Embed Size (px)
Citation preview

Comparative Biochemistry and Physiology, Part B 164 (2013) 221–228
Contents lists available at SciVerse ScienceDirect
Comparative Biochemistry and Physiology, Part B
j ourna l homepage: www.e lsev ie r .com/ locate /cbpb
Superoxide dismutases from larvae of the camel tickHyalomma dromedarii
Mahmoud A. Ibrahim a,⁎, Mona M. Mohamed b, Abdel-Hady M. Ghazy a, Hassan M.M. Masoud a
a Molecular Biology Department, National Research Centre, El-Tahrir st., Dokki, Cairo, Egyptb Zoology Department, Faculty of Science, Cairo University, Cairo, Egypt
Abbreviations: BSA, Bovine serum albumin; PAGE, Posis; NBT, Nitroblue tetrazolium; PMS, Phenazine methosspecies; SOD, Superoxide dismutase; TLSOD, Tick larvae⁎ Corresponding author. Tel.: +20 2 33335980; fax: +
E-mail address: [email protected] (M.A. Ibr
1096-4959/$ – see front matter © 2013 Elsevier Inc. Allhttp://dx.doi.org/10.1016/j.cbpb.2013.01.002
a b s t r a c t
a r t i c l e i n f oArticle history:Received 8 October 2012Received in revised form 8 January 2013Accepted 8 January 2013Available online 16 January 2013
Keywords:Superoxide dismutaseCamel tick larvaePurification and characterization
Three superoxide dismutases (EC 1.15.1.1) (TLSOD1, TLSOD2 and TLSOD3) were purified from larvae of thecamel tick Hyalomma dromedarii by ammonium sulfate precipitation, ion exchange and gel filtration columns.SDS-PAGE revealed that the subunit molecular masses of the SODs are 40±2 kDa, 67±1.5 kDa and 45±2.6 kDa for TLSOD1, TLSOD2 and TLSOD3, respectively. TLSOD1 and TLSOD2 are monomeric proteins, whileTLSOD3 isoenzyme exhibits dimeric structurewith nativemolecularmass of 90 kDa. The pI values are estimatedat pH 8.0, pH 7.2 and pH 6.6 for the three SODswhich displayed pH optima at 7.6, 8.0 and 7.8, respectively. CuCl2and ZnCl2 increase the activity of TLSOD2and TLSOD3,whileMnCl2 increases the activity of TLSOD1. KCN inhibitsthe activity of TLSOD2 and TLSOD3, while a remarkable resistance of TLSOD1 isoenzymewas detected. TLSOD1 issuggested to be a manganese containing isoenzyme while TLSOD2 and TLSOD3 are suggested to be copper/zinc-containing isoenzymes. These results indicate the presence of three different forms of SODs in the larvalstage of camel tick. This finding will contribute to our understanding of the physiology of these ectoparasitesand the development of non-traditional methods to control them.
© 2013 Elsevier Inc. All rights reserved.
1. Introduction
Antioxidant defense molecules fall into one of two categories: en-zymatic or nonenzymatic. Specifically, enzymatic defenses includemolecules that are capable of removing, neutralizing, or scavengingreactive oxygen species (ROS) that include hydroxyl radical, superox-ide anion, hydrogen peroxide, and singlet oxygen. Antioxidant de-fense mechanisms may also include the inhibition of ROS formation,the binding of metal ions needed for catalysis of ROS generation,and the up-regulation of antioxidant defense activity, suggestingthat the removal of free radicals by cells is not likely accomplishedby a single pathway but composed of several cascades of intricatelyrelated events (Mruk et al., 2002). The enzymatic antioxidant systemis represented by enzymes such as superoxide dismutase (SOD), cat-alase (CAT), glutathione peroxidases (GPxs) (Rikans and Hornbrook,1997) and glutathione reductase (GR) (Sies, 1993).
Superoxide dismutases (SOD; superoxide oxidoreductase EC1.15.1.1)are ubiquitous metalloenzymes important for aerobic and anaerobicorganisms that protect against oxidative stress from superoxide radicals
lyacrylamide gel electrophore-ulphate; ROS, Reactive oxygensuperoxide dismutase.20 2 33370931.
ahim).
rights reserved.
in living cells (Osatomi et al., 2001; Babitha et al., 2002; Yao et al.,2004). They catalyze the conversion of superoxide radicals to molecu-lar oxygen and hydrogen peroxide to protect organisms against toxicradicals produced during oxidative processes (Bao et al., 2009; Ghoshet al., 2010). In addition, hydrogen peroxide is safely decomposed bycatalase to water and oxygen (Terevinto et al., 2010). SOD is found invirtually all oxygen-consuming organisms (Yao et al., 2004; Törö etal., 2009). Superoxide dismutases are classified according to the metalspecies occurring in the active site of the enzyme into four groups(Bittencourt et al., 2004), Cu/Zn-SOD, Mn-SOD, Fe-SOD (Miao et al.,2009) and Ni-SOD (Ghosh et al., 2010). Cu/Zn-SOD is present in nearlyall eukaryotic organisms (Weisiger and Fridovich, 1973; Ekanayake etal., 2006) and in the periplasm of a significant number of bacteria(Törö et al., 2009). It is a characteristic of the cytosol of eukaryotes,fungi, and mammalian cells (Palma et al., 1997; Oztürk-Urek andTarhan, 2001). MnSODs are found in prokaryotes and in mitochondriaof eukaryotes (Kawaguchi et al., 1989). FeSODs are mainly found inprokaryotes and chloroplasts (Halliwell and Gutteridge, 1999). Ni-SODwas found in the filamentous gram-positive bacteria Streptomyces sp.(Whittaker, 2010).
Ticks (Arachnida) are blood-sucking arthropods that infest a widearray of species (Sauer et al., 1995), including humans and almostevery animal of economic importance, and cause significant losses tolivestock production (Bowman et al., 1996). Studies about ecology,behavior and physiology of ticks afford a better understanding ofthese organisms and therefore become important tools to developnew control methods. Due to the rapid increase in pesticide-resistant

222 M.A. Ibrahim et al. / Comparative Biochemistry and Physiology, Part B 164 (2013) 221–228
tick populations (Davey and George, 1998), the study of tick physiologyhas gained increasing importance regarding the mechanisms involvedin detoxification of toxins (Hemingway, 2000; Kostaropoulos et al.,2001). These mechanisms, in general, not only act against specifictoxicmolecules, but also help in themaintenance of physiologic homeo-stasis, avoiding oxidative damage generated by ROS (Freitas et al.,2007).
Therefore, this study aims at purification and characterization ofSOD from the larvae of the camel tick Hyalomma dromedarii to inves-tigate its role as an antioxidant enzyme. Such study will establish thebasis for better understanding of some of the antioxidant defense sys-tems in the tick for developing new effective and untraditional bio-chemical control of tick.
2. Materials and methods
2.1. Ticks
Engorged camel tick H. dromedarii females were collected from aCamel market near Cairo and held at 28 °C and 85% relative humidity.Eggs were collected daily from fertilized oviposition female ticks andeither frozen immediately (−40 °C) or incubated under the samecondition until the appropriate age and transferred to frozen storageat intervals of three days (0, 3, 6, 9 etc.). The hatched larvae were col-lected at day 27.
2.2. Chemicals
Phenylmethylsulfonylfluoride (PMSF), xanthine sodium salt, xan-thine oxidase enzyme, diethylaminoethyl-cellulose (DEAE-cellulose),nitroblue tetrazolium (NBT), phenazine methosulphate (PMS), iso-electric focusing (IEF) standardmarkers mixture pI 3.6–9.3, molecularweight marker kits for gel filtration and Sephacryl S-300 were pur-chased from Sigma-Aldrich Chemical Co. All other chemicals were ofanalytical grade.
2.3. Assay of superoxide dismutase
The assay of SOD activity was carried out according to the methoddescribed by McCord and Fridovich, 1969. The assay reaction mixturecontained in a total volume of 1.0 mL of 50 mM potassium phosphatebuffer pH 7.8, with 0.01 mM cytochrome c, 0.1 mM EDTA and0.05 mM sodium xanthine. The reaction was started by adding21 mU xanthine oxidase. One unit of SOD activity is defined as theamount of enzyme giving 50% inhibition of cytochrome c reductionat 550 nm.
2.4. Purification of camel tick larval superoxide dismutase
2.4.1. Preparation of crude extractCamel tick larvae (2 g) were homogenized in 10 mL of 20 mM K-
phosphate buffer pH 7.4, using a Teflon-pestled homogenizer. Celldebris and insoluble materials were removed by centrifugation at12,000 g for 20 min and the supernatant was saved and designated ascrude extract.
2.4.2. Ammonium sulfate precipitationThe crude extract was brought to 80% saturation by gradually
adding solid (NH4)2SO4 and stirred for 30 min at 4 °C. The pelletwas obtained by centrifugation at 12,000 g for 30 min and dissolvedin 20 mM K-phosphate buffer (pH 7.4) and dialyzed extensivelyagainst the same buffer.
2.4.3. DEAE-cellulose column chromatographyThe dialyzed sample was chromatographed on a DEAE-cellulose col-
umn (12×2.4 cm i.d.) previously equilibratedwith 20 mM K-phosphate
buffer (pH 7.4). The adsorbed proteins were elutedwith a stepwise NaClgradient ranging from 0 to 1 M prepared in the equilibration buffer at aflow rate of 60 mL/h. Fractions (5 mL) were collected and the fractionscontaining SOD activity were pooled and lyophilized.
2.4.4. Sephacryl S-300 column chromatographyThe concentrated solution containing the SOD activity was applied
onto a Sephacryl S-300 column (142 cm×1.75 cm i.d.). The columnwas equilibrated and developed with 20 mM K-phosphate buffer(pH 7.4) at a flow rate of 30 mL/h and 2 mL fractions were collected.
2.5. Electrophoretic analysis
Native gel electrophoresis was carried out with 7% PAGE accordingto Smith (1969). SDS-PAGE was performed with 12% polyacrylamidegel following to Laemmli (1970). The subunit molecular weights ofthe purified SOD isoenzymes were determined by SDS-PAGE as de-scribed by Weber and Osborn (1969). Electrofocusing was performedaccording to O'Farrell (1975) and the isoelectric point (pI) valueswere calculated from a calibration curve as described by Ubukaet al. (1987). Proteins were stained with 0.25% Coomassie brilliantblue R-250.
2.6. Protein determination
Protein was determined by the dye binding assay method ofBradford (1976) using bovine serum albumin as a standard.
3. Results
3.1. Changes in SOD specific activity and isoenzyme pattern duringembryogenesis
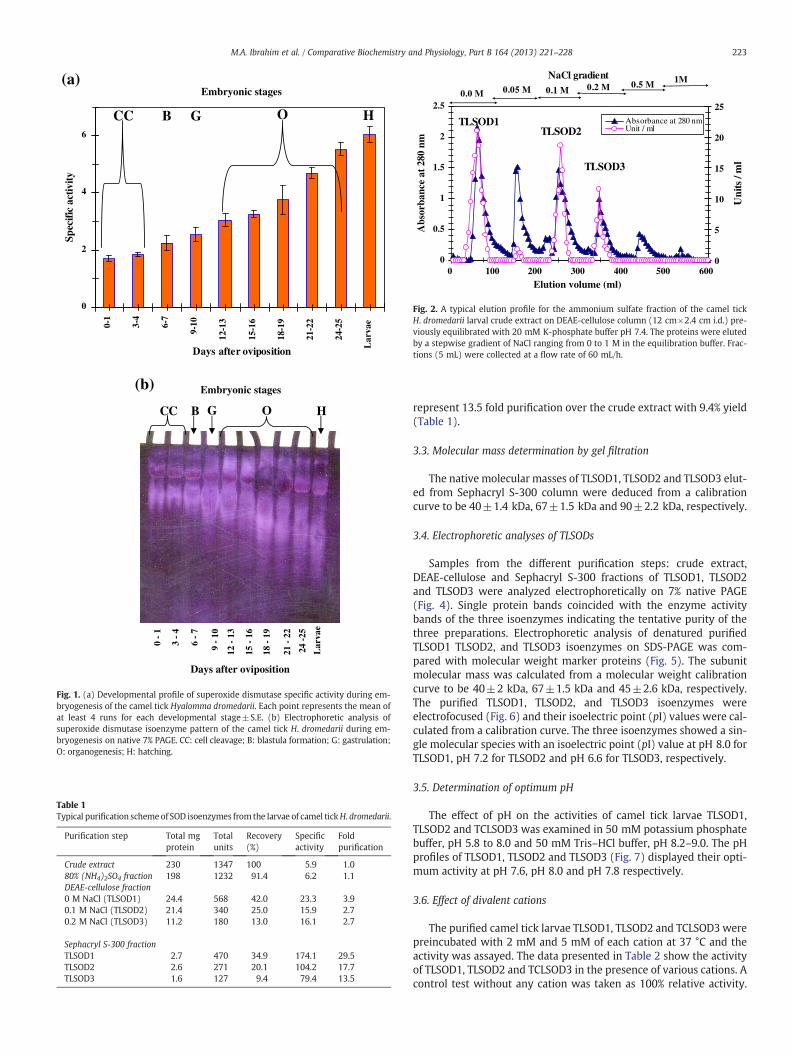
A developmental profile of SOD specific activity was constructedduring embryogenesis of the camel tick H. dromedarii which revealedthat the SOD specific activity was increased gradually and reached itshighest level at the larval stage (6.05±0.12 units/mg protein) andwas confirmed by SOD isoenzyme pattern on native 7% PAGE(Fig. 1). The just oviposited eggs contained only two SOD isoenzymes,TLSOD1 and TLSOD2, from the maternal origin. At the beginning ofblastula formation, the amount of both isoenzymes increased graduallyuntil the gastrulation stage. A new third SOD isoenzyme, TLSOD3, isexpressed at the start of organogenesis and its amount increased tilllarval hatching stage (Fig. 1b).
3.2. Purification of SOD isoenzymes from camel tick larvae
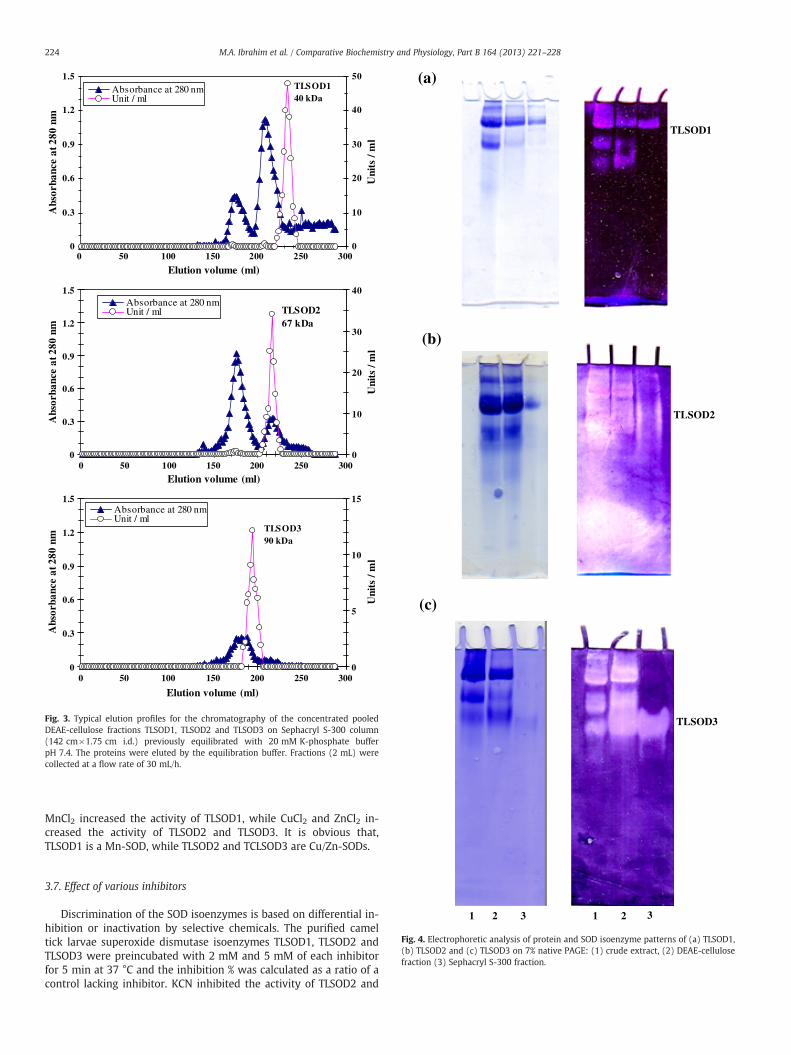
The SOD specific activity of the larval crude extract was foundto be 5.9 units/mg protein. A typical purification scheme of SODfrom the camel tick H. dromedarii larvae is presented in Table 1.After ammonium sulfate precipitation, most of the SOD activitywas precipitated so that 91.4% of the activity was recovered. TheDEAE-cellulose elution profile (Fig. 2) revealed the presence ofthree major peaks containing SOD activity designated as TLSOD1,TLSOD2 and TLSOD3 and eluted with 0.0 M NaCl, 0.1 M NaCl and0.2 M NaCl. The DEAE-cellulose fractions of each peak were pooled,concentrated by lyophilization and applied onto a Sephacryl S-300column. The elution profiles of TLSOD1, TLSOD2 and TLSOD3 on theSephacryl S-300 column (Fig. 3) revealed the presence of one peakof the enzyme activity. The specific activity of TLSOD1 was increasedto 174.1 units/mg protein which represent 29.5 fold purification overthe crude extract with 34.9% yield. The specific activity of TLSOD2was increased to 104.2 units/mg protein which represent 17.7-foldpurification over the crude extract with 20.1% yield. Also, the specificactivity of TLSOD3 was increased to 79.4 units/mg protein which
0
2
4
6
0-1
3-4
6-7
9-10
12-1
3
15-1
6
18-1
9
21-2
2
24-2
5
Lar
vae
Days after oviposition
Spec
ific
activ
ity(a)
(b)
Lar
vae
24 -
25
Days after oviposition
21 -
22
18 -
19
15 -
16
12 -
13
9 -
10
6 -
7
3 -
4
0 -
1
CC B G O H
Embryonic stages
CC B G O H
Embryonic stages
Fig. 1. (a) Developmental profile of superoxide dismutase specific activity during em-bryogenesis of the camel tick Hyalomma dromedarii. Each point represents the mean ofat least 4 runs for each developmental stage±S.E. (b) Electrophoretic analysis ofsuperoxide dismutase isoenzyme pattern of the camel tick H. dromedarii during em-bryogenesis on native 7% PAGE. CC: cell cleavage; B: blastula formation; G: gastrulation;O: organogenesis; H: hatching.
Table 1Typical purification schemeof SOD isoenzymes from the larvae of camel tickH. dromedarii.
Purification step Total mgprotein
Totalunits
Recovery(%)
Specificactivity
Foldpurification
Crude extract 230 1347 100 5.9 1.080% (NH4)2SO4 fraction 198 1232 91.4 6.2 1.1DEAE-cellulose fraction0 M NaCl (TLSOD1) 24.4 568 42.0 23.3 3.90.1 M NaCl (TLSOD2) 21.4 340 25.0 15.9 2.70.2 M NaCl (TLSOD3) 11.2 180 13.0 16.1 2.7
Sephacryl S-300 fractionTLSOD1 2.7 470 34.9 174.1 29.5TLSOD2 2.6 271 20.1 104.2 17.7TLSOD3 1.6 127 9.4 79.4 13.5
0
0.5
1
1.5
2
2.5
0 100 200 300 400 500 600
Elution volume (ml)
Abs
orba
nce
at 2
80 n
m
0
5
10
15
20
25
Uni
ts /
ml
Absorbance at 280 nmUnit / ml
0.0 M 0.05 M 0.1 M 0.2 M 0.5 M 1M
TLSOD1TLSOD2
TLSOD3
NaCl gradient
Fig. 2. A typical elution profile for the ammonium sulfate fraction of the camel tickH. dromedarii larval crude extract on DEAE-cellulose column (12 cm×2.4 cm i.d.) pre-viously equilibrated with 20 mM K-phosphate buffer pH 7.4. The proteins were elutedby a stepwise gradient of NaCl ranging from 0 to 1 M in the equilibration buffer. Frac-tions (5 mL) were collected at a flow rate of 60 mL/h.
223M.A. Ibrahim et al. / Comparative Biochemistry and Physiology, Part B 164 (2013) 221–228
represent 13.5 fold purification over the crude extract with 9.4% yield(Table 1).
3.3. Molecular mass determination by gel filtration
The native molecular masses of TLSOD1, TLSOD2 and TLSOD3 elut-ed from Sephacryl S-300 column were deduced from a calibrationcurve to be 40±1.4 kDa, 67±1.5 kDa and 90±2.2 kDa, respectively.
3.4. Electrophoretic analyses of TLSODs
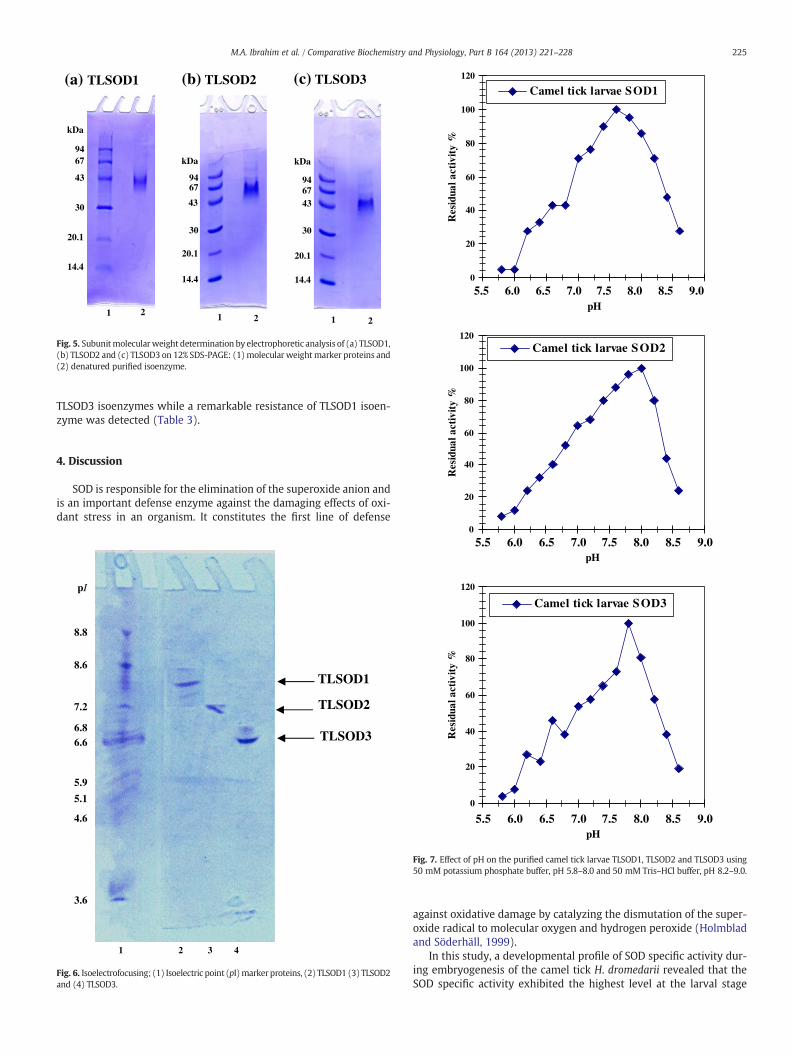
Samples from the different purification steps: crude extract,DEAE-cellulose and Sephacryl S-300 fractions of TLSOD1, TLSOD2and TLSOD3 were analyzed electrophoretically on 7% native PAGE(Fig. 4). Single protein bands coincided with the enzyme activitybands of the three isoenzymes indicating the tentative purity of thethree preparations. Electrophoretic analysis of denatured purifiedTLSOD1 TLSOD2, and TLSOD3 isoenzymes on SDS-PAGE was com-pared with molecular weight marker proteins (Fig. 5). The subunitmolecular mass was calculated from a molecular weight calibrationcurve to be 40±2 kDa, 67±1.5 kDa and 45±2.6 kDa, respectively.The purified TLSOD1, TLSOD2, and TLSOD3 isoenzymes wereelectrofocused (Fig. 6) and their isoelectric point (pI) values were cal-culated from a calibration curve. The three isoenzymes showed a sin-gle molecular species with an isoelectric point (pI) value at pH 8.0 forTLSOD1, pH 7.2 for TLSOD2 and pH 6.6 for TLSOD3, respectively.
3.5. Determination of optimum pH
The effect of pH on the activities of camel tick larvae TLSOD1,TLSOD2 and TCLSOD3 was examined in 50 mM potassium phosphatebuffer, pH 5.8 to 8.0 and 50 mM Tris–HCl buffer, pH 8.2–9.0. The pHprofiles of TLSOD1, TLSOD2 and TLSOD3 (Fig. 7) displayed their opti-mum activity at pH 7.6, pH 8.0 and pH 7.8 respectively.
3.6. Effect of divalent cations
The purified camel tick larvae TLSOD1, TLSOD2 and TCLSOD3 werepreincubated with 2 mM and 5 mM of each cation at 37 °C and theactivity was assayed. The data presented in Table 2 show the activityof TLSOD1, TLSOD2 and TCLSOD3 in the presence of various cations. Acontrol test without any cation was taken as 100% relative activity.
0
0.3
0.6
0.9
1.2
1.5
Elution volume (ml)
Abs
orba
nce
at 2
80 n
m
0
10
20
30
40
50
Uni
ts /
ml
Absorbance at 280 nmUnit / ml
TLSOD140 kDa
0
0.3
0.6
0.9
1.2
1.5
Elution volume (ml)
Abs
orba
nce
at 2
80 n
m
0
10
20
30
40
Uni
ts /
ml
Absorbance at 280 nmUnit / ml TLSOD2
67 kDa
0
0.3
0.6
0.9
1.2
1.5
0 50 100 150 200 250 300
0 50 100 150 200 250 300
0 50 100 150 200 250 300
Elution volume (ml)
Abs
orba
nce
at 2
80 n
m
0
5
10
15
Uni
ts /
ml
Absorbance at 280 nmUnit / ml
TLSOD390 kDa
Fig. 3. Typical elution profiles for the chromatography of the concentrated pooledDEAE-cellulose fractions TLSOD1, TLSOD2 and TLSOD3 on Sephacryl S-300 column(142 cm×1.75 cm i.d.) previously equilibrated with 20 mM K-phosphate bufferpH 7.4. The proteins were eluted by the equilibration buffer. Fractions (2 mL) werecollected at a flow rate of 30 mL/h.
(a)
(b)
(c)
TLSOD1
TLSOD2
TLSOD3
224 M.A. Ibrahim et al. / Comparative Biochemistry and Physiology, Part B 164 (2013) 221–228
MnCl2 increased the activity of TLSOD1, while CuCl2 and ZnCl2 in-creased the activity of TLSOD2 and TLSOD3. It is obvious that,TLSOD1 is a Mn-SOD, while TLSOD2 and TCLSOD3 are Cu/Zn-SODs.
1 21 332
Fig. 4. Electrophoretic analysis of protein and SOD isoenzyme patterns of (a) TLSOD1,(b) TLSOD2 and (c) TLSOD3 on 7% native PAGE: (1) crude extract, (2) DEAE-cellulosefraction (3) Sephacryl S-300 fraction.
3.7. Effect of various inhibitors
Discrimination of the SOD isoenzymes is based on differential in-hibition or inactivation by selective chemicals. The purified cameltick larvae superoxide dismutase isoenzymes TLSOD1, TLSOD2 andTLSOD3 were preincubated with 2 mM and 5 mM of each inhibitorfor 5 min at 37 °C and the inhibition % was calculated as a ratio of acontrol lacking inhibitor. KCN inhibited the activity of TLSOD2 and
1 2
kDa
9467
43
30
20.1
14.4
(a) TLSOD1
kDa
946743
30
20.1
14.4
kDa
9467
43
30
20.1
14.4
1 2 1 2
(b) TLSOD2 (c) TLSOD3
Fig. 5. Subunitmolecularweight determination by electrophoretic analysis of (a) TLSOD1,(b) TLSOD2 and (c) TLSOD3 on 12% SDS-PAGE: (1)molecular weightmarker proteins and(2) denatured purified isoenzyme.
0
20
40
60
80
100
120
pH
Res
idua
l act
ivit
y %
Camel tick larvae SOD1
80
100
120
5.5 6.0 6.5 7.0 7.5 8.0 8.5 9.0
ctiv
ity
%
Camel tick larvae SOD2
225M.A. Ibrahim et al. / Comparative Biochemistry and Physiology, Part B 164 (2013) 221–228
TLSOD3 isoenzymes while a remarkable resistance of TLSOD1 isoen-zyme was detected (Table 3).
20
40
60
Res
idua
l a
4. DiscussionSOD is responsible for the elimination of the superoxide anion andis an important defense enzyme against the damaging effects of oxi-dant stress in an organism. It constitutes the first line of defense
pI
8.8
8.6
7.2
6.6
4.6
3.6
5.1
5.9
6.8
TLSOD1
TLSOD3
TLSOD2
1 2 3 4
Fig. 6. Isoelectrofocusing; (1) Isoelectric point (pI)marker proteins, (2) TLSOD1 (3) TLSOD2and (4) TLSOD3.
0
20
40
60
80
100
120
pH
Res
idua
l act
ivit
y %
Camel tick larvae SOD3
0
5.5 6.0 6.5 7.0 7.5 8.0 8.5 9.0
5.5 6.0 6.5 7.0 7.5 8.0 8.5 9.0pH
Fig. 7. Effect of pH on the purified camel tick larvae TLSOD1, TLSOD2 and TLSOD3 using50 mM potassium phosphate buffer, pH 5.8–8.0 and 50 mM Tris–HCl buffer, pH 8.2–9.0.
against oxidative damage by catalyzing the dismutation of the super-oxide radical to molecular oxygen and hydrogen peroxide (Holmbladand Söderhäll, 1999).
In this study, a developmental profile of SOD specific activity dur-ing embryogenesis of the camel tick H. dromedarii revealed that theSOD specific activity exhibited the highest level at the larval stage

Table 2Effect of divalent cations on the purified camel tick larvae TLSOD1, TLSOD2 andTLSOD3.
Reagent Final concentration(mM)
Residual activity (%)
TLSOD1 TLSOD2 TLSOD3
Control – 100 100 100CaCl2 2.0 88.2 100.0 97.2
5.0 76.5 106.9 91.6CoCl2 2.0 94.1 103.4 102.3
5.0 85.3 110.3 108.3CuCl2 2.0 97.1 110.3 111.1
5.0 94.1 117.2 122.2FeCl2 2.0 52.9 72.4 58.3
5.0 44.1 62.2 38.8MgCl2 2.0 70.6 86.2 52.7
5.0 61.7 70.0 44.4MnCl2 2.0 111.8 100.0 94.4
5.0 123.5 96.5 97.2NiCl2 2.0 91.2 100.0 105.5
5.0 79.4 106.8 113.8ZnCl2 2.0 97.1 103.4 108.3
5.0 100.0 117.2 116.6
These values represent % of the control and the means of triplicate experiments.
226 M.A. Ibrahim et al. / Comparative Biochemistry and Physiology, Part B 164 (2013) 221–228
which was confirmed by SOD isoenzyme pattern on native 7% PAGE(Fig. 1). The tick embryo evolved a second Cu/Zn SOD at the start oforganogenesis suggesting its necessity after the larval hatching, startfeeding and exposure to environmental toxins (Fig. 1b). This studypresents a simple and reproducible purification method for SOD iso-enzymes from the larvae of the camel tick H. dromedarii. The purifica-tion procedure was carried out by ammonium sulfate precipitation,ion exchange chromatography on DEAE-cellulose column and gel fil-tration chromatography on Sephacryl S-300 column. Similar purifica-tion procedures of SODs were reported: SOD was purified from a teaclone (Vyas and Kumar, 2005), from muscle tissue of the shrimpMacrobrachium nipponense (Yao et al., 2004) and from hog erythro-cytes (Bartkowiak et al., 1978).
In the present study, the chromatography on DEAE-cellulose re-solved the SOD activity conveniently into three isoenzymes TLSOD1,TLSOD2 and TLSOD3 (Fig. 2). The chromatography on SephacrylS-300 column eluted each isoenzyme as a single enzyme activity
Table 3Effect of inhibitors on the purified camel tick larvae TLSOD1, TLSOD2 and TLSOD3.
Reagent Final concentration(mM)
Inhibition (%)
TLSOD1 TLSOD2 TLSOD3
Control – 0 0 0Potassium cyanide (KCN) 2.0 6.8 72.7 77.3
5.0 15.9 90.1 100.0Hydrogen peroxide (H2O2) 2.0 40.9 63.6 31.8
5.0 63.6 81.8 54.5Sodium azide (NaN3) 2.0 27.3 9.1 4.6
5.0 50.0 18.2 6.8Sodium dodecyl sulfate (SDS) 2.0 22.7 9.1 14.6
5.0 36.4 13.6 27.3Ethylenediamine tetraaceticacid (EDTA)
2.0 11.4 18.2 13.45.0 22.7 33.4 27.7
DL-Dithiothreitol (DTT) 2.0 4.6 12.6 4.65.0 13.6 22.7 9.1
β-Mercaptoethanol 2.0 18.2 4.5 22.75.0 24.7 13.6 31.8
1,10-Phenanthroline 2.0 13.6 18.2 18.25.0 18.2 25.3 29.5
Phenylmethylsulfonylfluoride (PMSF)
2.0 0.0 6.8 4.65.0 4.6 13.6 9.1
Iodoacetamide 2.0 0.0 0.0 0.05.0 0.0 4.0 4.6
Potassium dichromate(K2Cr2O7)
2.0 77.3 63.6 45.55.0 90.1 81.8 68.2
These values represent % of the control and the means of triplicate experiments.
peak and the deduced molecular masses from their elution volumeswere found to be 40±1.4 kDa, 67±1.5 kDa and 90±2.2 kDa forTLSOD1, TLSOD2 and TLSOD3 respectively (Fig. 3). The overall yieldof the three isoenzymes from the Sephacryl S-300 column is 64.4%(Table 1). A large variety of purification fold and recovery percent ofSOD were reported; the SOD was purified from hog liver 25.7-foldwith 29.6% yield (Bartkowiak et al., 1978), from a tea clone 51-foldwith 3.77% yield (Vyas and Kumar, 2005), from yeast 28.5-fold with53% yield (Nedeva et al., 2009) and from chicken liver 285.8-foldwith 7.3% yield (Oztürk-Urek and Tarhan, 2001).
All of the SOD isoenzymes turned out the presence of a major pro-tein band which coincided with the enzyme activity band confirmingthat the single protein band is the enzyme band as shown for TLSOD1,TLSOD2 and TLSOD3 (Fig. 4). Comparison of subunit molecular masswith that of native intact protein determined by gel filtration revealedthat TLSOD1 and TLSOD2 isoenzymes are monomeric proteins com-posed of single polypeptide chain while the TLSOD3 isoenzymeexhibited dimeric structure with two identical subunits each is45 kDa (Fig. 5). Many SODs were reported to have a monomericstructure such as; common carp liver SOD with molecular mass of36.0 kDa (Vig et al., 1989) and oriental river prawn SOD with a66.1 kDa (Yao et al., 2007). There are also many SODs which werereported to have a dimeric structure such as that of pumpkin(Cucurbita moschata) pulp SOD with two identical subunits of17.4 kDa (Qin et al., 2012), chicken erythrocyte SOD with subunitmolecular mass of 15 kDa (Aydemir and Tarhan, 2001), hog liverSOD with subunit molecular mass of 16 kDa (Bartkowiak et al.,1978) and chicken liver SOD with subunit molecular mass of 16 kDa(Oztürk-Urek and Tarhan, 2001). TLSOD2 molecular mass is almost67 kDa and this is consistent with that of oriental river prawn SOD(Yao et al., 2007) and TLSOD1 molecular mass is 40 kDa which is con-sistent with that of common carp liver SOD (Vig et al., 1989).
The isoelectric point (pI) values were estimated for the purifiedSOD isoenzymes from camel tick larvae (Fig. 6). Similar isoelectricpoint pI values were reported for Cryptococcus neoformans var.neoformans bacteria, four SOD isoenzymes with pIs of 5.9, 6.15, 6.35,and 6.6, and in Cryptococcus neoformans var. gattii bacteria, one SODisoenzyme with a pI value of 7.5 (Hamilton and Holdom, 1997), andin Paralichthys olivaceus fish, three SOD isoenzymes with pIs of 5.9,6.0 and 6.2 (Osatomi et al., 2001).
The camel tick larvae SOD isoenzymes TLSOD1, TLSOD2 and TLSOD3displayed their optimumpHat 7.6, 8.0 and 7.8 (Fig. 7). Similarly, the op-timum pH of the SOD was found at pH 7.8 in mung bean and corn(Giannopolitis and Rios, 1977), at pH 7.5–8.5 in yeast (Nedeva et al.,2009), at pH 7.0 in garlic (Liu et al., 2011), and at pH 8.0 in pearl millet(Babitha et al., 2002) and tea (Vyas and Kumar, 2005).
CuCl2, ZnCl2, NiCl2 and CoCl2 increased the activity of TLSOD2 andTLSOD3, while MnCl2 increased the activity of TLSOD1. FeCl2 andMgCl2 inhibited the activity of the three isoenzymes (Table 2). These re-sults emphasize the requirement of Cu+2 and Zn+2 for TLSOD2 andTLSOD3 activities and Mn+2 for TLSOD1 activity. ZnCl2 is required forthe SOD activity from Marinomonas sp. bacteria (Zheng et al., 2006)and from the muscle tissue of the shrimp M. nipponense (Yao et al.,2004).
The SOD types can be identified by their susceptibility to inhibi-tion by potassium cyanide (KCN) and hydrogen peroxide (H2O2)(Babitha et al., 2002; Yao et al., 2004; Ken et al., 2005; Wang et al.,2011, 2012). The presence of copper and zinc ions at the active siteof the enzyme was confirmed on the basis of inhibition by KCN andH2O2 (Chen et al., 2000).
In this study, KCN is found to be the potent inhibitor of the activity ofTLSOD2 and TLSOD3, while a remarkable resistance of the TLSOD1 iso-enzyme was detected toward it. Although the three isoenzymes wereinhibited by Sodium azide (NaN3) and Sodium dodecyl sulfate (SDS),the isoenzyme TLSOD1 was the most sensitive to both inhibitors(Table 3). It was reported that, copper/zinc containing isoenzymes are

227M.A. Ibrahim et al. / Comparative Biochemistry and Physiology, Part B 164 (2013) 221–228
sensitive to both KCN and H2O2 (Hoffner and Coleman, 1973; Chou andTan, 1990; Yun and Lee, 2003; Sheng et al., 2004; He et al., 2008). Theisoenzyme which resists the inhibition with KCN and H2O2 andinactivated with sodium dodecyl sulfate is a manganese containing iso-enzyme (Kawaguchi et al., 1989; Rhie et al., 1999; Osatomi et al., 2001;Yao et al., 2004; Vyas and Kumar, 2005; Yao et al., 2007). Azide inhibitsthe enzyme activity in the following orderMn-SOD>Cu/Zn-SOD (Misraand Fridovich, 1978; Chen, et al., 2000; Haddad and Yuan, 2005). In thisreport, the presented data confirm that TLSOD2 and TLSOD3 are copper/zinc containing SOD isoenzymes and TLSOD1 is manganese containingSOD isoenzyme.
EDTA and 1, 10-phenanthroline inhibited the three SODs indicatingthat all are metalloenzymes. β-Mercaptoethanol and dithiothreitolinhibited the TLSOD activity which indicates that thiol groups are in-volved in the isoenzymes structure and play an important role fortheir activity similar to SOD from Radix lethospermi seed (Haddad andYuan, 2005). The inhibition of camel tick larvae SODs with K2Cr2O7
was probably due to the strong oxidizing power of K2Cr2O7 that maycause oxidation of metal prosthetic groups in the enzyme that are im-portant to enzyme activity and this is consistentwith SOD ofmuscle tis-sue of the shrimpM. nipponense (Yao et al., 2004).
In conclusion, this study is the first report on SOD from camel tickembryonic and larval stages. It presents a simple and convenientmethod for the purification of two Cu/Zn-SODs and one Mn-SODfrom the camel tick larvae. These SOD isoenzymes might be essentialfor avoiding oxidative damage generated by reactive oxygen speciesor by the wide use of pesticides. Therefore, targeting these SOD isoen-zymes might be an important tool to develop new control methodsfor the rapid increase in pesticide-resistant tick populations.
References
Aydemir, T., Tarhan, L., 2001. Purification and partial characterisation of superoxidedismutase from chicken erythrocytes. Turk. J. Chem. 25, 451–459.
Babitha, M.P., Prakash, H.S., Shetty, H.S., 2002. Purification and partial characterizationof manganese superoxide dismutase from downy mildew resistant pearl milletseedlings inoculated with Sclerospora graminicola. Plant Sci. 163, 917–924.
Bao, Y., Li, L., Xu, F., Zhang, G., 2009. Intracellular copper/zinc superoxide dismutasefrom bay scallop Argopecten irradians: its gene structure, mRNA expression and re-combinant protein. Fish Shellfish Immunol. 27, 210–220.
Bartkowiak, A., Leyko,W., Fried, R., 1978. A comparative characterization of cytosolic super-oxide dismutase from hog liver and erythrocytes. Comp. Biochem. Physiol. B 62, 61–66.
Bittencourt, S.E.T., Castro, L.A., Farias, S.E., Bao, S.N., Schrank, A., Vainstein, M.H., 2004.Purification and ultrastructral localization of copper/zinc superoxide dismutasefrom entomopathogenic and a caricide fungus Metarhizium anisopliae. Res.Microbiol. 155, 681–687.
Bowman, A.S., Dillwith, J.W., Sauer, J.R., 1996. Tick salivary prostaglandins: presence,origin and significance. Parasitol. Today 12, 388–396.
Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgramquantities of protein utilizing the principle of protein-dye binding. Anal. Biochem.72, 248–254.
Chen, J.R., Weng, C.N., Ho, T.Y., Cheng, I.C., Lai, S.S., 2000. Identification of the copper–zinc superoxide dismutase activity in Mycoplasma hyopneumoniae. Vet. Microbiol.73, 301–310.
Chou, F.I., Tan, S.T., 1990. Manganese (II) induces cell division and increases in super-oxide dismutase and catalase activities in an aging Deinococcal culture. J. Bacteriol.172, 2029–2035.
Davey, R.B., George, J.E., 1998. In vitro and in vivo evaluations of a strain of Boophilusmicroplus (Acari: Ixodidae) selected for resistance to permethrin. J. Med. Entomol.35, 1013–1019.
Ekanayake, P.M., Kang, H.S., Zyosa, M.D., Jee, Y., Lee, Y.H., Lee, J., 2006. Molecular clon-ing and characterization of Mn-superoxide dismutase from disk abalone (Haliotisdiscus discus). Comp. Biochem. Physiol. B 145, 318–324.
Freitas, D.R.J., Rosa, R.M., Moraes, J., Campos, E., Logullo, C., Da Silva, V.J.I., Masuda, A.,2007. Relationship between glutathione S-transferase, catalase, oxygen consump-tion, lipid peroxidation and oxidative stress in eggs and larvae of Boophilusmicroplus (Acarina: Ixodidae). Comp. Biochem. Physiol. A 146, 688–694.
Ghosh, K., Tyagi, N., Kumar, P., Singh, U., Goel, N., 2010. Stabilization ofMn (II) andMn (III)inmononuclear complexes derived from tridentate ligandswith N2O donors: synthe-sis, crystal structure, superoxide dismutase activity and DNA interaction studies.J. Inorg. Biochem. 104, 9–18.
Giannopolitis, C.N., Rios, S.K., 1977. Superoxide dismutases: purification and quantita-tive relationship and soluble protein in seedlings. Plant Physiol. 59, 315–318.
Haddad, N., Yuan, Q., 2005. Purification and some properties of Cu/Zn superoxidedismutase from Radix lethospermi seed, kind of Chinese traditional medicine.J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 818, 123–131.
Halliwell, B., Gutteridge, J.M.C., 1999. Free Radicals in Biology and Medicine. OxfordUniversity Press, New York.
Hamilton, A.J., Holdom, M.D., 1997. Biochemical comparison of the Cu/Zn superoxidedismutases of Cryptococcus neoformans var. neoformans and Cryptococcusneoformans var. gattii. Infect. Immun. 65, 488–494.
He, N., Li, Q., Sun, D., Ling, X., 2008. Isolation, purification and characterization of super-oxide dismutase from garlic. Biochem. Eng. J. 38, 33–38.
Hemingway, J., 2000. The molecular basis of two contrasting metabolic mechanisms ofinsecticide resistance. Insect Biochem. Mol. Biol. 30, 1009–1015.
Hoffner, E.K., Coleman, J.E., 1973. Cu (II)-carbon binding in cyanide complexes of cop-per enzymes. J. Biol. Chem. 248, 6626–6629.
Holmblad, T., Söderhäll, K., 1999. Cell adhesion molecules and antioxidative enzymesin a crustacean, possible role in immunity. Aquaculture 172, 111–123.
Kawaguchi, T., Nojh, S., Uda, T., 1989. A monoclonal antibody against COOH-terminalpeptide of human liver manganese superoxide dismutase. J. Biol. Chem. 264,5762–5767.
Ken, C.F., Lee, C.C., Duan, K.J., Lin, C.T., 2005. Unusual stability of manganese superoxidedismutase from a new species, Tatumella ptyseosct: its gene structure, expressionand enzyme properties. Protein Expr. Purif. 40, 42–50.
Kostaropoulos, I., Papadopoulos, A.I., Metaxakis, A., Boukouvala, E., Papadopoulou-Mourkidou, E., 2001. Glutathione S-transferase in the defence against pyrethroidsin insects. Insect Biochem. Mol. Biol. 31, 313–319.
Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head ofbacteriophage T4. Nature 227, 680–685.
Liu, J., Wang, J., Yin, M., Zhu, H., Lu, J., Cui, Z., 2011. Purification and characterization ofsuperoxide dismutase from garlic. Food Bioprod. Process. 89, 294–299.
McCord, J.M., Fridovich, I., 1969. Superoxide dismutase: an enzymic function forerythrocuprein (hemocuprein). J. Biol. Chem. 244, 6049–6055.
Miao, L., Daret, K., Clair, S., 2009. Regulation of superoxide dismutase genes: implica-tions in disease. Free Radic. Biol. Med. 47, 344–356.
Misra, H.P., Fridovich, I., 1978. Inhibition of superoxide dismutase by azide. Arch.Biochem. Biophys. 189, 317–322.
Mruk, D.D., Silvestrini, B., Mo,M., Cheng, C.Y., 2002. Antioxidant superoxide dismutase— areview: its function, regulation in the testis, and role in male fertility. Contraception65, 305–311.
Nedeva, T., Dolashka-Angelova, P., Moshtanskab, V., Voelter, W., Petrova, V.,Kujumdzieva, A., 2009. Purification and partial characterization of Cu/Zn superoxidedismutases from Kluyveromyces marxianus yeast. J. Chromatogr. B 877, 3529–3536.
O'Farrell, P.H., 1975. High resolution two-dimensional electrophoresis of proteins.J. Biol. Chem. 250, 4007–4021.
Osatomi, K., Masuda, Y., Hara, K., Ishihara, T., 2001. Purification, N-terminal amino acidsequence and some properties of Cu/Zn superoxide dismutase from Japaneseflounder (Paralichthys olivaceus) hepato-pancreas. Comp. Biochem. Physiol. B128, 751–760.
Oztürk-Urek, R., Tarhan, L., 2001. Purification and characterization of superoxidedismutase from chicken liver. Comp. Biochem. Physiol. B 128, 205–212.
Palma, J.M., Pastori, G.M., Bueno, P., Distefano, S., Delrio, L.A., 1997. Purification andproperties of cytosolic copper/zinc superoxide dismutase from watermelonCitrullus vulgaris shard cotyledons. Free Radic. Res. Commun. 26, 83–91.
Qin, X., Zhang, M., Qin, J., Yuan, S., Hou, Y., Liu, J., 2012. Two-step purification of Cu, Zn-superoxide dismutase from pumpkin (Cucurbita moschata) pulp. Sep. Purif.Technol. 87, 79–83.
Rhie, G., Hwang, C.S., Brady, M.J., Kim, S.T., Kim, Y.R., Huh, W.K., Baek, Y.N., Lee, B.H.,Lee, J.S., Kang, O.S., 1999. Manganese-containing superoxide dismutase and itsgene from Candida albicans. Biochim. Biophys. Acta 1426, 409–419.
Rikans, L.E., Hornbrook, K.R., 1997. Lipid peroxidation, antioxidant protection andaging. Biochim. Biophys. Acta 1362, 116–127.
Sauer, J.R., McSwain, J.L., Bowman, A.S., Essenberg, R.C., 1995. Tick salivary gland phys-iology. Annu. Rev. Entomol. 40, 245–267.
Sheng, L., Zheng, X., Tong, H., Liu, S., Du, J., Liu, Q., 2004. Purification and characteriza-tion of cytosolic isoenzyme III of Cu/Zn superoxide dismutase from tobacco leave.Plant Sci. 167, 1235–1241.
Sies, H., 1993. Strategies of antioxidant defence. Eur. J. Biochem. 215, 213–219.Smith, I., 1969. Acrylamide gel disc electrophoresis. In: Smith, I. (Ed.), Electrophoretic
Techniques. Academic Press, New York, pp. 365–515.Terevinto, A., Ramos, A., Castroman, G., Cabrera, M., Saadoun, A., 2010. Oxidative status,
in vitro iron-induced lipid oxidation and superoxide dismutase, catalase and gluta-thione peroxidase activities in rhea meat. Meat Sci. 84, 706–710.
Törö, I., Petrutz, C., Pacello, F., Orazio, M.D., Battistoni, A., Carugo, K.D., 2009. Structuralbasis of heme binding in the Cu/Zn superoxide dismutase from Haemophilusducreyi. J. Mol. Biol. 386, 406–418.
Ubuka, T., Masuoka, N., Yoshida, S., Ishino, K., 1987. Determination of isoelectric pointvalue of 3-Mercaptopyruvate sulfurtransferase by isoelectric focusing using ribo-nuclease A-glutathione mixed disulfides as standards. Anal. Biochem. 167,284–289.
Vig, E., Gabrielak, T., Leyko, W., Nemcsok, J., Matkovics, B., 1989. Purification and char-acterization of Cu/Zn superoxide dismutase from common carp liver. Comp.Biochem. Physiol. B 94, 395–397.
Vyas, D., Kumar, S., 2005. Purification and partial characterization of a low temperatureresponsive Mn-SOD from tea (Camellia sinensis (L.) O. Kuntze). Biochem. Biophys.Res. Commun. 329, 831–838.
Wang, Y., Osatomi, K., Nagatomo, Y., Yoshida, A., Hara, K., 2011. Purification,molecular clon-ing, and some properties of a manganese-containing superoxide dismutase fromJapanese flounder (Paralichthys olivaceus). Comp. Biochem. Physiol. B 158, 289–296.
Wang, S., Shao, B., Liu, S., Ye, X., Rao, P., 2012. Purification and characterization of Cu,Zn-superoxide dismutase from black soybean. Food Res. Int. 47, 374–379.

228 M.A. Ibrahim et al. / Comparative Biochemistry and Physiology, Part B 164 (2013) 221–228
Weber, K., Osborn, M., 1969. The reliability of molecular weight determinations bydodecyl sulfate-polyacrylamide gel electrophoresis. J. Biol. Chem. 244, 4406–4412.
Weisiger, R.A., Fridovich, I., 1973. Superoxide dismutase: organelle specificity. J. Biol.Chem. 248, 3582–3592.
Whittaker, J.W., 2010. Metal uptake by manganese superoxide dismutase. Biochim.Biophys. Acta 1804, 298–307.
Yao, C., Wang, A., Wang, W., Sun, R., 2004. Purification and partial characterization ofMn superoxide dismutase from muscle tissue of the shrimp Macrobrachiumnipponense. Aquaculture 241, 621–631.
Yao, C., Wang, A., Wang, Z., Wang, W., Sun, R., 2007. Purification and partial character-ization of Cu, Zn superoxide dismutase from haemolymph of oriental river prawnMacrobrachium nipponense. Aquaculture 270, 559–565.
Yun, Y.S., Lee, Y.N., 2003. Production of superoxide dismutase by Deinococcusradiophilus. J. Biochem. Mol. Biol. 36, 282–287.
Zheng, Z., Jiang, Y.H., Miao, J.L., Wang, Q.F., Zhang, B.T., Li, G.Y., 2006. Purification andcharacterization of a cold-active iron superoxide dismutase from a psychrophilicbacterium, Marinomonas sp. NJ522. Biotechnol. Lett. 28, 85–88.





























