Embed Size (px)
Citation preview

Synaptic plasticity in sleep: learning,homeostasis and diseaseGordon Wang1,2, Brian Grone1, Damien Colas3, Lior Appelbaum4 andPhilippe Mourrain1,5
1 Department of Psychiatry and Behavioral Sciences, Center for Sleep Sciences, Beckman Center, Stanford University, Palo Alto, CA
94305, USA2 Department of Molecular and Cellular Physiology, Beckman Center, Stanford University, Palo Alto, CA 94305, USA3 Department of Biology, Stanford University, Palo Alto, CA 94305, USA4 Faculty of Life Sciences, Bar-Ilan University, Ramat Gan, 52900, Israel5 INSERM 1024, Ecole Normale Superieure, Paris, 75005, France
Review
Sleep is a fundamental and evolutionarily conservedaspect of animal life. Recent studies have shed lighton the role of sleep in synaptic plasticity. Demonstra-tions of memory replay and synapse homeostasis sug-gest that one essential role of sleep is in theconsolidation and optimization of synaptic circuits toretain salient memory traces despite the noise of dailyexperience. Here, we review this recent evidence andsuggest that sleep creates a heightened state of plasticity,which may be essential for this optimization. Further-more, we discuss how sleep deficits seen in diseases suchas Alzheimer’s disease and autism spectrum disordersmight not just reflect underlying circuit malfunction, butcould also play a direct role in the progression of thosedisorders.
IntroductionWhile we all experience sleep, and so believe we knowwhatit is, sleep remains a scientific enigma. A conclusive defi-nition of sleep has eluded researchers and probably willcontinue to do so until the function of sleep is fully eluci-dated. Nevertheless, a working description of sleep as anelectrophysiologically and behaviorally defined state hasbeen established since the middle of the 20th century [1,2].In animals with a developed neocortex, including mam-mals and birds, sleep states are defined by specific patternsof whole-brain activity detected by an electroencephalo-graph (EEG), along with eye movement electrooculogram(EOG) and muscle tone electromyogram (EMG) patterns.Non-rapid eyemovement sleep (NREM) is characterized byhigh-voltage synchronized slow waves of electrical activitythroughout the cortex and is referred to as slow-wave sleep(SWS) in its most synchronized form. Rapid eye movement(REM) sleep is characterized by rapid eye movement,muscle paralysis and low-voltage irregular EEG wavessimilar to waves observed during wakefulness [3].
During the early 1980s, Irene Tobler extended thisdefinition of sleep using additional behavioral criteria[4–6]: (i) decreased behavioral activity (immobility); (ii) sitepreference (e.g. bed); (iii) specific posture (e.g. lying); (iv)
Corresponding authors: Wang, G. ([email protected]); Mourrain, P.([email protected]).
452 0166-2236/$ – see front matter � 2011 Elsevier Ltd. All rights reserved
rapid reversibility (unlike coma); and, most importantly,(v) increased arousal threshold (offline state, no perceptionof the environment); and (vi) homeostatic control (sleeprebound after sleep deprivation). As of today, using theabove criteria, sleep has been documented and studied ina wide range of vertebrates and invertebrates [7] andthere is currently no clear evidence of an animal speciesthat does not sleep [8]. The existence of an ancestralsleep state, combined with evidence that prolonged sleepdeprivation leads to death in rats [9], fruit flies [10] andhumanswith fatal familial insomnia [11], strongly supportsthe hypothesis that sleep function serves a universalphysiological need.
Using the above electrophysiological and behavioralcriteria, major progress has been made in decipheringthe mechanisms regulating sleep and wake states. Brainnuclei, circuits, neurotransmitters and genes involved insleep–wake regulation and state switch have been identi-fied [12,13], but the most fundamental question remains:why do we sleep? Diverse theories have been postulated toaccount for the restorative effect of sleep and the impor-tance of sleep for cognitive performance [14–19]. Sleepprobably has multiple functions, but the strongest experi-mental evidence supports a primary role for sleep in theregulation of brain plasticity and cognition. Sleep depriva-tion impairs performance in motor and cognitive tasks [20]and sleep strengthens cognitive functions, including visualdiscrimination [21], motor learning [22] and insight (gain-ing explicit understanding of an implicit rule) [23]. Evi-dence has been gathered at the behavioral, neuronal,synaptic and molecular levels indicating that sleep pro-motes neural plasticity. Recent work in mammalian andnon-mammalianmodels highlights the importance of sleepfor synaptic remodeling and homeostasis (Table 1). In thisreview, we focus on the evidence for the role of sleep insynapse plasticity, a function conserved across animalphyla and critical for learning and memory as well assynaptic function and homeostasis.
Learning, memory and plasticity consolidationThe facilitation of memory retention is the most widelyaccepted and experimentally supported hypothesisexplaining the neuronal need for sleep. Although learning
. doi:10.1016/j.tins.2011.07.005 Trends in Neurosciences, September 2011, Vol. 34, No. 9

Table 1. Species commonly used in the study of neural plasticity during sleepa
Speciesb Year in which sleep
state was first described
and the criteria used
Evidence for the role of
sleep in neural plasticity
Experimental approaches usedc
[TD$INLINE]
Humans (Homo sapiens)
1935 [125]; EEG � Learning leads to reactivation
of hippocampus during SWS [43]
� Odors can enhance hippocampal
activation and memory retention
[43]
� Dreaming of a task is associated
with enhanced memory [22,38]
� fMRI and EEG data in subjects
following learning trials
� Increased spatial memory
performance
[TD$INLINE]
Domestic cat (Felis catus)
1936 [126]; EEG Ocular dominance column
plasticity following monocular
deprivation is enhanced by
sleep [51]
Microelectrode recordings and
optical imaging of visual cortex
[TD$INLINE]
Brown rat (Rattus norvegicus)
1961 [127]; EEG � Specific microcircuits activated
during wake are reactivated
during sleep [42]
� Circuit activity is synchronized
between hippocampus and
cortex [41]
� Sleep regulates neurogenesis
and genes associated with neural
function and plasticity [128]
� Multi-electrode array recordings
from hippocampus and cortex
of rats during sleep following
spatial learning tasks
� Microarray gene expression
profiling
� BrDU labeling for neurogenesis
[TD$INLINE]
Mouse (Mus musculus)
1964 [129]; EEG Wakefulness shown to induce
synaptic plasticity in HCRT
neurons [130]
Extracellular and whole-cell
voltage clamp recording of
mouse hypothalamic slices
[TD$INLINE]
Fruit fly (Drosophila melanogaster)
2000 [131]
Behavioral criteria:
immobility; prone position;
arousal threshold;
homeostatic regulation
Sleep reduces synapse number
throughout the CNS and in
specific sets of neurons [81–83]
Measurement of synaptic
components throughout the
CNS and quantification of
synapses in specific neurons
using EGF-labeled synaptic
proteins
[TD$INLINE]
Zebrafish (Danio rerio)
2001 [132]
Behavioral criteria:
immobility; floating
head-down or horizontal;
decreased arousal in
response to gentle tapping;
homeostatic regulation
Homeostatic and circadian plasticity
demonstrated in the hypocretin
circuit connecting the hypothalamus
to pineal and hindbrain [84]
Two-photon fluorescence
microscopy of EGFP-labeled
synapses in living larvae
[TD$INLINE]
Worm (Caenorhabditis elegans)
2008 [94]
Behavioral criteria:
immobility; arousal
threshold; homeostatic
regulation
Conserved EGF signaling induces
Lethargus [133], a sleep-like state
correlated with synapse
remodeling [98,99]
Currently correlational only
aOver the past 75 years, studies of sleep have been extended from EEG-based definitions in mammals to behaviorally based sleep criteria encompassing animals with
simpler nervous systems. Studies in each species have contributed valuable new information about the effects of sleep on neural plasticity and learning.
bImage credits (from top to bottom): Katharine Mach; emptysound; Eddy Van 3000; bundu; Max Westby; Brian Grone; and A.J. Cann.
cAbbreviations: BrdU, 5-Bromodeoxyuridine; fMRI, functional magnetic resonance imaging.
Review Trends in Neurosciences September 2011, Vol. 34, No. 9
453

Review Trends in Neurosciences September 2011, Vol. 34, No. 9
mostly occurs during wake, sleep is of critical importancefor memory processes. Sleep greatly enhances both theencoding and consolidation of memory [18,19,24]. Ade-quate sleep is necessary, both before and after an event,for that event to be properly encoded and stored in long-term memory [18,19,25]. Sleep-deprived humans havesignificantly impaired memory retention and degradedperformance in memory encoding [26–28]. Long periodsof sleep are clearly beneficial but gains in memorizationperformance have also been reported after short sleepperiods. Recall of events is stronger and more accurateafter a daytime nap as brief as a few minutes, as comparedto a similar wake period [29–31].
Quality of memory consolidation is not only a function oftime spent asleep, but can also vary depending on the typeof memories, the relevance of the memorized event and themotivation to remember. Following sleep, proceduralmemories (i.e. memorization of cognitive and motor skills)have been shown to benefit more than declarative memo-ries (i.e. recollection of experiences and information)[19,32]. Furthermore, sleep had a stronger stabilizingeffect on memories of tasks or events when there was aconscious effort or an incentive to memorize those. Simplyput, conscious learning of a motor task associated with apotential reward generates memories that profit the mostfrom sleep-dependent consolidation, in contrast to uncon-scious and/or unmotivated learning of the same task[19,32]. This uneven and contextual influence of sleep ondifferent classes of memory suggests an intriguing possi-bility that sleep-dependent and sleep-independent plastic-ity coexist and interact in the circuits and brain regionsresponsible for encoding and storing the different memorytypes.
Although behavioral observations have shown thatsleep as a whole is clearly important for memory consoli-dation, the roles of the different sleep phases are still beingdeciphered. Because of its relationship with dreams, REMsleep was first suspected to be critical for memory forma-tion, but most of the EEG studies performed so far havereported that NREM, especially SWS, sleep is critical formemory retention. SWS and/or NREM sleep deprivationafter learning prevents subsequent consolidation and en-hancement of memories [19,24]. Consistent with this ob-servation, stimulation of slow-wave oscillations duringsleep enhances the retention of same-day memory tracesfor next-day retrieval [33]. Although SWS seems to have aprimary role in memory formation, it is still unclear howother sleep phases participate in memory encoding andconsolidation. NREM sleep spindles, for example, havebeen shown to be important for consolidation [34] andmore recently encoding and/or learning capabilities [25].REM sleep has also been associated with emotion-relatedmemories [18]. Finally, in opposition to a dichotomous viewassociating a specific sleep stage with a specific type ofmemory, it has also been postulated that the sequence inwhich phases appear in normal sleep (i.e. NREM–REMsuccession) could be more important for optimal consolida-tion, whatever the memory type, than the duration of eachstage [35]. A better understanding of the molecular andphysiological mechanisms generating the different sleepstages should shed light on their roles in hippocampal
454
and/or cortical circuit plasticity and the different typesof memory.
An intriguing and important mechanism proposed forthe facilitation of memory consolidation is the replay ofmemory traces in hippocampal and cortical circuits duringsleep (reviewed in [19,36,37]). Firing patterns recordedduring wakefulness can be replayed during the followingNREM sleep period [19,37] and sometimes REM [38]. Inneurons of the zebra finch song system, replay of patternsof bursts corresponding to singing sequence was observedduring sleep [39,40]. In rats, neuronal activation patternsrecorded during maze learning are recreated during SWS[41,42]. The human hippocampus is similarly reactivatedduring SWS following learning of a spatial task and thestrength of this reactivation is associated with fidelity oflearning [43]. Importantly, the reactivation of memories inhumans by presenting, during SWS, odor or noise cues thatwere also present during learning leads to enhanced mem-ory consolidation [44–46] and increased resistance of thatmemory to interference [46]. During a NREM nap, mentalactivity related to a spatial memory task is associated withenhanced memory consolidation [38]. Consistently, reacti-vation in SWSwas correlated to activations of hippocampaland neocortical regions critical to learning and memory[46]. Interestingly, replay happens during the first15–30 min of sleep, when mammals are in SWS. Duringthis SWS period, reactivated circuits undergo synapticconsolidation according to the replay hypothesis, whereasothers could be pruned according to the synaptic homeo-stasis hypothesis (SHH; see below). One could speculatethat both hypotheses are not exclusive and that replaymechanisms could be important to protect fragile circuitsagainst global synaptic downscaling.
Although these recent reinstatement data are compel-ling, replay as a sleep-dependent mechanism for memoryconsolidation still remains to be fully established. Replayhas mostly been studied in extensively trained rodents,except in a few cases [47] and, thus, it might also reflect thefiring of well-entrained circuitry. Moreover, replay is ex-tremely transient and labile, and only a few studies havesuccessfully investigated its function in memory transferfrom the hippocampus to the neocortex (e.g. [48]). It isimportant to mention here that replay also occurs duringwake, when it can similarly affect learning and memoryconsolidation [49,50]. Reactivation of memories by odor-ants during sleep and during wake, however, activatesdifferent brain regions and elicits very different memoryresponses. Odor cues that were present during learningactivate hippocampal and posterior cortical regions andstrengthen object-location memories when presented dur-ing sleep, but weaken those memories and activate mainlyprefrontal cortical regions when presented during wake-fulness [46]. Clearly, more work needs to be done to uncov-er the molecular and circuit properties of sleep–wakegating of brain activity and effects of memory reactivationon consolidation.
Consistent with the replay and reactivation studies,sleep is believed to consolidate synaptic connections re-quired for encoding and retention of memories. Currently,the mechanisms underpinning synaptic consolidationduring sleep in these hippocampal and cortical memory

Review Trends in Neurosciences September 2011, Vol. 34, No. 9
storage circuits remain unknown. Sleep has, however,already proven to be critical for consolidation of oculardominance plasticity (ODP), a type of cortical plasticitywidespread in mammals and particularly well studied incats. In ODP, deprivation of vision in one eye (monoculardeprivation, MD) leads to increased rewiring of visualcortex by the non-deprived eye [51–53]. Interestingly,when MD is followed by just a few hours of sleep, corticalresponses to non-deprived eye stimulation are strength-ened [51]. Furthermore, cortical consolidation was found tobe correlated with the amount of NREM [51]. This findingsuggests that sleep, especially NREM, has a critical func-tion in cortical synaptic remodeling.
More recently, sleep-dependent consolidation in ODPwas disrupted when major molecular actors of synapticpotentiation and plasticity such as NMDA receptors(NMDARs) and protein kinase A (PKA) were antagonized[53]. Increased phosphorylation and activation of down-stream targets of these pathways [e.g. extracellular sig-nal-regulated kinase (ERK), Ca2+-calmodulin-dependentprotein kinase (CaMKII) and the AMPA receptor (AMPAR)GluR1subunit]were observed onlyafterpost-MDsleep [53].These data show that sleep can change the strength ofneuronal connections and that pathways involved in synap-tic plasticity are activated. Furthermore, all these datasuggest that synchronous reactivation of behaviorally rele-vant neural circuits during sleep can mediate meaningfuland functionally relevant changes in the brain, and furtherdissection of the molecular mechanisms underlying theseactivity states is critical to the understanding of sleep in theconsolidation and optimization of brain circuit function.
Homeostatic control of synaptic plasticityThe synaptic homeostasis hypothesis
The memory consolidation hypothesis proposes a specificmechanism of synapse modification by which the encodingof memory traces is rendered more efficient through mod-ification of relevant synapses. Recently a new hypothesis,the SHH, has emerged that postulates that sleep globallydownscales all synapses to compensate for the net increasein synapse formation and strength during wake [14,54](Figure 1a). The SHH assumes that wakefulness causesnet cortical synaptic potentiation throughout the brain,and this potentiation drives slow-wave activity (SWA)during NREM sleep. This SWA-mediated downscaling ofsynaptic strength has a beneficial effect on neuronal effi-ciency and function. Indeed, mathematical modeling sug-gests that changes in synaptic strength can explain suchchanges in SWS intensity [55]. The formulation of the SHHis based on the observation that synaptic density and am-plitude of long-term potentiation (LTP) increase duringexploration of enriched environments [56,57] and followingextendedmechanical stimulation of sensorymodalities [58].This daily type of anatomical and physiological increase insynaptic function is also associated with the modification ofgene expression and neural chemical systems that arecritical to the expression and maintenance of synapticpotentiation. Changes in cAMP-responsive element bindingprotein (CREB), activity-regulated cytoskeleton-associatedprotein (Arc), brain-derived neurotrophic factor (BDNF),AMPAR subunits [59], Homer, neuronal activity regulated
pentraxin (NARP) [60–62], acetylcholine [63] and norepi-nephrine [64] have been observed, although it should benoted that Arc and Homer are also required for long-termdepression (LTD) and/or homeostatic downscaling. Thus,theSHHprovides a compelling set of testablehypotheses fordescribing the synaptic changes in sleep.
Waking is not merely associated with increases in syn-aptic strength; the duration of wake time also controls theamplitude and duration of SWA. Moreover, sensory ormechanical stimulation that extends wakefulness leadsto higher-amplitude slow waves in NREM sleep withsteeper slopes and fewer multi-peak waves [65]. Further-more, this increase in SWA appears to be locally regulatedas behavioral tasks designed to activate a single corticalregion elevated the level of SWA specifically in that regionduring SWS [66]. Thus, there is support for the first twotenets of the SHH: synaptic strength is probably increasedduring wake, and this increase does regulate the amount ofSWA during SWS. The final conjecture that SWA is asso-ciated with synaptic downscaling is yet to be directlyobserved in mammalian species. Still, there is suggestiveevidence in rodents from cortical evoked responses andlocal field potential recording during sleep [59] and recentanalysis of spontaneous synaptic events [67,68] showingthat synaptic downscaling may occur. Nonetheless, themost convincing demonstration of this sleep-dependentdecrease in synaptic strength and connectivity appearsin non-mammalian and non-cortical neural circuits in fruitflies and zebrafish (Figure 1). These recent data show thathomeostasis of synaptic strength during sleep is neither anovel invention of mammals nor the sole purview of thecortex, but a more ancestral function, inherent in synapticcircuits, that is essential for the proper maintenance andefficient operation of networks of connected neurons.
The idea that synapses can be homeostatically regulat-ed is not unique to SHH; it is a documented phenomenon inthe field of synaptic scaling. In synaptic scaling, bidirec-tional changes of strength of individual synapses inducecompensatory changes in synaptic strength that are pro-portional across multiple synapses of a given postsynapticneuron [69–73]. Thus, modulation of network activity leadsto the uniform scaling of synaptic strength across groups ofsynapses or entire neurons [74,75]. This allows neurons tonormalize their output without changing the relative sig-naling strength of individual synapses, thus presumablymaintaining the information fidelity of the system. Synap-tic scaling can occur presynaptically or postsynaptically,and can involve changes in intrinsic excitability, inhibitoryand/or excitatory synaptic strength and number, or meta-plasticity (adjusting the extent of other forms of plasticity).Molecular mechanisms mediating synaptic scaling includesoluble factors [e.g.BDNFand tumornecrosis factor (TNF)],trans-synaptic signalingandcell adhesionmolecules [e.g.b3integrin and major histocompatibility complex (MHC1)],and intracellular signaling molecules [e.g. CaMKs, Arc,polio-like kinase 2 (PLK2), and cyclin-dependent kinase 5(CDK5)]. According to the synaptic scaling model, loweractivity levels or quiescent network states should increasesynaptic strength rather than downscale them, whichappears to be counterintuitive. One must, however, notethat sleep isnot aquiescent state: the energyuse of thebrain
455

Eye
HypPG HB
OT
ALOL
MB
ALOL
MB
Adult Drosophila brain
Larval zebrafish
CC
CC
DAY NIGHT
SLEEP
Synaptic homeostasisMemory consolidationCircadian regulation
WAKESynaptic potentiationLearningCircadian regulation
RestedSleep deprived
IsolatedEnriched
Decreased synaptic density and strength
High synaptic densityand strength
(a)
(c)
(b)
(f)
ALOL
MB
(d)
Day Night
Day Night
Sleep deprived Sleep
CC
Eye
HypPG HB
OT
(e)
Syn
apse
str
engt
h
TRENDS in Neurosciences
Figure 1. Summary of recent data in support of the synaptic homeostasis hypothesis (SHH). (a) Synapses, like learning and memories, are known to be affected by circadian
rhythms and homeostatic regulation. The SHH posits that synapse accumulation during the day drives a need for synaptic downscaling, which preferentially occurs during
sleep. (b–f) Recent studies from diurnal fruit fly Drosophila melanogaster and zebrafish Danio rerio have demonstrated increased synapse components or synapse numbers
following wake, or sleep deprivation. Images are not all to the same scale. (b) Bruchpilot (BRP; an essential constituent of the active zone of all synapses) levels were
measured in antennal lobes (AL), b lobes of the mushroom bodies (MB) and the ellipsoid body of the central complex (CC) in Drosophila [81]. BRP immunofluorescence was
found to be increased in animals sleep deprived for 16 h compared to rested controls (shown false-colored on a quantitative scale, with yellow indicating highest levels). (c)
Following social enrichment, sleep deprivation was found to lead to the retention of more synaptic terminals in Drosophila olfactory lobes (OL). Discs-large (DLG), a
postsynaptic protein, was fused to GFP expressed in pigment dispersing factor (PDF) neurons via a GAL4;UAS approach (i.e. pdf-GAL4/1::UAS-dlgWT-gfp/1). Social
enrichment led to increased numbers of GFP-positive terminals that recovered to baseline levels following sleep but not following sleep deprivation [83]. (d) Synaptotagmin
(a presynaptic protein) was fused to enhanced GFP (EGFP) and expressed in the g lobe of the MB. Right panels show a higher magnification of the area indicated by the
yellow square in the left panel. Sleep-deprived flies were found to contain larger GFP-positive puncta in the MB compared to sleeping controls [82]. Scale bar = 10 mm. (e)
Both circadian clock and sleep regulate synapse number rhythmicity in zebrafish. Sleep deprivation interferes with homeostatic downscalling of synapse number in larval
zebrafish (7 days old) [84]. Live transgenic fish expressing synaptophysin (SYP) fused to EGFP in hypocretin neurons (i.e. HCRT:SYP-EGFP) displayed significantly more
EGFP puncta in axons projecting to the pineal gland (PG) during diurnal wakefulness compared to the nocturnal sleep period. Red arrows depict additional synapses that
were not observed during the sleep period in the same fish. (f) Live larval zebrafish expressing HCRT:SYP-EGFP also display rhythmic EGFP puncta in hindbrain (HB)
projections from the hypocretin neurons [84]. Red arrows indicate additional synapses that were not observed during the sleep period in the same fish. Abbreviations: Hyp,
hypothalamus; OT, optic tectum. Reproduced, with permission, from [81] (b), [83] (c) and [82] (d).
Review Trends in Neurosciences September 2011, Vol. 34, No. 9
456

Review Trends in Neurosciences September 2011, Vol. 34, No. 9
during sleep is not significantly lower than duringwake andappears to even increase during the onset of SWS [76].Furthermore, the apparent synchrony and slow EEG oscil-lations of SWSdonot indicate thatneurons are firing less; infact, extracellular recordings in the cortex during NREMsleep show that there is an increase in high-frequency firing(>50 Hz with a peak at approximately 100 Hz) and low-frequency firing (<15 Hz with a peak at 3–5 Hz) with adecrease inmedium firing rate (15–50 Hz) [59]. Interesting-ly, the 100 Hz and 5 Hz firing rates that are exaggerated insleepareexactly the stimulation frequency for inducingLTPand LTD, respectively, in a broad range of neuronal pre-parations both in vivo and in vitro. Thus, we suggest thatsynapses are pruned, retuned and even added during sleepto optimize the information stored in the nervous system bymechanisms that might include synaptic scaling as well asHebbian plasticity.
Circadian and homeostatic control of synaptic plasticity
in the fruit fly and zebrafish
Although the SHH was originally postulated based onmammalian electrophysiological observations, its firstdemonstration at the molecular and neuronal levels camefrom studies in two non-mammalian species, namely, thefruit fly Drosophila melanogaster and the zebrafish Daniorerio. Several studies in Drosophila had previously shownthe existence of a day–night rhythm of neuronal structuralplasticity at the cellular level (Table 2). In the pigmentdispersing factor (PDF) circuit, rhythmic remodeling inaxonal terminals was reported [77]. Similarly, the mor-phology of flight neuromuscular terminals changes be-tween day and night [78], with a rhythm in synapticbouton size [79]. Finally, in the fly visual system, the sizeand morphology of monopolar cell arborization also variesrhythmically over a 24-h cycle [80]. In all these studies, thestructural plasticity rhythm was found to be regulated bythe circadian rhythm, because rhythmicity was main-tained in constant darkness [78,79], or controlled by clockgenes [77,80]. These data supported clock-controlled
Table 2. Current evidence for circadian and sleep regulation of strumodelsa
Morphological and structural
changes and regulatory process
Circuit and animal model
Circadian changes in the size of nuclei,
caliber of the axon and morphology
and length of the dendritic tree
Monopolar L2 cells in the visua
system in Drosophila
Circadian changes in synaptic
bouton size
Flight motor neurons in Drosop
Circadian remodeling in axonal
terminals
Pacemaker PDF neurons in
Drosophila
Homeostatic sleep-dependent control
of synapse number
PDF neurons in Drosophila
Homeostatic sleep-dependent control
of synapse size or number
PDF, visual system VS1 and mu
body gamma neurons in Droso
Circadian and homeostatic control of
synapse number
Hypocretin and/or orexin neuro
in larval zebrafish
aCurrently, the primary demonstration of rhythmic changes in synaptic circuits come
morphology and/or synaptic bouton density and structure have been shown to vary acco
the idea that overall synapse number decreases during sleep.
bAbbreviations: IHC, immunohistochemistry; mCD8, mouse lymphocyte marker CD8.
plasticity, but did not directly investigate the influenceof sleep on synapses.
Recent studies in the fruit fly demonstrate a sleep-dependent homeostatic process responsible for downregu-lation of synaptic components [81], downscaling of synapsenumber and a decrease in synapse volume and dendriticcomplexity (Figure 1 and Table 2) [82,83]. Quantification inthe fly central nervous system (CNS) of presynaptic andpostsynaptic proteins, including Bruchpilot (BRP) andDiscs-large (DLG), the Drosophila homolog of the verte-brate postsynaptic density protein PSD-95, showed thatsynaptic protein levels increase throughout wake but de-crease during sleep independently of the circadian time[81]. Sleep-deprived animals had up to 40% higher synap-tic component levels than animals allowed to sleep, strong-ly suggesting that sleep has a role throughout the CNS inrenormalizing synapses for the day to come (Figure 1b)[81]. This global sleep-dependent regulation of synapticmarkers was confirmed and strengthened by circuit-specificstudies investigating the precise influence of wake andsleep on synaptic terminals in the PDF circuit (Figure 1c)[83], as well as the g lobe of the mushroom bodies and thedendrites of a unique giant tangential neuron of the lobulaplate vertical system (VS1) (Figure 1d) [82,83]. Synapticclusterswere imaged and counted in transgenic flies expres-sing enhanced GFP (EGFP)-tagged constructs of either thepresynaptic proteins synaptobrevinand synaptotagmin, thepostsynaptic protein DLG, or actin to reveal the dendriticspines. Interestingly, synaptic terminalnumberandvolumeincreased in flies maintained in an enriched environmentand the number and volume of synaptic terminals werereduced during sleep [82,83]. This decline in synapticterminals was prevented by sleep deprivation [82,83]. Thisfinding, consistent with the SHH, revealed that sleepcan downscale structural synaptic connections that arepotentiated during waking experience.
A similar structural synaptic plasticity in a zebrafishneural circuit is regulated by both circadian and homeostat-ic processes. Transgenic fish expressing the presynaptic
ctural synaptic and circuit changes in non-mammalian animal
Experimental technique and/or
synaptic markersbRefs
l IHC on fixed tissue using membrane mCD8-GFP
and nuclear S65T-GFP fusion proteins
[78,80]
hila IHC using neuronal markers and the presynaptic
protein synaptotagmin
[79]
IHC using membrane mCD8-GFP and the
presynaptic fusion protein synaptobrevin-GFP
[77]
IHC using the postsynaptic localized fusion
protein DLG-GFP and the presynaptic
synaptobrevin-GFP fusion protein
[83]
shroom
phila
IHC using the presynaptic synaptotagmin-EGFP
marker and an actin-GFP fusion protein to
reveal the dendritic spines
[82]
ns Time-lapse two-photon live imaging using the
presynaptic synaptophysin-EGFP fusion protein
[84]
s from studies in diurnal Drosophila and zebrafish. In those systems, dendritic
rding to a daily rhythm, and the magnitude and direction of these changes support
457

Box 1. Current and future contributions of imaging modalities to studies of sleep and synapse modulation
Confocal and two-photon imaging
In vivo live imaging of Drosophila and zebrafish has already
contributed significantly to the study of synapse modulation by sleep
and circadian rhythms [82–84] (Figure I). However, live imaging of
mice and rat cortex is required to further extend these findings into
mammals, where the majority of sleep physiology has been done in
the past. Furthermore, network imaging using Ca2+ indicators in
sleeping and awake animals should provide an added level of detail
on the network firing patterns of the brain during the different periods
of sleep and wake.
Array tomography
The synaptic plasticity in sleep is probably mediated by changes in
protein expression on a global level, and the quantification of such
changes will be essential for furthering understanding of sleep. Array
tomography is a recent proteomic imaging technique [88,137] (Figure I).
Its advantages include the ability to visualize dozens of proteins across
entire cortical columns at the synaptic level of resolution and, thus,
should be a valuable tool for performing quantitative comparisons of
synaptic proteomic changes between tissues collected at different time
points during the day–night cycle.
Stochastic optical reconstruction microscopy (STORM)/photo-acti-
vated localization microscopy (PALM) and stimulated emission deple-
tion (STED)
Proteins that are regulated during sleep (e.g. kinases, channels and
receptors) are shuttled and modulated on a subsynaptic level.
STORM/PALM and STED are super-resolution imaging technologies
that enable single molecule resolution [138–142] (Figure I). These
technologies will provide the level of resolution needed to decipher
the actual molecular modifications occurring at synapses during
sleep.[(Figure_I)TD$FIG]
• Live imaging of spines and synaptic markers in mammals• Single neuron and network
analysis of neuronal function during sleep and wake using Ca2+ imaging
Synaptic punta in Drosophilaand zebrafish varies inaccordance with sleepand the circadian rhythm[82-84]
N/A
(a)
(b)
(c)
Tw
o-p
ho
ton
con
foca
lA
rray
tom
og
rap
hy
Today Tomorrow
N/A
ST
OR
M/
PA
LM
, ST
ED
• Characterization of sleep modulation across specific synapse subpopulation• Region-specific changes in synapse density and composition
• Subsynaptic analysis of changes in channel and protein kinases in sleep and wake• Causal molecular mechanism for synaptic modification in sleep, and whether it is strictly different from wake
TRENDS in Neurosciences
Figure I. Representative images to illustrate the type of images obtained using the different imaging modalities. (a) Two-photon image of transgenic zebrafish
expressing enhanced GFP in all hypothalamic hypocretin neurons. (b) Array tomography reconstruction of mouse cortical dendrite stained with yellow fluorescent
protein (green) and synaptotagmin (red) to label synapses. (c) Segmented microtubule bundles in mouse cortical white matter as imaged using stochastic optical
reconstruction microscopy (STORM). The colors are pseudocolors to separate out each individual microtubule. Reproduced, with permission, from Gordon Wang (a,b),
Nicholas C. Weiler and Xiaowei Zhuang (c).
Review Trends in Neurosciences September 2011, Vol. 34, No. 9
protein synaptophysin (SYP) fused to EGFP are a usefulmeans of following synaptic structures in transparentzebrafish [84]. This fusion protein was targeted to thehypothalamic hypocretin (HCRT) neurons (Figure 1e,f), awell-studied circuit involved in sleep–wake regulation[85–87]. Results from array tomography [88], a new proteo-mic imaging technique (Box 1), showed that the majority(>85%) of EGFP-labeled SYP presynaptic boutons were injuxtaposition with postsynaptic PSD 95, confirming thatSYP puncta represent good markers of structural synapses[84]. The optical clarity of larval zebrafish and infraredlight-based two-photon imaging allowed longitudinal anal-ysis in living zebrafish of the synaptic density in different
458
regions of the HCRT circuits (Box 1; Figure 1e,f). HCRTneuron synapse density waxed and waned according to acircadian rhythm [84]. Critically, synapse number was alsohomeostatically regulated by sleep [84]. Thus, sleep-de-prived animals were deficient in night-time synapticdownscaling. Overall, validation of the SHH in such phylo-genetically distant species as zebrafish and Drosophilastrongly suggests that the cellular processes demonstratedin synaptic potentiation and homeostasis have been con-served across evolution.
The aforementioned experiments demonstrate thatsynapses are dynamic during sleep andwake, and supporta sleep-dependent synaptic homeostasis. However,

Review Trends in Neurosciences September 2011, Vol. 34, No. 9
supplementary evidence needs to be gathered to validatethe SHH fully. First, none of the studies mentioned aboveproved a functional change in synaptic transmission orshowed whether the changes in synaptic density actuallyaffected the function of the circuit or the neurons withinthat circuit. This type of functional analysis will be criticalfor extending understanding of synapse modification dur-ing sleep to explain the physiological role of sleep. Itwill beimportant to demonstrate that synapses are lost or gainedby a selective mechanism that effectively reduces thephysical footprint of memories without losing the detailsof that memory. Second, in the first studies that directlyshowed changes in synapse density, the synapses in ques-tion are in circuits (i.e. PDF [83] and HCRT [84]) known tobe involved in circadian and sleep rhythm regulation.Although the recent evidence in mushroom bodies andthe visual system of flies is a great step forward [82], it willbe important to extend studies of sleep regulation ofsynapse density throughout the nervous system. Finally,the SHH was primarily formulated on observations madein mammalian cortex. Thus, it is critical that sleep-medi-ated synapse density changes in mammalian neocortex beconvincingly demonstrated, and that this change is medi-ated by sleep homeostatic pressure and is positively cor-related with the amount of SWS. Recent advances inmolecular and live imaging techniques (Box 1) shouldenable unprecedented access to the fundamental mecha-nisms involved in synaptic changes during sleep.
What could be the primary ancestral function of sleep?Theaccumulation of evidence linking sleep to synaptic andcircuit plasticity in vertebrates and, more recently, inver-tebrates (Table 1) allows informed speculations aboutwhat could be the ancestral and primary role of sleep.Across distantly related animal models, sleep has beenshown to have a critical role in at least three mainmanifestations of circuit plasticity: brain and nervoussystem development, learning and memory, and synaptichomeostasis. Based on this observation, one convergenthypothesis is that sleep is primarily a plastic state for thedevelopment and remodeling of neural circuits. In view ofthese commonalities, sleep might be compared to a neu-rodevelopmental state: a functional state that has beenevolutionary preserved from simple circuits to neocorticalcomplex networks. In this hypothesis, the sleep stateallows critical plasticity mechanisms to be brought on-line to facilitate the making and breaking of connectionswithin neural circuits that, during the desynchronizedand unpredictable synaptic environment of wake, coulddisrupt behavior or learning.
In mammals, the amount of sleep is highest early in lifewhen maximal amounts of neural development are occur-ring [89,90]. Newborns spend the majority of their time in asleep state, and sleep has been shown to be critical fornervous system maturation [89,90]. Sleep deprivationstudies in young rodents led to a loss of brain plasticityassociatedwith reduced learning performance and negativelong-term cognitive and behavioral effects [91]. NREMseems particularly important as human neonates respondto sleep deprivation with compensatory increases only inNREM but not REM sleep time [92,93]. The critical role of
sleep during mammalian nervous system developmentmight reflect a highly evolutionary conserved process. In-deed, at the other extremity of the animal evolution ladder,sleep and development could be not only associated, but alsoessentially identical. In the wormCaenorhabditis elegans, adevelopmental stage called lethargus has also been charac-terized as a sleep-like state [94]. This developmental stageoccurs before each of the four larval molts. Interestingly,lethargus can be induced by the epidermal growth factor(EGF) signaling pathway [95], known for its involvement inneuronal differentiation and synaptic plasticity in mam-mals [96,97]. Although synaptic remodeling of the wormGABArgic system is known to occur during the first moltbefore the larvalL1–L2 transition [98,99], nodemonstrationfor a direct function of lethargus in this remodeling has beenshown in the worm yet. It is noteworthy that lethargus orsleep, similar to any developmental process, is preciselytimed. The timing of the molts has been correlated withthe oscillation of the C. elegans ortholog of the well-knowncircadian factor Period [100]. It is tempting to speculatebased on these correlations that sleep as a behavioral stateand its circadian regulation could originate from an ances-tral developmental state and its developmental timing pro-gram.
The mammalian and worm studies, coupled with thedemonstration of conserved synaptic homeostasis andrhythmic plasticity during sleep in both zebrafish larvae[84] and adult flies [81,83], also support the idea that theancestral sleep function could be the same during devel-opment and adulthood. Furthermore, sleep as a recurrentstate in normal brain function can be considered as anabridged version of brain development that recapitulates,on a limited scale, the activity-dependent global pruningand refining of connectivity following the increase in syn-apse density and strength during the earliest part of braindevelopment. Each day, sleep provides the same functionas provided during development by this early window ofpruning, rewiring synaptic networks guided by salientneurological activity and, thus, selectively potentiatingcertain important synapses while simultaneously down-scaling non-essential synaptic connections.
So, with the experimental knowledge gathered to datefrom memory consolidation, visual cortex wiring, and syn-aptic homeostasis studies, it is safe to acknowledge thatsleep, on a synaptic level, is a specific type of plastic stateprobably conserved across circuits, developmental stagesand evolution. This critical state is not only important forthe proper function of the nervous system, but is itself alsodependent on the prior activity and connectivity of thenervous system. Although the effects of sleep on synapticplasticity during normal physiological conditions will re-quire extensive studies for many years to come, pathologi-cal conditions such as observed in neurodegenerative andneurodevelopmental disorders should also shed light onthe association of abnormal sleep and cognition im-pairment.
Sleep abnormalities in cognitive disorders and relatedanimal modelsOur discussion thus far has focused on the role of sleep as amajor organizer of synapse and circuit plasticity in the
459
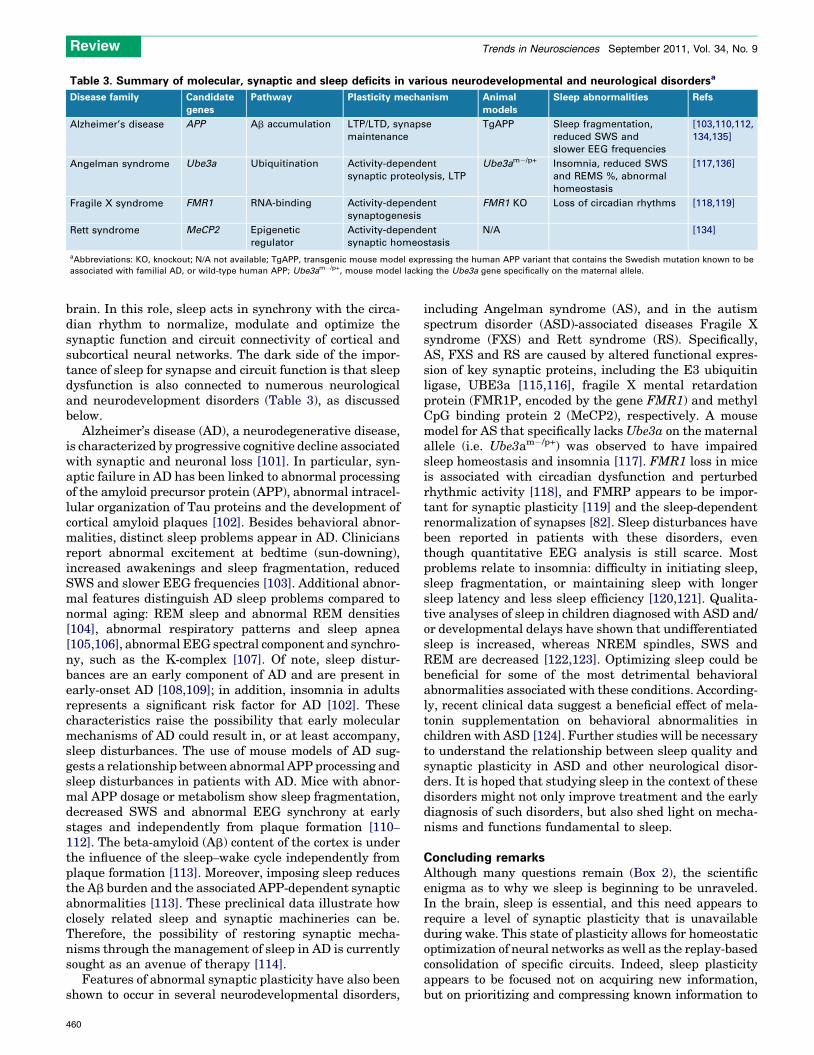
Table 3. Summary of molecular, synaptic and sleep deficits in various neurodevelopmental and neurological disordersa
Disease family Candidate
genes
Pathway Plasticity mechanism Animal
models
Sleep abnormalities Refs
Alzheimer’s disease APP Ab accumulation LTP/LTD, synapse
maintenance
TgAPP Sleep fragmentation,
reduced SWS and
slower EEG frequencies
[103,110,112,
134,135]
Angelman syndrome Ube3a Ubiquitination Activity-dependent
synaptic proteolysis, LTP
Ube3am�/p+ Insomnia, reduced SWS
and REMS %, abnormal
homeostasis
[117,136]
Fragile X syndrome FMR1 RNA-binding Activity-dependent
synaptogenesis
FMR1 KO Loss of circadian rhythms [118,119]
Rett syndrome MeCP2 Epigenetic
regulator
Activity-dependent
synaptic homeostasis
N/A [134]
aAbbreviations: KO, knockout; N/A not available; TgAPP, transgenic mouse model expressing the human APP variant that contains the Swedish mutation known to be
associated with familial AD, or wild-type human APP; Ube3am�/p+, mouse model lacking the Ube3a gene specifically on the maternal allele.
Review Trends in Neurosciences September 2011, Vol. 34, No. 9
brain. In this role, sleep acts in synchrony with the circa-dian rhythm to normalize, modulate and optimize thesynaptic function and circuit connectivity of cortical andsubcortical neural networks. The dark side of the impor-tance of sleep for synapse and circuit function is that sleepdysfunction is also connected to numerous neurologicaland neurodevelopment disorders (Table 3), as discussedbelow.
Alzheimer’s disease (AD), a neurodegenerative disease,is characterized by progressive cognitive decline associatedwith synaptic and neuronal loss [101]. In particular, syn-aptic failure in AD has been linked to abnormal processingof the amyloid precursor protein (APP), abnormal intracel-lular organization of Tau proteins and the development ofcortical amyloid plaques [102]. Besides behavioral abnor-malities, distinct sleep problems appear in AD. Cliniciansreport abnormal excitement at bedtime (sun-downing),increased awakenings and sleep fragmentation, reducedSWS and slower EEG frequencies [103]. Additional abnor-mal features distinguish AD sleep problems compared tonormal aging: REM sleep and abnormal REM densities[104], abnormal respiratory patterns and sleep apnea[105,106], abnormal EEG spectral component and synchro-ny, such as the K-complex [107]. Of note, sleep distur-bances are an early component of AD and are present inearly-onset AD [108,109]; in addition, insomnia in adultsrepresents a significant risk factor for AD [102]. Thesecharacteristics raise the possibility that early molecularmechanisms of AD could result in, or at least accompany,sleep disturbances. The use of mouse models of AD sug-gests a relationship between abnormal APP processing andsleep disturbances in patients with AD. Mice with abnor-mal APP dosage or metabolism show sleep fragmentation,decreased SWS and abnormal EEG synchrony at earlystages and independently from plaque formation [110–
112]. The beta-amyloid (Ab) content of the cortex is underthe influence of the sleep–wake cycle independently fromplaque formation [113]. Moreover, imposing sleep reducesthe Ab burden and the associated APP-dependent synapticabnormalities [113]. These preclinical data illustrate howclosely related sleep and synaptic machineries can be.Therefore, the possibility of restoring synaptic mecha-nisms through the management of sleep in AD is currentlysought as an avenue of therapy [114].
Features of abnormal synaptic plasticity have also beenshown to occur in several neurodevelopmental disorders,
460
including Angelman syndrome (AS), and in the autismspectrum disorder (ASD)-associated diseases Fragile Xsyndrome (FXS) and Rett syndrome (RS). Specifically,AS, FXS and RS are caused by altered functional expres-sion of key synaptic proteins, including the E3 ubiquitinligase, UBE3a [115,116], fragile X mental retardationprotein (FMR1P, encoded by the gene FMR1) and methylCpG binding protein 2 (MeCP2), respectively. A mousemodel for AS that specifically lacksUbe3a on the maternalallele (i.e. Ube3am�/p+) was observed to have impairedsleep homeostasis and insomnia [117]. FMR1 loss in miceis associated with circadian dysfunction and perturbedrhythmic activity [118], and FMRP appears to be impor-tant for synaptic plasticity [119] and the sleep-dependentrenormalization of synapses [82]. Sleep disturbances havebeen reported in patients with these disorders, eventhough quantitative EEG analysis is still scarce. Mostproblems relate to insomnia: difficulty in initiating sleep,sleep fragmentation, or maintaining sleep with longersleep latency and less sleep efficiency [120,121]. Qualita-tive analyses of sleep in children diagnosed with ASD and/or developmental delays have shown that undifferentiatedsleep is increased, whereas NREM spindles, SWS andREM are decreased [122,123]. Optimizing sleep could bebeneficial for some of the most detrimental behavioralabnormalities associated with these conditions. According-ly, recent clinical data suggest a beneficial effect of mela-tonin supplementation on behavioral abnormalities inchildren with ASD [124]. Further studies will be necessaryto understand the relationship between sleep quality andsynaptic plasticity in ASD and other neurological disor-ders. It is hoped that studying sleep in the context of thesedisorders might not only improve treatment and the earlydiagnosis of such disorders, but also shed light on mecha-nisms and functions fundamental to sleep.
Concluding remarksAlthough many questions remain (Box 2), the scientificenigma as to why we sleep is beginning to be unraveled.In the brain, sleep is essential, and this need appears torequire a level of synaptic plasticity that is unavailableduring wake. This state of plasticity allows for homeostaticoptimization of neural networks as well as the replay-basedconsolidation of specific circuits. Indeed, sleep plasticityappears to be focused not on acquiring new information,but on prioritizing and compressing known information to

Box 2. Outstanding questions
� Is sleep-dependent synaptic plasticity in the mammalian brain
highly governed by the circadian clock, as has been observed in
Drosophila and zebrafish? Or is the mammalian cortex different in
terms of sleep plasticity, being more sleep dependent and less
clock dependent?
� Are there specific epochs of synaptic plasticity in the brain? Are
there quantitative differences between synaptic and structural
plasticity during sleep versus wake?
� Does sleep plasticity occur similarly throughout the brain? More
specifically, is there one cycle of synaptic strengthening and
elimination, or are there multiple rhythms spread across different
brain regions?
� Is sleep-dependent plasticity in the neocortex different from in
deeper brain regions? How is plasticity correlated with EEG
measurements, and is the type of synchrony in the cortex
measured via EEG a widespread phenomenon or specific to the
cortex?
� How is sleep plasticity behaviorally adaptive? For instance, does
sleep optimize function based on prior environmental experience?
Review Trends in Neurosciences September 2011, Vol. 34, No. 9
maintain optimal network function. Based on availabledata, we postulate that this optimization requires a stateof structural and molecular plasticity that would be detri-mental to sensory processing or long-term stability of mem-ory in the asynchronous and unpredictable neuralenvironment of wake. Thus, this optimization is facilitatedin sleep during periods of highly synchronous activity.
Sleep resembles critical plastic periods during develop-ment and is an essential, recurring state of the brain that isrequired to maintain an optimal set point of connectivitythat is sensitive to both environmental enrichment andgenetic background. So, it is no surprise that when theunderlying structure of the brain is perturbed by neuronaldegeneration, or as occurs during aberrant neuronal de-velopment, sleep dysfunction arises as an early indicationof such problems. Thus, sleep is universal because it is acritical plastic state that consolidates prior informationand prioritizes network activity so that the brain functionsefficiently in whatever new world we wake up in.
AcknowledgmentOur work is supported by the National Institutes of Health (NS062798,DK090065).
References1 Aserinsky, E. and Kleitman, N. (1953) Regularly occurring periods of
eye motility, and concomitant phenomena, during sleep. Science 118,273–274
2 Jouvet, M. et al. (1959) On a stage of rapid cerebral electrical activityin the course of physiological sleep. C. R. Seances Soc. Biol. Fil. 153,1024–1028 (in French)
3 Carskadon, M.A. and Dement, W.C. (2005) Normal human sleep: anoverview. In Principles and Practice of Sleep Medicine (Kryger, M.H.et al., eds), pp. 13–23, Saunders
4 Tobler, I. (1983) Effect of forced locomotion on the rest-activity cycle ofthe cockroach. Behav. Brain Res. 8, 351–360
5 Campbell, S.S. and Tobler, I. (1984) Animal sleep: a review of sleepduration across phylogeny. Neurosci. Biobehav. Rev. 8, 269–300
6 Tobler, I. and Borbely, A.A. (1985) Effect of rest deprivation on motoractivity of fish. J. Comp. Physiol. A 157, 817–822
7 Hartse, K.M. (2011) The phylogeny of sleep. Handb Clin. Neurol. 98,97–109
8 Cirelli, C. and Tononi, G. (2008) Is sleep essential? PLoS Biol. 6, e2169 Rechtschaffen, A. et al. (1983) Physiological correlates of prolonged
sleep deprivation in rats. Science 221, 182–184
10 Shaw, P.J. et al. (2002) Stress response genes protect against lethaleffects of sleep deprivation in Drosophila. Nature 417, 287–291
11 Fiorino, A.S. (1996) Sleep, genes and death: fatal familial insomnia.Brain Res. Brain Res. Rev. 22, 258–264
12 Lu, J. et al. (2006) A putative flip-flop switch for control of REM sleep.Nature 441, 589–594
13 Saper, C.B. et al. (2010) Sleep state switching. Neuron 68, 1023–
104214 Tononi, G. and Cirelli, C. (2006) Sleep function and synaptic
homeostasis. Sleep Med. Rev. 10, 49–6215 Frank, M.G. (2006) The mystery of sleep function: current
perspectives and future directions. Rev. Neurosci. 17, 375–39216 Mignot, E. (2008) Why we sleep: the temporal organization of
recovery. PLoS Biol. 6, e10617 Krueger, J.M. et al. (2008) Sleep as a fundamental property of
neuronal assemblies. Nat. Rev. Neurosci. 9, 910–91918 Walker, M.P. (2009) The role of sleep in cognition and emotion. Ann.
N. Y. Acad. Sci. 1156, 168–19719 Diekelmann, S. and Born, J. (2010) The memory function of sleep.
Nat. Rev. Neurosci. 11, 114–12620 Pilcher, J.J. and Huffcutt, A.I. (1996) Effects of sleep deprivation on
performance: a meta-analysis. Sleep 19, 318–32621 Stickgold, R. et al. (2000) Visual discrimination learning requires
sleep after training. Nat. Neurosci. 3, 1237–123822 Walker, M.P. et al. (2002) Practice with sleep makes perfect: sleep-
dependent motor skill learning. Neuron 35, 205–21123 Wagner, U. et al. (2004) Sleep inspires insight. Nature 427, 352–35524 Walker, M.P. and Stickgold, R. (2006) Sleep, memory, and plasticity.
Annu. Rev. Psychol. 57, 139–16625 Mander, B.A. et al. (2011) Wake deterioration and sleep restoration of
human learning. Curr. Biol. 21, R183–R18426 Harrison, Y. and Horne, J.A. (2000) Sleep loss and temporal memory.
Q. J. Exp. Psychol. A 53, 271–27927 Drummond, S.P. et al. (2000) Altered brain response to verbal
learning following sleep deprivation. Nature 403, 655–65728 Drummond, S.P. and Brown, G.G. (2001) The effects of total sleep
deprivation on cerebral responses to cognitive performance.Neuropsychopharmacology 25, S68–S73
29 Lahl, O. et al. (2008) An ultra short episode of sleep is sufficient topromote declarative memory performance. J. Sleep Res. 17, 3–10
30 Mednick, S. et al. (2003) Sleep-dependent learning: a nap is as good asa night. Nat. Neurosci. 6, 697–698
31 Tucker, M.A. et al. (2006) A daytime nap containing solely non-REMsleep enhances declarative but not procedural memory. Neurobiol.Learn Mem. 86, 241–247
32 Stickgold, R. and Walker, M.P. (2007) Sleep-dependent memoryconsolidation and reconsolidation. Sleep Med. 8, 331–343
33 Marshall, L. et al. (2006) Boosting slow oscillations during sleeppotentiates memory. Nature 444, 610–613
34 Nader, R. and Smith, C. (2003) A role for stage 2 sleep in memoryprocessing. In Sleep and Brain Plasticity (Maquet, P. et al., eds), pp.87–99, Oxford University Press
35 Giuditta, A. et al. (1995) The sequential hypothesis of the function ofsleep. Behav. Brain Res. 69, 157–166
36 Girardeau, G. and Zugaro, M. (2011) Hippocampal ripples andmemory consolidation. Curr. Opin. Neurobiol. 21, 452–459
37 O’Neill, J. et al. (2010) Play it again: reactivation of waking experienceand memory. Trends Neurosci. 33, 220–229
38 Wamsley, E.J. et al. (2010) Dreaming of a learning task is associatedwith enhanced sleep-dependent memory consolidation. Curr. Biol. 20,850–855
39 Dave, A.S. and Margoliash, D. (2000) Song replay during sleep andcomputational rules for sensorimotor vocal learning. Science 290,812–816
40 Margoliash, D. (2010) Sleep, learning, and birdsong. ILAR J. 51,378–386
41 Wilson, M.A. and Ji, D.Y. (2007) Coordinated memory replay in thevisual cortexandhippocampusduring sleep.Nat.Neurosci.10,100–107
42 Wilson, M.A. and Mcnaughton, B.L. (1994) Reactivation ofhippocampal ensemble memories during sleep. Science 265, 676–
67943 Peigneux, P. et al. (2004) Are spatial memories strengthened in the
human hippocampus during slow wave sleep? Neuron 44, 535–545
461

Review Trends in Neurosciences September 2011, Vol. 34, No. 9
44 Rasch, B. et al. (2007) Odor cues during slow-wave sleep promptdeclarative memory consolidation. Science 315, 1426–1429
45 Rudoy, J.D. et al. (2009) Strengthening individual memories byreactivating them during sleep. Science 326, 1079
46 Diekelmann, S. et al. (2011) Labile or stable: opposing consequencesformemory when reactivated during waking and sleep.Nat. Neurosci.14, 381–386
47 Peyrache, A. et al. (2009) Replay of rule-learning related neuralpatterns in the prefrontal cortex during sleep. Nat. Neurosci. 12,919–926
48 Girardeau, G. et al. (2009) Selective suppression of hippocampalripples impairs spatial memory. Nat. Neurosci. 12, 1222–1223
49 Foster, D.J. and Wilson, M.A. (2006) Reverse replay of behaviouralsequences in hippocampal place cells during the awake state. Nature440, 680–683
50 Carr, M.F. et al. (2011) Hippocampal replay in the awake state: apotential substrate for memory consolidation and retrieval. Nat.Neurosci. 14, 147–153
51 Frank, M.G. et al. (2001) Sleep enhances plasticity in the developingvisual cortex. Neuron 30, 275–287
52 Frank, M.G. et al. (2006) Blockade of postsynaptic activity in sleepinhibits developmental plasticity in visual cortex. Neuroreport 17,1459–1463
53 Aton, S.J. et al. (2009) Mechanisms of sleep-dependent consolidationof cortical plasticity. Neuron 61, 454–466
54 Tononi, G. and Cirelli, C. (2003) Sleep and synaptic homeostasis: ahypothesis. Brain Res. Bull. 62, 143–150
55 Esser, S.K. et al. (2007) Sleep homeostasis and corticalsynchronization: I Modeling the effects of synaptic strength onsleep slow waves. Sleep 30, 1617–1630
56 Sale, A. et al. (2009) Enrich the environment to empower the brain.Trends Neurosci. 32, 233–239
57 van Praag, H. et al. (2000) Neural consequences of environmentalenrichment. Nat. Rev. Neurosci. 1, 191–198
58 Zito, K. and Svoboda, K. (2002) Activity-dependent synaptogenesis inthe adult mammalian cortex. Neuron 35, 1015–1017
59 Vyazovskiy, V.V. et al. (2008) Molecular and electrophysiologicalevidence for net synaptic potentiation in wake and depression insleep. Nat. Neurosci. 11, 200–208
60 Silva, A.J. (2003)Molecular and cellular cognitive studies of the role ofsynaptic plasticity in memory. J. Neurobiol. 54, 224–237
61 Wallace, C.S. et al. (1995) Correspondence between sites of Ngfi-aInduction and sites of morphological plasticity following exposure toenvironmental complexity. Mol. Brain Res. 32, 211–220
62 Ying, S.W. et al. (2002) Brain-derived neurotrophic factor induceslong-term potentiation in intact adult hippocampus: requirement forERK activation coupled to CREB and upregulation of Arc synthesis. J.Neurosci. 22, 1532–1540
63 Rasch, B. et al. (2009) Impaired off-line consolidation of motormemories after combined blockade of cholinergic receptors duringREM sleep-rich sleep. Neuropsychopharmacology 34, 1843–1853
64 Sara, S.J. (2010) Reactivation, retrieval, replay and reconsolidation inand out of sleep: connecting the dots. Front. Behav. Neurosci 4, 185
65 Huber, R. et al. (2000) Topography of EEG dynamics after sleepdeprivation in mice. J. Neurophysiol. 84, 1888–1893
66 Ghilardi, M.F. et al. (2000) Patterns of regional brain activationassociated with different forms of motor learning. Brain Res. 871,127–145
67 Liu, Z.W. et al. (2010) Direct evidence for wake-related increases andsleep-related decreases in synaptic strength in rodent cortex. J.Neurosci. 30, 8671–8675
68 Lante, F. et al. (2011) Removal of synaptic Ca(2)+-permeable AMPAreceptors during sleep. J. Neurosci. 31, 3953–3961
69 O’Brien, R.J. et al. (1998) Activity-dependent modulation of synapticAMPA receptor accumulation. Neuron 21, 1067–1078
70 Turrigiano, G.G. et al. (1998) Activity-dependent scaling of quantalamplitude in neocortical neurons. Nature 391, 892–896
71 Wierenga, C.J. et al. (2005) Postsynaptic expression of homeostaticplasticity at neocortical synapses. J. Neurosci. 25, 2895–2905
72 Kilman, V. et al. (2002) Activity deprivation reduces miniature IPSCamplitude by decreasing the number of postsynaptic GABA(A)receptors clustered at neocortical synapses. J. Neurosci. 22, 1328–
1337
462
73 Stellwagen, D. and Malenka, R.C. (2006) Synaptic scaling mediatedby glial TNF-alpha. Nature 440, 1054–1059
74 Turrigiano, G.G. and Nelson, S.B. (2004) Homeostatic plasticity in thedeveloping nervous system. Nat. Rev. Neurosci. 5, 97–107
75 Turrigiano, G.G. (2008) The self-tuning neuron: synaptic scaling ofexcitatory synapses. Cell 135, 422–435
76 Dworak, M. et al. (2010) Sleep and brain energy levels: ATP changesduring sleep. J. Neurosci. 30, 9007–9016
77 Fernandez, M.P. et al. (2008) Circadian remodeling of neuronalcircuits involved in rhythmic behavior. PLoS Biol. 6, e69
78 Gorska-Andrzejak, J. et al. (2005) Structural daily rhythms in GFP-labelled neurons in the visual system of Drosophila melanogaster.Photochem. Photobiol. Sci. 4, 721–726
79 Mehnert, K.I. et al. (2007) Circadian changes in Drosophila motorterminals. Dev. Neurobiol. 67, 415–421
80 Weber, P. et al. (2009) Circadian control of dendrite morphology in thevisual system of Drosophila melanogaster. PLoS ONE 4, e4290
81 Gilestro, G.F. et al. (2009)Widespread changes in synaptic markers asa function of sleep and wakefulness in Drosophila. Science 324,109–112
82 Bushey, D. et al. (2011) Sleep and synaptic homeostasis: structuralevidence in Drosophila. Science 332, 1576–1581
83 Donlea, J.M. et al. (2009) Use-dependent plasticity in clock neuronsregulates sleep need in Drosophila. Science 324, 105–108
84 Appelbaum, L. et al. (2010) Circadian and homeostatic regulation ofstructural synaptic plasticity in hypocretin neurons.Neuron 68, 87–98
85 Faraco, J.H. et al. (2006) Regulation of hypocretin (orexin) expressionin embryonic zebrafish. J. Biol. Chem. 281, 29753–29761
86 Yokogawa, T. et al. (2007) Characterization of sleep in zebrafish andinsomnia in hypocretin receptor mutants. PLoS Biol. 5, 2379–2397
87 Appelbaum, L. et al. (2009) Sleep-wake regulation and hypocretin-melatonin interaction in zebrafish. Proc. Natl. Acad. Sci. U.S.A. 106,21942–21947
88 Micheva, K.D. and Smith, S.J. (2007) Array tomography: a new toolfor imaging the molecular architecture and ultrastructure of neuralcircuits. Neuron 55, 25–36
89 Graven, S. (2006) Sleep and brain development. Clin. Perinatol. 33,693–706 vii
90 Ednick, M. et al. (2009) A review of the effects of sleep during the firstyear of life on cognitive, psychomotor, and temperament development.Sleep 32, 1449–1458
91 Maquet, P. et al. (2003) Sleep and Brain Plasticity, Oxford UniversityPress
92 Anders, T.F. and Roffwarg, H.P. (1973) The effects of selectiveinterruption and deprivation of sleep in the human newborn. Dev.Psychobiol. 6, 77–89
93 Thomas, D.A. et al. (1996) The effect of sleep deprivation on sleepstates, breathing events, peripheral chemoresponsiveness andarousal propensity in healthy 3 month old infants. Eur. Respir. J.9, 932–938
94 Raizen, D.M. et al. (2008) Lethargus is aCaenorhabditis elegans sleep-like state. Nature 451, 569–572
95 Van Buskirk, C. and Sternberg, P.W. (2007) Epidermal growth factorsignaling induces behavioral quiescence in Caenorhabditis elegans.Nat. Neurosci. 10, 1300–1307
96 Huang, Y.Z. et al. (2000) Regulation of neuregulin signaling byPSD-95 interacting with ErbB4 at CNS synapses. Neuron 26, 443–
45597 Kwon, O.B. et al. (2005) Neuregulin-1 reverses long-term potentiation
at CA1 hippocampal synapses. J. Neurosci. 25, 9378–938398 White, J.G. et al. (1978) Connectivity changes in a class of
motoneurone during the development of a nematode. Nature 271,764–766
99 Hallam, S.J. and Jin, Y. (1998) lin-14 regulates the timing of synapticremodelling in Caenorhabditis elegans. Nature 395, 78–82
100 Jeon, M. et al. (1999) Similarity of the C. elegans developmentaltiming protein LIN-42 to circadian rhythm proteins. Science 286,1141–1146
101 Wilson, R.S. et al. (2011) Cognitive decline in prodromal Alzheimerdisease and mild cognitive impairment. Arch. Neurol. 68, 351–
356102 Osorio, R.S. et al. (2011) Greater risk of Alzheimer’s Disease in older
adults with insomnia. J. Am. Geriatr. Soc. 59, 559–562

Review Trends in Neurosciences September 2011, Vol. 34, No. 9
103 Petit, D. et al. (2004) Sleep and quantitativeEEG in neurodegenerativedisorders. J. Psychosom. Res. 56, 487–496
104 Dykierek, P. et al. (1998) The value of REM sleep parameters indifferentiating Alzheimer’s disease from old-age depression andnormal aging. J. Psychiatr. Res. 32, 1–9
105 Daulatzai, M.A. (2010) Early stages of pathogenesis in memoryimpairment during normal senescence and Alzheimer’s disease. J.Alzheimers Dis. 20, 355–367
106 Cooke, J.R. et al. (2009) Sustained use of CPAP slows deterioration ofcognition, sleep, and mood in patients with Alzheimer’s disease andobstructive sleep apnea: a preliminary study. J. Clin. Sleep Med. 5,305–309
107 Hoch, C.C. and Reynolds, C.F., 3rd (1989) Electroencephalographicsleep in late-life neuropsychiatric disorders. Int. Psychogeriatr. 1,51–62
108 Bird, T.D. (1993) Early-onset familial Alzheimer disease. InGeneReviews (Pagon, R.A. et al., eds), University of Washington
109 Yesavage, J.A. et al. (2004) Sleep/wake disruption in Alzheimer’sdisease: APOE status and longitudinal course. J. Geriatr.Psychiatry Neurol. 17, 20–24
110 Colas, D. et al. (2004) Sleep-wake architecture in mouse models forDown syndrome. Neurobiol. Dis. 16, 291–299
111 Colas, D. et al. (2008) Sleep and EEG features in genetic models ofDown syndrome. Neurobiol. Dis. 30, 1–7
112 Jyoti, A. et al. (2010) EEG, activity, and sleep architecture in atransgenic AbetaPPswe/PSEN1A246E Alzheimer’s disease mouse.J. Alzheimers Dis. 22, 873–887
113 Kang, J.E. et al. (2009) Amyloid-beta dynamics are regulated byorexin and the sleep-wake cycle. Science 326, 1005–1007
114 Cardinali, D.P. et al. (2010) Clinical aspects of melatonin interventionin Alzheimer’s disease progression.Curr. Neuropharmacol. 8, 218–227
115 Bourgeron, T. (2009) A synaptic trek to autism.Curr. Opin. Neurobiol.19, 231–234
116 Horsthemke, B. and Wagstaff, J. (2008) Mechanisms of imprintingof the Prader-Willi/Angelman region. Am. J. Med. Genet. A 146A,2041–2052
117 Colas, D. et al. (2005) Sleep disturbances in Ube3a maternal-deficientmice modeling Angelman syndrome. Neurobiol. Dis. 20, 471–478
118 Zhang, J. et al. (2008) Fragile X-related proteins regulate mammaliancircadian behavioral rhythms. Am. J. Hum. Genet. 83, 43–52
119 Soden, M.E. and Chen, L. (2010) Fragile X protein FMRP is requiredfor homeostatic plasticity and regulation of synaptic strength byretinoic acid. J. Neurosci. 30, 16910–16921
120 Malow, B.A. et al. (2006) Characterizing sleep in children with autismspectrum disorders: a multidimensional approach. Sleep 29, 1563–
1571121 Miano, S. and Ferri, R. (2010) Epidemiology and management of
insomnia in children with autistic spectrum disorders. Paediatr.Drugs 12, 75–84
122 Buckley, A.W. et al. (2010) Rapid eye movement sleep percentage inchildren with autism compared with children with developmental
delay and typical development. Arch. Pediatr. Adolesc. Med. 164,1032–1037
123 Limoges, E. et al. (2005) Atypical sleep architecture and the autismphenotype. Brain 128, 1049–1061
124 Doyen, C. et al. (2011) Melatonin in children with autistic spectrumdisorders: recent and practical data. Eur. Child Adolesc. Psychiatry20, 231–239
125 Loomis, A.L. et al. (1935) Potential rhythms of the cerebral cortexduring sleep. Science 81, 597–598
126 Derbyshire, A.J. et al. (1936) The effects of anesthetics on actionpotentials in the cerebral cortex of the cat. Am. J. Physiol. 116,577–596
127 Michel, F. et al. (1961) Etude polygraphique du sommeil chez le rat.C.R. Soc. Biol. 155, 2389–2392
128 Cirelli, C. and Tononi, G. (2000) Gene expression in the brain acrossthe sleep-waking cycle. Brain Res. 885, 303–321
129 Weiss, T. and Fifkova, E. (1964) Sleep cycles in mice. Physiol.Bohemoslov. 13, 242–245
130 Gao, X.B. et al. (2007) Prolonged wakefulness induces experience-dependent synaptic plasticity in mouse hypocretin/orexin neurons. J.Clin. Invest. 117, 4022–4033
131 Hendricks, J.C. et al. (2000) Rest in Drosophila is a sleep-like state.Neuron 25, 129–138
132 Zhdanova, I.V. et al. (2001) Melatonin promotes sleep-like state inzebrafish. Brain Res. 903, 263–268
133 Sternberg, P.W. and Van Buskirk, C. (2007) Epidermal growth factorsignaling induces behavioral quiescence in Caenorhabditis elegans.Nat. Neurosci. 10, 1300–1307
134 Gonzales, M.L. and LaSalle, J.M. (2010) The role of MeCP2 in braindevelopment and neurodevelopmental disorders. Curr. PsychiatryRep. 12, 127–134
135 Bertram, L. et al. (2010) The genetics of Alzheimer disease: back to thefuture. Neuron 68, 270–281
136 Yashiro, K. et al. (2009) Ube3a is required for experience-dependentmaturation of the neocortex. Nat. Neurosci. 12, 777–783
137 Micheva, K.D. et al. (2010) Single-synapse analysis of a diversesynapse population: proteomic imaging methods and markers.Neuron 68, 639–653
138 Betzig, E. et al. (2006) Imaging intracellular fluorescent proteins atnanometer resolution. Science 313, 1642–1645
139 Hell, S.W. and Wichmann, J. (1994) Breaking the diffractionresolution limit by stimulated emission: stimulated-emission-depletion fluorescence microscopy. Opt. Lett. 19, 780–782
140 Hess, S.T. et al. (2006) Ultra-high resolution imaging by fluorescencephotoactivation localization microscopy. Biophys. J. 91, 4258–4272
141 Huang, B. et al. (2008) Three-dimensional super-resolutionimaging by stochastic optical reconstruction microscopy. Science319, 810–813
142 Willig, K.I. et al. (2006) STEDmicroscopy reveals that synaptotagminremains clustered after synaptic vesicle exocytosis. Nature 440,935–939
463







![Review Article Is Sleep Essential for Neural Plasticity in ...downloads.hindawi.com/journals/np/2013/103949.pdf], and by an increase of synaptic density [ ]. e synaptic homeostasis](https://img.dokumen.tips/doc/110x75/5f7a9a297422022fa8445184/review-article-is-sleep-essential-for-neural-plasticity-in-and-by-an-increase.jpg)





















