Embed Size (px)
Citation preview

OFFICIAL ORGAN OF THE RADIATION RESEARCH SOCIETY
RADIATION RESEARCH
M A N A G i N G EDITOR: ODDVAR F. NYGAARD
Volume 52, 1972
Academic Press • New York and London

Copyright © , 1972, by A C A D E M I C P R E S S , INC.
A L L R I G H T S R E S E R V E D
N o p a r i of this publication may be reproduced or I r a n s m i t t e d i n any f o r m , or by any means, electronic or mechanical, i n c l u d i n g photocopy, recording, or any i n f o r m a l i o n slorage and r e t r i e v a l System, w i t h -out permission i n w r i t i n g f r o m the Copyright owner.
Made in the United States of America
( \N Bayerische
Staatsbibliothek München

RADIATION RESEARCH
OFFICIAL ORGAN OF THE RADIATION RESEARCH SOCIETY
BOARD OF EDITORS: Managing Editor: ODDVAR F . N Y G A A R D , Department of Radiology, Case Western Reserve University, Cleveland, Ohio 44106.
G. E. ADAMS, Mt. Vernon Hospital, England
M . J . BERGER, National Bureau of Standards
J. G. CARLSON, University of Tennessee
R. A . CONARD, Brookhaven National Laboratory
C. C. CONGDON, Oak Ridge National Laboratory
S. B. CURTIS, University of California E . R. E P P , Sloan-Kettering Institute
for Cancer Research T. M . FLIEDNER, Universität Ulm, Ger-
many .1. A . GHORMLEY, Oak Ridge National
Laboratory
M . L . G R I E M , University of Chicago R . H . H A Y N E S , York University,
Canada J . JAGGER, University of Texas R . F . K A L L M A N , Stanford University P. RIESZ, National Institutes of Health W. C. SNIPES, Pennsylvania State
University H . D. SUIT, University of Texas J . K . THOMAS, University of Notre
Dame J . F . THOMPSON, Argonne National
Laboratory L. J . TOLMACH, Washington University G . M . WOODWELL, Brookhaven Na
tional Laboratory
OFFICERS OF THE SOCIETY: President, A. D. CONGER, Temple University, Philadelphia, Pennsylvania.
Vice President (and President Elect), V. P.. BOND, Brookhaven National Laboratory, Upton, Long Island
Secretary-Treasurer, M A X R.. Z E L L E , Dept. Rad. and Rad. BioL Colorado State University, Ft. Collins, Colorado 80521
Managing Editor, ODDVAR F. NYGAARD, Case Western Reserve University, Cleveland, Ohio
Executive Secretary. RICHARD J. B U R K , JR. , 4211 39th Street, N . W., Washington, D. C. 20016
Assistant to Managing E d i t o r : DOROTHY D. SCHOTTELIUS, University of Iowa, Iowa City, Iowa
ANNUAL MEETINGS: 1973: April 29-May 3, St. Louis, Missouri 1974: July 13-20, V International Congress Radiation Research, Seattle,
Washington
VOLUME 52, 1972
Copyright © 1972, by Academic Press, Inc., New York, N . Y . 10003, U . S. A.
Second class postage paid at Baltimore, M d . 21202

Councilors Radiation Research Society 1971-1972
PHYSICS
R. J. Shalek, University of Texas S. B. Curtis, University of California
BlOLOGY
R. F. Kallman, Stanford University G. W. Casarett, University of Rochester
MEDICINE
H . Spencer, V. A. Hospital, Hines, Illinois C . C. Lushbaugh, Oak Ridge Associated Universities
CHEMISTRY
Jack Schubert, University of Pittsburgh W . M . Garrison, University of California
A T - L A R G E
E. L. Alpen, Battelle Memorial Institute G. F. Whitmore, Ontario Cancer Institute

RADIATION R E S E A R C H 52, (1972)
CONTENTS OF VOLUME 52 NüMBER 1, OCTOBER 1972
J A M E S C . P E R S O N . Isotope Effects on M o l e c u l a r I o n i z a t i o n Yields. A n Extension of the Platz m a n M o d e l 1
J . A . K N I G H T . Radiolysis of Nilrobenzene 17 J . M . H E L L E R , J R . , R . D . B I R K H O F F , M . W . W I L L I A M S , A N D L . R . P A I N T E R . Optical
Properties of L i q u i d Glycerol i n the V a c u u m U l t r a v i o l e t 25 ß+, E C
R I C H A R D M . L A M B R E C H T A N D A L F R E D P . W O L F . The mTe(*He, 3n)l23Xe ' , 1 2 3 J Z . l tir,
Generator 32 L . W . S I E C K A N D P . A U S L O O S . Reactions of the Acetone C a t i o n . I m p l i c a t i o n s for the Radi-
olysis of Acetone 47 K . T . W H E E L E R A N D J . T . L E T T . F o r m a l i o n and R e j o i n i n g of D N A Strand Breaks i n
I r r a d i a t e d Neurons: I n V i v o 59 A . V A N P E L T A N D C . C . C O N G D O N . R a d i a t i o n L e u k e m i a i n G u i n e a Pigs 68 M A R I O N J . M E E K S , W A Y N E V . K E S S L E R , A N D J A M E S N . A R V E S E N . The Effect of Whole-
Body I r r a d i a t i o n on the M e l a b o l i s m of 5 - F l u o r o u r a c i l - 2 - u C i n the R a t 82 E . J . H A L L , W . G R O S S , R . F . D V O R A K , A . M . K E L L E R E R , A N D H . H . R O S S I . Survival Curves
and Age Response Functions for Chinese Hamster Cells Exposed lo X Rays or H i g h L E T A l p h a - P a r l i c l e s 88
C H R I S T O P H E R D . T O W N , K E N D R I C C . S M I T H , A N D H E N R Y S. K A P L A N . Influence of Ultra-
fast Repair Processes (Independent of D N A Polymerase I ) on the Y i e l d of D N A Single-Strand Breaks i n Escherichia coli K - 1 2 X - I r r a d i a t e d i n the Presence orAbsence of Oxygen 99
E . A . P U R O A N D G . M . C L A R K . T h e Effect of Exposure Rate on A n i m a l Lethality and Spleen Colony C e l l S u r v i v a l 115
S E I - I C H I N A K A M U R A . Effect of C a r b o n Particles on H e m a l o l o g i c a l Recovery i n I r r a d i a t e d M i c e 130
J A M E S P . O K U N E W I C K , D O R I S F U L T O N , A R N O L D M . M A R K O E , A N D E L E A N O R E L . P H I L L I P S .
I n t e r r e l a t i o n s h i p of E r y t h r o p o i e l i c Recovery, M a r r o w Recovery, C o l o n y - F o r m i n g U n i t s , and Erythropoiesis-Stimulaling Factors After Sublethal X - I r r a d i a t i o n 138
J . C . H A M P T O N A N D B . R O S A R I O . T h e F i n e S l r u c t u r e of I n l e s t i n e and Liver i n I r r a d i a t e d M i c e Düring F a t Absorption 152
C U R T I S P . S I G D E S T A D , R A L P H M . S C O T T , R O N A L D F . H A G E M A N N , A N D E D G A R B . D A R D E N ,
J R . I n t e s t i n a l Crypt Survival: T h e Effect of Cobalt-60, 2 5 0 kVcp X - r a y s and Fission Neutrons 168
S. P I I Y L L I S S T E A R N E R A N D E M I L Y J . B . C H R I S T I A N . L a t e Effects of I o n i z i n g Radiations i n the C h i c k e n : Survival, Body Weight, and Pathology 179
A . S A S S E N , F . K E N N E S , A . R E U T E R , A N D M . T E L I A . Catabolism of Serum A l b u m i n i n X - I r radiated M i c e 197
W I L L I A M F . B R A N D O M , G E N O S A C C O M A N N O , V I C T O R E . A R C H E R , P H I L I P G . A R C H E R , A N D
M A R I L Y N E . C O O R S . Chromosome Aberrations i n U r a n i u m M i n e r s Occupationally Exposed to i22Radon 204
Obüuary for Joseph Weiss 216
Copyright © 1972 by Academic Press, Inc. All rights of reproduction in any form reserved.

N U M B E R 2, N O V E M B E R 1972
E . M . F I E L D E N , S. C . L I L L I C R A P , A N D G . J . S M I T H . H i g h Energy E l e c t r o n I n d u c e d Emission i n Low Tempera i u r e Glasses C o n l a i n i n g Adenine 219
J . E . T U R N E R , J . D U T R A N N O I S , H . A . W R I G H T , R. N . H A M M , J . B A A R L I , A . H . S U L L I V A N , M .
J . B E R G E R , A N D S. M . S E L T Z E R . T h e C o m p u l a l i o n of P i o n Depth-Dose Curves i n Water and Comparison with E x p e r i m e n t 229
T . W . A R M S T R O N G A N D K . C . C H A N D L E R . M o n t e C a r l o C a l c u l a l i o n s of the Dose I n d u c e d by Charged Pions and Comparison with E x p e r i m e n t 247
S A M U E L S. K U R O H A R A A N D G E O R G E W . C A S A R E T T . Effects of Single T h o r a c i c X - R a y E x posure i n Rats 263
P . K . H O R A N , K A T H Y H I R D , A N D E . C . P O L L A R D . A S t r a i n of Escherichia C o l i wüh M i n i mum P o s l i r r a d i a t i o n D e g r a d a t i o n Proper lies 291
A . F . M C F E E , M . W . B A N N E R , A N D M A R Y N . S H E R R I L L . Behavior of Swine Leukocyles in
C u l t u r e öfter I n V i v o or I n V i t r o G a m m a I r r a d i a t i o n 301 J . H . H E N D R Y ' A N D L . G . L A J T H A . T h e Responses of Hemopoietic C o l o n y - F o r m i n g U n i t s lo
Repeated Doses of X - R a y s 309 J . C . H A M P T O N . Acute R a d i a l i o n Effects i n Kidney 316 E D W A R D R. E P P , H E R B E R T W E I S S , B O Z I D A R D J O R D J E V I C , A N D A N N S A N T O M A S S O . The
Radiosensitivity of C u l t u r e d M a m m a l i a n Cells Exposed to Single H i g h Intensity Pxdses of Electrons i n V a r i o u s Concentrations of Oxygen 324
L . F . W I N A N S , W . C . D E W E Y , A N D C . M . D E T T O R . Repair of Sublethal and PotentiaUy L e l h a l X - R a y D a m a g e i n Synchronous Chinese Hamster Cells 333
C . M . D E T T O R , W . C . D E W E Y , L . F . W I N A N S , A N D J . S. N O E L . Enhancement of X-Ray
D a m a g e i n Synchronous Chinese Hamster Cells by H y p e r t o n i e Treatments 352 W . C . D E W E Y , J . S. N O E L , A N D C . M . D E T T O R . Changes i n Radiosensitivity and Disper
sion of C h r o m a t i n Düring the C e l l Cycle of Synchronous Chinese Hamster Cells 373 A G N A R N I L S S O N , L A S Z L Ö R ß v E s z , A N D K A R L H U G O E R I K S S O N . Anligenicity of Radio-
slrontium-Induced Osleosarcomas 395 W . H O W A R D C Y R A N D E R N E S T C . P O L L A R D . I o n i z i n g R a d i a t i o n - I n i t i a t e d D e g r a d a t i o n of
D N A i n G e r m i n a l i n g Spores 409 Obituary for W r i g h t H . L a n g h a m 419 O b i l u a r y for D r . Viimos Värteresz 422 B O O K R E V I E W 424
A N N O U N C E M E N T S 426
E R R A T U M 427
N U M B E R 3, D E C E M B E R 1972
H A R V E Y E I S E N , M A R V I N R O S E N S T E I N , A N D J O S E P H S I L V E R M A N . Electron Depth-Dose Dis
t r i b u t i o n Measurements i n Two-Layer Slab Absorbers 429 R A N D A L L S. C A S W E L L A N D J . J O S E P H C O Y N E . Inleraclion of Neutrons and Sccondary
Charged Particles with Tissue: Secondary P a r t i c l e Spectra 448 P . N E T A . E l e c t r o n Spin Resonance Study of Radicals Produced by O n e - E l e c l r o n Reductior.
of P y r i d i n es 471 A . L . B R O O K S , R. O . M C C L E L L A N A N D S. A . B E N J A M I N . The Effects of ^ C - l u P r on th<
Metaphase Chromosomes of the Chinese Hamster Liver Cells in Vivo 481 J U L I A G I D Ä L I , I . F E H Ä R , A N D V . V Ä R T E R E S Z . Differences in the Growth and Differentiatior,
of the C F U F r a c i i o n Surviving Sublethal I r r a d i a t i o n 499 C . K . Y u A N D W . K . S I N C L A I R . Polyploidy I n d u c e d by X - R a y s Düring the C e l l Cycle of
Chinese Hamster Cells in Vitro 509 J A C K A . W I X S T E A D A N D S H I R L E Y A . M O S S . G a m m a - I r r a d i a l e d F l a v i n Adenine D i n u c l e o l i d e '
A. D - A m i n o Acid Oxidase I n h i b i t o r 520

J O S E P H L . B E L S K Y , K A Z U O O K O C H I , T O R A N O S U K E I S H I M A R U , A N D H O W A R D B . H A M I L T O N .
Hepatitis Associated Antigen and Antibody i n A - B o m b Survivors and Nonexposed Sub-jects i n H i r o s h i m a and Nagasaki 528
J O H N F . M C G R K G O R A N D H O W A R D B . N E W C O M B E . Decreased Risk of Embryo M o r t a l i t y F o l -lowing Low Doses of R a d i a t i o n to T r o u t Sperm 536
C . G . M C I I A L E A N D G . N . C A T R A V A S . T h e Response of R a t Liver Enzymes Involved i n F a t l y Acid Synlhesis to I n c r e a s i n g Doses of I o n i z i n g R a d i a t i o n in Vivo 545
D A V I D E . H A R R I S O N . L i f e s p a r i n g Ability ( i n Lethally I r r a d i a t e d M i c e ) of W / W Mouse M a r r o w with N o Macroscopic Colonies 553
J . H . M Y E R S , L . H . B L A C K W E L L , A N D R . R . O V E R M A N . E a r l y F u n c t i o n a l Hemodynamic I m -pairment i n Baboons After 1 0 0 0 R or Less of G a m m a R a d i a t i o n as Revealed by H e m o r -r h a g i c Slress 564
J A N A D A M N O R R I S , M . A . R O B K I N , T . H . S H E P A R D , A N D T . T A N I M U R A . Prompt Effects of
R a d i a t i o n on the H e a r t Rate on a M a m m a l i a n Embryo 579 L . K . M E E , S. J . A D E L S T E I N A N D G . S T E I N . I n a c t i v a t i o n of Ribonuklease by the P r i m a r y
Aqueous Radicals 588 F . A . H O D G E A N D D . S. N A C H T W E Y . X - R a y - I n d u c e d D e l a y of C e l l D i v i s i o n i n Sijnchronized
T e t r a h y m e n a pyriformis 603 P E G G Y L . O L I V E , W . R O D G E R I N C H , A N D R O B E R T M . S U T H E R L A N D . The Effect of Triace-
t o n e a m i n e - N - O x y l on Oxygenation and Radiocurability of a Mouse M a m m a r y C a r c i n o m a 618 P . H . M . L O H M A N , D . B O O T S M A , A N D A . H . H E Y . T h e I n f l u e n c e of 5-Bromodeoxyuridine on
the I n d u c t i o n of Breaks i n the Deoxyribonucleic Acid of C u l t i v a l e d H u m a n Cells by X - I r r a d i a t i o n and U l t r a v i o l e t L i g h t . 627
Obituary for H u g o F r i c k e 642 A U T H O R I N D E X 647
S U B J E C T I N D E X 649

RADIATION RESEARCH
OFRCIAL ORGAN OF THE RADIATION RESEARCH SOCIETY
BOARD OF EDITORS
M a n a g i n g Editor: O D D V A R F . N Y G A A R D , Department of Radiology, Case Western Reserve University, Cleveland, Ohio 44106
G . E . A D A M S , M t . Vernon Hospital, England
M . J . B E R G E R , National Bureau of Standards
J . G . C A R L S O N , University of Tennessee
R. A . C O N A R D , Brookhaven National Laboratory
C . C . C O N G D O N , Oak Ridge National Laboratory
S. B . C U R T I S , University of California
E . R. E P P , Sloan-Kettering Institute for Cancer Research
T . M . F L I E D N E R , U n i v e r s i t ä t U l m , Germany
J . A . G H O R M L E Y , Oak Ridge National Laboratory
M . L . G R I E M , University of Chicago
R. H . H A Y N E S , York University, Canada
J . J A G G E R , University of Texes
R. F . K A L L M A N , Stanford University
P. R I E S Z , National Institutes of Health
W. C . SNIPES, Pennsylvania State U n i versity
II. D . SUIT, Harvard University
J . K . T H O M A S , University of Notre Dame
J . F . T H O M S O N , Argonne National L a b oratory
L . J . T O L M A C H , Washington University
G . M . W O O D W E L L , Brookhaven National Laboratory
OFFICERS OF THE SOCIETY
President: A . D . C O N G E R , Temple University, Philadelphia, Pennsylvania
V i c e President (and President Elect): V . P. B O N D , Brookhaven National Laboratory, Upton, Long Island, New York
Secretary-Treasurer: M A X R. Z E L L E , Dept. Rad. and Rad. Biol. , Colorado State University, F t . Collins, Colorado 80521
M a n a g i n g Editor: O D D V A R F . N Y G A A R D , Case Western Reserve University, Cleveland, Ohio
Executive Secrelary: R I C H A R D J . B U R K , J R . , 4211 39th Street, N . W . , Washington, D . C . 20016
Assistant to M a n a g i n g Editor: D O R O T H Y D . S C H O T T E L I U S , University of Iowa, Iowa City , Iowa
ANNUAL MEETINGS 1973: Apri l 29-May 3, St. Louis, Missouri
1974: July 13-20, Fifth International Congress of Radiation Research, Seattle, Washington
VOLUME 52, NUMBER 1, OCTOBER 1972
Copyright © 1972, by Academic Press, Inc., New York, N . Y . 10003, U . S. A.
Second class postage paid at Baltimore, M d . 21202

Radiation Research Volume 52, No. 1, October 1972
CONTENTS
Jarnos C. Porson. Isotope Effects on Molecular I o n i z a t i o n Yields. A n E x t e n s i o n of the P l a t z m a n Model 1
J. A. Knight. R a d i o l y s i s of Nitrobenzene 17 J. M . Hollor, Jr., R. D. Birkhoff, M . W. Williams, and L. R. Paintcr
Optical Proper lies of L i q u i d Glycerol i n the V a c u u m Ultraviolet 25 Richard M . Lambrecht and Alfrod P. Wolf. The m T e ( 4 H e , Sn)mXe
ß+ VC \ ! t
m I Generator 32 2.1 Irr, L. W. Siock and P. Ausloos. R e a c t i o n s of the Acetone C a t i o n . I m -
p l i c a t i o n s for the R a d i o l y s i s of Acetone 47 K . T. Whoolor and J. T. Lett. F o r m a t i o n and Rejoining of D N A
S t r a n d B r e a k s i n Irradiated N e u r o n s : I n Vivo 59 A. Van Polt and C. C. Congdon. R a d i a t i o n L e u k e m i a i n Guinea P i g s 68 Marion J. Mceks, Wayno V. Kosslor, and James X . Arvesen. The Ef
fect of Whole-Body I r r a d i a t i o n on the Metabolism of 5-Fluorouracil-2 - l 4 C i n the Rat 82
E. J. Hall, W. Gross, R, F. Dvorak, A. M . Kellerer, and H . H . Rossi. S u r v i v a l Curves and Age Response F u n c t i o n s for Chinese Hamster Cells Exposed to X Rays or H i g h L E T A l p h a - P a r t i c l e s 88
Christophor D. Town, Kondric C. Smith, and Henry S. Kaplan. I n -fluence of Ultrafast R e p a i r Processes (Independent of D N A Polymerase I ) on the Yield of D N A S i n g l e - S t r a n d B r e a k s i n Escherichia i coli K - 1 2 X - I r r a d i a t e d i n the Presence or Absence of Oxygen 99
E. A. Puro and G. M . Clark. The Effect of Exposure Rate on A n i m a l l L e t h a l i t y and Spleen Colony Cell S u r v i v a l ] ] 1 lo
Sci-Ichi Nakamura. Effect of Carbon P a r t i c l e s on Hematological Recovery i n I r r a d i a t e d Mice ] 3 13C
James P. Okunewick, Doris Eulton, Arnold M . Markoe, and Eleanore L. Phillips. I n t e r r e l a t i o n s h i p of Erythropoietic Recovery, Marrow Recovery, Colony-Forming U n i t s , and E r y t h r o p o i e s i s - S t i m u l a t i n g / Factors After S u b l e t h a l X - I r r a d i a t i o n ] 13£
J. C. Hampton and B. Rosario. The F i n e S t r u c t u r e of I n t e s t i n e and ' l L i v e r i n I r r a d i a t e d Mice Düring Fat A b s o r p t i o n 15.
Curtis P. Sigdestad, Ralph M . Scott, Ronald F. Hagemann, and 1 Edgar B. Darden, Jr. I n t e s t i n a l Crypt S u r v i v a l : The Effect of Cobalt-60, 2 5 0 kVcp X - r a y s and F i s s i o n N e u t r o n s
S. Phyllis Stearner and Emily J. B . Christian. L a i e Effects of I o n i z i n g R a d i a t i o n s i n the C h i c k e n : S u r v i v a l , Body Weight, and Pathology
A. Sassen, F. Kermes, A . Reuter, and M . Telia. Catabolism of Serum A l b u m i n i n X - I r r a d i a t e d Mice
William F. Brandom, Gcno Saccomanno, Victor E. Archer, Philip G.. . ... Archer, and Marilyn E . Coors. Chromosome A b e r r a t i o n s i n >, n U r a n i u m M i n e r s Occupationally Exposed to 2 2 2 R a d o n : 20
Obituary f or J oseph We iss 21
• / 16!
' < l ' 17(
19'
Published monthly at M t . Royal & Guilford Aves., Baltimore, M d . 22121 22120' by Academic Press, Inc., 111 Fifth Avenue, New York, N . Y . 10003.
In i972, Volumes 49-52 will be published. Price of each volume $20.00. All correspondence and subscription Orders should be addressed to the ofhce oof of oof tr
Publishers at 111 Fifth Avenue, New York, N . Y . 10003. Send notices of change of address to the office of the Publishers at least 4 weeekeleeeks i
advance. Please include both old and new addresses. © 1972 by Academic Press, Inc. 1973: Volumes 53-56. Price per volume: $20.00 U .S .A . ; $22.50 outside U .S .A .

Radiation Research
Volume 52, No. 1, October 1972 Copyright © 1972 by Academic Press, Inc.
Al l rights reserved
No part of this publication may be reproduced or transmitted in any form, or by any means, electronic or mechanical, including photocopy, re-cording, or any information storage and retrieval System, without permission in writing from the Copyright owner.

R A D I A T I O N R E S E A R C H 52, 88-98 (1972)
Survival Curves and Age Response Functions for Chinese Hamster Cells Exposed to X-Rays or High LET Alpha-Particles1
E. J. H A L L , W. GROSS, R. F. DVORAK, A. M . K E L L E R E R , AND H . H . ROSSI
Radiological Research Laboratory, Department of Radiology, College of Physicians and Surgeom of C o l u m b i a University, New York, New Y o r k 1 0 0 3 2
H A L L , E. J., GROSS, W., DVORAK, R. F., K E L L E R E R , A. M . , AND ROSSI, H . H . Survival Curves and Age Response Functions for Chinese Hamster Cells Exposed to X-rays or High L E T Alpha Particles. R a d i a l . Res. 52, 8S-98 (1972).
Chinese hamster cells were synchronized with hydroxyurea, and ex-posed to either 800 rads of x-rays, or 250 rads of alpha particles at different points of the cell cycle. The pattern of response to x-rays is similar to that previously reported, namely, an increase in radioresistance to a maximum in late S, followed by a sensitive period in the late G 2 and M phases of the cycle. The pattern of response to alpha particles is quali-tatively very similar; the Variation between most sensitive and most re-sistant phases of the cell cycle is less than in the case of x-rays, but is still appreciable. The alpha particles have a L E T of approximately 90 keV/jum, and it was thought possiblc that the age response funetion would be flat for such densely ionizing radiation; however, this was not found to be the case.
I N T R O D U C T I O N
It lias been known for some years that survival curves for mammalian cells ex-posed to high L E T radiation, approximatc exponential funetions of dose (1). This finding contrasts sharply to the Situation for low L E T radiations for which survival curves can have a large initial Shoulder. For many cell lines, and in particular for the Chinese hamster cell which has been studied in great detail by Sinclair and ofchers (2), the sensitivity to x-rays varies as a funetion of age in the cell cycle, and much
1 Based on work performed under Contract AT-(30-1)-2740 for the United States Ar.omic Energy Commission.
88
Copyright © 1972 by Academic Press, Inc. All rights of reproduetion in any form reserved.

A L P H A P A R T I C L E S A N D T H E C E L L C Y C L E 89
of this Variation is duc to a changc in the Shoulder of the survival curve. Conse-quently it might be expected that the response to high L E T radiations might show significantly less change through the cell cycle than the response for low L E T radiation, sirtce there is little or no Shoulder in the first place. Sinclair (3) exposed syn-chronized Chinese hamster cells to fission neutrons from the Janus reactor, and indeed found that the response as a funetion of cell age varied less than for x-rays. Howcver, there was still an appreciable Variation through the cell cycle. Hall (4) condueted similar experiments with hamster cells using 14-MeV neutrons and also found that the pattern of response through the cycle was preserved, though it was less pronounced than in the case of x-rays. The neutrons used in both studies re-ferred to above can be considered to be of intermediate L E T .
Recently Bird and Burki (o) exposed synchronized Chinese hamster cells to three different beams of high-LET charged particles; carbon ions (190 k e V / p m ) , neon (650 k e V / p m ) , and argon ions (2000 k c V / p m ) . In all cases, the radiation response did not vary with cell age in the mitotic cycle. Elkind (6) has recently discussed unpublished data of Scarsgard in which synchronized hamster cells were irradiated with boron ions at a L E T of 127 keV/pm. These data indicate that an age Variation still exists, although it is much smaller than in the case of x-rays. The present report describes experiments in which synchronized hamster cells were exposed to mono-energetic alpha particles with a L E T of about 90 keV/jum; a quantitative comparison with the previous experimental results for various radiation qualities will be given.
The question of age response funetions with high L E T radiations is of interest in two connections. First, it has important implications in an understanding of the mechanisms of cell killing by ionizing radiations. Second, it may have some rele-vance to recent trends in radiotherapy, since high L E T modalities are being actively considered as an alternative or Supplement to x- and 7-rays. The bulk of clinical experience has been accumulated with sparsely ionizing radiations, whcre the response to a dose of radiation varies widely with the cell age. The effect of a frac-tionated regimen almost certainly involves partial synchronization of the exposed cell populations, although the Situation is very complicated and poorly understood at the present time. lf there is no dependence of sensitivity on age for high L E T radiations, this would be yet another altered variable when those radiations are used instead of x- or 7-rays, and possible changes in fractionation patterns may be de-sired.
M A T E R I A L S A N D M E T H O D S
C u l t u r e of the Cells V79 Chinese hamster cells were used, the original culture was supplied by Dr. M .
M . Elkind of the Brookhaven National Laboratory. The cells were grown in nu-trient medium F10 (7) supplemented with 10% fetal calf serum. The day before an

90 H A L L E T A L .
irradiation experiment, cells from a partly confluent actively growing stock' culture were prepared into a single cell Suspension by convontional methods (£), and plated out into Falcon plastic flasks for irradiation, or into specially eonstructed Containers with a thin Mylar window for exposure to alpha particles. These special Containers consist of a cylinder of stainless stcel, with entry and exit ports for medium and gas; thin sheets of Mylar (0.82 mg/cm2) were stretched over the end of this cylinder and held in tension by means of stainless-steel end-plates.
After overnight attachment for about 17 h, asynchronous populations of cells were irradiated with 210-kV x-rays or with alpha particles. At the time of irradiation, parallel cultures were fixed and stained and the mean number of cells per micro-colony (the multiplicity) was determined under a low-power microscope.
To obtain a synchronized population cells were allowed to attach overnight, after which hydroxyurea (H.U.) was added to the growth medium to a final con-centration of 1.25 mil/ and allowed to remain in contact with the cells for 3j^ h. The medium containing the H.U. was then removed, the cells were washed, and fresh growth medium added. Hydroxyurea blocks D N A synthesis while allowing protein and R N A synthesis to continue in a normal fashion. In the concentration used, it is toxic to hamster cells synthesizing D N A (S cells) while allowing free passage of cells in the remaining phases of the cycle to the Gi/S interface where they are blocked. When the drug is removed, a wave of cells passes into S and con-stitutcs a synchronized population ( 9 - 1 1 ) . The cell cultures were then returned to the 37° C incubator and after various time intervals, as the cells movedthroughtheir mitotic cycle, samples were exposed to x-rays or alpha particles.
Methods of I r r a d i a t i o n The source of x-rays was a 210-kV therapy machine, which at a treatment dis-
tance of 50 cm, produced a dose-rate of 70 rads/min. To expose cells to alpha particles of reasonably uniform LET, a special source was prepared which is illustrated in Fig. 1. The active deposit from 2 2 0 Em is plated on a pin in a device described in a previous report {12). This mixture of nuclides which has a half-life of 10.0 h, emits alpha particles at two energies, 8.78 and 6.07 AleV, but the patli from the source to the cells is designed to stop completely alpha particles of the lower energy. This path (see Fig. 1) is 11.6 cm overall, a distance required in order to achieve acccpta-ble uniformity of dose to the cells which are attached in a monolayer to a thin disc of Mylar 4.1 cm in diameter. This path consists of four parts. The first, insidc the body of the irradiator, is through helium at atmospheric pressure and has a thick-ness of about 1.7 mg/cm2. Next comes a Mylar window of 1.65 mg/cm2, then 0.69 cm of air at atmospheric pressure (0.73 mg/cm2), and finally 0.82 mg/cm2 of Mylar, which constitutes the surface to which the cells are attached. The material between source and cells is equivalent to 4.5 mg/cm2 of tissue. This is sufficient to absorb completely the 6.07-MeV alpha particlc the ränge of which is 4.4 mg/cm2.

A L P H A P A R T I C L E S A N D T H E C E L L C Y C L E 91
" H E L I U M AT A T M O S P H E R I C P R E S S U R E "
- S O U R C E S U R F A C E
- S O U R C E HOLDER
S C A L E (cm)
F I G . 1. Arrangement for irradiating cells with alpha particles. The radioactive isotope ( 2 2 0 Em) is plated on the conical pin. This nuclide emits alpha particles at two energies—8.78 and 6.07 MeV. The cells grow attached to a Mylar disc. The path betweeu the source and the cells consists of helium, two thicknesses of Mylar, and a small air gap. This pathlength is sufficient to filter out the lower energy alpha particles before they reach the cells.
The spectrum of alpha particles which reach the position occupied Iry the mam-malian cells is shown in Fig. 2. This was obtained with a lithium-drifted Silicon solid-state detector and multichannel analyzer. It is of interest to note that, while there is a small spread due to straggling, the beam is still very nearly monoenergetic. In addition, the remaining energy (5.2 MeV) is sufficient to permit the alpha particles to traverse the thickness of the cells, which is not more than 10 microns. Over this distance the L E T increases from 96 to 117 KoV//xm. This insures a reasonably uniform dose throughout the cells.
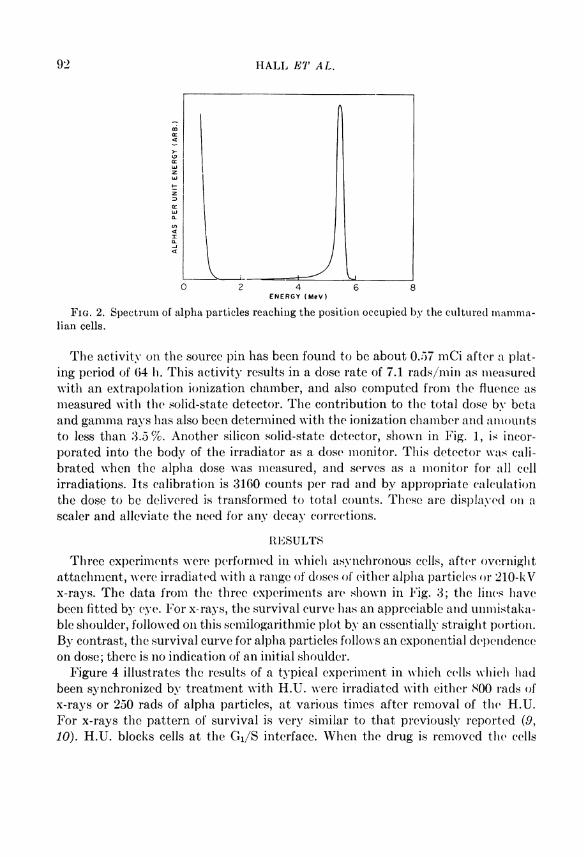
92 H A L L E T A L .
\ J\ \ 0 2 4 6 8
E N E R G Y ( M e V )
F I G . 2. Spectrum of alpha particles reaching the position occupied by the cultured mamma-lian cells.
The activity on the source pin has been found to be about 0.57 mCi after a plat-ing period of (54 h. This activity results in a dose rate of 7.1 rads/min as measured with an extrapolation ionization Chamber, and also computed from the fluence as measured with the solid-state detector. The contribution to the total dose by bcta and gamma rays has also been determined with the ionization Chamber and amouiits to less than 3.5%. Another Silicon solid-state detector, shown in Fig. 1, is incor-porated into the body of the irradiator as a dose monitor. This detector was cali-brated whcn the alpha dose was measured, and serves as a monitor for all cell irradiations. Its calibration is 3160 counts per rad and by appropriate calculation the dose to be delivered is transformed to total counts. These are displayed on a scaler and alleviate the need for any decay corrections.
R E S U L T S
Three experiments were performed in which asynchronous cells, after overnight attachment, were irradiated with a ränge of doses of either alpha particles or 210-kV x-rays. The data from the three experiments are shown in Fig. 3; the lines have been fitted by eye. For x-rays, the survival curve has an appreciable and unmistaka-ble Shoulder, followed on this semilogarithmic plot by an esscntially straight portion. By contrast, the survival curve for alpha particles follows an exponential dependence on dose; there is no indication of an initial Shoulder.
Figure 4 illustrates the results of a typical experiment in which cells which had been synchronized by trcatment with H.U. were irradiated with either 800 rads of x-rays or 250 rads of alpha particles, at various times after removal of the H.U. For x-rays the pattern of survival is very similar to that previously reported (9, 10). H.U. blocks cells at the Gi/S interface. When the drug is removed the cells
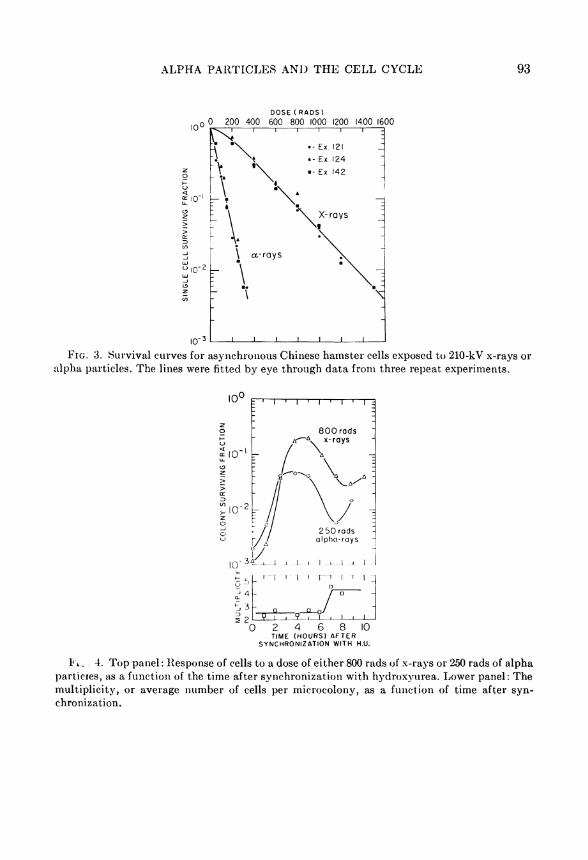
A L P H A P A R T I C L E S A N D T H E C E L L C Y C L E 93
D O S E ( R A D S )
0 0 200 400 600 800 1000 1200 1400 1600
F I G . 3. Survival curves for asynchronous Chinese hamster cells exposed to 210-kV x-rays or alpha particles. The lines were fitted by eye through data from three repeat experiments.
0 2 4 6 8 10 TIME ( H O U R S ) A F T E R
S Y N C H R O N I Z A T l O N WITH H.U.
F i - 4. Top panel: Response of cells to a dose of either 800 rads of x-rays or 250 rads of alpha particles, as a funetion of the time after synchronization with hydroxyurea. Lower panel: The multiplicity, or average number of cells per microcolony, as a funetion of time after synchronization.

94 H A L L E T A L .
move into S and become progressively more resistant; maximum resistance is ob-servod in late S, this is followed by an increase in sensitivity as the cells progress through G 2 into mitosis. For alpha particles the change of radiosensitivity as the cells move through the cell cycle is qualitatively similar to that for x-rays, but the extent of the Variation is less.
C O M P A R I S O N W I T H P R E V I O U S E X P E R I M E N T A L F I N D I N G S
A comparison of experimental data on the Variation of sensitivity throughout the cell cycle has been given by Bird and Burki (5), who plotted "survival Variation in life cycle compared with x-rays" as a funetion of L E T . Although no explicit exposi-tion of the meaning of this quantity is given we infer from the discussion that this Parameter is defined as
, = R ( D ) M D ) R X ( D X ) / S X ( D X ) '
where R ( D ) / S ( D ) is the ratio of the surviving fractions of the most resistant to the most sensitive subpopulations at dose D of the test radiation and R X ( D X ) / S X ( D X ) is the corresponding ratio for a dose D x of x-rays. In particular, Bird and Burki chose doses of the two types of radiation to produce equal survival level for the most sensitive cells so that S ( D ) = S X ( D X ) . Then one has
, = R ( D ) R X ( D X ) '
The disadvantage of this parameter is that it is dose dependent even for high L E T where the survival curves are approximately exponential. It is, therefore, practica 1 to choose a modified parameter which will be designated by V:
In R ( D ) In S ( D ) '
If the survival curves for the sensitive and the resistant subpopulations are exponential viz:
S ( D ) = e-a°D
R{D) = e~a'D
Then:
The parameter V can assume values between 1 and 0, and in this special case in which resistant and sensitive subpopulations exhibit exponential survival curves it is independent of dose.
A L P H A P A R T I C L E S A N D T H E C E L L C Y C L E 95
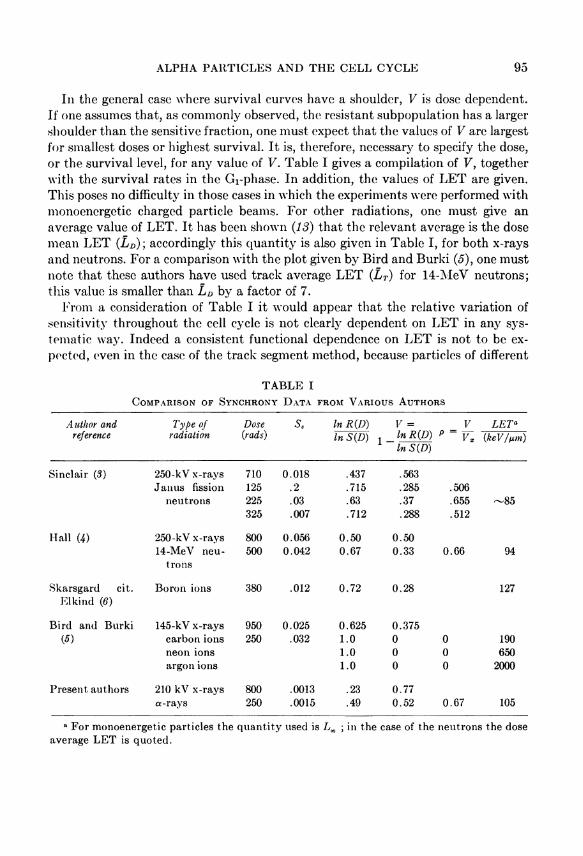
In the general case where survival curves have a Shoulder, V is dose dependent. If one assumes that, as commonly observed, the resistant subpopulation has a larger Shoulder than the sensitive fraction, one must expect that the values of V are largest for smallest doses or highest survival. It is, therefore, necessary to specify the dose, or the survival level, for any value of V. Table I gives a compilation of V, together with the survival rates in the G r phase. In addition, the values of L E T are given. This poses no difflculty in those cases in which the experiments were performed with monoenergetic charged particle beams. For other radiations, one must give an average value of L E T . It has been shown (13) that the relevant average is the dose mean L E T ( L D ) ; accordingly this quantity is also given in Table I, for both x-rays and neutrons. For a comparison with the plot given by Bird and Burki (5), one must note that these authors have used track average L E T ( L T ) for 14-MeV neutrons; this value is smaller than L D by a factor of 7.
From a consideration of Table I i t would appear that the relative Variation of sensitivity throughout the cell cycle is not clearly dependent on L E T in any sys-tematic way. Indeed a consistent functional dependence on L E T is not to be ex-pected, even in the case of the track segment method, because particles of different
T A B L E I
C O M P A R I S O N O F S Y N C H R O N Y D A T A F R O M V A R I O U S A U T H O R S
Author and reference
Type of r a d i a t i o n
Dose {rads)
s. In R ( D ) In S { D )
V = In R { D ) I n S ( D )
V P = K
L E T a
{keV/ixm)
Sinclair ( 3 ) 250-kV x-rays 710 0.018 .437 .563 Janus fission 125 .2 .715 .285 .506
neutrons 225 .03 .63 .37 .655 - 8 5 325 .007 .712 .288 .512
Hall W) 250-kV x-rays 800 0.056 0.50 0.50 14-MeV neu 500 0.042 0.67 0.33 0.66 94
trons
Skarsgard cit. Boron ions 380 .012 0.72 0.28 127 Elkind ( 6 )
Bird and Burki 145-kV x-rays 950 0.025 0.625 0.375 ( 5 ) carbon ions 250 .032 1.0 0 0 190
neon ions 1.0 0 0 650 argon ions 1.0 0 0 2000
Present authors 210 kV x-rays 800 .0013 .23 0.77 a-rays 250 .0015 .49 0.52 0.67 105
a For monoenergetic particles the quantity used is L x ; in the case of the neutrons the dose average L E T is quoted.

96 H A L L E T A L .
Charge and different velocity can produce the same value of L D . Such particles can have different radial profiles of energy deposition around their tracks. Microdosi-metric considerations show that, the use of L D as an indicator of radiation quality is even more seriously limited for a mixed radiation field.
Another complication in the attempt to find a functional dependence of sensitivity Variation on L E T , results from the fact that different experiments are not always strictly comparable, because the experimentally observed values of V are usually less than maximal because of imperfect synchronization of the cells. This accounts in part for the differences in the various observed values of V for x-rays given in Table I. For this rcason it is practical to normalize the Parameters V relative to the value for x-rays obtained in the same experiment. Owing to the limita-tion of the experimental data, one must be content with an approximate comparison, and for this reason the parameter
V p = v x
is also listed in Table I. Because Vx is expected to be largest at highest survival levels, p must be smallest at highest survival levels. For each value of p, the survival level for Gi cells exposed to x-rays is given ( S 8 ) .
It should be noted that the limitations of the comparison in Table I are due mainly to the fact that the change of the survival curve with cell age cannot be described realistically by the Variation of one parameter. However, in most cases an analysis in terms of several parameters cannot be performed, because the survival throughout the cell cycle has been determined at only a limited number of dose levels.
The survival curve for an asynchronous cell population exposed to a-particles is observed to be an exponential funetion of dose within experimental uncertainty (see Fig. 3). This result might be considered to be surprising since the survival curves for synchronized cells in various phases of the cycle each appear to be exponential (5). The curve for asynchronous cells is a composite of those for the various phases of the cycle. Superposition of exponential curves of different slope results in a curve which is coneave from above. Accordingly the survival curve for asynchronous cells would have to be nonexponential.
A quantitative evaluation shows, however, that the deviation from the exponential survival cannot be expected to be observed experimentally. Figure 5 shows an estimate of the cellular sensitivity throughout the cell cycle which would result if the curve in Fig. 4 were corrected for increase in cellular multiplicity and the loss of S3^nchronization with cell age. If one uses this curve and assumes that at any given cell age survival is exponential, one can calculate the resulting curve for a nonsyn-chronized population in logarithmic growth. The result is shown in Fig. 6 as a solid curve. One notes that the curve indeed deviates from the exponential shape. Despite the fact that the slope of the curve decreases by about a factor of 2 with increasing

A L P H A P A R T I C L E S A N D T H E C E L L C Y C L E 97
IC"
10"
' I ' I ' I ' I 1 I 1
2 5 0 R A D S a - r a y s
0 4 6 8 T(HOURS)
_l_ 10 12
F I G . 5. Survival of synchronized Chinese hamster cells exposed to 250 rads of a-particles at different times after Synchronisation. The curve represents an estimate of the data in Fig. 4 corrected for multiplicity increase and loss of synchrony with time.
o , 0 3
10 100 2 0 0 3 0 0 4 0 0 5 0 0
A B S O R B E D DOSE ( R A D ) 6 0 0 7 0 0
F I G . 6. Survival curve (solid line) for a nonsynchronized population in logarithmic growth under the assumption that the survival curves for all individual cell ages are exponential. The broken line is inserted to indicate the deviations from an exponential funetion.
dose, the deviation from the exponential shape is certainly within the limits of experimental accuraey. This is indicated by a comparison with the exponential funetion represented by the broken line. This consideration also implies that there may be cumulative damage which is masked by the limited experimental accuraey and the sensitivity variations. R E C E I V E D : S March 1972
Bayerische Staatsbibliothek]
München

98 H A L L E T A L .
R E F E R E N C E S
1. G . W . B A R E N D S K X , T . L . J . B E U S K E R , A. J . V E R G R O E S E N , and L . B U D K E , Effects of different
ionizing radiations on human cells in tissue culture. II. Biological experiments. Radial. Res. 13, 841-849 (1960).
2. W. K . S I N C L A I R and R. A. M O R T O N , Variation in response during the division cycle of par-tially synchronized Chinese hamster cells in culture. N a t u r e London 199,1158-1100 (1902).
3. W. K . S I N C L A I R , Dependence of radiosensitivity in cell age. In Time and Dose Relationships i n Radiation Biology as Applied to Radiotherapy. B N L Report 50203 (C-57), pp. 97-107, 1969.
4. E . J . H A L L , Radiobiological measurements with 14 MeV neutrons. Brit. J. Radiol. 42, 805-813 (1909).
5. R. B I R D and J . B U R K I , Inactivation of mammalian cells at different stages of the cell cycle as a funetion of radiation linear energy transfer. In Biophysical Aspects of Radiation Quality, International Atomic Energy Agency, Vienna, pp. 241-250, 1971.
6. M . M . E L K I N D , Damage registration and repair following neutron irradiation, In Proc. Conf. Neutrons i n Radialion Biology, U S A E C Conf. 691100, 1909.
7. R. G . H A M , An improved nutrient Solution for diploid Chinese hamster and human cell lines. E x p . C e l l Res. 28 , 515-526 (1963).
8. R. G . H A M and T . T . P U C K , Quantitative colonial growth of isolated mammalian cells. In Methods i n Enzymology (S. P. Colowick and N . O. Kaplan, eds.) Vol . V, pp. 90-119, Academic Press, New York, 1962.
9. W. K . SINCLAIR, Hydroxyurea; differential lethal effects on cultured mammalian cells during the cell cycle. Science 150,1729-1731 (1965).
10. M. M . E L K I N D , H . S U T T O N G I L B K R T , W . B. M O S E S , and C . K A M P E R , Sublethal and lethal
radiation damage. N a t u r e LondonZL4,1088-1092 (1967). 1 1 . C . W . Y O U N G and S. H O D A S , Hydroxyurea; inhibitory effect on D N A metabolism. Science
146, 1172-1174 (1964). 12. H . H . Rossi, L . J . G O O D M A N , and M . P E A R L M A N , Device for plating of active deposits.
Annual Report on Research Project NYO-2740-3, pp. 143-140, January 1, 1966. 13. A . M . K E L L E R E R and H . H . Rossi, R B E and the primary mechanisms of radiation action.
Radiat. Res. 47 , 15-34 (1971).





























