Characterizelibrary in a
simple circuit.
a b
Figure 1Flowchart for designing and implementing genetic circuits. (a) “Design, then mutate” strategy and(b) combining the “plug and play” and “design, then mutate” strategies.
the model indicates that multiple targets mustbe perturbed to make the circuit functional,then the concern again arises that the evo-lutionary search space may be prohibitivelylarge, reducing the likelihood that a randomsearch will produce a functional circuit.
Combining the “Plug and Play” and“Design, Then Mutate” Strategies
Given these problems, screening for functionusing only the nominal circuit may not bethe best strategy. An insightful work by Alperet al. (2) suggests an alternative approach: sys-tematically perturbing and characterizing in-dividual components (in effect screening onthe level of a simple circuit), and then us-ing these perturbed components to gain ad-ditional insight into the functionality of thenominal circuit. In this work, the transcrip-tional strength of the constitutive PL-λ pro-
moter was perturbed by directed evolution.Selected promoter mutants were systemati-cally characterized using two simple circuits.In the first circuit, green fluorescent proteinwas constitutively expressed from the pro-moter, and the circuits were characterized us-ing both flow cytometry to measure proteinexpression levels and reverse transcriptase-polymerase chain reaction to measure mRNAtranscription levels. In the second circuit, con-stitutive expression of an antibiotic resistancegene permitted measurement of the minimuminhibitory concentration required for antibi-otic resistance. Using these results, Alper et al.(2) selected a set of constitutive promoterswith well-characterized strengths. They sub-sequently employed these promoter mutantsto study more complex circuits, including op-timizing expression of enzymes required forlycopene production. Here, as the promoterstrength of mutants increased, lycopene
www.annualreviews.org • Synthetic Gene Circuits 5
Ann
u. R
ev. B
ioph
ys. B
iom
ol. S
truc
t. 20
07.3
6:1-
19. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
05/
08/0
7. F
or p
erso
nal u
se o
nly.

ANRV311-BB36-01 ARI 1 April 2007 9:12
production either increased monotonically orexhibited a maximum, depending on the ge-netic configuration.
The underlying concept is that the nomi-nal circuit should be characterized by system-atically perturbing a single component (cor-responding to a single model parameter). Webriefly outline this approach, which is pre-sented as a flowchart in Figure 1b. (Step 1)Implement and experimentally characterizethe synthetic circuit. (Step 2) Estimate modelparameters by optimally fitting the modelto experimental data. Given these estimates,model analysis will prioritize targets that cangenerate the desired functional behavior withminimal manipulation. (Step 3) Create a li-brary of targeted components with a range ofresponses via directed evolution and/or ratio-nal design. Screening and characterization ofthe library should be performed in a simplecircuit rather than the nominal circuit. (Step4) Incorporate the members of the resulting li-brary into the nominal circuit and check eachindividual circuit for functional behavior. Ifthe circuit does not function as desired, refinethe model and return to Step 2.
The benefits of this approach are fourfold.First, some circuit properties such as oscilla-tions and population-level responses are notsuitable for high-throughput screens or se-lections, so the directed evolution must beperformed in the context of a simple circuit.Second, performing directed evolution in thecontext of a simple circuit ensures that theresulting library covers a wide range of pa-rameter values and facilitates troubleshooting.Third, the generated library consists of inter-changeable components that (a) enable “plugand play” alteration of the circuit by simplyexchanging one component for another andthat (b) can be applied readily to other ge-netic circuits. The underlying assumption isthat mutants of a component are locally per-turbed and have a greater likelihood of retain-ing the same circuit and cellular contexts thando components taken from different contexts.Fourth, examining the circuit behavior overa library of components with widely varying
parameter values facilitates model validationand refinement. Although one could imple-ment the nominal circuit using only the com-ponent predicted to yield functional behavior,such implementation presumes that the modelis accurate. Most likely, there will be some dis-crepancy between the model and the experi-ment. Checking the nominal circuit for func-tional behavior with components spanning arange of parameter values is useful for over-coming such discrepancies.
A potential problem of this strategy is thatcertain circuit properties may not be indepen-dent. Changing one property often affects an-other. For example, changing stability oftenaffects expression level. A possible solutionto avoid this problem is to screen or selectfor multiple properties simultaneously. Re-cent works (18, 50) have demonstrated suc-cessful selection for multiple properties (seebelow). The success of this method dependson the ability to design a screen or selectionthat satisfactorily reports on all properties ofinterest.
Also, the parameter space accessible byDNA mutation is not well understood. Forexample, Mayo et al. (43) demonstrated howthe design of the lac operon transforms auniformly distributed parameter space into anon-uniform phenotype space (i.e., the circuitfunction). That is, the circuit design limits theset of possible phenotypes, and some of thepossible phenotypes are more likely than oth-ers. Interestingly, the wild-type lac operon and16 mutants (one to seven nucleotides differ-ence) fall in a sparsely populated region of thephenotype space. These results suggest thatthe fitness landscape for DNA sequences thatencode functions significantly biases the pa-rameter space.
USING DIRECTED EVOLUTIONTO PERTURB CIRCUITS
Synthetic gene circuits almost always requirefine-tuning for proper function. A brief lookat the experimental methods used to con-struct functional circuits illustrates this point.
6 Haseltine · Arnold
Ann
u. R
ev. B
ioph
ys. B
iom
ol. S
truc
t. 20
07.3
6:1-
19. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
05/
08/0
7. F
or p
erso
nal u
se o
nly.

ANRV311-BB36-01 ARI 1 April 2007 9:12
In some cases, synthetic gene circuits appearto work by assembling natural (pre-existing)components (9, 10, 40, 41, 67); however,the experimental methods outlined in theseworks may not reflect all the steps actuallyperformed to constitute the circuits. Otherprojects have applied various technologies totune circuit performance, including (a) engi-neering of promoters via construction of novelhybrid promoters (e.g., to include repression)(6, 8, 36) and alteration of existing promotersto adjust transcription rates and transcriptionfactor binding (14, 33, 36, 55); (b) alteration ofproteins via use of ssrA tags to increase proteindegradation rates (7, 8, 24, 31), codon opti-mization (7), and recombination (39); (c) ma-nipulation of ribosome-binding sites (RBSs)to alter translation rates (8, 33, 64); (d ) elim-ination or rewiring of native metabolic func-tions (14, 27, 31); and (e) random recombina-tion of regulatory and output functions (35).
However, these methods for perturbingcircuit function are tedious because they makeonly a few changes and then check to see if thecircuit works. If the desired behavior can bescreened in a high-throughput fashion or se-lected, it is significantly more efficient to makemany altered circuits and to screen or selectfor desired behavior, accumulating mutationsover multiple generations, i.e., perform di-rected evolution. High-throughput screeningusually examines between 103 and 106 clonesper generation and (ideally) directly examinesthe desired phenotype. Genetic selections canexamine up to 108 clones but require that thedesired phenotype be linked to cell survival.Because the goal of a synthetic gene circuitby definition is to regulate a cellular function,selection is often a natural choice for iden-tifying desired phenotypes. In this case, theregulatory function of the circuit is coupledto cell survival, for example, by means of anantibiotic resistance gene, to achieve the evo-lutionary pressure required for an effective se-lection.
While screening and selection methodscan potentially test large numbers of mutantsfor functional behavior, these numbers pale in
comparison to the astronomically large num-ber of mutants in a given sequence space. Con-sider, for example, using directed evolution toalter the properties of a relatively small pro-tein consisting of one hundred amino acidresidues. The number of proteins possible forthis one sequence length is 20100 ≈ 10130, aspace that could never be explored exhaus-tively. In addition, the number of sequencesexpected to encode a solution to a specificproblem is an infinitesimal fraction of thisspace. So how does directed evolution workto optimize circuit performance and generatenew functions?
One reason evolution (and directed evolu-tion) works is that the space is highly dimen-sional: There are many ways to mutate evena single small protein. There are many moreways to mutate a circuit. If some of these path-ways enhance the desired performance, theniterative rounds of random mutation of thecircuit and screening for the desired prop-erties may provide steady improvements infunction. The key is that the process is evolu-tionary: Small changes accumulated over mul-tiple generations can lead to a good circuitimplementation. This only works if the ini-tial design is close to (i.e., within a few mu-tations of ) a better design. Because biologicalcomponents and systems are the products ofexactly such an evolutionary search, they areapparently easily tuned in the same fashion.
Important to this search strategy are theideas of protein promiscuity and evolution-ary plasticity (sometimes called evolvability).Proteins that perform a native function oftenalso perform numerous promiscuous func-tions, and small numbers of mutations can in-duce large changes in promiscuous functions(i.e., plasticity) often while having little effecton the native function (1, 62). This theory jus-tifies the standard practice of choosing a pro-tein with a function closely related to the de-sired function as the starting point for directedevolution. In addition, if residues endowingplasticity are known, then this trait can beexploited by targeting mutagenesis to spe-cific residues (e.g., inserting each amino acid
www.annualreviews.org • Synthetic Gene Circuits 7
Ann
u. R
ev. B
ioph
ys. B
iom
ol. S
truc
t. 20
07.3
6:1-
19. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
05/
08/0
7. F
or p
erso
nal u
se o
nly.

ANRV311-BB36-01 ARI 1 April 2007 9:12
into the targeted sites) to perturb the proteinfunction.
Directed evolution is a relatively maturefield. We refer the interested reader to reviewsdiscussing methods for generating moleculardiversity (12) and screening for desired phe-notypes (16, 46). Instead, we focus on the rolethat directed evolution serves in engineeringsynthetic gene circuits, first in tuning the per-formance of synthetic circuits and second inengineering novel regulatory functions. Wealso assess the promise and limitations of di-rected evolution for circuit engineering.
Altering the Performance of a Circuit
Directed evolution is a powerful method foraltering circuit performance; however, it doeshave some limitations. To investigate theselimitations further, we review works that havetuned circuits by employing directed evolu-tion to identify strategies for selecting muta-tional targets.
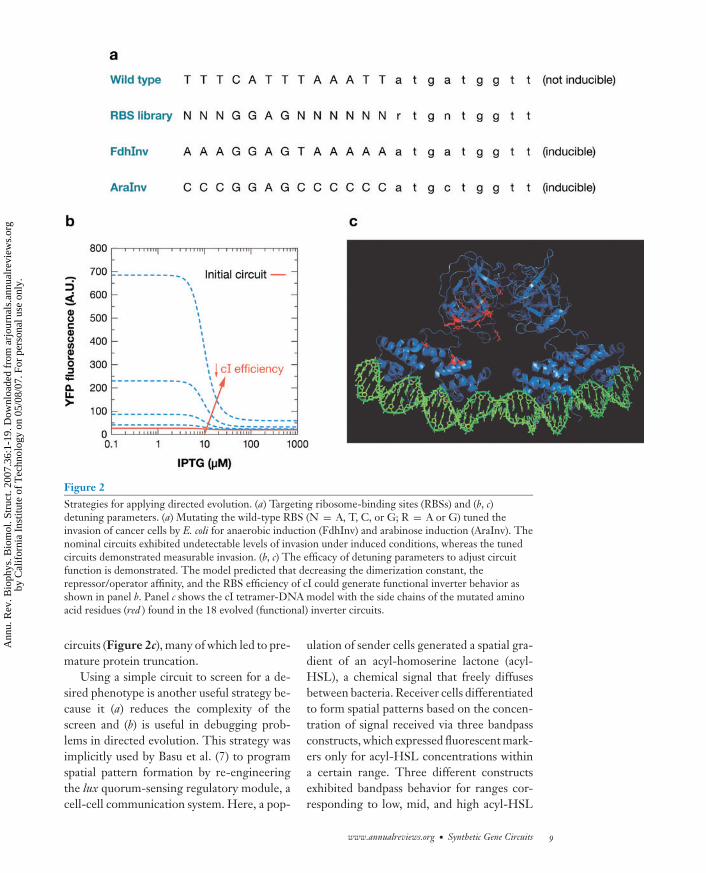
Targeting functionally rich segments ofthe DNA is critical for successfully alteringcircuit function, since the entire circuit se-quence space is too large to explore except inthe most cursory fashion. One such rich seg-ment for circuit function is the RBS, whichdictates protein expression levels. For exam-ple, the codons between positions -7 and -13in the 5′ to AUG initiation of mRNA appearto cause most of the variation in RBS strengthin Escherichia coli (54). The size of the resultinglibrary containing all possible variants is 47 =16,384, a size well within the capability of ei-ther screens or selections to cover completely.The design of a genetic circuit typically dic-tates the function; nonfunctional circuits areoften the result of improperly matched ex-pression levels rather than a flaw in the ba-sic design. For example, Anderson et al. (3)used random mutagenesis of the RBS to engi-neer E. coli to invade cancer cells. The bacteriawere programmed to express the gene invasin,which facilitates adhesion and invasion ofmammalian cells expressing β1-integrins, un-der induced, hypoxic, and density-dependent
conditions. Because the last two environmen-tal conditions indicate that the bacteria havelocalized in the tumor, the circuit thus targetsgene expression only in the tumor. By con-structing appropriate RBS libraries to tuneinvasin expression (see Figure 2a), Ander-son et al. (3) generated circuits that enablebacteria to invade cancer cells under theseconditions.
Targeting the RBS for mutation is an ef-fective strategy for tuning circuit behavior foranother reason: RBS mutations nominally af-fect only the expression level of a protein. Pro-tein transcription factors, on the other hand,tend to encompass numerous functions (forexample, DNA binding and activation or re-pression). Point mutations to such proteinsmay alter these functions in addition to geneexpression.
Another feature of mutagenesis is that per-turbed circuit components are more likely tobe destabilized or detuned than to exhibitimprovements in function. For proteins, thefraction of random mutants retaining wild-type function decreases exponentially withincreasing numbers of amino acid substitu-tions (13, 22, 60). An effective strategy forchoosing mutational targets is to select targetsthat yield functional behavior when detuned.Yokobayashi et al. (66) used this strategy tocorrect a nonfunctional inverter circuit. Inthis circuit, LacI protein was constitutivelyexpressed and repressed expression of cI. cIin turn repressed expression of the output,a fluorescent protein. As a result, the cir-cuit should naturally fluoresce, and exter-nal addition of isopropyl β-d-thiogalactoside,an inducer that interferes with LacI func-tion, should reduce fluorescence. However,the nominal circuit exhibited no fluorescenceeven in the absence of IPTG. An analysis ofa mathematical model describing the inverterindicated that decreasing either the cI repres-sor operator affinity, RBS strength, or dimer-ization could generate functional circuits (seeFigure 2b). Yokobayashi et al. (66) randomlymutagenized the cI gene and its RBS. Numer-ous mutations turned out to yield functional
8 Haseltine · Arnold
Ann
u. R
ev. B
ioph
ys. B
iom
ol. S
truc
t. 20
07.3
6:1-
19. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
05/
08/0
7. F
or p
erso
nal u
se o
nly.
ANRV311-BB36-01 ARI 1 April 2007 9:12
Figure 2Strategies for applying directed evolution. (a) Targeting ribosome-binding sites (RBSs) and (b, c)detuning parameters. (a) Mutating the wild-type RBS (N = A, T, C, or G; R = A or G) tuned theinvasion of cancer cells by E. coli for anaerobic induction (FdhInv) and arabinose induction (AraInv). Thenominal circuits exhibited undetectable levels of invasion under induced conditions, whereas the tunedcircuits demonstrated measurable invasion. (b, c) The efficacy of detuning parameters to adjust circuitfunction is demonstrated. The model predicted that decreasing the dimerization constant, therepressor/operator affinity, and the RBS efficiency of cI could generate functional inverter behavior asshown in panel b. Panel c shows the cI tetramer-DNA model with the side chains of the mutated aminoacid residues (red ) found in the 18 evolved (functional) inverter circuits.
circuits (Figure 2c), many of which led to pre-mature protein truncation.
Using a simple circuit to screen for a de-sired phenotype is another useful strategy be-cause it (a) reduces the complexity of thescreen and (b) is useful in debugging prob-lems in directed evolution. This strategy wasimplicitly used by Basu et al. (7) to programspatial pattern formation by re-engineeringthe lux quorum-sensing regulatory module, acell-cell communication system. Here, a pop-
ulation of sender cells generated a spatial gra-dient of an acyl-homoserine lactone (acyl-HSL), a chemical signal that freely diffusesbetween bacteria. Receiver cells differentiatedto form spatial patterns based on the concen-tration of signal received via three bandpassconstructs, which expressed fluorescent mark-ers only for acyl-HSL concentrations withina certain range. Three different constructsexhibited bandpass behavior for ranges cor-responding to low, mid, and high acyl-HSL
www.annualreviews.org • Synthetic Gene Circuits 9
Ann
u. R
ev. B
ioph
ys. B
iom
ol. S
truc
t. 20
07.3
6:1-
19. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
05/
08/0
7. F
or p
erso
nal u
se o
nly.

ANRV311-BB36-01 ARI 1 April 2007 9:12
concentrations. Wild-type luxR and a hyper-sensitive mutant were used to construct themid- and low-detect bandpasses required forspatial pattern formation. Collins et al. (17)constructed the hypersensitive mutant by ran-domly mutagenizing the acyl-HSL signal-binding domain of luxR and subsequentlyscreening for gene activation in a simple cir-cuit. Screening in the nominal bandpass cir-cuit would have been significantly more diffi-cult because it would have required screeningfor behavior at a wide range of acyl-HSL con-centrations to confirm bandpass behavior.
Engineering Novel Regulation
Another benefit of directed evolution is theability to construct novel regulatory func-tions altogether. For example, recent workhas focused on engineering the specificity oftranscriptional regulators (32). Two effectivestrategies for engineering novel regulation in-clude selecting for orthogonal properties andexploiting modularity.
Engineering orthogonal properties. Apowerful capability of directed evolution isthe ability to select for multiple orthogonalphenotypes. We define orthogonal in thiscontext to mean that the phenotypes arewell separated, e.g., one variant exhibitsfunction A but not function B, while anothermight exhibit function B but not A (seeFigure 3a). Rackham & Chin (50) evolvedorthogonal ribosome-mRNA pairs thatselectively translated mRNAs with specificRBSs not recognized by the native E. coliribosomes. The pairs were generated by firstselecting RBSs not recognized by the nativeribosomes and then selecting for mutant16S ribosomal RNA that recognized thenonnatural RBSs. Collins et al. (18) evolvedorthogonal transcriptional activators byrefocusing the specificity of the wild-typeluxR to a noncognate signal. This specificitywas obtained by first selecting mutants thatresponded to the desired noncognate signaland then selecting from the surviving mutants
those that did not respond to the cognate sig-nal. Other works have re-engineered nuclearhormone receptors (NHRs) to be specificand orthogonal to naturally occurring NHRs[see the recent review by Chockalingam et al.(15)].
Exploiting modularity to engineer novelfunctions. The modularity of regulatoryproteins offers the tantalizing prospect of gen-erating novel input-output relationships bysimply recombining domains from differentregulators (11). Figure 3b presents a simpleexample of such a swap, illustrating how re-combination can generate two novel input-output pairings from two existing pairings.For example, the diverse array of eukaryoticsignaling proteins is highly modular in thatthe protein’s input (signal) and output (cat-alytic function) appear to be physically andfunctionally separable. Bhattacharyya et al.(11) recently reviewed rewiring of eukaryoticregulation via recombination. The zinc-fingerdomain presents another possible scaffold forengineering novel DNA-binding proteins ineukaryotes (19). A single zinc finger typicallyrecognizes only three DNA bases, but link-ing multiple fingers can lead to recognition oflonger sequences. Such modularity theoreti-cally permits generating DNA-binding pro-teins of arbitrary sequence specificities via fu-sion of premade fingers recognizing any oneof the 64 possible DNA triplets. Papworthet al. (47) reviewed recent progress in thisfield by both rational design and selectionstrategies.
Modularity is a feature of certain prokary-otic regulatory proteins as well. Nativemembrane-bound sensors in bacteria are typ-ically modular and consist of a membrane-bound, extracellular sensor and an intracel-lular response regulator. Levskaya et al. (42)exploited such modularity to create a syn-thetic phytochrome by fusing a cyanobac-terial photoreceptor to an E. coli intracel-lular histidine kinase domain. By importingthis novel gene along with genes required toproduce the photoreceptor, they engineered
10 Haseltine · Arnold
Ann
u. R
ev. B
ioph
ys. B
iom
ol. S
truc
t. 20
07.3
6:1-
19. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
05/
08/0
7. F
or p
erso
nal u
se o
nly.

ANRV311-BB36-01 ARI 1 April 2007 9:12
a
b
Mutate
Select for B
Select for not A
A
Notfunctional
A B
Notfunctional
A B
Notfunctional
A BResponse
A
Signal A Signal B Signal A
ResponseB
ResponseB
ResponseA
Recombination
Orthogonalbehavior
Signal B
Figure 3Two effective strategies for engineering novel regulation using directed evolution include (a) selecting fororthogonal properties and (b) exploiting modularity. Selecting for orthogonal properties (a) first involvesgenerating diversity through, for example, random mutagenesis. Thus, a parent starting with function Acould yield mutants that are nonfunctional, retain function A, acquire a new function B, or acquire a newfunction B while retaining function A. To ensure orthogonality of functions, two selections must beperformed: first, selection for function B, and then selection for not retaining function A. Extrapolationof this logic to engineering orthogonality of multiple functions is straightforward. Exploiting themodularity of regulatory proteins (b) permits synthesis of new proteins with altered input-outputbehavior. For example, two regulatory proteins, each consisting of two domains, a signaling receptor, andan output effector, could theoretically permit swapping of input-output behavior.
E. coli to photograph light. The modularityof the LuxR protein, whose signal-specificityand DNA-binding functions are encoded inseparate domains, has been exploited to cre-ate a variant that responds to a new sig-nal by activating at a new promoter (C.A.Tracewell, C.H. Collins & F.H. Arnold,manuscript in preparation). This modular-ity allows the subfunctions to evolve sepa-rately and then recombine to efficiently createnovel, well-characterized, signal-dependenttranscriptional activators with a wide range ofbehaviors that can be used interchangeably inother circuits. Guntas et al. (37) used directedevolution to fuse a maltose-binding proteinto a β-lactamase, effectively building modu-
larity. This novel protein switch required mal-tose to hydrolyze β-lactam.
THE ROLE OF MATHEMATICALMODELING
In the previous section, we have demonstratedthe utility and limitations of using directedevolution to perturb gene circuits. In partic-ular, the three primary limitations are that(a) the evolutionary search space for a geneticcircuit composed of many genes is generallytoo large to explore efficiently; (b) detuningparameters (reducing function) is much easierthan improving function; and (c) although se-lecting for independent properties is possible,
www.annualreviews.org • Synthetic Gene Circuits 11
Ann
u. R
ev. B
ioph
ys. B
iom
ol. S
truc
t. 20
07.3
6:1-
19. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
05/
08/0
7. F
or p
erso
nal u
se o
nly.

ANRV311-BB36-01 ARI 1 April 2007 9:12
it usually requires setting up multiple roundsof screening or selection. Using mathemati-cal models to suggest mutational targets canhelp overcome each of these limitations. Iden-tifying mutational targets in this way makesthe evolutionary search reasonable by exploit-ing the modularity of the circuit design todivide the original problem into simpler sub-problems, thus overcoming the first limita-tion. Other problems of comparable size (e.g.,evolution of a multiprotein assembly) cannotbe subdivided in this way unless they are com-parably modular and the interactions betweenmodules can be described with some accu-racy. Biasing selection of the mutational tar-gets suggested by the model can account forthe second and third limitations (i.e., choosingtargets that require detuning or independenttargets such as RBSs).
We now focus more in depth on how to usemathematical models to identify mutationaltargets for directed evolution. Mathematicalmodels are analogous to a road map in thatthey contain a concise abstraction of the locallandscape. As with planning any excursion, itis critical to know both the current locationand the location of the desired destination.Continuing in this analogy, parameter esti-mation can determine the current location,bifurcation analysis is useful in identifying thedesired destination (robust, stable, or oscilla-tory states, for example), and sensitivity anal-ysis and optimization serve as a local com-pass (pointing out which directions are easiestto travel) and as a trip optimizer (the short-est distance to the final destination given tripconstraints), respectively. Two types of math-ematical models have proven particularly use-ful in describing synthetic gene circuits: deter-ministic and stochastic reaction models.
Overview of Deterministic Methodsfor Identifying Mutational Targets
In a deterministic setting, mass balances areused to determine evolution equations for thequantities of interest. For this type of model,the combination of parameters and initial con-
ditions uniquely determines the system evolu-tion. The most commonly used assumptionsare that the system is well mixed and that it issegregated by time, leading to models gener-ally of the form
d (NV )dt
= F(N, t;θ), 1.
in which N is a vector of the reacting compo-nent concentrations, V is the system volume,θ is a vector of the model parameters, and F
(N, t; θ) is a vector containing the productionrates and accumulation rates due to transportof components into and out of the system.
Models governed by Equation 1 are sim-ulated dynamically or analyzed for steady-state behavior. Dynamic simulation (or timeintegration) is generally straightforward, andmany numerical software packages such asMATLAB and Octave contain tools for solv-ing models governed by Equation 1. Such sim-ulation has been used, for example, to exam-ine how cells might spatially control pulsesof gene expression (8) or regulate populationdensity (67). The steady states of Equation1 are determined by solving for the reactingcomponents when the time derivatives are setto zero, i.e.,
0 = F(N, t;θ). 2.
Continuation methods offer a powerful meansfor mapping steady-state circuit performanceas a function of key parameters. Bifurcationanalysis complements continuation methodsby examining how changes in the parame-ter values affect the stability of the steadystates; Strogatz (58) provides a nice introduc-tory overview of this topic. Packages such asAUTO (21) and CL MATCONT (20) com-bine both of these functionalities. Excellentexamples of using bifurcation analysis to an-alyze synthetic gene circuits include deter-mination of regions of bistability for con-structing a genetic toggle switch (33) andcalculation of regions generating sustainedoscillations (24, 31).
12 Haseltine · Arnold
Ann
u. R
ev. B
ioph
ys. B
iom
ol. S
truc
t. 20
07.3
6:1-
19. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
05/
08/0
7. F
or p
erso
nal u
se o
nly.

ANRV311-BB36-01 ARI 1 April 2007 9:12
Parameter estimation. Once a model de-scribing the system of interest has been con-structed, the next step is to use parameterestimation to identify the exact location in pa-rameter space. This problem can be formu-lated as an optimization problem,
minθ
�(θ), 3.
subject to model predictions of the experi-mental measurements. The goal is to mini-mize the scalar objective function �(θ) by ad-justing the parameters θ, thereby reconcilingthe experimental data to the model predic-tions. In the event that the optimal parametersdo not satisfactorily predict all the experimen-tal data, the model should be refined and newparameters should be estimated.
Often, the function �(θ) correspondsto a weighted sum-of-squares error, sincethis formulation arises from a probabilisticsetting given appropriate assumptions (57).Of particular importance is the analysis ofthe Hessian of the objective function at theoptimal solution, i.e.,
H = ∂2�
∂θ∂θT . 4.
The analysis of the Hessian reveals whichcombination of parameters can and cannot beestimated from the experimental data. In thelatter case, the number of model parameterscan often be refined either by fixing parametervalues based on previously reported literatureestimates or by reducing the model by using,for example, equilibrium or quasi-steady-stateapproximations. Rawlings & Ekerdt (52) pro-vide a nice introduction to these methodsalong with several instructive examples.
Sensitivity analysis. Having pinpointed thelocation of the current circuit behavior inthe model parameter space, sensitivity anal-ysis offers a first approximation for determin-ing which parameters are critical for alteringcircuit behavior (assuming that the desired be-havior can be written quantitatively). Both lo-cal and global sensitivity analyses are useful inthis regard. Local sensitivity analysis seeks to
determine how a small perturbation to a givenparameter affects the desired circuit response.For example, we might ask how the period ofoscillation, τ , changes with respect to a givenparameter, k. Then the local sensitivity s isdefined as
s = ∂τ
∂k. 5.
More generically, the period τ could be writ-ten as a function of the state N, i.e., g(N ),yielding the general definition:
s = ∂g(N )∂θ
. 6.
Subsequently, one can prioritize which pa-rameter has the greatest effect on the de-sired response by examining the magnitudeof the values in the sensitivity vector s (thegreater the magnitude, the greater the effect).As noted above, prioritizing mutational tar-gets should take into account the directional-ity of the sensitivity.
Alternatively, global sensitivity analysis canbe used to identify mutational targets. Thebasic idea is to determine the effect of pa-rameters on the desired output for rangesof parameter values to elucidate generic orglobal parameter functions. Feng et al. (28)used the random sampling–high dimensionalmodel representation algorithm to addressthis task. Here, parameter values are ran-domly sampled from biologically relevantranges, and the sensitivities are calculated byaveraging the model responses over differ-ent parameter sets. When calculating globalsensitivities, care must be taken when themodel exhibits multiple steady states, peri-odic behavior without steady states, or chaoticbehavior.
The choice of which sensitivity to usemay depend on the complexity of the model.Local sensitivity appears better suited for in-terrogating simple models in which all or mostparameters can be uniquely determined fromexperimental data. In this case, the local sen-sitivity should accurately reflect the circuitbehavior in the experiment. For more com-plex models in which many combinations of
www.annualreviews.org • Synthetic Gene Circuits 13
Ann
u. R
ev. B
ioph
ys. B
iom
ol. S
truc
t. 20
07.3
6:1-
19. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
05/
08/0
7. F
or p
erso
nal u
se o
nly.
ANRV311-BB36-01 ARI 1 April 2007 9:12
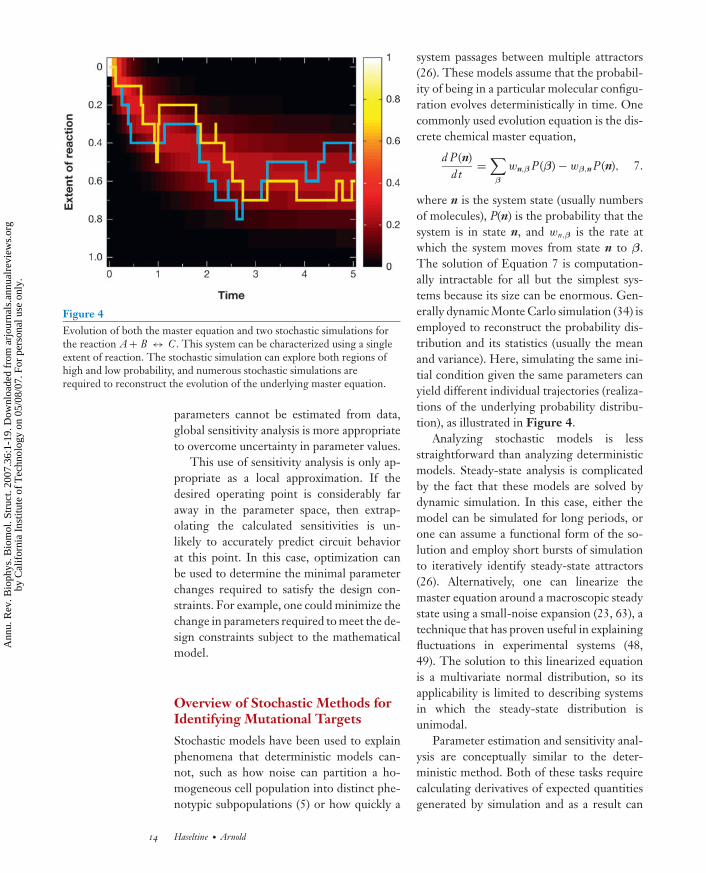
Figure 4Evolution of both the master equation and two stochastic simulations forthe reaction A + B ↔ C . This system can be characterized using a singleextent of reaction. The stochastic simulation can explore both regions ofhigh and low probability, and numerous stochastic simulations arerequired to reconstruct the evolution of the underlying master equation.
parameters cannot be estimated from data,global sensitivity analysis is more appropriateto overcome uncertainty in parameter values.
This use of sensitivity analysis is only ap-propriate as a local approximation. If thedesired operating point is considerably faraway in the parameter space, then extrap-olating the calculated sensitivities is un-likely to accurately predict circuit behaviorat this point. In this case, optimization canbe used to determine the minimal parameterchanges required to satisfy the design con-straints. For example, one could minimize thechange in parameters required to meet the de-sign constraints subject to the mathematicalmodel.
Overview of Stochastic Methods forIdentifying Mutational Targets
Stochastic models have been used to explainphenomena that deterministic models can-not, such as how noise can partition a ho-mogeneous cell population into distinct phe-notypic subpopulations (5) or how quickly a
system passages between multiple attractors(26). These models assume that the probabil-ity of being in a particular molecular configu-ration evolves deterministically in time. Onecommonly used evolution equation is the dis-crete chemical master equation,
d P (n)dt
=∑
β
wn,β P (β) − wβ,n P (n), 7.
where n is the system state (usually numbersof molecules), P(n) is the probability that thesystem is in state n, and wn,β is the rate atwhich the system moves from state n to β.The solution of Equation 7 is computation-ally intractable for all but the simplest sys-tems because its size can be enormous. Gen-erally dynamic Monte Carlo simulation (34) isemployed to reconstruct the probability dis-tribution and its statistics (usually the meanand variance). Here, simulating the same ini-tial condition given the same parameters canyield different individual trajectories (realiza-tions of the underlying probability distribu-tion), as illustrated in Figure 4.
Analyzing stochastic models is lessstraightforward than analyzing deterministicmodels. Steady-state analysis is complicatedby the fact that these models are solved bydynamic simulation. In this case, either themodel can be simulated for long periods, orone can assume a functional form of the so-lution and employ short bursts of simulationto iteratively identify steady-state attractors(26). Alternatively, one can linearize themaster equation around a macroscopic steadystate using a small-noise expansion (23, 63), atechnique that has proven useful in explainingfluctuations in experimental systems (48,49). The solution to this linearized equationis a multivariate normal distribution, so itsapplicability is limited to describing systemsin which the steady-state distribution isunimodal.
Parameter estimation and sensitivity anal-ysis are conceptually similar to the deter-ministic method. Both of these tasks requirecalculating derivatives of expected quantitiesgenerated by simulation and as a result can
14 Haseltine · Arnold
Ann
u. R
ev. B
ioph
ys. B
iom
ol. S
truc
t. 20
07.3
6:1-
19. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
05/
08/0
7. F
or p
erso
nal u
se o
nly.

ANRV311-BB36-01 ARI 1 April 2007 9:12
be computationally demanding to performif the simulation is computationally expen-sive to evaluate. A key concept is to evalu-ate these derivatives using the same trajec-tory (practically, the same string of randomnumbers) to significantly reduce the varianceof the sensitivity estimates (30). Supplemen-tal Figure 1 (follow the Supplemental Mate-rial link from the Annual Reviews home pageat http://www.annualreviews.org) demon-strates the difference between evaluat-ing derivatives using the same trajectoryand evaluating derivatives using randomtrajectories.
CONCLUSIONS AND FUTUREDIRECTIONS
The dream is that well-characterized com-ponents can be easily assembled to generatenovel genetic regulatory circuits. The real-ity is that this is hard to accomplish. Thecomponents and their assemblies are contextdependent: Synthetic circuits do not func-tion outside the cellular context and may be-have differently when the context changes.Directed evolution presents a powerful toolfor overcoming this problem, but the evolu-tionary search space can be large. Mathemat-ical modeling can reduce this space to a rea-sonable size by identifying mutational targets.The modular nature of synthetic gene cir-cuits and the mathematical model permit thisreduction.
Screening of the circuit for the desiredfunction is a necessity. However, screeningindividual components for altered functionin simple circuits is a subtly different andpowerful complementary approach. By eval-uating circuit behavior for components witha well-characterized range of functionality,the nominal circuit can be systematically per-turbed, permitting model validation and re-finement. In addition, the library of compo-nents can be used interchangeably with othercircuits. A key requirement of this method-ology is the ability to engineer componentswith independent properties, that is, to per-
turb only a single parameter or combinationof parameters while holding the remaining pa-rameters constant. It is unclear whether al-teration of biophysical components such asproteins and DNA will actually permit suchperturbations. However, recent works havedemonstrated the feasibility of selecting formultiple properties, suggesting that such per-turbations may indeed be possible at the priceof designing more complicated screens orselections.
The reviewed works employing directedevolution to alter circuit performance suggesttwo additional strategies for selecting muta-tional targets. First, targets that can be de-tuned to yield functional circuit behavior aremore likely to be generated and identified dur-ing directed evolution than are targets that re-quire increases in efficiency. Although manymutations can detune function, mutations thatimprove a function are usually rare and there-fore require more screening effort. Second,mutagenizing RBSs provides a generic and ef-ficient strategy for manipulating some aspectsof circuit function.
Directed evolution also has the potentialfor constructing novel regulatory functionsaltogether. Here, multiple rounds of selec-tion can be used to construct circuit compo-nents that meet multiple, possibly orthogonal,design considerations. Also, exploiting themodularity of proteins shows promise in de-signing efficiently regulated enzymes, con-structing new signaling pathways, and en-gineering novel regulation in eukaryoticcells.
Identifying mutational targets from math-ematical models involves first using param-eter estimation to pinpoint the location ofthe implemented circuit in the parameterspace and then sensitivity analysis or opti-mization to suggest mutational targets. Thesetasks are straightforward for deterministicmodels. Because stochastic models are gen-erally solved by simulation, these tasks canbe performed efficiently by evaluating re-quired derivatives using the same simulationtrajectories.
www.annualreviews.org • Synthetic Gene Circuits 15
Ann
u. R
ev. B
ioph
ys. B
iom
ol. S
truc
t. 20
07.3
6:1-
19. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
05/
08/0
7. F
or p
erso
nal u
se o
nly.

ANRV311-BB36-01 ARI 1 April 2007 9:12
SUMMARY POINTS
1. Synthetic circuit components and their assemblies are context dependent: They donot function outside the cellular context and may behave differently when the contextchanges.
2. Context dependence can be overcome by combining directed evolution with math-ematical modeling. Using mathematical modeling to identify mutational targets re-duces the evolutionary search space for directed evolution, which manipulates circuitperformance without requiring mechanistic or detailed structural information.
3. Screening individual components for altered function in simple circuits allows system-atic perturbation of the nominal circuit, permitting model validation and refinementwhile generating a library of components that can be used interchangeably with othercircuits.
4. RBSs and targets that detune function are two effective mutational targets that altercircuit performance.
5. Selecting for orthogonal properties and exploiting the modularity of regulatory pro-teins facilitate construction of novel regulatory functions by directed evolution.
FUTURE ISSUES
1. Is systematic perturbation of circuit components a useful and generic strategy fortuning circuit function? Also, do individual components tuned in one context behavesimilarly once integrated into the context of the nominal circuit?
2. The parameter space accessible by random mutation of the DNA is not well un-derstood. As a result, it is unclear what circuit functions are accessible by directedevolution.
3. Directed evolution can be used to construct libraries of well-characterized compo-nents spanning a wide range of parameter values. Will these libraries enable “plugand play” alteration of future circuits?
4. What are the scalability issues involved in fine-tuning synthetic circuits? Can a fewsmall changes tune circuit behavior when the number of components becomes large?
ACKNOWLEDGMENTS
ELH is supported by the Caltech Center for Biological Circuit Design. FHA acknowledges sup-port from the NSF and NIH. We thank R. Weiss and L. You for comments on the manuscript.
LITERATURE CITED
1. Aharoni A, Gaidukov L, Khersonsky O, McQ Gould S, Roodveldt C, Tawfik DS. 2005.The ‘evolvability’ of promiscuous protein functions. Nat. Genet. 37:73–76
2. Alper H, Fischer C, Nevoigt E, Stephanopoulos G. 2005. Tuning genetic control throughpromoter engineering. Proc. Natl. Acad. Sci. USA 102:12678–83
16 Haseltine · Arnold
Ann
u. R
ev. B
ioph
ys. B
iom
ol. S
truc
t. 20
07.3
6:1-
19. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
05/
08/0
7. F
or p
erso
nal u
se o
nly.

ANRV311-BB36-01 ARI 1 April 2007 9:12
3. Anderson JC, Clarke EJ, Arkin AP, Voigt CA. 2006. Environmentally controlled invasionof cancer cells by engineered bacteria. J. Mol. Biol. 355:619–27
4. Andrianantoandro E, Basu S, Karig DK, Weiss R. 2006. Synthetic biology: new engineeringrules for an emerging discipline. Mol. Syst. Biol. 2:2006.0028
5. Arkin A, Ross J, McAdams H. 1998. Stochastic kinetic analysis of developmental pathwaybifurcation in phage lambda-infected Escherichia coli cells. Genetics 149:1633–48
6. Atkinson M, Savageau MA, Myers JT, Ninfa AJ. 2003. Development of genetic circuitryexhibiting toggle switch or oscillatory behavior in Escherichia coli. Cell 113:597–607
7. Basu S, Gerchman Y, Collins CH, Arnold FH, Weiss R. 2005. A synthetic multicellularsystem for programmed pattern formation. Nature 434:1130–34
8. Basu S, Mehreja R, Thiberge S, Chen M-T, Weiss R. 2004. Spatiotemporal control ofgene expression with pulse-generating networks. Proc. Natl. Acad. Sci. USA 101:6355–60
9. Becskei A, Seraphin B, Serrano L. 2001. Positive feedback in eukaryotic gene networks:cell differentiation by graded to binary response conversion. EMBO J. 20:2528–35
10. Becskei A, Serrano L. 2000. Engineering stability in gene networks by autoregulation.Nature 405:590–93
11. Bhattacharyya RP, Remenyi A, Yeh BJ, Lim WA. 2006. Domains, motifs, and scaffolds:the role of modular interactions in the evolution and wiring of cell signaling circuits. Annu.Rev. Biochem. 75:655–80
12. Bloom JD, Meyer MM, Meinhold P, Otey CR, MacMillan D, Arnold FH. 2005. Evolvingstrategies for enzyme engineering. Curr. Opin. Struct. Biol. 15:447–52
13. Bloom JD, Silberg JJ, Wilke CO, Drummond DA, Adami C, Arnold FH. 2005. Thermo-dynamic prediction of protein neutrality. Proc. Natl. Acad. Sci. USA 102:606–11
14. Bulter T, Lee S, Wong WW, Fung E, Connor MR, Liao JC. 2004. Design of artificialcell-cell communication using gene and metabolic networks. Proc. Natl. Acad. Sci. USA101:2299–304
15. Chockalingam K, Zhao H. 2005. Creating new specific ligand-receptor pairs for transgeneregulation. Trends Biotechnol. 23:333–35
16. Cohen N, Abramov S, Dror Y, Freeman A. 2001. In vitro enzyme evolution: the screeningchallenge of isolating the one in a million. Trends Biotechnol. 19:507–10
17. Collins CH, Arnold FH, Leadbetter JR. 2005. Directed evolution of Vibrio fischeri LuxRfor increased sensitivity to a broad spectrum of acyl-homoserine lactones. Mol. Microbiol.55:712–23
18. Collins CH, Leadbetter JR, Arnold FH. 2006. Dual selection enhances the signaling speci-ficity of a variant of the quorum-sensing transcriptional activator LuxR. Nat. Biotechnol.24:708–12
19. Collins CH, Yokobayashi Y, Umeno D, Arnold FH. 2003. Engineering proteins that bind,move, make and break DNA. Curr. Opin. Biotechnol. 14:371–78
20. Dhooge A, Govaerts W, Kuznetsov YA, Mestrom W, Riet AM. 2004. CL MATCONT: acontinuation toolbox in Matlab. http://users.ugent.be/∼ajdhooge/research.html#new
21. Doedel EJ, Paffenroth RC, Champneys AR, Fairgrieve TF, Kuznetsov YA, et al. 2002.AUTO 2000: continuation and bifurcation software for ordinary differential equations(with HomCont). Tech. Rep., Concordia Univ., Montreal, Can.
22. Drummond DA, Iverson BL, Georgiou G, Arnold FH. 2005. Why high-error-rate ran-dom mutagenesis libraries are enriched in functional and improved proteins. J. Mol. Biol.350:806–16
23. Elf J, Ehrenberg M. 2003. Fast evaluation of fluctuations in biochemical networks withthe linear noise approximation. Genome Res. 13:2475–84
www.annualreviews.org • Synthetic Gene Circuits 17
Ann
u. R
ev. B
ioph
ys. B
iom
ol. S
truc
t. 20
07.3
6:1-
19. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
05/
08/0
7. F
or p
erso
nal u
se o
nly.

ANRV311-BB36-01 ARI 1 April 2007 9:12
24. Elowitz MB, Leibler S. 2000. A synthetic oscillatory network of transcriptional regulators.Nature 403:335–38
25. Endy D. 2005. Foundations for engineering biology. Nature 438:449–5326. Erban R, Kevrekidis IG, Adalsteinsson D, Elston TC. 2006. Gene regulatory net-
works: a coarse-grained, equation-free approach to multiscale computation. J. Chem. Phys.124:084106
27. Farmer WR, Liao JC. 2000. Improving lycopene production in Escherichia coli by engi-neering metabolic control. Nat. Biotechnol. 5:533–37
28. Feng X, Hooshangi S, Chen D, Li G, Weiss R, Rabitz H. 2004. Optimizing genetic circuitsby global sensitivity analysis. Biophys. J. 87:2195–202
29. Francois P, Hakim V. 2004. Design of genetic networks with specified functions by evo-lution in silico. Proc. Natl. Acad. Sci. USA 101:580–85
30. Fu M, Hu JQ. 1997. Conditional Monte Carlo: Gradient Estimation and Optimization Appli-cations. Boston: Kluwer
31. Fung E, Wong WW, Suen JK, Bulter T, Lee S, Liao JC. 2005. A synthetic gene-metabolicoscillator. Nature 435:118–22
32. Galvao TC, de Lorenzo V. 2006. Transcriptional regulators a la carte: engineering neweffector specificities in bacterial regulatory proteins. Curr. Opin. Biotechnol. 17:34–42
33. Gardner TS, Cantor CR, Collins JJ. 2000. Construction of a genetic toggle switch inEscherichia coli. Nature 403:339–42
34. Gillespie DT. 1976. A general method for numerically simulating the stochastic timeevolution of coupled chemical reactions. J. Comput. Phys. 22:403–34
35. Guet CC, Elowitz MB, Hsing W, Leibler S. 2002. Combinatorial synthesis of geneticnetworks. Science 296:1466–70
36. Guido NJ, Wang X, Adalsteinsson D, McMillen D, Hasty J, et al. 2006. A bottom-upapproach to gene regulation. Nature 439:856–60
37. Guntas G, Mansell TJ, Kim JR, Ostermeier M. 2005. Directed evolution of proteinswitches and their application to the creation of ligand-binding proteins. Proc. Natl. Acad.Sci. USA 102:11224–29
38. Hasty J, McMillen D, Collins JJ. 2002. Engineered gene circuits. Nature 420:224–3039. Hyun Park S, Zarrinpar A, Lim WA. 2003. Rewiring MAP kinase pathways using alterna-
tive scaffold assembly mechanisms. Science 299:1061–6440. Kobayashi H, Kaern M, Araki M, Chung K, Gardner TS, et al. 2004. Programmable cells:
interfacing natural and engineered gene networks. Proc. Natl. Acad. Sci. USA 22:8414–1941. Kramer BP, Viretta AU, Daoud-El-Baba M, Aubel D, Weber W, Fussenegger M. 2004.
An engineered epigenetic transgene switch in mammalian cells. Nat. Biotechnol. 22:867–7042. Levskaya A, Chevalier AA, Tabor JJ, Simpson ZB, Lavery LA, et al. 2005. Synthetic biology:
engineering Escherichia coli to see light. Nature 438:441–4243. Mayo AE, Setty Y, Shavit S, Zaslaver A, Alon U. 2006. Plasticity of the cis-regulatory input
function of a gene. PLoS Biol. 4:e4544. McAdams HH, Arkin A. 2000. Gene regulation: towards a circuit engineering discipline.
Curr. Biol. 10:R318–2045. McDaniel R, Weiss R. 2005. Advances in synthetic biology: on the path from prototypes
to applications. Curr. Opin. Biotechnol. 16:476–8346. Olsen M, Iverson B, Georgiou G. 2000. High-throughput screening of enzyme libraries.
Curr. Opin. Biotechnol. 11:331–3747. Papworth M, Kolasinska P, Minczuk M. 2006. Designer zinc-finger proteins and their
applications. Gene 366:27–38
18 Haseltine · Arnold
Ann
u. R
ev. B
ioph
ys. B
iom
ol. S
truc
t. 20
07.3
6:1-
19. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
05/
08/0
7. F
or p
erso
nal u
se o
nly.

ANRV311-BB36-01 ARI 1 April 2007 9:12
48. Paulsson J. 2004. Summing up the noise in gene networks. Nature 427:415–1849. Pedraza JM, van Oudenaarden A. 2005. Noise propagation in gene networks. Science
307:1965–6950. Rackham O, Chin JW. 2005. A network of orthogonal ribosome x mRNA pairs. Nat. Chem.
Biol. 1:159–6651. Rao CV, Arkin AP. 2001. Control motifs for intracellular regulatory networks. Annu. Rev.
Biomed. Eng. 3:391–41952. Rawlings JB, Ekerdt JG. 2002. Chemical Reactor Analysis and Design Fundamentals. Madison,
WI: Nob Hill Publ.53. Reed JL, Palsson BO. 2003. Thirteen years of building constraint-based in silico models
of Escherichia coli. J. Bacteriol. 185:2692–9954. Shultzaberger RK, Bucheimer RE, Rudd KE, Schneider TD. 2001. Anatomy of Escherichia
coli ribosome binding sites. J. Mol. Biol. 313:215–2855. Solem C, Jensen PR. 2002. Modulation of gene expression made easy. Appl. Environ.
Microbiol. 68:2397–40356. Sprinzak D, Elowitz MB. 2005. Reconstruction of genetic circuits. Nature 438:443–4857. Stewart WE, Caracotsios M, Sørensen JP. 1992. Parameter estimation from multiresponse
data. AIChE J. 38:641–5058. Strogatz SH. 1994. Nonlinear Dynamics and Chaos: With Applications to Physics, Biology,
Chemistry and Engineering. Cambridge, MA: Westview59. Suel GM, Garcia-Ojalvo J, Liberman LM, Elowitz MB. 2006. An excitable gene regulatory
circuit induces transient cellular differentiation. Nature 440:545–5060. Suzuki M, Christians FC, Kim B, Skandalis A, Black ME, Loeb LA. 1996. Tolerance of
different proteins for amino acid diversity. Mol. Divers. 2:111–1861. Deleted in proof62. Umeno D, Tobias AV, Arnold FH. 2005. Diversifying carotenoid biosynthetic pathways
by directed evolution. Microbiol. Mol. Biol. Rev. 69:51–7863. van Kampen NG. 1992. Stochastic Processes in Physics and Chemistry. Amsterdam, The Neth.:
Elsevier. 2nd ed.64. Weiss R, Basu S. 2002. The device physics of cellular logic gates. Presented at 1st Workshop
Non-Silicon Comput., Cambridge, MA65. Weiss R, Homsy GE, Knight TF Jr. 1999. Toward in vivo digital circuits. In Dimacs
Workshop on Evolution as Computation, ed. L Landweber, E Winfree, R Lipton, S Freeland,pp. 275–95. Princeton, NJ: Springer
66. Yokobayashi Y, Weiss R, Arnold FH. 2002. Directed evolution of a genetic circuit. Proc.Natl. Acad. Sci. USA 9:16587–91
67. You L, Cox RS III, Weiss R, Arnold FH. 2004. Programmed population control by cell-cellcommunication and regulated killing. Nature 428:868–71
www.annualreviews.org • Synthetic Gene Circuits 19
Ann
u. R
ev. B
ioph
ys. B
iom
ol. S
truc
t. 20
07.3
6:1-
19. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
05/
08/0
7. F
or p
erso
nal u
se o
nly.

Contents ARI 12 April 2006 20:14
Annual Reviewof Biophysics andBiomolecularStructure
Volume 35, 2006Contents
FrontispieceMartin Karplus � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �xii
Spinach on the Ceiling: A Theoretical Chemist’s Return to BiologyMartin Karplus � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 1
Computer-Based Design of Novel Protein StructuresGlenn L. Butterfoss and Brian Kuhlman � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �49
Lessons from Lactose PermeaseLan Guan and H. Ronald Kaback � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �67
Evolutionary Relationships and Structural Mechanisms of AAA+ProteinsJan P. Erzberger and James M. Berger � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �93
Symmetry, Form, and Shape: Guiding Principles for Robustness inMacromolecular MachinesFlorence Tama and Charles L. Brooks, III � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 115
Fusion Pores and Fusion Machines in Ca2+-Triggered ExocytosisMeyer B. Jackson and Edwin R. Chapman � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 135
RNA Folding During TranscriptionTao Pan and Tobin Sosnick � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 161
Roles of Bilayer Material Properties in Function and Distribution ofMembrane ProteinsThomas J. McIntosh and Sidney A. Simon � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 177
Electron Tomography of Membrane-Bound Cellular OrganellesTerrence G. Frey, Guy A. Perkins, and Mark H. Ellisman � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 199
Expanding the Genetic CodeLei Wang, Jianming Xie, and Peter G. Schultz � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 225
Radiolytic Protein Footprinting with Mass Spectrometry to Probe theStructure of Macromolecular ComplexesKeiji Takamoto and Mark R. Chance � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 251
v
Ann
u. R
ev. B
ioph
ys. B
iom
ol. S
truc
t. 20
07.3
6:1-
19. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
05/
08/0
7. F
or p
erso
nal u
se o
nly.

Contents ARI 12 April 2006 20:14
The ESCRT Complexes: Structure and Mechanism of aMembrane-Trafficking NetworkJames H. Hurley and Scott D. Emr � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 277
Ribosome Dynamics: Insights from Atomic Structure Modeling intoCryo-Electron Microscopy MapsKakoli Mitra and Joachim Frank � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 299
NMR Techniques for Very Large Proteins and RNAs in SolutionAndreas G. Tzakos, Christy R.R. Grace, Peter J. Lukavsky, and Roland Riek � � � � � � � � � � 319
Single-Molecule Analysis of RNA Polymerase TranscriptionLu Bai, Thomas J. Santangelo, and Michelle D. Wang � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 343
Quantitative Fluorescent Speckle Microscopy of CytoskeletonDynamicsGaudenz Danuser and Clare M. Waterman-Storer � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 361
Water Mediation in Protein Folding and Molecular RecognitionYaakov Levy and José N. Onuchic � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 389
Continuous Membrane-Cytoskeleton Adhesion Requires ContinuousAccommodation to Lipid and Cytoskeleton DynamicsMichael P. Sheetz, Julia E. Sable, and Hans-Günther Döbereiner � � � � � � � � � � � � � � � � � � � � � � � 417
Cryo-Electron Microscopy of Spliceosomal ComponentsHolger Stark and Reinhard Lührmann � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 435
Mechanotransduction Involving Multimodular Proteins: ConvertingForce into Biochemical SignalsViola Vogel � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 459
INDEX
Subject Index � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 489
Cumulative Index of Contributing Authors, Volumes 31–35 � � � � � � � � � � � � � � � � � � � � � � � � � � � 509
Cumulative Index of Chapter Titles, Volumes 31–35 � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 512
ERRATA
An online log of corrections to Annual Review of Biophysics and Biomolecular Structurechapters (if any, 1997 to the present) may be found athttp://biophys.annualreviews.org/errata.shtml
vi Contents
Ann
u. R
ev. B
ioph
ys. B
iom
ol. S
truc
t. 20
07.3
6:1-
19. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
alif
orni
a In
stitu
te o
f T
echn
olog
y on
05/
08/0
7. F
or p
erso
nal u
se o
nly.


![synthetic biological circuits - arXivarXiv:1306.4493v1 [cs.LO] 19 Jun 2013 A temporal logic approach to modular design of synthetic biological circuits Ezio Bartocci1, Luca Bortolussi2,3,](https://img.dokumen.tips/doc/110x75/5f0ab9fa7e708231d42d0a99/synthetic-biological-circuits-arxiv-arxiv13064493v1-cslo-19-jun-2013-a-temporal.jpg)


![Tools and Principles for Microbial Gene Circuit Engineeringwang.bio.ed.ac.uk/wang/sites/sbsweb2.bio.ed.ac.uk... · program synthetic gene circuits [7,8]. Synthetic biology has often](https://img.dokumen.tips/doc/110x75/5ec793b1581c6f7c9e1752d4/tools-and-principles-for-microbial-gene-circuit-program-synthetic-gene-circuits.jpg)























