Embed Size (px)
Citation preview

STUDIE VAN FACTOREN DIE
POLLENKWALITEIT BIJ COURGETTE
BEÏNVLOEDEN
Aantal woorden: 15 641
Amelien Lybaert
Stamnummer: 01203130
Promotor: Prof. dr. Marie-Christine Van Labeke
Tutor: Ir. Simon Craeye
Masterproef voorgelegd voor het behalen van de graad master in de richting Master of
Science in de biowetenschappen: land- en tuinbouwkunde
Academiejaar: 2016 - 2017


STUDIE VAN FACTOREN DIE
POLLENKWALITEIT BIJ COURGETTE
BEÏNVLOEDEN
Aantal woorden: 15 641
Amelien Lybaert
Stamnummer: 01203130
Promotor: Prof. dr. Marie-Christine Van Labeke
Tutor: Ir. Simon Craeye
Masterproef voorgelegd voor het behalen van de graad master in de richting Master of
Science in de biowetenschappen: land- en tuinbouwkunde
Academiejaar: 2016 - 2017

Auteursrechtelijke bescherming
“De auteur en de promotor geven de toelating deze scriptie voor consultatie beschikbaar te
stellen en delen van de scriptie te kopiëren voor persoonlijk gebruik. Elk ander gebruik valt
onder de beperkingen van het auteursrecht, in het bijzonder met betrekking tot de
verplichting de bron uitdrukkelijk te vermelden bij het aanhalen van resultaten uit deze
scriptie.”
“The author and the promoter give the permission to use this thesis for consultation and to
copy parts of it for personal use. Every other use is subject to the copyright laws, more
specifically the source must be extensively specified when using the results from this thesis”.

Dankwoord
In dit dankwoord zou ik graag een aantal mensen willen bedanken die mij geholpen hebben
met het tot stand brengen van deze thesis.
Als allereerste bedank ik ten zeerste mijn promotor, Prof. dr. Marie-Christine Van Labeke, die
mij zeer goed begeleid heeft gedurende het gehele jaar. Haar continue begeleiding en
bijsturing hebben ervoor gezorgd dat ik gemotiveerd bleef en dat ik deze thesis tot een goed
einde kon brengen.
Naast mijn promotor, bedank ik ook graag mijn tutor, Ir. Simon Craeye. Hij stond steeds
paraat voor mijn vragen en zijn advies en begeleiding tijdens de proeven, heeft ervoor
gezorgd dat alle experimenten goed verliepen. Zonder zijn hulp was dit niet gelukt.
Tot slot wil ik graag mijn ouders, broers, zussen en mijn vriend bedanken voor hun steun
tijdens dit proces.

Abstract
In de courgetteteelt zijn veel problemen met de bevruchting van de vrouwelijke bloemen op
het einde van de teelt. Dit komt omdat in deze fase bijna geen mannelijke bloemen worden
gevormd. Het bewaren van pollen met behoud van vitaliteit zou dit probleem kunnen
oplossen.
In deze masterproef wordt een overzicht gegeven van de verschillende mogelijke
bewaarmethodes van het pollen van Cucurbita pepo L. (courgette) en het effect van deze
bewaring op de pollenvitaliteit en de bevruchting.
Als eerste experiment werd gezocht naar een medium dat gebruikt kon worden om het
kiemvermogen van het pollen te onderzoeken. Hiervoor werden verschillende basismedia
voor pollenkieming onderzocht. Er werden ook verschillende componenten die pollenkieming
stimuleren toegevoegd aan het medium en onderzocht. Er werd geen goed medium
gevonden, waardoor pollenkwaliteit enkel via een vitaliteitskleuring kon worden bepaald.
Vervolgens werd de vitaliteit van het pollen van de rassen Bengal en Tosca getest na een
korte bewaring bij -80°C, waarna ook de pollenvitaliteit bij bewaring bij -20°C, bij 6°C en op
kamertemperatuur werd getest. De reservestoffen, die een rol bij de houdbaarheid van pollen
spelen, werden bij 6 rassen in kaart gebracht. De lage suikerconcentraties zijn indicatief voor
een slechte houdbaarheid.
Om na te gaan of bewaard pollen ook tot een goede bevruchting en goede vruchten leidt,
werden in een latere proef vrouwelijke bloemen manueel bestoven met het bewaarde pollen.
De courgetteplanten die gebruikt werden voor de experimenten, werden geteeld op Inagro.
Het oorspronkelijke idee om tot een goede bewaarmethode van het pollen te komen om zo
de telers te kunnen helpen, is niet behaald doordat het pollen van courgette zeer moeilijk
bewaard en de vitaliteit te snel afneemt in tijd. Bovendien is gebleken dat een behoorlijke
pollenvitaliteit niet gelijk staat aan een goede bevruchting en kwalitatief goede vruchten.
Kernwoorden: Cucurbita pepo L., pollenkieming, pollenvitaliteit, bewaring

Abstract
There are many problems with the fertilization of the female flowers in the cultivation of
zucchini, especially at the end of the cultivation. This is due to the lack of male flowers at this
production stage. Storage of pollen while maintaining a good vitality could solve this problem.
In this master study an overview of the different conservation methods for the pollen of the
Cucurbita pepo L. (zucchini), and the effect of this conservation on the pollen vitality and
fructification will be given.
In a first experiment, we looked for a medium that could be used in order to investigate the
germinability. Different basal media for pollen germination, as well as different components
enhancing the germination power were investigated. We did not find a good germination
medium, therefore the vitality was only tested by a pollen staining method.
Next, we tested the vitality for Bengal and Tosca, two zucchini varieties after a short
conservation at -80°C. In parallel, we tested also the pollen vitality at -20°C, at +6°C and at
room temperature. For six varieties, we surveyed the carbohydrate reserve who were
contributing to the longevity of the pollen. Low sugar concentrations herein revealed a bad
longevity of the pollen.
In order to assess if stored pollen could lead to a good fertilization and a good fruit quality
afterwards, , we pollinated the female flowers manually with the different stored pollen. The
plants used for the experiments were grown at Inagro.
The original idea that we could be able to help the growers by offering them a good
conservation method of the pollen was not supported by our test results because of the
difficulty of conservation pollen and a very fast decrease of pollen vitality by time. In addition,
it has turned out that a good pollen vitality does not automatically lead to a good fertilization
and a good fruit quality.
Keywords: Cucurbita pepo L., spore germination, spore vitality, conservation

1
Inhoud
Inleiding ................................................................................................................................. 3
1 Literatuurstudie .............................................................................................................. 4
1.1 Cucurbita pepo ........................................................................................................ 4
1.2 Het pollen ................................................................................................................ 5
1.2.1 Morfologie ........................................................................................................ 5
1.2.2 Pollencomponenten.......................................................................................... 6
1.2.3 Inhoud van een pollenkorrel ............................................................................. 7
1.3 Pollenvitaliteit .......................................................................................................... 9
1.3.1 Methodes ter bepaling van de pollenkwaliteit ................................................... 9
1.3.2 Factoren die de vitaliteit beïnvloeden ..............................................................10
1.4 In vitro pollenkieming ..............................................................................................11
1.4.1 In vivo kieming versus in vitro kieming ............................................................11
1.4.2 Voorbehandelingen .........................................................................................12
1.4.3 Kiemingsmedium .............................................................................................12
1.5 Vorming pollenkiembuis en bevruchting .................................................................17
1.5.1 Morfologie pollenkiembuis ...............................................................................17
1.5.2 Bevruchting .....................................................................................................17
1.6 Bewaring van pollenkorrels ....................................................................................23
1.6.1 Temperatuur en relatieve vochtigheid .............................................................23
1.6.2 Watergehalte van pollenkorrels .......................................................................23
1.6.3 Suikergehalte van pollenkorrels ......................................................................25
2 Materiaal en methode ....................................................................................................26
2.1 Kiemingstesten .......................................................................................................26
2.2 Zetmeel- en suikergehalte ......................................................................................29
2.3 Pollenvitaliteit na bewaring bij -80°C ......................................................................30
2.4 Pollenvitaliteit na bewaring bij verschillende temperaturen .....................................30
2.5 Manuele bestuiving ................................................................................................31
2.6 Statistische verwerking ...........................................................................................32

2
3 Resultaten .....................................................................................................................33
3.1 Pollenkieming .........................................................................................................33
3.1.1 Algemene observaties .....................................................................................33
3.1.2 Basismedium...................................................................................................33
3.1.3 Effect van verschillende toevoegingen ............................................................34
3.2 Zetmeelgehalte ......................................................................................................35
3.3 Suikergehalte .........................................................................................................35
3.3.1 Sucrosegehalte ...............................................................................................35
3.3.2 Glucosegehalte ...............................................................................................35
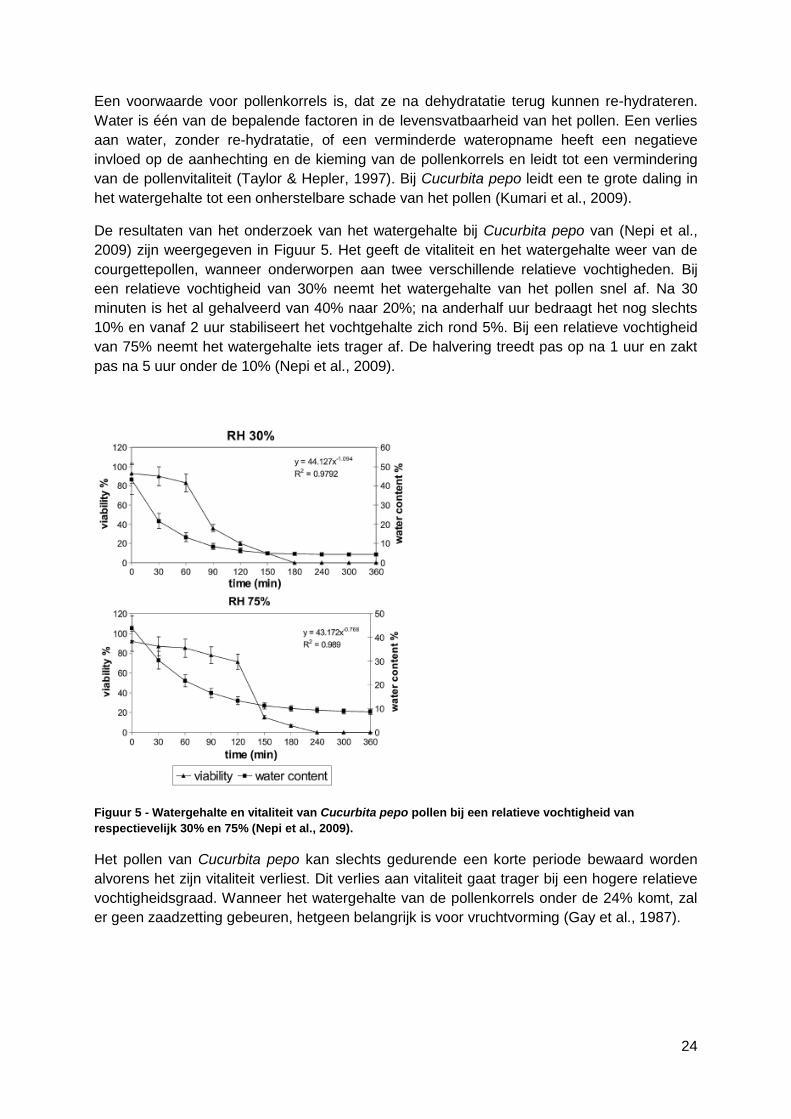
3.3.3 Fructosegehalte ..............................................................................................36
3.4 Pollenvitaliteit na bewaring bij -80°C ......................................................................37
3.5 Pollenvitaliteit na bewaring bij verschillende temperaturen .....................................37
3.5.1 Vers pollen ......................................................................................................37
3.5.2 Bewaring op kamertemperatuur ......................................................................37
3.5.3 Bewaring in de frigo (6°C) ...............................................................................38
3.5.4 Bewaring in diepvries (-20°C) ..........................................................................38
3.6 Manuele bestuiving ................................................................................................39
4 Discussie ...................................................................................................................41
4.1 Pollenkieming .........................................................................................................41
4.2 Zetmeel- en suikergehalte ......................................................................................41
4.3 Pollenvitaliteit na bewaring bij -80°C ......................................................................42
4.4 Pollenvitaliteit na bewaring bij verschillende temperaturen .....................................43
4.4.1 Vers pollen ......................................................................................................43
4.4.2 Bewaring op kamertemperatuur ......................................................................43
4.4.3 Bewaring in de frigo (6°C) ...............................................................................43
4.4.4 Bewaring in diepvries (-20°C) ..........................................................................44
4.5 Manuele bestuiving ................................................................................................44
5 Besluit ...........................................................................................................................45
6 Bibliografie ....................................................................................................................46
7 Bijlagen .........................................................................................................................50
7.1 Zetmeelanalyse ......................................................................................................50
7.2 Pollenvitaliteit .........................................................................................................52

3
Inleiding
Uit gegevens van de Vlaamse veilingen (REO en BelOrta) blijkt dat het aantal geteelde
courgettes jaarlijks toeneemt. In 2007 werden ongeveer 25 400 000 courgettes geteeld voor
de versmarkt. In 2016 waren dat er al 49 100 000. Sinds 2005 is de omzet op de REO-veiling
verzesvoudigd, van 1,64 mio in 2005 naar 6,85 mio in 2016. Op de BelOrta-veiling was de
stijging minder spectaculair (2,9 mio in 2007 en 3,3 mio in 2016).
Deze stijging van het aantal geteelde courgettes weerspiegelt zich in de areaalgegevens
verzameld door het Vlaams departement Landbouw en Visserij. De totale teeltoppervlakte
van courgette schommelt sinds 2011 tussen 629 ha en 740 ha. Tot 2015 nam de
teeltoppervlakte voor de industrie af, maar in 2016 was er opnieuw een lichte stijging tot 335
ha. Bij de courgettes, geteeld voor de vers markt, neemt sinds 2011 de oppervlakte jaarlijks
toe, met als hoogtepunt 359 ha in 2016.
In de courgetteteelt zijn er heel veel problemen bij de bevruchting van de vrouwelijke
bloemen. Het grote probleem ligt bij een tekort aan mannelijke bloemen op het einde van de
teelt. Hierdoor is er onvoldoende stuifmeel voorhanden om alle vrouwelijke bloemen van
voldoende pollen te voorzien. Wanneer de vrouwelijke bloemen onvoldoende bevrucht
worden, ontstaan er vruchten die niet mooi gelijkvormig zijn, de zogenaamde spitsvruchten.
Deze spitsvruchten zijn niet gewild op veilingen doordat ze zeer gevoelig zijn voor vruchtrot
en hierdoor zeer slecht bewaren.
Uit een bevraging van 92 telers, uit West-Vlaanderen en Antwerpen, blijkt dat bij 78%
jaarlijks puntvruchten voorkomen. Het tijdstip waarop dit gebeurt is afhankelijk van de teelt: in
de serreteelt in vollegrond is dit in het voorjaar, in de hydroteelt en de openluchtteelt komen
de puntvruchten gedurende het hele jaar voor. Niet alleen het onvoldoende aanwezig zijn
van pollen of een te lage activiteit van de bestuivers is volgens de telers een probleem, ook
het klimaat speelt een belangrijke rol bij het voorkomen van puntvruchten.
Indien er een geschikte methode voor de bewaring van het pollen zou worden gevonden,
kan het pollen verzameld worden op momenten waarop het overvloedig aanwezig is. Het
bewaarde pollen zou dan aangebracht worden op de vrouwelijke bloemen wanneer er
onvoldoende pollen geproduceerd wordt. Tijdens de problematische weken worden door een
aantal serretelers de vrouwelijke bloemen manueel bestoven, maar ze beschikken niet altijd
over voldoende pollen hiervoor.
In deze thesis wordt gezocht naar een manier om pollen te bewaren zonder al te veel
kwaliteitsverlies. Deze methode zou door telers toegepast kunnen worden om periodes met
weinig stuifmeelproductie te overbruggen en het voorkomen van puntvruchten te reduceren.
Om de kwaliteit van het pollen te bepalen kan een vitaliteitskleuring of een in vitro kieming
worden toegepast. De in vitro kieming geeft een correcter beeld van de pollenkwaliteit.
Daarom wordt in deze thesis ook gezocht naar een goed kiemingsmedium voor het pollen
van courgette.

4
1 Literatuurstudie
1.1 Cucurbita pepo
De courgette - of Cucurbita pepo L. - behoort tot de Cucurbitaceae, beter gekend als de
komkommerfamilie. Tot het geslacht Cucurbitaceae behoren vijf dominante soorten en
ongeveer 22 wilde soorten (Decker, 1988). Er bestaan twee grote subfamilies binnen de
Cucurbitaceae: de Cucurbitoideae en de Zanonioideae, door sommige auteurs ook wel
Nhandiroboideae genoemd. Cucurbita pepo behoort tot de Cucurbitoideae, meer bepaald tot
de stam Cucurbiteae (Jeffrey, 1980). De leden van de Cucurbitaceae zijn wijd verspreid met
zowel tropische als subtropische soorten die vorstgevoelig zijn (Schaefer & Renner, 2011)
(Jeffrey, 1980). Tot de Cucurbitaceae behoren zowel belangrijke sierplanten, als land- en
tuinbouwgewassen zoals komkommer, pompoen, meloen en courgette (Schaefer & Renner,
2011).
Alle Cucurbitaceae bevatten cucurbitacine (verantwoordelijke voor de bitterhied en giftig in
hoge concentraties) en andere triterpenoïden in de vegetatieve plantendelen en vruchten.
Sommige geslachten bevatten ook citruline en andere ninhydrine componenten. Niet alleen
de vruchten, maar ook de zaden van Cucurbitaceae hebben een nutritionele waarden door
hun hoge olie- en proteïnegehaltes (Jeffrey, 1980).
Cucurbita pepo is één van de meest verspreide soorten van de Cucurbitaceae. De oudste
zaden van Cucurbita pepo zijn teruggevonden in Mexico en dateren van 9000 jaar geleden
(Decker, 1988).
De vruchten van Cucurbita pepo hebben geen bittere smaak en kunnen rond of langwerpig
zijn met een gele, groene of oranje kleur. Bij oudere vruchten kan de schil verhard zijn
(Decker, 1988).
Bij de Cucurbitaceae is 50% van de soorten eenhuizig, de overige 50% is tweehuizig
(Schaefer & Renner, 2011). Cucurbita pepo is een eenhuizige plant, met zowel mannelijke
als vrouwelijke bloemen. De verhouding mannelijke/vrouwelijke bloemen is afhankelijk van
het ras en de klimaatomstandigheden (Ciampolini et al., 1993).

5
1.2 Het pollen
1.2.1 Morfologie
Volwassen pollenkorrels van Angiospermen zijn geen gewone vegetatieve cellen. Het zijn
cellen die zich ontwikkelen in de antheren van de mannelijke bloemen en spermacellen
bevatten. De spermacellen zijn omgeven door een celwand en celwand componenten. Deze
configuratie wordt bereikt kort na de meiose, wanneer een asymptotische mitosedeling een
grote cel produceert, die zijn kleine zuster cel omgeeft (Edlund et al., 2004). Na deze eerste
mitotische deling is het pollen bicellulair (Eady et al., 1994). De grote cel is de gametofyt en
heeft als functie de synthese en de vorming van de pollenkiembuis celwand (Mascarenhas,
1993). De kleinere cel is de generatieve cel, die na de eerste mitotische deling nog een
tweede mitotische deling ondergaat waardoor een tweede spermacel gevormd wordt. Deze
tweede spermacel is nodig voor de dubbele bevruchting. Bij tricellulaire pollen zijn al deze
delingen gedaan alvorens de pollenkorrels de antheren verlaten. Bicellulair pollen ondergaat
een laatste mitotische deling tijdens de groei van de pollenkiembuis (Edlund et al., 2004).
Tijdens de ontwikkeling van de pollenkorrels op de antheren zorgen de tapetumcellen, die de
pollenkorrels omgeven, voor de nodige voedingselementen. Deze voedingselementen
voorzien het pollen van energie, die nodig is voor de metabolische activiteiten (Jóhannsson
& Stephenson, 1998). De degeneratie van de tapetumcellen bij Cucurbita pepo start al in het
vroegere bicellulaire stadium van het pollen, wat vroeger is dan bij andere plantensoorten
(Ciampolini et al., 1993).
De mannelijke bloemen van Cucurbita pepo hebben 5 buisvormige antheren, die samen
ongeveer 16 500 pollenkorrels produceren. Bij het dwars opensnijden van de antheren van
Cucurbita pepo, zijn er twee rijen pollenkorrels zichtbaar. Tijdens de anthese vormen deze
twee rijen één laag, die wordt samengehouden door de pollenkit (Nepi & Pacini, 1993). Elke
pollenkorrel heeft 12 poriën. Een deel van de poriën steekt tijdens de anthese boven het
oppervlak uit. Dit is te zien op Figuur 1 (Nepi et al., 2009).
Figuur 1 - Morfologie van een pollenkorrel bij Cucurbita pepo L. met de poriën die boven het oppervlak steken (Nepi & Pacini, 1993).

6
Pollenkorrels van Cucurbitaceae zijn meestal monaden, hoewel ook tetraden voorkomen
(Perveen & Qaiser, 2008). De subfamilie Zanonioideae is gekernmerkt door uniforme en
tricellulaire pollenkorrels die relatief klein zijn (tot 40 μm) (Jeffrey, 1964) (Lira et al., 1998)
(Van Der Ham & Van Heuven, 2003). De subfamilie Cucurbitoideae heeft eerder grotere
pollenkorrels, tot 200 μm (Jeffrey, 1980) (Van Der Ham et al., 2010). Cucurbita pepo heeft
bicellulair pollen, dat bolvormig is, met een diameter van 180 μm tot 200 μm (Nepi et al.,
2009) (Gay et al., 1987) (Carvajal et al., 2013).
Bij de leden van de Cucurbitaceae produceert elke pollenkorrel 1 tot 3 pollenkiembuizen.
Eén pollenkiembuis hiervan wordt dominant en zal verder uitgroeien terwijl de andere
kiembuizen afsterven (Vasil, 1960).
1.2.2 Pollencomponenten
1.2.2.1 Pollencelwand
Pollenkorrels zijn omgeven door een celwand, waarvan de verschillende cellagen zich
geleidelijk ontwikkelen (Edlund et al., 2004). Deze celwand is heel belangrijk voor de
pollenkorrels omdat het zorgt voor de signalisatie bij de aanhechting van het pollen op de
stempel. Ook bij de kieming speelt de celwand een belangrijke rol (Taylor & Hepler, 1997).
De celwand bestaat uit drie lagen, waarvan de samenstelling verschilt naargelang de
plantensoort (Edlund et al., 2004). De dwarsdoorsnede van de celwand is weergegeven op
Figuur 2.
Figuur 2 – Dwarsdoorsnede van een pollenkorrel (P) en een stempel cel (S) (Edlund et al., 2004).

7
De buitenste cellaag van de celwand, ook wel de exinelaag genoemd, is meerlagig en is
opgebouwd uit een chemisch resistent polymeer, sporopollenine. Ze heeft een oranjeachtige
kleur en is olieachtig, wat zorgt voor de plakkerige structuur van het pollen (Edlund et al.,
2004) (Vasil, 1960). De exinelaag is een opslagplaats voor proteïnen afkomstig van
tapetumcellen en bevat meerdere poriën (Howlett et al., 1973)(Edlund et al., 2004). Deze
buitenste laag zorgt voor de stevigheid en de elasticiteit van de pollenwand en is betrokken
bij de interacties met insecten (Edlund et al., 2004). De synthese van de exinelaag start in
het tetradenstadium tijdens de sporogenese (Shi & Yang, 2010).
De middelste laag van de celwand is een intinelaag, die eveneens meerlagig is en
voornamelijk bestaat uit cellulose (Edlund et al., 2004). De intinelaag wordt na de synthese
van de exinelaag gesynthetiseerd (Shi & Yang, 2010). Verschillende diktes van exine- of
intinelagen en een ander patroon van poriën zorgen voor een grote diversiteit in de
samenstelling van de pollenkorrel (Perveen & Qaiser, 2008).
1.2.2.2 Pollencoat
De term pollencoat verwijst naar de vetachtige substantie die volwassen pollenkorrels
omgeeft (Shi & Yang, 2010). Het komt voor op het pollen van bijna alle Angiospermen, maar
bij Gymnospermen is het afwezig (Pacini & Hesse, 2005). Het grootste bestanddeel van de
pollencoat is de pollenkit, een hydrofoob mengsel dat grotendeels bestaat uit verzadigde en
onverzadigde vetten, carotenoïden, flavonoïden, proteïnen en koolhydraten. De exacte
samenstelling van de pollenkit is verschillend voor verschillende plantensoorten (Pacini &
Hesse, 2005) (Shi & Yang, 2010). Naast de pollenkit bevat de pollencoat ook nog lipiden,
proteïnen, pigmenten en aomatische componenten (Shi & Yang, 2010) (Edlund et al., 2004).
De lipiden in de pollencoat zijn betrokken in de cel-cel herkenning en de aanhechting en zijn
ook noodzakelijk voor de hydratatie van het pollen (Taylor & Hepler, 1997) (Edlund et al.,
2004). De pollencoat vult de openingen in de exinelaag op (Edlund et al., 2004). De
verdeling van de pollencoat over het oppervlak van het pollen is verschillend van
plantensoort tot plantensoort (Pacini & Hesse, 2005). De pollencoat beschermt het pollen
tegen uitdroging, UV-straling en aanvallen van pathogenen (Edlund et al., 2004).
1.2.3 Inhoud van een pollenkorrel
1.2.3.1 Koolhydraten
Koolhydraten zijn een van de belangrijkste bestanddelen van pollenkorrels en bestaan
voornamelijk uit polysacchariden zoals zetmeel en celwandmateriaal (Campos et al., 2008).
Kleine pollenkorrels bevatten over het algemeen weinig zetmeel en meer vetten, grote
pollenkorrels hebben vaak een groter zetmeelgehalte. Doordat zetmeel meer plaats inneemt
dan vetten, kan de grootte van de pollenkorrel gerelateerd worden met het zetmeelgehalte
ervan. Plantensoorten die door insecten bestoven worden hebben pollen met een groter
suikergehalte en een lager zetmeelgehalte dan planten die door de wind bestoven worden
(Baker & Baker, 1979).

8
Ook de temperatuur en de relatieve vochtigheid bepalen het zetmeelgehalte van het pollen.
Pollen dat ontwikkelt bij 25°C bevat minder zetmeel dan pollen dat bij 15°C ontwikkelt. Hoe
hoger de relatieve vochtigheid van de omgeving, hoe lager het zetmeelgehalte in het pollen.
Deze afname is te verklaren door een verhoogde ademhaling die resulteert in een hoger
verbruik van de reservestoffen, waaronder ook zetmeel. Het pollen van de Cucurbitaceae
heeft over het algemeen niet veel zetmeel, hoewel er ook species zijn die een hoger
zetmeelgehalte hebben (Baker & Baker, 1979). Het cytoplasma van de pollenkorrels van
Cucurbita pepo is rijk aan zetmeel (Nepi et al., 2009).
Het analytisch berekende koolhydratengehalte, dat alle aanwezige koolhydraten in een
pollenkorrel weergeeft, toont een hoger percentage dan de hoeveelheid koolhydraten die via
chemische analyses berekend wordt. Dit komt doordat vezels en celwand materiaal ook tot
de koolhydraten behoren, maar niet worden gedetermineerd bij chemische analyses. Het
gehalte aan vezels varieert van 7g/100g tot 20g/100g (Campos et al., 2008).
1.2.3.2 Proteïnen en aminozuren
Het proteïnegehalte van pollen is afhankelijk van de botanische origine van de plant en
varieert tussen 2,5% en 61% van het drooggewicht van het pollen (Campos et al., 2008)
(Roulston et al., 2000). Er zijn zeventien aminozuren aanwezig in het pollen waarvan
proline, glutamine en leucine de belangrijkste zijn (Campos et al., 2008).
De variatie in het proteïnegehalte bepaalt mee de mate waarin de planten bestoven worden
door insecten. Bijen, trips, mijten… verkiezen pollen met een hoger proteïnegehalte omwille
van de hogere nutritionele waarde. De meeste proteïnen in pollen zijn enzymen die
belangrijk zijn voor de groei van de pollenkiembuis en de bevruchting (Roulston et al., 2000).
1.2.3.3 Flavonoïden
Flavonolen, een specifieke klasse van flavonoïden, zijn secundaire metabolieten die zowel
pigmenten als kleurloze componenten bevatten. Ze zijn betrokken in de bevruchting,
zaadverspreiding en de bescherming tegen UV licht en spelen een grote rol in de kieming
van het pollen. Flavonolen komen voor in het pollen van verschillende Angiospermen en
Gymnospermen, maar ook in de sporen van mossen en varens (Ylstra, et al., 1992). Alle
pollenkorrels bezitten hoge concentraties aan flavonolen, die gesynthetiseerd worden in de
tapetumcellen en al aanwezig zijn voordat het pollen rijp is. Planten die niet in staat zijn om
flavonolen te synthetiseren, zijn bijgevolg niet in staat pollen te produceren dat kiemt (Taylor
& Hepler, 1997).
1.2.3.4 Mineralen en sporenelementen
Het hoofdelement in bijenpollen (een verzameling pollen van ongekende oorsprong), is
kalium, goed voor 60% van het totale mineraalgehalte. Andere belangrijke elementen zijn
magnesium, natrium en calcium. Vitaminen zijn ook aanwezig in kleine hoeveelheden
(Campos et al., 2008).

9
1.3 Pollenvitaliteit
Met pollenvitaliteit wordt gedoeld op het aandeel pollenkorrels dat levensvatbaar is. Dit
percentage is een maat voor de kwaliteit van het geproduceerde pollen en kan dienen als
indicatie voor de fitness van de plant. Vaak wordt de pollenkwaliteit gelijkgesteld aan de
pollenvitaliteit (Kelly et al., 2002). Bij de interpretatie van de pollenvitaliteitstesten, moet men
rekening houden met het feit dat de resultaten kunnen verschillen naargelang de gebruikte
test.
1.3.1 Methodes ter bepaling van de pollenkwaliteit
1.3.1.1 Door middel van kleuring
Kleuringsmethodes zijn zeer populair voor het aantonen van de vitaliteit van het pollen. Ze
tonen enzymatische activiteit en de integriteit van de membranen aan (Vizintin & Bohanec,
2004).
Eén van de meest gebruikte kleuringsmethodes in het onderzoek naar pollenvitaliteit, is de
FCR-test (Fluorochrome reactie), ook wel de FDA-kleuring (Fluorescein diacetate) genoemd.
Deze test is een uitstekende tool om de kieming te voorspellen (Heslop-Harrison et al.,
1983). De FCR-test steunt op de aanwezigheid van een cytoplasmatische inhoud en test de
esterase activiteit van het pollen (Vizintin & Bohanec, 2004). Bij de FDA-kleuring wordt er
FDA toegevoegd aan een sucroseoplossing. Nadien moet het pollen incuberen in het
medium alvorens de vitaliteit kan geanalyseerd worden (Nepi & Pacini, 1993). Enkel vitale
cellen worden gekleurd door de FDA-kleuring, dode cellen niet. Dit komt doordat enkel in
levende cellen het FDA, door esteraseactiviteit, wordt gehydrolyseerd tot fluorescine, dat in
het celmembraan wordt vastgehouden. Als de cellen belicht worden met blauw licht (480
nm), kleuren de intacte cellen groen door de aanwezigheid van het fluorescine (Jones &
Senft, 1985). Nadelen van deze test methode zijn dat het enkel kan gebruikt worden bij
volwassen, rijp pollen en dat het niet op het veld kan worden toegepast. Indien onrijp pollen
gebruikt wordt, zal de FCR-test een zeer hoge kiemkracht aangeven, terwijl het in
werkelijkheid zeer slecht zal kiemen (Heslop-Harrison et al., 1983).
Een andere kleuring is deze met tetrazolium zout (MTT) gebaseerd op een dehydrogenase
activiteit (Vizintin & Bohanec, 2004). Daarnaast heb je nog de kleuring met aniline blauw. Bij
deze methode worden de pollenkorrels verzameld en in een oplossing van aniline blauw
gebracht. Pollenkorrels die levensvatbaar zijn, zullen de blauwe kleurstof opnemen en
bijgevolg blauw kleuren terwijl niet-levensvatbare pollenkorrels niet zullen verkleuren (Kelly
et al., 2002). De kleuring met aniline maakt gebruik van de aanwezigheid van callose in de
celwand van het pollen (Vizintin & Bohanec, 2004). De aniline methode geeft een zeer goed
beeld van het aantal pollenkorrels en de levensvatbaarheid van de pollenkorrels, maar is
zeer arbeidsintensief. Hierdoor wordt er vaak voor een andere methode gekozen (Kelly et al.,
2002).

10
Een andere veelgebruikte kleuring is de Alexander kleuring. Deze methode wordt ook de
carmine kleuring genoemd. Bij deze kleuringsmethode wordt de aanwezigheid van een
cellulaire matrix gelinkt met de vitaliteit van het pollen. Omdat de aanwezigheid van een
cellulaire matrix niet gelijk staat aan metabolische activiteit, is het niet mogelijk om het
kiemingspercentage van de pollenkorrels te voorspellen aan de hand van de carmine
kleuring. De Alexander kleuring geeft vaak een te hoog vitaliteitspercentage weer (Duarte-
Silva et al. , 2011).
1.3.1.2 Door middel van een kiemingstest
De vitaliteit van het pollen kan ook getest worden door het pollen in vitro te laten kiemen op
het gepast kiemingsmedium. Het is de beste test om aan te geven wat de vitaliteit van het
pollen is (Duarte-Silva et al., 2011). De optimale samenstelling van dit kiemingsmedium
varieert naargelang de plantensoort en de onderzoeker (Jayaprakash & Sarla, 2001). De
nadelen van de in vitro kieming is dat deze, net als de FCR test, enkel kan toegepast worden
op volwassen pollenkorrels en dat de in vitro kieming onderhevig is aan een aantal factoren
waaronder het genotype, de temperatuur en de relatieve vochtigheid (Vizintin & Bohanec,
2004).
1.3.2 Factoren die de vitaliteit beïnvloeden
De vitaliteit van het pollen is niet altijd even groot. Regen, extreme droogte en hoge
temperaturen dragen bij tot het verlies van vitaliteit bij het pollen, ook wanneer deze condities
heersen tijdens de ontwikkeling van het pollen op de moederplant (Hoekstra, 1995).
Onderzoek bij Trichosanthes dioica, een species die tot de Cucurbitaceae behoort, toont aan
dat de vitaliteit het grootst is op het moment van de anthese, dit zowel bij de in vitro kieming
als bij de FCR test. Het hoogste vitaliteitspercentage werd bekomen bij de FCR test. Uit deze
test blijkt ook dat na 30 uur bewaring nog slechts 20% tot 25% van het pollen vitaal is
(Kumari et al., 2009).
De vitaliteit van het pollen bij Cucurbita pepo daalt naarmate de tijd verstrijkt. Bij het openen
van de bloemen is 92% van het pollen vitaal. Dit is maar 75% meer op het moment dat de
bloemen sluiten (Nepi et al., 2001). Na 24 uur bewaring is nog slechts 1% van het pollen van
Cucurbita pepo vitaal. Deze snelle daling is te wijten aan de dehydratatie van het pollen
(Kumari et al. , 2009).

11
1.4 In vitro pollenkieming
Algemeen wordt gesteld dat pollenkorrels gekiemd zijn indien de lengte van de gevormde
pollenkiembuis, gelijk is aan of groter is dan de diameter van de pollenkorrel (Vizintin &
Bohanec, 2004). Kieming van bicellulair pollen, zoals bij courgette, vertoont een tragere
kieming dan tricellulair pollen (Hoekstra, 1995).
Het pollen, waarmee in vitro kiemingsproeven worden uitgevoerd, kan best geoogst worden
net na het openspringen van de antheren. Dan is het pollen immers optimaal wat betreft
vitaliteit. Dit is meestal ’s ochtends vroeg, hoewel er plantensoorten bestaan waarbij dit op
andere momenten plaatsvindt (Vasil, 1960). Bij het pollen van Cucurbita pepo L., moet men
snel zijn om het pollen te oogsten omdat de bloem slechts zes uur open is (Nepi & Pacini,
2001). Op warme dagen sluit de bloem soms nog sneller. De reden hiervoor is dat de
kroonbladeren instaan voor de regeling van het microklimaat rondom de stamper. Lange
blootstelling aan de droge omgeving op warme dagen zou de vitaliteit van het pollen en de
receptiviteit van de stamper snel doen afnemen (Gay et al., 1987).
1.4.1 In vivo kieming versus in vitro kieming
Wanneer het pollen op een kiemingsmedium wordt aangebracht, waarvan enkele essentiële
componenten ontbreken, zal de kieming niet doorgaan. Kiemingstesten op een suboptimaal
medium zijn dus eerder een waarde-indicatie van het medium dan een kwaliteitsbepaling van
het pollen. Pollen dat in vivo kiemt, bezit deze essentiële componenten wel, wat zal
resulteren in een groter kiemingspercentage (Ylstra et al., 1992). Bij in vitro kieming zal de
pollenkiembuis slechts 30% tot 40% van de lengte van een in vivo gekiemde pollenkiembuis
bereiken (Taylor & Hepler, 1997).
In vivo kiemingstesten zijn een stuk technischer en arbeidsintensiever dan in vitro kieming.
Om snel tot resultaten te komen, wordt daarom beter gebruik gemaakt van in vitro
kiemingstesten.
Uit onderzoek blijkt dat bij de in vitro groei van Cucumis sativus L. minder en kleinere
bloemen worden gevormd, vergeleken met de groei in kassen. Er is ook aangetoond dat het
pollen, afkomstig van in vitro opgegroeide planten, minder vitaal is dan het pollen van
planten opgegroeid in serres (Kiełkowska & Havey, 2012).

12
1.4.2 Voorbehandelingen
Soms kan het nodig zijn om het pollen te behandelen alvorens ze in vitro te doen kiemen.
Niet elke voorbehandeling heeft een positief effect op de pollenkieming. Een
warmtebehandeling van 40°C gedurende 30 minuten of van 60°C gedurende 20 minuten,
beïnvloedt de pollenkieming niet. Een hydratatie vooraf levert eveneens geen hoger
kiemingspercentage op (Jayaprakash & Sarla, 2001). Sommige onderzoekers laten het
pollen eerst gedurende 60 minuten incuberen in een klimaatkast bij 24°C (Vizintin &
Bohanec, 2004). Een incubatie van het pollen in organische solventen levert positieve
resultaten op voor de kieming van bepaalde Cucurbitaceae. Een incubatieperiode van 60
minuten in benzeen resulteert bij Citrullus lanatus L., (Cucurbitaceae) in het hoogste
kiemingspercentage (Khan & Perveen, 2010).
1.4.3 Kiemingsmedium
Wanneer pollenkorrels op een geschikt voedingsmedium geplaatst worden, zullen ze kiemen
en een pollenkiembuis vormen (Vasil, 1960). Bij het plaatsen van de pollenkorrels op een
medium kan men bij sommige plantensoorten, waaronder tomaat, na 90 minuten bepalen of
het pollen al dan niet een pollenkiembuis heeft gevormd (Abdul-Baki, 1992).
Algemeen wordt gesteld dat een medium kan gebruikt worden in onderzoek naar
pollenbewaring, bestuiving en bevruchting wanneer het in staat is om 90% van het vitale
pollen te doen kiemen (Jayaprakash & Sarla, 2001).
Kiemingsmedia kunnen zowel vast als vloeibaar zijn en de optimale samenstelling varieert
naargelang de plantensoort (Vasil, 1960). In de meeste onderzoeken wordt gebruik gemaakt
van vloeibare media. Het pollen van een groot aantal plantensoorten kan kiemen in
gedestilleerd water, maar het aantal gekiemde pollenkorrels en de lengte van de
pollenkiembuis is vaak minder dan op media die suiker bevatten (Vasil, 1960). In een
vloeibaar medium zijn vaak calcium, boor en een osmoticum aanwezig. Toevoeging van
deze elementen leidt tot hogere kiemingspercentages, maar zelfs met de meest
geoptimaliseerde kiemingsmedia verloopt de in vitro kieming van pollenkorrels anders dan
de in vivo kieming. Bij in vitro kieming bereiken de pollenkiembuizen slechts 30 tot 40% van
de lengte van de pollenkiembuizen van de in vivo kieming (Taylor & Hepler, 1997).

13
1.4.3.1 Pollenbelasting
Kleine pollenpopulaties zullen zelden goed kiemen in een suikermedium of een suiker-agar
medium. De optimale pollenpopulatie voor de kieming is verschillend voor elke plantensoort.
Dit is ook te zien in Figuur 3 (Brewbaker & Kwack, 1963).
Figuur 3 - Relatie tussen het aantal pollenkorrels en het kiemingspercentage getest bij verschillende
sierplantensoorten (Brewbaker & Kwack, 1963).
1.4.3.2 Osmoticum
Naast het aantal pollenkorrels, is ook de samenstelling van het kiemingsmedium zeer
belangrijk. Wanneer een PEG (polyethyleenglycol) medium met een sucrose medium
vergeleken wordt, is de kieming op het PEG medium hoger dan op het sucrose medium
(Jayaprakash & Sarla, 2001). Op het PEG medium barst de pollenkiembuis niet, wat wel
voorkomt op een sucrose medium. Dit komt doordat PEG metabolisch inert is en de cellen
niet kan binnendringen. Sucrose geeft wel aanleiding tot een verhoging van de interne
concentratie, wat zorgt voor een verandering in de membraanpermeabiliteit van de
groeiende pollenkiembuis waardoor ionen en metabolieten in het medium terechtkomen. Bij
Angiospermen leidt een verhoogde sucroseconcentratie tot een verhoogde
ethanolfermentatie, hetgeen de groei van de pollenkiembuis remt. Het is nog ongekend of er
ook ethanolfermentatie optreedt bij de in vivo groei (Taylor & Hepler, 1997).
De sucroseconcentratie mag niet hoger dan 40% zijn. Een hoger sucrosegehalte zorgt voor
een duidelijke daling van de groei. Een sucroseconcentratie lager dan 2% leidt dan weer tot
veel gebarsten pollenkorrels (Brewbaker & Kwack, 1963). Pollenkorrels die later in het
seizoen geoogst worden, vertonen een grotere suikerbehoefte dan pollenkorrels die vroeger
worden geoogst in het seizoen (Vasil, 1960).
Onderzoek naar de pollenkieming en de groei van de pollenkiembuis bij Trichosanthus
dioica, toont aan dat in een medium met 1% suiker al kieming optreedt. Maximale kieming en
maximale lengte van de pollenkiembuis komen voor bij een 6% sucroseoplossing waaraan
0.01% boorzuur is toegevoegd. Bij verdere verhoging van het suikergehalte, was er een
daling in het kiemingspercentage en de pollenkiembuislengte te zien (Kumari, Komal,
Rajesh, & Pandey, 2009).

14
De kieming van het pollen van komkommer verloopt beter wanneer de sucrose in het
medium wordt vervangen door maltose (Vizintin & Bohanec, 2004). In Tabel 1 zijn de
resultaten van een onderzoek naar de optimale suikerconcentratie in het kiemingsmedium
van verschillende Cucurbitaceae weergegeven (Vasil, 1960).
Tabel 1 - Optimaal kiemingspercentage en pollenkiembuislengte voor verschillende Cucurbitaceae, in
suikeroplossing met 0.01% boorzuur (Vasil, 1960).
Naam
%suiker
%kieming
Gemiddelde pollenkiembuislengte
in micrometer
Benincasa hispida 10 60 5472 Citrullus vulgaris 7.5 70 2232 Cucumis melo 20 90 3960 C. melo var. momordica 7.5 92 2919 C. melo var. utilissimus 12.5-15 80 2970 Lagenaria vulgaris 35 80 2180 Luffa acutangula 10 70 2779 L. cylindrica 15 91 3312 Momordica charantia 10 82 2988
Uit Tabel 1 blijkt dat de optimale suikerconcentratie afhankelijk is van de plantensoort. De
optimale kieming en de langste pollenkiembuizen bij verschillende Cucurbitaceae worden
bekomen bij een suikerconcentratie tussen 7.5% en 20%. Er is ook gebleken dat, hoe lager
de osmotische concentratie van het kiemingsmedium, hoe groter het aantal opengebarsten
pollenkorrels (Vasil, 1960).
1.4.3.3 Veel gebruikte elementen
Boor
Een heel belangrijk element in een voedingsmedium, is boor en dan voornamelijk onder de
vorm van boraat. De stempel van bloemen bevat van nature boor, waardoor de kieming van
het pollen bevorderd wordt. Bij de in vitro kieming van pollen dient er boraat aan het medium
toegevoegd te worden (Johri & Vasil, 1961). Boraat verbetert de groei van de pollenkiembuis
bij een groot aantal species (Brewbaker & Kwack, 1963). Dit komt doordat boor de absorptie
van suikers, nodig voor de metabolische processen, bevordert door vorming van het suiker-
boraatcomplex. Daarnaast zorgt het boraat ook voor een verhoogde zuurstofopname en is
het betrokken in de pectinesynthese, noodzakelijk voor de vorming van de celwand van de
groeiende pollenkiembuis (Kumari et al., 2009) (Vasil, 1960).
Calcium
Toevoeging van calcium aan het medium bevordert de kieming en de groei van de
pollenkiembuis. Niet zomaar elk calciumzout kan hiervoor gebruikt worden. Zo leidt
overvloedige aanwezigheid van calciumoxalaat tot het openbarsten van pollenkorrels. Een
veel gebruikt calciumzout is Ca(N03)2.4H20, in concentraties die variëren van 50 tot 5000
ppm. Toevoeging van K+, Mg2+ en Na+ zorgt voor een betere pollenkiembuisgroei door de
calciumactiviteit in goede banen te leiden (Brewbaker & Kwack, 1963).

15
1.4.3.4 Andere elementen
Het toevoegen van agar aan een kiemingsmedium heeft geen significante effecten op de
kieming van het pollen. In sommige plantensoorten, waaronder Cucurbitaceae,
Leguminoseae en Oleaceae, zorgt agar wel voor een langere pollenkiembuis (Brewbaker &
Kwack, 1963).
Plantenhormonen worden ook soms toegevoegd aan een kiemingsmedium. Gibberelline
zorgt in Lilium longiflorum voor een snellere pollenkieming en versnelde groei van de
pollenkiembuis (Kato, 1955). De kieming van de pollenkorrels en de groei van de
pollenkiembuis bij Cucumis melo var. utilissimus wordt positief beïnvloed door toevoegen
van indolazijnzuur (Vasil, 1960). Kinetine heeft ook positieve effecten op de pollenkieming.
Bij Cucumis sativus L. is de vitaliteit en het kiemingspercentage hoger bij pollen dat wordt
geplaatst op een Murashige en Skoog medium waaraan 6 μM kinetine is toegevoegd
(Kiełkowska & Havey, 2012).
Andere groeiregulator bij pollen zijn flavonolen. De flavonolen quercetine, kaempferol en
myricetine hebben een zeer positief effect op zowel het kiemingspercentage als de lengte
van de pollenkiembuis bij Nicotiana tabacum L. Andere flavonolen hebben geen effect en de
positieve effecten zijn ook minder uitgesproken bij andere plantensoorten (Ylstra et al.,
1992).
Naast groeihormonen en groeiregulatoren zijn ook bepaalde antibiotica voordelig voor de
pollenkieming. Penicilline en streptomycine zijn gunstig voor de kieming en de groei van het
pollen van Cucumis melo var. utilissimus (Vasil, 1960).
Bepaalde vitaminen, waaronder biotine, bevorderen de kieming van de pollenkorrels en de
groei van de pollenkiembuis in Cucumis melo var. utilissimus. De effecten van de vitaminen
zijn wel niet zo uitgesproken als deze van andere groeiregulatoren. Biotine heeft het meeste
effect bij een concentratie van 1,25 ppm (Vasil, 1960).
Kokosnootmelk (optimum rond 2-10%) heeft bij Petunia en Ornithogalum een positief effect
op de kieming van pollenkorrels (Brewbaker & Kwack, 1963). Bij Cucumis sativus zorgt
toevoeging van gist voor een kleine stijging van het kiemingspercentage (Johri & Vasil,
1961).

16
1.4.3.5 Bestaande media
Een veel gebruikt medium voor het kiemen van pollen, is het medium van Brewbaker en
Kwack. Het medium bestaat uit: 10% sucrose, 100 ppm H3B03, 300 ppm Ca(N03)2.4H20, 200
ppm MgSO4. 7H20 en 100 ppm KNO3. Toevoegen van agar aan dit medium zorgt niet voor
een verbetering van de kieming van de pollenkorrels van verschillende plantenfamilies,
waaronder Cucurbitaceae en Oleaceae (Brewbaker & Kwack, 1963).
Voor de kieming van Cucumis sativus L. hebben Kiełkowska en Havey een eigen medium
uitgeprobeerd. Dit medium bestaat uit 15% maltose, 0,16 mM boorzuur, 1 mM calciumnitraat,
bij pH 7, en 0,25% Phytagel (Kiełkowska & Havey, 2012).
Voor de kieming van pollenkorrels van Cucumis sativus L. kan ook een eenvoudig medium
gebruikt worden. Dit medium bestaat uit: 15% sucrose, 0,25 mg/l H3BO3 en 1 mM Ca(NO3)2 ,
bij een pH 7 (Vizintin & Bohanec, 2004).

17
1.5 Vorming pollenkiembuis en bevruchting
1.5.1 Morfologie pollenkiembuis
De pollenkiembuis is essentieel bij het doorgeven van het DNA en zorgt ervoor dat de
mannelijke gameet tot bij de vrouwelijke gameet geraakt en de bevruchting kan optreden
(Taylor & Hepler, 1997) (Edlund et al., 2004).
In de apicale zone van de pollenkiembuis bestaat de celwand uit één laag. In de meer basale
zone van de pollenkiembuis bestaat de celwand uit twee lagen. De binnenste cellaag wordt
later gevormd dan de buitenste cellaag. De specifieke samenstelling van de pollenkiembuis
celwand is nog niet bekend. Wel is geweten dat er cellulose, pectine, hemicellulose en
callose in aanwezig is. In de apicale zone van de pollenkiembuis ontbreekt de cellulose
(Mascarenhas, 1993).
In de apex van de pollenkiembuis zijn secretorische cellen aanwezig. Deze secretorische
cellen bevatten de componenten voor de elongatie van de cellen en de uitbreiding van de
celwand. Actine microfilamenten zorgen voor het transport van deze componenten (Taylor &
Hepler, 1997). In de pollenkiembuis is de groei beperkt tot de apex. Het cytoplasma in de
zone die niet deelt, bevat een overmaat aan ER, vesikels, dictyosomen en mitochondriën. De
delende apicale zone bevat heel veel vesikels en zeer weinig andere cel componenten. Er
bestaan twee soorten vesikels in de apex: grote vesikels met een diameter van 300 nm en
vesikels met een diameter van 50 nm. De grootste vesikels zijn afkomstig van het Golgi
apparaat en bevatten de precursoren voor de pollenkiembuis celwand (Mascarenhas, 1993).
De gestockeerde elementen in het pollen zijn afkomstig van de tapetumcellen en worden
tijdens de pollenkiembuisgroei gemobiliseerd (Jóhannsson & Stephenson, 1998). Onderzoek
heeft aangetoond dat kiembuizen die sneller groeien, meer kans maken op bevruchting dan
traag groeiende pollenkiembuizen. De onderlinge competitie is groter bij toenemende
hoeveelheid pollenkorrels op de stempel (Winsor & Stephenson, 1995).
1.5.2 Bevruchting
Tijdens de bevruchting versmelt het mannelijke plantenmateriaal met het vrouwelijke
plantenmateriaal. Omdat Cucurbita pepo een éénhuizige plant is, dienen pollen van de ene
bloem naar de andere bloem te worden getransporteerd (Ciampolini et al., 1993).
Wanneer de pollenkorrels rijp zijn, hebben ze twee spermacellen geproduceerd, die alle
nodige informatie voor de bevruchting van de eicel bevatten (Edlund et al., 2004)
(Mascarenhas, 1993).

18
1.5.2.1 Vrouwelijke bloemen
De bloemen van de Cucurbitoideae hebben één stempel (Jeffrey, 1980). De vrouwelijke
bloemen van Cucurbita pepo zijn gemakkelijk te onderscheiden van de mannelijke. De bloem
heeft een langwerpige, onderstandige basis, een opgezwollen stijl en drie gestigmatiseerde
lobben (das Graças Vidal et al., 2006). De bloemstengel is veel kleiner in vergelijking met de
mannelijke bloemen. De bloemen verschijnen vanaf het vierde tot het vijfde blad (Nepi &
Pacini, 1993). Twee dagen voor de anthese verkleuren de petalen van de vrouwelijke
bloemen aan de randen. Ze krijgen een geel tot oranje achtige kleur. Na de anthese kleurt de
stempel bruin, waardoor je visueel de ontvankelijkheid van de stempel kan waarnemen. De
ontvankelijkheid van de stempel duurt vier dagen: van een dag voor de anthese tot twee
dagen nadien. Indien het pollen later op de stempel landt, zal de pollenkiembuis nog
beginnen groeien, maar zal deze afsterven voordat er bevruchting kan optreden (Nepi &
Pacini, 2001) (Nepi & Pacini, 1993).
De vrouwelijke bloemen van een plant bestaan uit een stempel, stijl en vruchtbeginsel. Er
bestaan twee soorten stempels, natte stempels en droge stempels. Men spreekt van een
natte stempel als de oppervlaktecellen een vloeistof bestaande uit lipiden, proteïnen en
pigmenten, secreteren. Droge stempels zijn nog eens extra omgeven door een waslaag. Het
is de stempel die de pollenkorrels ontvangt. Sommige stempels inhiberen de aanhechting en
kieming van pollenkorrels van dezelfde plant om zelfbestuiving te voorkomen (Edlund et al.,
2004). Bij courgette kan er wel zelfbestuiving optreden (Nepi & Pacini, 1993).
Wanneer het pollen op een natte stempel landt, worden de pollenkorrels onmiddellijk
omgeven door een waterlaag. Pollenkorrels die op droge stempels landen, mobiliseren hun
pollen coat om zo een verbinding tussen de twee cellagen te vormen. Water, nutriënten en
andere moleculen worden snel naar de pollenkorrels getransporteerd volgens een
mechanisme dat nog niet volledig duidelijk is. Wanneer de pollenkorrels te vroeg
gehydrateerd worden, kan dit leiden tot kieming van het pollen op de antheren (Edlund et al.,
2004).
De reproductie van Angiospermen is heel complex. Zo is het vrouwelijke weefsel in staat om
verschillende soorten pollenkorrels te herkennen en enkel het pollen van aanverwante
soorten te laten kiemen. Deze selectiviteit gaat gepaard met een heel grote diversiteit van
celoppervlakken van de vrouwelijke en mannelijke voortplantingsorganen bij een plant
(Edlund et al., 2004).

19
1.5.2.2 Mannelijke bloemen
De mannelijke bloemen bij Cucurbita pepo lijken sterk op de vrouwelijke bloemen. Ze zijn
even groot en eveneens oranjegeel. Een eerste punt waarin ze verschillen is de afwezigheid
van een vruchtbeginsel; de bloemen staan op een slanke steel. Ook verschillen ze van de
vrouwelijke bloemen door de aanwezigheid van de mannelijke geslachtskenmerken: vijf
meeldraden waarvan de filamenten en de antheren vergroeid zijn tot één centrale helmknop.
Bij Cucurbita pepo produceren zowel de mannelijke als de vrouwelijke bloemen nectar (das
Graças Vidal et al., 2006). De geproduceerde hoeveelheid nectar is kleiner dan deze van de
vrouwelijke bloemen. De nectarsamenstelling is bij beide geslachten gelijk (Nepi et al., 2001)
(Ciampolini et al., 1993).
1.5.2.3 Eigenlijke bevruchting
Wanneer de pollenkorrels op de stempel van de vrouwelijke bloemen vallen, zullen deze
hydrateren (Nepi et al., 2001). Hydratatie van de pollenkorrels zorgt voor een transformatie
van een niet-polaire cel naar een polaire cel. Na de hydratatie zullen het cytoplasma en het
cytoskelet zich reorganiseren waardoor de interne turgordruk verhoogt en een verweking van
de exinelaag optreedt. Dit zorgt ervoor dat de pollenkiembuis door de exinelaag kan breken,
waarna de pollenkiembuis de pollenkorrel zal verlaten. Het DNA, de aanwezige proteïnen en
kleine bioactieve moleculen van het pollen zorgen voor de snelle kieming van de pollenkorrel
(Edlund et al., 2004). De pollenkiembuis groeit vervolgens doorheen de stijl tot aan het
vruchtbeginsel. Deze bevindt zich onder de bladeren van de vrouwelijke bloem en bevat de
ovules of zaadknoppen. De ovules zullen uitgroeien tot de vrouwelijke gametofyten, die de
embryozakken bevatten. In elke embryozak zitten de eicel (n), de centrale cel (2n), de twee
synergide cellen en de drie antipodale cellen (Van Labeke, 2016). De pollenkiembuis zal
doorheen de micropylle, een opening in de integumenten die de embryozak omgeven, de
embryozak binnendringen waar hij met één van de synergide cellen fuseert. Door deze fusie
zullen de vegetatieve kern van de pollenkorrel en de synergidekern degenereren (Edlund et
al., 2004).
De bevruchting is een tweeledig proces. De fusie van de eicel met de ene zaadcel leidt tot de
zygote. Uit de zygote ontstaat het embryo. De fusie van de centrale cel en de tweede
spermacel leidt tot het ontstaan van het endosperm (Van Labeke, 2016).
De pollenkiembuisgroei, van kieming tot bevruchting, duurt bij Cucurbita pepo L. twaalf tot
achtenveertig uur. De gemiddelde groei is groter dan 2 mm/uur (Nepi & Pacini, 2001). In het
begin van de groeicyclus is de cel elongatie groter dan op het einde van de groeicyclus
(Vasil, 1960). Deze groeisnelheid is afhankelijk van de vitaliteit van het pollen alsook de
omgevingstemperatuur. Boven 40°C en onder 10°C is pollenkiembuisgroei niet meer
mogelijk (Nepi & Pacini, 2001). Bij Cucurbita pepo, heeft pollen dat kiemt bij warme
temperaturen (30°C) een grotere kiembuislengte dan pollen dat kiemt bij 20°C (Jóhannsson
& Stephenson, 1998).

20
De pollenkieming en kiembuisgroei zijn verdeeld in vier fases: de imbibitie fase waarin het
pollen hydrateert en toeneemt in volume, de initiatie fase waarin de aanmaak van de
pollenkiembuis start en tenslotte de elongatie fase waarin de pollenkiembuis groeit door
celdelingen. De lengte van iedere fase is afhankelijk van de plantensoort (Kumari et al.,
2009) (Taylor & Hepler, 1997).
De ontvankelijkheid van de stempel, de stijl en het vruchtbeginsel moeten overlappen
voordat er bevruchting kan optreden. Het pollen dat dan op de stempel landt, moet vitaal zijn.
Met vitaal wordt hier bedoeld dat het pollen in staat moet zijn om te kiemen, de stempel en
de stijl te doorboren om uiteindelijk het vruchtbeginsel met de eicel te bereiken (Nepi &
Pacini, 2001).
1.5.2.4 Beïnvloedende factoren bevruchting
Vrucht- en zaadvorming in bloeiende planten wordt beïnvloed door verscheidene abiotische
en biotische factoren (Nepi & Pacini, 2001).
Bestuivers
Bestuivers spelen een belangrijke rol in het behoud van plantensoorten en de bevruchting
van land- en tuinbouwgewassen. Insecten, en dan voornamelijk bijen, zijn de belangrijkste
bestuivers wereldwijd. Voor de courgetteteelt in Vlaanderen wordt ofwel uitsluitend, ofwel
aanvullend op honingbijen, beroep gedaan op hommelkolonies. Daarnaast leveren ook wilde
soorten een belangrijke bijdrage in de teelten in open lucht. Een daling van de
honingbijpopulatie resulteert in een verminderde bestuiving en navenante opbrengst van het
gewas. De laatste jaren hebben de bijenpopulaties sterk te lijden onder de aanwezigheid van
de parasitaire mijt Varroa destructor (Potts et al., 2010).
Voor de verspreiding van het pollen doet Cucurbita pepo beroep op bestuivers want de wind
zorgt voor onvoldoende verspreiding. Deze moeten liefst overvloedig aanwezig zijn tijdens
de bloei van de bloemen van Cucurbita pepo L (Nepi & Pacini, 2001). Bestuivers worden
aangetrokken tot bepaalde bloemen door een combinatie van factoren. De kleur, grootte,
vorm en toegankelijkheid zijn belangrijk in het beslissingsproces van de bestuiver, maar ook
de hoeveelheid en de samenstelling van het nectar of de aanwezigheid van pollen
(Ciampolini et al., 1993).
Tijdstip van openen
De bloemen van Cucurbita pepo zijn slechts zes uur per dag open (Nepi & Pacini, 2001).
Zowel de mannelijke als de vrouwelijke bloemen openen in de zomer tussen 5h00 en 6h00,
dit is het uur waarop de zon opkomt. In de andere seizoenen gaan de bloemen later open,
door de later opkomen van de zon. Rond de middag sluiten de bloemen, om nooit meer open
te gaan. Het sluiten van de bloemen is eveneens seizoensafhankelijk (Nepi & Pacini, 1993).

21
Bodemcondities
De kwaliteit en de hoeveelheid zaden die een plant produceert, zijn afhankelijk van de
vruchtbaarheid en vochtigheid van de bodem. De omstandigheden waarin de sporofyt groeit,
beïnvloeden naast de pollenproductie, ook de vitaliteit en de kieming van het pollen (Lau &
Stephenson, 1993).
Het stikstofgehalte in de bodem blijkt een belangrijke factor in de pollenkieming. Dit komt
doordat stikstof in veel macromoleculen voorkomt, die zowel de kieming van het pollen
alsook de groei van de pollenkiembuis beïnvloeden. Stikstof is zeer vaak de beperkende
factor in de groei en reproductie van courgette. Planten die een hogere concentratie stikstof
toegediend krijgen, produceren tot 14% meer pollen en zwaardere zaden dan
courgetteplanten die behandeld zijn met een lagere concentratie stikstof. Bovendien kiemen
de pollenkorrels sneller en zal de pollenkiembuis sneller groeien bij een hogere
stikstofconcentratie dan bij een behandeling met een lagere stikstofconcentratie (Lau &
Stephenson, 1993).
Pollenbelasting
De lengte en het gewicht van de vrucht, alsook het aantal zaden dat de vrucht bevat, is
afhankelijk van de ‘pollen load’ dit is het aantal pollen dat op de stempel wordt aangebracht.
Een lage pollen load resulteert in onvoldoende bevruchte ovules en bijgevolg spitse vruchten
(Nepi & Pacini, 2001).
Klimaat
Natte of koude weersomstandigheden zorgen voor een vertraging van de bloemontwikkeling.
Dit heeft een rechtstreeks effect op de aanwezigheid van bestuivers. Hoe later de anthese,
hoe later er bestuivers nodig zijn (Bertin & Sholes, 1993). Door overvloedige regenval kan
het ook zijn dat de antheren later opengaan, waardoor het pollen reeds in de antheren begint
te kiemen of waardoor teveel pollen zullen openbarsten (Hoekstra, 1995) (Vasil, 1960).
Tijdens de meiose zijn de microsporen zeer gevoelig voor hoge temperaturen, alsook tijdens
de anthese. Negatieve gevolgen van een temperatuur boven 33°C bij paprika zijn kleine,
gekrompen pollenkorrels en lege korrels. Deze afwijkingen zijn ook te zien bij andere
plantensoorten, waaronder bonen en sojabonen, wanneer ze aan hoge temperaturen worden
blootgesteld (Reddy & Kakani, 2007). Pollen dat geoogst wordt op een warme dag, zal een
zeer lage vitaliteit bezitten en bijgevolg ook slecht kiemen (Vasil, 1960). Een temperatuur
hoger dan 32.8°C leidt tot, een kortere pollenkiembuis, een verminderde bevruchting en
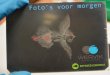
minder vruchtvorming. Dit is ook te zien in Figuur 4 (Reddy & Kakani, 2007).
22
Figuur 4 – Invloed van de temperatuur op (A) pollen kiemingspercentage and (B) pollenkiembuislengte bij verschillende Capsicum soorten (Reddy & Kakani, 2007).
Naast de temperatuur en het tijdstip van oogsten, is ook het moment in de teelt bepalend
voor het kiemingspercentage. Pollen van Cucurbitaceae, dat later in het seizoen geoogst
wordt, zal een verminderde kieming vertonen. Bovendien zal de lengte van de
pollenkiembuis ook duidelijk kleiner zijn dan deze van pollenkorrels van vroeger in het
seizoen (Vasil, 1960). Bij duivenerwt ligt het kiemingspercentage van het pollen hoger
wanneer er minder bloemen aanwezig zijn op de plant (Jayaprakash & Sarla, 2001).

23
1.6 Bewaring van pollenkorrels
Doordat er zoveel problemen zijn in de bestuiving van de vrouwelijke bloemen van
courgetteplanten, zijn onderzoekers sterk geïnteresseerd naar de eventuele mogelijkheden
tot bewaring van pollenkorrels. Indien men tot een goede bewaring van pollenkorrels kan
komen, kunnen problemen met planten die op een verschillend tijdstip of op een andere
plaats bloeien, opgelost worden (Perveen & Ali, 2011). Concreet betekent dit voor courgettes
dat bij aanvang van de teelt, wanneer mannelijke bloemen nog veelvuldig voorkomen, pollen
geoogst zou kunnen worden, om dan in tijden van pollentekort ingezet te worden. Er werd
reeds onderzoek uitgevoerd naar de mate waarin pollenkorrels kunnen bewaard worden in
klimaatkasten en wat de invloed hiervan was op het watergehalte, de vitaliteit en het
suikergehalte van de pollenkorrels.
1.6.1 Temperatuur en relatieve vochtigheid
De twee belangrijkste factoren bij de bewaring van pollen zijn de temperatuur en de relatieve
vochtigheid van de omgeving. Deze factoren bepalen in grote mate de vitaliteit van het pollen
(Khan & Perveen, 2010). Algemeen kan gesteld worden dat een lage relatieve vochtigheid
van 0% tot 30% tijdens de bewaring van het pollen, zorgt voor een maximale
bewaarbaarheid, tot zelfs een verdrievoudiging (Johri & Vasil, 1961) (Holman & Brubaker,
1926). Bij Cucurbita pepo blijft de pollenvitaliteit bij 30% RV de eerste 60 minuten hoog om
vervolgens snel af te nemen en 0 te worden na 180 minuten. Bij 75% RV treedt de sterke
daling in vitaliteit pas op na 120 minuten en wordt de vitaliteit 0 na 4 uur. Dit is weergegeven
op Figuur 5 (Nepi et al, 2009).
Over de optimale bewaarstrategie van het pollen van Cucurbita pepo bestaat er onenigheid
tussen de verschillende auteurs. Volgens (Vasil, 1960) wordt het pollen van Cucurbitaceae
het best bewaard bij een relatieve vochtigheid van 20% tot 50% en een temperatuur tussen -
5°C en 8°C (Vasil, 1960). Andere onderzoekers tonen aan dat pollen van Cucurbitaceae een
hoger kiemingspercentage hebben als ze bewaard worden bij nog lagere temperaturen, met
een optimum rond -30°C (Perveen & Ali, 2011). Pollen van Cucurbita pepo, dat bewaard
wordt bij 5°C of hoger, is na vier dagen niet meer vitaal (Gay et al., 1987).
1.6.2 Watergehalte van pollenkorrels
Pollenkorrels worden verspreid in een gedeeltelijke gedehydrateerde toestand. Pollen van
verschillende plantensoorten wordt in verschillende klasses opgedeeld, naargelang het
watergehalte ze bezitten. Een eerste groep heeft een gemiddeld watergehalte tussen 5% en
20%, waardoor ze enkele dagen zonder water kunnen overleven. Daarnaast bestaat er ook
een groep waarvan pollenkorrels een hoger watergehalte bevatten, tot wel 50%. Deze
pollenkorrels worden ook wel gedeeltelijke gehydrateerde pollenkorrels genoemd. Het zijn
grote pollenkorrels met grote poriën. Ze vertonen een zeer snelle kieming, hebben ze een
lage sucroseconcentratie en hebben een hoge vitaliteit (Nepi et al., 2009). Cucurbita pepo
heeft gedeeltelijk gehydrateerde pollenkorrels. Het watervolume van het pollen bij Cucurbita
pepo bedraagt ongeveer 55% bij het openspringen van de antheren en zakt naar 35% op het
einde van de anthese (Nepi et al., 2001).
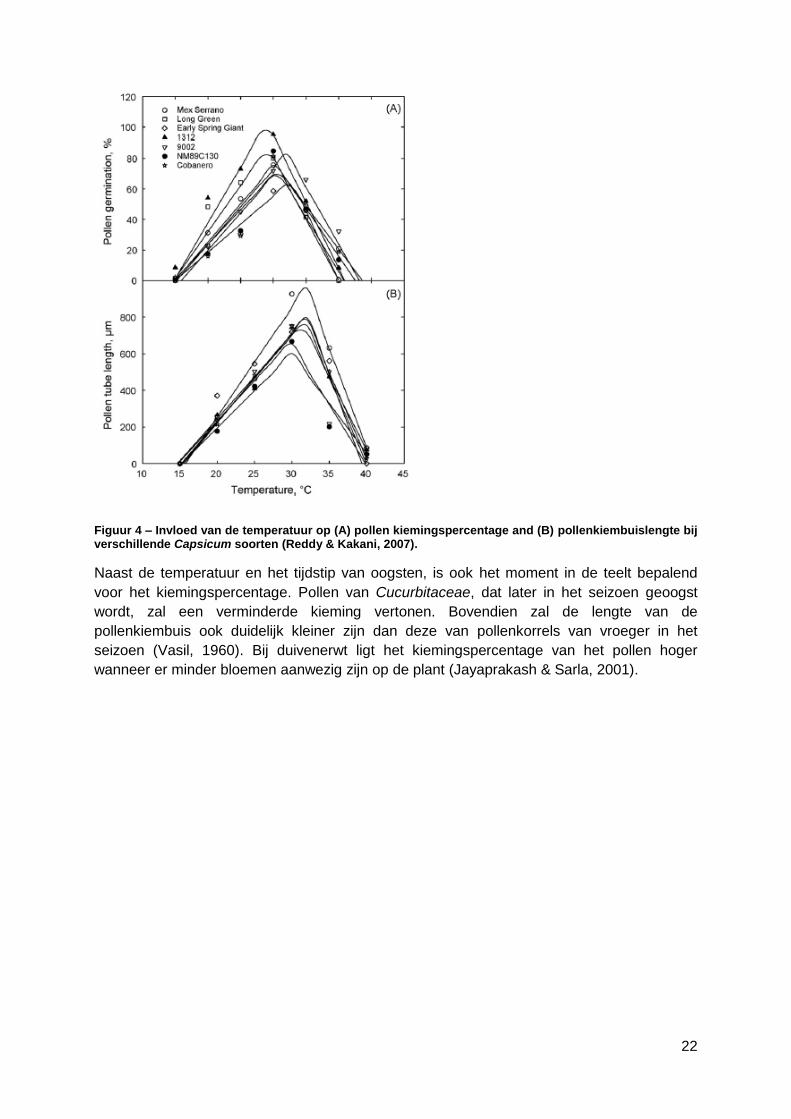
24
Een voorwaarde voor pollenkorrels is, dat ze na dehydratatie terug kunnen re-hydrateren.
Water is één van de bepalende factoren in de levensvatbaarheid van het pollen. Een verlies
aan water, zonder re-hydratatie, of een verminderde wateropname heeft een negatieve
invloed op de aanhechting en de kieming van de pollenkorrels en leidt tot een vermindering
van de pollenvitaliteit (Taylor & Hepler, 1997). Bij Cucurbita pepo leidt een te grote daling in
het watergehalte tot een onherstelbare schade van het pollen (Kumari et al., 2009).
De resultaten van het onderzoek van het watergehalte bij Cucurbita pepo van (Nepi et al.,
2009) zijn weergegeven in Figuur 5. Het geeft de vitaliteit en het watergehalte weer van de
courgettepollen, wanneer onderworpen aan twee verschillende relatieve vochtigheden. Bij
een relatieve vochtigheid van 30% neemt het watergehalte van het pollen snel af. Na 30
minuten is het al gehalveerd van 40% naar 20%; na anderhalf uur bedraagt het nog slechts
10% en vanaf 2 uur stabiliseert het vochtgehalte zich rond 5%. Bij een relatieve vochtigheid
van 75% neemt het watergehalte iets trager af. De halvering treedt pas op na 1 uur en zakt
pas na 5 uur onder de 10% (Nepi et al., 2009).
Figuur 5 - Watergehalte en vitaliteit van Cucurbita pepo pollen bij een relatieve vochtigheid van
respectievelijk 30% en 75% (Nepi et al., 2009).
Het pollen van Cucurbita pepo kan slechts gedurende een korte periode bewaard worden
alvorens het zijn vitaliteit verliest. Dit verlies aan vitaliteit gaat trager bij een hogere relatieve
vochtigheidsgraad. Wanneer het watergehalte van de pollenkorrels onder de 24% komt, zal
er geen zaadzetting gebeuren, hetgeen belangrijk is voor vruchtvorming (Gay et al., 1987).

25
1.6.3 Suikergehalte van pollenkorrels
Naast het watergehalte en de vitaliteit van de pollenkorrels, is ook het suikergehalte en het
zetmeelgehalte van de pollenkorrels belangrijk voor bewaring op lange termijn. De resultaten
van dit onderzoek zijn weergegeven in Figuur 6. Het gehalte aan fructose, glucose, sucrose
en zetmeel bleef gedurende de gehele bewaarperiode constant (Nepi et al., 2009). Een
verhoogd gehalte aan dissaccharides, zorgt voor een betere weerstand tegen uitdroging van
het pollen. Dit komt doordat sucrose water vervangt door OH-groepen en hierdoor de
membranen beschermt tegen wijzigingen in membraanstructuur door uitdroging. Bovendien
zorgen opgeloste stoffen ook voor een vriespuntverlaging wat de pollenkorrel beschermt
tegen kristallisatie van de polleninhoud bij invriezen (Hoekstra, 1995).
Figuur 6 - Sucrose-, fructose-, glucose- en zetmeelgehalte bij Cucurbita pepo pollen bij een relatieve
vochtigheid van 30% (Nepi et al., 2009).

26
2 Materiaal en methode
Het pollen, gebruikt voor de experimenten, is afkomstig van zes verschillende
courgetterassen. De onderzochte rassen zijn: Bengal (Seminis), Precioza (Hazera Seeds),
Tosca (Clause), Noriac (Gaultier), Storrs Green (Seminis) en Milos (Syngenta).
De planten werden gezaaid begin december 2016 op Inagro, waar ze in een donkere ruimte
tot kieming werden gebracht. Tot het eerste echte bladstadium stonden de planten in een
stookruimte, onder 100 μmol belichting en met een gemiddelde opkweektemperatuur van
17,5°C tussen 19 december 2016 en 9 januari 2017. Op 27 december werden de
kiemplanten opgepot in potten met kokossubstraat van het merk Copertiz, met een pH
waarde van 5,9, een granulometrie van 0-12 mm en een waterretentiecapaciteit van 350 ml/l.
Meteen na het oppotten kregen de planten een eerste preventieve Vertimecbehandeling
tegen trips.
Op 9 januari 2017 werden de planten getransporteerd naar de serres van de UGent, waar ze
opgebonden werden en A. swirskii geïntroduceerd werd als biologische bestrijder tegen trips
(F. occidentalis). Er werden 1,5 planten per m2 geplant. Bij alle planten werden de
vrouwelijke vruchtjes voor de bloei weggenomen. De voeding van de planten was de
samengestelde meststof Solufeed met NPK 19-8-16 + 4MgO + micro-elementen tot een EC-
waarde van 2,3 mS/cm werd bereikt. Dit werd aangevuld met CaNO3 totdat een EC-waarde
van 3 mS/cm bereikt werd. Fertigatie gebeurde volgens de goede landbouwkundige praktijk.
De water- en nutriëntenvoorziening was steeds voldoende.
2.1 Kiemingstesten
Voor alle kiemingstesten werd pollen van het ras Bengal gebruikt. Om het
kiemingspercentage van het pollen na te gaan, werden verschillende basismedia getest
(Tabel 2 - Overzicht van de geteste kiemingsmedia). Ieder medium werd in vier herhalingen
getest. Per medium werden ongeveer 150 - 200 pollenkorrels getest. Het beste basismedium
werd verder gebruikt om het effect van toevoegingen van sporenelementen na te gaan. De
kieming van het pollen werd na 3 uur en 24 uur incuberen in het medium geëvalueerd. Een
pollenkorrel werd als gekiemd beschouwd als de pollenkiembuis groter was dan de diameter
van de pollenkorrel.

27
Tabel 2 - Overzicht van de geteste kiemingsmedia
Mediumnummer Samenstelling medium
1
10% sucrose 100 ppm H3B03 300 ppm Ca(N03)2.4H20 200 ppm MgSO4. 7H20 100 ppm KNO3
2
10% maltose 100 ppm H3B03 300 ppm Ca(N03)2.4H20 200 ppm MgSO4. 7H20 100 ppm KNO3
3
10% PEG 100 ppm H3B03 300 ppm Ca(N03)2.4H20 200 ppm MgSO4. 7H20 100 ppm KNO3
4
15% sucrose 0,25 mg/l H3BO3
1 mM Ca(NO3)2
5
2 - 20% maltose 100 ppm H3B03 300 ppm Ca(N03)2.4H20 200 ppm MgSO4. 7H20 100 ppm KNO3
6
20% maltose 100 ppm H3B03 300 ppm Ca(N03)2.4H20 200 ppm MgSO4. 7H20 100 ppm KNO3
5 – 20% agar
7
20% maltose 100 ppm H3B03 300 ppm Ca(N03)2.4H20 200 ppm MgSO4. 7H20 100 ppm KNO3 100 – 250 ppm gist
8
20% maltose 100 ppm H3B03 300 ppm Ca(N03)2.4H20 200 ppm MgSO4. 7H20 100 ppm KNO3 1,29 – 2,58 ppm kinetine
9 20% maltose 100 ppm H3B03 300 ppm Ca(N03)2.4H20 200 ppm MgSO4. 7H20 100 ppm KNO3 1,25 ppm biotine

28
10
20% maltose 100 ppm H3B03 300 ppm Ca(N03)2.4H20 200 ppm MgSO4. 7H20 100 ppm KNO3 1,25 – 2,50 ppm indolazijnzuur
11
20% maltose 100 ppm H3B03 300 ppm Ca(N03)2.4H20 200 ppm MgSO4. 7H20 100 ppm KNO3 1,29 ppm kinetine 100 ppm gist
12
20% maltose 100 ppm H3B03 300 ppm Ca(N03)2.4H20 200 ppm MgSO4. 7H20 100 ppm KNO3 1,29 ppm kinetine 1,25 ppm indolazijnzuur
13
20% maltose 100 ppm H3B03 300 ppm Ca(N03)2.4H20 200 ppm MgSO4. 7H20 100 ppm KNO3 2,58 ppm kinetine 1,25 ppm indolazijnzuur
14
20% maltose 100 ppm H3B03 300 ppm Ca(N03)2.4H20 200 ppm MgSO4. 7H20 100 ppm KNO3 1,29 ppm kinetine 1,25 ppm indolazijnzuur 1,25 ppm biotine
Nadat er gekozen was voor medium 2 als basismedium, werd in medium 5 gekeken of er
een verschil in pollenkieming was bij toenemende maltoseconcentraties. De geteste
concentraties waren: 2%, 5%, 10%, 15% en 20% maltose.
Om na te gaan of toevoegen van agar tot een hoger kiemingspercentage leidde, werden in
medium 6 verschillende agarconcentraties getest. Voor agar waren de geteste concentraties:
5%, 10%, 15% en 20%.
In medium 7 werd het effect van gist getest, in concentraties gaande van 100 ppm tem 250
ppm, met stappen van 50 ppm. Ook in medium 8 werden verschillende concentraties getest.
Kinetine werd getest in 1,29 ppm en 2,58 ppm. In medium 10 werd IAA ook in dubbele
concentratie getest.

29
2.2 Zetmeel- en suikergehalte
Voor de zetmeel- en suikeranalyses van het pollen, werd van 13 februari tot 1 maart iedere
dag pollen geoogst tussen 8u en 9u ’s ochtends, dit was kort na het opengaan van de
bloemen (tussen 7 en 8u ’s ochtends). Het oogsten van het pollen gebeurde door de
helmknop van de mannelijke bloemen tegen de rand van een epje te strijken. Het pollen
werd per ras samengebracht en bewaard bij -80°C, dit in de veronderstelling dat bij deze
temperatuur geen wijzigingen in het zetmeel- en suikergehalte van het pollen plaatsvindt.
Van vier rassen werden 10 stalen verzameld, die getest werden op hun suiker- en
zetmeelgehalte. Bij Noriac en Precioza werden respectievelijk 9 en 7 stalen geanalyseerd.
Dit gebeurde via het standaardprotocol van het labo plantaardige productie.
In een eerste fase van de analyse vond een extractie met ethanol plaats, waarbij het gewicht
van de stalen afhing van de hoeveelheid pollen dat kon geoogst worden op de
desbetreffende dagen. Suikers gaan in oplossing in ethanol, maar het zetmeel niet. Het
zetmeel, dat in het pellet van de ethanol extractie zat, werd opgelost door middel van een
zure hydrolyse met HCl. Na de zure hydrolyse werd gebruik gemaakt van de reagentiakit van
Roche om de zetmeelkwantificatie te scoren met de spectrofotometer. Een uitgebreide
beschrijving van het protocol is weer te vinden in de bijlages.
Het supernatans van de ethanolextractie werd gebruikt voor de analyse van mono- en
disacchariden met de HPLC (scheidingskolom PA-20, Dionex).

30
2.3 Pollenvitaliteit na bewaring bij -80°C
In een eerste experiment werd de vitaliteit van het pollen van de rassen Bengal en Noriac
onderling vergeleken. Het pollen werd geoogst op verschillende dagen, werd vervolgens
bewaard bij -80°C, om na de gewenste bewaarperiode het vitaliteitspercentage van het
pollen te testen. Van elk ras werden vier herhalingen genomen. De vitaliteit werd nagegaan
door middel van de FDA-kleuring.
Voor de FDA-kleuring werd gebruik gemaakt van een FDA-stock, die in de diepvries
bewaard werd. De FDA-stock bestond uit 5 mg FDA, dat werd opgelost in 1 ml aceton. Er
werd 8 μl FDA-stock toegevoegd aan 1 ml van een 20% sucroseoplossing. Per well van een
96-well plaat werd 200 μl oplossing aangebracht waarin 100 tot 200 pollen werden
toegevoegd met een fijn penseel. Nadien werden de platen omhuld met aluminiumfolie
gedurende 1 uur. Na 1 uur incuberen vond de analyse plaats onder de microscoop, zowel
onder BF als GFP, bij 488 nm. Voor de vitaliteitanalyse werd een programma gebruikt, dat
geschreven werd door Ir. Simon Craeye. De gebruikte programma’s staan in de bijlage.
2.4 Pollenvitaliteit na bewaring bij verschillende temperaturen
Het tweede experiment was een vergelijking tussen vier verschillende bewaarmethodes:
bewaring bij kamertemperatuur, bewaring in de frigo (6°C), bewaring in de diepvries (- 20°C)
en bewaring bij -80°C. Op 23 maart 2017 werd pollen geoogst in de serres van de UGent
van de rassen Noriac en Bengal en werd de vitaliteit van vers pollen van beide rassen
vergeleken. Enkele dagen later, op 27 maart 2017, werden alle stalen, zowel op het ILVO als
van de serres van de UGent, verzameld voor de bewaring bij verschillende temperaturen. Bij
dit experiment werd enkel pollen van het ras Bengal gebruikt. De stalen voor de vitaliteitstest
van vers pollen zijn afkomstig van planten op het ILVO. Voor de bewaring bij verschillende
temperaturen werden stalen van de serres op de UGent genomen. Van deze stalen werd de
vitaliteit van het vers pollen niet nagegaan, deze stalen werden meteen bewaard bij de
vooropgestelde temperatuur. Op verschillende tijdstippen (na 1 dag, 2 dagen, 4 dagen en 7
dagen bewaring) werd de vitaliteit van het pollen nagegaan door gebruik te maken van de
FDA-kleuring. De FDA-kleuring werd uitgevoerd zoals beschreven onder 2.3.

31
2.5 Manuele bestuiving
Om na te gaan wat het effect van bewaring van het pollen op de bevruchting en de uitgroei
van de vruchten is, werden enkele bloemen manueel bestoven met bewaard pollen. Deze
proef ging door in de plastiekserres 11 en 12 van het proefcentrum Inagro. Het gewas (ras
Tosca) werd geplant op 7 maart 2017 in vollegrond (zandleembodem) en kan als een
voorjaarsteelt bestempeld worden. Overdag werd een temperatuur van 16°C aangehouden,
’s nachts daalde de temperatuur tot 10°C. De watergift gebeurde door middel van
druppelirrigatie met T-tape en was gedoseerd volgens goede landbouwkundige praktijk.
Pollen werd op verschillende dagen geoogst en bewaard bij 5°C. Voor de manuele
bestuiving werd vers geoogst pollen gebruikt, naast pollen dat respectievelijk 5 dagen, 10
dagen en 15 dagen bewaard was. Tot slot werden ook enkele bloemen onbestoven gelaten.
Elk van deze objecten werd in 10 herhalingen getest.
Om zeker te zijn dat er geen ongewenste bezoeken door bestuivers zouden optreden,
werden de vrouwelijke bloemen, de avond voor de manuele bestuiving, afgedekt met plastic
zakjes met microperforaties. De dag van de bloei werd het plastic zakje voorzichtig
afgenomen, pollen manueel op de stempel van de vrouwelijke bloemen aangebracht en werd
de bloem terug afgesloten met behulp van een tomatenclips. Dit is te zien op Figuur 7. Een
week na het aanbrengen van het pollen, werden de courgettes beoordeeld op vruchtgrootte
en vruchtgewicht.
Figuur 7 – (a) Foto van een vrouwelijke bloem afgedekt met een plastic zakje met microperforaties en (b) van een bloem gesloten met een tomatenclip
a b

32
2.6 Statistische verwerking
Voor de berekening van de gemiddelden en standaardafwijkingen van de experimenten werd
gebruik gemaakt van SPSS. Bij het vergelijken van het vitaliteitpercentage van de rassen
Noriac en Bengal, werd gebruikt gemaakt van de t-test om na te gaan of er een significant
verschil was tussen beide rassen (p=0,05). Voor de bewaring bij verschillende temperaturen
en de analyses van het suiker- en zetmeelgehalte van alle onderzochte rassen, werden de
resultaten onderling vergeleken in een One-Way ANOVA analyse (p=0,05). Indien uit de
ANOVA analyse bleek dat er significante verschillen waren tussen de verschillende
temperaturen en rassen, werd een post-hoc analyse (Tukey test, p=0,05) gedaan om te zien
waar de verschillen zich bevonden.

33
3 Resultaten
3.1 Pollenkieming
3.1.1 Algemene observaties
Na 24 uur incuberen in het kiemingsmedium liggen de pollen meer gegroepeerd dan na 3
uur incuberen. Bovendien zijn er na 24 uur meer pollenkorrels die een beginnende
pollenkiembuis, die niet langer dan de diameter van de pollenkorrels is, gevormd hebben.
3.1.2 Basismedium
Om na te gaan welk basismedium het beste was, werden medium 1, medium 2, medium 3
en medium 4 onderling vergeleken (zie Tabel 2). Medium 1 met sucrose vertoonde geen
kieming, maar wel veel opengebarsten pollenkorrels. Wanneer het sucrose vervangen werd
door maltose (medium 2), was er eveneens geen kieming, maar de pollenkorrels sprongen,
op enkele korrels na, niet open. In medium 2 waren er wel pollenkorrels die een beginnende
kiembuis gevormd hadden. Medium 3, waarin sucrose vervangen werd door PEG, vertoonde
gelijkaardige resultaten wat betreft de vorming van een beginnende pollenkiembuis, maar
bevatte meer gebarsten pollenkorrels. Medium 4 werd ook getest als basismedium, maar
vertoonde ook geen kieming. Het kiemingspercentage en het percentage opengebarsten
pollenkorrels per medium zijn terug te vinden in
Tabel 3.
Doordat er in alle geteste basismedia geen kieming optrad, werd er gekozen voor het
medium met het kleinste aantal opengebarsten pollenkorrels, dit is een medium op basis van
maltose (medium 2).
Tabel 3 - Kiemingspercentage en percentage opengebarsten pollenkorrels van de verschillende basismedia
Mediumnummer Kiemingspercentage Percentage opengebarsten pollenkorrels
1 0% 85% 2 0% 6% 3 0% 10% 4 0% 75%

34
In medium 5 werden verschillende maltoseconcentraties getest. Bij een maltoseconcentratie
van 5% waren nagenoeg alle pollenkorrels gebarsten. In het medium met 10% maltose
waren slechts enkele pollenkorrels opengebarsten. Een maltoseconcentratie van 15% of
20% zorgde ook niet voor kieming van het pollen, maar er waren meer beginnende
pollenkiembuizen gevormd en minder opengebarsten pollenkorrels bij 20% maltose. De
resultaten van de kiemingstesten met verschillende maltoseconcentraties zijn weergegeven
in Tabel 4.
Voor de verdere kiemingstesten werd gekozen voor een maltoseconcentratie van 20%.
Tabel 4 - Effect van de maltoseconcentratie op het kiemingspercentage en het percentage opengebarsten pollenkorrels
Maltoseconcentratie Kiemingspercentage Percentage opengebarsten pollenkorrels
5% 0% 99% 10% 0% 6% 15% 0% 5% 20% 0% 3%
3.1.3 Effect van verschillende toevoegingen
In medium 6 werd het effect van agar in het medium getest, dit in concentraties gaande van
5% tot en met 20%. Dit medium gaf geen andere resultaten dan een medium zonder agar.
Verhogen of verlagen van de concentratie boor in het basismedium (medium 2 met 20%
maltose) had geen invloed op het kiemen van het pollen. Hetzelfde is waar te nemen bij het
toevoegen van calcium.
Geen enkele gistconcentratie zorgde voor kieming van het pollen. Hetzelfde geldt voor
kinetine, hetgeen getest werd in medium 8. Indolazijnzuur werd toegevoegd in medium 10, in
concentraties van 1,25 ppm en 2,50 ppm, en had geen effect op de kieming van het pollen.
Biotine werd toegevoegd in medium 9 in een concentratie van 1,25 ppm en zorgde voor
geen veranderingen in het kiemingspercentage.
Vrouwelijke bloemen 24 uur laten incuberen in het kiemingsmedium alvorens de pollen
worden aangebracht, vertoont geen verschil in kiemingspercentage in vergelijking met een
medium zonder deze incubatie.
Omdat al de voorgaande elementen afzonderlijk geen positieve effecten hadden op het
kiemingspercentage van het pollen, werden combinaties van de elementen getest. Deze
combinaties werden steeds toegevoegd aan medium 5.
De combinatie van gist en kinetine (medium 11) zorgde niet voor kieming van het pollen.
Hetzelfde werd vastgesteld bij media 12, 13 en 14, waarin andere combinaties van
toevoegingen werd getest.

35
3.2 Zetmeelgehalte
Het zetmeelgehalte van de zes rassen ligt tussen 11,49% bij Precioza en 13,66% bij Bengal.
Dit is weergegeven in Tabel 5.
Tabel 5 - Gemiddeld zetmeelgehalte voor de zes onderzochte rassen
Ras Gemiddeld zetmeelgehalte Standaardafwijking
Bengal 13,66%NS 2,24 Precioza 11,49%NS 2,97 Tosca 13,07%NS 1,64 Noriac 13,09%NS 1,52 Storrs Green 12,81%NS 3,32 Milos 12,72%NS 1,56 NS: niet significant verschillend volgens One-Way ANOVA (p=0,05)
3.3 Suikergehalte
3.3.1 Sucrosegehalte
Uit Tabel 6 blijkt dat de gemiddelde sucrosegehaltes voor de zes onderzochte rassen tussen
0,65% (Milos) en 0,95% (Precioza) liggen.
Tabel 6 - Gemiddeld sucrosegehalte voor de zes onderzochte rassen
Ras Gemiddeld sucrosegehalte Standaardafwijking
Bengal 0,86%NS 0,61% Precioza 0,95%NS 0,51% Tosca 0,91%NS 0,50% Noriac 0,91%NS 0,48% Storrs Green 0,76%NS 0,43% Milos 0,65%NS 0,52% NS: niet significant verschillend volgens One-Way ANOVA (p=0,05)
3.3.2 Glucosegehalte
Het hoogste glucosegehalte, 0,52%, is terug te vinden in het pollen van Bengal. Precioza
heeft met 0,39% glucose het laagste percentage. Dit is weergegeven in Tabel 7.
Tabel 7 - Gemiddeld glucosegehalte voor de zes onderzochte rassen
Ras Gemiddeld glucosegehalte Standaardafwijking
Bengal 0,52%NS 0,28% Precioza 0,39%NS 0,32% Tosca 0,45%NS 0,24% Noriac 0,44%NS 0,30% Storrs Green 0,49%NS 0,31% Milos 0,51%NS 0,26% NS: niet significant verschillend volgens One-Way ANOVA (p=0,05)
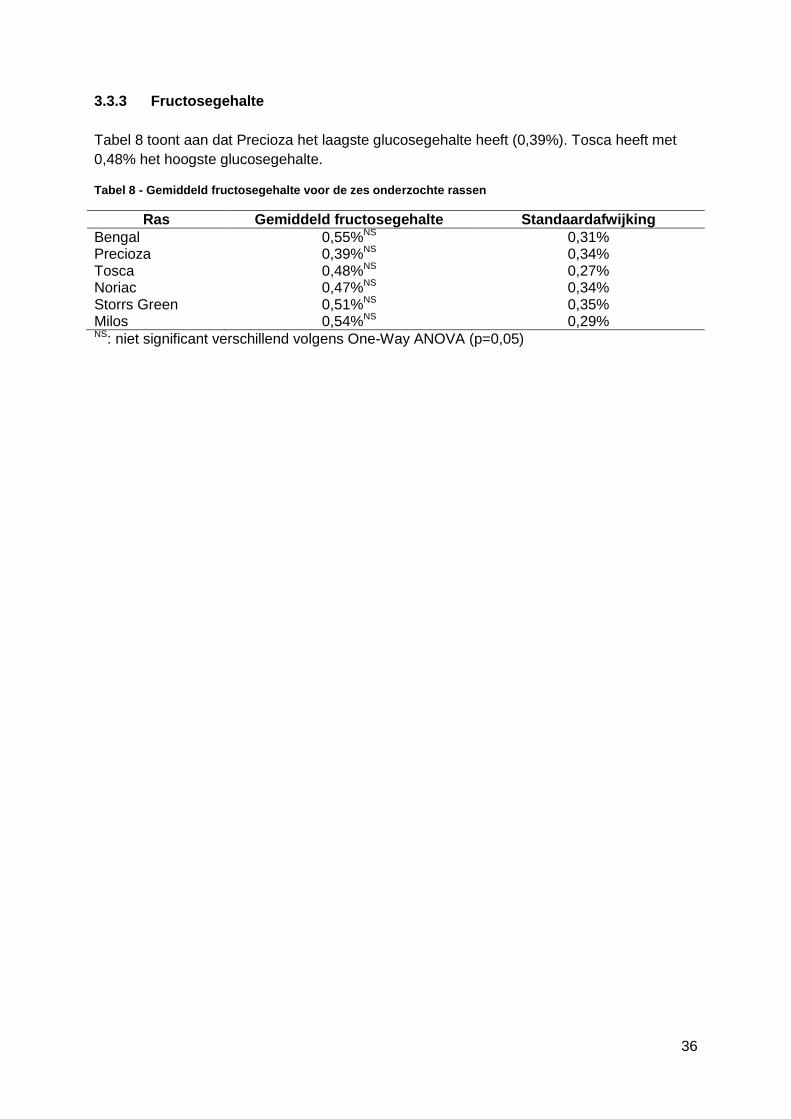
36
3.3.3 Fructosegehalte
Tabel 8 toont aan dat Precioza het laagste glucosegehalte heeft (0,39%). Tosca heeft met
0,48% het hoogste glucosegehalte.
Tabel 8 - Gemiddeld fructosegehalte voor de zes onderzochte rassen
Ras Gemiddeld fructosegehalte Standaardafwijking
Bengal 0,55%NS 0,31% Precioza 0,39%NS 0,34% Tosca 0,48%NS 0,27% Noriac 0,47%NS 0,34% Storrs Green 0,51%NS 0,35% Milos 0,54%NS 0,29% NS: niet significant verschillend volgens One-Way ANOVA (p=0,05)

37
3.4 Pollenvitaliteit na bewaring bij -80°C
Het testen van de pollenvitaliteit na een bewaring in -80°C bleek niet mogelijk, toch niet op
de manier die onder 2.3 werd beschreven. Het pollen barstte telkens open. Naarmate het
pollen meer tijd krijgt om te acclimatiseren, vooraleer het in het medium wordt gebracht,
verloopt het openbarsten trager, het openbarsten is onvermijdelijk. Hierdoor was het niet
mogelijk om vitaliteit te bepalen.
3.5 Pollenvitaliteit na bewaring bij verschillende temperaturen
3.5.1 Vers pollen
Tabel 9 geeft de resultaten weer van de vitaliteitstest van de stalen verzameld op 23 maart
2017. Er is een verschil in vitaliteit tussen de rassen Bengal en Noriac. Bengal heeft, met
een gemiddelde vitaliteit van 70%, het hoogste vitaliteitspercentage. Noriac heeft een
gemiddeld vitaliteitspercentage van 59%. In de vier herhalingen van elk ras zijn er ook vrij
grote verschillen in vitaliteit zoals blijkt uit de grote standaardafwijking
Tabel 9 - Vergelijking vitaliteitspercentage van vers pollen tussen de rassen Bengal en Noriac
Ras Gemiddeld vitaliteitspercentage Standaardafwijking
Bengal 69,75%NS 7,80% Noriac 59,00%NS 4,24%
NS: niet significant verschillend volgens de Levene’s test (p=0,05)
Tabel 10 geeft het resultaat weer van de vitaliteitstest van het pollen (Bengal) verzameld op
27 maart 2017 en levert een heel ander resultaat op dan de eerste vitaliteitstest. Het
gemiddelde vitaliteitspercentage ligt hier beduidend lager en is slechts 42%.
Tabel 10 - Vitaliteitspercentage van vers pollen van het ras Bengal
Ras Gemiddeld vitaliteitspercentage Standaardafwijking
Bengal 42,04% 3,98%
3.5.2 Bewaring op kamertemperatuur
Uit Tabel 11 valt op te maken dat de vitaliteit van het pollen (Bengal) bij bewaring op
kamertemperatuur na twee dagen nog gemiddeld 80% bedraagt. Na vier dagen is de vitaliteit
zeer sterk gedaald en bedraagt deze nog maar 2%.
Tabel 11 - Vitaliteitspercentage van pollen van het ras Bengal bij bewaring op kamertemperatuur
Bewaringsduur Gemiddeld vitaliteitspercentage Standaardafwijking
1 dag 92,96%a 4,49% 2 dagen 79,59%b 7,50% 4 dagen 1,83%c 2,22%
a, b, c: onderling significant verschillend volgens ANOVA analyse (p=0,05)

38
3.5.3 Bewaring in de frigo (6°C)
Tabel 12 toont dat de vitaliteit van het pollen (Bengal) bij bewaring in de frigo (6°C) na vier
dagen nog 29% bedraagt.
Tabel 12 - Vitaliteitspercentage van pollen van het ras Bengal bij bewaring in de frigo (6°C)
Bewaringsduur Gemiddeld vitaliteitspercentage Standaardafwijking
1 dag 69,00%a 14,76% 2 dagen 58,50%b 12,40% 4 dagen 29,00%b 4,69%
a, b: onderling significant verschillend volgens ANOVA analyse (p=0,05)
3.5.4 Bewaring in diepvries (-20°C)
De vitaliteitstest op pollen (Bengal) dat bij -20°C werd bewaard kende dezelfde problemen
als deze beschreven onder 3.4. Doordat de pollen niet intact bleven na aanbrengen in het
kleuringsmedium, kon de vitaliteit niet bepaald worden.

39
3.6 Manuele bestuiving
Een week na de manuele bestuiving van de vrouwelijke bloemen bij Tosca werden alle
courgettes beoordeeld op vruchtgrootte en –gewicht. De gemiddelde gewichten worden
weergegeven in Tabel 13. Uit de tabel blijkt dat enkel bloemen die met vers pollen bestoven
werden, uitgroeien tot vruchten met een voldoende kwaliteit. 9 van de 10 herhalingen, van de
bestuiving met vers pollen, leverde vermarktbare vruchten op. Slechts één vrucht groeide
niet goed uit en bleef een zwartje. Dit is te zien op Figuur 8.
Tabel 13 - Resultaten van de manuele bestuiving bij Tosca met pollen van verschillende bewaarduur
Bloemen bestoven met Gemiddeld gewicht in g Vruchtgrootte
Vers pollen 470,58 Normale vruchten 5 dagen bewaring 43,30 Zwartje*
10 dagen bewaring 24,00 Zwartje* 15 dagen bewaring 28,72 Zwartje*
Onbestoven 19,94 Zwartje*
*Zwartje is de telersomschrijving voor een vrucht die niet voldoet aan de kwaliteitseisen
Figuur 8 - Resultaten van bestuiving bij Tosca met vers pollen

40
Bestuiving met bewaard pollen leidde steeds tot te kleine vruchten, zogenaamde zwartjes, of
puntvruchten die niet voldoen aan de kwaliteitseisen. De resultaten van de bestuiving met
pollen van 5 dagen oud, zijn te zien op Figuur 9.
Figuur 9 - Resultaten van bestuiving bij Tosca met pollen van 5 dagen oud

41
4 Discussie
4.1 Pollenkieming
De kieming van het pollen verliep niet zoals verwacht. Het openbarsten van de pollenkorrels
op een medium met sucrose komt doordat sucrose zorgt voor een verhoging van de interne
sucroseconcentratie, wat zorgt voor een verandering in membraanpermeabiliteit
(Jayaprakash & Sarla, 2001). Bij maltose komen veel minder opengebarsten pollenkorrels
voor. Dit kan komen doordat maltose bestaat uit twee moleculen glucose die aan elkaar
gebonden zijn (sucrose bestaat uit een fructose en een glucose molecule) waardoor de
interne concentratie minder verhoogd wordt en het pollen intact blijft. Het gebruik van PEG
als osmoticum in het kiemingsmedium zorgt ervoor dat de wateropname, nodig voor de
hydratatie van de pollenkorrels, trager verloopt waardoor lekkage van de cel minder optreedt
(Hoekstra, 1995).
Ondanks alle geteste toevoegingen aan het medium slaagden we er niet in een goed
pollenkiemingsmedium te bekomen. Ook media, die volgens de literatuur tot betere
kiemingspercentages moeten leiden, waren niet succesvol bij de geteste courgette-rassen.
4.2 Zetmeel- en suikergehalte
Er bestaat een correlatie tussen het sucrose-, fructose- en glucosegehalte van het pollen. Bij
elk ras is het percentage aan glucose en fructose ongeveer gelijk, maar hoe hoger het
glucose- of fructosegehalte, hoe lager het sucrosegehalte. Dit komt doordat sucrose wordt
omgezet in fructose en glucose (Pacini et al., 2006). Het lage gehalte aan suikers in het
pollen van Cucurbita pepo is ook gemeld door Nepi et al. (2001). Figuur 10 toont een
histogram met de suikergehaltes van 13 verschillende species met gedeeltelijk gehydrateerd
pollen, zoals Cucurbita pepo.
Figuur 10 - Histogram van glucose, fructose, sucrose en totaal suikergehalte in pollenkorrels van 13 species met gedeeltelijk gehydrateerd pollen (Nepi et al., 2001)

42
Voor bewaringsproeven is een hoog gehalte aan sucrose wenselijk omdat het zorgt voor een
betere bewaring van het pollen en het beschermt tegen uitdroging (Pacini et al., 2006). Hoge
sucrosegehaltes verhinderen ook kristallisatie van de waterige polleninhoud wanneer het
pollen wordt ingevroren. Lage concentraties bieden blijkbaar weinig bescherming.
Het zetmeelgehalte en het suikergehalte van het pollen vertonen ook een correlatie. Hierbij
geldt dat, hoe hoger het zetmeelgehalte van het pollen is, hoe lager het sucrosegehalte. Dit
was te verwachten want tijdens de ontwikkeling van het pollen wordt het zetmeel
gehydrolyseerd tot sucrose, fructose en glucose (Pacini et al., 2006). Een hoog
zetmeelgehalte, met daarbij gepaard een laag gehalte aan mono- en disacchariden, wordt
bevestigd door Nepi et al. (2001).
4.3 Pollenvitaliteit na bewaring bij -80°C
Het pollen van Cucurbita pepo is gedeeltelijk gehydrateerd, waardoor het pollen nog een
behoorlijke hoeveelheid water bevat bij het oogsten, tot 35% water op het einde van de
anthese (Nepi et al., 2001). Bij het invriezen van het pollen, vormt het aanwezige water
ijskristallen, die meer plaats innemen dan water, waardoor barsten kunnen ontstaan in de
celmembraan (Hoekstra, 1995). Wanneer het pollen, na een bewaring in de diepvries, terug
ontdooit, zorgen de barsten ervoor dat het cytoplasma uit de pollenkorrel stroomt bij
aanbrengen in een medium.
Een andere reden voor het mislukken van de bewaring van het pollen, is het feit dat het
pollen niet op voorhand gedroogd is. Voor bewaring is het belangrijk het watergehalte in het
pollen zo snel als mogelijk te verminderen, dit gebeurt het best op kamertemperatuur. Een
watergehalte van 5% tot 8% is voldoende voor bewaring in de diepvries of bij -80°C. Hogere
watergehaltes zorgen voor de vorming van ijskristallen bij bewaring bij -20° of lager. Een
oplossing voor dit probleem is waarschijnlijk de bewaring van het pollen in organische
solventen (Hoekstra, 1995).

43
4.4 Pollenvitaliteit na bewaring bij verschillende temperaturen
4.4.1 Vers pollen
4.4.1.1 Vergelijking vitaliteit van het pollen van de rassen Bengal en Noriac
Het ras Bengal bevat een lagere sucroseconcentratie dan Noriac, waardoor volgens (Pacini
et al., 2006) Noriac beter bestand zou moeten zijn tegen uitdroging en langer bewaart. Toch
heeft Noriac een lager vitaliteitspercentage dan Bengal, wat kan wijzen op sneller uitdrogen
van het pollen van Noriac.
4.4.1.2 Testen van de vitaliteit van vers pollen van het ras Bengal
Bij het vergelijken van het vitaliteitspercentage van het pollen van het eerste en het tweede
experiment, is er een groot verschil waar te nemen. In het eerste experiment bedraagt de
vitaliteit van Bengal gemiddeld 70%, maar in de tweede test is dit slechts 42%. Dit is te
verklaren door het feit dat voor het eerste experiment pollen gebruikt werd van planten die op
de faculteit zelf stonden. De tijd tussen het oogsten van het pollen en het aanbrengen in de
suikeroplossing was maximaal 10 minuten. Voor het tweede experiment is pollen gebruikt
van planten die op het ILVO stonden. Hierdoor is er ongeveer 1 uur verlopen tussen het
oogsten van het pollen en het aanbrengen in de suikeroplossing. Het pollen was al meer
gedehydrateerd op het moment van testen, wat het afwijkende vitaliteitspercentage kan
verklaren.
4.4.2 Bewaring op kamertemperatuur
Het vitaliteitspercentage van het pollen is na twee dagen bewaring nog steeds zeer hoog,
zo’n 80%. Doordat de vitaliteit van het vers pollen van de stalen voor de bewaring (afkomstig
van de serres op de UGent) niet nagegaan is, kan het niet vergeleken worden met de
vitaliteit van vers pollen. De vitaliteit bij bewaring op kamertemperatuur valt echter zeer snel
terug, na vier dagen is de vitaliteit zo goed als 0%. Bewaring op kamertemperatuur van het
pollen kan dus een zeer tijdelijke oplossing zijn, wanneer men het pollen binnen de twee
dagen kan aanbrengen op de vrouwelijke bloemen. Om een langere periode te overbruggen,
moet een andere bewaarmethode gekozen worden.
4.4.3 Bewaring in de frigo (6°C)
Bij de bewaring in de frigo bij 6°C, blijft de vitaliteit van het pollen gedurende 4 dagen goed.
Daarna is er een snelle terugval. De vitaliteit blijft beter behouden dan op kamertemperatuur.
Dit kan komen door het feit dat het pollen minder snel uitdroogt in de frigo waardoor de
vitaliteit hoger blijft.

44
4.4.4 Bewaring in diepvries (-20°C)
De bewaring bij -20°C is even problematisch als bij -80°C. Er worden eveneens ijskristallen
in het pollen gevormd, die ervoor zorgen dat bij ontdooien het cytoplasma uit de cel vloeit.
Bewaren van courgette pollen bij negatieve temperaturen is dus waarschijnlijk niet mogelijk.
Eventueel zouden nog licht negatieve temperaturen zoals -1°C kunnen getest worden. De
keuze voor -20°C was echter ingegeven door het feit dat de bedrijven, deze temperatuur wel
gemakkelijk kunnen toepassen.
4.5 Manuele bestuiving
Uit de bestuivingsproef kan besloten worden dat enkel vers pollen gebruikt kan worden voor
de manuele bestuiving van vrouwelijke bloemen. Pollen dat enkele dagen bewaard werd in
de frigo bij 6°C, levert geen goede vruchten op. Doordat de vitaliteit van het pollen, dat
gebruikt is voor de manuele bestuiving (pollen van in de serres op het ILVO), niet nagegaan
is, kan er geen link gelegd worden tussen het vitaliteitspercentage van het pollen en de
resultaten van de manuele bestuiving.
Door deze resultaten is er geen oplossing gevonden voor het stuifmeeltekort tijdens de
problematische weken in de courgetteteelt. Om het stuifmeeltekort op te vangen, moet er via
de veredeling gekeken worden of er rassen zijn die langer voldoende mannelijke bloemen
produceren, zodat deze rassen eventueel tussen de courgetteplanten geplaatst kunnen
worden.

45
5 Besluit
Dit onderzoek slaagde er niet in om een geschikt kiemingsmedium voor courgettepollen te
ontwikkelen. Toch werden interessante inzichten verworven met betrekking tot de
verschillende elementen die nodig zijn in dergelijk medium. In een kiemingsmedium voor
pollen van Cucurbita pepo is sucrose geen goed osmoticum. Er wordt best gekozen voor
maltose of PEG. De meerwaarde van verschillende toevoegingen kon niet worden bevestigd.
De zetmeel- en suikergehaltes in het pollen vertonen een duidelijke correlatie waarbij een
laag zetmeelgehalte zorgt voor een hoger suikergehalte. Glucose en fructose komen steeds
in het pollen in ongeveer dezelfde concentratie voor, het sucrosegehalte is lager naarmate
het fructose en glucosegehalte hoger is. Hoewel een ras met een hoger sucrosegehalte
beter bestand zou moeten zijn tegen uitdroging, met dus een hoger vitaliteitspercentage als
gevolg, was dit niet het geval in deze experimenten.
Om de problematische weken tijdens de teelt te overbruggen, is er geen mogelijkheid om het
pollen gedurende enkele dagen te bewaren, bij welke temperatuur ook. Bewaring bij -20°C of
-80°C zorgt voor de vorming van ijskristallen in het pollen. Hierdoor is het celmembraan te
veel beschadigd en zal bij ontdooien nadien, de celinhoud uit de pollenkorrel vloeien. Licht
negatieve temperaturen werden niet getest. Bewaring in de frigo (6°C) en op
kamertemperatuur leverde wel positieve resultaten op. Op kamertemperatuur kan het pollen
twee dagen bewaard worden, zonder een te groot verlies aan vitaliteit. Na 4 dagen bewaring
is er 0% vitaliteit. Bij bewaring in de frigo, blijft de vitaliteit gedurende enkele dagen hoog,
maar neemt nadien snel af.
Of een behoorlijk vitaliteitspercentage garant staat voor een goede bevruchting, kon niet
nagegaan worden doordat de vitaliteit van het gebruikte pollen voor de manuele bestuiving,
niet getest is. Uit de manuele bestuiving is wel gebleken dat enkel bloemen die met vers
pollen bestoven zijn, uitgroeien tot vermarktbare vruchten. Het pollen dient dan wel best van
dichtbij te komen, want anders daalt de vitaliteit tijdens het transport te snel. Indien bewaard
pollen wordt gebruikt, groeien de vruchten niet goed uit en ontstaan puntvruchten of
zwartjes.
Aangezien de bewaringsproeven geen goede resultaten voor de telers opleverden, zal er
gezocht moeten worden naar een andere manier om gedurende het hele groeiseizoen
voldoende pollen te voorzien. Zo kan de teler ervoor kiezen om andere rassen, die meer
en/of langer stuifmeel produceren, als tussenras aan te planten of om een deel van zijn
gewas later op te planten zodat de piek in stuifmeelproductie van het ene deel, het tekort
opvangt in het andere deel.

46
6 Bibliografie
Abdul-Baki, A. (1992). Determination of pollen viability in tomatoes. In J. Amer. Soc. Hort.
Sci., 117(3) (pp. 473 - 476).
Baker, H., & Baker, I. (1979). Starch in Angiosperm pollen grains and its evolutionary
significance. In Amer. J. Bot., 66(5) (pp. 591 - 600).
Bertin, R., & Sholes, O. (1993). Weather, pollination and the phenology of Geranium
maculatum. In The American Midland Naturalist, Vol. 129, No. 1 (pp. 52 - 66).
Brewbaker, J., & Kwack, B. (1963). The essential role of calcium ion in pollen germination
and pollen tube growth. In American Journal of Botany, Vol. 50, No. 9 (pp. 859-865).
Campos, M., Bogdanov, S., Bicudo de Almeida-Muradian, L., Szczesna, T., Mancebo, Y.,
Frigerio, C., & Ferreira, F. (2008). Pollen composition and standardisation of
analytical methods. In Journal of Apicultural Research, 47:2 (pp. 154 - 161).
Carvajal, F., Garrido, D., Jamilena, M., & Rosales, R. (2013). Cloning and characterisation of
a putative pollen-specific polygalacturonase gene (CpPG1) differentially regulated
during pollen development in zucchini (Cucurbita pepo L.). In Plant Biology.
Ciampolini, F., Nepi, M., & Pacini, E. (1993). Tapetum development in Cucurbita pepo
(Cucurbitaceae). In V. 7. Plant Syst. Evol..
das Graças Vidal, M., de Jong, D., Morse, R., & Wien, H. (2006). Nectar and pollen
production in pumpkin (Cucurbita pepo L.). In Rev. bras. Bot., vol.29, no.2.
Decker, D. (1988). Evolution and systematics of Cucurbita pepo (Cucurbitaceae). In
Economic Botany, Vol. 42, No. 1 (pp. 4 - 15).
Duarte-Silva, E., Rodrigues, L., & Mariath, J. (2011). Contradictory results in pollen viability
determination of Valeriana scandens L. In Geneconserve 40 (pp. 234-242).
Eady, C., Lindsey, K., & Twell, D. (1994). Differential activation and conserved vegetative
cellspecific activity of a late pollen promoter in species with bicellular and tricellular
pollen. In The Plant Journal, 5(4) (pp. 543 - 550).
Edlund, A., Swanson, R., & Preuss, D. (2004). Pollen and stigma structure and function: the
role of diversity in pollination. In The Plant Cell, Vol. 16 (pp. 84-97).
Erdtman, G. (1986). Pollen morphology and plant taxonomy.
Esquinas-Alcazar, J., & Gulick, P. (1983). Genetic resources of Cucurbitaceae.
Gay, G., Kerhoas, C., & Dumas, C. (1987). Quality of a stress-sensitive Cucurbita pepo L.
pollen. In Planta, 171 (pp. 82-87).
Heslop-Harrison, J., Heslop-Harrison, Y., & Shivanna, K. (1983). The evaluation of pollen
quality, and a further appraisal of the fluorochromatic (FCR) test procedure. In Theor
Appl Genet (1984), 67 (pp. 367-375).

47
Hoekstra, F. (1995). Collecting pollen for genetic resources conservation. In Collecting plant
genetic diversity: technical guidelines (pp. 527 - 550 ).
Holman, R., & Brubaker, F. (1926). On the longevity of pollen. In Publ. Bot., 13 (pp. 179 -
204).
Howlett, B., Knox, R., & Heslop-Harrison , J. (1973). Pollen-wall proteins: release of the
allergen antigen E from intine and exine sites in pollen grains of Ragweed and
Cosmos. In J. Cell. Sci., 13 (pp. 603 - 619).
Jayaprakash, P., & Sarla, N. (2001). Development of an improved medium for germination of
Cajanus cajan (L.) Millsp. pollen in vitro. In Journal of Experimental Botany, Vol. 52,
No. 357 (pp. 851-855).
Jeffrey, C. (1964). A note on pollen morphology in Cucurbitaceae. In Kew Bulletin, Vol. 17,
No. 3 (pp. 473 - 477).
Jeffrey, C. (1980). A review of the Cucurbitaceae. In Botanical journal of the Linnean society,
81 (pp. 233 - 247).
Jóhannsson, M., & Stephenson, A. (1998). Effects of temperature during microsporogenesis
on pollen performance in Cucurbita pepo L. (Cucurbitaceae). In International Journal
of Plant Sciences, Vol. 159, No. 4 (pp. 616-626).
Johri, B., & Vasil, I. (1961). Physiology of pollen. In Botanical Review, Vol. 27, No. 3 (pp. 325
- 381).
Jones, K., & Senft, J. (1985). An improved method to determine cell viability by simultaneous
staining with fluorescein diacetate - propidium iodide. In The Journal of
Histochemistry and Cytochemistry (pp. 77 - 79).
Kato, Y. (1955). Responses of plant cells to gibberellin. In Botanical Gazette 117, no. 1 (pp.
16 - 24).
Kelly, J., Rasch, A., & Kalisz, S. (2002). A method to estimate pollen viability from pollen size
variation. In American Journal of Botany, 89(6) (pp. 1021–1023).
Khan, S., & Perveen, A. (2010). In vitro pollen germination capacity of Citrullus Lanatus L.
(Cucurbitaceae). In Pak. J. Bot., 42(2) (pp. 681-684).
Kiełkowska, A., & Havey, M. (2012). In vitro flowering and production of viable pollen of
cucumber. In Plant Cell Tiss Organ Cult, 109 (pp. 73–82).
Kumari, A., Komal, R., Rajesh, R., & Pandey, A. (2009). In vitro pollen germination, pollen
tube growth and pollen viability in Trichosanthes dioica Roxb. (Cucurbitaceae). In The
International Journal of Plant Reproductive Biology, 1(2) (pp. 147-151).
Lau, T.-C., & Stephenson, A. (1993). Effects of soil nitrogen on pollen production, pollen
grain size, and pollen performance in Cucurbita pepo (Cucurbitaceae). In American
Journal of Botany, Vol. 80, No. 7 (pp. 763-768).

48
Lira, R., Alvarado, J., & Ayala‐Nieto, M. (1998). Pollen morphology in Sicydium
(Cucurbitaceae, Zanonioideae). In Grana 37 (pp. 215 - 221).
Mascarenhas, J. (1993). Molecular mechanisms of pollen tube growth and differentiation. In
The Plant Cell, Vol. 5 (pp. 1303-1314).
Nepi, M., & Pacini, E. (1993). Pollination, pollen viability and pistil receptivity in Cucurbita
pepo. In Annuals of Botany, 72 (pp. 527 - 536).
Nepi, M., & Pacini, E. (2001). Effect of pistil ageon pollen tube growth, fruit development and
seed set in Cucurbita pepo L. In Acta Societatis Botanicorum Poloniae, Vol.70, No.3
(pp. 165-172).
Nepi, M., Cresti, L., Guarnieri, M., & Pacini, E. (2009). Effect of relative humidity on water
content, viability and and carbohydrate profile of Petunia hybrida and Cucurbita pepo
pollen. In Plant Syst. Evol. (2010), 284 (pp. 57-64).
Nepi, M., Franchi, G., & Pacini, E. (2001). Pollen hydration status at dispersal:
cytophysiological features and strategies. In Protoplasma, 216 (pp. 171-180).
Nepi, M., Guarnieri, M., & Pacini, E. (2001). Nectar secretion, reabsorption and sugar
composition in male and female flowers of Cucurbita pepo. In Int. J. Plant Sci., 162(2)
(pp. 353 – 358).
Pacini, E., & Hesse, M. (2005). Pollenkit – its composition, forms and functions. In Flora 200
(pp. 399 - 415).
Pacini, E., Guarnieri, M., & Nepi, M. (2006). Pollen carbohydrates and water content during
development, presentation, and dispersal: a short review. In Protoplasma, 228 (pp.
73 - 77).
Perveen, A., & Ali, S. (2011). Pollen germination capacityand maintenance of pollen in
Praecitrullus Fistulosus (Stocks) Pangola (Cucurbitaceae). In Pak. J. Bot., 43(1) (pp.
47-50).
Perveen, A., & Qaiser, M. (2008). Pollen flora of Pakistan - LVI Cucurbitaceae. In Pak. J.
Bot., 40(1) (pp. 9 - 16).
Potts, S., Biesmeijer, J., Kremen, C., Neumann, P., Schweiger, O., & Kunin, W. (2010).
Global pollinator declines: trends, impacts and drivers. In Trends in Ecology and
Evolution, Vol.25, No.6 (pp. 345 - 353).
Reddy, K., & Kakani, V. (2007). Screening Capsicum species of different origins for high
temperature tolerance by in vitro pollen germination and pollen tube length. In
Scientia Horticulturae, 112 (pp. 130–135).
Roulston, T., Cane, J., & Buchmann, S. (2000). What governs protein content of pollen:
pollinator preferences, pollen-pistil interactions, or phylogeny? In Ecological
Monographs, Vol. 70, No. 4 (pp. 617 - 643).

49
Schaefer, H., & Renner, S. (2011). Phylogenetic relationships in the order Cucurbitales and a
new classification of the gourd family (Cucurbitaceae). In Taxon 60 (1) (pp. 122 -
138).
Shi, D.-Q., & Yang, W.-C. (2010). Pollen germination and tube growth. In Plant
Developmental Biology - Biotechnological Perspectives, Vol. 1.
Taylor, L., & Hepler, P. (1997). Pollen germination and tube growth. In Annu. Rev. Plant
Physiol. Plant Mol. Biol., 48 (pp. 461-491).
Van Der Ham, R., & Van Heuven, B. (2003). A new pollen type in Old World Cucurbitaceae.
In Grana 42 (pp. 88 - 90).
Van Der Ham, R., Mennes, C., & Van Heuven, B. (2010). Fevilleoideae pollen
(Cucurbitaceae): a study in striate ornamentation. In Grana 49 (pp. 157 – 169).
Van Labeke, M. (2016). Ontwikkeling: kieming van zaden. In Groei en
ontwikkelingsbeheersing bij planten (pp. 8-11).
Vasil, I. (1960). Studies on pollen germination of certain Cucurbitaceae. In American Journal
of Botany, Vol. 47, No. 4 (pp. 239-247).
Vizintin, L., & Bohanec, B. (2004). In vitro manipulation of cucumber (Cucumis sativus L.)
pollen and microspores: Isolation procedures, viability, tests, germination, maturation.
In Acta Biologica Cracoviensia Series Botanica, 46 (pp. 177–183).
Winsor, J., & Stephenson, A. (1995). Demographics of pollen tube growth in Cucurbita pepo.
In Can. J. Bot., 73 (pp. 583 - 589 ).
Ylstra, B., Touraev, A., Benito Moreno, R., Stoger, E., van Tunen, A., Vicente, O., . . .
Heberle-Bors, E. (1992). Flavonols stimulate development, germination, and tube
growth of Tobacco pollen. In Plant Physiol., 100 (pp. 902-907).

50
7 Bijlagen
7.1 Zetmeelanalyse
Voor de analyse van het zetmeel- en suikergehalte van het pollen, werd een protocol
gebruikt van het labo Plantaardige Productie van de UGent.
Voordat begonnen werd met de extracties, werden alle stalen gewogen. De gewichten
varieerden van 22,1 mg tot 190 mg.
De zetmeelconcentratie werd afgeleid uit de mate van absorbantie van het staal, gemeten
met de spectrofotometer. Een aantal reagentia waren nodig voor het meten van de
absorbantie
1. TRA-buffer 0.75 mol/l (100ml)
- 14 g Triëthanolamine.hydrochloride
- 250 mg MgSO4.7H2O)
- Beide worden samen opgelost in 70 ml H2O
- Aanpassen van de pH met NaOH (50%) tot een pH 7,6
- Het geheel wordt bijgevuld met H2O tot 100 ml in kolfje
2. NADP+ (nicotinamide-adenine dinucleotide fosfaat)
- 100 mg NADP-NA2 oplossen in 10 ml H2O
3. ATP (adenosine-5-trifosfaat)
- 500 mg ATP-Na2H2
- 500 mg NaHCO3
- Beide oplossen in 10 ml H2O
4. Glucosestandaard
- 50 mg glucose oplossen in 50 ml water
Na het aanmaken van de oplossingen, wordt de absorbantie, in een cuvet van 4,5 ml
gemeten.
- 1920 µl verdund staal : 100µl staal + 1820µl H2O
Voor standaard: 100µl glucosestandaard + 1820µl H2O
Voor blanco: 1920 µl H2O
- 1200µl reagentiamengsel (1000µl TRA-buffer, 100µl NADP+, 100µl ATP)

51
Na 3 minuten wachten werd de absorbantie (A1) gemeten met de spectrofotometer bij een
golflengte van 340 nm. Daarna werd aan de cuvet een 20 µl enzymmengsel van hexokinase
en glucose-6-fosfaatdehydrogenase toegevoegd. Na 15 minuten wachten werd de
absorbantie (A2) opnieuw gemeten bij 340 nm. De zetmeelconcentratie kon vervolgens
bepaald worden uit onderstaande formule
V. MW c = . ∆A D-glucose (mg D-glucose/ml oplossing)
Ɛ.d.v.1000
V = 3,140 ml (finaal volume van het cuvet)
MW = MW glucose – MW water = 180,16 g/mol – 18 g/mol = 162,16 g/mol
Ɛ = 6,3 l mmol-1 cm-1 (extinctiecoëfficiënt van NADPH bij 340nm)
D = 1 cm (afstand van het lichtpad)
V = 0,1 ml (staalvolume)
∆A staal = ∆A2,staal D-glucose - ∆A1,staal D-glucose
∆A blanco= ∆A2,blanco D-glucose - ∆A1,blanco D-glucose
∆A D-glucose= ∆A staal - ∆A blanco

52
7.2 Pollenvitaliteit
Voor de analyse van de pollenvitaliteit van het pollen, werd van elke well meerdere foto’s
genomen die tot 1 geheel werden herleid. Nadien werden de foto’s geanalyseerd door een
programma, geschreven door Ir. Simon Craeye. Op deze manier werd de vitaliteit bij alle
stalen op dezelfde manier berekend.
In een eerste fase werden alle pollenkorrels geteld:
run("Duplicate...", "title=A.jpg");
selectWindow("A.jpg");
run("8-bit");
setAutoThreshold("Default");
//run("Threshold...");
setThreshold(0, 90);
setOption("BlackBackground", false);
run("Convert to Mask");
run("Watershed");
run("Analyze Particles...", "size=4000-14000 circularity=0.20-1.00 show=Outlines display
summarize");
Nadien werden alle vitale pollenkorrels geteld:
run("Duplicate...", "title=A.jpg");
run("Conversions...", "scale");
run("16-bit");
run("Duplicate...", "title=B.jpg");
setAutoThreshold("Default");
//run("Threshold...");
setThreshold(30, 255);
run("Convert to Mask");
run("Watershed");
run("Set Measurements...", "area mean min redirect=A.jpg decimal=3");
run("Analyze Particles...", "size=4000-14000 circularity=0.20-1.00 show=Outlines display
summarize");