Embed Size (px)
Citation preview

Romanian Biotechnological Letters Vol. 19, No. 4, 2014 Copyright © 2014 University of Bucharest Printed in Romania. All rights reserved
ORIGINAL PAPER
9517 Romanian Biotechnological Letters, Vol. 19, No. 4, 2014
Stress Effects on Cutaneous Nociceptive Nerve Fibers and Their Neurons of Origin in Rats
Received for publication, February6, 2014 Accepted, February 21, 2014
CONSTANTIN CARUNTU1,2, DANIEL BODA2*, SORIN MUSAT3, ANA CARUNTU4, ELENA POENARU5, BOGDAN CALENIC6, ILINCA SAVULESCU-FIEDLER7, ALINA DRAGHIA8, MARIA ROTARU9, ANCA IOANA BADARAU1
1Physiology Department, “Carol Davila” University of Medicine and Pharmacy, Bucharest, Romania 2Dermatology Research Laboratory, “Carol Davila” University of Medicine and Pharmacy, Bucharest, Romania 3Themis Pathology SRL, Bucharest, Romania 4“Dan Theodorescu” Oral and Maxillofacial Surgery Hospital, Bucharest, Romania 5Department of Medical Informatics and Biostatistics, “Carol Davila” University of Medicine and Pharmacy, Bucharest, Romania 6Department of Biochemistry, Faculty of Dental Medicine, “Carol Davila” University of Medicine and Pharmacy, Bucharest, Romania 7Internal Medicine Department - ”Coltea” Clinical Hospital, ”Carol Davila” University of Medicine and Pharmacy, Bucharest, Romania 8Department of Cellular and Molecular Medicine, "Carol Davila" University of Medicine and Pharmacy, Bucharest, Romania 9Department of Dermatology, “Lucian Blaga” University of Sibiu, Romania *Address correspondence to: Daniel Boda,Dermatology Research Laboratory, “Carol Davila” University of Medicine and Pharmacy, 22-24 Gr. Manolescu, 0111234 Bucharest, Romania; Tel: +40757079117; Fax: +40212221310; E-mail: [email protected]
Abstract
Stress effect on nociception is a topic of particular interest in scientific research. Our objective is to evaluate the influence of stress on dorsal root ganglion (DRG) nociceptive neurons and on cutaneous nerve fibers arising from these neurons, from both glabrous and hairy skin.
The study included adult male Wistar rats divided into four groups: Stress 1 day, Stress 10 days, Stress 21 days and Control. Stress was administered by restraining the rats for 2 h per day. Subsequently, skin samples and DRGs were collected for immunohistochemical evaluation of cutaneous nociceptive nerve fibers and of primary sensory neurons, respectively.
Our results show that stress exposure induces a moderate increase of dermal nociceptive free nerve endings density in glabrous skin. The stress-induced increase of nociceptive activity, as evidenced by an increased expression of peripherin and TRPV1 (VR1) receptor, was more prominent in primary sensory neurons. Besides the impact on pain sensitivity, increased activity of nociceptive nervous structures might be associated with activation of neurogenic inflammatory processes. Thus, the impact of stress on nervous structures involved in nociception may explain both the role of stress in modulating nociceptive sensitivity and stress involvement in triggering or worsening of many inflammatory skin diseases.
Keywords:nociception, peripherin, TRPV1 (VR1), skin, DRG neurons
1. Introduction
Stress, “the nonspecific response of the body to any demand”, as it was defined by Hans Selye [1, 2], has become a common part of modern lifestyle, in which the human body has to face more frequently and more intense challenges for which to adapt. The action of stressors is associated with significant changes in homeostatic mechanisms of the body, but also has a complex emotional impact [3]. In turn, pain has a physical and an emotional component, being defined as „an unpleasant sensory and emotional experience associated

CONSTANTIN CARUNTU, DANIEL BODA, SORIN MUSAT, ANA CARUNTU, ELENA POENARU, BOGDAN CALENIC, ILINCA SAVULESCU-FIEDLER, ALINA DRAGHIA, MARIA ROTARU,
ANCA IOANA BADARAU
Romanian Biotechnological Letters, Vol. 19, No. 4, 2014 9518
with actual or potential tissue damage or described in terms of such damage” [4]. Thus, even at a conceptual level, a complex relationship between stress and pain is to be expected.
In this area of research, a topic of particular interest is the influence of stress on nociception. Even though numerous studies have investigated this topic, the results are conflicting. Some researchers have shown that stress associates with a reduced nociceptive sensitivity, while others report its augmentation. Stress effect on nociceptive threshold seems to be dependent on the character of the exposure to stress: acute or chronic, the type of the stressing agent, and on various individual factors [5-8]. Previous studies have shown that chronic psychoemotional stress reduces the pain threshold in humans [9]. Moreover, most previous experimental studies have shown that chronic exposure to stress is associated with a hiperalgesic effect for thermal, mechanical and chemical nociceptive stimuli [7, 10-13]. This effect is probably dependent on the sympato-adrenal system, involving neuroplasticity and phenotypical changes in nociceptors [7, 10, 14-16]. However, other studies have shown opposing effects, such as an increased nociceptive threshold induced by repeated exposure to stress in rats [17]. Conflicting results on the effect of acute stress on nociception have also been described. Thus, there are studies on human subjects suggesting an increased sensitivity to pain after psychogenic stress induction [18-20]. Also, numerous studies conducted on laboratory animals have shown that acute stress inhibits nociceptive reflex responses induced by chemical, thermal or mechanical stimuli [5, 11, 12, 21-34]. As a consequence, a stress-induced analgesic effect was suggested, probably mediated by the activation of descending inhibitory pain pathways [8, 31, 35]. However, other studies suggest that acute stress may be associated with an increase in nociceptive reflex responses [36-38]. It was also shown that acute stress stimulates learned escape responses induced by nociceptive stimulation, suggesting a hyperalgesic effect [5].
Considering that assessment of impact exhibited by stress on the process of skin nociception is associated with so many controversies, a study of stress-induced morphological changes of nervous structures involved in nociception may provide a new insight into the complex relationship between stress and pain. One of the best methods to identify dermal nociceptive nerve fibers and to assess the impact of stress on their density is by using peripherin as an immunohistochemical marker. Peripherin is a protein that belongs to the type III intermediate filaments and is a marker of nociceptive thin myelinated and unmyelinated nerve fibers and of their neurons of origin [39, 40]. In order to investigate the effects of stress on DRG (dorsal root ganglion) nociceptive neurons, a very efficient technique is the immunohistochemical assessment of peripherin and TRPV1 (Transient Receptor Potential Vanilloid 1) (VR1) receptor expression. Previous immunohistochemical studies have shown that in adult rats more than half of the DRG neurons are immunoreactive to peripherin, and that most of them are small-diameter neurons [41]. TRPV1 (VR1) is a marker of nociceptive nerve structures and is mainly expressed by a subpopulation of small diameter DRG neurons that are the origin of nociceptive unmyelinated fibers [42]. The same receptor was identified in the neurons of origin for thin myelinated nociceptive fibers [43]. In adult rats, most of the DRG neurons are immunoreactive for TRPV1 (VR1) and over 70% of the TRPV1 (VR1) positive DRG neurons are also immunoreactive for peripherin [40].
In most previous experimental studies, stress exposure was limited to a single application, and the effects on skin were investigated immediately after stress exposure [44], at 1 day [45] or 2 days post-stress [46-48]. A single study used repeated stress exposure for 1 hour per day, for 10 consecutive days [49]. Moreover, the great majority of previous studies have investigated the effect of stress on nociceptive nerve fibers only in hairy skin.

Stress Effects on Cutaneous Nociceptive Nerve Fibers and Their Neurons of Origin in Rats
9519 Romanian Biotechnological Letters, Vol. 19, No. 4, 2014
The objective of our study is to evaluate the influence of short-term and repeated stress exposure on the DRG nociceptive neurons and on the cutaneous nerve fibers arising from these neurons in both hairy and glabrous skin.
2. Materials and methods
2.1. Experimental protocol. Experiments were performed in accordance with European Guidelines on Laboratory Animal Care and the methodology approved by the "Carol Davila"University of Medicine and Pharmacy Ethics Committee.
The study included 29 adult male Wistar rats, weighing 250-350 g, that were randomly divided into four groups: Stress 1 day group (n = 8), Stress 10 days group (n = 7), Stress 21 days group (n = 6) and Control group (n = 8). Rats were kept in clean cages, maximum of 5 rats per cage in standard experimental conditions, including light - dark cycles of 12h/12h, temperature 22 ± 2° C and free access to water and food.
The protocol of stress induction consisted in placing the rats in a well ventilated restrainer that did not allow their movement without causing any compression. The stress induced by this technique is considered to be of moderate intensity and of a mainly psychological nature [50-52]. The 2 hour/day restraining procedure was administered in a separate room, for 1, 10 and 21 consecutive days (Stress 1 day, Stress 10 days and Stress 21days groups, respectively). The rats from the Control group were kept in their standard under normal housing conditions.
Subsequently, the animals were euthanized by ethyl ether overdose, and hairy and glabrous skin samples were collected for the immunohistochemical examination of cutaneous nociceptive nerve fibers. Glabrous skin samples were harvested from the plantar region of the posterior members and the hairy skin samples from the ears. Also, for the immunohistochemical evaluation of primary sensory neurons, DRGs from thoracolumbar region were collected from two rats for each group, each ganglion being considered as a separate entity. The total number (n) of DRGs analyzed for each group was as follows: Stress 1 day group n = 7, Stress 10 days group n = 5, Stress 21 days group n = 5 and Control group n = 4.
2.2. Preparation of tissue samples for light microscopy. Tissue samples were collected within 2 minutes after the animals were sacrificed for skin samples and within 5 minutes for DRGs. After harvesting, samples were fixed in 4% formaldehyde in 150 mM phosphate buffer, pH 7.5, for 24±1 hours. Representative fragments of glabrous and hairy skin and DRGs were dehydrated through a graded series of ethanols (70, 90, 96, 100%), clarified in n-butanol and embedded in paraffin. Representative tissue fragments were dissected from the donor paraffin blocks and placed in a recipient sectionable vegetal matrix, (maximum 12 samples per block), using the "tissue microarray" technique [53]. The resulting blocks were sectioned semiserially at 5 m and prepared for staining.
2.3. Imunohistochemistry. The resulting slides were baked for 60 minutes at 59°C, dewaxed in xylene and rehydrated through a graded series of ethanols, as customary. Antigen retrieval was performed using Tris -EDTA solution, pH 9.0, for 5 minutes, at 126° C. The slides were blocked for non-specific staining for 1 hour with 5% normal goat serum in TBS (Tris buffered saline, 50mM, pH 7.4, supplemented with 0.05% Tween 20) and incubated at 4° C for 18 h with primary antibody in the following concentrations: 1/1000 for anti-Peripherin – marker for nociceptive neurons, and 1/500 for anti-VR1. The slides were rinsed with TBS and endogenous peroxidases were inactivated with 3% hydrogen peroxide for 30 minutes at room temperature. The slides were rinsed and then incubated for 30 minutes with Envision Flex HRP (Dako, Glostrup, Denmark). The color reaction was developed with DAB + (Dako, Glostrup, Denmark) for 5 minutes. Nuclear counterstaining was performed with 1%

CONSTANTIN CARUNTU, DANIEL BODA, SORIN MUSAT, ANA CARUNTU, ELENA POENARU, BOGDAN CALENIC, ILINCA SAVULESCU-FIEDLER, ALINA DRAGHIA, MARIA ROTARU,
ANCA IOANA BADARAU
Romanian Biotechnological Letters, Vol. 19, No. 4, 2014 9520
chlorofom-purified methyl green in acetate buffer, pH 4.0 for 20 minutes. The slides were rinsed with distilled water, dehydrated briefly with n-butanol, clarified with xylene, mounted in Cytoseal 60 and stored for further analysis.
Figure 1.(A)Dermal peripherin positive nerve fibers enclosed in peripheral nerve ( ) from
glabrous skin; (B) Free nerve endings at the base of a hair follicle (*) and dermal peripherin positive nerve fibers ( ) in hairy skin. Original magnification 400×.
2.4. Histomorphomeric analysis of images by light microscopy. Light microscopy
images were acquired and stored digitally using an Olympus CKX41 microscope equipped with a SLR Olympus E330 digital camera. Evaluation of light microscopy images was performed in a blinded fashion, using the image analysis software ImageJ 1.45. Histomorphometric analysis of skin tissue was performed at a magnification of 400× and the epidermis, dermis and hypodermis were examined separately. Since the most conclusive data were obtained at the dermis level, this skin compartment was selected for further evaluation. For both glabrous and hairy skin, we determined the density of peripherin positive nervous structures – nerve fibers enclosed in peripheral nerves and free nerve endings (see Figure 1), per mm2 of dermal tissue. Dermal area of each evaluated skin sample section was delineated and quantified using microscopic calibration scales. Subsequently, all peripherin positive nerve structures detected in the examined dermal area were evaluated, and their number was quantified in relation to the total area of dermal tissue.
For histomorphometric analysis of DRGs, we examined images acquired at 100× magnification. We performed a semiquantitative assessment of immunohistochemical staining for peripherin in primary sensory neurons. Cells were segregated in four categories: negative, low-positive, positive and highly-positive. We analysed all the neurons identified on the tissue slides of each examined ganglion and we determined the percentage of neurons from each category compared to the total number of neurons. For each ganglion we examined at least 40 cells. A similar procedure was conducted for TRPV1 (VR1) immunohistochemistry (for each ganglion a minimum of 80 cells being examined).
2.5. Chemicals. All reagents were purchased from Sigma-Aldrich (St. Louis, Missouri, USA), unless otherwise stated. The primary antibodies used in

Stress Effects on Cutaneous Nociceptive Nerve Fibers and Their Neurons of Origin in Rats
9521 Romanian Biotechnological Letters, Vol. 19, No. 4, 2014
immunohistochemical study were: anti-Peripherin antibody - nociceptive neuron marker (ab4666, Abcam, Cambridge, UK) and anti-VR1 antibody (ab6166, Abcam, Cambridge, UK).
2.6. Statistical analysis. We used Bartlett's test for the assessment of the normality and homogeneity of data distribution. The differences between experimental groups were evaluated using one-way between groups analysis of variance (ANOVA) followed by Tukey post hoc test, (when data had normal distribution and homogeneity), and Kruskall-Wallis test followed by 2-tailed Multiple Comparison Test (when data did not express a normal distribution and homogeneity), respectively. For each group, the results were expressed as mean ± standard deviation (SD). P values <0.05 were considered significant.
3. Results
3.1. Assessment of stress effects on the density of dermal peripherin positive nervous structures
Figure 2.Comparative analysis between groups of the density of dermal peripherin positive
nerve fibers and free nerve endings in glabrous and hairy skin. In glabrous skin stress exposure had no impact on (A) the density of dermal peripherin positive nerve fibers, but induced an increase of (B) peripherin positive free nerve endings. In hairy skin, there was no significant difference between groups for (C) peripherin positive dermal nerve fibers, nor for (D) dermal peripherin positive free nerve endings.
In glabrous skin there was no statistically significant difference between groups for the
density of dermal nerve fibers enclosed in peripheral nerves (P = 0.342595, ANOVA). However, stress exposure induced an increase of dermal peripherin positive free nerve endings from 5.63061 ± 4.28291 / mm2 for Control group, to 11.76151 ± 6.18606 / mm2 for Stress 1 day group, 14.54485 ± 10.45456 / mm2 for Stress 10 days group, and 15.78646 ± 6.38119 / mm2 Stress 21 days group (P = 0.049864, ANOVA). However, the comparative

CONSTANTIN CARUNTU, DANIEL BODA, SORIN MUSAT, ANA CARUNTU, ELENA POENARU, BOGDAN CALENIC, ILINCA SAVULESCU-FIEDLER, ALINA DRAGHIA, MARIA ROTARU,
ANCA IOANA BADARAU
Romanian Biotechnological Letters, Vol. 19, No. 4, 2014 9522
analysis between groups did not reveal differences with statistical significance (all P values >0.061604, Tukey post hoc test) (see Figure 2).
In hairy skin, no statistically significant difference between groups was found for the dermal nerve fibers enclosed in peripheral nerves (P = 0.068166, ANOVA), nor for the dermal free nerve endings (P = 0.735535, ANOVA) (see Figure 2).
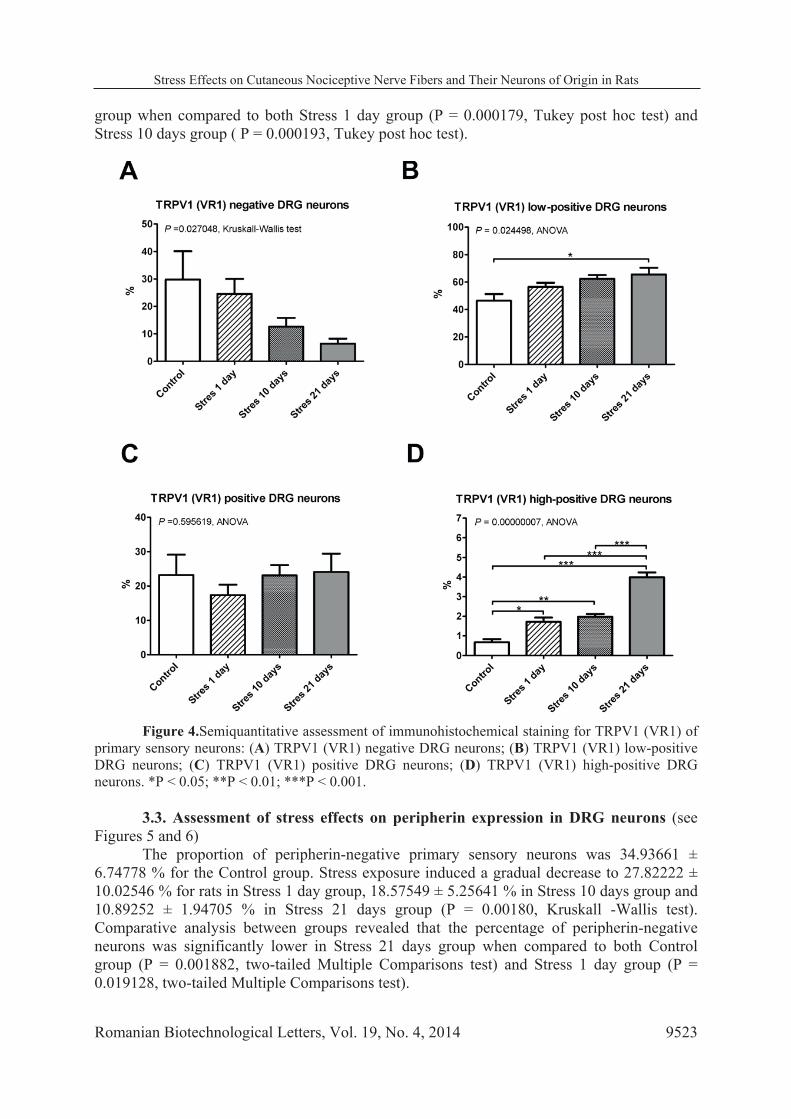
3.2. Assessment of stress effects on TRPV1 (VR1) expression in DRG neurons (see Figures 3 and 4)
Figure 3.Immunohistochemical evaluation of TRPV1 (VR1) expression in primary sensory
neurons; DRGs harvested from (A) Control; (B) Stress 1 day; (C) Stress 10 days; (D) Stress 21 days groups. Original magnification 100×.
In Control group the proportion of TRPV1 (VR1) negative primary sensory neurons
was 29.74185 ± 20.61401 % . Stress exposure was associated with a gradual decrease in their proportion to 24.50735 ± 14.51920 % in Stress 1 day group, 12.59128 ± 7.18412 % in Stress 10 days group and 6.43266 ± 4.08520 % in Stress 21 days group (P = 0.027048, Kruskall-Wallis test). However, comparative analysis between groups showed no statistical significance (all P values > 0.05882, two-tailed Multiple Comparisons test).
The percent of TRPV1 (VR1) low-positive neurons increased gradually from 46.41394 ± 9.64744 % in Control group to 56.45082 ± 8.04705 % in Stress 1 day group, 62.37468 ± 6.15587 % in Stress 10 days group and 65.47957 ± 11.011 % in Stress 21 days group (P = 0.024498 , ANOVA). A statistically significant difference was seen between Control rats and rats exposed to stress for 21 days (P = 0.022458, Tukey post hoc test).
Proportion of TRPV1 (VR1) positive neurons was similar for all experimental groups (P = 0.595619, ANOVA). In contrast, exposure to stress led to an important increase in the percentage of high-positive neurons, from 0.67262 ± 0.32357 % in Control group to 1.71578 ± 0.57948 % in Stress 1 day group, 1.97046 ± 0.32703 % in Stress 10 days group and 3.98790 ± 0.53883 % in Stress 21 days group (P = 0.00000007, ANOVA) . When compared to the Control group, this difference was statistically significant even after a single session of stress exposure (P = 0.014266, Tukey post hoc test), and increased even further for Stress 10 days group ( P = 0.004481, Tukey post hoc test) and for Stress 21 days group (P = 0.000178, Tukey post hoc test). In addition, statistically significant changes were found in Stress 21 day
Stress Effects on Cutaneous Nociceptive Nerve Fibers and Their Neurons of Origin in Rats
9523 Romanian Biotechnological Letters, Vol. 19, No. 4, 2014
group when compared to both Stress 1 day group (P = 0.000179, Tukey post hoc test) and Stress 10 days group ( P = 0.000193, Tukey post hoc test).
Figure 4.Semiquantitative assessment of immunohistochemical staining for TRPV1 (VR1) of
primary sensory neurons: (A) TRPV1 (VR1) negative DRG neurons; (B) TRPV1 (VR1) low-positive DRG neurons; (C) TRPV1 (VR1) positive DRG neurons; (D) TRPV1 (VR1) high-positive DRG neurons. *P < 0.05; **P < 0.01; ***P < 0.001.
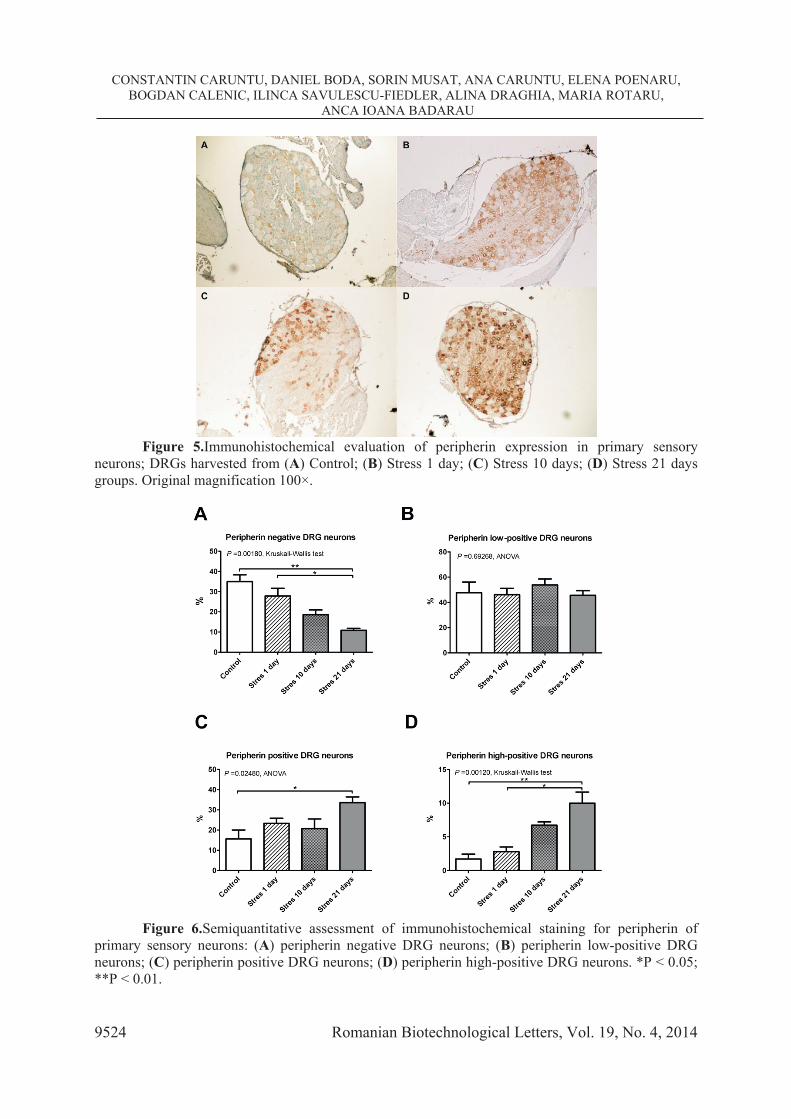
3.3. Assessment of stress effects on peripherin expression in DRG neurons (see
Figures 5 and 6) The proportion of peripherin-negative primary sensory neurons was 34.93661 ±
6.74778 % for the Control group. Stress exposure induced a gradual decrease to 27.82222 ± 10.02546 % for rats in Stress 1 day group, 18.57549 ± 5.25641 % in Stress 10 days group and 10.89252 ± 1.94705 % in Stress 21 days group (P = 0.00180, Kruskall -Wallis test). Comparative analysis between groups revealed that the percentage of peripherin-negative neurons was significantly lower in Stress 21 days group when compared to both Control group (P = 0.001882, two-tailed Multiple Comparisons test) and Stress 1 day group (P = 0.019128, two-tailed Multiple Comparisons test).
CONSTANTIN CARUNTU, DANIEL BODA, SORIN MUSAT, ANA CARUNTU, ELENA POENARU, BOGDAN CALENIC, ILINCA SAVULESCU-FIEDLER, ALINA DRAGHIA, MARIA ROTARU,
ANCA IOANA BADARAU
Romanian Biotechnological Letters, Vol. 19, No. 4, 2014 9524
Figure 5.Immunohistochemical evaluation of peripherin expression in primary sensory
neurons; DRGs harvested from (A) Control; (B) Stress 1 day; (C) Stress 10 days; (D) Stress 21 days groups. Original magnification 100×.
Figure 6.Semiquantitative assessment of immunohistochemical staining for peripherin of
primary sensory neurons: (A) peripherin negative DRG neurons; (B) peripherin low-positive DRG neurons; (C) peripherin positive DRG neurons; (D) peripherin high-positive DRG neurons. *P < 0.05; **P < 0.01.

Stress Effects on Cutaneous Nociceptive Nerve Fibers and Their Neurons of Origin in Rats
9525 Romanian Biotechnological Letters, Vol. 19, No. 4, 2014
The proportion of peripherin low-positive neurons was similar in all studied groups (P = 0.69268, ANOVA). In contrast, the percentage of peripherin positive neurons was augmented by stress exposure, increasing from 15.61254 ± 8.83484 % in Control group to 23.28804 ± 6.71772 % in Stress 1 day group, 20.79931 ± 10.49907 % in Stress 10 days group and 33.54464 ± 6.45696 % in Stress 21 days group (P = 0.02480, ANOVA). Comparative analysis between groups showed that the percentage of peripherin positive neurons was significantly higher in Stress 21 days group compared to Control group (P = 0.019900, Tukey post hoc test).
The proportion of peripherin highly-positive neurons showed an important increase after stress exposure, from 1.71008 ± 1.44825 % in the Control group to 2.80435 ± 1.84391 % in Stress 1 day group, 6.73242 Stress ± 1.07920 % in Stress 10 days group and 9.99349 ± 3.69508 % in Stress 21 days group (P = 0.00120, Kruskall -Wallis test). Comparative analysis between groups demonstrated a statistically significant increase for Stress 21 days group when compared to Control group (P = 0.008736, two -tailed Multiple Comparisons test) and Stress 1 day group (P = 0.011367, two -tailed Multiple Comparisons test).
4. Discussions
We have investigated the influence of stress on the density of dermal nerve structures with positive immunohistochemical labeling for peripherin and the effects of stress exposure on the expression of peripherin and TRPV1 (VR1) receptor in primary sensory neurons. In addition to previous studies, we analyzed both samples of glabrous and hairy skin, and we investigated the impact of repeated stress exposure.
Stress exposure does not induce major changes in the density of nociceptive nerve structures immunoreactive for peripherin in glabrous and hairy skin, and is associated with only a discrete increase of dermal peripherin positive free nerve endings in the glabrous skin.
In our study we performed an analysis of nociceptive nerve endings as a whole, using peripherin as marker. However, most previous studies that investigated stress impact on nociceptive cutaneous nerve fibers have analyzed the expression of neuropeptides such as SP (Substance P) and CGRP (Calcitonin Gene Related Peptide). Their results have shown that stress increases the number of cutaneous nerve fibers containing SP, a phenomenon that occurs rapidly, in the first 24 hours after stress exposure and appears to be related to a nerve growth process [46, 47]. Regarding the effect of stress on CGRP positive nerve fibers, previous studies have provided conflicting results. A recent study showed that stress exposure is associated with a significant decrease in number of CGRP immunoreactive nerve fibers but in the context of an overall increase of nerve fibers, immunoreactive for PGP (Protein Gene Product) 9.5, in dermis and epidermis of hairy skin [45]. On the other hand, the results of other studies have shown that stress induces an increased CGRP expression in nerve endings [54]. The differences between our findings and previous research data could be related to the different methodology employed, because, although most nociceptive cutaneous nerve fibers contain SP and CGRP [55, 56], there is not a perfect overlapping between nociceptive nerve structures that express SP and CGRP and those immunoreactive for peripherin [41]. In addition, assessment of stress impact on the number of nociceptive nerve structures expressing SP and CGRP can be influenced by the release of neuropeptides from nerve endings, as a direct effect of stress-induced neuronal activation.
The investigation of stress effects on the TRPV1 (VR1) expression in primary sensory neurons showed that stress exposure is associated with a gradual decrease in proportion of TRPV1 (VR1) negative DRG neurons, even though differences between groups were not statistically significant.

CONSTANTIN CARUNTU, DANIEL BODA, SORIN MUSAT, ANA CARUNTU, ELENA POENARU, BOGDAN CALENIC, ILINCA SAVULESCU-FIEDLER, ALINA DRAGHIA, MARIA ROTARU,
ANCA IOANA BADARAU
Romanian Biotechnological Letters, Vol. 19, No. 4, 2014 9526
Stress exposure induced a gradual increase of low-positive TRPV1 (VR1) neurons proportion, significantly higher in rats exposed to stress for 21 days compared to Control. In addition, both short-term and repeated stress exposure led to a marked increase of TRPV1 (VR1) highly-positive neurons.
These results are confirmed by recent investigations, which also showed a stress-induced increase of TRPV1 (VR1) expression in DRGs [49]. The mechanisms by which stress exposure induces an amplification of TRPV1 (VR1) expression in DRG neurons could be related to sympathetic activation and protein kinase pathways [57, 58]. TRPV1 (VR1) is a non-selective cation channel activated by vanilloids such as capsaicin, as well as by heat, acidosis and various endogenous proinflammatory substances. In this respect, TRPV1 acts as molecular integrator for various nociceptive physical and chemical stimuli in primary sensory neurons. [59-63]. Increased expression of TRPV1 (VR1) in primary sensory neurons may be associated with the anterograde transport of this receptor to peripheral terminals, increasing nociceptive sensitivity [64].
Assessment of stress effects on peripherin expression in DRG neurons showed that prolonged stress exposure induces a gradual but significant decline of peripherin negative primary sensory neurons proportion. The proportion of peripherin low-positive neurons was similar in all experimental groups, however, the percentage of peripherin positive neurons was augmented by prolonged stress exposure. Peripherin highly-positive neurons showed also a significant increase after stress exposure.
Peripherin is a marker of nociceptive neurons [39, 40] and an increase of peripherin expression indicates the amplification of nociceptors activity. Although its exact function has not yet been determined, peripherin might be involved in nerve growth and cytoskeletal stabilization [65-68]. Thus, increased expression of this protein could be a marker for nociceptive nerve network growth.
Free nerve endings of nociceptive fibers contain various neuropeptides, such as SP, CGRP [69-71]. Thus, the increased activity of nociceptive nervous structures might be associated with a consequent increased release of proinflamatory neuropeptides in the skin, activating a neurogenic inflammatory process characterized by increased capillary permeability and vasodilation [72-74]. Besides this, stress can play a role in the onset or aggravation of many inflammatory skin diseases such as psoriasis, atopic dermatitis, seborrheic eczema, acne, prurigonodularis, lichen planus, chronic urticaria, rosacea etc. [75-79] an effect that might be explained by the stress-induced activation of cutaneous nociceptive system.
5. Conclusions
Our results have shown an increased nociceptive activity induced by stress exposure and this effect was more prominent in the primary sensory neurons. The impact of stress on nervous structures involved in nociception may explain the role of stress in modulation of both nociceptive sensitivity and skin inflammatory processes.
6. Acknowledgments
Research funded by grant PN-II-RU-TE-2011-3-0249 from the National University Research Council, Romania. Authors wish to thank Professor Leon Zagrean from the Center for Excellence in Neuroscience, “Carol Davila” University of Medicine and Pharmacy, Bucharest and Professor Bogdan Amuzescu from the Dept. Biophysics & Physiology, Faculty of Biology, University of Bucharest for their support during the course of this work.

Stress Effects on Cutaneous Nociceptive Nerve Fibers and Their Neurons of Origin in Rats
9527 Romanian Biotechnological Letters, Vol. 19, No. 4, 2014
References 1. H. SELYE, A syndrome produced by diverse nocuous agents. Nature, 138, 32 (1936). 2. H. SELYE, Forty years of stress research: principal remaining problems and misconceptions. Can. Med.
Assoc. J., 115(1),53, 56 (1976). 3. N. BOLGER, A. DELONGIS, R.C. KESSLER, E.A. SCHILLING, Effects of daily stress on negative
mood. J. Pers. Soc. Psychol., 57(5),808, 818 (1989). 4. J.D. LOESER, R.D. TREEDE, The Kyoto protocol of IASP Basic Pain Terminology. Pain, 137(3),473,
477 (2008). 5. C.D. KING, D.P. DEVINE, C.J. VIERCK, J. RODGERS, R.P. YEZIERSKI, Differential effects of
stress on escape and reflex responses to nociceptive thermal stimuli in the rat. Brain Res., 987(2),214, 222 (2003).
6. H. IMBE, Y. IWAI-LIAO, E. SENBA, Stress-induced hyperalgesia: animal models and putative mechanisms. Front. Biosci.,11,2179, 2192 (2006).
7. S.G. KHASAR, O.A. DINA, P.G. GREEN, J.D. LEVINE, Sound stress-induced long-term enhancement of mechanical hyperalgesia in rats is maintained by sympathoadrenalcatecholamines. J. Pain., 10(10),1073, 1077 (2009).
8. P. CARRIVE, M. CHURYUKANOV, D. LE BARS, A reassessment of stress-induced "analgesia" in the rat using an unbiased method. Pain, 152(3),676, 686 (2011).
9. I.Y.A. ASHKINAZI, E.A. VERSHININA, Pain sensitivity in chronic psychoemotional stress in humans. Neurosci. Behav. Physiol., 29(3),333, 337 (1999).
10. T. HATA, T. KITA, E. ITOH, A. KAWABATA, The relationship of hyperalgesia in SART (repeated cold)-stressed animals to the autonomic nervous system. J. Auton. Pharmacol.,8(1),45, 52 (1988).
11. G.D. GAMARO, M.H. XAVIER, J.D. DENARDIN, J.A. PILGER, D.R. ELY, M.B. FERREIRA, C. DALMAZ, The effects of acute and repeated restraint stress on the nociceptive response in rats. Physiol. Behav., 63(4),693, 697 (1998).
12. L. QUINTERO, M. MORENO, C. AVILA, J. ARCAYA, W. MAIXNER, H. SUAREZ-ROCA, Long-lasting delayed hyperalgesia after subchronic swim stress. Pharmacol. Biochem. Behav., 67(3),449, 458 (2000).
13. C. RIVAT, C. BECKER, A. BLUGEOT, B. ZEAU, A. MAUBORGNE, M. POHL, J.J. BENOLIEL, Chronic stress induces transient spinal neuroinflammation, triggering sensory hypersensitivity and long-lasting anxiety-induced hyperalgesia. Pain, 150(2),358, 368 (2010).
14. S.G. KHASAR, J. BURKHAM, O.A. DINA, A.S. BROWN, O. BOGEN, N. ALESSANDRI-HABER, P.G. GREEN, D.B. REICHLING, J.D. LEVINE, Stress induces a switch of intracellular signaling in sensory neurons in a model of generalized pain. J. Neurosci., 28(22),5721, 5730 (2008).
15. D.B. REICHLING, J.D. LEVINE, Critical role of nociceptor plasticity in chronic pain. Trends Neurosci., 32(12),611, 618 (2009).
16. P.G. GREEN, X. CHEN, P. ALVAREZ, L.F. FERRARI, J.D. LEVINE, Early-life stress produces muscle hyperalgesia and nociceptor sensitization in the adult rat. Pain, 152(11),2549, 2556 (2011).
17. N. SHANKAR, N. AWASTHY, H. MAGO, O.P. TANDON, Analgesic effect of environmental noise: a possible stress response in rats. Indian J. Physiol. Pharmacol., 43(3),337, 346 (1999).
18. H. LOGAN, S. LUTGENDORF, P. RAINVILLE, D. SHEFFIELD, K. IVERSON, D. LUBAROFF, Effects of stress and relaxation on capsaicin-induced pain. J. Pain, 2(3),160, 170 (2001).
19. A. JONES, H. SPINDLER, M.M. JØRGENSEN, R. ZACHARIAE, The effect of situation-evoked anxiety and gender on pain report using the cold pressor test. Scand. J. Psychol., 43(4),307, 313 (2002).
20. L.E. CARTER, D.W. MCNEIL, K.E. VOWLES, J.T. SORRELL, C.L. TURK, B.J. RIES, D.R. HOPKO, Effects of emotion on pain reports, tolerance and physiology. Pain Res. Manag., 7(1),21, 30 (2002).
21. S AMIR, Z AMIT, Endogenous opioid ligands may mediate stress-induced changes in the affective properties of pain related behavior in rats. Life Sci., 23(11),1143, 1151 (1978).
22. R.J. BODNAR, D.D. KELLY, M. BRUTUS, M. GLUSMAN, Stress-induced analgesia: neural and hormonal determinants. Neurosci. Biobehav. Rev., 4(1),87, 100 (1980).
23. D.J. CALCAGNETTI, S.G. HOLTZMAN, Potentiation of morphine analgesia in rats given a single exposure to restraint stress immobilization. Pharmacol. Biochem. Behav.,41(2),449, 453 (1992).
24. D.J. CALCAGNETTI, J.L. STAFINSKY, T. CRISP, A single restraint stress exposure potentiates analgesia induced by intrathecally administered DAGO. Brain Res., 592(1-2),305, 309 (1992).
25. W.T. CHANCE, G.M. KRYNOCK, J.A. ROSECRANS, Antinociception following lesion-induced hyperemotionality and conditioned fear. Pain, 4(3),243, 252 (1978).
26. H.S. CHEN, F.P. LI, X.Q. LI, B.J. LIU, F. QU, W.W. WEN, Y. WANG, Q. LIN, Acute stress regulates nociception and inflammatory response induced by bee venom in rats: possible mechanisms. Stress,

CONSTANTIN CARUNTU, DANIEL BODA, SORIN MUSAT, ANA CARUNTU, ELENA POENARU, BOGDAN CALENIC, ILINCA SAVULESCU-FIEDLER, ALINA DRAGHIA, MARIA ROTARU,
ANCA IOANA BADARAU
Romanian Biotechnological Letters, Vol. 19, No. 4, 2014 9528
16(5),557, 563 (2013). 27. G.B. CHESHER, B. CHAN, Footshock induced analgesia in mice: its reversal by naloxone and cross
tolerance with morphine. Life Sci., 21(11),1569, 1574 (1977). 28. H.S. HAGEN, K.F. GREEN, Effects of time of testing, stress level, and number of conditioning days on
naloxone sensitivity of conditioned stress-induced analgesia in rats. Behav. Neurosci.,102(6),906, 914 (1988).
29. R.L. HAYES, G.J. BENNETT, P.G. NEWLON, D.J. MAYER, Behavioral and physiological studies of non-narcotic analgesia in the rat elicited by certain environmental stimuli. Brain Res., 155(1),69, 90 (1978).
30. F.J. HELMSTETTER, M.S. FANSELOW, Strain differences in reversal of conditional analgesia by opioid antagonists. Behav. Neurosci.,101(5),735, 737 (1987).
31. J.W. LEWIS, J.T. CANNON, J.C. LIEBESKIND, Opioid and nonopioid mechanisms of stress analgesia. Science, 208(4444),623, 625 (1980).
32. J. 4TH MADDEN, H. AKIL, R.L. PATRICK, J.D. BARCHAS, Stress-induced parallel changes in central opioid levels and pain responsiveness in the rat. Nature, 265(5592),358, 360 (1977).
33. C.A. PORRO, G. CARLI, Immobilization and restraint effects on pain reactions in animals. Pain, 32(3),289, 307 (1988).
34. L.R. WATKINS, D.J. MAYER, Organization of endogenous opiate and nonopiate pain control systems. Science, 216(4551),1185, 1192 (1982).
35. R.K. BUTLER, D.P. FINN, Stress-induced analgesia. Prog. Neurobiol.,88(3),184, 202 (2009). 36. C. VIDAL, J.J. JACOB, Stress hyperalgesia in rats: an experimental animal model of
anxiogenichyperalgesia in human. Life Sci., 31(12-13),1241, 1244 (1982). 37. E. JØRUM, Analgesia or hyperalgesia following stress correlates with emotional behavior in rats. Pain,
2(3),341, 348 (1988). 38. E. JØRUM, Noradrenergic mechanisms in mediation of stress-induced hyperalgesia in rats. Pain,
32(3),349, 355 (1988). 39. K.H. CHEN, C.H. YANG, J.T. CHENG, C.H. WU, W.D. SY, C.R. LIN, Altered neuronatin expression
in the rat dorsal root ganglion after sciatic nerve transection. J. Biomed. Sci., 17(1),41 (2010). 40. S. HONG, J.W. WILEY, Early painful diabetic neuropathy is associated with differential changes in the
expression and function of vanilloid receptor 1. J. Biol. Chem., 280(1),618, 627 (2005). 41. M.E. GOLDSTEIN, S.B. HOUSE, H. GAINER, NF-L and peripherin immunoreactivities define
distinct classes of rat sensory ganglion cells. J. Neurosci. Res., 30(1):92-104 (1991). 42. M. TOMINAGA, M.J. CATERINA, Thermosensation and pain. J. Neurobiol., 61(1),3, 12 (2004). 43. Q.P. MA, Expression of capsaicin receptor (VR1) by myelinated primary afferent neurons in rats.
Neurosci. Lett.,319(2),87, 90 (2002). 44. S. KAWANA, Z. LIANG, M. NAGANO, H. SUZUKI, Role of substance P in stress-derived
degranulation of dermal mast cells in mice. J. Dermatol. Sci., 42(1),47, 54 (2006). 45. C.E. KLEYN, L. SCHNEIDER, R. SARACENO, C. MANTOVANI, H.L. RICHARDS, D.G.
FORTUNE, M. CUMBERBATCH, R.J. DEARMAN, G. TERENGHI, I. KIMBER, C.E. GRIFFITHS, The effects of acute social stress on epidermal Langerhans' cell frequency and expression of cutaneous neuropeptides. J. Invest. Dermatol., 128(5),1273, 1279 (2008).
46. P.C. ARCK, B. HANDJISKI, E.M. PETERS, A.S. PETER, E. HAGEN, A. FISCHER, B.F. KLAPP, R. PAUS, Stress inhibits hair growth in mice by induction of premature catagen development and deleterious perifollicular inflammatory events via neuropeptide substance P-dependent pathways. Am. J. Pathol., 162(3),803, 814 (2003).
47. E.M. PETERS, A. KUHLMEI, D.J. TOBIN, S. MÜLLER-RÖVER, B.F. KLAPP, P.C. ARCK, Stress exposure modulates peptidergic innervation and degranulates mast cells in murine skin. Brain Behav. Immun., 19(3),252, 262 (2005).
48. S. PAVLOVIC, M. DANILTCHENKO, D.J. TOBIN, E. HAGEN, S.P. HUNT, B.F. KLAPP, P.C. ARCK, E.M. PETERS, Further exploring the brain-skin connection: stress worsens dermatitis via substance P-dependent neurogenic inflammation in mice. J. Invest. Dermatol., 128(2),434, 446 (2008).
49. Y.B. YU, J. YANG, X.L. ZUO, L.J. GAO, P. WANG, Y.Q. LI, Transient receptor potential vanilloid-1 (TRPV1) and ankyrin-1 (TRPA1) participate in visceral hyperalgesia in chronic water avoidance stress rat model. Neurochem. Res., 35(5),797, 803 (2010).
50. F. BERKENBOSCH, D.A. WOLVERS, R. DERIJK, Neuroendocrine and immunological mechanisms in stress-induced immunomodulation. J. Steroid. Biochem. Mol. Biol., 40(4-6),639, 647 (1991).
51. G.B. GLAVIN, W.P. PARÉ, T. SANDBAK, H.K. BAKKE, R. MURISON, Restraint stress in

Stress Effects on Cutaneous Nociceptive Nerve Fibers and Their Neurons of Origin in Rats
9529 Romanian Biotechnological Letters, Vol. 19, No. 4, 2014
biomedical research: an update. Neurosci. Biobehav. Rev., 18(2),223, 249 (1994). 52. F.S. DHABHAR, B.S. MCEWEN, Acute stress enhances while chronic stress suppresses cell-mediated
immunity in vivo: a potential role for leukocyte trafficking. Brain. Behav. Immun., 11(4),286, 306 (1997).
53. S. MUSAT-MARCU, H.E. GUNTER, B.I. JUGDUTT, J.C. DOCHERTY, Inhibition of apoptosis after ischemia-reperfusion in rat myocardium by cycloheximide. J. Mol. Cell. Cardiol., 31(5),1073, 1082 (1999).
54. Y. KAWAGUCHI, T. OKADA, H. KONISHI, M. FUJINO, J. ASAI, M. ITO, Reduction of the DTH response is related to morphological changes of Langerhans cells in mice exposed to acute immobilization stress. Clin. Exp. Immunol., 109(2),397, 401 (1997).
55. P. HOLZER, Local effector functions of capsaicin-sensitive sensory nerve endings: involvement of tachykinins, calcitonin gene-related peptide and other neuropeptides. Neuroscience, 24(3),739, 768 (1988).
56. J.D. RICHARDSON, M.R. VASKO, Cellular mechanisms of neurogenic inflammation. J. Pharmacol. Exp. Ther., 302(3),839, 845 (2002).
57. X. XU, P. WANG, X. ZOU, D. LI, L. FANG, Q. LIN, Increases in transient receptor potential vanilloid-1 mRNA and protein in primary afferent neurons stimulated by protein kinase C and their possible role in neurogenic inflammation. J. Neurosci. Res., 87(2),482, 494 (2009).
58. X. XU, P. WANG, X. ZOU, D. LI, L. FANG, K. GONG, Q. LIN, The effects of sympathetic outflow on upregulation of vanilloid receptors TRPV(1) in primary afferent neurons evoked by intradermal capsaicin. Exp. Neurol., 222(1),93, 107 (2010).
59. M.J. CATERINA, M.A. SCHUMACHER, M. TOMINAGA, T.A. ROSEN, J.D. LEVINE, D. JULIUS, The capsaicin receptor: a heat-activated ion channel in the pain pathway. Nature, 389(6653),816, 824 (1997).
60. A. SZALLASI, P.M. BLUMBERG, Vanilloid (Capsaicin) receptors and mechanisms. Pharmacol. Rev., 51(2),159, 212 (1999).
61. E. BODÓ, T. BÍRÓ, A. TELEK, G. CZIFRA, Z. GRIGER, B.I. TÓTH, A. MESCALCHIN, T. ITO, A. BETTERMANN, L. KOVÁCS, R. PAUS, A hot new twist to hair biology: involvement of vanilloid receptor-1 (VR1/TRPV1) signaling in human hair growth control. Am. J. Pathol., 166(4),985, 998 (2005).
62. K. BÖLCSKEI, Z. HELYES, A. SZABÓ, K. SÁNDOR, K. ELEKES, J. NÉMETH, R. ALMÁSI, E. PINTÉR, G. PETHO, J. SZOLCSÁNYI, Investigation of the role of TRPV1 receptors in acute and chronic nociceptive processes using gene-deficient mice. Pain, 117(3),368, 376 (2005).
63. B.A. CHIZH, M.B. O'DONNELL, A. NAPOLITANO, J. WANG, A.C. BROOKE, M.C. AYLOTT, J.N. BULLMAN, E.J. GRAY, R.Y. LAI, P.M. WILLIAMS, J.M. APPLEBY, The effects of the TRPV1 antagonist SB-705498 on TRPV1 receptor-mediated activity and inflammatory hyperalgesia in humans. Pain, 132(1-2),132, 141 (2007).
64. R.R. JI, T.A. SAMAD, S.X. JIN, R. SCHMOLL, C.J. WOOLF, p38 MAPK activation by NGF in primary sensory neurons after inflammation increases TRPV1 levels and maintains heat hyperalgesia. Neuron, 36(1),57, 68 (2002).
65. M.M. OBLINGER, J. WONG, L.M. PARYSEK, Axotomy-induced changes in the expression of a type III neuronal intermediate filament gene. J. Neurosci., 9(11),3766, 3775 (1989).
66. M. ESCURAT, K. DJABALI, M. GUMPEL, F. GROS, M.M. PORTIER, Differential expression of two neuronal intermediate-filament proteins, peripherin and the low-molecular-mass neurofilament protein (NF-L), during the development of the rat. J. Neurosci., 10(3),764, 784 (1990).
67. R.C. LARIVIÈRE, M.D. NGUYEN, A. RIBEIRO-DA-SILVA, J.P. JULIEN, Reduced number of unmyelinated sensory axons in peripherin null mice. J. Neurochem., 81(3),525, 532 (2002).
68. K.S. ERIKSSON, S. ZHANG, L. LIN, R.C. LARIVIÈRE, J.P. JULIEN, E. MIGNOT, The type III neurofilament peripherin is expressed in the tuberomammillary neurons of the mouse. BMC Neurosci., 9,26 (2008).
69. J.C. ANSEL, C.A. ARMSTRONG, I. SONG, K.L. QUINLAN, J.E. OLERUD, S.W. CAUGHMAN, N.W. BUNNETT, Interactions of the skin and nervous system. J. Investig. Dermatol. Symp. Proc., 2(1),23, 26 (1997).
70. T. SCHOLZEN, C.A. ARMSTRONG, N.W. BUNNETT, T.A. LUGER, J.E. OLERUD, J.C. ANSEL, Neuropeptides in the skin: interactions between the neuroendocrine and the skin immune systems. Exp. Dermatol., 7(2-3),81, 96 (1998).
71. D. ROOSTERMAN, T. GOERGE, S.W. SCHNEIDER, N.W. BUNNETT, M. STEINHOFF, Neuronal control of skin function: the skin as a neuroimmunoendocrine organ. Physiol. Rev., 86(4),1309, 1379 (2006).

CONSTANTIN CARUNTU, DANIEL BODA, SORIN MUSAT, ANA CARUNTU, ELENA POENARU, BOGDAN CALENIC, ILINCA SAVULESCU-FIEDLER, ALINA DRAGHIA, MARIA ROTARU,
ANCA IOANA BADARAU
Romanian Biotechnological Letters, Vol. 19, No. 4, 2014 9530
72. N. JANCSÓ, A. JANCSÓ-GÁBOR, J. SZOLCSÁNYI, Direct evidence for neurogenic inflammation and its prevention by denervation and by pretreatment with capsaicin. Br. J. Pharmacol. Chemother., 31(1),138, 151 (1967).
73. L.F. CHAPMAN, Mechanisms of the flare reaction in human skin. J. Invest. Dermatol.,69(1),88, 97 (1977).
74. C. C RUNTU, D. BODA, Evaluation through in vivo reflectance confocal microscopy of the cutaneous neurogenic inflammatory reaction induced by capsaicin in human subjects. J. Biomed. Opt., 17(8),085003 (2012).
75. J. KOO, A. LEBWOHL, Psycho dermatology: the mind and skin connection. Am. Fam. Physician, 64(11),1873, 1878 (2001).
76. P. ARCK, R. PAUS, From the brain-skin connection: the neuroendocrine-immune misalliance of stress and itch. Neuroimmunomodulation,13(5-6),347, 356 (2006).
77. F. TAUSK, I. ELENKOV, J. MOYNIHAN, Psychoneuroimmunology. Dermatol. Ther., 21(1),22, 31 (2008).
78. C. C RUNTU, M.A. GHI , A. C RUNTU, D. BODA, The role of stress in the multifactorial etiopathogenesis of acne. Ro. Med. J., 58(2),98, 101 (2011).
79. C. C RUNTU, C. GRIGORE, A. C RUNTU, A. DIACONEASA, D. BODA, The role of stress in skin diseases. Internal Medicine-Medicina Intern , 8(3),73, 84 (2011).





























