Embed Size (px)
Citation preview

HAL Id: hal-00895656https://hal.archives-ouvertes.fr/hal-00895656
Submitted on 1 Jan 2007
HAL is a multi-disciplinary open accessarchive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come fromteaching and research institutions in France orabroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, estdestinée au dépôt et à la diffusion de documentsscientifiques de niveau recherche, publiés ou non,émanant des établissements d’enseignement et derecherche français ou étrangers, des laboratoirespublics ou privés.
Studies of casein micelle structure: the past and thepresent
Phoebe X. Qi
To cite this version:Phoebe X. Qi. Studies of casein micelle structure: the past and the present. Le Lait, INRA Editions,2007, 87 (4-5), pp.363-383. �hal-00895656�

Lait 87 (2007) 363–383 Available online at:c© INRA, EDP Sciences, 2007 www.lelait-journal.orgDOI: 10.1051/lait:2007026
Review
Studies of casein micelle structure:the past and the present
Phoebe X. Qi*
Eastern Regional Research Center, Agricultural Research Service, U.S. Department of Agriculture,600 East Mermaid Lane, Wyndmoor, PA 19038, USA
Abstract – At the heart of the milk system are the colloidal casein-calcium-transport complexestermed the casein micelles. The application of physical chemical techniques such as light, neutron,and X-ray scattering, and Electron Microscopy (EM) has yielded a wealth of experimental detailconcerning the structure of the casein micelle. From these experimental data bases have arisen twoconflicting models for the internal structure of the casein micelle. One model emphasizes proteinsubmicellar structures as the dominant feature, while the other proposes that inorganic calciumphosphate nanoclusters serve this function. In this study, these models are critically examined inlight of the two current primary dogmas of structural biology which are: protein structure givesrise to function and that competent and productive protein-protein interactions (associations) willlead to efficient transit through the mammary secretory apparatus. In this light an overwhelmingargument can be made for the formation of proteinaceous complexes (submicelles) as the formativeagents in the synthesis of casein micelles in mammary tissue. Whether these submicelles persist inmilk has been questioned. Recently we have carried out studies on casein micelles and submicellesusing Atomic Force Microscopy (AFM) and high resolution Transmission Electron Microscopy(TEM) to gain insights on the nature of protein-protein interactions in submicelles and micellesfrom a structural biology perspective. The results provide experimental evidence that protein-proteininteractions are important in the formation and stabilization of casein micelles.
milk / micelle / casein / structure / protein-protein interaction
摘摘摘要要要 –酪酪酪蛋蛋蛋白白白胶胶胶束束束结结结构构构研研研究究究的的的回回回顾顾顾与与与展展展望望望。。。酪蛋白胶束是由酪蛋白与钙盐形成胶体,酪蛋白胶束是乳系统的核心。采用物理化学方法,如光、中子、X-射线散射及电子显微镜技术等方法对酪蛋白胶束结构给予了详细的分析。现有的资料对酪蛋白内部结构的解释已经形成了两个互相矛盾的模型,一种是强调酪蛋白结构的亚胶束模型,而另一种是认为无机磷酸钙和酪蛋白形成的纳米簇结构。在本研究中基于两种目前公认的结构生物学理论,即蛋白的结构赋予其功能性和高效的蛋白-蛋白之间相互作用 (凝聚)导致其通过乳腺分泌器官进行有效的转移过程,对这些模型进行了严格的验证。目前更多的理论观点认为在乳腺组织中的酪蛋白胶束是以蛋白质复合物也就是亚胶束结构存在。但是亚胶束结构观点受到了质疑。近来我们采用原子力显微镜 (AFM)和高分辨率透射电镜 (TEM)从结构生物学性质的角度对酪蛋白胶束和亚胶束中蛋白质之间的相互作用进行了研究。结果证明了蛋白-蛋白的相互作用是胶束形成和稳定的重要因素。
乳乳乳 /胶胶胶束束束 /酪酪酪蛋蛋蛋白白白 /结结结构构构 /蛋蛋蛋白白白-蛋蛋蛋白白白相相相互互互作作作用用用
* Corresponding author (通讯作者): [email protected] of brand or firm name does not constitute an endorsement by the U.S. Department ofAgriculture over others of a similar nature not mentioned.
Article published by EDP Sciences and available at http://www.lelait-journal.org or http://dx.doi.org/10.1051/lait:2007026

364 P.X. Qi
Résumé – Structure de la micelle de caséines : études passées et actuelles. Les micelles de ca-séines, complexes colloïdaux caséine/calcium, sont au cœur de l’organisation moléculaire du lait.L’application de techniques physico-chimiques comme la diffraction de lumière, de neutrons et derayons X et la microscopie électronique a permis d’obtenir une mine de renseignements expérimen-taux concernant la structure de la micelle de caséines. Ces résultats ont abouti à l’élaboration dedeux modèles opposés sur la structure interne de la micelle de caséines. L’un d’eux met en avantdes structures sub-micellaires des caséines comme caractéristique prédominante, tandis que l’autrepropose une organisation sous forme de nano-clusters (amas) de phosphate de calcium inorganique.Dans cette étude, ces modèles sont examinés et discutés à la lumière des deux dogmes élémen-taires de la biologie structurale actuelle, à savoir que la structure protéique détermine la fonction,et que des interactions (associations) protéines/protéines adéquates conduiront à un acheminementefficace au sein de l’appareil sécréteur de la glande mammaire. À la lumière de ces éléments, onpeut suggérer que la sub-micelle est l’élément de base dans la synthèse des micelles de caséinesdans le tissu mammaire. La question reste de savoir si ces sub-micelles persistent dans le lait. Nousavons conduit récemment des études sur les micelles et les sub-micelles à l’aide de la microsco-pie à force atomique (AFM) et la microscopie électronique à transmission haute résolution (TEM)pour avoir une visualisation de la nature des interactions protéines/protéines dans les sub-micelleset les micelles du point de vue de la biologie structurale. Les résultats obtenus démontrent que lesinteractions protéines/protéines sont importantes dans la formation et la stabilisation des micellesde caséines.
lait / micelle / caséine / structure / interaction protéique
1. INTRODUCTION
The virtual image of milk, which wouldbe constructed by most people, is that ofa creamy white fluid. The lubricity andtaste of milk are related to this perceptionand are based upon three unique biologicalstructures: the colloidal calcium-proteincomplexes (the casein micelles), the milkfat globules with their limiting membrane,and the milk sugar: lactose [51]. The com-plexity of these structures is necessitatedby the fact that milk is in essence predom-inantly water. It is the accommodation ofthese ingredients to an aqueous environ-ment that forms the basis for the structureof milk at the molecular level and calls forthe unique secretory process: milk synthe-sis [51].
Although these colloids have been thesubject of extensive research for manyyears [15, 26, 28, 30, 36, 47, 48, 50, 52, 54,62, 68] the structural details of the caseinmicelles on the molecular level remain elu-sive [24]. Biochemical and physical studiesof these colloids have focused on: the sizeand properties of the colloids, their pro-tein and mineral composition, and stepwise
reconstitution of the micelles. Conflictingmodels for the structure of the casein mi-celles have arisen from differing interpre-tations of the core data bases developed inthese studies. This manuscript will focuson the recent advancement in the field ofstructural biology and protein-protein in-teractions will be applied to attempt to dis-cern the biologically competent route forthe formation of the casein micelles.
2. BRIEF REVIEW OF PROPOSEDMODELS FOR CASEINMICELLES
2.1. Protein composition of skim milk
The dominant constituent in skim milkis the casein micelle (Fig. 1a). It isthis unique supramolecular aggregate im-parts the opalescence characteristic of skimmilk. The chief function of the micelle is tofluidize the casein molecules and solubilizethe calcium and phosphate [16]. In gen-eral, when milks that contain > 2% proteinare analyzed, the accompanying inorganicphosphate and calcium levels found would

Casein micelle structure 365
(a)
(b)
Figure 1. Transmission Electron Micrograph of bovine milk casein micelles (a) fixed with glu-taraldehyde and stained with uranyl acetate and lead citrate (pH 7.0). Inset at the upper right showsan enlarged single micelle. (b) The submicellar model for casein micelles showing proteinaceousstructures and surface arrangement of κ-casein; reprinted from Schmidt [54].

366 P.X. Qi
Table I. Casein distribution (%) in variousmilks.
αs1- αs2- β- κ-Goats 5 to 17 6 to 20 50 15Cows 38 10 40 12Human Trace Trace 70 27
by themselves, yield insoluble precipitates(apatite or brushite depending upon thepH). Conversely in the absence of thesesalts, the casein components, as a result oftheir open structures, have a high viscosity.The formation of the casein colloidal com-plexes, micelles, averts these two prob-lems.
Four major casein components in cow’smilk, αs1-, αs2-, β- and κ-casein, have beenwell characterized in the past [59]. Ca-seins have been found to be homologousto these proteins in their gene and primaryprotein sequences in all species examinedto date [23]. However, the proportions ofthe various caseins vary widely. β-Caseinis the primary casein in human milk and ingoats’ milk it comprises 40% to 50% of thecasein, for example. In goats’ milk thereis also a high degree of variance in caseinproportions among animals, which appearsto be genetically controlled (Tab. I). De-spite the variations in casein componentsall species competently form colloidal ca-sein micelles for the transport of calciumand phosphate. At the ultra structural level,the casein micelles of most species appearquite similar.
The αs1-, αs2- and β-caseins are pre-cipitated by calcium (calcium-sensitive)binding to their phosphoserine residues atthe concentrations of protein and calciumfound in most milks. However, κ-caseinis not only soluble in calcium (calcium-insensitive), but also interacts with and sta-bilizes the calcium insoluble caseins to ini-tiate formation of the stable colloidal state.It has been generally recognized by stud-ies [7,10,13,29] using numerous chemical,
enzymatic and immunological techniquesthat while the majority of the κ-casein re-side on the surface of the casein micelles,other caseins might also occur there aswell. In all models for casein structure,κ-casein is thought to predominate on themicellar surface (Fig. 1b). In milk clot-ting in the neonate stomach, the enzymechymosin (rennin) specifically cleaves onebond in κ-casein to initiate aggregation ofthe micelles. It has been clearly demon-strated recently [55] by κ-casein gene nullmutation experiments in mice that κ-caseinis essential for the assembly of the caseinmicelles and for lactation to occur in vivo.
2.2. The submicelle theory of caseinstructure
For many years the most accepted the-ory [15, 54] of the structure of the caseinmicelle was that it was composed of spher-ical aggregates of the caseins (submicelles)held together by calcium-phosphate link-ages (Fig. 1b). The submicelle hypothe-sis has been historically supported by bio-chemical and biophysical studies [39, 40,42, 57] on the individual casein compo-nents, reconstitution of micelles from theircomponent caseins [27, 41, 56], as well aselectron microscopy of the micelles them-selves [8] and partially decomposed mi-celles. Early studies on the purified ca-seins demonstrated that in the absence ofcalcium they formed rather large aggre-gates (submicelles) and that these aggre-gates formed colloidal complexes in thepresence of added calcium. By gradualdialysis and direct addition of EDTA to re-move calcium from casein micelles, Linet al. [45] proposed possible emergenceof casein micellar framework or subunitsas later substantiated by physical tech-niques and freeze fracture electron mi-croscopy [41]. These particles had phys-ical properties similar to the aggregatesfound in whole casein preparations in the

Casein micelle structure 367
absence of calcium and were consideredas submicellar in nature. Based on thesedata, Schmidt proposed the “submicelle”model [54] in Figure 1b. In summary thismodel stipulates protein-protein interac-tions, traps calcium phosphate and formsthe colloidal complexes.
In recent years this theory has beenchallenged by concepts [30, 35] arisingfrom the study of the casein-calcium-phosphate interactions, the micelles them-selves and physical chemical studies of theindividual proteins at interfaces.
2.3. Casein micelle models with aninternal gel matrix
Two more recently proposed models forthe casein micelle have emerged that refutethe notion of discrete submicellar struc-tures within the micelle. The first model todepart from the submicelle theory was thatof de Kruif and Holt [12]. During a seriesof studies on casein-calcium-phosphate in-teractions Holt and coworkers [31, 33] dis-covered that the phosphopeptide fractionof β-casein could bind to and stabilizecalcium-phosphate aggregates resulting inthe formation of nanoclusters of a dis-crete size and composition; without thepeptides the calcium phosphate structureswould grow randomly and precipitate. Thisdiscovery led de Kruif and Holt to pro-pose that such nanoclusters are the cen-terpiece of casein micelle structure [14].The formation of nanoclusters with a ra-dius of 2.3 nm would drive micelle forma-tion by randomly binding phosphoproteinscausing an inverted micelle, and then moreproteins could coat this new hydrophobicsurface and in turn, bind more calciumphosphate until a size limited colloid isformed. There are about 800 of these amor-phous calcium phosphate nanoclusters inan average sized casein micelle (∼100 nmin diameter), see Figure 2a. This nanoclus-ter model is supported by the earlier rheo-
morphic theory of casein structure [32]and the recent SAXS and USAXS data byPignon et al. [53]. In this view, the un-structured proteins form about the amor-phous inorganic species and their functionof binding to the calcium-phosphate givesrise to their structure, hence no specificprotein secondary structures or protein-protein interactions are invoked, exceptthat a surface position for κ-casein is re-quired.
The casein micelle model proposed byHorne [35] (Fig. 2b) considered the sur-face chemistry of the individual caseinsand concluded that protein-protein interac-tions were indeed important, but in essencethe model retains the rheomorphic con-cept. In this view the amphiphilic natureof the caseins causes them to act more asblock copolymers of alternating charge andhydrophobicity, that is, a charged phos-phopeptide loop and a hydrophobic trainfor β-casein, the reverse for κ-caseins, andan N-terminal hydrophobic train, followedby a charged loop and a final C-terminalhydrophobic train for αs1-casein. Individ-ual secondary structural elements of theeach casein are ignored and only gross hy-drophobic interactions are considered. Inthis model, the growth of the calcium phos-phate nanoclusters begins the process ofmicelle formation, but it is limited by bind-ing to the phosphopeptide loop regions ofthe caseins. Once bound to the amorphousinorganic matrix, further protein-proteininteractions are related to the hydropho-bic blocks and polymerization proceeds byrepeating the entire process. Micelle for-mation leads to an internal gel-like struc-ture with embedded nanoclusters of cal-cium and phosphate, and the reaction ofκ-casein which contains only one phos-phoserine residue limits micellar growthby acting as a dead end capping unit inanalogy with the growth of synthetic poly-mers. Note that this model shows onlymonomeric κ-casein molecules.

368 P.X. Qi
(a)
(b)
Figure 2. (a) The nanocluster model for casein micelles. Casein monomers are thread-like, whilethe dark circles represent calcium phosphate nanoclusters; reprinted from [12] with permissionof Springer, NL. (b) The model of Horne – dual bonding model of casein micelles using caseinmonomers as indicated. Protein-protein interactions occur between hydrophobic regions (rectangu-lar bars) while the protein hydrophilic regions (loops) bind to calcium phosphate clusters (triangles).κ-Casein is monomeric and on the surface; reprinted with permission from Elsevier.

Casein micelle structure 369
Interestingly, these two distinctly differ-ent views of the internal structure of thecasein micelles-submicelles versus gel ma-trices arise essentially from the same bio-chemical and physical chemical data bases.For a further exposition on their similar-ities and differences see the excellent re-view of Walstra [67].
3. SYNTHESIS AND SECRETIONOF CASEINS
3.1. Cell physiology
The evolution of the mammary gland,presumably from external sweat glands,has yielded a great variety of exterior ap-pearances in many species [51], but at thetissue level there is a common organiza-tional theme as shown in Figure 3a. Mam-mary secretory cells are epithelial in na-ture and are arranged in alveoli which areconnected to ductal tissue. The secretoryepithelial cells (SEC) are surrounded by alayer of myoepithelial cells, which are ableto contract and expel milk into the ductsin response to the hormone oxytocin. Thealveoli are highly vascularized to ensure aconstant flow of the metabolic precursorsneeded for milk synthesis and secretion.Finally the vascularized alveoli are embed-ded in an extracellular matrix. This matrixnot only supports the cells, but also throughcell-cell interactions is responsible for thefull expression of the genes that controlmilk synthesis [51].
3.2. Protein synthesis and secretion:overview
Adaptation of milk components to theirultimate aqueous environment begins dur-ing secretion. Lipid and protein synthe-sis are partitioned from the start. Aminoacids and their metabolic precursors are ac-tively transported into the SEC and assem-bled into proteins on the ribosomes of the
highly developed rough endoplasmic retic-ulum [51]. All milk proteins of mammaryorigin have conserved leader sequenceswhich cause insertion of the nascent pro-teins into the lumen of the endoplasmicreticulum (ER) shown in Figure 3b. Theproteins are then transported through theGolgi apparatus [15, 51] as shown in Fig-ure 3b; presumably the globular proteinsof milk are folded during this period. Inthe Golgi apparatus, the caseins, which arethe major milk proteins in most species,appear to be spherical complexes of about10 nm in diameter. The caseins are phos-phorylated by a calcium-activated mem-brane bound kinase to begin the processof calcium transport [3]. A membrane as-sociated ATPase delivers calcium to thevesicles [5]. The gradual intercalation ofcalcium, casein, and phosphate into thesubmicellar structures leads to the forma-tion of casein micelles and insures the ef-fective transport of these vital minerals.This process can be visualized in Fig-ure 4 (top) where small submicellar parti-cles are seen in the secretory vesicles near-est the trans Golgi. Through the bindingof calcium and the accretion of phosphate,the colloidal casein micelles are formedand finally secreted by reverse pinocytosis(Fig. 4, bottom). Overall this view stronglysupports the involvement of proteinaceoussubmicellar structures in the synthesis andsecretion of casein micelles.
3.3. Protein synthesis and secretion:details in the ER lumen
The process of casein secretion withinthe lumen of the ER has not been stud-ied in specific detail [24]; however refer-ence to recent information on cell biologyand protein folding in other systems mayshed more light on the issues of caseinmicelle formation. The issue of qualitycontrol of protein folding has become awidely researched area in cell biology [49].

370 P.X. Qi
b
c br
d
a
p
c
Fig
ure
3.C
ell
phys
iolo
gyof
lact
atin
gm
amm
ary
glan
d.A
sing
leal
veol
us(a
)co
nsis
ting
ofla
ctat
ing
epit
heli
alce
lls
(SE
C)
surr
ound
ing
the
lum
en.
Aty
pica
lla
ctat
ing
cell
(b)
indi
cati
ngac
tive
secr
etio
nof
prot
ein
and
lipi
dby
dist
inct
mec
hani
sms;
repr
inte
dw
ith
perm
issi
onof
Sci
enti
fic
Am
eric
anan
dS
.Pat
ton.
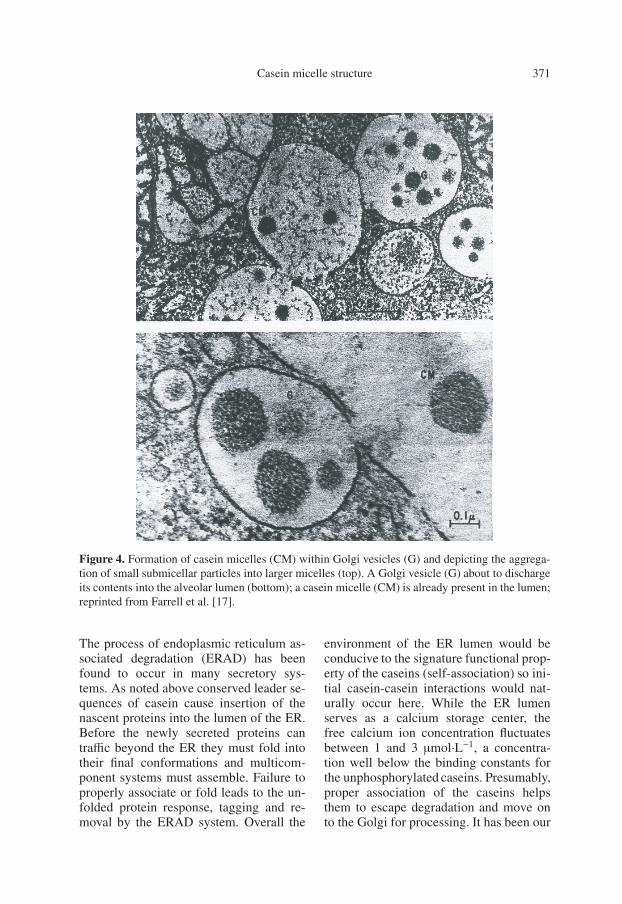
Casein micelle structure 371
Figure 4. Formation of casein micelles (CM) within Golgi vesicles (G) and depicting the aggrega-tion of small submicellar particles into larger micelles (top). A Golgi vesicle (G) about to dischargeits contents into the alveolar lumen (bottom); a casein micelle (CM) is already present in the lumen;reprinted from Farrell et al. [17].
The process of endoplasmic reticulum as-sociated degradation (ERAD) has beenfound to occur in many secretory sys-tems. As noted above conserved leader se-quences of casein cause insertion of thenascent proteins into the lumen of the ER.Before the newly secreted proteins cantraffic beyond the ER they must fold intotheir final conformations and multicom-ponent systems must assemble. Failure toproperly associate or fold leads to the un-folded protein response, tagging and re-moval by the ERAD system. Overall the
environment of the ER lumen would beconducive to the signature functional prop-erty of the caseins (self-association) so ini-tial casein-casein interactions would nat-urally occur here. While the ER lumenserves as a calcium storage center, thefree calcium ion concentration fluctuatesbetween 1 and 3 μmol·L−1, a concentra-tion well below the binding constants forthe unphosphorylated caseins. Presumably,proper association of the caseins helpsthem to escape degradation and move onto the Golgi for processing. It has been our

372 P.X. Qi
contention that conserved sequences of theindividual caseins give rise to selected sec-ondary structural elements and that theseelements lead to self-association of the ca-seins without classical protein folding [20,21], that is to say, caseins contain littleor no tertiary structure and proceed di-rectly from secondary structure formationto quaternary structures exhibiting bothrigid and flexible elements. This is in linewith the basic tenet of structural biologythat protein structure gives rise to biolog-ical function (the Anfinsen hypothesis [2]).Recent development in the field of pro-tein folding has shed light on understand-ing protein self-assembly and aggregationas delineated by Jaenicke and Lilie [37].Self-associations are those on-line and pro-ductive reactions which lead to competentbiological protein assemblies such as amy-loid [38], whereas non-productive aggrega-tions lead to often mis-assembled proteincomplexes. The latter would be targetedfor the ERAD process. The question nowarises as to which of the many known invitro reactions of the caseins studied todate are important in this context. It may beof interest to examine those “productive”self-associations which may lead to the for-mation of casein micelles and those “non-productive” aggregations formed throughvarious protein-protein interactions.
The self-association of bovine β-caseinhas been studied by many (for exam-ple [36]). The most commonly acceptedmechanism is that the β-casein moleculesform rather spherical polymers of lim-ited size following a critical micelle path-way. Interestingly the dimensions of thispolymer are rather fixed, but its molecu-lar weight is highly dependent on ionicstrength and temperature. In species suchas human where β-casein is the predomi-nate protein, this process could be viewedas a prominent on-line self-association inthe ER lumen. It must be noted thatthe unphosphorylated form of the humanβ-casein has nearly the same propensity
for self-association as its phosphorylatedforms [6]. The weight average molecularweight of bovine β-casein at 37 ◦C in theabsence of calcium is 1 250 000 as shownin Table II. This is almost three times theestimated values for the submicellar struc-tures observed in Figure 4 and could poten-tially lead to ER stress and ERAD tagging.
Studies on the polymerization ofαs1-casein had previously shown [1]that the molecule exhibits a progres-sive consecutive association to dimers,tetramers, hexamers, etc., and this processis highly dependent on pH and ionicstrength. Most of the early studies [54]on the polymerization of αs1-casein wereconducted at or below 25 ◦C; because ofits hydrophobic nature it was expectedthat polymerization would be accentuatedat 37 ◦C. This clearly is not the case at37 ◦C as shown in Table II; all three majorgenetic variants depolymerize [46] andbehave essentially as dimers. The dimerformation is centered on its C-terminal halfwith a strong selective interaction betweenresidues 136 and 160 of each monomer,and there appears to be no involvement ofN-terminal hydrophobic train of αs1-caseinat normal ionic strength and 37 ◦C. Inaddition, the latter region is positivelycharged and could participate in phosphatebinding. This appears to contradict oneof the modes of crosslinking postulatedby Horne [35] as part of the casein gelnetwork. Finally it has been shown [4]that both native and dephosphorylatedαs1-caseins undergo similar aggregationand precipitation reactions. Thus in theER lumen the αs1-casein molecule wouldnot be greatly polymerized and the higherorder polymers observed in vitro are theproducts of an aggregation process, whichwould likely be “non-productive”.κ-Casein is the calcium-soluble stabiliz-
ing protein of the casein micelles; it is alsothe only casein whose disulfide bonds playa significant role in casein structure. Asisolated from milk, the protein displays a
Casein micelle structure 373
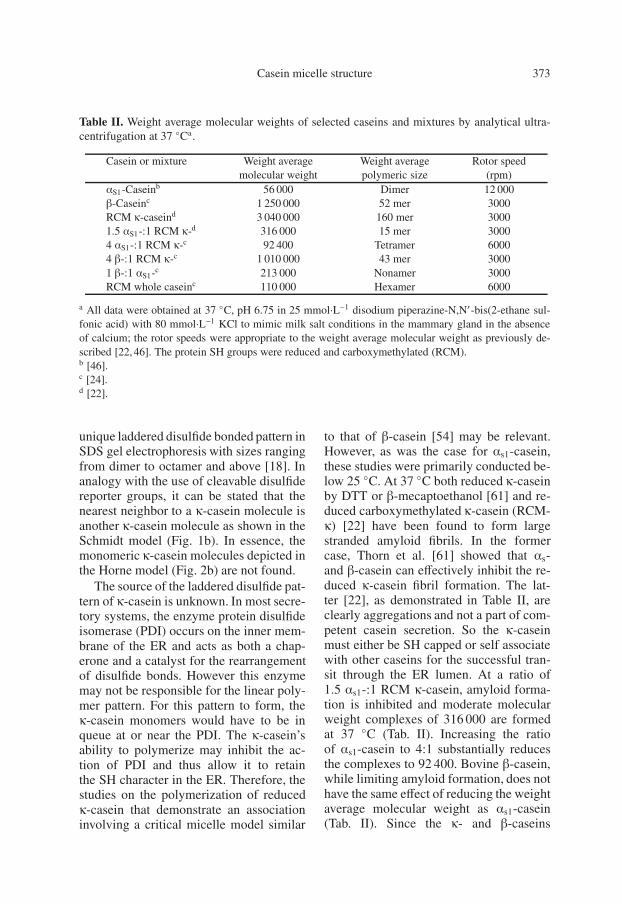
Table II. Weight average molecular weights of selected caseins and mixtures by analytical ultra-centrifugation at 37 ◦Ca.
Casein or mixture Weight average Weight average Rotor speedmolecular weight polymeric size (rpm)
αS1-Caseinb 56 000 Dimer 12 000β-Caseinc 1 250 000 52 mer 3000RCM κ-caseind 3 040 000 160 mer 30001.5 αS1-:1 RCM κ-d 316 000 15 mer 30004 αS1-:1 RCM κ-c 92 400 Tetramer 60004 β-:1 RCM κ-c 1 010 000 43 mer 30001 β-:1 αS1-c 213 000 Nonamer 3000RCM whole caseinc 110 000 Hexamer 6000
a All data were obtained at 37 ◦C, pH 6.75 in 25 mmol·L−1 disodium piperazine-N,N′-bis(2-ethane sul-fonic acid) with 80 mmol·L−1 KCl to mimic milk salt conditions in the mammary gland in the absenceof calcium; the rotor speeds were appropriate to the weight average molecular weight as previously de-scribed [22, 46]. The protein SH groups were reduced and carboxymethylated (RCM).b [46].c [24].d [22].
unique laddered disulfide bonded pattern inSDS gel electrophoresis with sizes rangingfrom dimer to octamer and above [18]. Inanalogy with the use of cleavable disulfidereporter groups, it can be stated that thenearest neighbor to a κ-casein molecule isanother κ-casein molecule as shown in theSchmidt model (Fig. 1b). In essence, themonomeric κ-casein molecules depicted inthe Horne model (Fig. 2b) are not found.
The source of the laddered disulfide pat-tern of κ-casein is unknown. In most secre-tory systems, the enzyme protein disulfideisomerase (PDI) occurs on the inner mem-brane of the ER and acts as both a chap-erone and a catalyst for the rearrangementof disulfide bonds. However this enzymemay not be responsible for the linear poly-mer pattern. For this pattern to form, theκ-casein monomers would have to be inqueue at or near the PDI. The κ-casein’sability to polymerize may inhibit the ac-tion of PDI and thus allow it to retainthe SH character in the ER. Therefore, thestudies on the polymerization of reducedκ-casein that demonstrate an associationinvolving a critical micelle model similar
to that of β-casein [54] may be relevant.However, as was the case for αs1-casein,these studies were primarily conducted be-low 25 ◦C. At 37 ◦C both reduced κ-caseinby DTT or β-mecaptoethanol [61] and re-duced carboxymethylated κ-casein (RCM-κ) [22] have been found to form largestranded amyloid fibrils. In the formercase, Thorn et al. [61] showed that αs-and β-casein can effectively inhibit the re-duced κ-casein fibril formation. The lat-ter [22], as demonstrated in Table II, areclearly aggregations and not a part of com-petent casein secretion. So the κ-caseinmust either be SH capped or self associatewith other caseins for the successful tran-sit through the ER lumen. At a ratio of1.5 αs1-:1 RCM κ-casein, amyloid forma-tion is inhibited and moderate molecularweight complexes of 316 000 are formedat 37 ◦C (Tab. II). Increasing the ratioof αs1-casein to 4:1 substantially reducesthe complexes to 92 400. Bovine β-casein,while limiting amyloid formation, does nothave the same effect of reducing the weightaverage molecular weight as αs1-casein(Tab. II). Since the κ- and β-caseins

374 P.X. Qi
share a similar self-association mecha-nism, it would appear that in mixed asso-ciations the RCM κ-casein can be insertedinterchangeably into the self-associationreaction of β-casein; but the resulting over-all size is somewhat smaller than that ofβ-casein alone (Tab. II). Such large com-plexes might not allow the associated pro-teins to escape ERAD and move on to theGolgi apparatus. In fact Chanat et al. [9]have studied the transport of caseins fromthe ER to the Golgi apparatus in mammaryepithelial cells. Their data suggest that foranimals with a high casein content, αs1-casein must interact with the other caseinsfor efficient transport to the Golgi. In cellsthat completely lack αs1-casein the accu-mulation of β-casein (or κ-β-mixtures?) isobserved in the ER. In the long term thiscauses ER stress, activates the ERAD sys-tem and impedes secretion. Recent workby Shekar et al. [55] demonstrated thatκ-casein is essential for the formation ofcasein micelles as well as for lactation tooccur.
To test the efficacy of αs1-casein at re-ducing the size of β-casein aggregates invitro, 1:1 mixtures of the two proteins werestudied by analytical ultracentrifugationat 37 ◦C. The weight average molecularweights of the complexes were speed de-pendent, increasing with decreasing speed,indicating strong hydrophobic interactions;see Table II. The weight average molecularweight of the 1:1 complexes was 213 000which represents a six fold reduction ofthe β-casein aggregate or a four fold in-crease over that of the αs1-casein alone.This result is of importance because thereare few studies of the mixed associationsof these two caseins at 37 ◦C and condi-tions close to that of the ER. Here the αs1-casein acts to diminish the size of either theβ- or κ-casein aggregates; in this sense itmay be considered a molecular detergentfor the other caseins. Thus these in vitrodata confirm the in vivo observations thatαs1-casein can reduce aggregated species
and allow the associated particles to escapethe ER. For human milk, the small amountof α-caseins present may help reduce theaggregates of β-casein; also the net ca-sein content in human milk is only 17% ofthat in bovine milk (Tab. I) so smaller ag-gregates would be favored. Finally, RCM-derived whole bovine casein, with the stan-dard ratios of the four caseins, has a weightaverage molecular weight of 110 000 at37 ◦C under the same conditions of pH andin the absence of calcium. From all of thesebiological and physical chemical studies itwould appear that for the competent syn-thesis and secretion of casein: preformedcasein complexes of the size of the puta-tive casein submicelles must form throughprotein-protein interactions – triggered byconserved protein sequences – and emergefrom the ER for efficient transit via secre-tory vesicles to the Golgi apparatus.
3.4. Protein synthesis and secretion:details in the Golgi vesicles
From the discussion above it seemsmost likely that the individual caseinmolecules undergo significant self-association in the ER and are thentransported in vesicles to the cis face ofthe Golgi apparatus. In this region threesignificant events occur in the process ofcasein secretion. The first event is mostlikely an increase in calcium concentra-tion accomplished by an ATPase-drivenpump [5]. The second most likely nextstep is the phosphorylation of the asso-ciated caseins by a membrane associatedcasein kinase which uses calcium-ATP assubstrate and is specific for Ser residuespreceded at the n + 2 position by Glu ora serine phosphate (SerP) residue [23].The casein kinase [3] responsible forthis reaction has not been purified, but inGolgi preparations the enzyme requiresa surprisingly high calcium ion concen-tration (KM ∼ 20 mmol·L−1) so that at

Casein micelle structure 375
the time of phosphorylation, calcium ionsmay be almost immediately bound to thecaseins (KD ∼ 5 mmol·L−1). The thirdand often overlooked step is that one ofthe byproducts of the kinase reaction,ADP, retains bound calcium, so that whenthis is converted by membrane associ-ated diphosphatases to phosphate andAMP [17], both calcium and phosphateare released near the interior membranesurface where the casein proteins are stillbeing phosphorylated. Studies by West andClegg [69] showed that phosphorylationof casein is still proceeding in large Golgivesicles as is most likely calcium transport.Thus both calcium and phosphate mayautomatically be bound to the submicellarcomplexes, as seen in Figure 4, prior tomicelle formation. In general, when milksthat contain > 2% protein are analyzed, theaccompanying inorganic phosphate andcalcium levels found would yield insolubleprecipitates (apatite or brushite) in theabsence of casein. But the question arisesare the concentrations of these compoundsever high enough or concentrated enoughto form nanoclusters in the Golgi vesicles?Additionally, would the energy gainedby coating these precipitating clusters besufficient to depolymerize the preformedcasein complexes in the manner suggestedby the Horne model? Veis [65] has sug-gested that, in general, mineralization inmammalian systems such as collagen anddentin matrices is directed and controlled(assembled) by the structural proteinspresent; inorganic direction appears to belimited to simpler systems such as thecrystalline shells of corals.
Based on the average composition ofthe colloidal caseinate [15] the averageconcentrations of calcium and phosphatewithin the colloidal complexes are 18.7and 15.2 mmol·L−1, respectively. In turnthe average concentration of casein withinthe colloidal complexes is 1 mmol·L−1 butthe average casein molecule has 6.5 phos-phate groups thus the concentration is
6.5 mmol·L−1 SerP. Moreover all fourcaseins have selected areas of positive sur-face [59] which may bind phosphate af-ter calcium binds to the protein as sug-gested for αs1-casein [46]. In all of theabove models, the binding of phosphatehas been largely neglected. The prominentpositive patches of the caseins are: 1–10 ofαs1- (+6); 165–199 of αs2- (+11); 97–113of β- (+6) and 97–116 of κ-casein (+6);these too average out to 6.5 mmol·L−1. Ifeach serine phosphate binds one calciumwhich in turn binds one inorganic phos-phate, then this would result in a doublelayer. Additionally if each positive chargein the above mentioned areas then boundone phosphate and one calcium in a sec-ond double layer, then the possible con-centrations of unbound calcium and phos-phate within the micelle is further reduced.It can be seen in a molecular dynamicsstudy [19] that both the smaller calciumions and the larger chloride ions bind tothe peptide and then associated ions tend toform a charged double layer about the pep-tide chain. In this case there is limited at-traction between the aqueous calcium andchloride ions, but complexes between cal-cium and phosphate would be more numer-ous. In vivo the formation of such a doublelayer of calcium phosphate would reducethe unbound (uninfluenced) concentrationsof these two ions to 5.7 and 2.2 mmol·L−1
respectively. This is far from the concentra-tions used by Holt and coworkers [31, 33]to form nanoclusters in vitro (37 mmol·L−1
calcium and 30 mmol·L−1 phosphate and3 mmol·L−1 phosphopeptide). In this samecontext the average submicelle would have65 negative charges [46] due to clustersof SerP groups and 65 positive chargesdue to clusters of basic amino acids, intotal it could carry 260 ions as a sim-ple double layer; the standard nanoclus-ter has about 355 ions at its core [31, 33].The coalescence of two submicelles withtheir bound (influenced) ions then wouldterminate any possible calcium phosphate

376 P.X. Qi
growth and begin micelle formation. Itmay be that nanoclusters then have aninteresting and informative chemistry butactually represent a process which is aninorganic aggregation similar to amyloidformation by κ-casein rather than an on-line productive biological process. Thusfrom the point of view of the synthesis andsecretion of casein micelles the Schmidtmodel may be representative of the biolog-ical process, although the stoichiometry ofthe inorganic “cement” is probably incor-rect based upon the latest physical chemi-cal data [11, 13] which indicates a type ofapatite is the most likely candidate for themolecular structure within the micelles.
4. STUDIES ON THE STRUCTUREOF CASEIN MICELLES
From the above discussion it is clearthat proteinaceous submicelles (here de-fined as associated casein species with anaverage size of 10 nm, not necessarilyspherical, Figs. 1a and 4) play a majorrole in the on-line formation of casein mi-celles in mammary tissue. Additionally itwould also appear that αs1-casein acts as atype of detergent to limit the size of thesesubmicellar particles in order to defeat theunfolded response and escape the ERADsystem. The formation of these controlledaggregates (productive association) allowsfor and facilitates transfer from the ER tothe Golgi apparatus. Once present in theGolgi, micelle formation occurs. Past re-search on understanding the detailed struc-ture of the micelles has centered on us-ing electron microscopy, neutron scatteringand X-ray scattering techniques. AtomicForce Microscopy (AFM) [25,64] has onlyrecently been applied to the study of thecasein micelles.
4.1. Electron microscopy
Electron microscopy (EM) represents apowerful tool for elucidation of biological
ultrastructures as seen in Figures 1a and 4.The main problem with this technique liesin the fixatives and metal staining used toaccentuate the particular features they re-act with and visualize, usually at the ex-pense of other features. In contrast, whenuranyl oxalate is used as a positive stainfor proteins [48] a more uniform distribu-tion of material is seen because the stainis binding to the caseins, particularly theSerP and accentuating the protein distribu-tion. However, more details can be visu-alized from our studies of casein micellesand sodium caseinate (Figs. 1a and 6).Similar strand-like protein structures witha “knot” about 10 nm (diameter) can befound in skim milk (Fig. 1a) and sodiumcaseinate, a common commercial caseinproduct without calcium (Fig. 5), the lat-ter perhaps exemplifies the prevalent ex-istence of casein-casein interactions evenafter the complete destruction of the mi-celles. In addition, Figure 4 shows whatcould be “submicellar” casein aggregatesin rat Golgi vesicles. Clearly, EM imagesare influenced by the stains used in the ex-periments. One is tempted to employ thescientific dialectic here and say that nei-ther the thesis (submicelles) nor the an-tithesis (no submicelles) is correct, but thatsynthesis is needed. Walstra [67] has pro-posed that the submicelles reemerge in EMrepresentations of products such as cheese.Work on a variety of cheeses [63] demon-strates the dynamic nature of the submicel-lar structures of the cheese protein matrix.
4.2. Micelle dissociation studies
Studies on micelle dissociation wereamong the first to indicate the existence ofsubmicelles and Schmidt [54] drew heav-ily on these and on reconstitution stud-ies in framing the model shown in Fig-ure 1b. Clearly the Walstra hypothesis onmicelle equilibria is at play in these ex-periments (Fig. 5). Those components in

Casein micelle structure 377
m
m c
Figure 5. Proposed scheme of the most important dynamic equilibria between casein micelles andmilk serum from Walstra [67]; reprinted with permission from Elsevier.
rapid equilibrium will quickly exchangeand yield one result, while those slow toequilibrate will accentuate another feature.It has also been suggested that calciumbinding to caseinates must precede phos-phate binding [66]. Temperature plays an-other role in that the aggregation of αs1-casein is, as noted above, accentuated atlower temperatures.
Finally, Holt [31] studied the effect ofκ-casein on micelle dissociation; he ex-pected that added κ-casein would causedissociation of the micelles, and it didnot. The experiment conducted was simi-lar to what Talbot and Waugh termed mi-celle transformation [60]. In all previousstudies of κ-casein content versus size,the more κ-casein present, the smaller themicelles. Addition of purified κ-casein tomicelles causes a shift to smaller sizesnot complete dissociation. It should alsobe noted that the purified κ-casein usedin these experiments represents an SH-capped laddered polymer and not a reactivereduced monomeric species as discussedabove (Tab. II).
4.3. Scattering studies
Both small-angle X-ray (SAXS) scatter-ing [34, 43, 53] and small angle neutron
(SANS) scattering [28, 58] have been ap-plied to the casein micelles. These tech-niques both provide information on theelectron density of the sample relative tothe solvent. Because of technical limita-tions, regarding the wavelength of the ra-diation relative to the total particle size,SAXS methodologies in essence provide aviewing window on the micelles. The datathen must be interpreted in terms of thedensity of the average particles observedwithin the micelles. To circumvent thisproblem it has been common to study firstthe sodium caseinate, which is totally con-tained within the experimental window andso determine its scattering density. Com-parison of the density difference is thenmade with that observed for the windowon the micelle. When this is done carefully,good inferences into the nature of the par-ticles within the overall micelle structurecan be made. Using these concepts and en-hanced experimental techniques, the twoSANS studies came to very similar con-clusions that the micelles have within themparticles with electron densities (scatter-ing centers) similar to those found for thesodium caseinate (caseins only). For theSAXS data the electron density differencefor the sodium caseinate is extremely lowrelative to globular proteins (9.9 e·nm−3 for

378 P.X. Qi
Figure 6. Transmission Electron Micrograph of sodium caseinate in imidazole buffer (pH 7.0), fixedwith glutaraldehyde and stained with uranyl acetate and lead citrate.
casein versus 67 e·nm−3 for α-lactalbumin)and the particles within the micelles havethis same low electron density; similarcalculations can be done for the SANSdata. Where these calculations differ is inthe mathematical models used to fit thedata for the proteins; basically the scatter-ing centers within the micelles display agood deal of heterogeneity leading Hansenet al. [28] to conclude a polydisperse dis-tribution of submicelles, while Stothartand Cebula [58] postulated submicelles ofmore closely packed nature. Kumosinskiet al. [43] fitted their data for sodiumcaseinate to a somewhat lopsided spherewithin a sphere, basically a spherical hy-drophobic core and a loose hydrophilicshell reminiscent of Schmidt’s submicellemodel given in Figure 1b. However, thedata could only be fitted well for the re-formed micelles when there was signifi-cant overlap among the casein moleculescontained in adjacent submicelles. Thesestudies arrived at the same conclusion:proteinaceous submicellar structures existwithin the casein micelles. Where they
differ are on the possible arrangement ofthe casein monomers within the clustersand the possibility of overlap.
Holt et al. [33] studied calcium phos-phate nanoclusters with both SANS andSAXS. These clusters also gave the bestfit to the shell core model used byKumosinski et al. [43], and this led Holtto speculate that the calcium phosphate“clusters rather than putative submicellescould be solely responsible for the het-erogeneous structure revealed by electronmicroscopy, neutron scattering and X-rayscattering”. On the surface this interpreta-tion appears plausible, but it is not. First thecalcium phosphate nanoclusters studied byHolt et al. [33] yielded an electron densityof 700 to 900 e·nm−3, nearly 100 times thatof the caseins as noted above. Secondlythe SAXS data by Kumosinski et al. [43]were for reformed casein micelles in a non-phosphate buffer thus nanoclusters wereabsent. The agreement between the SANSdata with calcium phosphate and the SAXSdata without is therefore all the more strik-ing indicating the presence of submicellar

Casein micelle structure 379
structures of a proteinaceous nature andruling out calcium phosphate nanoclustersas the central feature of the casein micelle.
Considering the nature of the submi-celles which may not be globular withinthe micelle, the overlap of the structuresmakes good sense particularly if the cal-cium phosphate double layers surroundthem at the time of coalescence in theGolgi vesicles. This would allow for the re-arrangements seen in the alveolar lumen aswell as the clustering of the κ-caseins intodisulfide linked polymeric chains.
4.4. AFM studies
Atomic Force Microscopy tech-niques [25, 44, 64] have recently beenapplied to study the effects of pH andpressure on the casein micelles. To under-stand the important role of protein-proteininteractions in the formation and stabi-lization of the micelles, we investigatednative casein micelles using AFM intapping mode in comparison to sodiumcaseinate which may serve as a modelfor submicelles (proteinaceous particles).The results were shown in Figure 7.Under dilute native micelle concentration,the particle size distribution centered on∼10 nm in diameter (Fig. 7a). It should benoted that these particles were persistentlyfound in other caseinate systems suchas calcium caseinate. We speculate thiscategory of particles might be the “basic”necessary but not sufficient proteinaceousunit formed through protein-proteininteractions; they may or may not un-dergo binding calcium and assemblinginto micelles successfully. The particlesize distribution in sodium caseinate, onthe other hand, exhibited nearly doublesized particles with severe overlappingand clustering. Clearly, protein-proteininteractions are the dominating feature inthis system. Our TEM results seemed tosuggest a similarity among the “random
strands” (protein aggregates) in skimmilk (Fig. 1a), the proteinaceous particlessecreted in the Golgi (prior to calciumbinding, Fig. 4), and the sodium caseinatesystem (Fig. 6). This is consistent with thefindings by Gebhardt et al. [25] in theirwork on pressure-induced dissociation ofcasein micelles. It is, therefore, reasonableto assume that these 20 nm sized particlesmight be attributable to the much debatedterm “submicelles”. We recognize thatfurther research is needed to fully under-stand the driving force in the formationand stabilization of casein micelles.
5. CONCLUSIONS
The common attribute that defines thevertebrate class mammalia is the produc-tion of milk, as the primary nutrient forthe neonate, by mammary tissue. Whilethe morphology and physiology of themammary gland varies considerably fromspecies to species, at the ultrastructurallevel the mammary epithelial cells havea common cellular motif. This cellularmotif is quite adaptable and by the regu-lation of its elements, each species can re-spond to a variety of nutritional circum-stances and efficiently produce a milk witha composition suited to the requirementsof its neonate. This is particularly truefor the calcium-phosphate-transport com-plexes, the casein micelles. Despite a widevariety of genetic influences that can al-ter the ratios of the individual caseins, ca-sein micelles are formed in a biologicallycompetent fashion to allow the secretion ofthe completed micelles. The combinationof past research on the details of this bi-ological process and recent developmentsfrom the studies of protein-protein interac-tions in the field of protein science leads tothe following conclusions:
• Selective and productive protein-protein interactions (electrostatic and

380 P.X. Qi
(a)
(b)
Figure 7. Atomic Force Microscopy images of native micelles (7a) and sodium caseinate (7b).Original protein concentration applied onto the mica was 0.3 mg·mL−1. Mica was washed threetimes to remove unbound proteins with water before imaging. The scan scale is 500 nm × 500 nmand 1 μm × 1 μm, and the height bar is 5 nm and 10 nm for 7a and 7b, respectively.

Casein micelle structure 381
hydrophobic, etc.) are the driving forcein the formation of casein micelles.• On transport to the Golgi appara-
tus, the pre-formed proteinaceous par-ticles (submicelles) are phosphorylated(rather slowly) and calcium and phos-phate intercalated into these particles.• Casein association/aggregation occurs
in the Golgi vesicles through thecoupling of the “submicelles” like(∼20 nm) complexed with calcium andphosphate-casein micelles.• It is likely that casein micelle forma-
tion is a hierarchal process-originatedfrom a basic protein-protein interactionunit (∼9.0–11 nm), which may or maynot lead to the successful formation ofmicelles.
Acknowledgements: The author would like tothank Dr. Harold M. Farrell, Jr., for fruitful dis-cussions and acknowledge Dr. Peter H. Cookeand Ms. Winnie Yee for their technical assis-tance in obtaining the high resolution TEM andAtomic Force Microscopy Images presented inthis work.
REFERENCES
[1] Alaimo M.H., Wickham E.D., Farrell H.M.,Jr., Effect of self-association of αs1-caseinand its cleavage fractions αs1-casein(136–196) and αs1-casein(1–197), on aromatic cir-cular dichroic spectra: comparison with pre-dicted models, Biochim. Biophys. Acta 1431(1999) 395–409.
[2] Anfinsen C.B., Principles that govern thefolding of protein chains, Science 181 (1973)223–230.
[3] Bingham E.W., Farrell H.M., Jr., Casein ki-nase from the Golgi apparatus of lactatingmammary gland, J. Biol. Chem. 249 (1974)3647–3651.
[4] Bingham E.W., Farrell H.M., Jr., Carroll,R.J., Properties of dephosphorylated αs1-casein. Precipitation by calcium ions andmicelle formation, Biochemistry 11 (1972)2450–2454.
[5] Bingham E.W., McGranaghan M.B.,Wickham E.D., Leung C.T., Farrell H.M.,Jr., Properties of [Ca2+ + Mg2+]-adenosinetriphosphatases in the Golgi apparatus and
microsomes of the lactating mammaryglands of cows, J. Dairy Sci. 76 (1993)393–400.
[6] Bu H., Hu Y., Sood S.M., Slattery C.W.,Comparison of native and recombinant non-phosphorylated human β-casein: further evi-dence for a unique β-casein folding pattern,Arch. Biochem. Biophys. 415 (2003) 213–220.
[7] Carroll R.J., Farrell H.M., Jr.,Immunological approach to location ofκ-casein in the casein micelle by elec-tron microscopy, J. Dairy Sci. 66 (1983)679–686.
[8] Carroll R.J., Thompson M.P., Nutting G.C.,Glutaraldehyde fixation of casein micellesfor microscopy, J. Dairy Sci. 51 (1968)1903–1908.
[9] Chanat E., Martin P., Ollivier-Bousquet M.,α(S1)-Casein is required for the efficienttransport of β- and κ-casein from the endo-plasmic reticulum to the Golgi apparatus ofmammary epithelial cells, J. Cell Sci. 112 (Pt19) (1999) 3399–3412.
[10] Dalgleish D.G., A mechanism for thechymosin-induced flocculation of casein mi-celles, Biophys. Chem. 11 (1980) 147–155.
[11] Dalgleish D.G., Spagnuolo P.A., Goff H.D.,A possible structure of the casein mi-celle based on high-resolution field-emissionscanning electron microscopy, Int. Dairy J.14 (2004) 1025–1031.
[12] de Kruif C.G., Holt C., Casein micellestructure, functions and interactions, in:Fox P.F. and McSweeney P.L.H. (Eds.),Advanced Dairy Chemistry: Proteins,Kluwer Academic/Plenum Publishers,New York, NY, 2003, 1, pp. 233–276.
[13] de Kruif C.G., Zhulina E.B., κ-Casein as apolyelectrolyte brush on the surface of caseinmicelles, Colloid Surface A 117 (1996) 151–159.
[14] de Kruif C.G., Tuinier R., Holt C., TimminsP.A., Rollema H.S., Physicochemical studyof κ- and β-casein dispersions and the ef-fect of cross-linking by transglutaminase,Langmuir 18 (2002) 4885–4891.
[15] Farrell H.M., Jr., Physical equilibria:Proteins, in: Wong N.B. (Ed.), Fundamentalsof Dairy Chemistry, 3rd edn., Van Nostrand,Reinhold, NY, 1988, pp. 461–510.
[16] Farrell H.M., Jr., Thompson M.P., Caseins ascalcium binding proteins, in: Thompson M.P.(Ed.), Calcium binding proteins, CRC Press,Boca Raton, FL, 1988, II, pp. 117–137.
[17] Farrell H.M., Jr., Leung C.T., Wickham E.D.,Distribution of ADPase activity in the lactat-ing rat mammary gland and its possible role

382 P.X. Qi
in an ATP cycle in the Golgi apparatus, Arch.Biochem. Biophys. 292 (1992) 368–375.
[18] Farrell H.M., Jr., Cooke P.H., King G.,Hoagland P.D., Groves M.L., KumosinskiT.F., Chu B., Particle sizes of casein sub-micelles and purified κ-casein: Comparisonsof dynamic light scattering and electron mi-croscopy with predictive three-dimensionalmolecular models, in: Parris N., Kato A.,Creamer L.K., Pearce J. (Eds.), ACSSymposium Series 650: MacromolecularInteractions in Food Technology, Chapter 5,Washington, DC, 1996, pp. 61–95.
[19] Farrell H.M., Jr., Kumosinski T.F., MalinE.L., Brown E.M., The caseins of milkas calcium binding proteins, Methods Mol.Biol. 172 (2002) 97–140.
[20] Farrell H.M., Jr., Qi P.X., Brown E.M.,Cooke P.H., Tunick M.H., Wickham E.D.,Unruh J.J., Molten globule structures inmilk proteins: Implications for potential newstructure-function relationships, J. Dairy Sci.85 (2002) 459–471.
[21] Farrell H.M., Jr., Brown E.M., HoaglandP.D., Malin E.L., Higher order structuresof the caseins: A paradox?, in: Fox P.F.,McSweeney P.L.H. (Eds.), Advanced DairyChemistry - 1: Proteins, 3rd edn., KluwerAcademic/Plenum Publishers, New York,NY, 2003, Part A, pp. 203–231.
[22] Farrell H.M., Jr., Cooke P.H., Wickham E.D.,Piotrowski E.G., Hoagland P.D.,Environmental influences on bovineκ-casein: Reduction and conversion to fibril-lar (amyloid) structures, J. Protein Chem. 22(2003) 259–273.
[23] Farrell H.M., Jr., Jimenez-Flores R., BleckG.T., Brown E.M., Butler J.E., CreamerL.K., Hicks C.L., Hollar C.M., Ng-Kwai-Hang K.F., Swaisgood H.E., Nomenclatureof the proteins of cows’ milk-sixth revision,J. Dairy Sci. 87 (2004) 1641–1674.
[24] Farrell H.M., Jr., Malin E.L., Brown E.M.,Qi P.X., Casein micelle structure: What canbe learned from milk synthesis and structuralbiology? Curr. Opin. Colloid Interface Sci.11 (2006) 135–147.
[25] Gebhardt R., Doster W., Friedrich J., KulozikU., Size distribution of pressure-decomposedcasein micelles studied by dynamic lightscattering and AFM, Eur. Biophys. J. 35(2006) 503–509.
[26] Griffin M.C., Roberts G.C., A 1H-n.m.r.study of casein micelles, Biochem. J. 228(1985) 273–276.
[27] Griffin M.C., Lyster R.L., Price J.C., The dis-aggregation of calcium-depleted casein mi-celles, Eur. J. Biochem. 174 (1988) 339–343.
[28] Hansen S., Bauer R., Lomholt S.B., BruunQvist K., Pedersen J.S., Mortensen K.,Structure of casein micelles studiedby small-angle neutron scattering, Eur.Biophys. J. 24 (1996) 143–147.
[29] Hill R.J., Wake R.G., Amphiphile nature ofκ-casein as the basis for its micelle stabiliz-ing property, Nature 221 (1969) 635–639.
[30] Holt C., Structure and stability of bovine ca-sein micelles, Adv. Protein Chem. 43 (1992)63–151.
[31] Holt C., Casein micelle substructure andcalcium phosphate interactions studied bySephacryl colume chromatography, J. DairySci. 81 (1998) 2994–3003.
[32] Holt C., Sawyer L., Caseins as rheomor-phic proteins: interpretation of primary andsecondary structures of the αs1-, β- andκ-caseins, J. Chem. Soc. Faraday Trans. 89(1993) 2683–2692.
[33] Holt C., Timmins P.A., Errington N., LeaverJ., A core-shell model of calcium phos-phate nanoclusters stabilized by beta-caseinphosphopeptides, derived from sedimenta-tion equilibrium and small-angle X-ray andneutron-scattering measurements, Eur. J.Biochem. 252 (1998) 73–78.
[34] Holt C., de Kruif C.G., Tuinier R., TimminsP.A., Substructure of bovine casein micellesby small-angle X-ray and neutron scattering,Colloid Surface A 213 (2003) 275–284.
[35] Horne D.S., Casein interactions: casting lighton the black boxes, the structure of dairyproducts, Int. Dairy J. 8 (1998) 171–177.
[36] Horne D.S., Casein structure, self-assemblyand gelation, Curr. Opin. Colloid InterfaceSci. 7 (2002) 456–461.
[37] Jaenicke R., Lilie H., Folding and associa-tion of oligomeric and multimeric proteins,Adv. Protein Chem. 53 (2000) 329–401.
[38] Jahn T.R., Parker M.J., Homans S.W.,Radford S.E., Amyloid formation underphysiological conditions proceeds via anative-like folding intermediate, Nat. Struct.Mol. Biol. 13 (2006) 195–201.
[39] Jang H.D., Swaisgood H.E., Characteristicsof the interaction of calcium with casein sub-micelles as determined by analytical affinitychromatography, Arch. Biochem. Biophys.283 (1990) 318–325.
[40] Kakalis L.T., Kumosinski T.F., Farrell H.M.,Jr., A multinuclear, high-resolution NMRstudy of bovine casein micelles and submi-celles, Biophys. Chem. 38 (1990) 87–98.
[41] Knoop A.M., Knoop E., Wiechen A., Sub-structure of synthetic casein micelles, J.Dairy Res. 46 (1979) 347–350.

Casein micelle structure 383
[42] Kumosinski T.F., Pessen H., Prestrelski S.J.,Farrell H.M., Jr., Water interactions withvarying molecular states of bovine casein:2H NMR relaxation studies, Arch. Biochem.Biophys. 257 (1987) 259–268.
[43] Kumosinski T.F., Pessen H., Farrell H.M.,Jr., Brumberger H., Determination of thequaternary structural states of bovine ca-sein by small-angle X-ray scattering: submi-cellar and micellar forms, Arch. Biochem.Biophys. 266 (1988) 548–561.
[44] Lencki R.W., Evidence for fibril-like struc-ture in bovine casein micelles, J. Dairy Sci.90 (2007) 75–89.
[45] Lin S.H., Leong S.L., Dewan R.K.,Bloomfield V.A., Morr C.V., Effect ofcalcium ion on the structure of native bovinecasein micelles, Biochemistry 11 (1972)1818–1821.
[46] Malin E.L., Brown E.M., Wickham E.D.,Farrell H.M., Jr., Contributions of terminalpeptides to the associative behavior of αs1-casein, J. Dairy Sci. 88 (2005) 2318–2328.
[47] McMahon D.J., Brown R.J., Composition,structure, and integrity of casein micelles: areview, J. Dairy Sci. 67 (1984) 499–512.
[48] McMahon D.J., McManus W.R., Rethinkingcasein micelle structure using electron mi-croscopy, J. Dairy Sci. 81 (1998) 2985–2993.
[49] Meusser B., Hirsch C., Jarosch E., SommerT., ERAD: the long road to destruction, Nat.Cell Biol. 7 (2005) 766–772.
[50] Noble R.W., Jr., Waugh D.F., Casein mi-celles. Formation and structure, I, J. Am.Chem. Soc. 87 (1965) 2236–2245.
[51] Patton S., Milk: Its Remarkable Contributionto Human Health and Well-Being,Transaction Publishers, New Brunswick, NJ,2004.
[52] Payens T.A., Casein micelles: the colloid-chemical approach, J. Dairy Res. 46 (1979)291–306.
[53] Pignon F., Belina G., Narayanan T.,Paubel X., Magnin A., Gesan-Guiziou G.,Structure and rheological behavior of caseinmicelle suspensions during ultrafiltra-tion process, J. Chem. Phys. 121 (2004)8138–8146.
[54] Schmidt D.G., Association of caseins andcasein micelle structure, in: Fox P.F. (Ed.),Developments in Dairy Chemistry: 1,Applied Science Publishers, London, UK,1982, Proteins, pp. 61–86.
[55] Shekar P.C., Goel S., Rani S.D., Sarathi D.P.,Alex J.L., Singh S., Kumar S., κ-Casein-deficient mice fail to lactate, Proc. Natl.Acad. Sci. USA 103 (2006) 8000–8005.
[56] Slattery C.W., A phosphate-induced sub-micelle-micelle equilibrium in reconstitutedcasein micelle systems, J. Dairy Res. 46(1979) 253–258.
[57] Stothart P.H., Subunit structure of casein mi-celles from small-angle neutron-scattering, J.Mol. Biol. 208 (1989) 635–638.
[58] Stothart P.H., Cebula D.J., Small-angle neu-tron scattering study of bovine casein mi-celles and sub-micelles, J. Mol. Biol. 160(1982) 391–395.
[59] Swaisgood H.E., Chemistry of the caseins,in: Fox P.F., McSweeney P.L.H. (Eds.),Advanced Dairy Chemistry: Proteins,Kluwer Academic/Plenum Publishers,New York, NY, 2003, 1, pp. 139–201.
[60] Talbot B., Waugh D.F., Micelle-formingcharacteristics of monomeric and covalentpolymeric kappa caseins, Biochemistry 9(1970) 2807–2813.
[61] Thorn D.C., Meehan S., Sunde M., RekasA., Gras S.L., Macphee C.E., Dobson C.M.,Wilson M.R., Carver J.A., Amyloid fib-ril formation by bovine milk κ-casein andits inhibition by the molecular chaperonesα(s)- and β-casein, Biochemistry 44 (2005)17027–17036.
[62] Tuinier R., de Kruif C.G., Stability of caseinmicelles in milk, J. Chem. Phys. 117 (2002)1290–1295.
[63] Tunick M.H., Cooke P.H., Malin E.L., SmithP.W., Holsinger V.H., Reorganization of ca-sein submicelles in mozzarella cheese duringstorage, Int. Dairy J. 7 (1997) 149–155.
[64] Uricanu V.I., Duits M.H., Mellema J.,Hierarchical networks of casein proteins: anelasticity study based on atomic force mi-croscopy, Langmuir 20 (2004) 5079–5090.
[65] Veis A., Enhanced: A Window onBiomineralization, Science 307 (2005)1419–1420.
[66] Visser J., Schaier R.W., van Gorkom M., Therole of calcium, phosphate and citrate ions inthe stabilization of casein micelles, J. DairyRes. 46 (1979) 333–335.
[67] Walstra P., Casein sub-micelles: do they ex-ist? Int. Dairy J. 9 (1999) 189–192.
[68] Waugh D.F., Creamer L.K., Slattery C.W.,Dresdner G.W., Core polymers of casein mi-celles, Biochemistry 9 (1970) 786–795.
[69] West D.W., Clegg R.A., Casein kinase ac-tivity in rat mammary gland Golgi vesicles.Phosphorylation of endogenous caseins, Eur.J. Biochem. 137 (1983) 215–220.

![Casein micelles in milk revisited · Casein micelles per 17liter - N cm 1.14 10 [L 1] Calculated Nanocluster r nc 1.72 to 2.27 [nm] SANS/SAXS Nanoclusters/ Casein micelle N nc 285](https://img.dokumen.tips/doc/110x75/5fd572d92d5adf1c9e637683/casein-micelles-in-milk-revisited-casein-micelles-per-17liter-n-cm-114-10-l.jpg)



























