Embed Size (px)
Citation preview

Exp Brain Res (2005) 160: 38–59DOI 10.1007/s00221-004-1984-0
RESEARCH ARTICLES
Steven T. Moore . Bernard Cohen . Theodore Raphan .Alain Berthoz . Gilles Clément
Spatial orientation of optokinetic nystagmus and ocular pursuitduring orbital space flight
Received: 10 November 2003 / Accepted: 10 May 2004 / Published online: 28 July 2004# Springer-Verlag 2004
Abstract On Earth, eye velocity of horizontal optokineticnystagmus (OKN) orients to gravito-inertial acceleration(GIA), the sum of linear accelerations acting on the headand body. We determined whether adaptation to micro-gravity altered this orientation and whether ocular pursuitexhibited similar properties. Eye movements of fourastronauts were recorded with three-dimensional video-oculography. Optokinetic stimuli were stripes movinghorizontally, vertically, and obliquely at 30°/s. Ocularpursuit was produced by a spot moving horizontally orvertically at 20°/s. Subjects were either stationary or werecentrifuged during OKN with 1 or 0.5 g of interaural ordorsoventral centripetal linear acceleration. Average eyeposition during OKN (the beating field) moved into thequick-phase direction by 10° during lateral and upwardfield movement in all conditions. The beating field did notshift up during downward OKN on Earth, but there was astrong upward movement of the beating field (9°) duringdownward OKN in the absence of gravity; this likely
represents an adaptation to the lack of a vertical 1-g biasin-flight. The horizontal OKN velocity axis tilted 9° in theroll plane toward the GIA during interaural centrifugation,both on Earth and in space. During oblique OKN, thevelocity vector tilted towards the GIA in the roll planewhen there was a disparity between the direction of stripemotion and the GIA, but not when the two were aligned.In contrast, dorsoventral acceleration tilted the horizontalOKN velocity vector 6° in pitch away from the GIA. Rolltilts of the horizontal OKN velocity vector toward the GIAduring interaural centrifugation are consistent with theorientation properties of velocity storage, but pitch tiltsaway from the GIAwhen centrifuged while supine are not.We speculate that visual suppression during OKN mayhave caused the velocity vector to tilt away from the GIAduring dorsoventral centrifugation. Vertical OKN andocular pursuit did not exhibit orientation toward the GIAin any condition. Static full-body roll tilts and centrifuga-tion generating an equivalent interaural accelerationproduced the same tilts in the horizontal OKN velocitybefore and after flight. Thus, the magnitude of tilt in OKNvelocity was dependent on the magnitude of interaurallinear acceleration, rather than the tilt of the GIA withregard to the head. These results favor a ‘filter’ model ofspatial orientation in which orienting eye movements areproportional to the magnitude of low frequency interaurallinear acceleration, rather than models that postulate aninternal representation of gravity as the basis for spatialorientation.
Keywords Microgravity . OKN . Otoliths . Smoothpursuit . Velocity storage
Abbreviations Ag: Acceleration of gravity . Ac:Centripetal acceleration . CCW: Counterclockwise . CW:Clockwise . FD- X: Flight day X . g: Gravity . GIA:Gravito-inertial acceleration . H: Horizontal . LED: Left-ear-down . LEO: Left-ear-out . LOB: Lying-on-back . L-X: Launch minus X days . NCM: No-chair-motion . ND:Nose-down . NU: Nose-up . OCR: Ocular counter-colling . OKAN: Optokinetic after-nystagmus . OKN:
S. T. Moore (*) . B. CohenDepartment of Neurology, Mount Sinai School of Medicine,1 E 100th St.,New York, NY 10029, USAe-mail: [email protected].: +1-212-2419306Fax: +1-212-8311610
B. CohenDepartment of Physiology and Biophysics, Mount Sinai Schoolof Medicine,New York, NY, USA
T. RaphanDepartment of Computer and Information Science, BrooklynCollege,Brooklyn, NY, USA
A. BerthozLaboratoire de Physiologie de la Perception et de l’Action,Collège de France,Paris, France
G. ClémentCentre de Recherche Cerveau et Cognition, CNRS/UPS,Toulouse, France

Optokinetic nystagmus . OKS: Optokinetic stimulus . pos:Position . REO: Right-ear-out . R+ X: Recovery plus Xdays . T: Torsional . V: Vertical . vel: Velocity
Introduction
On Earth, the pull of gravity, experienced as an equivalentupward linear acceleration of 9.8 m/s2, is fundamental forestablishing spatial orientation. Due to the universality ofgravity on Earth, however, it is difficult to determine itsprecise role relative to other factors such as transient linearaccelerations during turns (Imai et al. 2001) and theinternal sense of the body vertical (Clément et al. 2001).During orbital spaceflight, the acceleration of gravity isalmost exactly counterbalanced by the radial centripetalacceleration generated by circling the Earth at a highangular velocity. This results in microgravity (10−6 g)within the spacecraft, which is a unique environment inwhich to study the components contributing to spatialorientation. Prolonged exposure to microgravity results indeficits in posture, gaze control and orthostatic toleranceupon return to Earth (for review see Clément and Reschke1996; Moore et al. 2004). The underlying causes for thesedeficits are not well understood, but imply adaptivechanges in the body’s response to linear acceleration (seeMoore et al. 2001, 2003, 2004). In 1998, we performed aseries of experiments aboard the orbiter Columbia as partof the Neurolab (STS-90) mission. The goal was to studythe effects of adaptation to microgravity on vestibularfunction and spatial orientation. Here we describeexperiments relating to the effect of microgravity exposureon spatial orientation of the eye velocity of optokineticnystagmus (OKN) and of smooth pursuit. Effects of orbitalspaceflight on ocular counter-rolling (OCR) and percep-tion of the upright in the same astronauts have beenreported previously (Clément et al. 2001; Moore et al.2001).
The optokinetic reflex, widely distributed across spe-cies, responds to movement of the visual surround toproduce OKN and optokinetic after-nystagmus (OKAN)(Purkinje 1820; Ter Braak 1936; Collewijn 1969; Hoodand Leech 1974; Cohen et al. 1977). The response tomovement of the visual field has been modeled in humansand monkeys by two components (Cohen et al. 1977;Robinson 1977; Raphan et al. 1979). A fast componentgenerates a short latency response that stabilizes gazeduring rapid changes in surround velocity due to highfrequency movements of the head or visual surround. Aslow component stabilizes visual following and produceseye velocities that can long outlast the stimulus, generatingOKAN (Ter Braak 1936; Cohen et al. 1977). This slowcomponent has been modeled by an integrator with a timeconstant that ‘stores’ activity related to surround motion inlight and discharges slowly in darkness (Cohen et al.1977). Termed ‘velocity storage’, this integrative processis common to both OKN and to the angular vestibulo-ocular reflex (VOR) (Raphan et al. 1979). An importantaspect of velocity storage is its spatial orientation (Harris
1987; Dai et al. 1991; Raphan and Sturm 1991; Angelakiand Hess 1994, 1995), which tends to orient the axis ofeye velocity during horizontal OKN (Raphan and Cohen1988; Raphan and Sturm 1991; Dai et al. 1991; Gizzi et al.1994) or horizontal vestibular nystagmus (Harris 1987;Harris and Barnes 1987; Angelaki and Hess 1994, 1995)towards gravito-inertial acceleration (GIA), the vector sumof all linear accelerations, including gravity, that act on thehead (for review see Raphan and Cohen 2002).
There is evidence that spatial orientation of eye velocityto the GIA might not be maintained after adaptation tomicrogravity. Two of four monkeys that flew on RussianCOSMOS missions in 1988 and 1993 for periods up to2 weeks exhibited changes consistent with a shift of theorientation properties of velocity storage from the GIAtowards the body vertical. Just after flight, one monkeylost the ability to modify the time constant of post-rotatory(vestibular) nystagmus when it was tilted out of the planeof the ongoing nystagmus (Cohen et al. 1991). Termed ‘tiltdumping’ (Benson 1974; Raphan et al. 1981; Fetter et al.1996), this is part of the process that helps align the axis ofhorizontal eye velocity to the spatial vertical for bothvestibular nystagmus and OKAN (Dai et al. 1991; Merfeldet al. 1993). In a second animal, the axis of horizontalOKAN velocity, which had aligned with the spatialvertical during roll tilts with regard to gravity beforeflight, was shifted 28° toward the long body axis afterflight (Dai et al. 1994). There were also long-lasting post-flight decrements in OCR and vergence in response tointeraural and naso-occipital linear accelerations in thesemonkeys (Dai et al. 1994, 1996).
The first aim of the experiments was to determinewhether the spatial orientation of the eye velocity ofhorizontal OKN, which exhibits a tendency to orienttowards the GIA on Earth, shifted towards a bodycoordinate frame in microgravity. Some evidence for achange to a body reference frame was observed in theCOSMOS monkey data described above, but more recentOCR and perception data from the Neurolab astronautssuggested that spatial orientation maintained its orientationto the GIA on orbit (Clément et al. 2001; Moore et al.2001). We also wished to confirm previous findings on theorientation of human horizontal OKN to gravity duringstatic roll tilts of the head (Gizzi et al. 1994) using a moreadvanced measurement technique, i.e., 3-D video-oculog-raphy, and to determine whether there was orientation ofOKN towards the GIA in the pitch (sagittal) plane duringlinear acceleration along the Gz (dorsoventral) axis. Suchorientation in pitch occurred during OKAN in the monkey,but the velocity axis had tilted away from the GIA duringOKN (Raphan and Cohen 1988). Whether horizontalOKN velocity orients to tilts of the GIA relative to thehead in the pitch plane has not been studied in humans.
How the central nervous system incorporates theconstant linear acceleration of gravity into spatial orien-tation is a critical question. A number of Earth-basedinvestigations have hypothesized that the vestibularsystem can distinguish between translation of the headalong interaural or naso-occipital axes, and tilts of the head
39

with regard to the spatial vertical, both of which impose alinear acceleration in the head-horizontal plane (Mayne1974; Paige 1991; Merfeld 1995; Merfeld and Young1995; Hess and Angelaki 1999; Merfeld et al. 1999;Angelaki et al. 1999, 2001; Paige and Seidman 1999); forreview see Raphan and Cohen (2002). Based on thishypothesis, models have been formulated combining aninternal estimate of gravity with internal models ofsemicircular canal dynamics, neurodynamics that mimicthe physics of motion, and the GIA, to separate head tiltsrelative to gravity from linear accelerations generated bytranslation (Merfeld 1995; Merfeld and Young 1995;Angelaki et al. 1999, 2000). This internal gravity estimatehas also been postulated as a critical vector governingperception of spatial orientation and its associated eye,body and limb movements (Merfeld 1995; Merfeld andYoung 1995; Angelaki et al. 1999, 2000). Models of thelinear vestibulo-ocular reflex (lVOR) have been formu-lated that lead to simpler realizations of how the centralvestibular system might separate tilts from translation ofthe head, without requiring an internal estimate of gravity.These models transform tilts of the head relative to theGIA about a roll axis into a low frequency orientingcommand related directly to the magnitude of interaurallinear acceleration (Raphan et al. 1996; Seidman et al.1998; Paige and Seidman 1999). These are referred to hereas the ‘filter’ models.
A key difference between the ‘internal gravity estimate’and ‘filter’ models is the role that gravity plays indetermining orientation. Models that rely on an internalestimate of gravity would be devoid of a spatial referencefor generating orienting responses in the absence ofgravity, leading to the ‘reinterpretation’ of low-frequencylinear acceleration as translation. Thus, orientation wouldbe linked to the body/head rather than a spatial frame, andconsequently, ocular responses would not exhibit spatialorientation. An empirical formulation of this premise,based on post-flight perceptual reports of astronauts, is the‘otolith-tilt-translation-reinterpretation’ (OTTR) hypothe-sis (Young et al. 1984; Parker et al. 1985). This posits thatafter adaptation to microgravity all afferent otolithinformation would be interpreted as translation. On returnto Earth, therefore, pitch and roll tilts of the head wouldinitially be perceived as naso-occipital and lateral trans-lation, respectively. In the ‘filter’ models, the interaurallinear acceleration component determines orientation inthe roll plane, and there would be an equivalent responseto the combined GIA vector whether or not it contained agravitational component. Findings from the Neurolabflight, demonstrating that OCR and perception of theupright remained oriented to the GIA during constantvelocity centrifugation in microgravity, support the filtermodels (Clément et al. 2001; Moore et al. 2001, 2003).The space experiments described in this study provided anadditional opportunity to test model predictions duringOKN. They also enabled a test of a basic empiricalassumption of the Raphan/Sturm model of spatial orien-tation of velocity storage, namely that cross-coupling onlyoccurs for horizontal eye velocity and that there is no
vertical-to-horizontal or torsional-to-horizontal cross-cou-pling (Dai et al. 1991; Raphan and Sturm 1991). This hasbeen verified in monkeys (Raphan and Cohen 1988; Dai etal. 1991) but not in humans.
On Earth, the otoliths produce static orientation of theeyes toward the GIA through changes in horizontal,vertical, and torsional eye position, but the otoliths canalso orient eye velocity through kinematic mechanisms(Helmholz 1867). For example, if the eyes are positionedvertically during rotatory yaw stimulation, there is a pitchtilt of the eye velocity vector during ocular pursuit andsaccades that obeys Listing’s Law (Tweed and Vilis 1987,1990; Crawford and Vilis 1991; Haslwanter et al. 1991).Vertical or horizontal repositioning of the eyes during rollstimulation also tilts the eye velocity vector in thedirection of gaze (Misslisch et al. 1994). Extensions ofthis idea, using a pulley model of the ocular muscles,would predict a tilt of the eye velocity axis in the roll planeas a result of changes in static torsional position of theeyes (Raphan 1998). Thus, the otoliths, by repositioningthe eyes in response to head orientation relative to theGIA, could orient eye velocity towards the GIA through apurely kinematic effect. Changes in mean eye position (thebeating field) during vertical OKN have been noted duringprevious microgravity studies. Average eye positionmoved down more strongly during upward OKN for thefirst 2 days during flight, with a post-flight reversal in thedirection of shift (Clément et al. 1992a, b). Therefore, itwas of interest to determine if changes in the OKN beatingfield were also present on orbit, and how these kinematiceffects on eye velocity might be manifest during OKN,either on Earth or in space.
The final component of this experiment involved theeffect of microgravity, and of tilts of the GIA with respectto the head, on smooth pursuit eye movements. The fastcomponent of OKN, which is produced over many of thesame pathways as ocular pursuit, is in head-fixedcoordinates and does not exhibit spatial orientation(Wearne et al. 1997; see also Yakushin et al. (2000) forreview). Thus it appeared likely that this would also holdtrue for visual tracking of a target.
In order to perform these experiments, a number of‘firsts’ were required. Two human centrifuges weredeveloped for the Neurolab mission for in-flight andground-based testing (Clément et al. 2001; Moore et al.2001). This was the first use of optokinetic stimulationduring centrifugation, either on Earth or in space, toobserve the effects of GIA tilts on the eye velocity axis ofOKN in humans. Furthermore, the generation of sustained1-g linear acceleration was the first application of‘artificial gravity’ during manned space flight, and enabledus to evaluate the orientation properties of the VOR, OKNand ocular pursuit during orbital spaceflight.
40

Methods
Subjects
Experiments were carried out on the National Aeronautics andSpace Administration (NASA) Neurolab STS-90 mission, whichtook place on the orbiter Columbia from 17 April to 2 May 1998.The equipment was installed inside the Spacelab module, which wasa reusable laboratory developed by the European Space Agency,mounted in the shuttle cargo bay. The study was approved by theInstitutional Review Boards at Johnson Space Center in Houstonand the Mount Sinai School of Medicine and was performed inaccordance with the ethical standards of the 1964 Declaration ofHelsinki. Subjects gave informed consent prior to their inclusion inthe study. Four payload crewmembers served both as subjects andin-flight operators. The astronauts were exposed to 1 g of interauralor dorsoventral centripetal acceleration during centrifugation 90, 60,and 15 days prior to launch (L-90, L-60, and L-15), five timesduring flight (Flight Days (FD) 2, 5, 10, 11, and 16), and three timesafter flight (24 h after recovery [R+1] and on R+2 and R+9). Allfour astronauts were also exposed to 0.5-g centrifugation on L-30and R+4 and two received 0.5-g centrifugation in-flight on FD 7 andFD 12. In addition, using the tilt capability of the ground centrifuge,OKN and ocular pursuit were also measured in the four subjectsduring static roll tilt on L-90, L-60, L-30 and R+1, R+4 and R+9,and in a static tilt chair at Kennedy Space Center before flight (L-2)and within 4 h of landing (R+0).
Coordinate frames
A head-fixed coordinate frame {Xh, Yh, Zh} was defined such thatXh was parallel to the naso-occipital axis (positive forward), Yh wasparallel to the interaural axis (positive left), and Zh was normal to theXh-Yh plane (positive upwards). The origin of the head coordinateframe was the intersection of these axes, located on the interauralaxis at a point midway between the vestibular labyrinths. The Xh-Yhplane is equivalent to the stereotaxic horizontal plane. The Xh, Yh,Zh axes are the roll, pitch, and yaw axes of the head, respectively.According to the right-hand rule, the optokinetic stimulus velocityaxis was directed along the positive Zh axis for leftward stimulationand along the negative Zh axis for rightward stimulation. Thestimulus axis was along the positive Yh axis for downward stripemovement and along the negative Yh axis during upward stimuli.On Earth, the acceleration of gravity was aligned with the Zh axiswhen sitting upright and with the Xh axis when supine. An eye-fixedcoordinate frame {Xe, Ye, Ze} was also defined with the origin at thecenter of the eye and Xe passing through the center of the pupil(positive forward) and normal to Ye-Ze plane. Eye rotations in thehead were defined as the relative rotations of the eye coordinate axeswith respect to the head coordinate axes. Positive directions forhorizontal, vertical and torsional eye movements were to the left,down and clockwise from the subject’s point of view.
Centrifuges
Two centrifuges, a flight (European Space Agency) and a ground-based model (Neurokinetics Inc., Pittsburgh, PA, USA), wereutilized in this experiment (for a detailed description of thecentrifuges see Clément et al. 2001; Moore et al. 2001). The two
Fig. 1a–c Centrifuge configurations used for the Neurolab (STS-90) mission. a Left-Ear-Out (LEO) b Right-Ear-Out (REO) c Lying-on-Back (LOB). On Earth, a 1-g centripetal acceleration, Ac, sumswith the acceleration of gravity, Ag, to create a GIA vector of
magnitude 1.4-g tilted 45° with respect to the head. A 0.5-gcentripetal acceleration generates a GIA of magnitude 1.1 g tilted27°. In space, the gravitational vector Ag is negligible and the GIA isequivalent to the 1- or 0.5-g centripetal acceleration
41

machines were functionally identical except that the groundcentrifuge allowed subjects to be statically tilted in roll. Subjectssat upright with their long body (Zh) axis parallel to the axis ofrotation with either their left-ear-out (Fig. 1a) or right-ear-out(Fig. 1b) from the rotation axis, with the centripetal accelerationdirected along the interaural (Yh) axis (Gy centrifugation). In thelying-on-back configuration (Fig. 1c) subjects were supine with therotation axis through the lumbar region, and the centripetalacceleration was directed along the dorsoventral (Zh) axis (-Gzcentrifugation). During Gy centrifugation, the midpoint between thetwo labyrinths was 50 cm from the axis of rotation (Fig. 1a, b).When lying-on-back for -Gz centrifugation, the interaural axis was65 cm from the rotation axis (Fig. 1c). A 5-point harness as well asthigh, shoulder and neck pads and a knee strap held the body firmlyin place. A custom-made facemask, consisting of a fiberglass frontand back shell that were molded to the bony features of eachsubject’s skull, restrained the head. The mask was oriented so thatthe stereotaxic horizontal (Xh-Yh) plane was orthogonal to both thegravitational vertical (on Earth) and the axis of rotation whenupright in the left- or right-ear-out positions, and parallel to the axisof rotation when lying-on-back.Constant velocity rotations of 254 and 180°/s during Gy
centrifugation produced centripetal accelerations of 1 and 0.5 g atthe midpoint between the labyrinths, directed along the interaural(Yh) axis. On Earth, when combined with the upward acceleration ofgravity (Ag), the resultant GIA vectors had magnitudes of 1.4 and1.1 g and were tilted in the roll plane by 45 and 27° relative to thesubject’s head dorsoventral (Zh) axis, respectively (Fig. 1a, b). In thelying-on-back configuration, centripetal accelerations along thesubject’s head and body yaw axes (Zh) were oppositely directed atthe feet and head. Rotations at constant velocities of 223 and 158°/sgenerated 1 and 0.5 g of linear acceleration at the level of theinteraural axis, pitching the GIA 45 and 27° down with respect tothe subject’s naso-occipital (Xh) axis, respectively (Fig. 1c). Onlythe centripetal acceleration acting on the head is shown in Fig. 1c,but it was matched by an oppositely directed centripetal accelerationat the feet.During flight, the gravitational acceleration sensed by the otoliths
(Ag) was negligible (~10−6g) and the GIA was equivalent to thecentripetal linear acceleration (Ac) with magnitudes of 1 or 0.5 g(Fig. 1a–c). We used the same terminology, right-ear-out, left-ear-out and lying-on-back to describe the different conditions on orbit.Thus, in microgravity the GIA was always aligned with theinteraural or Yh axis during Gy centrifugation and was tilted 90°relative to the head yaw (Zh) axis with magnitudes of 1 or 0.5 g(Fig. 1a–c). Similarly, the GIA was along the dorsoventral (Zh) axisin the lying-on-back configuration. In this paper ‘1-g centrifugation’is defined as centrifugation with a centripetal acceleration of 1 g(both on Earth and in space); similarly ‘0.5-g centrifugation’ isdefined as centrifugation with a centripetal acceleration of 0.5 g.
Visual stimuli
The visual display, developed by the European Space Agency,consisted of a 158×167 mm liquid-crystal display screen mounted ina box directly in front of the subject’s face on the centrifuge chair,with a field of view of ±44° horizontally and ±40° vertically. Thenaso-occipital (Xh) axis was normal to the display screen.Measurements of the difference between the horizontal position ofthe left and right eyes during visual stimuli indicated a mean ocularvergence of 5.2°, suggesting that the optics generated a display thatwas seen at a distance of 68 cm. The optokinetic stimuli werehorizontal, vertical, or 45° oblique amber stripes of 5° width thatmoved at a constant velocity of 30°/s over a black background.Randomly generated dots were superimposed over the edges of thestripes to prevent visual tracking of the stripe edges as they weredrawn. Optokinetic stimuli were presented for 20 s followed by a10-s period of darkness in the following order: (1) up, (2) up and tothe right, (3) right, (4) down, (5) down and to the left, and (6) left. Inall instances, the direction of OKN was defined by the direction of
the slow phase velocity. Eye movements were calibrated by havingthe subjects look at 0.36° targets appearing at 25 successivelocations on two concentric circles (corresponding to gaze angles of10 and 20°) on the screen. The center point of the calibration patternwas directly ahead in binocular gaze, and subsequent eye positionwas referenced to this ‘zero’ point. Calibrations were performed atthe beginning of each test and were repeated each time the subjectsentered the chair. A ‘mini-calibration’ was also performed duringeach centrifuge run by having the subjects look at five targets (0°and ±10° horizontally and vertically) to ensure that the head had notmoved during the acceleration phase.Horizontal and vertical smooth pursuit was elicited by motion of a
0.36° black target on an amber background using a step ramp profile(Rashbass 1961). There were four trials for each direction of targetmotion (up, down, left, right) in random order. The target wasstationary at the center of the screen for periods ranging from 1.8 to2.4 s. It then stepped 5° to one side before moving at a constantspeed of 20°/s across the mid-position to a final position atamplitudes ranging from 25 to 35°. The duration of the appearanceof the target at the center of the screen and the final angular positionof the target were varied randomly to avoid anticipation. The size ofthe step was adjusted so that the target moved past the midpoint150 ms after the onset of target motion. Across subjects and trials,the latency to the onset of smooth pursuit was 132±36 ms. Thus, thetarget was foveated at the onset of pursuit. This provided optimumconditions for tracking to be initiated with a smooth acceleration ofthe eyes rather than with a saccade (Rashbass 1961).
Tilt stimuli
The chair of the ground-based centrifuge could also be tilted aroundan axis located under the subjects’ feet by means of an actuator,permitting OKN and ocular pursuit to be tested with subjects upright(0°) and statically tilted in roll left-ear-down (LED) at angles of 30,45 and 90°, corresponding to steady-state interaural linear accelera-tions of 0, 0.5, 0.7 and 1 g, respectively. Optokinetic and smoothpursuit stimuli were presented, as for centrifugation, on the visualdisplay described above at each angle of tilt. A separate tilt chair anddome, developed at the Mount Sinai School of Medicine by LeighMcGarvie (Neuro-otology Department, Royal Prince Alfred Hospi-tal, Sydney, Australia), was modified for use in the Neurolabexperiments (Neurokinetics, Inc.) and installed at the KennedySpace Center (Clément et al. 2001; Moore et al. 2001). Subjects satupright in an automobile-racing seat, firmly held in place by a 5-point safety harness, adjustable padded shoulder and neck supports,and a fiberglass back shell to support the head. A sturdy pin lockedthe chair in place at each angle. The optokinetic display wasprojected on a 3-m diameter white hemisphere, positioned directly infront of the subjects and centered at eye level. The optokineticstimulus was a full-field pattern of 5°-wide black and white stripes,moving at 30°/s in the same horizontal, vertical and obliquedirections and for the same duration (20 s) as during centrifugation.A static 5-point display, consisting of a center point and foureccentric points at ±10° horizontal and vertical gaze angles, wasused to calibrate eye movements. Subjects were tested in this devicejust prior to launch (L-2) and within 4 h of landing (R+0).
Eye movement recording
Binocular eye movements were recorded by two miniature NTSCvideo cameras mounted either side of the visual display unit. Thecameras generated video images at a frame rate of 30 Hz. Processingof each odd and even video field advanced the sampling rate of theeye movements to 60 Hz. Two rectangular banks of infrared light-emitting diodes (wavelength 950 nm) were attached to each cameraand illuminated the subject’s eyes. The diodes were not visible to thesubject. Images of the eyes were directed onto charge-coupleddevices of the video cameras via an infrared beam-splitter that wastransparent to light in the visible range, allowing the subjects a clear
42

view of the display. The cameras were positioned and focusedmanually by the operator before each centrifuge run using a smallvideo monitor mounted on the wall of the Spacelab to obtain optimalimages of the subject’s eyes. Eye movements, verbal perceptionreports and parameters relative to the centrifuge configuration andmotion (velocity, acceleration, mission-elapsed time) were recordedon Hi-8 tape recorders mounted on the centrifuge. During the flight,video images of the eyes and centrifuge parameters were transmittedin real-time to the Science Monitoring Facility at Johnson SpaceCenter in Houston, allowing the investigator team to verify dataquality.
Experimental protocol
Following a 25-point calibration, optokinetic stimuli and pursuittargets were presented with the centrifuge immobile (no-chair-motion or NCM) for control. This was typically followed by threecentrifuge profiles. First, the subject was oriented left-ear-out(Fig. 1a) in darkness, and the centrifuge was accelerated at 26°/s2
in a clockwise direction (subject facing motion) to a constantvelocity that produced a centripetal acceleration of 0.5 or 1 g.Vestibular nystagmus was recorded in darkness during the angularacceleration and for the first 60 s of constant velocity rotation. Then,with the chair still rotating, the subject performed a 5-point mini-calibration and next viewed optokinetic stimuli moving (1) up, (2)up and right, (3) right, (4) down, (5) down and left, and (6) left. Eachsubject was asked to verbally report their perception of the motion ofthe moving stripes relative to the head during the oblique (‘up andright’ and ‘down and left’) OKN stimuli, which was recorded on theaudio track of the Hi-8 tape and analyzed post-flight (Clément et al.2001). After a short period in darkness, the subjects performed the16 smooth pursuit trials. The centrifuge was then decelerated indarkness at 26°/s2 to rest.This profile was then repeated with the subject right-ear-out
(Fig. 1b) rotating counterclockwise, and when lying-on-back
(Fig. 1c) with clockwise rotation. A full calibration was performedprior to centrifugation in each case. Only one centripetal acceler-ation magnitude (0.5 or 1 g) was used in a test session on any givenday. Due to mission time constraints, presentation of all optokineticand smooth pursuit stimuli was not possible for all in-flight testsessions. Oblique optokinetic stimuli were not presented duringcentrifugation when lying-on-back and smooth pursuit was nottested in the right-ear-out orientation. Only an abbreviated test wasperformed in-flight on the first and last days of the sciencecomponent of the mission (FD2 and FD16), where the right-ear-outrun was eliminated, and subjects were centrifuged only in the left-ear-out and lying-on-back configurations. In addition, time con-straints limited the allowable time between presentation of eachoptokinetic stimuli to 10 s; therefore, it was not possible to recordoptokinetic after-nystagmus (OKAN) following each OKN run.Thus, OKAN will not be considered in this report. In pre- and post-flight testing, optokinetic and smooth pursuit stimuli were presentedduring centrifugation as well as during left-ear-down static roll tiltsat angles of 0, 30, 45 and 90°, using the tilt capability of the groundcentrifuge and the stand-alone tilt chair at Kennedy Space Center.
Data analysis
Horizontal, vertical, and torsional eye movements were determinedfrom videotape using a Fick rotation convention (rotation about eye-fixed axes using a horizontal, vertical, torsional sequence) with aresolution of 0.1° (Fick 1854; Moore et al. 1991, 1996; Zhu et al.1999, 2004). For a detailed description of the video-based eyemovement monitor see Moore et al. (1996, 2001). Eye velocitiesduring OKN were computed as angular velocity vectors along theroll (Xh), pitch (Yh) and yaw (Zh) axes of the head from the Fickcoordinates and their derivatives (Goldstein 1980; Haslwanter 1995;Yakushin et al. 1995). Velocities associated with saccades wereremoved by hand and slow phase eye velocity was extrapolatedacross the saccade. This technique is equivalent to those that obtain
Fig. 2a, b Typical pre-flight data during rightward horizontaloptokinetic stimulation. The subject was seated upright with thecentrifuge stationary. a From top to bottom the traces represent:optokinetic stimulus on, horizontal eye position (H pos, leftpositive), vertical eye position (V pos, down positive), torsionaleye position (T pos, clockwise from the subject’s point of viewpositive), horizontal velocity (H vel), vertical velocity (V vel), andtime calibration (5 s). The eye position calibration (10 deg) was thesame for all three components of eye position and is shown under Hpos. The velocity calibration (20 deg/s) is shown under H vel. Therewas vigorous horizontal OKN with the mean eye position deviated
towards the left, i.e., toward the origin of the moving stripes. Thehorizontal dashed line represents average eye position (the beatingfield) over the 20-s stimulus period. Vertical and torsional eyecomponents were small. b Phase plot of vertical re horizontal eyeposition seen from the point of view of the subject (insert). Each dotrepresents an eye position during the slow phases. The dashedvertical and horizontal lines indicate the center of the visual field.The vertical solid line (OKN axis) was calculated from a linear leastsquares fit to the eye position data, and shows the direction of thevelocity vector and marks the mean shift in the beating field to theleft
43

eye velocity from eye position representations using quaternions orrotation vectors (Haslwanter 1995). The beating field was calculatedfrom the mean eye position during OKN (Fig. 2a, H pos, dashedline). The OKN eye velocity vector was determined from itscomponents along the roll, pitch, and yaw axes of the head, fromwhich tilts of the vector relative to the GIA and the visual stimuluscould be computed. The magnitude of OKN tilt during centrifuga-tion was calculated relative to the OKN velocity axis in response tothe same visual stimulus direction with the centrifuge stationary.OKN direction was defined as the direction of the slow phases
and of slow phase velocity, not of the quick phases. Thus, upwardOKN was OKN in which both the eyes and the visual stimulusmoved upward. The same was true for downward, leftward orrightward OKN. Gains of OKN were defined in terms of thestimulus magnitude and direction. The magnitude of the stimulusvelocity was 30°/s for all tests. For a horizontal optokinetic stimulus,the horizontal gain was defined as the magnitude of the horizontaleye velocity/stimulus velocity. Similarly, vertical gain was definedas vertical eye velocity/stimulus velocity.OKN tilts from the four payload crew subjects during horizontal
optokinetic stimulation and 1-g centrifugation were pooled into pre-flight (six measures per subject), in-flight (eight measures persubject) and post-flight (six measures per subject) bins for eachcentrifuge condition (no-chair-motion NCM, left-ear-out LEO, right-ear-out REO, and lying-on-back LOB). Statistical analysis wasperformed using a one-dimensional repeated measures analysis ofvariance (ANOVA; LABVIEW, National Instruments, Austin, TX,USA) to determine if microgravity affected the OKN tilt for eachcentrifuge condition. Results were considered significant for P<0.05. This process was repeated for vertical and oblique optoki-netic stimulation.
Results
Mean eye position during OKN (beating field)
The general characteristics of horizontal OKN weresimilar on Earth and in-flight. A typical example of pre-flight horizontal OKN with the centrifuge stationary isshown in Fig. 2. The stripes moved to the right, evokingrightward slow phases. At the onset of stimulation, theeyes beat across the midline to the left, into the direction ofthe quick phases and against the direction of stimulusmotion, with a mean leftward eye position deviation of12.3° toward the origin of the stripes (Fig. 2a). There wasalso a small downward deviation of about 2°, but notorsional component (Fig. 2a). The shift in the horizontalbeating field is shown both in the raw data (Fig. 2a) and inthe phase plot of vertical re horizontal eye position(Fig. 2b). Similar shifts of the beating field in the directionof the quick phases of leftward or rightward OKN werepresent during Gy and -Gz centrifugation and were notaffected by microgravity (Fig. 3a). Over the 20-s period ofoptokinetic stimulation, the average deviation of thehorizontal beating field for all conditions was −7.2,−10.4 and −9.9° (pre-, in- and post-flight) to the rightduring leftward OKN, and 8.2, 8.8 and 12.2° to the leftduring rightward OKN (Fig. 3a). Differences between thethree flight conditions were not significant.
In contrast, there were significant effects of micro-gravity exposure and visual stimulus direction on meaneye position during vertical OKN (Fig. 3b). On Earth,when the stimulus field was moving up, the beating field
of the upward OKN moved down towards the origin of themoving stripes (Fig. 3b; Pre-flight, upward pointingtriangle), similar to horizontal OKN (Fig. 3a), with anaverage magnitude of 8.7°. When the stimulus movementwas downward, however, the beating field of the down-ward OKN remained close to the midline (−1.1°) (Fig. 3b;Pre-flight, downward pointing triangle). Sample data from90 days prior to launch demonstrates this tendency(Fig. 3c; V pos), where mean eye position deviated only0.6° up from the midline during downward OKN. Inspace, however, a significant (P <0.05) upward deviationof the eyes (−9°) developed during downward OKN(Fig. 3b; In-flight, downward pointing triangle). This shiftcan clearly be seen in data from the same subject on flightday (FD) 5, where the mean eye position during down-ward OKN was now shifted up by −9.6° (Fig. 3d; V pos).The downward shift during upward OKN, however, wasunchanged (Fig. 3b; In-flight, upward pointing triangle).Post-flight, the upward shift of the eyes during downwardOKN was reduced (P <0.01) to close to the pre-flightlevels (−3.8°) (Fig. 3b; Post-flight, downward pointingtriangle), but there was a significant increase (P <0.01) indownward deviation of the beating field during upwardOKN (Fig. 3b; Post-flight, upward pointing triangle) to13.4°. Thus, upward deviation of the beating field wassuppressed during downward OKN on Earth, but emergedin-flight in the absence of gravity. Post-flight re-adaptationto gravity induced an increase in the downward shift inmean eye position during upward OKN, and suppressionof the beating field shift of downward OKN was againevident.
Horizontal orienting responses to interaural linearacceleration have recently been described in monkeysand humans (Cohen et al. 2001; Kushiro et al. 2002). Weevaluated whether the addition of the 1 or 0.5 g ofinteraural centripetal acceleration during Gy centrifugationprovoked a shift in the beating field. A comparison ofaverage eye position during horizontal OKN with thecentrifuge stationary and in motion revealed no differences(Fig. 3a). Therefore, the addition of centripetal accelera-tion did not alter the shifts of the beating field ofhorizontal OKN, either on Earth or in-flight.
OKN gain
The average gains of horizontal, vertical and oblique OKNfor the four astronauts with the centrifuge stationary andduring centrifugation are shown in Table 1. With thecentrifuge stationary (No Chair Motion, first three rows),horizontal slow phase velocity to the left was slightlygreater than to the right. This produced a small asymmetryin the gain of horizontal OKN, which was maintainedduring and after flight. The cause for this asymmetry isunknown, and it was considered to be a characteristic ofthe group.
In general, there was little effect of space flight on OKNgain, with one exception. Exposure to microgravity alteredthe gains of vertical OKN, producing a reversal of the
44

Table 1 Gain of optokineticnystagmus (OKN). (Mean andSE of all trials for four subjects,data pooled into pre-, in- andpost-flight bins)
UP DOWN LEFT RIGHT UP/RIGHT DOWN/LEFT
No Chair Motion (NCM)Pre-flight 0.65±0.16 0.73±0.14 0.89±0.04 0.77±0.13 0.82±0.15 0.77±0.09In-flight 0.74±0.15 0.72±0.15 0.89±0.06 0.83±0.08 0.84±0.13 0.80±0.12Post-flight 0.70±0.17 0.75±0.16 0.93±0.04 0.79±0.13 0.77±0.18 0.83±0.12
Left-Ear-Out/Right-Ear-Out (LEO/REO) 1-g CentrifugationPre-flight 0.73±0.16 0.81±0.13 0.87±0.09 0.90±0.10 0.87±0.14 0.81±0.12In-flight 0.76±0.15 0.77±0.16 0.92±0.05 0.89±0.09 0.90±0.13 0.84±0.10Post-flight 0.67±0.14* 0.84±0.12* 0.90±0.07 0.87±0.10 0.85±0.14 0.86±0.08
Lying-On-Back (LOB) 1-g CentrifugationPre-flight 0.79±0.15 0.81±0.14 0.89±0.04 0.84±0.11 X XIn-flight 0.83±0.14 0.65±0.17 0.89±0.06 0.81±0.11 X XPost-flight 0.83±0.13 0.86±0.13 0.94±0.04 0.86±0.07 X X
Fig. 3a–d Mean position of the beating field during horizontal andvertical OKN for all four subjects, with the centrifuge stationary(NCM) and during Gy (LEO) and -Gz (LOB) centrifugation. aDuring both rightward and leftward OKN, the mean eye positiondeviated approximately 10° towards the origin of the moving stripes.Neither microgravity nor centrifugation (LEO, LOB) affected themagnitude of the shifts in mean horizontal eye position. The lateraltriangles show means of all conditions ±1 SD. The inserts illustratethe direction of stripe motion. The eyes show the direction ofdeviation of the beating field in response to the stimulus to thesubject’s right (upper panel) and left (lower panel). b Schemesimilar to that in a, showing vertical deviation of the beating fieldduring upward OKN (upper panel) and downward OKN (lower
panel). Inserts illustrate the direction of stripe motion and thevertical deviation of the eyes during OKN. During upward OKNthere was a 10° downward shift in mean eye position (upwardtriangle) towards the origin of the moving stripe pattern that wasmaintained in-flight. There was a significant (*) increase in thisdownward shift post-flight. During downward OKN on Earth therewas no upward shift in the beating field pre- and post-flight(downward triangle), with mean eye position close to the midline.In-flight, however, there was a significant (9°) upward shift towardsthe origin of the stripes. c Typical downward OKN 90 days prior tolaunch. The mean vertical eye position was close to the midline(−0.6°). d After 5 days on orbit, the mean eye position shiftedupward by 9.6° during downward OKN
45

vertical asymmetry. On Earth, downward OKN had ahigher gain (0.81±0.13) than upward OKN (0.72±0.15),both with the centrifuge stationary and with the centrifugein motion (P <0.05; Table 1, mean of all upward anddownward pre- and post-flight OKN [first two columns]).This asymmetry was lost in-flight, and the gain of upwardOKN (0.79±0.14) was slightly larger than downwardOKN (0.72±0.16). The difference was not statisticallysignificant, however. A similar reversal in up/down gainasymmetry of the aVOR has been noted during previousorbital space flights (Berthoz et al. 1986; Clément 1998;Clarke et al. 2000).
Spatial orientation of OKNwith the centrifuge stationary (No Chair Motion)
There was no effect of test day on tilts in the axes of eyevelocity during OKN with the centrifuge stationary (P>0.05). Therefore, the data were grouped into pre-, in- andpost-flight bins and averaged across the four subjects(Fig. 4). A small downward eye velocity component waspresent on Earth during horizontal OKN that wasindependent of the direction of the stimulus. This down-ward vertical component tilted the axis of eye velocity anaverage of 7.4±2.1° from the stimulus axis during pre- andpost-flight tests. There was a small decrease in this verticalbias in-flight (P <0.05), with the axis of OKN tilted 5±2.1°(Fig. 4a–d). We conjectured that this tilt might have been
Fig. 4a–l Axes of eye velocity (means [ black lines] and SE [ greylines]) of four subjects portrayed against the stimulus axes of OKNduring horizontal (top row, b–d), vertical (middle row, f–h) andoblique (bottom row, j–l) optokinetic stimulation. Subjects werestationary (no centrifuge motion; NCM). The orientation of theoptokinetic stimuli are shown in the insets (a, e, i). Duringhorizontal optokinetic stimulation (a–d) a small downward eyemovement was present throughout pre-, in- and post-flight testing.
This tilted the OKN axis up to 7° from the optokinetic stimulus axis.During vertical optokinetic stimulation (e–h), the axis of OKN wasclosely aligned with the stimulus axes. There was a markedasymmetry in the OKN response to oblique OKS (i–l). The OKNaxis was more closely aligned with the stimulus axis when the visualfield moved down and to the left. There was a significant (*)increase in the deviation of the OKN axis from the obliquestimulation axis when the stimulus was directed up and to the right
46

related to the fact that the leading edges of the stripes weredrawn from top to bottom of the visual display. As statedin the Methods, however, the edges of the stripes wereobscured with a random dot pattern, and there was noright-to-left tendency during vertical OKN, which mayhave been expected if subjects were tracking the edge ofthe stripes as they were drawn. A similar downward biasduring horizontal OKN with the head upright wasobserved in two out of six subjects in a previous studyusing solid black and white stripes, but was absent in theother four subjects (Gizzi et al. 1994). The cause for thisdeviation of the axis of horizontal OKN is unknown.
During vertical optokinetic stimulation (Fig. 4e), eyevelocity was closely aligned with the axis of stimulationboth on Earth and in-flight (1.1±2.6°; Fig. 4f–h). Incontrast, the axis of eye velocity rotated clockwise awayfrom the axis of stimulation during oblique stimulation(Fig. 4i) toward the subjects’ dorsoventral (Zh) axis by 9.9±4.1° for stripes moving down and to the left both onEarth and in-flight (Fig. 4j–l; upper-left quadrants). Therewas a significant (P <0.01) increase in this clockwise tilt to23.0±4.2° when the visual stimulus was moving up and tothe right (Fig. 4 j–l; lower-right quadrants). This did notappear to reflect an orientation of the OKN axis to gravity,as these oblique OKN velocity tilts were maintained in-flight. The large asymmetry in OKN axis tilts duringoblique stimulation may be due in part to the downwardbias observed during horizontal OKN (Fig. 4b–d), whichtended to drive the OKN axis closer to the axis ofstimulation when the stripes moved down and to the left.
Thus, in general, there was little effect of microgravityon the orientation of the OKN axis with the centrifugestationary. In subsequent analyses, OKN tilts duringcentrifugation were calculated relative to the OKNvelocity axis tilt with the centrifuge stationary from thesame test session.
Spatial orientation of OKN during Gy (left- and right-ear-out) centrifugation
Horizontal OKN
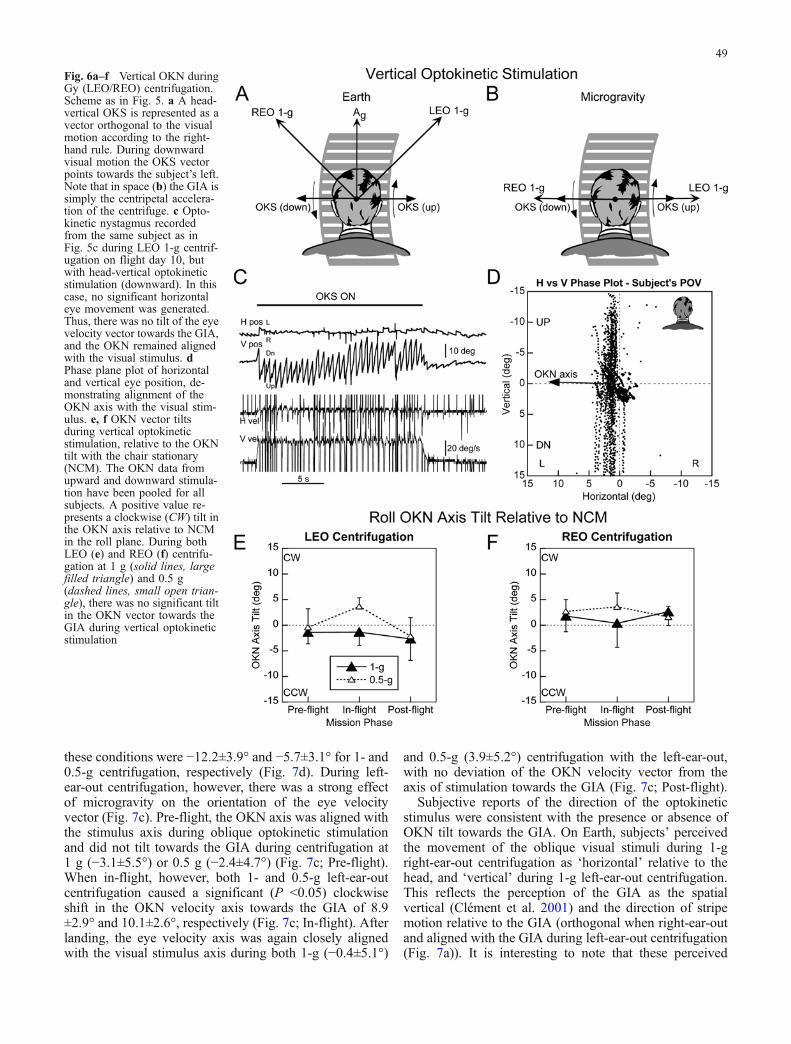
On Earth, constant velocity centrifugation tilted the GIA inthe roll (Yh-Zh) plane away from the gravitational verticalwhen upright and oriented left- and right-ear-out out by 27and 45° for centripetal accelerations of 0.5 and 1 g,respectively (Fig. 5a). During flight, gravity was negli-gible and the GIA was equivalent to the 0.5- or 1-gcentripetal acceleration, and was solely directed along theinteraural (Yh) axis (Fig. 5b). In response to these tilts ofthe GIA, vertical components appeared in horizontal OKN(Fig. 5c; V pos), tilting the axis of eye velocity toward theGIA (Fig. 5d, arrow). A vertical downward eye positionshift of about 7° (in the direction of the slow phases) wasalso present in this condition (Fig. 5d).
There was no significant difference in the tilts of theOKN velocity vector on Earth and in space during either1- or 0.5-g Gy centrifugation. During 1-g Gy centrifuga-
tion, the mean OKN axis tilts in the roll plane, relative tothe OKN axis tilt with the centrifuge stationary, were 8.9±3.5° (mean and SE of all pre-, in- and post-flight data)towards the GIA for left-ear-out centrifugation and −8.9±4.3° during right-ear-out centrifugation (Fig. 5e, f, solidlines). During 0.5-g centrifugation, the mean OKNvelocity vector tilts were 6.7±2.2° when left-ear-out and−6.4±2.7° during right-ear-out centrifugation (Fig. 5e, f;dashed lines). Although the mean tilt values during 0.5-gcentrifugation were less than the 1-g trials, the differenceswere not significant due to the limited data set (only twoin-flight subjects at 0.5 g). These data confirm the resultsof a previous study of horizontal OKN during lateral headtilts on Earth (Gizzi et al. 1994), demonstrating robustcross-coupling from horizontal-to-vertical eye movementsduring roll-tilts of the GIA. They further demonstrate thatthe shift in the axis of eye velocity towards the GIA wasmaintained during and after space flight, indicating thatadaptation to microgravity had not altered the orientationproperties of velocity storage.
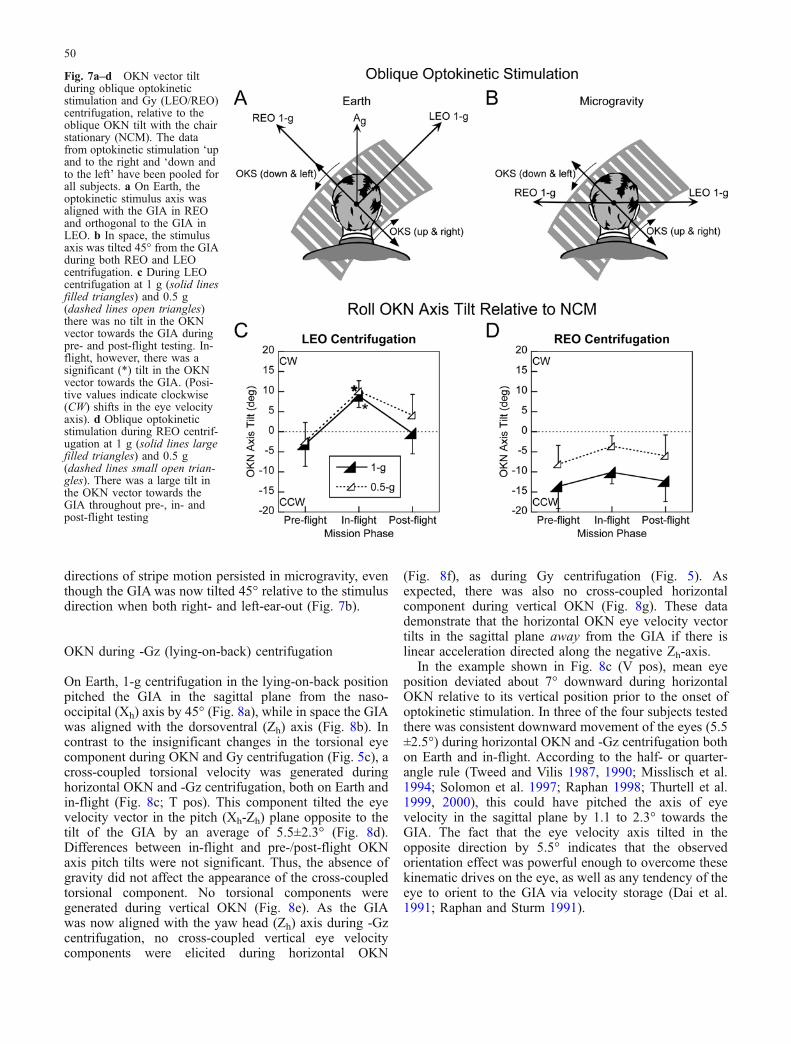
Vertical OKN
Only vertical OKN components were induced duringvertical optokinetic stimulation and Gy centrifugation,with no significant horizontal or torsional componentsboth on Earth (not shown) and in-flight (Fig. 6c, d). Asnoted above, there was a tendency for the mean position ofthe beating field to move up during downward OKN inspace (Fig. 6c; V pos). Due to the absence of cross-coupled components, the eye velocity axis of verticalOKN was aligned with the pitch (Yh) axis in all conditions(Fig. 6d), and tilts of the eye velocity axis were notsignificantly different from the axis of stimulation (P>0.05). The mean deviations of the axes of eye velocityfrom the stimulus velocity axis for 1-g centrifugation were−1.8±3.0° and 1.7±3.4° for left- and right-ear-out centrif-ugation, respectively (Fig. 6e, f; solid lines). Similarly,there were insignificant shifts of the OKN velocity vectorsduring 0.5-g centrifugation with the left- (−0.3±3.4°) andright-ear-out (2.4±2.2°) (Fig. 6e, f; dashed lines). Thesedata show that the axis of vertical OKN did not tilt towardthe GIA during Gy centrifugation, confirming theassumption that there is no vertical-to-horizontal crosscoupling from velocity storage during vertical OKN whenthe GIA is tilted relative to the yaw axis of the head in theroll plane (Dai et al. 1991; Raphan and Sturm 1991).
Oblique OKN
In contrast to the OKN response during horizontal andvertical optokinetic stimulation, there was a strikingasymmetry in the response to oblique visual stimulationduring centrifugation. The counterclockwise shift of theOKN axis in the roll plane toward the GIA during both0.5- and 1-g right-ear-out centrifugation was unaffected byspace flight. The overall mean tilts towards the GIA in
47

Fig. 5a–f Horizontal OKN during Gy (LEO/REO) centrifugation.a A head-horizontal optokinetic stimulus (OKS) is represented as avector orthogonal to the visual motion according to the right-handrule. Thus, during leftward visual motion the OKS vector pointsupward. The REO 1-g and LEO 1-g arrows denote the orientation ofthe GIA relative to the head during centrifugation on Earth (a) andon orbit (b). Note that in space (b) the GIA is now simply thecentripetal acceleration of the centrifuge. c Optokinetic nystagmusrecorded from a subject in response to head-horizontal optokineticstimulation (rightward) during LEO 1-g centrifugation on flightday 10. Although the visual stimulus was head-horizontal there wasa significant downward vertical eye velocity component, whichtilted the OKN axis towards the GIA. Note that there was notorsional eye movement component. d Phase plane plot of horizontaland vertical eye position, demonstrating the roll tilt of the OKN axis
towards the GIA of approximately 10°. Each dot represents an eyeposition during the OKN slow phases. The vertical dotted lineshows the axis of stimulation, and the solid line the OKN eyevelocity axis in the roll (Yh-Zh) plane. The arrow denotes the shift inthe axis of eye velocity towards the GIA due to the centripetalacceleration. e, f OKN vector tilt during horizontal optokineticstimulation, relative to the OKN axis tilt with the chair stationary(NCM). The data from leftward and rightward horizontal stimulationhave been pooled for all subjects. A positive value represents aclockwise (CW) tilt in the OKN axis in the roll plane from thesubject’s point of view. During both LEO (e) and REO (f)centrifugation at 1 g (solid lines, large filled triangle) and 0.5 g(dashed lines, small open triangle) there was a robust tilt in theOKN vector towards the GIA during horizontal optokineticstimulation
48
these conditions were −12.2±3.9° and −5.7±3.1° for 1- and0.5-g centrifugation, respectively (Fig. 7d). During left-ear-out centrifugation, however, there was a strong effectof microgravity on the orientation of the eye velocityvector (Fig. 7c). Pre-flight, the OKN axis was aligned withthe stimulus axis during oblique optokinetic stimulationand did not tilt towards the GIA during centrifugation at1 g (−3.1±5.5°) or 0.5 g (−2.4±4.7°) (Fig. 7c; Pre-flight).When in-flight, however, both 1- and 0.5-g left-ear-outcentrifugation caused a significant (P <0.05) clockwiseshift in the OKN velocity axis towards the GIA of 8.9±2.9° and 10.1±2.6°, respectively (Fig. 7c; In-flight). Afterlanding, the eye velocity axis was again closely alignedwith the visual stimulus axis during both 1-g (−0.4±5.1°)
and 0.5-g (3.9±5.2°) centrifugation with the left-ear-out,with no deviation of the OKN velocity vector from theaxis of stimulation towards the GIA (Fig. 7c; Post-flight).
Subjective reports of the direction of the optokineticstimulus were consistent with the presence or absence ofOKN tilt towards the GIA. On Earth, subjects’ perceivedthe movement of the oblique visual stimuli during 1-gright-ear-out centrifugation as ‘horizontal’ relative to thehead, and ‘vertical’ during 1-g left-ear-out centrifugation.This reflects the perception of the GIA as the spatialvertical (Clément et al. 2001) and the direction of stripemotion relative to the GIA (orthogonal when right-ear-outand aligned with the GIA during left-ear-out centrifugation(Fig. 7a)). It is interesting to note that these perceived
Fig. 6a–f Vertical OKN duringGy (LEO/REO) centrifugation.Scheme as in Fig. 5. a A head-vertical OKS is represented as avector orthogonal to the visualmotion according to the right-hand rule. During downwardvisual motion the OKS vectorpoints towards the subject’s left.Note that in space (b) the GIA issimply the centripetal accelera-tion of the centrifuge. c Opto-kinetic nystagmus recordedfrom the same subject as inFig. 5c during LEO 1-g centrif-ugation on flight day 10, butwith head-vertical optokineticstimulation (downward). In thiscase, no significant horizontaleye movement was generated.Thus, there was no tilt of the eyevelocity vector towards the GIA,and the OKN remained alignedwith the visual stimulus. dPhase plane plot of horizontaland vertical eye position, de-monstrating alignment of theOKN axis with the visual stim-ulus. e, f OKN vector tiltsduring vertical optokineticstimulation, relative to the OKNtilt with the chair stationary(NCM). The OKN data fromupward and downward stimula-tion have been pooled for allsubjects. A positive value re-presents a clockwise (CW) tilt inthe OKN axis relative to NCMin the roll plane. During bothLEO (e) and REO (f) centrifu-gation at 1 g (solid lines, largefilled triangle) and 0.5 g(dashed lines, small open trian-gle), there was no significant tiltin the OKN vector towards theGIA during vertical optokineticstimulation
49
directions of stripe motion persisted in microgravity, eventhough the GIAwas now tilted 45° relative to the stimulusdirection when both right- and left-ear-out (Fig. 7b).
OKN during -Gz (lying-on-back) centrifugation
On Earth, 1-g centrifugation in the lying-on-back positionpitched the GIA in the sagittal plane from the naso-occipital (Xh) axis by 45° (Fig. 8a), while in space the GIAwas aligned with the dorsoventral (Zh) axis (Fig. 8b). Incontrast to the insignificant changes in the torsional eyecomponent during OKN and Gy centrifugation (Fig. 5c), across-coupled torsional velocity was generated duringhorizontal OKN and -Gz centrifugation, both on Earth andin-flight (Fig. 8c; T pos). This component tilted the eyevelocity vector in the pitch (Xh-Zh) plane opposite to thetilt of the GIA by an average of 5.5±2.3° (Fig. 8d).Differences between in-flight and pre-/post-flight OKNaxis pitch tilts were not significant. Thus, the absence ofgravity did not affect the appearance of the cross-coupledtorsional component. No torsional components weregenerated during vertical OKN (Fig. 8e). As the GIAwas now aligned with the yaw head (Zh) axis during -Gzcentrifugation, no cross-coupled vertical eye velocitycomponents were elicited during horizontal OKN
(Fig. 8f), as during Gy centrifugation (Fig. 5). Asexpected, there was also no cross-coupled horizontalcomponent during vertical OKN (Fig. 8g). These datademonstrate that the horizontal OKN eye velocity vectortilts in the sagittal plane away from the GIA if there islinear acceleration directed along the negative Zh-axis.
In the example shown in Fig. 8c (V pos), mean eyeposition deviated about 7° downward during horizontalOKN relative to its vertical position prior to the onset ofoptokinetic stimulation. In three of the four subjects testedthere was consistent downward movement of the eyes (5.5±2.5°) during horizontal OKN and -Gz centrifugation bothon Earth and in-flight. According to the half- or quarter-angle rule (Tweed and Vilis 1987, 1990; Misslisch et al.1994; Solomon et al. 1997; Raphan 1998; Thurtell et al.1999, 2000), this could have pitched the axis of eyevelocity in the sagittal plane by 1.1 to 2.3° towards theGIA. The fact that the eye velocity axis tilted in theopposite direction by 5.5° indicates that the observedorientation effect was powerful enough to overcome thesekinematic drives on the eye, as well as any tendency of theeye to orient to the GIA via velocity storage (Dai et al.1991; Raphan and Sturm 1991).
Fig. 7a–d OKN vector tiltduring oblique optokineticstimulation and Gy (LEO/REO)centrifugation, relative to theoblique OKN tilt with the chairstationary (NCM). The datafrom optokinetic stimulation ‘upand to the right and ‘down andto the left’ have been pooled forall subjects. a On Earth, theoptokinetic stimulus axis wasaligned with the GIA in REOand orthogonal to the GIA inLEO. b In space, the stimulusaxis was tilted 45° from the GIAduring both REO and LEOcentrifugation. c During LEOcentrifugation at 1 g (solid linesfilled triangles) and 0.5 g(dashed lines open triangles)there was no tilt in the OKNvector towards the GIA duringpre- and post-flight testing. In-flight, however, there was asignificant (*) tilt in the OKNvector towards the GIA. (Posi-tive values indicate clockwise(CW) shifts in the eye velocityaxis). d Oblique optokineticstimulation during REO centrif-ugation at 1 g (solid lines largefilled triangles) and 0.5 g(dashed lines small open trian-gles). There was a large tilt inthe OKN vector towards theGIA throughout pre-, in- andpost-flight testing
50

Fig. 8a–g OKN response during -Gz (LOB) centrifugation.Relative orientation of the GIA and visual stimulus on Earth (a)and in microgravity (b). Note that the optokinetic stimulus axesduring vertical stimulation (not shown) are orthogonal to the page. cHorizontal OKN in-flight during 1-g LOB centrifugation. In contrastto LEO/REO centrifugation (Fig. 5), there was no vertical OKNcomponent. Rather, a cross-coupled CCW torsional nystagmus wasgenerated. This negative torsional velocity tilted the resultant OKN
vector in the pitch plane away from the GIA (inset). d Averageorientation of the axis of horizontal OKN velocity in the pitch (Xh-Zh) plane during horizontal OKS. A positive value for tilt in thepitch plane indicates a nose-down (ND) tilt of the OKN axis fromthe subject’s point of view. The horizontal OKN axis pitched awayfrom the GIA. e The eye velocity axis did not tilt in the pitch planeduring vertical OKN. OKN velocity did not orient in the roll (Yh-Zh)plane during either horizontal (f) or vertical (g) stimulation
51

Static tilt
Orientation of the horizontal OKN velocity vector togravity was also observed during static roll tilt while left-ear-down during pre- and post-flight testing. There was noeffect of test day (P >0.1) and pre- and post-flight datawere expressed relative to the OKN tilt in the uprightposition (0°) and averaged across subjects for each tiltangle (30, 45 and 90°). There was a significant effect ofstatic tilt angle (P <0.01), with the tilt in the velocity ofOKN increasing from 6.3±0.7° at 30° of tilt (0.5-ginteraural linear acceleration stimulus) to 8.2±0.7° at 45°(0.7 g) and 9.5±1.3° at 90° (1 g). The shift of eye velocityduring horizontal OKN toward the GIA increased linearlywith interaural linear acceleration during static tilt (Fig. 9a;open triangles, dashed line, 9.8°/g, R=0.98). Themagnitude of axis tilts during horizontal OKN and left-ear-out centrifugation at 0.5 g (6.6±1.4°; mean and SD ofall pre-, in- and post-flight data) and 1 g (9.0±1.5°) wasnot significantly different from that induced by static tiltson Earth with equivalent interaural linear accelerations(Fig. 9a; filled squares). This suggests that the magnitudeof tilt of horizontal OKN velocity is dependent on theamount of interaural linear acceleration. There was nosignificant tilt in the velocity vector of vertical or obliqueOKN when the subjects were statically tilted (Fig. 9b).
Ocular pursuit
Target motion at 20°/s in the horizontal and verticaldirections produced brisk eye accelerations at meanlatencies of 142 and 171 ms, respectively (Fig. 10a, b).Mean eye velocity during pursuit was calculated over a500-ms period at a gaze angle of 10°. Both horizontal andvertical velocities were unaffected by space flight(Fig. 10c; P >0.05). Consequently, data from pre-, in-and post-flight tests were combined. Horizontal pursuit
had a mean velocity of 18.6±0.4°/s, corresponding to anaverage gain of 0.93 (Fig. 10c). Vertical pursuit velocitieswere lower than horizontal velocities (Fig. 10b, c), withupward velocity (15.0±0.9°/s) being significantly greaterthan downward velocity (13.1±0.8°/s; P <0.01). Thiscorresponded to gains of 0.75 and 0.66, respectively.There was a significant decrease in downward velocityduring the first 48 h after landing to 11.7±1.6°/s (Fig. 10c;R+1, R+2; P <0.01), but this returned to baseline by thefourth day (R+4). Thus, apart from a transient downwardgain decrease for the first 48 h after landing, there wasrelatively little effect of adaptation to microgravity oneither horizontal or vertical mean pursuit velocity.
There was a greater effect of adaptation to microgravityon saccadic intrusions during pursuit. On Earth, catch-upsaccades, i.e., saccades in the same direction as targetmotion, were present during both horizontal and verticalpursuit, but were more frequent during vertical movement(Fig. 10a, b; L-30). Saccadic intrusions increased in spaceduring both horizontal and vertical pursuit, but as onEarth, were more frequent during vertical movement(Fig. 10a, b; FD 5). This difference was reflected in thecumulative amplitude of forward saccades, which wassignificantly greater in-flight (P <0.01) for both horizontal
Fig. 9a, b OKN velocity vec-tor tilts towards the GIA duringleft-ear-down (LED) static tilts,relative to OKN tilt in theupright position. a During hor-izontal visual stimulation theOKN vector tilt (open triangles)increased with tilt angle, andwas linearly related (dashedline; R=0.98) to the interaurallinear acceleration, i.e., to thesine of the tilt angle. The tilts ofthe OKN velocity vectors whenLED had the same magnitudeand direction as during LEOcentrifugation (average of allpre-, in- and post-flight data)with an equivalent interaurallinear acceleration (filledsquares). b Vertical and obliqueoptokinetic stimulation gener-ated no significant tilt of theOKN velocity vectors in LEDpositions
"Fig. 10a–d Typical eye position and velocity data during trackingof a target moving along head-horizontal (a) or head-vertical (b)axes during pre-flight (L-30 days) and in-flight (flight day 5) testing.Note the increased number of saccades during vertical tracking in-flight. c Mean eye velocity during head-horizontal (right, left) andhead-vertical (up, down) pursuit. There was no change in eyevelocity in-flight or after landing during horizontal pursuit. Therewas a significant (*) decrease in post-flight downward pursuitvelocity that persisted for the first 48 h after landing. (Note: * underL-30, FD07, FD12 and R+4 denotes sessions using 0.5-g centrif-ugation). d Cumulative amplitude of saccades (number of saccades× amplitude) during tracking of a target moving along head-horizontal (right, left) and head-vertical (up, down) axes. There wasa significant (*) increase in saccades in-flight for both horizontal andvertical pursuit. (c and d represent mean ± SD of all four subjectsand tests during NCM, LEO and LOB centrifugation.)
52

53

(pre-flight 1.7±0.8°; in-flight 3.6±1.1°; post-flight 2.3±1.0°) and vertical (pre-flight 8.1±1.6°; in-flight 10.9±1.9°; post-flight 8.8±1.6°) pursuit (Fig. 10d).
No cross-coupled horizontal or vertical eye velocitieswere present when subjects visually tracked the targetduring centrifugation, and the eyes exactly followed thetargets in both the horizontal and vertical directions(Fig. 11). There were no dynamic torsional movements ofthe eyes during pursuit. Thus, eye movements obeyedListings’ law (Haslwanter et al. 1991, 1992). During Gy(interaural) centrifugation, there was a sustained torsionaleye position generated by the OCR reflex (Moore et al.2001), which would act to shift Listing’s plane along theroll axis. The lack of dynamic torsional eye movementsduring pursuit with Gy centrifugation would similarlyconfine the axis of eye movement to this shifted Listing’sPlane (Haslwanter et al. 1992; Bockisch and Haslwanter2001). This was true on Earth and in space flight, as wellas during pre- and post-flight static roll tilt. Thus, ocularpursuit did not exhibit orientation properties during rolltilts of the GIA as observed during horizontal OKN.
Discussion
A number of important findings emerged from this study.The velocity axis of horizontal OKN oriented towards the
GIA in response to an applied interaural linear accelerationin all conditions. This confirms earlier results using activehead tilts and electro-oculography (Gizzi et al. 1994), andextends the results from a 1-g environment to micro-gravity. In contrast, visual tracking exhibited no spatialorientation properties, and the eyes followed the visualtarget regardless of the orientation of the GIA. There wasalso a gravity-dependence of the beating field duringvertical OKN. On Earth, the upward shift in mean eyeposition during downward OKN was suppressed, but inmicrogravity a significant upward shift appeared in thebeating field during downward OKN.
Spatial orientation of OKN during Gy centrifugation
The magnitude of the tilt in the OKN velocity axis wasdependent on the amount of interaural linear acceleration.Thus, the tilt of the eye velocity vector to the GIAproduced by 0.5 g (6.6°) was approximately 70% of thatproduced by 1 g (9°). This is consistent with thedependence of the magnitude of OCR on interaural linearacceleration observed in the same astronauts (Moore et al.2001). There was a difference between the averagehorizontal OKN axis tilt in our results (9°) and thosefound in the previous human study of OKN orientationduring active roll head tilts (18°) (Gizzi et al. 1994). This
Fig. 11a–d Horizontal andvertical eye position during vi-sual pursuit of 16 step-rampsalong head-horizontal (four tothe right, four to the left) andhead vertical (four up, fourdown) axes over a 60-s period. aPursuit data from flight day(FD) 16 with the centrifugestationary (NCM). b Pursuitdata from the same subject andtest session (FD 16) during 1-gLEO centrifugation. c Horizon-tal and vertical eye position andvelocity data during horizontalpursuit from the trial shown inpanel b. d Vertical eye positionand velocity data during verticalpursuit from the trial shown inpanel b. There was no cross-coupling from horizontal tovertical eye velocity during tiltsof the GIA, as observed duringhorizontal OKN. Thus, smoothpursuit did not exhibit spatialorientation properties
54

may be due to the contribution of neck stretch receptors inthe perceived spatial vertical during active head movement(Mittelstaedt and Glasauer 1993), or to variation insensitivity among subjects.
The maintenance of spatial orientation to the GIAduring and after space flight in the four Neurolab subjectsstands in contrast to the 70% reduction of OCR gain andvergence during off-vertical axis rotation, and a shift ineye velocity axis of horizontal OKAN of approximately28° away from the GIA toward the body axis in monkeysafter the COSMOS flight (Dai et al. 1994, 1996).Although a species difference is possible, the cause forthe disparity is likely to be that the motion experience ofthe two groups in space was quite different (Moore et al.2003). The monkeys were seated in primate chairs duringflight and could rotate but not translate their heads. Thus,they had little or no exposure to linear acceleration overthe 2-week flight. In contrast, the astronauts moved freelywithin the spacecraft and regularly experienced translationas they performed their daily in-flight activities. Moreover,they were regularly exposed to intermittent 1- and 0.5-gcentripetal acceleration on the short radius centrifuge overthe course of the 16-day flight. From the COSMOS andNeurolab results we infer that the gains of orientingotolith-ocular reflexes are not fixed, and that gain changescan occur relatively rapidly in the absence of linearacceleration. If correct, exposure to ‘artificial gravity’ on ashort radius centrifugation could be an important counter-measure to deconditioning of orienting reflexes in futurelong-duration manned space flights.
The experiment utilizing oblique OKN and centrifuga-tion provided a unique confirmation of the spatialorientation properties of OKN in the roll plane. Subjectsperceive the GIA as the spatial vertical during centrifu-gation (Clément et al. 2001). When the direction of theobliquely moving stripes was aligned with the GIA onEarth (left-ear-out), subjects sensed that stripe motion was‘vertical’ relative to the head, and there was no tilt in theOKN velocity axis. In contrast, when the direction ofstripe movement was orthogonal to the GIA (right-ear-out), the perceived stripe motion was ‘horizontal’ and theOKN axis tilted toward the GIA in the roll plane. In space,the direction of the oblique optokinetic stimulus was notaligned with the GIA during centrifugation in both the left-or right-ear-out positions, and the OKN velocity axis tiltedtowards the GIA in both cases.
Spatial orientation of OKN during -Gz centrifugation
The finding that the shift in the velocity axis of horizontalOKN was away from the GIA during dorsoventral (-Gzcentrifugation) linear acceleration was unexpected. Shiftsin the eye velocity axis (5.5°) in the pitch plane weresmaller than those in response to interaural acceleration inthe roll plane (9°). In both conditions, the shifts wererobust, occurring across all subjects both on Earth and inmicrogravity. The same pattern of horizontal OKN axistilts away from the GIA during dorsoventral linear
acceleration has been observed in monkeys (Dai et al.1991; Raphan and Cohen 1988). In the monkey, however,there was robust OKAN, and the OKAN velocity vectortilted in the opposite direction toward the GIA, consistentwith other findings that indicated that the orientation ofvelocity storage is truly three-dimensional.
It is unlikely that velocity storage contributed to theOKN tilt during -Gz centrifugation in the current study, asobserved during Gy centrifugation. In the monkey,velocity storage is expressed as OKAN, and the axis ofOKAN velocity aligns with the GIA regardless of whetherthe animal is supine (lying-on-back) or roll-tilted 90°(lying-on-side) (Raphan and Cohen 1988). This impliesthat the orientation of the eye velocity generated byvelocity storage would be toward the GIA in bothconditions. The fact that the tilt was opposite to the GIAduring horizontal OKN and -Gz centrifugation for bothhumans and monkeys argues against a dominant role ofvelocity storage in the observed orientation. Furthermore,the horizontal OKN axis tilt in the pitch plane in theastronauts persisted in microgravity, despite the fact thatthe optokinetic stimulus axis and the GIA were bothaligned with the dorsoventral axis. Rather, we speculatethat the orientation effects of horizontal OKN duringdorsoventral centrifugation may be due to a difference inhow the visual system responded to the retinal slip signals.During horizontal axis stimulation in the Gy configuration,the stripes were vertical relative to the subject, and theeyes could move up or down along the stripes withoutcontrasting visual stimuli that could suppress the cross-coupled vertical eye velocity component. In contrast,when a torsional eye velocity was induced during -Gzcentrifugation, there was visual contrast at the edge of thevertical stripes across the retina. This could havesuppressed any cross-coupled torsional eye velocitygenerated by velocity storage, or driven torsional eyevelocity in the opposite direction, as observed in thisstudy. Evidence for a suppressive effect is shown by thedirection of the subsequent OKAN axis tilt in the monkeyfollowing horizontal optokinetic stimulation while cen-trifuged in a supine position, which was opposite to thatduring OKN and towards the GIA (Raphan and Cohen1988). Since there was little or no OKAN in the humandata, we could not evaluate whether human OKANsimilarly oriented towards the GIA.
We considered whether kinematic effects could havegiven rise to the OKN eye velocity tilts described above,but discounted this on theoretical grounds. The axis of eyevelocity will tilt relative to a vertical eye position changein response to an imposed yaw axis head velocity by 50%(Helmholz 1867). Termed the ‘half-angle rule’ (Tweed andVilis 1987, 1990; Raphan 1998), in actuality, the tilts aresubject-dependent and vary between 20–50% (Misslisch etal.1994; Solomon et al.1997; Thurtell et al. 1999, 2000),leading to the ‘quarter-angle rule’ (Misslisch et al. 1994).Symmetry would suggest that static torsional position ofthe eyes would induce a roll tilt of the eye velocity vectorfor a horizontal optokinetic stimulus due to muscle pulleyaction, which would be in the direction observed in this
55

study, i.e., towards the GIA (Raphan 1998; Thurtell et al.2000; Raphan and Cohen 2002). However, the magnitudesof the predicted tilts due to kinematic effects were toosmall to account for the data. During 1-g Gy centrifuga-tion, which produces GIA tilts in the roll plane, ocularcounter roll was 6° on average on Earth and in space(Moore et al. 2001). According to the quarter- and half-angle rules, this static torsional deviation of the eye couldhave produced eye velocity tilts of 1–3° in the roll planetowards the GIA. In actuality, the tilt in the horizontalOKN velocity vector was 9° in this condition. Likewise,vertical eye position changes of about 6° during -Gzcentrifugation could also have contributed up to 3° to theaverage tilts of horizontal OKN velocity in the pitch planetoward the GIA, whereas the magnitude of pitch tilts of theOKN axis was 5.5° in the opposite direction (i.e., awayfrom the GIA). Thus, it is unlikely that kinematic effectsproduced the observed OKN axis tilts, although they mayhave augmented the tilts due to velocity storage during rolltilts of the GIA, and may account for the smaller axis tiltduring -Gz centrifugation.
Changes in the beating field and OKN gain
During horizontal OKN the beating field of nystagmusmoves against the direction of field motion (Borries 1926;Ter Braak 1936; Rademacher and Ter Braak 1948;Brucher 1964; Hood and Leech 1974). Similar movementof the horizontal beating field in the direction of headmotion also occurs during nystagmus induced by rotation(Ter Braak 1936; Hood 1967). This probably represents atendency to look into the direction of movement forvestibular activation (Chun and Robinson 1978), andtowards the origin of the visual field movement duringoptokinetic stimulation. This movement of the beatingfield was also present for horizontal OKN in the Neurolabastronauts and was not altered in microgravity. The samewas true for downward movement of the beating fieldduring upward OKN. In contrast, the beating field did notmove upward during downward OKN on Earth eitherbefore or after flight. This asymmetry may be related tothe visual flow in the downward direction experiencedwhen walking forwards on Earth, where the importantvisual cues are directly in front or on the ground below.Presumably, it would not be advantageous for gaze to shiftupwards during downward OKN generated by forwardlocomotion. The difference in mean eye position duringupward and downward OKN disappeared in microgravity,with the eyes now shifting upwards during downwardOKN. This may represent an adaptation to the lack of avertical 1-g bias in-flight in an environment where bipedallocomotion is not applicable.
Changes in the asymmetry of vertical OKN gain havebeen observed during both space and parabolic flight(Clément et al. 1992a, b), as well as during static tiltsrelative to gravity (Clément and Lathan 1991). OKN gaindata from the present study were consistent with pastresults, with an asymmetry on Earth (downward gain
larger) that was lost or reversed in-flight. This reversalmay have been due to unloading of the saccules in-flight.
Central coding of linear acceleration
These results have implications with regard to how linearacceleration is coded centrally. As noted above, themaintenance of spatial orientation of eye velocity to theGIA is consistent with other results from the same flight:OCR was maintained proportional to interaural linearacceleration during and after flight (Moore et al. 2001),and perception of the spatial vertical (the GIA) in-flightwas also sustained over the same period (Clément et al.2001). Each of these three functions, perception, OCR andspatial orientation of OKN eye velocity, involves proces-sing by different neural subsystems; perception ispredominantly a cortical phenomenon (Brandt 1999),OCR arises in the vestibular nuclei exclusive of thecerebellum (Markham 1989; Angelaki and Hess 1994;Uchino et al. 1996, 1997; Wearne et al. 1998) andorientation of OKN to the GIA is controlled by thenodulus and uvula of the vestibulo-cerebellum (Angelakiand Hess 1995; Wearne et al. 1998). Had the brain utilizedan internal model of gravity to produce these very differenttypes of orientation, we would have expected changes inthe eye movements and perceptual responses when gravitywas at an imperceptible level. Specifically, interauralcentripetal acceleration would have been sensed astranslation and produced deviation of horizontal eyeposition and/or horizontal nystagmus, and there wouldhave been no shift in the axis of horizontal OKN towardsthe GIA. Moreover, orienting responses to tilt relative togravity or to tilts of the GIA relative to the head and bodywould be reinterpreted perceptually as translation inmicrogravity. However, the orientation to low-frequencylinear acceleration was maintained. In the case of theperceptual responses, there was an initial underestimationof the tilt of the GIA on entry into microgravity and anoverestimation upon returning to Earth (Clément et al.2001), which could be modeled by assuming that theweighting of the body vertical was altered by adaptation tomicrogravity.
We conclude that the brain did not differentiate the 1-gcentripetal acceleration of the centrifuge in microgravityfrom gravitational linear acceleration. This suggests thatthe brain does not utilize an internal estimate of gravity(Merfeld 1995; Merfeld and Young 1995; Angelaki et al.1999, 2000) to generate perception of the upright, nororientation of eye velocity and position towards the GIA.These results favor a ‘filter’ model of spatial orientation inwhich orientation of horizontal eye position and velocity,and torsional eye movements, to the GIA are proportionalto the magnitude of low frequency interaural linearacceleration (Paige and Tomko 1991a, b; Raphan et al.1996; Seidman et al. 1998; Paige and Seidman 1999;Kushiro et al. 2002; Raphan and Cohen 2002).
56

Ocular pursuit
This study also provided the opportunity to study theeffect of gravity on both horizontal and vertical ocularpursuit. Listing’s plane would shift along the roll (Xh) axisby an amount equal to the sustained torsional position ofthe eyes (OCR) during tilts of the GIA in the roll plane(Haslwanter et al. 1992; Bockisch and Haslwanter 2001;Moore et al. 2001). Pursuit eye movements were confinedto this shifted plane in accordance with Listing’s law(Haslwanter et al. 1991, 1992), and there were no cross-coupled eye velocity components during horizontal pursuiton Earth or in-flight. Thus, pursuit did not exhibit spatialorientation toward the GIA. This is similar to the lack ofspatial orientation of the direct vestibular and optokineticpathways in monkeys (Wearne et al. 1998). Horizontal andvertical pursuit gains were unchanged in microgravity, butthere was a significant increase in saccades in-flight,particularly in the vertical direction, as noted in a previousspace study (Andre-Deshays et al. 1993). Also consistentwith previous studies (Baloh et al. 1988; Grasse andLisberger 1992), pursuit was smoother (less saccadic) inthe horizontal than in the vertical direction on Earth.
In conclusion, results from the Neurolab studies indicatethat spatial orientation and the corresponding tilts of theeyes to low frequency linear accelerations can be wellmaintained in space as on Earth. This holds true for OKN,OCR (Moore et al. 2001), for the perception of tilts of theGIA with regard to the head (Clément et al. 2001), andpossibly also for vestibulo-sympathetic reflexes (Moore etal. 2001, 2004). A practical outcome of this study is thatintermittent short radius centrifugation may help tomaintain the gain of otolith-ocular pathways and spatialorientation during long-duration space flight.
Acknowledgements We gratefully acknowledge the efforts of theNeurolab payload crew, Drs. Rick Linnehan, Dave Williams, JayBuckey and Jim Pawelczyk, for serving as subjects and operatorsduring the mission; the alternate payload specialists, Drs. AlexDunlap and Chiaki Mukai; Dr. Mel Buderer, Dan Harfe, GwennSandoz and Nasser Ayub (JSC); Jacqui Van Tweste (KSC); MikeCork and Dr. Thierry Dewandre (ESA/ESTEC); Frederic Bellossi(Aerospatiale); Dr. Ian Curthoys and Hamish MacDougall (Uni-versity of Sydney); Dr. Izumi Koizuka (St. Mariana University); Dr.Takeshi Kubo (Osaka University). Supported by NASA ContractsNAS 9-19441, NCC 9-128 and NNJ04HF51G (Drs. Moore andCohen) and the Centre National d’Etudes Spatiales (Drs. Clémentand Berthoz).
References
Andre-Deshays C, Israel I, Charade O, Berthoz A, Popov K,Lipshits M (1993) Gaze control in microgravity. 1. Saccades,pursuit and eye-head coordination. J Vestib Res 3:331–343
Angelaki DA, Wei M, Merfeld DM (2001) Vestibular discriminationof gravity and translational acceleration. Ann NY Acad Sci942:114–127
Angelaki DE, Hess BJM (1994) Inertial representation of angularmotion in the vestibular system of rhesus monkeys. I.Vestibuloocular reflex. J Neurophysiol 71:1222–1249
Angelaki DE, Hess BJM (1995) Inertial representation of angularmotion in the vestibular system of rhesus monkeys. II. Otolith-controlled transformation that depends on an intact cerebellarnodulus. J Neurophysiol 73:1729–1751
Angelaki DE, McHenry MQ, Dickman D, Newlands SD, Hess BJM(1999) Computation of inertial motion: neural strategies toresolve ambiguous otolith information. J Neuroscience 19:316–327
Angelaki DE, Merfeld DM, Hess BJM (2000) Low frequencyotolith and semicircular canal interactions after canal inactiva-tion. Exp Brain Res 132:539–549
Baloh R, Yee R, Honrubia V, Jacobson K (1988) A comparison ofthe dynamics of horizontal and vertical smooth pursuit innormal human subjects. Aviat Space Environ Med 59:121–124
Benson AJ (1974) Modification of the response to angularaccelerations by linear accelerations. In: Kornhuber H (ed)Handbook of sensory physiology. Springer, Berlin, pp 281–320
Berthoz A, Brandt T, Dichgans J, Probst T, Bruzek W, Vieville T(1986) European vestibular experiments on the Spacelab-1mission: 5. Contribution of the otoliths to the vertical vestibulo-ocular reflex. Exp Brain Res 64:272–278
Bockisch CJ, Haslwanter T (2001) Three-dimensional eye positionduring static roll and pitch in humans. Vis Res 41:2127–2137
Borries GVT (1926) Fixation and nystagmus. Linds, CopenhagenBrandt T (1999) Vertigo. Springer-Verlag, LondonBrucher JM (1964) L’aire oculogyre frontale du singe. Ses fonctions
et ses voies efferents. In: Editions Arscia. Librairie maloine,Bruxelles-Paris
Chun K-S, Robinson DA (1978) A model of quick phase generationin the vestibuloocular reflex. Biol Cybern 28:209–221
Clarke AH, Grigull J, Mueller R, Scherer H (2000) The three-dimensional vestibulo-ocular reflex during prolonged micro-gravity. Exp Brain Res 134:322–334
Clément G (1998) Alteration of eye movements and motionperception in microgravity. Brain Res Rev 28:161–172
Clément G, Lathan CE (1991) Effect of static tilt about the roll axison horizontal and vertical optokinetic nystagmus and optoki-netic after-nystagmus in humans. Exp Brain Res 84:335–341
Clément G, Reschke MF (1996) Neurosensory and sensory-motorfunctions. In: Moore D, Bie P, Oser H (eds) Biological andmedical research in space: an overview of life sciences researchin microgravity. Springer-Verlag, Heidelberg, pp 178–258
Clément G, Reschke MF, Verrett CM, Wood SJ (1992a) Effects ofgravitational force variation on optokinetic nystagmus and onperception of visual stimulus orientation. Aviat Space EnvironMed 63:771–777
Clément G, Wood SJ, Reschke MF (1992b) Effects of microgravityon the interaction of vestibular and optokinetic nystagmus inthe vertical plane. Aviat Space Environ Med 63:778–784
Clément G, Moore ST, Raphan T, Cohen B (2001) Perception of tilt(somatogravic illusion) in response to sustained linear acceler-ation during space flight. Exp Brain Res 138:410–418
Cohen B, Matsuo V, Raphan T (1977) Quantitative analysis of thevelocity characteristics of optokinetic nystagmus and optoki-netic after-nystagmus. J Physiol (Lond) 270:321–344
Cohen B, Raphan T, Solomon D, Helwig D, Cohen N, KozlovskayaIB, Sirota MG, Yakushin SB (1991) Effects of spaceflight in the“Cosmos” biosatellite 2044 on the VOR of Rhesus Monkey.Proceedings: International Meeting “Biosatellite Cosmos”,Leningrad 12–15 August. IMBP, Moscow, pp 113
Cohen B, Maruta J, Raphan T (2001) Orientation of the eyes togravito-inertial acceleration. Ann NY Acad Sci 942:241–258
Collewijn H (1969) Optokinetic eye movements in the rabbit: input-output relations. Vision Res 9:117–132
Crawford JD, Vilis T (1991) Axes of eye rotation and Listing’s lawduring rotations of the head. J Neurophysiol 65:407–423
Dai M, Raphan T, Cohen B (1991) Spatial orientation of thevestibular system: dependence of optokinetic after nystagmuson gravity. J Neurophysiol 66:1422–1438
Dai M, McGarvie L, Kozlovskaya IB, Raphan T, Cohen B (1994)Effects of spaceflight on ocular counterrolling and spatialorientation of the vestibular system. Exp Brain Res 102:45–56
57

Dai M, Raphan T, Kozlovskaya I, Cohen B (1996) Modulation ofvergence by off-vertical yaw axis rotation in the monkey:normal characteristics and effects of space flight. Exp Brain Res111:21–29
Fetter M, Heimberger J, Black RA, Hermann W, Sievering F,Dichgans J (1996) Otolith-semicircular canal interaction duringpostrotatory nystagmus in humans. Exp Brain Res 108:463–473
Fick A (1854) Die Bewegungen des menschlichen Augapfels. ZRationelle Med 4:101–128
Gizzi M, Raphan T, Rudolph S, Cohen B (1994) Orientation ofhuman optokinetic nystagmus to gravity: a model basedapproach. Exp Brain Res 99:347–360
Goldstein H (1980) Classical Mechanics. Reading, MA, Addison-Wesley
Grasse K, Lisberger S (1992) Analysis of naturally occurringasymmetry in vertical smooth pursuit in the monkey. JNeurophysiol 67:164–179
Harris LR (1987) Vestibular and optokinetic eye movements evokedin the cat by rotation about a tilted axis. Exp Brain Res 66:522–532
Harris LR, Barnes GR (1987) Orientation of vestibular nystagmus ismodified by head tilt. In: Graham MD, Kemink JL (eds) Thevestibular system: neurophysiologic and clinical research.Raven Press, New York, pp 539–548
Haslwanter T (1995) Mathematics of three-dimensional eyerotations. Vision Res 35:1727–1739
Haslwanter T, Straumann D, Hepp K, Hess BJM, Henn V (1991)Smooth pursuit eye movements obey Listing’s law in themonkey. Exp Brain Res 87:470–472
Haslwanter T, Straumann D, Hess BJM, Henn V (1992) Static rolland pitch in the monkey: shift and rotation in Listing’s plane.Vis Res 32:1341–1348
Helmholz HV (1867) Handbuch der Physiologichen Optik. LeopoldVoss, Leipzig
Hess BJM, Angelaki DA (1999) Inertial processing of vestibulo-ocular signals. Ann NY Acad Sci 871:148–161
Hood JD (1967) Observations upon the neurological mechanism ofoptokinetic nystagmus with special reference to the contribu-tion of peripheral vision. Acta Otolaryngol (Stockh) 63:208–215
Hood JD, Leech J (1974) The significance of peripheral vision in theperception of movement. Acta Otolarygol (Stockh) 1974:73–79
Imai T, Moore ST, Raphan T, Cohen B (2001) Interaction of thebody, head and eyes during walking and turning. Exp Brain Res136:1–18
Kushiro K, Dai M, Kunin M, Yakushin SB, Cohen B, Raphan T(2002) Compensatory and orienting eye movements induced byoff-vertical axis Rotation (OVAR) in monkeys. J Neurophysiol88:2445–2462
Markham CH (1989) Anatomy and physiology of otolith-controlledocular counterrolling. Acta Otolaryngol Suppl 468:263–266
Mayne R (1974) A systems concept of the vestibular organs. In:Kornhuber HH (ed) Handbook of vestibular physiology.Vestibular System. Springer-Verlag, New York, pp 493–580
Merfeld DM (1995) Modeling the vestibulo-ocular reflex of thesquirrel monkey during eccentric rotation and roll tilt. ExpBrain Res 106:123–134
Merfeld DM, Young LR (1995) The vestibulo-ocular reflex of thesquirrel monkey during eccentric rotation and roll tilt. ExpBrain Res 106:111–122
Merfeld DM, Young LR, Paige GD, Tomko DL (1993) Threedimensional eye movements of squirrel monkeys followingpostrotatory tilt. J Vest Res 3:123–141
Merfeld DM, Zupan L, Peterka RJ (1999) Humans use internalmodels to estimate gravity and linear acceleration. Nature398:615–618
Misslisch H, Tweed D, Fetter M, Sievering D, Koenig E (1994)Rotational kinematics of the human vestibuloocular reflex. III.Listing’s law. J Neurophysiol 72:2490–2502
Mittelstaedt H, Glasauer S (1993) Illusions of verticality inweightlessness. Clin Investig 71:732–739
Moore ST, McCoy SG, Curthoys IS (1991) VTM—an imageprocessing system for measuring ocular torsion. Comp MethProg Biomed 35:219–230
Moore ST, Haslwanter T, Curthoys IS, Smith ST (1996) Ageometric basis for measurement of three-dimensional eyeposition using image processing. Vis Res 36:445–459
Moore ST, Clément G, Raphan T, Cohen B (2001) Ocular counter-rolling induced by centrifugation during orbital spaceflight.Exp Brain Res 137:323–335
Moore ST, Clément G, Dai M, Raphan T, Solomon D, Cohen B(2003) Ocular and perceptual responses to linear acceleration inmicrogravity: alterations in otolith function on the COSMOSand Neurolab flights. J Vest Res 13:377–393
Moore ST, Diedrich A, Biaggioni I, Kaufmann H, Raphan T, CohenB (2004) Artificial gravity: a possible countermeasure for post-flight orthostatic intolerance. Acta Astronautica (in press)
Paige GD (1991) Linear vestibulo-ocular reflex (LVOR) andmodulation by vergence. Neurosci Lett 481[Suppl]: 282–286
Paige GD, Seidman SH (1999) Characteristics of the VOR responseto linear acceleration. Ann NY Acad Sci 871:123–135
Paige GD, Tomko DL (1991a) Eye movement responses to linearhead motion in the squirrel monkey. I. Basic Characteristics. JNeurophysiol 65:1170–1182
Paige GD, Tomko DL (1991b) Eye movement responses to linearhead motion in the squirrel monkey. II. Visual-vestibularinteractions and kinematic considerations. J Neurophysiol65:1183–1196
Parker DE, Reschke MF, Arrott AP, Homick JL, Lichtenberg BK(1985) Otolith tilt translation reinterpretation following pro-longed weightlessness: implications for preflight training. AviatSpace Environ Med 56:601–607
Purkinje JE (1820) Beiträge zur näheren Kenntnis des Schwindelsaus heutognostischen Daten. Med JB (Wien) 6:79–125
Rademacher GGJ, Ter Braak JWG (1948) On the central mechanismof some optic reactions. Brain 71:48–76
Raphan T (1998) Modeling control of eye orientation in threedimensions. I. Role of muscle pulleys in determining saccadictrajectory. J Neurophysiol 79:2653–2667
Raphan T, Cohen B (1988) Organizational principles of velocitystorage in three dimensions: the effect of gravity on cross-coupling of optokinetic after-nystagmus. Ann NY Acad Sci545:74–92
Raphan T, Cohen B (2002) The vestibulo-ocular reflex (VOR) inthree dimensions. Exp Brain Res 145:1–27
Raphan T, Sturm D (1991) Modelling the spatiotemporal organiza-tion of velocity storage in the vestibuloocular reflex byoptokinetic studies. J Neurophysiol 66:1410–1420
Raphan T, Matsuo V, Cohen B (1979) Velocity storage in thevestibulo-ocular reflex arc (VOR). Exp Brain Res 35:229–248
Raphan T, Cohen B, Henn V (1981) Effects of gravity on rotatorynystagmus in monkeys. Ann NY Acad Sci 374:44–55
Raphan T, Wearne S, Cohen B (1996) Modeling the organization ofthe linear and angular vestibulo-ocular reflexes. Ann NY AcadSci 781:348–363
Rashbass C (1961) The relationship between saccadic and smoothtracking eye movements. J Physiol 159:326–338
Robinson D (1977) Vestibular and optokinetic symbiosis: anexample of explaining by modeling. In: Baker R, Berthoz A(eds) Control of gaze by brain stem neurons. Elsevier/NorthHolland, Amsterdam, pp 49–58
Seidman SH, Telford L, Paige GD (1998) Tilt perception duringdynamic linear acceleration. Exp Brain Res 119:307–314
Solomon D, Straumann D, Zee DS (1997) Three-dimensional eyemovements during vertical axis rotation: effects of visualsuppression, orbital eye position and head position. In: FetterM, Haslwanter T, Misslisch H, Tweed D (eds) Three-dimen-sional kinematics of eye, head and limb movements. Harwoodacademic publishers, The Netherlands, pp 197–208
Ter Braak JWG (1936) Untersuchungen ueber optokinetischenNystagmus. Archives Neerlander Physiologie 21:309–376
58

Thurtell MJ, Black RA, Halmagyi GM, Curthoys IS, Aw ST (1999)Vertical eye position-dependence of the human vestibuloocularreflex during passive and active yaw head rotations. JNeurophysiol 81:2415–2428
Thurtell MJ, Kunin M, Raphan T (2000) Role of muscle pulleys inproducing eye position-dependence in the angular vestibuloo-cular reflex: a model-based study. J Neurophysiol 84:639–650
Tweed D, Vilis T (1987) Implications of rotational kinematics forthe oculomotor system in three dimensions. J Neurophysiol58:832–849
Tweed D, Vilis T (1990) Geometric relations of eye position andvelocity vectors during saccades. Vision Res 30:111–127
Uchino Y, Sasaki M, Sato H, Imagawa M, Suwa H, Isu N (1996)Utriculoocular reflex arc of the cat. J Neurophysiol 76:1896–1903
Uchino Y, Sato H, Sasaki M, Ikegami H, Isu N, Graf W (1997)Sacculocollic reflex arcs in cats. J Neurophysiol 77:3003–3012
Wearne S, Raphan T, Cohen B (1997) Contribution of vestibularcommissural pathways to velocity storage and spatial orienta-tion of the angular vestibuloocular reflex. J Neurophysiol78:1193–1197
Wearne S, Raphan T, Cohen B (1998) Control of spatial orientationof the angular vestibuloocular reflex by the nodulus and uvula.J Neurophysiol 79:2690–2715
Yakushin SB, Dai MJ, Suzuki J-I, Raphan T, Cohen B (1995)Semicircular canal contribution to the three-dimensional ves-tibulo-ocular reflex: a model-based approach. J Neurophysiol74:2722–2738
Yakushin SB, Gizzi M, Reisine H, Raphan T, Buttner-Ennever J,Cohen B (2000) Functions of the nucleus of the optic tract(NOT). II. Control of ocular pursuit. Exp Brain Res 131:416–432
Young LR, Oman CM, Watt DGD, Money KE, Lichtenberg BK(1984) Spatial orientation and readaptation to Earth’s gravity.Science 225:205–208
Zhu D, Moore ST, Raphan T (1999) Robust pupil center detectionusing a curvature algorithm. Comp Meth Prog Biomed 59:145–157
Zhu D, Moore ST, Raphan T (2004) Robust and real-time torsionaleye position calculation using a template matching technique.Comp Meth Prog Biomed 74:201–209
59



















![Alain Bashung - Alain Bashung [PVC Book]](https://img.dokumen.tips/doc/110x75/55cf868e550346484b98cd83/alain-bashung-alain-bashung-pvc-book.jpg)









