Embed Size (px)
Citation preview

LETTERdoi:10.1038/nature12732
Structural basis for recognition of synaptic vesicleprotein 2C by botulinum neurotoxin ARoger M. Benoit1, Daniel Frey1*, Manuel Hilbert1*, Josta T. Kevenaar2*, Mara M. Wieser1, Christian U. Stirnimann3,David McMillan4, Tom Ceska4, Florence Lebon5, Rolf Jaussi1, Michel O. Steinmetz1, Gebhard F. X. Schertler1,6,Casper C. Hoogenraad2, Guido Capitani1 & Richard A. Kammerer1
Botulinum neurotoxin A (BoNT/A) belongs to the most dangerousclass of bioweapons1. Despite this, BoNT/A is used to treat a widerange of common medical conditions such as migraines and a varietyof ocular motility and movement disorders2. BoNT/A is probablybest known for its use as an antiwrinkle agent in cosmetic applica-tions (including Botox and Dysport)3. BoNT/A application causeslong-lasting flaccid paralysis of muscles through inhibiting the releaseof the neurotransmitter acetylcholine by cleaving synaptosomal-associated protein 25 (SNAP-25) within presynaptic nerve terminals4.Two types of BoNT/A receptor have been identified, both of whichare required for BoNT/A toxicity and are therefore likely to coope-rate with each other5: gangliosides and members of the synapticvesicle glycoprotein 2 (SV2) family, which are putative transporterproteins that are predicted to have 12 transmembrane domains,associate with the receptor-binding domain of the toxin5. Recently,fibroblast growth factor receptor 3 (FGFR3) has also been reportedto be a potential BoNT/A receptor6. In SV2 proteins, the BoNT/A-binding site has been mapped to the luminal domain7, but the mole-cular details of the interaction between BoNT/A and SV2 are unknown.Here we determined the high-resolution crystal structure of theBoNT/A receptor-binding domain (BoNT/A-RBD) in complex withthe SV2C luminal domain (SV2C-LD). SV2C-LD consists of a right-handed, quadrilateral b-helix that associates with BoNT/A-RBDmainly through backbone-to-backbone interactions at open b-strandedges, in a manner that resembles the inter-strand interactions inamyloid structures. Competition experiments identified a peptidethat inhibits the formation of the complex. Our findings provide astrong platform for the development of novel antitoxin agents andfor the rational design of BoNT/A variants with improved thera-peutic properties.
The overall structure of BoNT/A-RBD in complex with SV2C-LD isshown in Fig. 1, and the data collection and refinement statistics arelisted in Extended Data Table 1. The convex interface formed by theamino- and carboxy-terminal BoNT/A-RB subdomains interacts withSV2C-LD. SV2C-LD forms a right-handed, quadrilateral b-helix, afold that is characteristic of pentapeptide-repeat proteins8 (ExtendedData Fig. 1). Based on a search of the Dali server9, the proteins thatare most structurally similar to SV2C-LD in the Protein Data Bank(PDB)10 are MfpA, a pentapeptide-repeat protein from Mycobacteriumtuberculosis11, and AlbG, an McbG-like protein from Xanthomonasalbilineans12.
It has been noted that the molecular architecture of b-helices issimilar to the structure of amyloid fibrils13. Like in amyloid fibrils, theregular b-sheet edges of b-helices are in a conformation that providesthe possibility of interacting with other b-strands and aggregating intohigher-order structures. To avoid the formation of such aggregates, theends of b-helices are typically capped by other secondary-structure
elements14,15. No additional secondary-structure elements are seenon the N-terminal side of SV2C-LD, but the first N-terminal turnof the SV2C-LD b-helix contains more charged amino acids in theinward facing positions than do the other coils. Such strategicallyplaced charges are known to prevent aggregation of single-sheet proteinsand b-propellers15. At its C terminus, SV2C-LD contains a short 310-helix (Extended Data Fig. 1), but this helix precedes the two b-strandsthat interact with BoNT/A-RBD. The interaction between the openb-strand of SV2C-LD and the b-strand edge of BoNT/A-RBD is domi-nated by backbone–backbone hydrogen bonds, resembling the inter-strand interactions in amyloid structures16,17. Our findings thereforeindicate that Clostridium botulinum has exploited a weakness in thestructure of SV2 proteins to transport BoNT/A into host cells.
*These authors contributed equally to this work.
1Laboratory of Biomolecular Research, Paul Scherrer Institut, CH-5232 Villigen PSI, Switzerland. 2Cell Biology, Faculty of Science, Utrecht University, 3584 CH Utrecht, The Netherlands. 3Swiss LightSource, Paul Scherrer Institut, CH-5232 Villigen PSI, Switzerland. 4UCB Celltech, UCB Pharma, UCB NewMedicines, Slough SL1 4EN, UK. 5UCB Pharma, UCB NewMedicines, B-1420 Braine-L’Alleud,Belgium. 6Department of Biology, ETH Zurich, CH-8093 Zurich, Switzerland.
ca
db
R115R115
F563F563F562F562 F557F557
C560C560
S1142S1142 M1144M1144
Y1149Y1149 T1145T1145
T1146T1146
R1156R1156
T1146T1146
T1145T1145
Y1149Y1149M114M114
S1142S1142
C560C560
N559N559F557F557
F562F562F563F563
C N
N
C
(D574)(D574)
(D574(D574)
(D456)(D456)
(D456)(D456)
ca
db
R1156
F563F562 F557
C560
S1142 M1144
Y1149 T1145
T1146
R1156
T1146
T1145
Y1149M1144
S1142
C560
N559F557
F562F563
C N
N
C
(D574)
(D574)
(D456)
(D456)
Figure 1 | The BoNT/A-RBD–SV2C-LD complex. a, Cartoon representationof the structure of BoNT/A-RBD (green) in complex with SV2C-LD (blue).Dotted lines indicate flexible regions of SV2C-LD that were not visible in thestructure. The terminal residues of the SV2C-LD expression construct areindicated. b, Close-up view of the interaction site (boxed area from a).Backbone–backbone and backbone–side chain hydrogen bonds are indicatedby dashed orange lines. Side chain–side chain interactions are shown as dashedpurple lines. c, The complex structure from a different view. d, Close-up ofthe interaction site (boxed region from c). b, d, Carbon is shown in light blue forSV2C-LD and green for BoNT/A-RBD. In both structures, dark blue denotesnitrogen, red denotes oxygen and yellow denotes sulphur.
0 0 M O N T H 2 0 1 3 | V O L 0 0 0 | N A T U R E | 1
Macmillan Publishers Limited. All rights reserved©2013

BoNT/A has been shown to bind to neurons not only through SV2proteins7, or potentially FGFR3 (ref. 6), but also in conjunction withgangliosides as co-receptors18. A superimposition of our BoNT/A-RBD–SV2C-LD complex crystal structure with the previously reported full-length BoNT/A structure19 and the BoNT/A-RBD–GT1b structure18
shows that the binding site for gangliosides is located in the C-termi-nal BoNT/A-RB subdomain almost opposite to the SV2C-binding site(Fig. 2). The two binding regions are separated by a distance of approxi-mately 40 A, illustrating that both receptors can simultaneously interactwith the toxin. This finding might have important functional implica-tions for the translocation of BoNT/A across the synaptic vesicle mem-brane and for subsequent cell intoxication, which are mechanisms ofmajor importance that are still poorly understood. Recent studies usingBoNT/B and BoNT/E have demonstrated that the low pH in the endo-somal lumen and binding to the ganglioside GT1b are both requiredfor the toxins to transform into oligomeric, hydrophobic membrane-associated channels20,21. Although the mechanism of channel formationhas not yet been studied at such detail for BoNT/A, additional bindingto SV2 proteins might be a prerequisite for toxin translocation. Alter-natively, binding to SV2 proteins might prevent the translocation ofBoNT/A by hindering the formation of oligomeric transmembranechannels. Such inhibition might be relieved by acidification in the endo-somal lumen and result both in the dissociation of the SV2–BoNT/Acomplex and the known inhibition of BoNT/A translocation by theRBD at neutral pH22. One promising candidate for a pH-sensing resi-due in SV2C-LD is H564, which is present at the interface of the complex.Consistent with this hypothesis, we observed an approximately five-fold decrease in the dissociation constant (Kd) of the complex between
pH 7.5 and pH 5 in fluorescence anisotropy experiments (ExtendedData Fig. 2).
Comparison of BoNT/A-RBD structures in the free state and SV2C-LD-bound state reveals no significant conformational changes in theSV2C-LD-interacting elements of BoNT/A-RBD on complex forma-tion. Ca superimposition of the BoNT/A-RBD23 onto the BoNT/A-RBD–SV2C-LD complex results in a root mean squared deviation(r.m.s.d.) of 0.54 A over 412 residues. At the interface, 15 SV2C-LDresidues and 19 BoNT/A-RBD residues are partially or fully buried,resulting in a contact area of approximately 596 A2. Several of theseburied residues are engaged in backbone–backbone hydrogen bondsbetween interacting b-strands (Extended Data Table 2). One prom-inent feature seen in the structure is the accumulation of positivelycharged surface residues at the BoNT/A-RBD interface, whereas thebinding surface of SV2C-LD is slightly negatively charged (Fig. 3). Theresidues are not engaged in inter-domain salt bridges; therefore, rela-tively few specific side-chain interactions are observed in the complexstructure. Among these, the interaction between R1156 of BoNT/A-RBD and F563 of SV2C-LD is of particular interest, as it is a cation–p-stacking interaction. The hydrogen bond between SV2C-LD N559 andBoNT/A-RBD Y1149 extends the interaction to the more distantb-strand of the toxin. Hydrogen bonds between SV2C-LD N559 andBoNT/A-RBD T1145, as well as between the backbone oxygen andnitrogen of SV2C-LD F557 and BoNT/A-RBD T1146, stabilize thecomplex downstream of the C-terminal boundary of the directly inter-acting toxin b-strand. R1294 is a residue that prominently contributesto the positive charge of the BoNT/A-RBD surface. Together, theseresidues seem to be important for shape and charge complementarityand consequently for binding specificity.
The importance of these prominent side-chain interactions wastested by site-directed mutagenesis (Extended Data Fig. 3). Mutationof the two BoNT/A-RBD threonines T1145 and T1146 completelyabrogated binding to SV2C-LD in pull-down assays, and mutationof R1294 or R1156 in BoNT/A-RBD significantly reduced this binding(Extended Data Fig. 3). Likewise, substitution of the phenylalanine atposition 563 of SV2C-LD with alanine strongly reduced the binding to
N565
GT1b
Lumen
Cytoplasm
D476
Y577Y577Q462Q462
Y577Q462
Figure 2 | Proposed model for simultaneous ganglioside and SV2C bindingby BoNT/A. Superimposition of full-length BoNT/A19, the BoNT/A-RBD–GT1b ganglioside complex18 and the BoNT/A-RBD–SV2C-LD complex. Theganglioside (white and atom colours; labelled in the figure) and SV2C-LD(blue) bind to opposite sides of the C-terminal BoNT/A-RB subdomain(dark green). The N-terminal BoNT/A-RB subdomain (pale green), thetranslocation domain (including the belt (red)) and the protease domain(yellow) point away from the binding site. The SV2C transmembrane portion(light grey) was modelled. The position of the plasma membrane isproposed. Linker regions are shown as black dotted lines. Linker boundariesare indicated.
BoNT/ABoNT/A
SV2CSV2C
R1294R1294
S1142S1142
Y1149Y1149
M1144M1144
T1146T1146
T1145T1145R1156R1156
C560C560
F557F557
F563F563
S519S519
F562F562
D539D539
N559N559
90º90º
ComplexComplex
90º90º
BoNT/A
SV2C
R1294
S1142
Y1149
M1144
T1146
T1145R1156
C560
F557
F563
S519
F562
D539
N559
90º
Complex
90º
Figure 3 | Open book representation of the BoNT/A-RBD–SV2C-LDinteraction site. The components of the complex are shown separately andcoloured according to their electrostatic potential (63 kT/e, where k is theBoltzmann constant, T is temperature and e is the elementary charge): bluedenotes basic residues, red denotes acidic residues and white denoteshydrophobic residues. The yellow ellipse indicates the approximate location ofthe hydrogen-bonding residues. The orange ellipse indicates the positivelycharged R1294 in BoNT/A-RBD and the mostly negatively charged surfaceregion in SV2C-LD. R1294 is not defined in the electron density of the complexstructure and hence does not participate in hydrogen bonds or salt bridges.
RESEARCH LETTER
2 | N A T U R E | V O L 0 0 0 | 0 0 M O N T H 2 0 1 3
Macmillan Publishers Limited. All rights reserved©2013

BoNT/A-RBD, whereas mutation of the SV2C-LD residue N559 hadno significant effect on interaction with the toxin. To further char-acterize the complex interaction, we performed fluorescence aniso-tropy experiments. Analysis of the wild-type proteins yielded a Kd
value of 0.26 6 0.2mM. Kd values obtained for the mutant proteinswere consistent with the pull-down results (Extended Data Fig. 4 andExtended Data Table 3).
It has been reported that SV2 peptides harbouring the toxin-bindingsite prevent BoNT/A from binding to neurons7. We therefore analysedthe potency of short BoNT/A-RBD- and SV2C-LD-derived peptides atinhibiting complex formation. Based on the interactions seen in thecrystal structure, the peptide sequences were designed to correspond tothe two SV2C-LD b-strands (peptide SV2C-A3), the BoNT/A-RBDb-strand (peptide BoNT/A-A5) and the BoNT/A-RBD b-sheet (pep-tide BoNT/A-A2) that interact in the complex (Extended Data Fig. 3g).We observed full inhibition of complex formation in the presence ofthe peptide corresponding to the BoNT/A-RBD b-sheet and partialinhibition in the presence of the peptide corresponding to the twoSV2C-LD b-strands, whereas no reduction in binding was seen in thepresence of the peptide corresponding to the single BoNT/A-RBDb-strand. These results indicate that the three-dimensional conforma-tion of SV2C-LD is crucial for the interaction. Consistent with thisconclusion, the shortest SV2C-LD fragment reported to bind to BoNT/A spans residues 529–566 of the SV2C sequence7. The fragmentextends over almost two turns of the b-helix and might therefore forma moderately stable three-dimensional structure. We next tested theBoNT/A-A2 b-sheet peptide for inhibition of BoNT/A-RBD bindingto and internalization by cultured striatal neurons (at 18 days in vitro)(Fig. 4 and Extended Data Fig. 5), which endogenously express SV2C,or HEK-293 cells that were transfected with Flag-tagged SV2C protein(Extended Data Fig. 6). In the absence of the toxin-derived peptide,BoNT/A-RBD co-localized with SV2C. Pre-incubation of the cells with5mM glutathione S-transferase (GST)-tagged BoNT/A-RBD peptideresulted in a significant decrease in toxin binding and internalization,whereas no inhibition of the complex interaction was observed in thepresence of a GST-only control.
The findings of our study have important implications for the develop-ment of novel antitoxin agents, as well as for the medical and cosmeticapplications of BoNT/A. The toxin has been described as one of the sixmost dangerous bioweapons1. Currently, the only available antidotes
for BoNT/A have severe side effects. Proposed alternative approachesinclude peptide inhibitors, and all three domains of the BoNTs havebeen considered as potential drug targets24. The detailed structuralinformation reported on the SV2C-LD–BoNT/A-RBD interactionshould therefore be of particular interest for the targeted developmentof BoNT/A-specific antibodies or high-affinity peptide inhibitorsdirected against the SV2-binding interface.
Depending on the type and area of medical or cosmetic application,the recommended dose of BoNT/A varies markedly25, carrying a sub-stantial risk of accidental BoNT/A overdose and poisoning. The crystalstructure of the BoNT/A-RBD in complex with SV2C-LD provides astrong structural basis for the rational design of BoNT/A variants withan attenuated capacity to bind to SV2 proteins. Such variants are pro-mising candidate proteins for broadening the very narrow therapeuticwindow, possibly resulting in much safer applications of the toxin.
METHODS SUMMARYAll of the methods are described in detail in the Methods. The proteins were clonedinto pET-based vectors and expressed in bacteria using auto-induction. Proteinpurification was performed using immobilized metal affinity chromatography(IMAC) or GST sepharose affinity chromatography followed by size-exclusionchromatography. The purified BoNT/A and SV2C domains were combined in a1:1 ratio. The complex was purified in an additional size-exclusion chromato-graphy step, concentrated to 8 mg ml21 and crystallized by vapour diffusion at20 uC. The reservoir solution consisted of 100 mM HEPES, pH 7.5, 6% PEG 8000,8% glycerol and 100 mM NaCl. For cryoprotection, the reservoir solution wassupplemented with an additional 10% glycerol. Diffraction data were collectedat the Swiss Light Source (SLS). The crystals belong to space group C 1 2 1 and arepseudomerohedrally twinned with twin law 2h, 2k, l. The structure was solved bymolecular replacement using PDB entry 3FUO as a search model. Pull-downassays were performed using IMAC with 63His-tagged BoNT/A-RBD anduntagged SV2C-LD. Kd determination was performed by fluorescence anisotropy.For functional assays, primary striatal cultures were prepared from embryonic day18 rat brains, as described for hippocampal neurons26. At 18 days in vitro, culturedstriatal neurons were stimulated using a high K1 buffer (70 mM KCl, 51.5 mMNaCl, 25 mM HEPES, 30 mM glucose, 2 mM CaCl2 and 2 mM MgCl2; pH 7.4)with or without the GST–BoNT/A-A2 peptide or a GST control, or neurons weretreated with a control buffer (2.5 mM KCl, 119 mM NaCl, 25 mM HEPES, 30 mMglucose, 2 mM CaCl2 and 2 mM MgCl2; pH 7.4), for 15 min at 37 uC. The neuronswere incubated with GFP–BoNT/A-RBD or GFP only for 10 min at 37 uC and thenwashed with high K1 or control buffer and then fixed. HEK293T cells werecultured in DMEM/Ham’s F-10 (50/50) medium supplemented with 10% FCS
GFP–BoNT/A-RBDSV2C
GFP–BoNT/A-RBD
SV2C
SV2C VGAT SV2C
VGAT
a
10
8
6
Co
-lo
caliz
ing
GF
P/V
GA
T p
un
cta
(%
)
4
2
GFP–BoNT/A-RBD
GFP
High K+
GST–A2
+ +
+++ +
+
–
–
––– –+–
––– –
–
+
+
+
–
***
***
NS
0
b
GST +––– – –
Merged
Merged
Merged
Merged
Figure 4 | The BoNT/A-A2 peptide inhibits the internalization of BoNT/A-RBD by striatal neurons (at 18 days in vitro). a, Co-localization ofendogenous SV2C with GFP–BoNT/A-RBD (top) or endogenous vesicularinhibitory amino acid transporter (VGAT) (bottom) in cultured striatalneurons. Scale bars, 50mm. Enlargements of the regions indicated by the whiterectangles are shown in the panels on the right. Scale bar, 10mm.b, Quantification of GFP–BoNT/A-RBD uptake by cultured striatal neurons.The number of co-localizing GFP–BoNT/A-RBD and VGAT puncta was
normalized to the total number of VGAT puncta. No GFP–BoNT/A-RBD,0.01% 6 0.00%; GFP, 0.03% 6 0.01%; no high K1 buffer, 0.57% 6 0.18%; noglutathione S-transferase (GST)–BoNT/A-A2 (GST–A2), 8.53% 6 0.09%;GST–BoNT/A-A2 (GST–A2), 3.42% 6 0.69%; GST, 6.84% 6 1.12%. n 5 20images per group from two cultures, Mann–Whitney U test; ***P , 0.001;NS, not significant. N 5 2 independent experiments. All tests were performedtwo-sided. Data are presented as mean 6 s.e.m.
LETTER RESEARCH
0 0 M O N T H 2 0 1 3 | V O L 0 0 0 | N A T U R E | 3
Macmillan Publishers Limited. All rights reserved©2013

and 1% penicillin/streptomycin at 37 uC in 5% CO2 and were transfected with theplasmid pGW2–mRFP (monomeric red fluorescent protein) and SV2C–Flag, orpGW2-mRFP only. After 24 h, the cells were incubated with fresh culture mediumcontaining GST–BoNT/A-A2 or a GST control for 15 min at 37 uC. The cells werethen incubated with BoNT/A-RBD for 10 min at 37 uC, washed with fresh culturemedium and fixed. Immunohistochemistry, confocal imaging, data representationand statistical analyses were performed as previously described27. Image analysiswas performed using ImageJ software.
Online Content Any additional Methods, Extended Data display items and SourceData are available in the online version of the paper; references unique to thesesections appear only in the online paper.
Received 11 January; accepted 3 October 2013.
Published online 17 November 2013.
1. Arnon, S. S. et al. Botulinum toxin as a biological weapon: medical and publichealth management. J. Am. Med. Assoc. 285, 1059–1070 (2001).
2. Carruthers, J. & Carruthers, A. Botox: beyond wrinkles. Clin. Dermatol. 22, 89–93(2004).
3. Markey, A. C. Botulinum A exotoxin in cosmetic dermatology. Clin. Exp. Dermatol.25, 173–175 (2000).
4. Montecucco, C. & Schiavo, G. Mechanism of action of tetanus and botulinumneurotoxins. Mol. Microbiol. 13, 1–8 (1994).
5. Montal, M. Botulinum neurotoxin: a marvel of protein design. Annu. Rev. Biochem.79, 591–617 (2010).
6. Jacky, B. P. et al. Identification of fibroblast growth factor receptor 3 (FGFR3) as aprotein receptor forbotulinumneurotoxin serotypeA (BoNT/A).PLoSPathogens9,e1003369 (2013).
7. Dong, M. et al. SV2 is the protein receptor for botulinum neurotoxinA. Science 312,592–596 (2006).
8. Vetting, M. W. et al. Pentapeptide repeat proteins. Biochemistry 45, 1–10 (2006).9. Holm, L. & Rosenstrom, P. Dali server: conservation mapping in 3D. Nucleic Acids
Res. 38, W545–W549 (2010).10. Bernstein, F. C. et al. The Protein Data Bank: a computer-based archival file for
macromolecular structures. J. Mol. Biol. 112, 535–542 (1977).11. Hegde, S. S. et al. A fluoroquinolone resistance protein from Mycobacterium
tuberculosis that mimics DNA. Science 308, 1480–1483 (2005).12. Vetting, M. W., Hegde, S. S., Zhang, Y. & Blanchard, J. S. Pentapeptide-repeat
proteins that act as topoisomerase poison resistance factors have a commondimer interface. Acta Crystallogr. F 67, 296–302 (2011).
13. Krishnan, R. & Lindquist, S. L. Structural insights into a yeast prion illuminatenucleation and strain diversity. Nature 435, 765–772 (2005).
14. Bryan, A.W. Jr, Starner-Kreinbrink, J. L.,Hosur, R., Clark,P. L.& Berger, B.Structure-based prediction reveals capping motifs that inhibit b-helix aggregation. Proc. NatlAcad. Sci. USA 108, 11099–11104 (2011).
15. Richardson, J. S. & Richardson, D. C. Natural b-sheet proteins use negative designtoavoid edge-to-edgeaggregation. Proc. Natl Acad.Sci. USA99, 2754–2759 (2002).
16. Ritter, C. et al. Correlation of structural elements and infectivity of the HET-s prion.Nature 435, 844–848 (2005).
17. Wasmer, C. et al. Amyloid fibrils of the HET-s(218–289) prion form a b solenoidwith a triangular hydrophobic core. Science 319, 1523–1526 (2008).
18. Stenmark, P., Dupuy, J., Imamura, A., Kiso, M. & Stevens, R. C. Crystal structureof botulinum neurotoxin type A in complex with the cell surface co-receptorGT1b—insight into the toxin-neuron interaction. PLoS Pathogens 4, e1000129(2008).
19. Lacy, D. B., Tepp, W., Cohen, A. C., DasGupta, B. R. & Stevens, R. C. Crystal structureof botulinum neurotoxin type A and implications for toxicity. Nature Struct. Biol. 5,898–902 (1998).
20. Sun,S., Tepp,W.H., Johnson, E.A.&Chapman,E.R.BotulinumneurotoxinsBandEtranslocate at different rates and exhibit divergent responses to GT1b and low pH.Biochemistry 51, 5655–5662 (2012).
21. Sun, S. et al. Receptor binding enables botulinum neurotoxin B to sense low pH fortranslocation channel assembly. Cell Host Microbe 10, 237–247 (2011).
22. Fischer, A., Mushrush,D. J., Lacy, D.B.& Montal,M.Botulinum neurotoxin devoidofreceptor binding domain translocates active protease. PLoS Pathogens 4,e1000245 (2008).
23. Fu, Z., Chen, C., Barbieri, J. T., Kim, J. J. & Baldwin, M. R. Glycosylated SV2 andgangliosides as dual receptors for botulinum neurotoxin serotype F. Biochemistry48, 5631–5641 (2009).
24. Cai, S. & Singh, B. R. Strategies to design inhibitors of Clostridium botulinumneurotoxins. Infect. Disord. Drug Targets 7, 47–57 (2007).
25. Anderson, E. R. Jr. Proper dose, preparation, and storage of botulinum neurotoxinserotype A. Am. J. Health Syst. Pharm. 61, S24–S29 (2004).
26. Kaech, S. & Banker, G. Culturing hippocampal neurons. Nature Protocols 1,2406–2415 (2006).
27. Spangler, S. A. et al. Liprin-a2 promotes the presynaptic recruitment and turnoverof RIM1/CASK to facilitate synaptic transmission. J. Cell Biol. 201, 915–928(2013).
Acknowledgements We thank A. Blanc for performing the mass spectrometry analysisof the recombinant proteins and L. Knecht for help with protein production. This workwas supported by UCB Pharma, UCB NewMedicines. This work was also supported bythe Swiss National Science Foundation (grant 310030B_138659), The NetherlandsOrganization for Scientific Research (NWO-ALW-VICI) and The NetherlandsOrganization for Health Research and Development (ZonMW-TOP).
Author Contributions R.M.B., M.H., D.M., T.C., F.L., M.O.S., G.F.X.S., C.C.H. and R.A.K.designed the research. R.M.B., D.F., M.H., J.T.K., M.M.W., C.U.S., R.J. and G.C. carried outthe research. R.M.B., M.H., J.T.K., C.C.H., G.C. and R.A.K. analysed the data. R.M.B. andR.A.K. wrote the paper with input from the other authors.
Author Information Atomic coordinates for BoNT/A-RBD–SV2C-LD have beendeposited in the Protein Data Bank database under accession number 4JRA. Reprintsand permissions information is available at www.nature.com/reprints. The authorsdeclare no competing financial interests. Readers are welcome to comment on theonline version of the paper. Correspondence and requests for materials should beaddressed to R.A.K. ([email protected]).
RESEARCH LETTER
4 | N A T U R E | V O L 0 0 0 | 0 0 M O N T H 2 0 1 3
Macmillan Publishers Limited. All rights reserved©2013

METHODSCloning and protein production. The cDNAs encoding human SV2C (Swiss-Prot Q496J9) amino acids 456–574 and BoNT/A (Swiss-Prot P10845) amino acids871–1296 were amplified by PCR using DNA templates that were codon-optimizedfor expression in Escherichia coli (GENEWIZ). The primer sequences used were:BoNT/A-RBD, 59-GTGCCGCGCGGATCCAAAAACATCATCAACACGTCCATTCTCAACCTC-39 and 59- TTGCTAAGTGAGCTCTCATTACAGCGGACGTTCACCCCAACC-39; and SV2C-LD, 59-GTGCCGCGCGGATCCGACGTAATCAAGCCGCTCCAGTC-39 and 59-TTGCTAAGTGAGCTCTCATTAGTCAAACGTGATTTGGCAACCCG-39. The BoNT/A-A2 (b-sheet) peptide insert encod-ing residues 1140–1153 was generated by using synthetic primers (59-CCACGAACATATATCTGAATTCCAGCTGATAATTGACAGAGCTCACTTAGCAAGATATAATACAAATCCGCCGAGC-39 and 59-CAGATATATGTTCGTGGTCATTACACTACCGCGACCGGATCCGGGTCCCTGGAACAGAACTTCCAGATCCGATTTTG-39). Cloning into in-house assembled, pET-based vectors con-taining an N-terminal 63His, 63His–GFP or GST tag was performed by co-transformation cloning28. Mutagenesis was performed as described previously29.All insert sequences were verified by DNA sequencing (by GATC).
The proteins were expressed separately in E. coli strain NiCo21(DE3) (NewEngland Biolabs). Precultures (100 ml Luria Bertani (LB) medium supplementedwith 50mg ml21 kanamycin) were grown overnight at 37 uC in a shaking incub-ator. ZYM-5052 medium for auto-induction30 (4 l supplemented with 50mg ml21
kanamycin) was inoculated 1:40 with the overnight culture, and the bacteria weregrown for 7 h at 37 uC. Subsequently, the temperature was lowered to 20 uC, andthe incubation was continued overnight. The GST control was expressed in LBmedium supplemented with 100mg ml21 ampicillin, using induction with 1 mMisopropylthiogalactoside (IPTG).
The cells were harvested by centrifugation. The cell pellets were resuspended in100 ml lysis buffer (50 mM Tris, pH 7.5, 500 mM NaCl, 10 mM imidazole, 10 mMb-mercapthoethanol and 1 cOmplete EDTA-free protease inhibitor cocktail tablet(Roche Diagnostics)). The cells were lysed on ice by ultrasonication. Lysate clear-ing was performed for 1 h at 25,000g, and the resultant supernatant was filtered(0.45mm filter). The proteins were subsequently purified by IMAC (on a 5 mlHisTrap FF Crude column, GE Healthcare) and gel filtration (HiLoad 16/60Superdex 200, GE Healthcare). The GST control was purified by GST sepharoseaffinity chromatography (GE Healthcare) followed by size-exclusion chromato-graphy. SV2C-LD and BoNT/A-RBD were combined in a 1:1 ratio, co-purified bysize-exclusion chromatography and concentrated to 8 mg ml21 for crystallization.Crystallization and structure elucidation. The BoNT/A-RBD–SV2C-LD com-plex was crystallized in 100 mM HEPES, pH 7.5, 6% PEG 8000, 8% glycerol and100 mM NaCl using a grid screen around the conditions described for the crys-tallization of BoNT/A-RBD alone23. For cryoprotection, the reservoir solution wassupplemented with an additional 10% glycerol. A complete 2.3 A resolution dataset was collected from a single crystal at 100 K at the X06DA beamline of the SwissLight Source using a wavelength of 1.000 A. The high-resolution limit was chosenaccording to the guidelines recently suggested by Karplus and Diederichs31. Dataprocessing was carried out with XDS32. The crystals belong to space group C 1 2 1and are pseudomerohedrally twinned with twin law 2h, 2k, l. The structure of theBoNT/A-RBD–SV2C-LD complex was solved by molecular replacement usingresidues 875–1295 of BoNT/A-RBD from PDB entry 3FUO as a search model23.Two BoNT/A-RBD molecules could be positioned in the C 1 2 1 asymmetric unitby using Phaser33. The initial atomic model was refined with phenix.refine34,taking into account the twin law. From the first refinement cycles, additionaldensity corresponding to SV2C-LD became visible. This allowed manual buildingof two copies of the domain, each one bound to a copy of BoNT/A-RBD in a 1:1complex. Coot35 was used for model building. The final refined structure exhibitsgood geometry and stereochemistry, as validated with MolProbity36 (residues infavoured regions 90.17% and outliers 0.81%). Structure figures were preparedusing PyMOL37. Electrostatic potentials were calculated using APBS38. Secondary-structure matching superimpositions39 were performed using Coot35. PROMOTIF40
was used for structure analysis. A homology model of the transmembrane part ofSV2C was generated using Phyre41. A PDB file of a lipid bilayer slice by Helleret al.42 was used for the generation of Fig. 2. Hydrogen bonds at the complexinterface were analysed using PDBePISA43 v1.37.Pull-down assays. Purified 63His-tagged BoNT/A-RBD and untagged SV2C-LDwere centrifuged separately at 16,000g for 15 min at 4 uC. The proteins were thencombined and incubated overnight at 4 uC in pull-down buffer (20 mM Tris,pH 7.8, 150 mM NaCl and 5 mM b-mercaptoethanol). The interactions of wild-type BoNT/A-RBD and mutant BoNT/A-RBD with wild-type SV2C-LD wereanalysed at 5mM and 40mM, respectively. The binding of wild-type SV2C-LDand mutant SV2C-LD to wild-type BoNT/A-RBD was analysed at 10mM and 5mM,respectively. After incubation, the samples were centrifuged, and the supernatantswere incubated with ,50ml Ni Sepharose 6 Fast Flow resin (GE Healthcare) for
1 h. The Ni Sepharose resin was washed four times with 2 ml ice-cold pull-downbuffer. Proteins were eluted in 100ml pull-down buffer supplemented with350 mM imidazole. The eluted solutions were analysed by SDS–PAGE.
Peptide competition experiments were performed in the same way exceptthat 50ml 10 mM peptide suspension in pull-down buffer was mixed with thepartner protein domain and incubated on ice for 5 min before adding the otherprotein domain. Final concentrations were 5mM BoNT/A-RBD, 20mM SV2C-LDand 2 mM peptide. Peptides were ordered from Charite. The peptide sequenceswere as follows: SV2C-A3, NH2-DSEFKNCSFFHNK-COOH; BoNT/A-A2 (b-sheet), NH2-RGSVMTTNIYLNSS-COOH; and BoNT/A-A5 (b-strand), NH2-RGSVMTTN-COOH.Protein labelling. The accessible cysteine residues of SV2C-LD were covalentlylabelled overnight with fluorescein maleimide (Molecular Probes) in 50 mM Tris,pH 7.5, 150 mM NaCl and 1 mM Tris(2-carboxyethyl)phosphine, using a 20-foldexcess of dye. Successful labelling and removal of free fluorescein by desalting(illustra NAP-5 columns, GE Healthcare) was confirmed by SDS–PAGE. Theprotein:fluorescein ratio was determined from the absorption at 280 nm and 493 nm.The absorption at 280 nm was corrected for the absorption of fluorescein to cal-culate the protein concentration. The labelling efficiency was approximately 50%.Kd determination. The binding affinities of SV2C-LD to BoNT/A-RBD (wild-type and mutants) were determined in 50 mM Tris, pH 7.5, and 150 mM NaCl byfluorescence anisotropy titration using 100 nM fluorescein-labelled SV2C-LD(SV2C-LD-F). The anisotropy signal was measured in a Cary Eclipse FluorescenceSpectrophotometer (Agilent Technologies) equipped with automated polarizers(lex 480 nm, lem 520 nm, slit 10 nm, 20 uC, 5 s signal acquisition, g 5 1.4495). Eachdata point was averaged from five measurements. To account for the reducedquantum efficiency of fluorescein at pH 5 (20 mM citrate and 150 mM NaCl),the slits were opened to 20 nm. The Kd values were determined by fitting the datato a one-site-binding model using Origin 7 (OriginLab). The binding affinity ofunlabelled SV2C-LD and its mutants was determined by displacing SV2C-LD-Ffrom the complex. The coupled equations for competitive binding were numer-ically fitted with DataFitter Software (D. Veprintsev, unpublished). The affinities ofSV2C-LD mutants were independently confirmed by determining the affinityfrom the apparent Kd of the BoNT/A-RBD–SV2C-LD-F complex in the presenceof unlabelled SV2C-LD44.Animals and cells. All animal experiments were performed in compliance with theguidelines for the welfare of experimental animals issued by the federal governmentof The Netherlands. All animal experiments were approved by the Animal EthicalReview Committee (DEC) of Utrecht University. HEK293T cells were obtainedfrom the American Type Culture Collection and tested for mycoplasma.DNA constructs and antibodies for functional studies. The SV2C–Flag con-struct was generated by PCR using the human full-length cDNA (Origene) as atemplate. The insert was cloned into a GW1-CMV vector, and the sequence wasverified. The pGW2-mRFP construct was generated as described45. The followingprimary antibodies were used: goat anti-SV2C (1:400, Santa Cruz Biotechnology,clone P-20, order no. sc-11946), rabbit anti-VGAT (1:400, Synaptic Systems, orderno. 131003), rabbit anti-Flag, (1:1,000, Sigma, order no. F7425), mouse anti-His(1:400, NeuroMab, clone N144/14, order no. 75-169) and mouse anti-tubulin-b3(TUBB3, 1:400, Sigma, order no. PRB-435P). The following secondary antibodieswere used: Alexa Fluor 488-conjugated goat anti-mouse antibodies (order no. A11029);Alexa Fluor 568-conjugated goat anti-rabbit antibodies (order no. A11036); AlexaFluor 647-conjugated goat anti-rabbit antibodies (order no. A21245) and AlexaFluor 647-conjugated goat anti-mouse antibodies (order no. A21236); Alexa Fluor488 (order no. A11055)- and Cy5 (order no. 705-175-147)-conjugated donkeyanti-goat antibodies; and Cy3-conjugated donkey anti-rabbit antibodies (orderno. 711-165-152). Alexa Fluor-conjugated antibodies were purchased fromInvitrogen, and cyanine-dye-conjugated antibodies were purchased from JacksonImmunoResearch. All secondary antibodies were used 1:400.Blocking of BoNT/A-RBD uptake by neurons. Primary striatal cultures wereprepared from embryonic day 18 rat brains, as described for hippocampal neurons26.Cells were plated on coverslips coated with 35mg ml21 poly-L-lysine (Sigma) and5mg ml21 laminin (Roche) at a density of 100,000 cells per well. Striatal cultureswere grown in neurobasal medium (NB, Invitrogen) supplemented with B27(Invitrogen), 0.5 mM glutamine (Invitrogen), 12.5mM glutamate (Sigma) andpenicillin/streptomycin mix (Invitrogen) at 37 uC in 5% CO2.
At 18 days in vitro (DIV), cultured striatal neurons were stimulated using a highK1 buffer (70 mM KCl, 51.5 mM NaCl, 25 mM HEPES, 30 mM glucose, 2 mMCaCl2 and 2 mM MgCl2; pH 7.4) with or without the GST–BoNT/A-A2 peptide(5mM) or the GST control (5mM), or neurons were treated with a control buffer(2.5 mM KCl, 119 mM NaCl, 25 mM HEPES, 30 mM glucose, 2 mM CaCl2 and2 mM MgCl2; pH 7.4), for 15 min at 37 uC. The neurons were incubated with GFP–BoNT/A-RBD (200 nM) or GFP only (200 nM) for 10 min at 37 uC. The cells were
LETTER RESEARCH
Macmillan Publishers Limited. All rights reserved©2013

then washed with high K1 or control buffer solution and fixed with 4% para-formaldehyde/4% sucrose in PBS for 10 min.Blocking of BoNT/A-RBD binding in HEK293T cells. HEK293T cells werecultured in DMEM/Ham’s F-10 (50/50) medium supplemented with 10% FCSand 1% penicillin/streptomycin at 37 uC in 5% CO2 and were transfected withpGW2–mRFP and SV2C–Flag, or pGW2–mRFP only, using the FuGENE 6reagent (Roche). After 24 h, cells were incubated with fresh culture medium con-taining GST–BoNT/A-A2 or the GST control for 15 min at 37 uC. Next, cells wereincubated with BoNT/A-RBD (100 nM) for 10 min at 37 uC, washed with freshculture medium and fixed with 4% paraformaldehyde/4% sucrose in PBS for10 min.Immunohistochemistry, confocal imaging and image analysis. After fixation,cells were washed three times with PBS for 5 min and incubated with the indicatedprimary antibodies in GDB buffer (0.2% BSA, 0.8 M NaCl, 0.5% Triton X-100 and30 mM phosphate buffer; pH 7.4) overnight at 4 uC. Neurons were then washedthree times for 5 min with PBS, incubated with secondary antibodies in GDBbuffer for 1 h at room temperature and washed again three times for 5 min withPBS. Coverslips were mounted using VECTASHIELD mounting medium (VectorLaboratories).
Confocal images were acquired using an LSM 700 confocal laser-scanningmicroscope (Zeiss) using a 403 (1.3 NA) oil objective. Each image was acquiredfrom a z-series of 5 images, each averaged twice, and was chosen to cover the entireregion of interest from top to bottom using a 0.47-mm interval. The resultantz-stack was ‘flattened’ into a single image using maximum intensity projection.All image acquisition settings were kept constant throughout the experiments.
Image analysis was performed using ImageJ software (National Institutes ofHealth). For analysis of the amount of synaptic GFP–BoNT/A-RBD uptake bycultured striatal neurons, the ImageJ plug-in ‘Puncta Analyzer’ was used to auto-matically count the number of VGAT- and GFP–BoNT/A-RBD positive puncta.Threshold values were held constant throughout the experiments. To excludenonspecific background signal, a filter excluding puncta smaller than 0.6mm2
and larger than 6.3mm2 was applied. The number of co-localizing VGAT andGFP–BoNT/A-RBD puncta was normalized to the total number of VGAT-positivepuncta to correct for changes in neuron density per image. For analysis of theamount of BoNT/A-RBD binding in HEK293T cells, the total area of BoNT/A-RBD was normalized to the total area of mRFP-transfected HEK293T cells abovean assigned threshold. Threshold values were kept constant throughout the experi-ments. Thresholded images were processed using a median filter to remove noise.Data representation and statistics. The number of cells analysed was based onprevious data sets27. For all experiments, N (number of replicates) represents bio-logical replicates. In all bar graphs, the data are presented as the mean values 6 s.e.m.
Statistical analysis was performed using SPSS Statistics v20 (IBM). The non-parametricMann–Whitney U test was used to test for statistical significance. P values below0.05 are considered significant (*P , 0.05; **P , 0.01; ***P , 0.001; NS, notsignificant). All tests were performed two-sided. The data samples were testedfor normal distribution with the Kolmogorov–Smirnov test and for heterogeneityof variance with Bartlett’s test.
28. Benoit, R. M., Wilhelm, R. N., Scherer-Becker, D. & Ostermeier, C. An improvedmethod for fast, robust, and seamless integration of DNA fragments into multipleplasmids. Protein Expr. Purif. 45, 66–71 (2006).
29. Olieric, N. et al. Automated seamless DNA co-transformation cloning with directexpression vectors applying positive or negative insert selection. BMC Biotechnol.10, 56 (2010).
30. Studier, F. W. Protein production by auto-induction in high density shakingcultures. Protein Expr. Purif. 41, 207–234 (2005).
31. Karplus, P. A. & Diederichs, K. Linking crystallographic model and data quality.Science 336, 1030–1033 (2012).
32. Kabsch, W. XDS. Acta Crystallogr. D 66, 125–132 (2010).33. McCoy, A. J.et al.Phasercrystallographic software. J. Appl. Crystallogr.40, 658–674
(2007).34. Afonine, P. V. et al. Towards automated crystallographic structure refinement with
phenix.refine. Acta Crystallogr. D 68, 352–367 (2012).35. Emsley, P., Lohkamp, B., Scott, W. G. & Cowtan, K. Features and development of
Coot. Acta Crystallogr. D 66, 486–501 (2010).36. Chen, V. B. et al. MolProbity: all-atom structure validation for macromolecular
crystallography. Acta Crystallogr. D 66, 12–21 (2010).37. DeLano, W. L. The PyMOL Molecular Graphics System. (DeLano Scientific, 2012).38. Baker, N. A., Sept, D., Joseph, S., Holst, M. J. & McCammon, J. A. Electrostatics of
nanosystems: application to microtubules and the ribosome. Proc. Natl Acad. Sci.USA 98, 10037–10041 (2001).
39. Krissinel, E. & Henrick, K. Secondary-structure matching (SSM), a new tool forfast protein structure alignment in three dimensions. Acta Crystallogr. D 60,2256–2268 (2004).
40. Hutchinson, E. G. & Thornton, J. M. PROMOTIF—a program to identify and analyzestructural motifs in proteins. Protein Sci. 5, 212–220 (1996).
41. Kelley, L. A. & Sternberg,M. J.Protein structureprediction on the Web: a case studyusing the Phyre server. Nature Protocols 4, 363–371 (2009).
42. Heller,H.,Schaefer,M.&Schulten, K.Moleculardynamics simulation ofa bilayerof200 lipids in the gel and in the liquid crystal phase. J. Phys. Chem. 97, 8343–8360(1993).
43. Krissinel, E. & Henrick, K. Inference ofmacromolecular assemblies from crystallinestate. J. Mol. Biol. 372, 774–797 (2007).
44. Zhang, Y. L. & Zhang, Z. Y. Low-affinity binding determined by titration calorimetryusing a high-affinity coupling ligand: a thermodynamic study of ligand binding toprotein tyrosine phosphatase 1B. Anal. Biochem. 261, 139–148 (1998).
45. Kapitein, L. C., Yau, K. W. & Hoogenraad, C. C. Microtubule dynamics in dendriticspines. Methods Cell Biol. 97, 111–132 (2010).
RESEARCH LETTER
Macmillan Publishers Limited. All rights reserved©2013
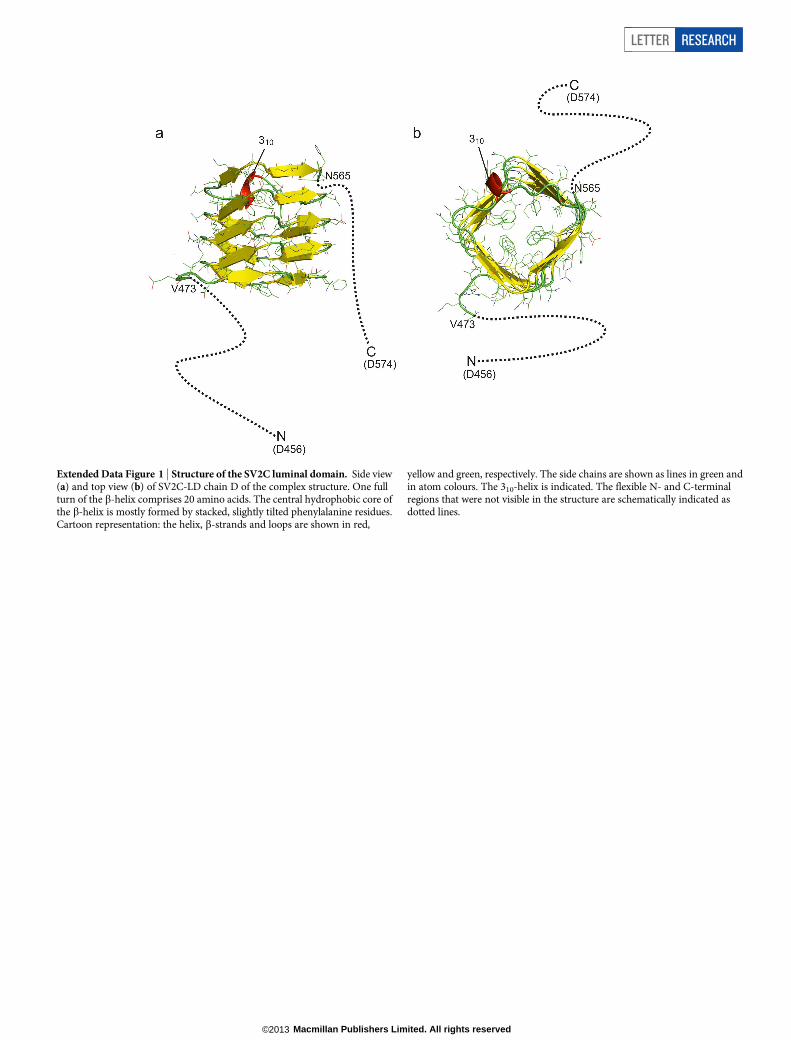
Extended Data Figure 1 | Structure of the SV2C luminal domain. Side view(a) and top view (b) of SV2C-LD chain D of the complex structure. One fullturn of the b-helix comprises 20 amino acids. The central hydrophobic core ofthe b-helix is mostly formed by stacked, slightly tilted phenylalanine residues.Cartoon representation: the helix, b-strands and loops are shown in red,
yellow and green, respectively. The side chains are shown as lines in green andin atom colours. The 310-helix is indicated. The flexible N- and C-terminalregions that were not visible in the structure are schematically indicated asdotted lines.
LETTER RESEARCH
Macmillan Publishers Limited. All rights reserved©2013

Extended Data Figure 2 | Binding of BoNT/A-RBD to SV2C-LD is reducedon acidification. Normalized fluorescence anisotropy titration ofBoNT/A-RBD with labelled SV2C-LD (a) and displacement with unlabelled
SV2C-LD at pH 7.5 (green) and pH 5 (red) (b). The affinity of SV2C-LD forBoNT/A-RBD at pH 5 is reduced by a factor of ,5. For values, see ExtendedData Table 3.
RESEARCH LETTER
Macmillan Publishers Limited. All rights reserved©2013

Extended Data Figure 3 | Interaction between SV2C-LD andBoNT/A-RBD. a, Overview of the prominent interactions that were analysedby site-directed mutagenesis. The colour code in Fig. 1 is used. b, SDS–PAGEanalysis of the pull-down assays. The 63His-tagged BoNT/A domain(,50 kDa) and the untagged SV2C domain (, 15 kDa) are indicated byarrows. c–f, Close-up views of specific interactions. c, The hydrogen bonds ofN559. d, The hydrogen bonds of T1145/T1146. e, The cation–p stackinginteraction between BoNT/A-RBD R1156 and SV2C-LD F563. f, The putativelong-range electrostatic interactions of BoNT/A-RBD R1294. R1294 is not
defined in the electron density of the complex structure and hence does notparticipate in hydrogen bonds or salt bridges. Nevertheless, mutagenesis ofR1294 to alanine strongly reduces the binding of BoNT/A-RBD to SV2C-LD.We speculate that long-range electrostatic interactions between the positivelycharged BoNT/A-RBD arginine (depicted as a surface coloured according tothe electrostatic potential) and the negatively charged regions in SV2C-LD havea role in complex formation. g, Sequences and schematic representations of thepeptides that were used for the complex inhibition studies.
LETTER RESEARCH
Macmillan Publishers Limited. All rights reserved©2013

Extended Data Figure 4 | Kd determination of wild-type and mutantBoNT/A-RBD and SV2C-LD proteins. The affinities of wild-type and mutantSV2C-LD for BoNT/A-RBD were determined by fluorescence anisotropytitration of labelled SV2C-LD (a, c, f) and subsequent displacement withunlabelled SV2C-LD (b), SV2C-LD F563A (d) or SV2C-LD N559A(g). Alternatively, the affinities of SV2C-LD mutants were calculated from the
apparent Kd of labelled SV2C-LD in the presence (green) or absence (red)of SV2C-LD F563A (22.6mM, e) or SV2C-LD N559A (18.2mM, h). Theaffinities of BoNT/A-RBD R1294A (i, j), BoNT/A-RBD R1156E (k, l) andBoNT/A-RBD T1145A/T1146A (m, n) for SV2C-LD were determinedaccordingly by anisotropy displacement titrations. For values, see ExtendedData Table 3.
RESEARCH LETTER
Macmillan Publishers Limited. All rights reserved©2013

Extended Data Figure 5 | The BoNT/A-A2 peptide inhibits theinternalization of BoNT/A-RBD by striatal neurons. Representativeimages of GFP–BoNT/A-RBD uptake by cultured striatal neurons (DIV18).Neurons were pre-incubated with the GST–BoNT/A-A2 or the GSTcontrol (5mM, 15 min) in high K1 or control buffer and treated with
GFP–BoNT/A-RBD or GFP only (200 nM, 10 min) and stained for theneuronal marker tubulin-b3 (TUBB3) and endogenous VGAT to labelpresynaptic terminals. Scale bar, 50mm. Representative images are from a totalof 20 images per group, N 5 2 independent experiments.
LETTER RESEARCH
Macmillan Publishers Limited. All rights reserved©2013

Extended Data Figure 6 | The BoNT/A-A2 peptide inhibits the binding ofBoNT/A-RBD in HEK293T cells. a, A typical example of BoNT/A-RBD(green) binding to SV2C–Flag (blue) in HEK293T cells. Cells were transfectedwith SV2C–Flag and mRFP (red) to highlight transfected cells, fixed andstained using Flag- and His-tagged antibodies. DAPI (49,6-diamidino-2-phenylindole, purple) was used for visualizing all cell nuclei. Scale bar, 50mm.b, Quantification of BoNT/A-RBD binding in SV2C–Flag expressingHEK293T cells. Cells were transfected with SV2C–Flag and mRFP, ormRFP only, and incubated with GST–BoNT/A-A2 peptide or GST control
(5mM, 15 min) and treated without (control) or with BoNT/A-RBD (100 nM,10 min). The total area of BoNT/A-RBD was normalized to the total area ofmRFP-transfected cells. No BoNT/A-RBD: 1.24 6 0.64%, n 5 8; noSV2C–Flag: 0.82 6 0.64%, n 5 5; no GST–BoNT/A-A2: 42.13 6 7.44%, n 5 9;5mM GST–BoNT/A-A2: 6.92 6 1.93%, n 5 9; 5mM GST: 24.55 6 8.01%,n 5 9; Mann–Whitney U test, *** P , 0.001, * P , 0.05, NS not significant,n 5 number of images analysed per group, N 5 2 independent experiments.All tests were performed two-sided. Data are presented as the mean 6 s.e.m.
RESEARCH LETTER
Macmillan Publishers Limited. All rights reserved©2013

Extended Data Table 1 | Data collection and refinement statistics
*The highest resolution shell is shown in parentheses.
LETTER RESEARCH
Macmillan Publishers Limited. All rights reserved©2013

Extended Data Table 2 | SV2C-LD–BoNT/A-RBD hydrogen bonds
The table summarizes the hydrogen bonds of the interface between chain A and chain D.
RESEARCH LETTER
Macmillan Publishers Limited. All rights reserved©2013

Extended Data Table 3 | Kd values of the interaction of wild-type and mutant proteins
aDisplacement with unlabelled protein. bIn the presence of the unlabelled mutant SV2C-LD. Please note that the affinity of SV2C-LD for BoNT/A-RBD R1156E and T1145A/T1146A is reduced to an extent thatallows only determination of a lower boundary for the affinity of unlabelled SV2C-LD (Extended Data Fig. 4l, n). However, these values are consistent with the reduced binding affinity of labelled SV2C-LD and thepull-down experiments.
LETTER RESEARCH
Macmillan Publishers Limited. All rights reserved©2013





























