Embed Size (px)
Citation preview

i
STUDIES ON WILDLIFE FORENSICS TO DETECT SPECIES
ORIGIN OF SAMBAR (Cervus unicolor) MEAT
By
K. VENKAT RAGAVAN B.V.Sc.
THESIS SUBMITTED TO THE
ACHARYA N.G. RANGA AGRICULTURAL UNIVERSITY
IN PARTIAL FULFILMENT OF THE REQUIREMENTS
FOR THE AWARD OF THE DEGREE OF
MASTER OF VETERINARY SCIENCE
IN THE FACULTY OF VETERINARY SCIENCE
DEPARTMENT OF VETERINARY PUBLIC HEALTH
COLLEGE OF VETERINARY SCIENCE, TIRUPATI
ACHARYA N.G. RANGA AGRICULTURAL UNIVERSITY
HYDERABAD
JUNE, 2003

ii
CERTIFICATE
Mr. K. VENKAT RAGAVAN has satisfactorily prosecuted the course of
research and that the thesis entitled “STUDIES ON WILDLIFE FORENSICS TO
DETECT SPECIES ORIGIN OF SAMBAR (Cervus unicolor) MEAT”
submitted is the result of original research work and is of sufficiently high standard
to warrant its presentation to the examination. I also certify that the thesis or part
thereof has not been previously submitted by him for a degree of any University.
Date : (Dr. S. UMAMAHESWARA RAO)
Major Advisor
Professor & University Head
Department of Veterinary Public Health
College of Veterinary Science
Tirupati – 517 502 (A.P)

iii
CERTIFICATE
This is to certify that the thesis entitled “STUDIES ON WILDLIFE
FORENSICS TO DETECT SPECIES ORIGIN OF SAMBAR (Cervus unicolor)
MEAT” submitted in partial fulfilment of the requirements for the degree of
MASTER OF VETERINARY SCIENCE of the Acharya N.G. Ranga Agricultural
University, Hyderabad, is a record of the bonafide research work carried out by Mr.
K. VENKAT RAGAVAN under my guidance and supervision. The subject of the
thesis has been approved by the Student’s Advisory Committee.
No part of the thesis has been submitted for any other degree or diploma. The
published part has been fully acknowledged. All assistance and help received during
the course of investigation have been duly acknowledged by the author of the thesis.
(Dr. S. UMAMAHESWARA RAO)
Chairman of the Advisory Committee
Thesis approved by the Student’s Advisory Committee
Chairman : (Dr. S. UMAMAHESWARA RAO) ______________
Professor and University Head
Department of Veterinary Public Health
College of Veterinary Science
Tirupati – 517 502
Member : (Dr. P. MASTHAN REDDY) ______________
Professor and Head
Department of Livestock Products Technology
College of Veterinary Science
Tirupati – 517 502
Member : (Dr. I. SANKARA REDDY) ______________
Associate Professor in Dairy Microbiology
Dairy Technology Programme
College of Veterinary Science
Tirupati – 517 502

iv
LIST OF CONTENTS
CHAPTER
NUMBER
TITLE PAGE
NUMBER
I 1. INTRODUCTION 1
II 2. REVIEW OF LITERATURE 5
2.1 Anatomical and histological methods 5
2.2 Physical, chemical and biochemical methods 6
2.3 Sodium dodecyl sulphate – Poly Acrylamide Gel
Electrophoresis (SDS-PAGE)
6
2.4 Immunochemical tools 8
2.4.1 Double Immuno Diffusion (DID) test 9
2.4.2 Single Radial Immuno Diffusion (SRID) test 14
2.4.3 Immuno Electrophoresis (IE) 15
2.4.4 Counter Immuno Electrophoresis (CIE) 16
2.5 Enzyme Linked Immuno Sorbent Assay (ELISA) 17
2.6 Western blot technique (Electroblot Immuno Assay) 19
2.7 Iso Electric Focusing (IEF) 19
2.8 Other methods 21
2.8.1 DNA probe Technology 21
2.8.2 Polymerase Chain Reaction (PCR) 22
2.8.3 Wildlife Forensics – Micro satellite analysis of
DNA
22
III 3 MATERIALS AND METHODS 24
3.1 Preparation of sambar muscle protein antigens 24
3.1.1 Collection and processing of sambar flesh 24
3.1.1.1 Materials 24
3.1.1.2 Method 25
1. Determination of protein concentration 25
2. Standardization of the method of processing for
antigen preparation
25

v
CHAPTER
NUMBER
TITLE PAGE
NUMBER
3. Processing of sambar muscle 25
3.2 Preparation of fresh and thermostable muscle
protein antigens (TMP) of field samples (cattle,
buffalo, sheep, goat, pig and rabbit)
28
3.3 Production and preparation of hyperimmune sera to
sambar muscle protein antigens
29
3.3.1 Materials 29
3.3.2 Method 29
1. Immunization of rabbits with fresh muscle protein
antigens of sambar
29
2. Immunization of rabbits with thermostable muscle
protein antigens (TMP) of sambar
30
3. Immunization of sheep with fresh muscle antigens
of sambar
31
4. Immunization of sheep with thermostable muscle
protein (TMP) antigen of sambar
31
3.4 Preparation of binary meat mixtures 32
3.5 Immunochemical tools to detect wildlife sambar
meat
33
3.5.1 Double Immuno Diffusion (DID) test 33
3.5.1.1 Materials 33
3.5.1.2 Method 33
1. Pre-coating of the slides 33
2. Preparation and casting of the gel 34
3. Washing drying and staining of the gels 34
3.5.2 Single Radial Immuno Diffusion (SRID) technique 35
3.5.2.1 Materials 35
3.5.2.2 Method 35
3.5.3 Counter Immuno Electrophoresis (CIE) 37

vi
CHAPTER
NUMBER
TITLE PAGE
NUMBER
3.5.3.1 Materials 37
3.5.3.2 Method 37
3.6 Sodium Dodecyl Sulphate Poly Acrylamide Gel
Electrophoresis (SDS – PAGE))
38
3.6.1 Preparation of reagents 38
1. Acrylamide Bis acrylamide stock solution (100ml) 38
2. Resolving gel buffer (1.5m Tris – HCl pH 8.8) 39
3. Stacking gel buffer (0.5m Tris HCl buffer pH 6.8) 39
4. 10% Sodium Dodecyl Sulphate (SDS) 39
5. Sample Buffer 39
6. Tank Buffer 40
7. 10% Ammonium per sulphate 40
8. Staining solution 40
9. Destaining solution 40
3.6.2 Gel preparation 41
3.6.3 Gel preparation procedure 41
3.6.4 Sample preparation 42
3.6.4.1 Fresh antigen 42
3.6.4.2 Heated antigen 42
3.6.5 Sample loading 42
3.6.6 Staining and destaining 44
3.6.7 Molecular weight determination 44
3.7 Electroblot immuno assay (semi–dry) 45
3.7.1 Materials 45
3.7.1.1 Preparation of Buffers 45
1. Anode buffer I, pH 10.4 45
2. Anode buffer II, pH 10.4 45
3. Cathode buffer, pH 9.4 46
4. Tris buffered saline (TBS), pH 7.5 46

vii
CHAPTER
NUMBER
TITLE PAGE
NUMBER
5. TBS – Tween 20 (TBS – T) 46
6. Blocking solution 47
7. Antibody buffer 47
8. HRP labeled goat anti rabbit Ig – G conjugate
1/5000)
47
9. Substrate buffer (0.5 M sodium citrate pH 5.2 for
HRP system)
47
10. Substrate solution for HRP system 47
3.7.2 Method 48
IV 4. RESULTS 51
4.1 Development, preparation and standardization of
immuno chemical reagents for identification of
wildlife sambar meat
51
4.1.1 Standardization of the method of processing for
antigen preparation and preparation of sambar
muscle protein antigens
51
4.1.2 Preparation of sambar fresh muscle protein antigens 52
4.1.3 Preparation of thermostable muscle protein (TMP)
antigen of sambar
52
4.1.4 Production and preparation of hyperimmune sera to
sambar fresh muscle protein antigens
53
4.1.4.1 Hyperimmune sera in rabbits 53
4.1.4.2 Hyperimmune sera in sheep 53
4.1.5 Production and preparation of hyperimmune sera to
thermostable muscle protein (TMP) antigens of
sambar
53
4.1.5.1 Hyperimmune sera in rabbits 53
4.1.5.2 Hyperimmune sera in sheep 54

viii
CHAPTER
NUMBER
TITLE PAGE
NUMBER
4.2 Preparation of fresh and thermostable muscle
protein (TMP) antigens of domestic species (field
samples)
54
4.3 Standardization of immunochemical methods to
detect wildlife sambar meat (fresh muscle antigen)
57
4.3.1 Standardization with rabbit antisera 57
4.3.1.1 Double Immuno Diffusion (DID) test 57
4.3.1.2 Counter Immuno Electrophoresis (CIE) 57
4.3.2 Standardization with sheep antisera 58
4.3.2.1 Double Immuno Diffusion (DID) test 58
4.3.2.2 Counter Immuno Electrophoresis (CIE) 58
4.3.2.3 Comparison of species of animals for optimum
responsiveness in the antibody production against
sambar fresh muscle antigen
58
4.4 Standardization of immunochemical methods to
detect wildlife sambar meat (thermostable muscle
protein antigen)
63
4.4.1 Standardization with rabbit antisera 63
4.4.1.1 Double Immuno Diffusion (DID) test 63
4.4.1.2 Single Radial Immuno Diffusion (SRID) test 63
4.4.1.3 Counter Immuno Electrophoresis (CIE) 64
4.4.2 Standardization with sheep antisera 64
4.4.2.1 Double Immuno Diffusion (DID) test 64
4.4.2.2 Single Radial Immuno Diffusion (SRID) test 65
4.4.2.3 Counter Immuno Electrophoresis (CIE) 65
4.4.2.4 Comparison of species of animals for optimum
responsiveness in the antibody production against
sambar thermostable muscle protein (TMP) antigen
65

ix
CHAPTER
NUMBER
TITLE PAGE
NUMBER
4.5 Determination of protein profile of muscle protein
of sambar and different species of animals and
sambar by SDS – PAGE
67
4.5.1 Comparative analysis of SDS – PAGE of fresh
muscle proteins of sambar and different species of
domestic animals
67
4.5.2 Comparative analysis of SDS – PAGE of TMP
antigens of sambar and different species of animals
69
4.5.3 Comparison of the protein profile of fresh and TMP
antigens of sambar muscle protein
69
4.5.4 Comparison of the protein profile of fresh and TMP
antigen of other domestic animals
70
4.6 Electroblot immunoassay (western blotting) of
cooked meat (TMP antigens) of sambar and other
domestic species
70
4.7 Comparison of SDS – PAGE analysis and western
blotting of TMP antigens of sambar and other
domestic species
72
4.8 Detection of adulterated wildlife (sambar) meat
with the meat of domestic species by Immuno
Diffusion and Immuno Electrophoresis
73
4.8.1 Detection of sambar fresh meat adulterated with the
meat of domestic species
73
4.8.1.1 Detection with hyperimmune sera raised in rabbits 73
4.8.1.1.1 Double Immuno Diffusion (DID) test 73
4.8.1.1.2 Counter Immuno Electrophoresis (CIE) 76
4.8.1.2 Detection with hyperimmune sera raised in sheep 76
4.8.1.2.1 Double Immuno Diffusion (DID) test 76
4.8.1.2.2 Counter Immuno Electrophoresis (CIE) 77

x
CHAPTER
NUMBER
TITLE PAGE
NUMBER
4.8.2 Detection of sambar cooked meat (TMP)
adulterated with the meat of domestic species
77
4.8.2.1 Detection with hyperimmune sera raised in rabbits
against sambar TMP antigens
77
4.8.2.1.1 Double Immuno Diffusion 77
4.8.2.1.2 Single Radial Immuno Diffusion (SRID) test 78
4.8.2.1.3 Counter Immuno Electrophoresis (CIE) 78
4.8.2.2 Detection with hyperimmune sera raised in sheep
against sambar TMP antigens
80
4.8.2.2.1 Double Immuno Diffusion (DID) test 80
4.8.2.2.2 Single Radial Immuno Diffusion (SRID) test 80
4.8.2.2.3 Counter Immuno Electrophoresis 80
4.9 Detection of sambar meat adulterated with the meat
of domestic animals by electroblot immunoassay
(western blotting)
83
4.10 Detection of sambar meat adulterated with buffalo
meat by Electroblot immunoassay (western blotting)
83
V 5 DISCUSSION 84
VI 6 SUMMARY 101
LITERATURE 108
APPENDICES 118

xi
LIST OF ILLUSTRATIONS
FIGURE
NUMBER
DESCRIPTION
PAGE
NUMBER
1. Schematic flow diagram for preparation of fresh sambar
muscle protein antigens
26
2. Schematic flow diagram for preparation of thermostable
muscle protein (TMP) antigens of sambar
27
3. Arrangement of sheets in semidry blot unit 48
4. Photograph of the DID reactions comparing the
precipitation reaction of sambar fresh muscle antigen
against rabbit anti sambar sera and sheep anti sambar
sera. Peripheral wells contain I rabbit anti sambar sera
and II sheep anti sambar sera while the central wells
contains sambar muscle (fresh) antigen.
55
5. CIE of sambar muscle (Fresh) antigen. 56
6. Photograph showing the negative precipitation reaction
between cooked muscle extracts of sambar and domestic
animals against rabbit anti sambar serum.
59
7. Photograph showing the precipitation reaction of
thermostable muscle protein antigen of sambar against
sheep anti sambar (heated) sera.
60
8. Standardization of SRID test for sambar Thermostable
muscle protein (TMP) antigen perceptible ring diameters
of precipitation reaction was noticed only with 50%
rabbit anti sambar sera incorporated in the gel.
61
9. CIE of Thermostable muscle protein (TMP) antigen of
sambar.
62
10. SDS – PAGE analysis of fresh muscle antigens of
sambar and other domestic animals muscle samples.
66

xii
FIGURE
NUMBER
DESCRIPTION PAGE
NUMBER
11. SDS – PAGE analysis of Thermostable muscle protein
(TMP) antigen of sambar and other domestic muscle
samples.
68
12. Immunoblot analysis of Thermostable muscle protein
(TMP) of antigens of sambar and other domestic muscle
heated samples.
71
13. Photograph showing the precipitation reaction between
rabbit anti sambar (fresh) serum and sambar muscle
(fresh) antigen experimentally adulterated with buffalo
muscle (fresh) extract.
74
14. CIE of sambar muscle (fresh) extract experimentally
adulterated with buffalo muscle (fresh) extract.
75
15. CIE of Thermostable muscle protein (TMP) antigen of
sambar experimentally adulterated with buffalo muscle
(treated) extract.
79
16. Immunoblot analysis of 1% adulterated Thermostable
muscle protein (TMP) of sambar with other domestic
species muscle extracts.
81
17. Immunoblot analysis of adulterated Thermostable muscle
protein (TMP) antigen of sambar with Buffalo muscle
extract.
82

xiii
LIST OF TABLES
TABLE
NUMBER
TITLE PAGE
NUMBER
1 Preparation of binary meat mixtures 32
2 Protein concentration of fresh muscle extracts of
different species
52
3 Comparative studies on protein concentration of fresh
and heated muscle extracts (TMP) of different species
54

xiv
ACKNOWLEDGEMENTS
I avail this unique opportunity with pride to express my whole
hearted indebtedness to my guide Dr. S. Umamaheswara Rao Ph.D.,
Professor and University Head, Department of Veterinary Public
Health, College of Veterinary Science, Tirupati for his inspiring and
able guidance and unfailing support during the course of this research
and throughout my M.V.Sc programme.
I have immense pleasure in expressing my heartfelt gratitude to
the member of the advisory committee, Dr. P. Masthan Reddy,
Professor and Head, Department of Livestock Products Technology,
College of Veterinary Science, Tirupati for rendering his genuine
cooperation and help during the course of this investigation.
I am highly indebted to Dr. I. Sankara Reddy, Associate
Professor in Dairy Microbiology, Dairy Technology Programme,
College of Veterinary Science, Tirupati for acting as a member of the
advisory committee and for his marked advice while going through the
manuscript of the thesis.
My sincere thanks are due to Dr. A.Seshagiri Rao, Associate
Dean , College of Veterinary Science, Tirupati and to Dr.C.R.K
Reddy, then Associate Dean, College of Veterinary Science,
Tirupati for providing necessary facilities to carry out the present
investigation.

xv
My grateful thanks are due to Sri. K. Subba Rao, Principal Chief
Conservator of Forests, Govt. of Andhra Pradesh for giving kind
permission to collect sambar flesh. I am extremely thankful to Sri E.
Narasimhulu, I.F.S Curator, S V Zoological Park Tirupati for giving
necessary permission to collect sambar flesh for my research work.
My sincere thanks are due to Dr. Ramana, Zoo Veterinarian
Sri Venkateswara Zoological Park ,Tirupati without whose willing
co-operation and help, the collection of required sambar flesh for the
current investigation would not have been possible.
With deep sense of gratitude I acknowledge the help and
cooperation received from Dr. D.V.R. Sai Gopal, Ph.D., Department
of Virology, S.V. University, Tirupati for his help, whole hearted
cooperation and for his kind permission to avail the facilities in his
department.
I am highly thankful to Sri M. R. L. Prabhu, Head of the
Department of Biochemistry, College of Veterinary Science,
Tirupati for his encouragement, support and cooperation.
I am extremely thankful to Dr. V.H Rao, Associate Professor
and Head, Department of Animal Physiology, College of Veterinary
Science, Tirupati for kindly providing experimental rabbits for my
research work.
I am very much thankful to Dr. J. Rama Prasad, Professor and
Head, Department of Animal Nutrition, College of Veterinary
Science, Tirupati for his whole hearted cooperation and for kindly
providing experimental sheep for my research.

xvi
I am extremely thankful to Dr. P.K. Sriraman Ph.D., Head
(Rtd), Department of Pathology, College of Veterinary Science,
Hyderabad for his invaluable help and for taking excellent photographs
of my research work.
I Wholeheartedly express my grateful thanks to Sree N. Bhaskara
Reddy, M.Lib.Science, Librarian, ANGRAU Regional Library,
Tirupati for his help and cooperation.
I sincerely thank Dr. A. Jagdeesh Babu, M.V.Sc., and Dr. T.
Srinivasa Rao, M.V.Sc., Assistant Professors, Department of
Veterinary Public Health College of Veterinary Science, Tirupati for
their affectionate encouragement, help and cooperation.
I express my heartfelt gratitude to Mr. Venkata Prasanna,
Research Scholar, Department of Virology, S.V. University, Tirupati
for his technical help, immense support and encouragement and I
thankfully acknowledge the invaluable help and affection extended by
my colleagues and friends, Tamilmani, Rajarajan, Manigandan,
Anandaraja, Sarvana Kumar, Siva, Kumareshan, Upendra.
I have no words to express my heartful thanks to my father Sri.
M. Kandasamy, mother Smt. K. Baby Rani and sister Miss.
Premalatha and Mrs. R. Jayachitra for their inspiration, moral
support, everlasting affection and encouragement during the course of
my study.
I express my sincere thanks to Mr. V. Chandra Sekhar for his
excellent typing of the thesis.
(K. VENKAT RAGAVAN)

xvii
Name of the author : K. VENKAT RAGAVAN
Title of the thesis : “STUDIES ON WILDLIFE FORENSICS
TO DETECT SPECIES ORIGIN OF
SAMBAR (Cervus unicolor) MEAT”
Degree to which it is
submitted : MASTER OF VETERINARY SCIENCE
Faculty : VETERINARY SCIENCE
Department : VETERINARY PUBLIC HEALTH
Major Advisor : Dr. S. UMAMAHESWARA RAO BVSc., MSc(Vety.)., Ph.D.
Professor and University Head
Department of Veterinary Public Health
College of Veterinary Science
Tirupati – 517 502
University : Acharya N.G. Ranga Agricultural
University
Year of Submission : June, 2003
ABSTRACT
Attempts were made to develop suitable wildlife forensic tools
with immunologic basis to detect and identify sambar (Cervus unicolor)
meat in the experimentally adulterated meat samples of domestic animal
species viz., buffalo, cattle, sheep, goat, pig and rabbit both fresh and
cooked (thermostable muscle protein (TMP)) meat.

xviii
PBS, pH 7.3 was found to be a suitable diluent for preparation of
antigens both from fresh and cooked meat of sambar, buffalo, cattle,
sheep, goat, pig and rabbit.
Two kinds of experimental animals viz. rabbit and sheep were
immunized with sambar muscle protein antigen both fresh and cooked
(TMP) to study their ability to develop specific antibody. The rabbit, a
distinct phylogenetic animal was found to be more suitable when
compared to sheep which is a phylogenetically related species as
evidenced by the immunocompetence of the sera to recognize and react
with TMP antigens leading to the formation of precipitation reaction.
The sambar flesh, the source of antigen for the current
investigation was collected immediately from the naturally dead sambar
(either by natural death or by accidental death without any infectious
etiology) from Sri Venkateswara Zoological Park, Tirupati.
The conventional immunochemical methods like DID and CIE
tests were used to detect wildlife sambar meat, both fresh and cooked
(TMP) muscle antigens, were standardized using hyperimmune sera

xix
raised in both, rabbit and sheep. The results of precipitation reaction
were quick within 6 hours of incubation for fresh muscle antigens with
rabbit and sheep antisera, while the precipitation reaction was delayed
by 72 hours for TMP antigens with rabbit antisera only but the reaction
was absent with sheep antisera.
The protein profile of sambar fresh and TMP antigens was studied
by subjecting to SDS – PAGE and a comparison was made with those of
other domestic animal species. The electrophoretogram revealed that
sambar fresh muscle protein was resolved into seven protein bands with
molecular weights of 60kd, 52kd, 44kd, 42kd, 30kd, 22kd and 12kd,
while the buffalo muscle protein profile resolved into five protein bands
with molecular weights of 60kd, 52kd, 44kd, 42kd and 22kd. The cattle
meat profile was resolved into three protein bands: 40kd, 30kd and 12kd,
while the sheep meat protein profile resolved into six bands: 60kd, 52kd,
44kd, 22kd and that of goat meat protein profile was resolved into five
protein bands: 60kd, 52kd, 44kd, 42kd and 12kd.
The electrophoretogram of TMP antigens of sambar and other
species of domestic animals like buffalo, sheep, goat and pig except
cattle showed a partially denatured two protein complex with diffused

xx
migration pattern (44kd and 42kd). The two low molecular weight
protein bands 22kd and 12kd were present only in sambar probably
species specific.
The electroblot immuno assay (western blot) technique could
identify serologically related TMP antigens of 44kd and 42kd of sambar,
buffalo, sheep, goat and pig. Another serologically related protein 40kd
was noticed only in cattle although not appreciated very well in sambar.
Two serologically identified proteins with molecular weight of 22kd and
12kd were present only in sambar, possibly species specific.
The tests like DID and CIE were carried out using rabbit or sheep
antiserum to detect adulteration of sambar fresh meat from binary
mixtures of fresh meat of buffalo, cattle, sheep, goat, rabbit and pig.
These tests could detect the fresh meat adulteration up to 1% level while
detection of sambar cooked meat (TMP) adulterated with cooked meats
(TMP) of domestic animals could be up to a level of 10% only with
rabbit antisera. The sheep antisera against TMP antigen of sambar did
not detect the adulteration.

xxi
Western blotting could identify cooked (TMP) sambar meat
adulterated with the cooked (TMP) meats of buffalo, cattle, sheep, goat
and pig as low as up to a level of 1%. The experimental adulteration of
cooked (TMP) sambar meat with cooked buffalo meat when subjected to
western blotting, the detection was as low as 0.1% but not at 0.05%
level.

xxii
LIST OF ABBREVIATIONS
ADS - Ammonium per Sulphate
AGIEF - Agarose Gel Iso Electric Focussing
B – C - Buffalo muscle extract – Cooked
B – F - Buffalo muscle extract – fresh
BE - Boiling resistant Ethanol
C – C - Cattle muscle extract – Cooked
C – F - Cattle muscle extract – fresh
CIE - Counter Immuno Electrophoresis
COE - Cross Over Electrophoresis
DAB - Di amino benzidine
DID - Double Immuno Diffusion
DNA - De Oxyribo Nucleic Acid
ELISA - Enzyme Linked Immuno Sorbent Assay
F – F - Fowl muscle extract – fresh
FCA - Freund’s Complete Adjuvant
G – C - Goat muscle extract – Cooked
G – F - Goat muscle extract – fresh
HA - Haem agglutination
HCl - Hydrochloric Acid
HRP - Horse Radish Peroxidase
IE - Immuno Electrophoresis
IEF - Iso Electric Focussing
Kg - Kilogram

xxiii
MW - Molecular Weight
P – C - Pig muscle extract – Cooked
P – F - Pig muscle extract – fresh
PBS - Phosphate Buffer Saline
PCR - Polymerase Chain Reaction
PVDF - Poly Vinylidine Diflouride
R – C - Rabbit muscle extract – Cooked
S – C - Sheep muscle extract – Cooked
S – F - Sheep muscle extract – fresh
SDS – PAGE - Sodium Dodecyl sulphate – Poly Acrylamide
Gel Electrophoresis
SRID - Single Radial Immuno Diffusion
TBS - Tris Buffered Saline
TBS - Tris Buffered Saline – Tween
TMP - Thermostable muscle protein
µl - Microlitre
µg - Microgram
V/V - Volume / Volume

xxiv
CHAPTER I
1. INTRODUCTION
Preservation of wildlife ecosystem and protection of wildlife
fauna and flora are imperative and uncompromised edicts to maintain
natural ecological balance.
International trade in wildlife and its products is worth around $17
billion a year (Agrawal, 2000). Unfortunately illegal trade in wildlife,
probably the world’s second largest illegitimate business, is on its
increase. In order to check this menace, a convention of international
trade in endangered species (CITES) of world flora and fauna was
brought into existence by International union for conservation of nature
and natural resources (IUCN). The government of India had also enacted
the wildlife (protection) Act 1972 to provide protection for the
endangered and threatened species and their hunting is prohibited.
However, the opening up of international market for meat foods
and also increased desire of man to taste new culinary dishes prepared
with wildlife meat may lead to fraudulent substitution of meat from
protected species of wildlife. The meat of such animal species at times
1

xxv
poached or often misrepresented for flesh of cattle, buffalo, sheep, goat
or pig.
Under such circumstances speciation of meats and meat products
is mandatory and it is therefore essential to develop an appropriate
methodology to detect the species origin of such meats in order to
contest in a court of law.
Determining the species of meat origin is an integral part of food
regulatory control with respect to economic fraudulence. Besides this,
correct species identification is important to the consumer for reasons of
specific food allergies or religious dietary restrictions.
In order to combat these problems, species identification of meats
acquired great importance and is a challenging task in food hygiene,
food quality control programmes, medico legal cases and veterinary
forensic medicine. Authentic detection of species origin of meat
however remains a nagging problem for food regulatory agencies at
national and global level.
In spite of decades research carried out in this field, the
problem still remains unsolved in the absence of a single reliable fool

xxvi
proof technique to detect such unscrupulous mixing. The potential of
immunoassay as a method of choice for replacing less specific and more
costly methods has been recently appreciated.
Sambar, black buck and other deer species are prohibited from
hunting in Andhra Pradesh as per wildlife (protection) Act, 1972 and
other reserve forest regulations. Sambar are frequent victims targeted by
poachers for their meat and hide. The meat of such animal species is
often misrepresented for flesh of cow, buffalo, sheep or goat leading to
medico legal cases (Umamaheswararao, 2003).
With scanty reported literature on wildlife forensics, a study was
under taken to develop a methodology for the detection of sambar meat
experimentally adulterated with the meat of other domestic animal
species using immunochemical techniques with the following objectives.
1. To study the efficacy of using fresh and thermostable muscle protein
(TMP) antigen of sambar to develop species specific antibodies in
rabbits and sheep in order to identify the fresh as well as cooked
sambar (Cervus unicolor) meat.

xxvii
2. To evaluate the suitability of immunochemical techniques like
double immuno diffusion (DID) test and counter immuno
electrophoresis (CIE) test etc., in identifying fresh as well as cooked
sambar muscle.
3. To apply the so developed wildlife immunochemical forensic tool to
detect sambar meat when adulterated with the meat of other domestic
animals.

xxviii
CHAPTER II
2. REVIEW OF LITERATURE
The identification of species origin of meat is not only imperative
in a given case but also the detection of level of adulteration is equally
essential for appropriate regulatory control of marketing of such meats.
Methods employed generally to detect the origin of meats are
anatomical and histological; physical, chemical and biochemical;
electrophoretic; immunochemical; enzyme linked immunosorbent assay
(ELISA); Immunoblotting and Isoelectric focusing techniques. Hence a
brief survey of the literature pertaining to these techniques would enable
the investigator to choose the most expedient procedure bearing in mind
the circumstances expected: whether the test should cover domestic
species, wild species or closely related species.
2.1 ANATOMICAL AND HISTOLOGICAL METHODS :
The species identification of meats based on anatomical and
histological methods such as length of carcass, ribs on the thorax and the
size of the muscle fibers is not always useful in the detection of
adulteration and lacks authenticity especially in forensic cases. 5

xxix
2.2 PHYSICAL, CHEMICAL AND BIOCHEMICAL METHODS:
Colour, consistency and texture of meats were among the physical
characteristics used for differentiation of meats which are highly variable
within same species (Ginsberg, 1948 and Krishna, 1998).
According to Sherikar et al (1988) chemical tests for species
identification of meats such as estimation of body fats, refractive index,
Iodine number, carotene content , fatty acid composition, linoleic acid
content, ratio of histidine dipeptides, glycogen content, myoglobin
content, muscle enzymes and muscle nitrogen fractions are highly
variable and cannot be used with accuracy to rely upon.
2.3 SODIUM DODECYL SULPHATE - POLY ACRYLAMIDE
GEL ELECTROPHORESIS (SDS-PAGE) :
SDS-PAGE electrophoresis was successfully applied by several
workers to study the band pattern of sarcoplasmic proteins and
myofibrillar proteins (Scopes and Penny, 1971 and Rattire and
Regenstien, 1977).
Ramadas (1972) reported species specific muscle protein band
patterns of bullock, buffalo, dog, pig and poultry while in case of sheep
and goat the band patterns were identical but different from other

xxx
species. Prasad (1974) using SDS – gel electrophoresis, established a
relationship of muscle protein band pattern of different species between
different age and sex groups.
Lawrie et al (1977) while studying the identification of meats in
fresh and processed foods by starch gel electrophoresis and PAGE,
indicated water insoluble myofibrillar and water soluble sarcoplasmic
muscle protein patterns for quantitative identification of meat proteins.
Babiker et al (1981) used a modified linear gradient poly
acrylamide gel electrophoresis technique and identified cooked horse
meat (1200C) from cooked beef on the basis of myofibrilar protein
pattern.
Yamamoto et al (1982) advocated the use of SDS – PAGE for
the detection of various non meat extenders in meat products.
Govindarajulu (1982) differentiated beef from mutton on the basis
of species specific electrophoretic pattern on starch gel electrophoresis.
Anneli Skrokki and Osmo Harmi (1994) quantitatively
differentiated beef, pork, venison, reindeer and mutton and detected
adulterated minced beef with pork by employing modified PAGE.

xxxi
Nath et al (1999) by employing SDS – PAGE made it possible to
identify the meats of sheep, goat, cattle, buffalo, pig, chicken based on
total number of protein bands, their relative position and concentration,
ionic charges together with differences in RF values.
2.4 IMMUNOCHEMICAL TOOLS :
The remarkable specificity of serological reactions was amply
demonstrated by Landsteiner (1945). Various immunochemical methods
adopted by different workers gave promising results in the identification
of species origin of meats.
Fresh meat antigens can be successfully used to produce species
specific antibodies for the identification of various meats. But the
identification of cooked meats through serological means will be
difficult as the muscle protein may get denatured during heating. The
main disadvantages of immunochemical techniques were cross reactions
among related species and failure to identify the origin of cooked meats
(Krishnamurthy, 1998).
2.4.1 Double Immuno Diffusion test (DID) :

xxxii
The application of double immuno diffusion (DID) or Agar gel
precipitation test (AGPT) was first carried out by Pike and Sulkin (1957)
and they could detect the adulteration of beef with horse meat.
Fugate and Shelton (1971) employed immuno diffusion technique
for the detection of mixed muscle samples.
Ramadas (1972) observed that agar gel immuno diffusion (AGID)
technique was a reliable technique for differentiating meats of closely
related species.
Poli et al (1977) differentiated animal proteins in cooked sausages
employing agar gel diffusion method.
Pandey and Pathak (1974) could not distinguish the cow meat
from buffalo meat by agar gel diffusion test.
Beljaare and Oleman (1977) compared PAGE and DID test for
detection of non meat protein in meat products and concluded that DID
was superior over PAGE.
Flego and Bor gheese (1979) made comparison of DID with that
of cross over electrophoresis (COE) method and concluded that COE

xxxiii
was 26-45 times more sensitive than DID in the detection of pork, beef,
horse and chicken meat.
Sherikar et al. (1979) concluded that inspite of cross reactions,
DID is useful in differentiating meats of various species viz, cattle,
buffalo, sheep, goat, pig and poultry.
Hayden (1979) used myoglobin of ovine, porcine and equine
muscle as antigens for antibody production in rabbits and successfully
detected the presence of meat of these species.
Hayden (1980) could successfully employ agar gel immuno
diffusion and could quantitate the pig, horse and rabbit flesh in ground
beef using troponin and myoglobin as test proteins.
Hayden (1981) could detect mammalian adrenal antigens in beef
sausages at 5% level of adulteration while the avian kidney – adrenal
antigens were detected at 10% level of adulteration by means of agar gel
immunodiffusion technique.
Nath (1986) experimented on the use of cross immunization in
phylogenetically related species using freeze dried muscle extractives as

xxxiv
antigen and successfully detected the species origin of fresh meats by
using DID.
Sherikar et al. (1987) used extracts of raw and partially heated
(700C) muscles of buffalo, cattle, sheep, goat, pig and poultry as antigen
and raised anti rabbit antibody to identify and differentiate their raw,
heated (700C) and boiled (100
0C) meats. They observed cross reactions
between related species but anti chicken sera and antisera to heated pig
muscle extracts gave species specific reaction. Boiled (1000C) meat
extracts did not elicit any antibody production and did not react with
these antisera. Antibody to raw meats stored at -200C for 9 years were
found continue to yield similar results as that of fresh antiserum.
Bansal and Mandokhot (1988) used DID, Counter
immunoelectrophoresis (CIE), Immunoelectrophoresis (IE) and micro
passive haemagglutination (HA) test to detect beef, sheep, goat, pig and
chicken meats and proved that DID and CIE were equally sensitive and
specific and concluded that DID was more suitable because of its
simplicity and reliability.

xxxv
Sherikar et al. (1988) identified species of origin in cooked (700C)
meat mixtures of closely related species by agar gel immuno diffusion
technique using specific antisera to adrenal heat stable antigens.
Radhakrishna et al. (1988) reported immunogenic nature of
thermostable muscle protein (TMP) of goat, sheep, ox and buffalo and
the sera raised against these proteins were made monospecific by cross
absorption technique and thereby they could identify the respective TMP
as well as the extractives from raw beef and from those canned spiced
mutton curry.
Karkare et al. (1988) studied species origin of meat in
commercially processed canned meat products viz, beef, kheema, corned
beef, mutton kheema, mutton chunk curry and pork mince. Species
specific antisera was raised in rabbits to heat stable antigens of adrenal
gland of cattle, buffalo, sheep, goat and pig and conducted DID and CIE
by using saline extracts and ethanol precipitated proteins of the canned
meat products as antigens. Their results revealed that rabbit anti adrenal
sera could identify the species origin of meats in canned products while
other components in the canned meat products viz, additives, condiments
etc., did not interfere in the test reactions.

xxxvi
Suryarao (1990) conducted experiments in identifying the species
origin of meats using testicular antigens of sheep, goat, cattle and buffalo
employing serological tests including DID, IE, CIE and rocket immuno
electrophoresis technique. He could detect adulterated meat as low as at
1% level in fresh meats and at 10-20% level in cooked meat mixtures.
Reddy et al. (1990) made the antisera raised against adrenal
boiling resistant and ethanol precipitable (BE) antigens of cattle and
sheep species specific by immuno absorption and detected cooked meat
at 1% level using DID.
Saisekhar and Reddy (1995) developed monospecific antisera to
troponin of cattle and buffalo and detected fresh beef at 10% level and
fresh buffalo meat at 1% level but these sera could not detect cooked
meats.
Reddy and Reddy (1995) could detect either fresh or cooked
meats as low as at 10% level in cooked state and at 20% level in fresh
condition from a mixture of beef or cara beef or mutton using DID
technique.

xxxvii
Sasidhar (1995) developed species specific sera against native as
well as heated porcine testicular extract antigens and detected cooked
pork up to a level of 2% and fresh pork up to 0.5% level with DID and
CIE techniques.
Sridharlingareddy (1996) concluded that, of the tests employed
DID and CIE were superior to detect adulteration in fresh meats of
cattle, buffalo and pig using brain extract antigens of respective species.
2.4.2 Single Radial Immuno diffusion (SRID) test :
Suryarao (1990) conducted single radial immuno diffusion (SRID)
to find out fresh buffalo meat antigen and concluded that SRID can be
used to determine the level of adulteration.
2.4.3 Immuno Electrophoresis (IE) :
Graber and Williams (1953) were the pioneers of this technique
and they could find out the use of immunoelectrophoresis (IE) for
analyzing the quality and purity of antibody rich serum fractions of
immunized animals.

xxxviii
Durand and Schneider (1962) used IE for species identification of
cattle, buffalo, horse, sheep, goat, donkey and camel by observing
precipitation lines and their number and position.
Ramadas and Misra (1981) employed IE for species
differentiation of bullock, buffalo, horse, goat, sheep, pig and chicken
according to the number and position of precipitation arcs by using alum
precipitated muscle extracts of the respective species as antigens.
Reddy (1986) successfully used IE and detected species specific
antibodies for identification of cooked beef and mutton. His studies
further revealed that two precipitation lines found to be common for all
the antigens tested viz., muscle extract antigens of cattle, buffalo, sheep
and goat.
2.4.4 Counter Immuno Electrophoresis (CIE) :
The diffusion of antigen and antibody is facilitated by the
application of low voltage current and by using this technique,
Schweiger et al. (1982) and Brehmer (1983) identified species origin of
meat in various mixed meat products.

xxxix
Nath (1986) used freeze dried muscle extracts as antigen for
immunization in phylogenetically related species and employed DID and
CIE for species identification and found that CIE was more sensitive
than DID.
Allsup (1987) concluded that CIE was more rapid and highly
sensitive in species identification of fresh and frozen meats and offals
using commercially produced bovine, porcine and equine antisera.
Karkare et al. (1988) employed DID and CIE using adrenal
boiling resistant and ethanol precipitable (BE) antigens for species
identification of cattle, buffalo, sheep, goat and pig and concluded that
CIE was most sensitive.
Lajon et al. (1989) detected added poultry meat in charcuterie
products by employing DID, CIE, Lourell IES and ELISA. CIE and
Lourell IES methods detected up to a level of 15% added poultry meat
while ELISA could detect up to 5% fresh meats. None of these
techniques could detect poultry meat in cooked samples.
Srinivasarao (1996) employed DID, CIE and IE tests and
concluded that DID and CIE were more sensitive than IE in the detection

xl
of adulteration using native brain extracts of sheep, goat and rabbit as
antigen.
Using CIE, species of origin in cooked meat mixtures of closely
related species have been identified by Sherikar et al. (1988), Reddy et
al. (1990), Suryarao (1990), Nath et al. (1999) and Sivakumar et al.
(1999).
2.5 ENZYME LINKED IMMUNO SORBENT ASSAY ( ELISA) :
Kang’ethe et al. (1984) and Manz (1985) employed ELISA for the
detection of adulteration in cooked meats both qualitatively and
quantitatively using heat treated beef and pork muscle containing antigen
component precipitated in the alpha 2 (α 2) globulin region.
Both competitive and indirect ELISA procedures were capable of
detecting as low as 5% pork or mutton in beef (Dincer et al., 1987).
Kang’ ethe and Gathuma (1987) used thermostable muscle antigen
and developed monospecific antisera in goats and detected adulteration
of cooked, sterilized and autoclaved meat and meat products. Kang’ethe
and Lindquist (1987) using ELISA could detect beef in pork sausages at
10% and 5% level but not at 1% level.

xli
Sawaya et al. (1990) using antisera in sheep raised against
autoclaved porcine muscle extract could detect pork at 1% level mixed
in beef and mutton at 0.5% level.
Sherikar et al. (1993) compared various immunological
techniques such as DID, CIE and ELISA in meat speciation using
thermostable species specific anti adrenal sera and found that ELISA
could detect adulteration as low as at the 1%level.
Gacheru et al. (1994) described a sandwich ELISA for speciation
of cooked meat samples of cattle, buffalo, camel, pig, horse and bush pig
and detected the adulterants at 1% level.
Mandokhot and Kotwal (1997) advocated the use of thermostable
antigens in the ELISA test to get promising results in the identification
of cooked meats.
2.6 WESTERN BLOT TECHNIQUE (ELECTROBLOT IMMUNO
ASSAY) :
By Western Blot technique, Chen and Hsieh (2001) characterized
a porcine specific thermostable muscle protein from cooked pork
successfully and the same was identified as troponin.

xlii
2.7 ISO ELECTRIC FOCUSING (IEF) :
Sinclair and Slattery (1982) reported that differentiation of buffalo
meat from beef as well as sheep, horse, kangaroo and pig was possible
by employing isoelectric focusing (IEF) but the differentiation of closely
related species such as sheep and goat or horse and donkey was not
possible by this method.
Isoelectric focusing was a sensitive and suitable technique for
identification of fresh meats according to Abraham and Leela Nayar
(1983).
Abraham (1987) found that agarose isoelectric focusing technique
was found to be effective for species identification of raw and cooked
meats. Species specific differences existed in the protein band profiles of
beef, cara beef, chevon and mutton on agarose gel isoelectric focusing
(AGIEF) using urea extracts of raw meats and this technique could be
used for rapid and reliable species identification in the raw meats.
Sherikar et al. (1988) successfully demonstrated that buffalo,
cattle, sheep, goat, pig and poultry meats could be differentiated by
using poly acrylamide gel isoelectric focusing. The bands of the raw and

xliii
heated (700C) meats behaved characteristically and distributed in a
particular pH range specific for respective species. However, the bands
of boiled and autoclaved meats were diffused and could not be
differentiated.
Mc Cormick et al. (1992) employed IEF technique in the
detection of meat from game and domestic species. Banding patterns in
meat from prong horn, mule, deer, white tailed deer, sheep, moose, pork
bison, elk, caribon, reindeer, beef and goat could be characterized with
IEF combined with specific enzyme staining for creatine kinase.
However the inability to separate some games species from domestic
species was the limitation of the application of this technique.
2.8 OTHER METHODS :
2.8.1 DNA probe Technology :
DNA assay technique was applied to the identification of meat
species specific sequence of DNA in the meat (Chikuni et al., 1990;
Wintero et al., 1990; Ebbehoj and Thomson, 1991 a & b). However the
probes prepared from genomic DNA showed cross reactions between
cattle, buffalo and sheep (Chikuni et al., 1990).

xliv
Ebbehoj and Thomson (1991a) concluded that the DNA
hybridization technique applying 32p labeled probes were useful for
quantitative determination in the quality control of heat treated meat
product.
Wintero et al. (1990) compared DNA hybridization with other
techniques and could detect raw pork in beef at 0.4% by CIE, 1% by
DID, 0.5% by IEF and DNA hybridization.
2.8.2 Polymerase Chain Reaction (PCR) :
Chikuni et al. (1994) by employing polymerase chain reaction
(PCR) technique showed 92% of homology between sheep and goat and
they were differentiated by APa1 digestion of the PCR products because
sheep had I A APa1 site and goat had no site in the PCR products.
Rasool et al. (1995) developed a simple method for the
identification of raw meats of various species based on repetitive DNA
sequence. They also further showed that raw meat derived from the
family bovidae can be identified using restriction enzyme analysis.

xlv
Matsunaga et al. (1999) could successfully employ PCR to
identify species origin of meats from cattle, pig, chicken, sheep, goat and
horse.
2.8.3 Wildlife Forensics – Micro satellite analysis of DNA :
Micro satellite DNA markers have been determined for population
samples of moose and white tailed deer and the genotyping of individual
samples have been automated using DNA sequence and gene scan and
genotype software, and these innovations facilitated rapid analysis of
evidence in criminal cases (Irv Kornfield 1999).
Coffin et al. (1999) developed micro satellite loci for a number of
big game species including pronghorn.
Wilson and White (1999) have developed a protocol using highly
repetitive satellite DNA markers to identify and quantify the amount of
game tissue present in a processed meat sample. They have used p32
radio actively labeled genomic DNA of the game species being detected
in hybridized to southern blotted DNA digested with species diagnostic
restriction enzymes which revealed species specific banding patterns
without cross homology to other game or domestic species.

xlvi
CHAPTER III
3. MATERIALS AND METHODS
All the glassware used in this work were of Corning (India) make.
The chemicals used to prepare different buffers and reagents were of
either Analar (BDH) or E-Merck. The fine chemicals used were of
Sigma make. Quartz distilled water (pH 6.8 to 6.9) was used throughout
this work to prepare buffers and reagents.
3.1 PREPARATION OF SAMBAR MUSCLE PROTEIN
ANTIGENS :
3.1.1 Collection and processing of sambar flesh :
3.1.1.1 Materials :

xlvii
1. PBS, pH 7.3 (Appendix I)
2. 0.15M saline
3. Sambar (Cervus unicolor) flesh
Four to five kilograms of flesh without any subcutaneous fat was
collected from the wildlife Sambar (Cervus unicolor) maintained at Sri
Venkateswara Zoological Park, Tirupati. The flesh was collected
immediately after the death of the animal (either natural death or death
due to accidents but without history of any infectious etiology) without
any subcutaneous fat, fascia and connective tissue. The samples were
transported to the laboratory immediately and washed with PBS and
stored at –200C in aliquots of 1 kg each till used.
3.1.1.2 Method :
1. Determination of protein concentration :
Protein concentration of samples was estimated as per the standard
method described by Lowry et al (1951).
2. Standardization of the method of processing for antigen
preparation :
In order to extract muscle proteins, two diluents viz., 0.15M saline
and Phosphate buffer saline (PBS) pH 7.3 were tried and standardization
was done depending upon the protein concentration in the solute as per
the method of Lowry et al (1951).
24

xlviii
3. Processing of sambar muscle :
The muscle was washed with PBS, pH 7.3 thrice, minced with
sterilized forceps and homogenized and the fresh muscle protein
antigens were prepared as illustrated in the flow diagram (Fig.1) while
the method for thermostable protein (TMP) antigens is depicted in
(fig.2).
Fig.1 SCHEMATIC FLOW DIAGRAM FOR PREPARATION OF
FRESH SAMBAR MUSCLE PROTEIN ANTIGENS
100 gm of muscle +200 ml of PBS, pH 7.3
Homogenate
Centrifugation@
, 5000 rpm/30 mts
Residue discarded Supernatant

xlix
Preserved with thiomersal
(1:10,000) and stored at -200C
__________________________________________________________
@ HITACHI – HIMAC CR 21 F
Fig. 2. SCHEMATIC FLOW DIAGRAM FOR PREPARATION OF
THERMOSTABLE MUSCLE PROTEIN (TMP) ANTIGENS OF
SAMBAR 50 gm of muscle + 100 ml of PBS, pH 7.3
Homogenate
Boiling water / 1hr
Centrifuged at 3000 rpm@
/ 15 min
Supernatant
Autoclaved at 15 lb psi and centrifuged at
15000 rpm @
/ 30 min at 40C
Supernatant
3 vol of ethyl alcohol / 18 hrs at room
temperature
Centrifuged at 5000 rpm @
/ 15 min at
40C

l
Precipitate
30 ml of PBS, pH 7.3, centrifuged at
2000 rpm@
/ 30 min at 40C
Supernatant (TMP)
__________________________________________________________ @ HITACHI – HIMAC CR 21 F
The supernatant contained thermostable muscle protein (TMP)
antigen and was stored at -200C in 1 ml aliquots and preserved with
thiomersal (1 in 10,000) in screw capped tubes.
3.2 PREPARATION OF FRESH AND THERMOSTABLE
MUSCLE PROTEIN ANTIGENS (TMP) OF FIELD
SAMPLES (CATTLE, BUFFALO, SHEEP, GOAT, PIG
AND RABBIT) :
Muscle antigens were prepared from field samples comprising
species of cattle, buffalo, sheep, goat, pig and chicken from the slaughter
houses of Tirupati municipality and other local slaughter houses. The
preparation of respective muscle protein antigens was carried out as
already described in 3.1. These antigens were catalogued and identified
as cattle muscle extract – fresh (C-F), cattle muscle extract – cooked (C-
C), buffalo muscle extract – fresh (B-F), buffalo muscle extract – cooked
(B – C), sheep muscle extract – fresh (S-F), sheep muscle extract –
cooked (S-C), goat muscle extract – fresh (G-F), goat muscle extract –

li
cooked (G – C), pig muscle extract – fresh (P-F), pig muscle extract –
cooked (P-C), rabbit muscle extract – fresh (R-F), rabbit muscle extract
cooked (R – C).
3.3 PRODUCTION AND PREPARATION OF HYPERIMMUNE
SERA TO SAMBAR MUSCLE PROTEIN ANTIGENS :
Hyperimmune sera to both fresh and heated antigens of sambar
muscle were prepared in different species of animals.
3.3.1 Materials :
1. Rabbits
2. Sheep
3. Freund’s Complete Adjuvant (FCA) *
4. Sambar fresh muscle antigen
5. Sambar thermostable muscle protein (TMP) antigen
6. Thiomersal (1 : 10,000)
3.3.2 Method :
1. Immunization of rabbits with fresh muscle protein antigens of
sambar :
Three healthy rabbits weighing about 1.2 kg were used for
immunization. The method followed was that of Williams and Chase
(1967) with slight modification. One initial intramuscular injection in

lii
divided doses was given at multiple sites with 1 ml of (1:1) mixture of
__________________________________________________________
*Sigma chemical company, St Louis, USA.
sambar muscle protein antigen with a protein concentration of 1.16
mg/ml. A similar injection was repeated 2 weeks later. Blood was
collected seven days after the last injection. If antibody titre of serum is
not satisfactory another injection was given and the animal was bled
after giving rest for one week. The respective sera were pooled and
preserved at –20 0C after adding thiomersal to a final concentration of 1
in 10,000.
2. Immunization of Rabbits with thermostable muscle protein
antigens (TMP) of sambar :
Three healthy rabbits weighing about 1.2 kg were used for
immunization. The method followed was that of Williams and Chase
(1961) with slight modification. One initial intramuscular injection in
divided doses was given at multiple sites with 1 ml of (1:1) mixture of
thermostable muscle protein antigen of sambar with a protein
concentration 0.88 mg/ml. Such similar injections were repeated at
fortnight intervals and a total of five injections were made with a total
protein concentration of 1.1 mg every time. After resting the animals for

liii
one week after last injection, the animal was bled, serum was separated
and after adding with thiomersal to a final concentration of 1 in 10,000
and preserved at – 200C.
3. Immunization of sheep with fresh muscle antigens of sambar :
A healthy adult sheep (ram) aged 3 years was immunized against
fresh sambar muscle protein, a similar procedure as described above
with a total protein concentration of 4.64 mg each time. A total of two
intramuscular injections were given. After resting the animals for one
week after last injection, the animal was bled, serum was separated and
preserved at –200C after adding thiomersal to a final concentration of 1
in 10,000.
4. Immunization of sheep with thermostable muscle protein (TMP)
antigen of sambar :
A healthy adult sheep (ram) aged about 3 years was immunized
against TMP antigen of sambar, a similar procedure as described above
with a total protein concentration of 3.52 mg / injection. A total of three
intramuscular injections were given. After resting the animals for one
week after last injection, the animal was bled, serum was separated and

liv
preserved at – 200C after adding thiomersal to a final concentration of 1
in 10,000.
3.4 PREPARATION OF BINARY MEAT MIXTURES :
Twenty percent muscle extracts either fresh or autoclaved muscles
of cattle, buffalo, goat, sheep pig and rabbit were prepared and binary
meat mixtures of one species meat with another was achieved by mixing
20% extract solutions in different proportions, so as to arrive at different
percentages of mixing as illustrated below.
Table 1. Preparation of binary meat mixtures
Sambar muscle
extract (ml)
Cattle muscle extract
(ml)
Percentage of
adulteration (%)
0.5 0.5 50
0.25 0.75 25
0.10 0.90 10
0.05 0.95 5.0
0.025 0.975 2.5
0.01 0.990 1.0
0.005 0.995 0.5
0.001 0.999 0.1
0.0005 0.9995 0.05

lv
Experimental adulteration was carried out similarly with muscle
extracts of buffalo, sheep, goat, pork and rabbit.
3.5 IMMUNOCHEMICAL TOOLS TO DETECT WILDLIFE
SAMBAR MEAT :
3.5.1 Double immuno diffusion (DID) test :
Double Immuno diffusion (DID) test employing agarose was
carried out to detect the presence of precipitating antibody to the sambar
muscle protein antigen, fresh or cooked, as per the method described by
Ouchterlony (1958).
3.5.1.1 Materials :
1. Glass slides, 75mm x 75mm
2. Eppendorf automatic pipette *
3. 0.7% Agarose@
4. Barbital buffer pH 8.6 (Appendix I)
5. 2% saline and physiological saline(Appendix I)
6. staining solutions (Appendix II)
(a) Amido Black stain
(b) Coomassie Brilliant blue stain
7. Destaining solutions (Appendix II)
3.5.1.2 Method :
1. Pre-coating of the slides :
Ten ml of 0.2% agarose solution was prepared while hot and glass
slides were smeared and dried.

lvi
_________________________________________________________
* Eppendorf, West Germany
@ Type I agarose, Sigma chemical company, St Louis, USA
2. Preparation and casting of the gel :
The method followed was that of Parulekar (1982). Four ml of the
hot dissolved agarose in buffer was spread over the pre-coated slide to
get uniform thickness of 1.3 mm and allowed to solidify. The gel was
allowed to harden by transferring the slides to a refrigerator in a humid
chamber. Then a five well pattern was made with a 3 mm distance
between the edges of adjacent well with a diameter of 3mm. Each well
was charged with antigen and antibody separately as the case may be
with Eppendorf automatic micropipette. Then the slides incubated at 40C
for 18 to 24 hr. After diffusion was complete, the precipitation patterns
were read directly. Some slides were washed, dried and stained.
3. Washing drying and staining of the gels :
After completion of precipitation reaction and before staining, the
unprecipitated proteins present in the gel were removed by washing in
2% saline followed by physiological saline and distilled water over the
day. For drying, the gel was covered with a Whatman no.1 filter paper

lvii
moistened with sterilized distilled water and left overnight in the
incubator at 370C.
The dried gel on the slides was stained with Amido black or
Coomassie Brilliant blue for 5 to 10 minutes followed by destaining with
destaining solution for about 15 minutes. Then rinsed in tap water and
dried.
3.5.2 Single Radial Immuno Diffusion (SRID) technique :
Single radial Immuno diffusion (SRID) technique developed by
Mancini et al (1965) with slight modifications was used to quantify
sambar thermostable muscle protein (TMP) antigens.
3.5.2.1 Materials :
1. Phosphate buffer saline, pH 7.3
2. Barital buffer pH 8.6
3. Agarose 0.7%@
4. Specific antiserum
3.5.2.2 Method :
Agarose dissolved in buffer (1.4%) was kept at 560C in a water
bath. Hyperimmune serum raised against sambar thermostable muscle
protein (TMP) antigens diluted in the buffer was added to equal volume
of agarose solution kept in the water bath to a final concentration

lviii
__________________________________________________________
@ Type I agarose, Sigma chemical company, St Louis, USA
of 5%. Similarly antiserum in buffer was also added to agarose to a final
concentration of 5%. These mixtures in appropriate quantities were over
layered on precoated glass slides so as to obtain a gel thickness 1.3 mm.
After solidification, wells of 3 mm diameter were punched out at a
distance of 12 mm between the wells.
The wells were charged with several dilutions of sambar muscle
protein (TMP antigen) and incubated at 40C in humid chamber for 24 to
48 hours.
The formation of antigen antibody precipitate rings around the
wells were observed and their diameters were measured. Some slides
were washed and stained by the method described already in 3.5.1.2.
Duplicate determinations were made for every sample on different
slides. A standard curve was constructed by plotting precipitate ring
diameters against the corresponding log of protein concentrations of the
antigen. This curve a linear one was used for calculation of sambar
muscle proteins (TMP) in the test sample.

lix
3.5.3 Counter Immuno Electrophoresis (CIE) :
Procedure of Jad (1982) was followed.
3.5.3.1 Materials :
1. Electrophoresis apparatus and power pack.#
2. Glass slides, 75mm x 75mm
3. Whatman No. 1 filter paper wicks
4. Agarose@
5. Barbital buffer pH 8.6
6. Hyper immune sera
7. Antigen
3.5.3.2 Method :
Agarose was prepared in barbital buffer, pH 8.6. Each
microscopic slide was layered with 2.5ml of agarose and after setting,
two wells with a diameter of 3mm were punched at a distance of 2mm
from each well. Ten µl of antigen was filled in the cathodal well and the
test serum in the anodal well with a micropipette.
The electrophoresis chambers were filled with 60 ml of barbital
buffer, pH 8.6. Four microscope glass slides were placed in the
_____________________________________________________
# Systronics Electrophoresis Supply 610
@ Type 1 agarose, Sigma chemical company, St Louis, USA

lx
electrophoresis chamber and contacts were made by means of Whatman
no.1 filter wicks. Electrophoresis was carried out for 5 minutes with a
constant current supply of 15 ma per slide. The slides were observed for
precipitation bands.
The electrophoresed slides were washed, dried and stained as
described earlier (3.5.1.2).
3.6 SODIUM DODECYL SULPHATE POLY ACRYLAMIDE
GEL ELECTROPHORESIS (SDS - PAGE) :
SDS – PAGE was performed by using the procedure of Laemmli (1970).
3.6.1 Preparation of reagents :
Stock solutions
1. Acrylamide Bis acrylamide stock solution (100 ml) :
Acrylamide – 30.0g
N’N1 methylene bis acrylamide – 0.8g
The above two chemicals were dissolved in 80 ml of distilled
water and made up to 100 ml of distilled water. Filtered the solution and
stored at 40C in an amber coloured bottle.
2. Resolving gel buffer (1.5m Tris – HCl pH 8.8) :
Tris base – 18.15 g

lxi
Tris base was dissolved in 80 ml of distilled water and pH was
adjusted to 8.8 with 0.1 N HCl and made up to 100 ml with distilled
water.
3. Stacking gel buffer (0.5m Tris HCl buffer pH 6.8) :
Tris base – 6.0 g
It was dissolved in 80 ml of distilled water and pH was adjusted to
6.8 with 0.1 N HCl and made up to 100 ml with distilled water.
4. 10% Sodium dodecyl sulphate (SDS) :
SDS – 10 g
Dissolved in 10 ml of distilled water and stored at room
temperature.
5. Sample Buffer :
0.5 M Tris – HCl buffer (6.8 pH) – 1.0 ml
Glycerol - 0.8 ml
10% SDS - 1.06 ml
2-mercaptoethanol - 0.4 ml
0.05% Bromophenol blue - 0.2 ml
Distilled water - 4.0 ml
The above solutions were mixed thoroughly and stored at room
temperature.

lxii
6. Tank Buffer :
Tris Base - 450 mg
Glycine - 2.16 g
SDS - 150 mg
Distilled water - 150 ml
7. 10% Ammonium per sulphate :
Ammonium per sulphate - 100 mg
Dissolved in 1 ml of distilled water and prepared just before use.
8. Staining solution :
Coomassie brilliant blue (R – 250) - 100 mg
Methanol - 50 ml
Glacial acetic acid - 7 ml
Dissolved and mixed the above chemicals thoroughly and made
up to 100 ml with distilled water.
9. Destaining solution :
Methanol - 40 ml
Glacial acetic acid - 14 ml
The above solutions were mixed and made up to 200 ml with
distilled water.
3.6.2 Gel preparation :

lxiii
Stock solution Resolving gel (12%) Stacking gel (4%)
30% Acrylamide 4.0 ml 1.3 ml
Resolving gel Buffer 5.0 ml -
Stacking gel buffer - 2.5 ml
10% SDS 0.2 ml 0.1 ml
10% APS (Ammonium
per sulphate
0.1 ml 0.05 ml
Distilled water 6.7 ml 6.04 ml
TEMED 20 µl 10 µl
3.6.3 Gel preparation procedure :
The vertical slab gel@
unit in the casting mould with 1 mm spacer
was used. Resolving gel solutions was prepared as given under
preparation of reagents. The solution was poured into the sandwich to a
level of 2 cm from the top. Distilled water was gently added along the
wall of the sandwich to form uniform gel surface after polymerization.
After polymerization the water on the resolving gel was poured off.
Stacking gel was prepared and overlayed on the resolving gel. The comb
was inserted into the sandwich of stacking gel. The gel was allowed to
polymerize for 30 min.
__________________________________________________________
@ Broviga Electrophoresis Apparatus, Chennai.
3.6.4 Sample preparation :
3.6.4.1 Fresh antigen :

lxiv
The sample was prepared by taking 50 µl of sambar fresh antigen
C-F, B-F, S-F, G-F and 50 µl of sample buffer mixed with each sample
and prepared.
3.6.4.2 Heated antigen :
The sample was prepared by taking 50 µl of thermostable muscle
protein (TMP) antigen of sambar, C-C, B-C, S-C, G-C, P-C and 50 µl of
sample buffer mixed with each sample and prepared.
3.6.5 Sample Loading :
Comb was slowly removed from the gel after polymerization from
the sandwich. The lower and upper chambers were filled with electrode
buffer. Fresh antigens were prepared and loaded in the wells as
mentioned below.
Lane: 1 Protein marker* (Phosphorylase B 97.4 kd, Bovine
serum albumin 68 kd, Ovalbumin 43 kd, Carbonic
unhydrase 29 kd, Soyabean Trypsin inhibitor 20 kd
Lysozyme 14.3kd).
__________________________________________________________
* GENEI, Bangalore
Lane: 2 Fresh muscle antigen of sambar
Lane: 3 Buffalo muscle extract – fresh (B-F)
Lane: 4 Cattle muscle extract – fresh (C-F)
Lane: 5 Sheep muscle extract – fresh (S-F)

lxv
Lane: 6 Goat muscle extract – fresh (G-F).
Another gel was prepared as mentioned above and the heated
muscle extracts (antigen) were prepared and loaded in the wells as
mentioned below.
Lane: 1 Protein marker (Phosphorylase B 97.4 kd, Bovine
serum albumin 68kd, Ovalbumin 43kd, Carbonic
unhydrase 29kd, Soyabean Trypsin inhibitor 20kd
Lysozyme 14.3kd).
Lane: 2 Thermostable muscle protein (TMP) antigen of
sambar.
Lane: 3 Buffalo muscle extract – Cooked (B-C)
Lane: 4 Cattle muscle extract – Cooked (C-C)
Lane: 5 Sheep muscle extract – Cooked (S-C)
Lane: 6 Goat muscle extract – Cooked (G-C)
Lane: 7 Pig muscle extract – Cooked (P-C)
The electrophoresis unit was connected to the power pack and
voltage was initially adjusted to 60 V and when the samples entered in
the resolving gel the current was adjusted to 100V. This was
disconnected when the moving dye touched the bottom edge of the gel.
3.6.6 Staining and Destaining :
The gel was carefully dismantled after completion of run and the
gel was immersed in a glass tray containing staining solution for 2 hours.

lxvi
Later the gel was destained in destaining solution until the back ground
of the gel became clear.
3.6.7 Molecular weight determination :
The marker proteins used in electrophoresis were Phosphorylase B
97.4 kd, Bovine serum albumin 68 kd, Ovalbumin 43 kd, Carbonic
unhydrase 29 kd, Soyabean Trypsin inhibitor 20 kd, Lysozyme 14.3 kd
used as the marker protein standards (GENEI)
The distance migrated by the meat proteins and marker proteins
were recorded and a standard curve was plotted by taking distance
migration on x-axis and molecular weights on y-axis. The molecular
weight of the meat proteins were calculated from the standard curve.
3.7 ELECTROBLOT IMMUNO ASSAY (SEMI – DRY) :
Electroblot immuno assay was performed according to the
procedure of Walker (1986)
3.7.1 Materials :
3.7.1.1 Preparation of Buffers :

lxvii
1. Anode buffer – I – pH 10.4
0.3 M Tris - 3.63 g
Distilled water - 80 ml
Methanol - 20 ml
Tris was dissolved in 50 ml of distilled water and pH was adjusted
to 10.4 with 0.1 N HCl. The solution was made up to 80 ml with distilled
water and to this 20 ml of methanol was added.
2. Anode buffer II, pH 10.4
0.0125 M Tris - 0.3 g
Distilled water - 80 ml
Methanol (20%) - 20 ml
Tris was dissolved in 50 ml of distilled water and pH was adjusted
to 10.4 with 0.1 N HCl. The solution was made up to 80 ml with distilled
water and to this 20 ml of methanol was added.
3. Cathode buffer, pH 9.4
0.02 M Tris - 0.3 g
0.04 M Glycine - 0.3 g
Distilled water - 80 ml
Methanol (20%) - 20 ml

lxviii
Tris and glycine were dissolved in 50 ml of distilled water made
up to 80 ml with distilled water and to this 20 ml of methanol was added.
4. Tris buffered saline (TBS), pH 7.5
0.02 M Tris - 2.42 g
0.15 M Nacl - 29.24 g
The above chemicals were dissolved in 900 ml of distilled water
and pH was adjusted to 7.5 with 0.1 N HCl. The solution was made up to
1000 ml with distilled water.
5. TBS – Tween 20 (TBS – T)
TBS - 1000 ml
Tween 20 - 0.5 ml
Mixed well and stored at 40C
6. Blocking solution
Non fat dried milk powder - 5 g
TBS - 100 ml
Mixed well by stirring on a magnetic stirrer for 10 min and stored
at 40C.
7. Antibody buffer
Non fat dried milk powder - 5 g
TBS – T - 100 ML
Mixed well by stirring on a magnetic stirrer for 10 min and stored
at 40C.
8. HRP labeled goat anti rabbit Ig – G conjugate (1/5000)

lxix
Conjugate - 4µl
Antibody buffer - 20 ml
9. Substrate buffer (0.5 M sodium citrate pH 5.2 for HRP system)
Tris sodium citrate - 735 mg
The above chemical was dissolved in 30 ml of distilled water and
pH was adjusted to 5.2 with 0.1N HCl and made up to 50 ml with
distilled water.
10. Substrate solution for HRP system
3, 31 diamino benzidine (DAB) - 6 mg
DAB was dissolved in 9 ml of substrate buffer and 1 ml of 0.3%
cobaltous chloride and 10 ml of 30% H2O2 were added and mixed well.
It was used immediately.
Fig. 3. Arrangement of sheets in semidry blot unit
(-)
Cathode plate
Cathode buffer filter papers
Slab gel
PDVF membrane
Anode II buffer filter papers
Anode I buffer filter papers
Anode plate
(+)

lxx
3.7.2 Method :
50 µl of TMP antigen of sambar, C – C, B – C, S – C, G – C, P –
C and 50 µl of sample buffer (1: V/V) mixed and electrophorosed by 12%
SDS – PAGE described earlier. Electroblotting was carried out by using
semi dry blot apparatus#.
__________________________________________________________
# Millipore Semidry Blot Apparatus.
The anode buffers I, II and cathode buffer were prepared and
taken separately in petriplates. The unstained gel was soaked in cathode
buffer and shaken for 20 minutes at room temperature. Seven sheets of
Whatman no.3 filter papers were cut in the size of the gel and soaked
two sheets in anode buffer I, two sheets in anode buffer – II and three
sheets in cathode buffer. The poly vinylidine diflouride (PVDF)
membrane was cut in the gel size and soaked in 100% methanol for 15
seconds and they soaked in deionized water for 15 seconds. Membrane
was equilibrated in anode buffer II for 5 minutes and the sheets were
arranged in semidry blot unit according to the fig.3 and the unit was
connected to power pack and current was adjusted to 54 mA per 1 hour.

lxxi
After the electrophoretic transfer of proteins, the membrane was
incubated on a shaker with blocking solution for 2h at room temperature.
The membrane was incubated for 1 hour in 1/40 dilution of homologous
antiserum (antibody) buffer. After 3 washings each 5 min in TBS – T,
the membrane was incubated for 1 hour in 1/1000 dilution of horse radish
peroxidase labeled goat anti rabbit antibodies in antibody buffer. The
membrane was washed thrice by 5 minutes interval in TBS – T and
transferred to a substrate solution. Colour development was recorded
visually and the reaction was stopped by washing the membrane in
distilled water. The membrane was dried and photograph was taken and
kept under dark condition.

lxxii
CHAPTER IV
4. RESULTS
4.1 DEVELOPMENT, PREPARATION AND STANDARDI-
ZATION OF IMMUNOCHEMICAL REAGENTS FOR
IDENTIFICATION OF WILDLIFE SAMBAR MEAT :
4.1.1 Standardization of the method of processing for antigen
preparation and preparation of sambar muscle protein
antigens :
As the soluble proteins of muscle are the potent antigens with
appropriate epitopes to stimulate the specific antibody production, the
solubility of sambar muscle proteins in two different solvents was tested
and standardized depending upon the total concentration of protein. The
same methodology was used in standardizing the appropriate buffer

lxxiii
system while extracting the muscle proteins of other domestic animals as
well. The comparative results of protein estimation of muscle proteins of
various domestic animals and sambar are presented in the table 2.
Table 2. Protein concentration of fresh muscle extracts of different
species
Species
Protein concentration
PBS pH 7.3
(mg / ml)
0.15 M NaCl
(mg / ml)
Sambar 1.16 0.56
Cattle 0.52 0.36
Buffalo 0.48 0.40
Sheep 0.42 0.36
Goat 0.48 0.44
Pig 0.36 0.32
Rabbit 0.40 0.36
4.1.2 Preparation of sambar fresh muscle protein antigens:
The sambar muscle was processed as illustrated in the fig.1 and
the fresh muscle protein antigens were prepared accordingly. The total
protein present was found to be 1.16 mg/ml of the fresh muscle antigen.
4.1.3 Preparation of thermostable muscle protein (TMP) antigen of
sambar :
51

lxxiv
The sambar muscle was processed and TMP antigens were
prepared as illustrated in the fig.2. The total protein present was found to
be 0.88 mg/ml of the TMP antigens.
4.1.4 Production and preparation of hyperimmune sera to sambar
fresh muscle protein antigens :
4.1.4.1 Hyperimmune sera in rabbits :
Satisfactory titre of antibody could be observed after giving three
injections of fresh sambar muscle antigen with a total protein
concentration of 3.48 mg / rabbit.
4.1.4.2 Hyperimmune sera in sheep:
Two injections of sambar fresh muscle antigen at fortnight interval
were sufficient to produce required titre of antibody in the serum of
sheep with a total protein concentration of 9.28 mg of antigen.
4.1.5 Production and preparation of hyperimmune sera to
thermostable muscle protein (TMP) antigens of sambar :
4.1.5.1 Hyperimmune sera in rabbits :

lxxv
Satisfactory titre of antibody could be observed after giving five
injections of TMP antigen of sambar with a total protein concentration of
4.4 mg/ rabbit.
4.1.5.2 Hyperimmune sera in sheep :
Three injections of TMP antigen of sambar with total protein
concentration of 10.56 mg were given to produce required titre of
antibody in the serum of sheep.
4.2 PREPARATION OF FRESH AND THERMOSTABLE
MUSCLE PROTEIN (TMP) ANTIGENS OF DOMESTIC
ANIMAL SPECIES (FIELD SAMPLES):
The field samples (cattle, buffalo, sheep, goat, pig and rabbit)
were processed, fresh and thermostable antigens were prepared and
protein concentration was estimated. The results of protein estimation of
muscle extracts (antigen) of field samples are presented in the table 3.
Table 3. Comparative studies on protein concentration of fresh and
cooked muscle extracts (TMP) of different species
Species
Protein concentration
Fresh (mg / ml) TMP (mg / ml)
Sambar 1.16 0.88
Cattle 0.52 0.34
Buffalo 0.48 0.32
Sheep 0.42 0.24
Goat 0.48 0.36

lxxvi
Pig 0.36 0.28
Rabbit 0.40 0.28
Fig. 4 Photograph of the DID reactions comparing the precipitation
reaction of sambar fresh muscle antigen against rabbit anti sambar sera
and sheep anti sambar sera. Peripheral wells contain I rabbit anti sambar
sera and II sheep anti sambar sera while the central wells contains
sambar muscle (fresh) antigen.

lxxvii
Fig. 5 CIE of sambar muscle (fresh) antigen.
Left wells - Sambar muscle (fresh) antigen
Right wells - 1, 2 – Rabbit anti sambar sera
3, 4 – Sheep anti sambar sera

lxxviii
4.3 STANDARDIZATION OF IMMUNOCHEMICAL METHODS
TO DETECT WILDLIFE SAMBAR MEAT (FRESH
MUSCLE ANTIGEN ) :
4.3.1 Standardization with rabbit antisera :
4.3.1.1 Double Immunodiffusion (DID) test :
Rabbit antisera raised against fresh antigen of sambar muscle
showed lines of identity when analyzed in Ouchterlony DID test within
six hours after incubation at 40C and this optimum titre could be
achieved after giving three injections with a total protein concentration
of 3.48 mg/rabbit.
The precipitation reaction between sambar fresh muscle extract
(antigen) and the hyperimmune sera is presented in the fig. 4.
4.3.1.2 Counter Immuno Electrophoresis (CIE):
The counter immuno electrophoresis (CIE) with sambar fresh
antigens revealed one sharp precipitation line between antigen and
antibody well and the same is presented in the fig. 5.
4.3.2 Standardization with sheep antisera :

lxxix
4.3.2.1 Double Immunodiffusion (DID) test:
Sheep antisera raised against fresh antigens of sambar meat
showed lines of identity when analyzed in Ouchterlony DID test within
six hours after incubation at 40C and this optimum titre could be
achieved after giving three injections with a total protein concentration
of 9.28 mg/sheep.
The precipitation reaction between sambar fresh muscle extract
(antigen) and the hyperimmuno sera is presented in the fig. 4.
4.3.2.2 Counter Immuno Electrophoresis (CIE):
The CIE with sambar fresh antigen revealed one sharp
precipitation line between antigen and antibody well and the same is
shown in the fig.5.
4.3.2.3 Comparison of species of animals for optimum
responsiveness in the antibody production against sambar
fresh muscle antigen :
The species of animals chosen for the production of antibody
against sambar fresh muscle antigen viz., rabbit and sheep apparently
responded similarly in the yield of satisfactory titre of antibody so
produced and hence the antisera produced in these two species of
animals were utilized in various serological tests conducted.

lxxx
Fig. 6. Photograph showing the negative precipitation reaction between
cooked muscle extracts of sambar and domestic animals against rabbit
anti sambar serum.
Central well - Sheep anti sambar serum
Peripheral wells - 1. Sambar muscle extract – TMP
2. Cattle muscle extract – Cooked
3. Buffalo muscle extract – Cooked
4. Sheep muscle extract – Cooked
5. Goat muscle extract – Cooked
6. Pig muscle extract – Cooked
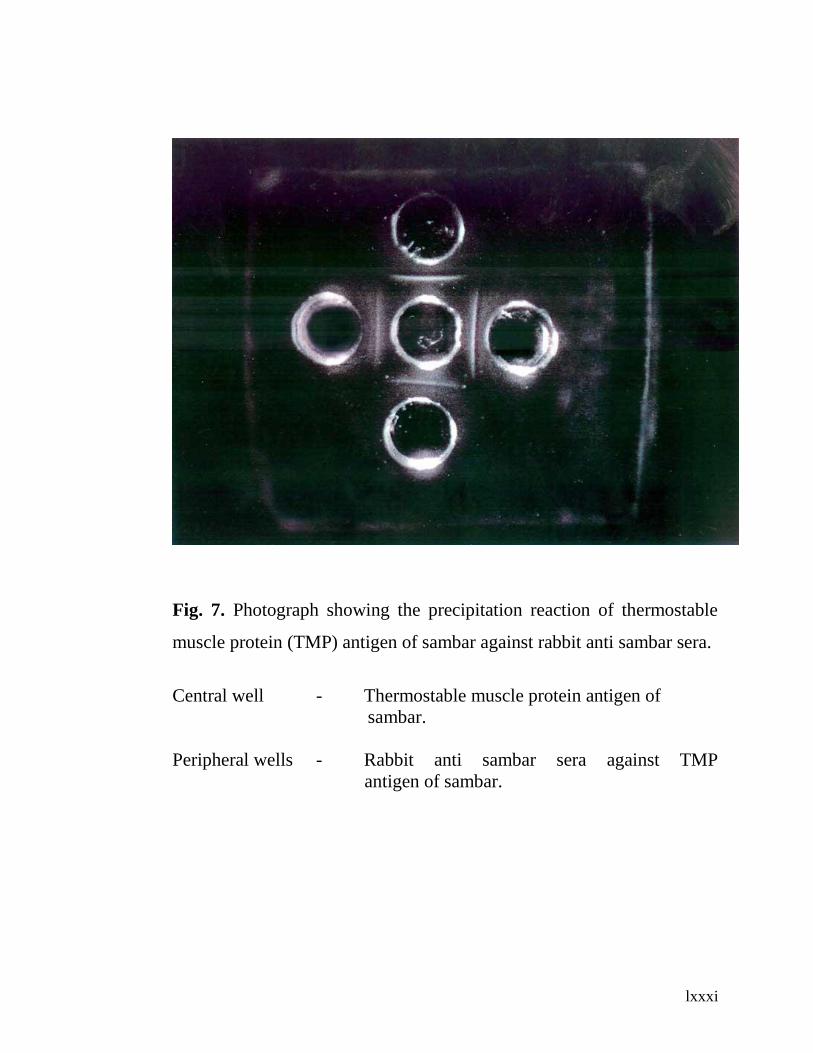
lxxxi
Fig. 7. Photograph showing the precipitation reaction of thermostable
muscle protein (TMP) antigen of sambar against rabbit anti sambar sera.
Central well - Thermostable muscle protein antigen of
sambar.
Peripheral wells - Rabbit anti sambar sera against TMP
antigen of sambar.

lxxxii
Fig.8. Standardization of SRID test for sambar TMP antigen.
Perceptible ring diameters of precipitation reaction were noticed only
with 50% rabbit anti sambar sera incorporated in the gel.

lxxxiii
Fig. 9. CIE of Thermostable muscle protein (TMP) antigen of Sambar
Left wells - TMP antigen of Sambar
Right wells - Rabbit antisambar sera

lxxxiv
4.4 STANDARDIZATION OF IMMUNOCHEMICAL METHODS
TO DETECT WILDLIFE SAMBAR MEAT
(THERMOSTABLE MUSCLE PROTEIN ANTIGEN) :
4.4.1 Standardization with rabbit antisera :
4.4.1.1 Double Immunodiffusion (DID) test:
Rabbit antisera raised against TMP antigens of sambar meat
showed perceptible lines of identity when analysed in DID test only after
72 hours when incubated at 40C and the same is presented in the fig. 7.
4.4.1.2 Single Radial Immunodiffusion (SRID) test :
Two buffer systems phosphate buffer saline pH 7.3 with ionic
strength of 0.01 and sodium barbital buffer pH 8.6 with ionic strength of
0.05 were tested to standardize the suitable buffer test system for single
radial immunodiffusion (SRID) test. The hyperimmune sera raised
against sambar fresh antigen in rabbits was tested. The ring diameters
were clear and sharp in the case of PBS than that of sodium barbital
buffer and hence the former test system was followed. The anti sambar
sera raised in rabbits reacted well and showed clear precipitin rings and
hence anti sambar sera raised in rabbits was used throughout the test.

lxxxv
As the test results would largely depend upon the antibody content
in the agarose gel, a series of dilutions of 5%, 10%, 25% and 50% of
rabbit antisera were carried out with known protein concentration of
fresh sambar meat to select the optimal dilutions which produced a clear
precipitation reaction as shown in the fig. 8.
The estimations were made with five dilutions of fresh
sambar meat and the average precipitation ring diameters so obtained
were used to construct a linear standard curve by plotting the ring
diameters against the log of protein concentration of fresh sambar meat.
4.4.1.3 Counter Immuno Electrophoresis (CIE):
The CIE with sambar TMP antigens revealed one sharp
precipitation line between antigen and antibody well and the same is
shown in the fig. 9.
4.4.2 Standardization with sheep antisera :
4.4.2.1 Double immunodiffusion (DID) test :
Sheep antisera raised against TMP antigens of sambar meat did
not show any precipitation reaction when analysed in DID test even after
72 hours of incubation.

lxxxvi
4.4.2.2 Single Radial Immunodiffusion (SRID) test :
Perceptible immuno diffusion could not be noticed around the
antigen wells even after incubating at 40C for 96 hr.
4.4.2.3 Counter Immuno Electrophoresis (CIE):
The CIE with sambar TMP antigens revealed no precipitation
reaction between antigen and antibody well.
4.4.2.4 Comparison of species of animals for optimum
responsiveness in the antibody production against sambar
thermostable muscle protein (TMP) antigen :
The TMP antigens of sambar did stimulate the antibody
production in rabbits but could not in sheep as evidenced by the
precipitation reaction observed in a variety of serological tests viz. DID,
SRID and CIE.
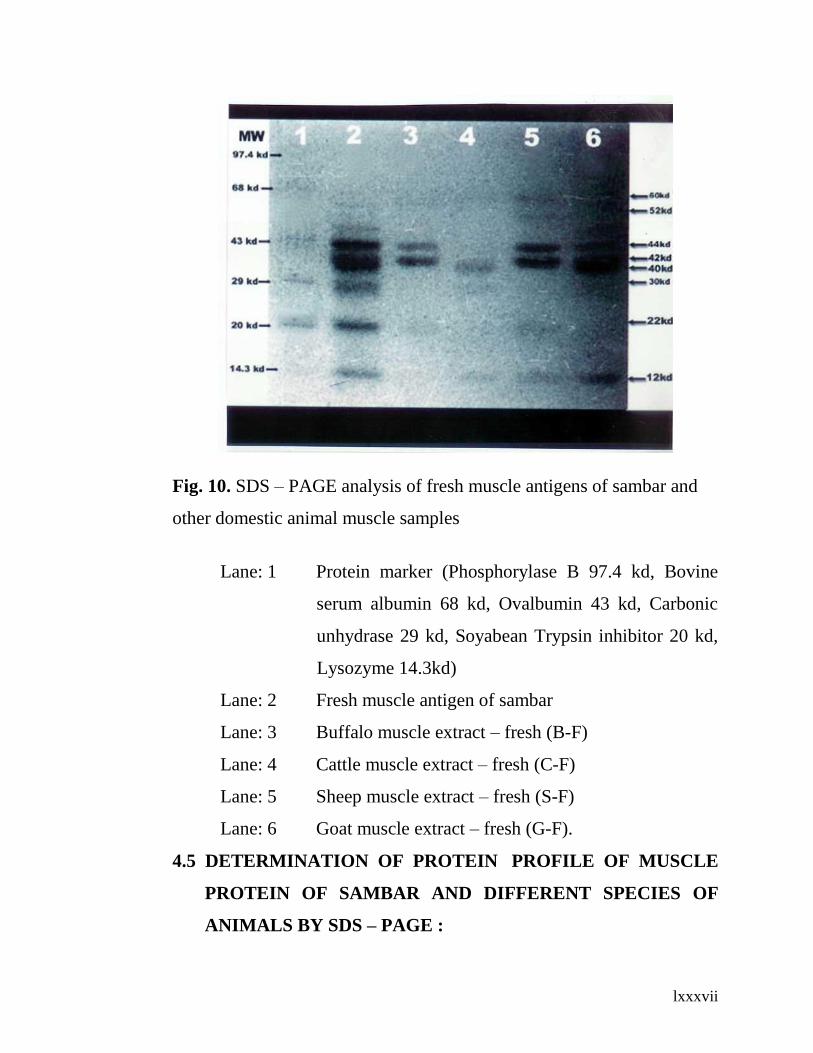
lxxxvii
Fig. 10. SDS – PAGE analysis of fresh muscle antigens of sambar and
other domestic animal muscle samples
Lane: 1 Protein marker (Phosphorylase B 97.4 kd, Bovine
serum albumin 68 kd, Ovalbumin 43 kd, Carbonic
unhydrase 29 kd, Soyabean Trypsin inhibitor 20 kd,
Lysozyme 14.3kd)
Lane: 2 Fresh muscle antigen of sambar
Lane: 3 Buffalo muscle extract – fresh (B-F)
Lane: 4 Cattle muscle extract – fresh (C-F)
Lane: 5 Sheep muscle extract – fresh (S-F)
Lane: 6 Goat muscle extract – fresh (G-F).
4.5 DETERMINATION OF PROTEIN PROFILE OF MUSCLE
PROTEIN OF SAMBAR AND DIFFERENT SPECIES OF
ANIMALS BY SDS – PAGE :

lxxxviii
The muscle proteins of different species of animals viz. buffalo,
cattle, sheep, goat, pig and sambar were subjected to SDS–PAGE to
assess their comparative molecular weights of protein profile so
delineated.
4.5.1 Comparative analysis of SDS – PAGE of fresh muscle proteins
of sambar and different species of domestic animals :
The electrophoretic separation of migrated proteins of sambar
muscle protein depending upon their molecular weight was resolved into
seven protein bands with molecular weights of 60 kd, 52 kd, 44 kd, 42
kd, 30 kd, 22 kd and 12 kd while the buffalo muscle protein profile
resolved into five protein bands with molecular weights of 60 kd, 52 kd,
44 kd, 42 kd and 22 kd. The cattle meat profile was resolved into three
protein bands with molecular weights of 40 kd, 30 kd and 12 kd while
the sheep meat protein profile resolved into six protein bands with
molecular weights of 60 kd, 52 kd, 44 kd, 42 kd and 22 kd and that of
goat meat protein profile was resolved into five protein bands 60 kd, 52
kd, 44 kd, 42 kd and 12 kd as shown in the fig. 10.

lxxxix
Fig. 11. SDS – PAGE analysis of thermostable muscle protein (TMP)
antigen of sambar and other domestic animal muscle samples.
Lane: 1 Protein marker (Phosphorylase B 97.4kd, Bovine serum
albumine 68kd, Ovalbumin 43kd, Carbonic unhydrase
29kd, Soyabean Trypsin inhibitor 20kd, Lysozyme 14.3 kd)
Lane:2 Thermostable muscle protein (TMP) antigen of sambar.
Lane: 3 Buffalo muscle extract –Cooked (B-C)
Lane: 4 Cattle muscle extract –Cooked(C-C)
Lane: 5 Sheep muscle extract –Cooked(S-C)
Lane: 6 Goat muscle extract –Cooked (G-C)
Lane: 7 Pig muscle extract –Cooked (P-C)
4.5.2 Comparative analysis of SDS – PAGE of TMP antigens of
sambar and different species of animals :

xc
The electrophoretic migration pattern of the protein bands of TMP
antigens of sambar was resolved into three bands, one major and two
minor. The major protein band comprising the proteins with molecular
weights of 44kd and 42kd was diffused in its migration probably due to
partial denaturation. The minor protein bands were 22kd and 12kd. A
similar pattern of diffused migration of two protein complex was
observed in the protein profile of the muscle proteins of other species
viz. buffalo, sheep, goat and pig except in cattle but conspicuously
absent were the two protein bands with molecular weights of 22 kd and
12 kd which were present in sambar only as shown in the fig. 11.
4.5.3 Comparison of the protein profile of fresh and TMP antigens
of sambar muscle protein :
The sambar fresh muscle protein profile when subjected to SDS –
PAGE resolved into seven bands and the same when subjected to SDS –
PAGE after cooking (TMP) resolved into only three protein bands one
major and two minor. The major protein band, a diffused one, possibly
reflects the denatured two protein complex viz. 44 kd and 42 kd while
the minor protein bands viz 22 kd and 12 kd were heat stable and the

xci
migration observed was similar in both the electrophoretic pattern of
analysis fresh muscle as well as heated (TMP) muscle protein.
4.5.4 Comparison of the protein profile of fresh and TMP antigen of
other domestic species :
The protein profile of fresh meat of buffalo, sheep and goat which
showed a similar electrophoretic pattern of migration of four protein
bands with 60 kd, 52 kd, 44 kd and 42 kd. However after subjecting to
heat treatment, three protein bands were noticed one major (comprising
44kd and 42kd of molecular weight) but merged into a single band
probably representing partial denaturation as evidenced by the diffused
pattern (fig. 10 and 11) and two minor protein bands with molecular
weights of 22kd and 12kd.
4.6 ELECTROBLOT IMMUNOASSAY (WESTERN BLOTTING)
OF COOKED MEAT (TMP ANTIGENS) OF SAMBAR AND
OTHER DOMESTIC SPECIES :
The electrophoretic migration of the cooked meat protein (TMP)
of sambar, buffalo, cattle, sheep, goat and pig followed by immunoassay
analysis were identified by using polyclonal antibodies raised in rabbits.
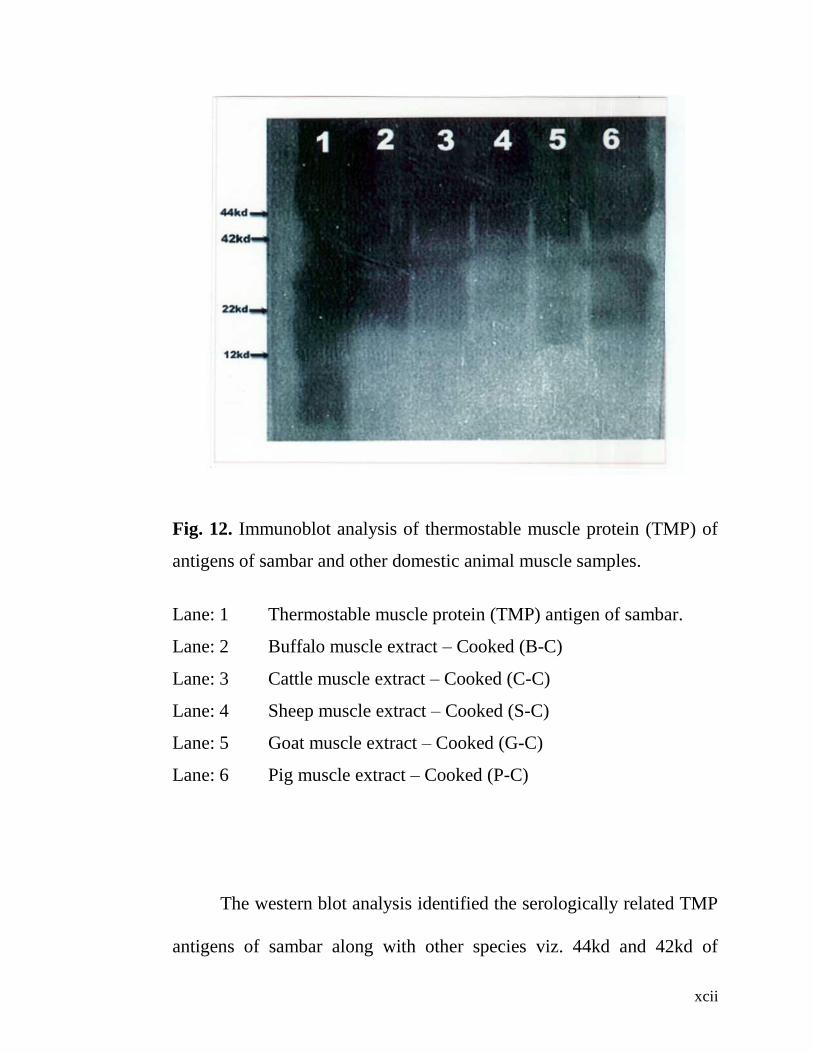
xcii
Fig. 12. Immunoblot analysis of thermostable muscle protein (TMP) of
antigens of sambar and other domestic animal muscle samples.
Lane: 1 Thermostable muscle protein (TMP) antigen of sambar.
Lane: 2 Buffalo muscle extract – Cooked (B-C)
Lane: 3 Cattle muscle extract – Cooked (C-C)
Lane: 4 Sheep muscle extract – Cooked (S-C)
Lane: 5 Goat muscle extract – Cooked (G-C)
Lane: 6 Pig muscle extract – Cooked (P-C)
The western blot analysis identified the serologically related TMP
antigens of sambar along with other species viz. 44kd and 42kd of

xciii
sambar, buffalo, sheep, goat and pig. Although 40kd protein band of
sambar could not be appreciated very well, it was identified as one of the
serologically related protein that was present in cattle.
The serologically identified proteins with molecular weight of
22kd and 12kd were present only in sambar.
4.7 COMPARISON OF SDS – PAGE ANALYSIS AND WESTERN
BLOTTING OF TMP ANTIGENS OF SAMBAR AND
OTHER DOMESTIC SPECIES :
A diffused two protein complex band comprising 44kd and 42kd
was observed in sambar, buffalo, sheep, goat and pig in SDS – PAGE
electrophoresis, and this was serologically identified common to all of
these animals by Western blotting.
The serologically identifiable protein specific to only sambar were
22kd and 12kd as seen in the western blotting (fig. 12).
4.8 DETECTION OF ADULTERATED WILDLIFE
(SAMBAR) MEAT WITH THE MEAT OF DOMESTIC
SPECIES BY IMMUNODIFFUSION AND IMMUNO
ELECTROPHORESIS :

xciv
4.8.1 Detection of sambar fresh meat adulterated with the meat
of domestic species :
4.8.1.1 Detection with hyperimmune sera raised in rabbits :
4.8.1.1.1 Double Immunodiffusion (DID) test :
Detection of experimental adulteration of sambar muscle extract
with that of cattle, buffalo, sheep, goat, pig and rabbit was done by the
standardized serological tests.
The presence of sambar meat could be established up to 1% level
of adulteration as evidenced by perceptible lines of identity in the DID
test when the anti sambar (fresh) sera raised in rabbits was allowed to
react with a series of muscle extracts of each species such as cattle,
buffalo, sheep, goat, pig and rabbit as illustrated in the table. 1. The
precipitation reaction, however could not be observed at 0.5% level.

xcv
Fig. 13. Photograph showing the precipitation reaction between rabbit
anti sambar (fresh) serum and sambar muscle (fresh) antigen
experimentally adulterated with buffalo muscle (fresh) extract.
Central well - Rabbit anti sambar serum (fresh)
Peripheral wells. 1 – 25%
2 – 10%
3 – 5%
4 – 2.5%
5 – 1%
6 – 0.5% of Sambar (fresh) muscle extract

xcvi
Fig. 14. CIE of sambar muscle (fresh) extract experimentally adulterated
with buffalo muscle (fresh) extract.
Left wells - 1 – 10%
2 – 5%
3 – 2.5%
4 – 1%
5 – 0.5 % of Sambar (fresh ) muscle extract
Right wells - Rabbit anti sambar sera.
4.8.1.1.2 Counter immuno electrophoresis (CIE):

xcvii
The results of CIE using different levels of adulteration are
furnished in the fig. 14. The antibody in the anti sambar hyper immune
sera raised in rabbits could react with the sambar meat as low as at 1%
level when adulterated with the meats of other species such as cattle,
buffalo, sheep, goat, pig and rabbits , within 5 minutes resulting in the
formation of a sharp line . Depending upon the presence of percentage of
sambar meat in the adulterated meats , the precipitation line found to be
thick to that of feeble and thin. However the sambar meat could not be
detected at 0.5% level of adulteration.
4.8.1.2 Detection with hyperimmune sera raised in sheep :
4.8.1.2.1 Double Immunodiffusion (DID) test :
The presence of sambar meat could be established up to 1%
level of adulteration as evidenced by perceptible lines of identity in the
DID test when the anti sambar (fresh) sera raised in rabbits was allowed
to react with a series of muscle extracts of each species such as cattle,
buffalo, sheep, goat, pig and rabbit as illustrated in the table.1. The
precipitation reaction, however could not be observed at 0.5% level.
4.8.1.2.2 Counter immuno electrophoresis (CIE):

xcviii
The antibody in the anti sambar hyper immune sera raised in
rabbits could react with the sambar meat as low as at 1% level when
adulterated with the meats of other species such as cattle, buffalo, sheep,
goat, pig and rabbit within 5 minutes. Depending upon the presence of
percentage of sambar meat in the adulterated meats, the precipitation line
found to be thick to that of feeble and thin. However the sambar meat
could not be detect at 0.5% level of adulteration.
4.8.2 Detection of sambar cooked meat (TMP) adulterated with the
meat of domestic species :
4.8.2.1 Detection with hyperimmune sera raised in rabbits against
sambar TMP antigens:
4.8.2.1.1 Double Immunodiffusion :
When the anti sambar hyper immune sera raised in rabbits was
allowed to react with TMP antigens of sambar meat, lines of identify
could be noticed only after 72 hours of incubation at 40C.
However the same precipitation reaction could not be noticed with
the adulterated meat of sambar with the meat of domestic animals such
as cattle, buffalo, sheep, goat, pig and rabbit. Therefore the hyper
immune sera as well as the adulterated meat mixtures were concentrated
to two folds by means of using poly ethylene glycol 600 and then the test
was run. The so concentrated hyper immune serum when allowed to

xcix
react with the concentrated adulterated meat mixtures could result in the
perceptible precipitation lines and lead to the detection of adulteration up
to 10% level but not at 5% level.
4.8.2.1.2 Single Radial Immunodiffusion (SRID) test :
The precipitation reaction around the antigen well could be
observed with the whole sambar muscle extract when the agarose gel
incorporated with 50% of the hyper immune sera. But none of the
sambar meat adulterated with meat of domestic species such as cattle,
buffalo, sheep, goat, pig and rabbit showed any precipitation reaction.
4.8.2.1.3 Counter Immuno Electrophoresis (CIE):
When the hyperimmune sera and the adulterated meat mixtures
were electrophorosed, appropriate precipitation reaction could be seen in
the form of a sharp precipitin line between antigen and antibody well
(fig.15), however depending upon the level of presence of percentage of
sambar meat in the adulterated meat mixtures, the precipitation reaction
found was formation of thick line to that of thin and faint. Thus the
adulteration could be detected up to a level of 10% but not at 5% level.

c
Fig. 15. CIE of thermostable muscle protein (TMP) antigen of sambar
experimentally adulterated with cooked buffalo muscle extract (TMP).
Left wells - 1 – 25%
2 – 10%
3 – 5%
4 – 2.5%
5 – 1% of TMP of sambar
Right wells - Rabbit anti sambar sera (heated)
4.8.2.2 Detection with hyperimmune sera raised in sheep against
sambar TMP antigens:

ci
4.8.2.2.1 Double Immuno Diffusion (DID) test :
The hyper immune sera raised in sheep when allowed to react with
TMP antigens of experimentally adulterated sambar meat with the meat
of domestic species, no perceptible reaction could be noticed even at
50% level even after incubating at 96 hours. The DID test conducted
with concentrated hyper immune sera and the adulterated meat mixtures
did not result in any perceptible precipitation reactions.
4.8.2.2.2 Single Radial Immuno Diffusion (SRID) test :
The anti sambar hyper immune sera raised in sheep against
sambar TMP antigens could not react even with concentrated sambar
TMP antigens and as such it could not detect any of the sambar meat
adulterated with the meat of other domestic species such as cattle,
buffalo, sheep, goat, pig and rabbit.
4.8.2.2.3 Counter Immuno Electrophoresis :
The hyper immune sera raised in sheep against sambar TMP
antigens when electrophoresed along with sambar TMP antigen in the
CIE test did not result in perceptible precipitation line and as such none
of the adulterated meat mixtures could be identified with the hyper
immune sera raised in sheep.

cii
Fig. 16. Immunoblot analysis of 1% adulterated thermostable muscle
protein (TMP) of sambar with other domestic species muscle extracts
(heated).
Lane 1 Thermostable muscle protein antigen of sambar
Lane 2 1% of sambar adulterated with cattle muscle extract
Lane 3 1% of sambar adulterated with buffalo muscle extract
Lane 4 1% of sambar adulterated with sheep muscle extract
Lane 5 1% of sambar adulterated with goat muscle extract
Lane 6 1% of sambar adulterated with pig muscle extract
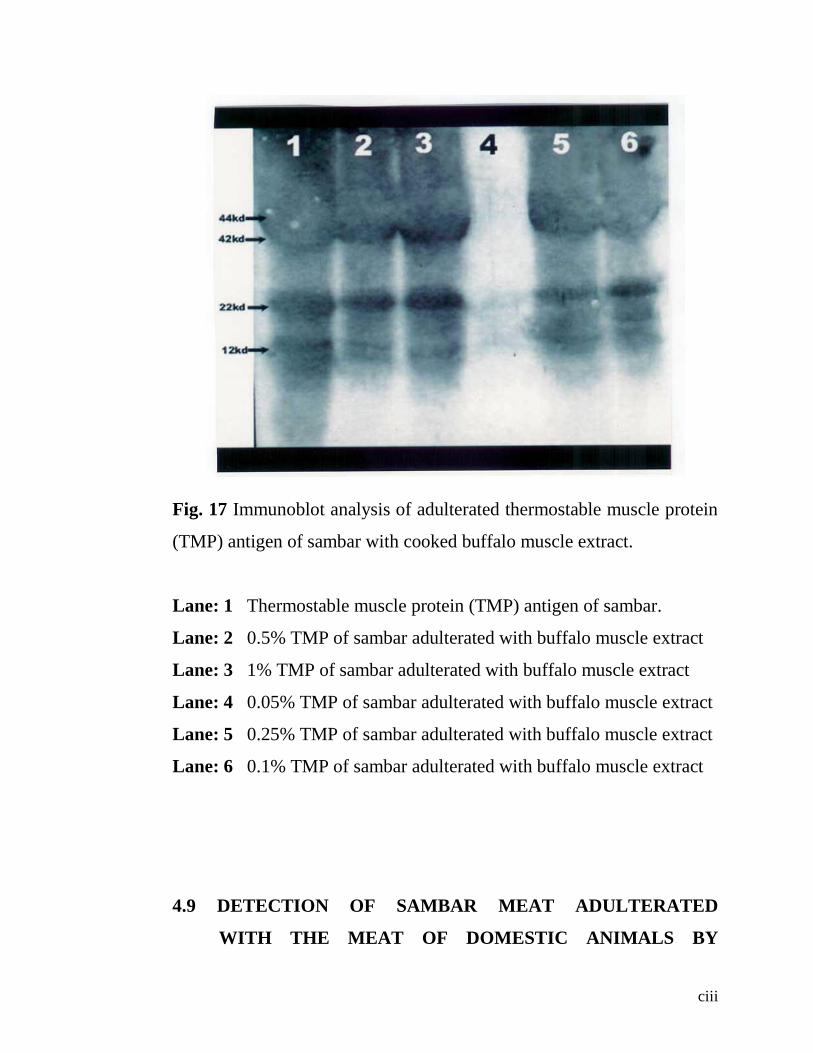
ciii
Fig. 17 Immunoblot analysis of adulterated thermostable muscle protein
(TMP) antigen of sambar with cooked buffalo muscle extract.
Lane: 1 Thermostable muscle protein (TMP) antigen of sambar.
Lane: 2 0.5% TMP of sambar adulterated with buffalo muscle extract
Lane: 3 1% TMP of sambar adulterated with buffalo muscle extract
Lane: 4 0.05% TMP of sambar adulterated with buffalo muscle extract
Lane: 5 0.25% TMP of sambar adulterated with buffalo muscle extract
Lane: 6 0.1% TMP of sambar adulterated with buffalo muscle extract
4.9 DETECTION OF SAMBAR MEAT ADULTERATED
WITH THE MEAT OF DOMESTIC ANIMALS BY

civ
ELECTROBLOT IMMUNOASSAY (WESTERN
BLOTTING) :
By western blot analysis, the adulteration of sambar cooked meat
(TMP) at 1% level with the meat of buffalo, sheep, goat and pig was
detected with distinct migration of protein with molecular weights of
44kd, 42kd, 22kd and 12kd.
4.10 DETECTION OF SAMBAR MEAT ADULTERATED
WITH BUFFALO MEAT BY ELECTROBLOT
IMMUNOASSAY (WESTERN BLOTTING)
The cooked buffalo meat (TMP) containing experimentally added
sambar cooked meat (TMP) at 1%, 0.5%, 0.25%, 0.1% and 0.05% when
subjected to western blotting with polyclonal serum raised in rabbits
against TMP antigen of sambar showed that this immunoassay could
detect sambar meat up to as low as 0.1% level but not at 0.05% level
as shown in the fig. 17.
CHAPTER V
5. DISCUSSION

cv
Certain species of wildlife like sambar and black buck are
protected species in Andhra Pradesh as they are declared as endangered
species and as such they are prohibited from hunting as per reserve
forest regulations. Now a days there is increased litigation over
misrepresentation of flesh of wildlife for flesh of domestic species like
cow, buffalo, sheep or goat (Umamaheswararao, 2003). Under such
circumstances development and standardization of wildlife forensic tools
are essential as authentic testing would be mandatory and essential in
order to detect the species origin of such meats and also to contest in a
court of law. This formed the back ground for this current study.
Till recent past, several studies were made on identification of
species origin of meats, involving various domestic species of food
animals (Reddy 1986, Giridharreddy 1994, Srinivasarao 1996,
Sridharlingareddy 1996, and Krishnamurthy 1998), however wildlife
species left untouched may be due to under reporting of poaching. In the
absence of adequate reported literature on wildlife forensics in India, any
investigator would have to fall back on immunological studies as the
case with other workers who investigated the meat speciation of
domestic animals. Therefore immunochemical determination of species
origin of meats of wildlife was attempted with the hypothesis that the
84

cvi
test antigens could be 1) obtainable in a reasonably pure form, 2)
sufficiently antigenic and foreign to the host animal, 3) thermostable to
withstand the heat treatment made during the purification process and 4)
sufficient enough to induce the production of specific antibody.
Besides conventional immunochemical methods for identification
of wildlife species (sambar) origin of meats, the present studies carried
out include certain advanced immunoassays like western blotting
(electro blot immuno assay) to detect the level of experimental
adulteration of sambar meat with the meat of other species of domestic
animals.
Trials were conducted with two diluents PBS, pH 7.3 and 0.15M
NaCl in order to standardize the suitable buffer system in which more
soluble protein goes into the solution without disrupting the antigenic
character of protein or without much loss of protein of test antigens of
the sambar meat system as evidenced by the concentration of proteins so
estimated with each diluent. The estimated protein concentration of meat
extract of different domestic animals including sambar is shown in the
table 2 which reveals that PBS, pH 7.3 is a preferred choice of buffer
system of solvent where the concentration of protein is more with every

cvii
species of the animal tested when compared with the other diluent
0.15M NaCl. Hence PBS pH 7.3 was used throughout the experiments
while extracting the muscle proteins either fresh or heated.
As a first step towards the development of wildlife forensic tools
for identification of sambar meat, the preparation of antigen from fresh
muscle extract (fig.1) and from heated muscle extract, the preparation of
thermostable proteins (fig.2) and the production of respective antibody
with appropriate immunization procedure was standardized.
The sera from the experimental animals (rabbits and sheep),
before immunizing them, were screened for the presence of antibodies
against the muscle protein antigens of sambar, cattle, buffalo, sheep,
goat, pig and rabbit and found to contain none.
Identification of species origin of meats of different food animals
depends on the production of species specific antibody by developing
antibody in phylogenetically related animals or by using species specific
protein fractions which are thermostable present in liver, testicles, brain,
pituitary and muscles as cited by Reddy et al (1990).

cviii
For the production of specific antibody against sambar muscle
antigen, two species of animals, rabbits and sheep were utilized in the
hyperimmunization experiment .Rabbit is chosen for immunization
because it is phylogenetically removed farther from the source of antigen
so that major antigenic differences will be recognized by immune
apparatus (Warnecke and Sarfle,1968; Pinto,1961;Ramadas and
Misra,1981) while sheep phylogenetically closely related to sambar for
generating immune response to the fine antigenic differences that exist
between isoantigens (Bansal and Mandokhot ,1988 ; Nath ,1986 and
Reddy ,1986 ).
However in our experiments, both of these animals responded
similarly in the antibody production to the sambar fresh muscle antigens.
But sheep failed in responding to the production of antibody against
TMP antigens of sambar, while rabbits responded adequately although
required pumping of antigen a few more times.
Possibly the reason for inadequate response of sheep to antibody
production against TMP antigens of sambar could be due to denaturation
of proteins having fine antigenic differences which could have
stimulated antibody production had the antigen been not heated. This

cix
interpretation gains strength further as evidenced by the results of the
serological experiments carried out as described in 4.3.
Therefore, TMP antigen was used to develop species specific
antibody for identification of sambar muscle besides fresh muscle
antigen in order to develop wildlife forensics utilizing the
immunochemical techniques.
Utilizing the hyperimmune sera raised in rabbits and sheep,
different conventional serological tests were tried and standardized for
the detection of sambar meat from a mixture (binary mixture) of
adulterated meats of different domestic species of animals.
The DID test carried out with sambar fresh meat antigen using
homologous antisera raised in rabbits and sheep showed precipitation
reaction with lines of identity (fig.4) when incubated at 40C within six
hours and is in full agreement with the findings of Giridhar reddy, 1996;
Sherikar et al.,1993; and Allsup,1987 who used rabbit antisera to
identify fresh muscle antigens of domestic species. However similar
results could be obtained with the hyperimmune sera raised in rabbits or
sheep when allowed to react with the fresh muscle antigen of buffalo and
cattle, while the precipitin lines were thin and feeble with the fresh

cx
muscle antigens of sheep and goat but not with pig. These results concur
with the findings of Sherikar et al. (1979) and Sherikar et al. (1987) who
noticed cross reactions between meats of heterologous species viz.,
cattle, buffalo, sheep, goat and pig and also with the findings of Pandey
and Pathak (1975), Hayden (1979) and Allsup (1987) who observed
cross reactions between cattle, horse, lamb and pig. Therefore it could be
inferred that sambar can also be considered as phylogenetically related
to other species viz., buffalo, cattle, sheep and goat but not with pig.
The DID test carried out with the hyperimmune sera raised in
rabbits and sheep against TMP antigens of sambar when tested with
homologous antigens and of heterologous species i.e., cattle, buffalo,
sheep, goat and pig resulted in interesting findings contrary to the
findings of the DID test using fresh muscle antigen. The rabbit antisera
reacted only with TMP antigens of homologous species (sambar) but not
with any of the heterologous species, and it could be due to denaturation
of serologically related proteins during heat treatment. These findings
are slightly deviated from the findings of earlier workers who noticed
faint reactions while working with speciation of meat of domestic
animals (Sherikar et al., 1987). However the hyperimmune sera raised in
sheep did not react even with homologous and heterologous species

cxi
antigen as the proteins specific for sambar might have been denatured
and hence could not elicit appropriate antibody production and there is
no reported literature to draw parallels. However, Carpenter (1965) was
of the view that heat caused denaturation of protein resulting in the loss
of antigenic fraction to form an antigen antibody complex leading to the
formation of precipitin line.
Ramdas and Misra (1981) and Sherikar et al. (1979) made
speciation of meats based on the number of precipitation lines using
antiserum raised in rabbits in DID test and immuno electrophoresis (IE).
In the present study also two precipitation lines were observed in the
DID test using rabbit antiserum with sambar fresh muscle antigen while
the DID test showed only one precipitation line with TMP antigens and
was species specific for sambar muscle only without any cross reactions
with TMP antigens of domestic animals.
The TMP antigen which was used to raise antibody in rabbits was
composed of heat stable proteins with molecular weight ranging from
44000 daltons to 12000 daltons. Some of these proteins are myofibrillar
protein fractions corresponding to tropomyosin and troponin fractions as
observed by Radhakrishna et al. (1989) pertaining to mutton and beef.

cxii
The rabbit antisera developed against TMP antigen was species specific
as it did not show cross reactions with other species of animals as
observed by Giridharreddy (1994) in the case of species specific TMP
antigens of pork.
In order to estimate the levels of presence of sambar meat in the
adulterated meats of other species, an attempt was made to standardize
single radial Immuno diffusion (SRID) test. The phosphate buffer saline
(PBS), pH 7.3 with ionic strength of 0.01 found to be the suitable test
system during the course of standardization with sodium barbital buffer,
pH 8.6 with ionic strength of 0.05. The anti sambar sera raised in rabbits
reacted well and showed clear precipitin rings around the antigen well
with 50% antiserum in agarose after incubating at 40C for 72 hours. The
SRID test conducted with sheep antiserum did not yield any visible
precipitin rings as in the case of DID test.
Although DID test is a reliable technique for differentiating meats
of different species (Ramadas 1972, Beljaar and Oleman 1977, Sherikar
et al., 1979, Hayden 1980, Nath 1986, and Bansal and Mandokhot
1988), it is cumbersome, time consuming and costly. Hence counter
immuno electrophoresis (CIE) was tried and standardized.

cxiii
The sambar fresh muscle antigen along with hyperimmune sera
raised in rabbit or sheep charged in the cathode well and anode well
respectively, when electrophoresed resulted in a clear sharp precipitin
line between antigen and antibody well (Fig 5 ). When the hyperimmune
serum raised in rabbits or sheep allowed to react with fresh muscle
antigens of cattle, buffalo, sheep and goat and pig reacted similarly in
the CIE test resulting in the formation of a sharp precipitin line which is
in agreement with the findings of Gridharreddy (1994); Allsup (1987)
and Sherikar et al. (1993) who obtained similar results while working
with the speciation of meats of domestic animals. However, there was no
visible precipitation reaction with that of pig in both the tests.
The TMP antigens of sambar when subjected to CIE test utilizing
sheep antisera raised against sambar TMP antigens, did not yield any
precipitation reaction unlike rabbit antiserum which had given a faint
precipitin line after running for 5 minutes followed by incubation at 40C
for 24 hours which resulted in increased sharpness of the precipitin line.
These results are in agreement with the findings of Giridharreddy,
(1994) who could observe similar results with porcine TMP antigens and

cxiv
Sherikar et al. (1993), using TMP antigens of cattle, buffalo, sheep, goat
and pig with homologous serum.
To delineate the sambar fresh muscle proteins with different
molecular weights, SDS – PAGE studies were carried out to examine
whether SDS – PAGE can be recommended as a standard procedure for
detection of adulteration. Comparative studies were also carried out with
muscle proteins of other species viz., cattle, buffalo, sheep and goat.
Upon comparison, the protein bands with similar molecular
weights viz 60 kd, 52 kd, 44 kd, and 42 kd were more or less similar in
sambar, buffalo, sheep and goat. Strangely cattle muscle protein profile
appears to be somewhat different in that these proteins are conspicuously
absent while it contained one protein band with 40 kd.
The protein band with a molecular weight of 30 kd was present
only in sambar and cattle, while it was absent in all other species.
The protein band with a molecular weight of 22 kd could be seen
in sambar, buffalo and sheep while it was absent in cattle and goat.

cxv
But a common low molecular protein band with a molecular
weight of 12kd was noticed in all the species of animals except in
buffalo.
Although work on wildlife is scanty, a similar type of work
involving meat samples of venison, beef and pork, when electrophoresed
showed species specific separation patterns (Anneli Skrokki, 1994).
Furthermore, our results are also in agreement with those of Ramadas
(1972), Prasad (1974), Govindarajulu (1982) and Nath et al. (1999) who
established a relationship of species specific muscle protein band
involving different species.However further work on SDS – PAGE is
required to identify the species specific proteins of sambar.
The electrophoretogram of TMP antigens of sambar with different
species of animals revealed a diffused protein band comprising protein
bands with molecular weights of 44kd and 42kd were seen in all the
species of animals including sambar except cattle, probably the diffused
pattern was due to partial denaturation. The specific protein bands
conspicuous only to sambar were low molecular weight protein bands of
22kd and 12kd, which were totally absent in any of the species of
animals when subjected to the SDS – PAGE as shown in the fig. 11.

cxvi
These results further prove that TMP antigens were species specific in
that they can be relied upon while detecting adulteration (Babiker et al.,
1981).
Identification of specific immunological proteins which are of
diagnostic significance can be made accurately by latest and advanced
technique of western blotting which gives a concrete evidence. This
latest technique was applied in the present study to identify the specific
immunological proteins which are species specific to sambar. This
technique involved separation of TMP antigens of sambar in gels
followed by electrophoretic transfer of the separated polypeptides to
nitrocellulose membrane for probing specific proteins by antisera raised
in rabbits.
The sambar TMP antigens when subjected to western blotting
resulted in the serologically identified TMP antigens of sambar i.e 44kd
and 42kd along with other species, buffalo, sheep, goat and pig but not
with cattle. Further the serologically identified proteins with molecular
weight of 22kd and 12kd were confined only to sambar. Although the
diffused pattern of the migration of the protein complex comprising
44kd and 42kd in sambar was serologically identified with other species

cxvii
of animals and the migration of this protein complex is diffused in all the
species possibly due to partial denaturation and as such the proteins with
22kd and 12kd molecular weight were probably species specific to
sambar and the same can be of diagnostic value in forensic studies. Our
findings are in agreement with those of Chen and Hsieh (2001) who
adopted western blotting technique and identified a thermostable 24kd
protein as species specific to pig.
The experimentally adulterated fresh meat of domestic animals
with fresh sambar meat (binary mixtures) when subjected to DID test
with rabbit or sheep antiserum, showed precipitation lines up to 1% level
of adulteration but not at 0.5% level and these results are in agreement
with those of Suryarao (1990) who used testicular antigens to detect
fresh meat of buffalo meat adulterated up to 10% level.
The DID test when used to detect adulterated cooked meats of
domestic animals with sambar could identify up to a level of 10% of
adulteration which is in agreement with those of Giridharreddy (1994)
who could detect adulteration of cooked pork up to a level of 20% and
Sherikar et al. (1993) who detected adulteration of cooked meats up to a
level of 5 -10%.

cxviii
Unsuccessful attempts were made to standardize single radial
immunodiffusion (SRID) test in order to estimate the levels of
adulteration. Although 50% of hyperimmune sera raised in rabbit was
required to be incorporated in agarose, it could not detect any of the
adulterated cooked meats.
Counter immunoelectrophoresis (CIE) was successfully
standardized as it is supposed to be sophisticated, reliable (without false
positive or false negative results) quick, rapid and economical.
The CIE test could detect the adulteration of sambar fresh meat as
low as up to 1% level with rabbit or sheep antisera which is in agreement
with those of Suryarao (1990).
The homologous hyperimmune serum raised in sheep could not
detect sambar meat in the adulterated cooked meats in CIE test similar
to that of DID test. The incompetence of the antibody to detect
adulteration could be due to failure of denatured proteins to stimulate
antibody production probably because of close phylogenetic relationship
with that of sambar.

cxix
The TMP antigens of sambar elicited adequate immune response
in rabbits which are phylogenetically distinct and the hyperimmune sera
so raised could detect adulteration up to a level of 10% but not at 5%
level and these results are in agreement with those of Giridhar reddy
(1994) and Suryarao (1990) who could detect adulteration up to a level
of 20% while Sherikar et al. (1993) detected up to a level of 5-10% of
adulteration.
Successful attempts were made to detect the presence of cooked
meat of sambar (TMP) in the experimentally adulterated cooked (TMP)
meats of domestic animals by using western blotting technique. Cooked
meats of phylogenetically related animals like buffalo, cattle, sheep and
goat and that of pig were experimentally adulterated at 1% level and
subjected to western blotting which could detect sambar cooked meat up
to 1% level in all the cooked meats of animal species tested. These
results could not be compared as the literature is lacking in this direction.
Western blotting studies were continued to identify the presence
of sambar cooked meat and its cut off level by selecting the cooked meat
of a domestic animal which is phylogenetically closely related and in
this case buffalo cooked (TMP) meat was experimentally adulterated

cxx
with TMP antigens of sambar with varying percentage. The polyclonal
antisera could detect serologically related proteins of sambar in these
binary mixtures as low as up to 0.1% level but not at 0.05% level as
shown in the fig. 17.
The binary mixtures of fresh meats adulterated with sambar fresh
meat antigen when subjected to immunological tests, the adulteration
could be detected at 1% level by DID and CIE, while DID and CIE tests
could detect the level of adulteration up to 10% of TMP antigens of
sambar. Western blotting analysis could detect the level of adulteration
of TMP antigens of sambar as low as 1% level when adulterated with the
meat of domestic animals like buffalo, cattle, sheep, goat and pig. The
western blotting could detect adulteration of cooked buffalo meat with
cooked sambar (TMP) meat as low as 0.1% level indicating the
reliability, sensitivity and specificity of the test.
However, continued studies are required in this direction
involving other wildlife also in order to standardize this technique as
dependable and fool proof tool in wildlife forensics.

cxxi
CHAPTER VI
6. SUMMARY
Experimental investigations were carried out to develop
appropriate wildlife forensic tools to detect species origin of sambar
(Cervus unicolor) meat for the first time.
Preparation of muscle protein antigens of sambar and other
species of domestic animals both fresh and thermostable proteins (TMP)
was standardized after conducting experimental trials with two diluents
PBS, pH 7.3 and 0.15 M NaCl. Most of the soluble proteins of the
muscle extracts which are potent antigens and necessary in forensic
work could be extracted more with PBS, pH 7.3 compared to 0.15 M
NaCl as evidenced by the estimated concentration of the protein.
Therefore PBS, pH 7.3 was selected and used in the experiments.
Suitability of experimental animals to develop specific antibody
against sambar muscle protein antigen was examined using rabbit, a

cxxii
phylogenetically distinct animal from the source of the antigen to have
major antigenic differences recognized by the immune apparatus while
sheep, a phylogenetically closely related to sambar to induce immune
response to the antigenic differences that exist between isoantigens.
Although both of these animals immunologically responded adequately
to the fresh muscle antigen of sambar, rabbit was found to be an ideal
choice for adequate induction of immune response to the thermostable
muscle proteins (TMP) when compared to sheep, probably because of
the immune apparatus in sheep failed to recognize the fine differences
between the isoantigens. Therefore, rabbit antisera against TMP antigens
of sambar was used throughout the present investigation.
Sambar fresh muscle antigens were prepared from the sambar
flesh collected immediately from the naturally dead sambar (either by
natural death or by accidental death without any infectious etiology)
from Sri Venkateswara Zoological Park, Tirupati.
Sambar fresh muscle antigens were prepared in PBS, pH 7.3,
while thermostable muscle protein (TMP) antigen was the boiling
resistant and ethanol precipitate of cooked meat extract.

cxxiii
Hyperimmune sera was raised to both sambar fresh muscle
antigens and TMP antigens in rabbits and sheep. Rabbit, a
phylogenetically distinct animal responded adequately to both fresh and
TMP antigen while sheep, a phylogenetically related species responded
to fresh antigens but not to TMP antigens.
The immunochemical methods like DID test and CIE test to detect
wildlife sambar fresh muscle antigens were standardized using
hyperimmune sera raised in both rabbit and sheep.
The DID test revealed lines of identity with sambar fresh muscle
antigen when allowed to react with homologous antiserum of rabbit and
two precipitation lines were noticed. The precipitation reaction with
sheep antisera showed lines of identity with only one precipitation band.
Lines of identity were also observed with the fresh muscle antigens of
buffalo, cattle, sheep, goat and rabbit but not with pig amply indicating
that these species except pig are closely related to sambar. When the
DID test was conducted with TMP antigens of sambar using
hyperimmune sera raised in rabbits, lines of identity were noticed after
prolonged incubation at 40C for 72 hours, but not with sheep antisera.

cxxiv
In order to improve upon the sensitivity, specificity and rapidity,
CIE test was standardized using both fresh muscle antigens and TMP
antigens of sambar.
The CIE test did give a sharp precipitation line between antigen
and antibody well with both the type of antigens when allowed to react
with respective sera raised in rabbits. The gel was electrophorosed for 5
min at 40C. The CIE test could detect the fresh antigens of species of
different domestic animals such as cattle, buffalo, sheep, goat, rabbit but
not pig, while such cross reactions were not noticed when tested against
TMP antigens of domestic animal species. The sheep antisera raised
against TMP antigens of sambar showed no cross reactions with any of
the phylogenetically related species of domestic animals and not even
with homologous antigen.
In order to detect the percentage level of sambar meat in the
experimentally adulterated meats of domestic animals, SRID test was
standardized using rabbit antisera. The SRID test could be standardized
with 50% of the hyperimmune sera incorporated in the agarose gel.
However SRID test could not detect the presence of sambar meat in any

cxxv
of the binary mixtures and as such SRID test was found to be unsuitable
as an immunochemical tool in wildlife forensics.
SDS – PAGE studies were also conducted to study the protein
profile of sambar fresh and TMP antigens and a comparison was made
with those of other domestic animal species. The electrophoretogram
revealed that sambar fresh muscle protein was resolved into seven
protein bands with molecular weights of 60kd, 52kd, 44kd, 42kd, 30kd,
22kd and 12kd while the buffalo muscle protein profile resolved into
five protein bands with molecular weights of 60kd, 52kd, 44kd, 42kd
and 22kd. The cattle meat profile was resolved into three protein bands:
40kd, 30kd and 12kd while the sheep meat protein profile resolved into
six bands: 60kd, 52kd, 44kd, 22kd and that of goat meat protein profile
was resolved into five protein bands: 60kd, 52kd, 44kd, 42kd and 12kd.
The electrophoretogram of TMP antigens of sambar and other
species of domestic animals (buffalo, sheep, goat and pig except cattle)
showed a partially denatured two protein complex with diffused
migration pattern (44kd and 42kd). Two protein bands with low
molecular weights 22kd and 12kd were present only in sambar but not in
other animals possibly species specific.

cxxvi
The electroblot immuno assay (western blot) identified
serologically related TMP antigens, 44kd and 42kd of sambar, buffalo,
sheep, goat and pig. Lone serologically related protein 40kd was noticed
in cattle although not appreciated very well in sambar. The serologically
identified proteins with molecular weight of 22kd and 12kd were present
only in sambar meat, possibly species specific.
The conventional immunochemical tests like DID and CIE were
carried out using rabbit or sheep antiserum to detect adulteration of
sambar fresh meat from binary mixtures of fresh meat of buffalo, cattle,
sheep, goat, rabbit and pig. These tests could detect the adulteration up
to 1% level, while detection in adulterated cooked meats of domestic
animals could be up to a level of 10% only with hyperimmune sera
raised in rabbits against TMP antigens. The sera raised in sheep against
TMP antigen of sambar could not detect the adulteration.
Western blotting did identify cooked (TMP) sambar meat
adulterated with the cooked meats of buffalo, cattle, sheep, goat and pig
as low as up to a level of 1%. The experimental adulteration of cooked
(TMP) sambar meat with cooked (TMP) buffalo meat, a
phylogenetically closely related domestic animal, detected as low as up

cxxvii
to a level of 0.1% but not at 0.05% level. Thus western blotting proved
to be a promising reliable, specific and sensitive technique in wildlife
forensics.

cxxviii
LITERATURE CITED
Abraham J 1987 Isoelectric focusing as a method for species
differentiation of raw and cooked meats. Ph.D. thesis, Andhra
Pradesh Agricultural University, Hyderabad.
Abraham J and Leela Nayar 1983 Effect of storage of meat in frozen
condition on the sarcoplasmic protein and patterns in agarose
isoelectric focusing. Kerala Journal of Veterinary Science 14: 76-
80.
Agrawal K C 2000 Wildlife of India conservation and management, pp
78. Nidhi publishers, India.
Allsup T N 1987 A comparison of the agar gel immunodiffusion (AGID)
and counter immunoelectrophoresis (CIE) tests for species
identification of imported red meat and offal. Meat Science 20:
119-128.
Anneli Skrokki and Osmo Hormi 1994 Composition of minced meat.
part A: Methods. Meat science 38: 497 – 501.
Babiker S A, Glover P A and Lawrie R A 1981 Improved determination
of horse meat in heated food stuffs. Meat Science 5: 473.
Bansal A K and Mandokhot U V 1988 Studies on differentiation of beef
from meat of other species of animals. I. Comparative specificity
and sensitivity of serum raised in buffalo calves against cattle
antigens. Journal of food science and technology 25 : 146-151.
Beljaare P R and Oleman W J 1977 Collaborative study on the detection
of caseinate and soy protein isolate in meat products. Annales de
la Nutrition et de l’Alimentation 31:603.
Brehmer H 1983 Positive results for soya protein meat products
detection in routine examination. Fleischwirtschaft 63: 1771.
108

cxxix
Carpenter P L 1965 Immunology and serology. 2nd
edition, pp 56. W B
Saunder’s company, London.
Chen F C and Hsieh Y H P 2001 Separation and characterization of
species specific thermostable muscle protein from cooked pork.
Journal of Food Science Vol. 66 No.5 799-803.
Chikuni K Ozutsumi K, Koishikawa T, and Koto S 1990 Species
identification of cooked meat by DNA hybridization assay. Meat
Science 27: 119.
Chikuni K Tabata T Kosugiyama M and Monma M 1994 Polymerase
chain reaction assay for detection of sheep and goat meats. Meat
Science 37: 337 – 345.
Coffin J.W., Corey S. Davis, Deedra Hawk, and curtis stroback, 1999 “A
DNA typing system for pronghorn Antelope and its use in a
poaching investigation from Wyoming” Northwest Association of
forensic scientists, Abstracts, Fall 1999.
Dincer B, Spearow J.L Cassens R.G and Greaser M.L 1987 The effects
of curing and cooking on the detection of species origin of meat
products by competitive and indirect ELISA techniques Meat
Science 20: 253 – 265.
Durand M Schneider R 1962 Immuno electrophoresis of meat juice and
its application to the detection of fraud. Arch. Inst. Pasteur Junic
39: 153 (Veterinary bulletin 33: 2188, 1963).
Ebbehoj K F and Thomsen P D 1991a Species identification of heated
meat products by DNA hybridization. Meat Science 30: 221.
Ebbehoj K F and Thomsen P D 1991b Differentiation of closely related
species by DNA hybridization. Meat Science 30: 221.
Flego R and Borghese R 1979 Analysis of meat production: critical
comparison of double immunodiffusion and COE methods for
minimum antigen detection. Bullettina Provinciali 5: 701-711.

cxxx
Fugate H G and Shelton P R 1971 Biochemical techniques-
Immunodiffusion technique for the identification of animal
species. Journal of AOAC 54: 1152.
Gacheru D N Kang’ethe E K Kaburia H F A and Njeruh F M 1994
Sandwich enzyme immunoassay for speciation of cooked meat
and for detecting trace amounts of adulterants in phylogenetically
related species. Agricultural and forestry Journal 59: 205 – 212.
Ginsberg A 1948 The differentiation of meats by precipitation test.
Veterinary Record 60: 683 – 685.
Govindarajulu M, Ranganatham M and Ramamurthy R 1982 Studies on
the differentiation of beef from mutton by starch gel
electrophoresis. Cherion 11: 6.
Graber, P and Williams C A 1953 Immunoelectrophoretic Analysis. In
Application to Human Biological fluids(eds Graber and Burtin)
Elsevier Publication Co. Amsterdam, London, New York.
Giridharreddy M 1994 Immunochemical detection of pork using
thermostable muscle protein antigens. M.V.Sc Thesis submitted to
the Acharya N.G. Ranga Agricultural University, Hyderabad.
Hayden A R 1979 Immunochemical detection of ovine, porcine and
equine flesh in beef products with antisera to species myoglobin.
Journal of Food Science 44: 494.
Hayden A R 1980 Immunochemical detection of species origin of meat
products. Ph.D Thesis, University of Wisconsin, Madison, W I.
Cited from Hayden A R 1981.
Hayden A R 1981 Use of antisera to heat stable antigens of adrenals for
species identification in thoroughly cooked beef sausages. Journal
of Food Science 46: 11810 – 11813.
Irv Kornfield 1999 Establishment of microsatellite databases for forensic
casework in the northwest united states. Northwest Association of
forensic scientists abstracts fall 1999.

cxxxi
Jad C Y 1982 Counter Immunoelectrophoresis. pp 66 – 107 In Gel
Immunodiffusion techniques in research and laboratory Medicine
(eds S S Kelkar and P M Khare) Popular Prakashan Private
Limited, Bombay.
Kang’ethe E K and Gathuma J M 1987 Species identification of
autoclaved meat samples using anti sera to thermostable muscle
antigens in an enzyme immunology. Meat Science 19: 265 – 270.
Kang’ethe E K and Lindquist K J 1987 Detection beef and pork in fresh
and heat treated meat products using enzyme immunology.
Journal of Food Science and Technology 24: 289 – 293.
Kang’ethe E K Lindquist and Gathuma J M 1984 In Biochemical
identification of meat species (Paterson RLS (ed.) Elsevier
Applied Science Publishers.pp129. cited from Kang’etha and
Gathuma 1987.
Karkare U D Sherikar A T Khot J B and Jaya rao B M 1988 Meat
speciation in commercial canned meats using species antisera to
thermostable antigens of adrenal glands. Indian journal of Meat
Science & Technology Vol 1(2).
Krishnamurthy G 1998 Identification of cooked meats using
thermostable brain antigens M.V.Sc Thesis Acharya N.G. Ranga
Agricultural University. Rajendranagar, Hyderabad.
Lammeli U K 1970 Cleavage of structural proteins during the assembly
of the head of Bacteriophage T4. Nature 227: 680 – 685.
Lajon A, Pauteau M L Demeulemester C Martin J L Pinson M and
Durand P 1989 Identification of added poultry meat in charcuterie
products. Viandes-et-produits-carnes 10: 203-206.
Landsteiner K 1945 Specificity of serological reactions. Dover
publications New York. Cited from Sherikar et al (1987).
Lawrie R A 1977 Meat identification in fresh and processed foods. Food
Technology Australia 20: 347 (FSTA 10, 4s, 505, 1978).

cxxxii
Lowry O H, Rosenbrough N J Farr A L and Randall R J 1951 Protein
measurement with Folin-phenol reagent Journal of Biological
Chemistry 193: 265 – 275.
Mancini G, Carbonara A O and Heremans J F 1965 Single radial
Immunodiffusion. PP 111 – 118. I Gel Immunodiffusion
techniques in research and Laboratory medicine (eds S S Kellar
and P M Khave 1982). Popular Prakshan Private Limited,
Bombay.
Mandokhot U V and Kotwal S K 1997 Enzyme linked immuno sorbant
assay in detection of species origin of meats – A critical appraisal.
Journal of Food Science and Technology 34: 369 – 380.
Manz J 1985 Detection of heat denatured muscle proteins by means of
ELISA. Determining beef and pork muscle proteins.
Fleiswirtschaft 65: 497 – 499.
Matsunaga T Chikuni K Tanabe R Muroya S Shibata K Yamda J and
Shimura Y 1999 A quick and simple method for the identification
of meat species and meat products by PCR assay. Meat Science
51: 143-148.
Mc Cormick R J, Collins D A, Field R A and Moore T D 1992
Identification of meat from game and domestic species. Journal of
Food Science 57: 516 – 517.
Nath D R 1986 Comparative studies on electrophoresis and
immunodiffusion technique for identification of fresh buffalo and
sheep meat. Ph.D., Thesis, Haryana Agricultural University,
Hisar.
Nath D R Chandir N K and Mandokhot U V 1999 Counter current
Immuno Electrophoresis Technique for Meat Specification and
detection of adulteration in meats. Souvenir cum proceedings of
National Seminar on processing of Meat, Poultry and by-products
for value addition.pp 231.
Nath D R Chandirmani N K and Mandokhot U V 1999 SDS-PAGE for
the different meat species. Proceedings of the National Seminar on

cxxxiii
Processing of Meat, Poultry and by-products for value addition.
pp 219.
Ouchterlony O 1958 Double diffusion in two dimensions PP 19. 16 – 10.
20. In Hand Book of experimental Immunology (ed D M Weir
1978) Volume I. Black well Scientific publications, Oxford.
Pandey R and Pathak R C 1975 Serological relationship of
taxonomically closely related domestic ruminants. Indian Journal
of Experimental Biology 13: 371.
Parulekar S D 1982 Agar gel micro Ouchtorlony radial double diffusion
(AGD). PP 32 – 65. In gel immuno diffusion techniques in
Research and Laboratory Medicine (eds S S Kelkar and P M
Khare). Popular Prakashan Private Limited, Bombay.
Pike R M and Sulkin S E 1957 The agar diffusion precipitation test as a
means of recording evidence of meat adulteration. Journal of
laboratory clinical Medicine 49: 657. Cited from Swart and Wilks
1982.
Pinto F C 1961 Serological identification of ox, buffalo, goat and deer
flesh. British Veterinary Journal 117 : 540.
Poli G, Point A, Balasari A and Cantom C 1977 Ind Aliment 16: 87 cited
from Swart and Wilks 1982.
Prasad V S S 1974 Studies on esterase and other proteins of raw meat of
different species of animals in relation to age and sex M.V.Sc
Thesis, G B Pant University of Agriculture and Technology,
Pantnagar, Uttar Pradesh, India.
Radhakrishna K Vijaya Rao D, Sharma T R 1989 Characterization of the
major component in thermostable muscle proteins. Journal of
Food Science and Technology 26:32.
Radhakrishna K, Vijaya Rao D and Sharma T R 1988 studies on the
electrophoretic and immunodiffusion methods in the
differentiation of mutton and beef subjected to severe thermal

cxxxiv
processing. Journal of Food Science and Technology 25: 280 –
284.
Ramadas P 1972 Physico – chemical and immunological differentiation
of meats of different species of animals. M.V.Sc. Thesis, G B Pant
University of Agriculture and Technology, Pant nagar, Uttar
Pradesh, India.
Ramadas P and Misra D S 1981 Immunoelectrophoretic identification of
meats of bullock, goat, sheep, pig, chicken. Indian Veterinary
Journal 58: 978 – 983.
Rasool T J, Appa Rao K B C, Bhat P P, Rawat A K and Bhat P N 1995
Simple method for differentiation of raw meat from various meat
animals. Potential technique for detection of meat adulteration
Journal of Animal Science 65: 684 – 687.
Rattire H W and Regenstein J M 1977 Action of crude papain on actin
and myosin heavy chains isolated from chicken breast muscle.
Journal of Food Science 42 : 1159.
Reddy P M 1986 Studies on the identification of cooked meats of cattle,
sheep / goat Ph.D. Thesis Haryana Agricultural University, Hisar.
Reddy P M and Reddy G M 1995 Immunochemical detection of pork
using thermostable muscle antigens. Journal of Food Science and
Technology 32: 221 – 223.
Reddy P M and Mandokhot U V and Chandirmani N K 1990 A
comparative study on heated adrenal and immunoglobin G –
antigen for identification of cooked meats of cattle and sheep.
Journal of Food Science and Technology 27: 221 – 223.
Saisekhar R and Reddy P M 1995 Use of troponin for species
identification of cattle and buffalo meats. Journal of Food Science
32: 68-70.
Sasidhar S 1995 Detection of pork by immunochemical techniques
M.V.Sc Thesis, Andhra Pradesh Agricultural University,
Hyderabad.

cxxxv
Saway W N, Maneesh M S, EL – Rayes, Hussain A and Dashi B 1990
Detection of pork in processed meat by enzyme linked immuno
sorbant assay using anti swine antisera. Journal of Food Science
55: 293.
Schweiger A, Hanning K, Gunther H O and Bandner S 1982
Immunological detection of meat from turkey. In Recent
developments on food analysis. Food Science and Technology
Abstracts 15 : 6S 806, 1983.
Scopes R K and Penny I F 1971 Sub unit sizes of muscle protein as
determined by sodium dodecyl sulphate gel electrophoresis,
Biochemi. Bipphys, Acta, 236:409.
Sherikar A T, Ajinkya S M Khot J B, and Vaidya A M 1979
Differentiation of meats by gel diffusion technique. Indian Journal
of Animal Science 49 : 350.
Sherikar A T Khot J B Jayarao B M and Pillai S R 1987 Species
differentiation in raw and heat treated meats using serological
methods (Agar gel precipitation test). Journal of Food Science and
Technology 24:293.
Sherikar A T, Karkare U D, Khot J B, Jayarao B M and Bhilogaonkar N
Y 1993 Studies on thermostable antigens production of species
specific anti – adrenal sera and comparison of immunological
techniques in meat speciation. Meat Science 33: 121 – 136.
Sherikar A T, Khot J B, Jayarao B M and Pillai S R 1988 Use of species
specific antisera to adrenal heat stable antigens for the
identification of raw and cooked meats by agar gel diffusion and
counter immunoelectrophoretic techniques. Journal of the Science
of Food and Agriculture 44: 63.
Sinclair A J and Slattery W J 1982 Identification of meat according to
species by isoelectric focusing. Australian Veterinary Journal 58:
79.

cxxxvi
Sivakumar K Shanmugam A M and Ramaswami A M 1999 Souvenir
Cum proceedings. National seminar on processing of meat,
poultry and Byproducts for value addition.
Sridharlingareddy V 1996 Use of Brain antigens for species
identification of beef, cara beef and pork, M.V.Sc. Thesis Andhra
Pradesh Agricultural University, Hyderabad.
Srinivasarao Z 1996 Immunochemical detection of mutton, chevon and
rabbit using brain antigens. M.V.Sc. thesis, Andhra Pradesh
Agricultural University, Hyderabad.
Suryarao K 1990 Identification of species origin of cooked meats using
antisera to testicular antigens Ph.D. Thesis Andhra Pradesh
Agricultural University, Hyderabad.
Umamaheswararao S 2003: Personal Communication.
Walker, J.M. 1986 Protein (Western) blotting with immuno detection. In
experiments in Molecular Biology, R.J Slater (ed), PP. 171 – 178.
Humana Press, Clifton, New Jersey U.SA.
Warnecke M O and Sarfle R L 1968 Serological identification of
proteins. Mode of injection and protein extracts for antibody
production Journal of Food Science 33: 131.
Williams C A and Chase M W 1967 Methods in Immunology and
Immunochemistry. PP 246. Volume I and Volume II. Academic
Press, New York and London.
Wilson P J and White N B 1999 “Identification and quantification of
game tissue in processed meat products”. Northwest Association
of Forensic Scientists, Abstracts, Fall 1999.
Wintero A K, Thomsen P D and Davies W 1990 A comparison of DNA
–hybridization, immuno diffusion, counter immuno
electrophoresis and isoelectric focusing for detecting the
admixture of pork to beef. Meat Science 27: 75 – 85.

cxxxvii
Yamamato K, Kobayashi N and Samazima K 1982 Detection of
Soyabean protein in meat products and SDS gel electrophoresis.
Journal of college of Dairying Natural Science 94: 25.

cxxxviii
APPENDICES
Appendix – I
Buffers and reagents
Phosphate buffered saline (PBS), pH 7.3 (Cruickshank, 1965)
NaCl - 8.0 g
K2HPO4 - 1.21 g
KH2PO4 - 0.34 g
Distilled water - 1 Litre
Sodium phosphate buffer saline
Solution A: 0.2 M Monobasic sodium phosphate (27.8 g / Lt NSS)
Solution B: 0.2 M Dibasic sodium phosphate (53.65 g / Lt NSS)
51 ml of solution A was mixed with 49 ml of solution B and pH
adjusted to 6.8 and was finally diluted to a total of 200 ml with NSS.
Sodium barbital buffer
Sodium barbital - 10.31 g
Barbituric acid - 1.84 g
Sodium acetate - 6.80 g
Distilled water to make - 1 Litre
Ionic strength - 0.1
pH - 8.6
The entire solution was diluted to 4 litre with the distilled water.
118

cxxxix
Physiological sodium chloride, ionic strength 0.15, pH 7.4 (Crowle,
1973)
Sodium chloride - 0.88 g
Distilled water - 100 ml
2% Sodium chloride
Sodium chloride - 2 g
Distilled water - 100 ml
Appendix – II
Staining solutions
(a) Amido black stain:
Amido black - 1.5 g
Methanol - 90 ml
Glacial acetic acid - 10 ml
Distilled water - 90 ml
(b) Coomassie brilliant blue stain:
Coomassie brilliant blue - 5 g
Ethanol - 450 ml
Glacial acetic acid - 100 ml
Distilled water - 450 ml
Destaining solutions
(a) For amido black stain:

cxl
Methanol - 45 ml
Acetic acid - 5 ml
Distilled water - 50 ml
(b) For coomassie brilliant blue stain:
Ethanol - 450 ml
Glacial acetic acid - 100 ml
Distilled water - 450 ml
120














![772 IEEE TRANSACTIONS ON INFORMATION FORENSICS AND ... Papers... · verification scheme (localization-based defense) was proposed to detect PUEA. In [10] and [11], the authors proposed](https://img.dokumen.tips/doc/110x75/5fd20540e5402e2403551a05/772-ieee-transactions-on-information-forensics-and-papers-veriication.jpg)













