8/10/2019 Stem cells article.pdf
5/7
can often only guess what the effects of such mutations will be
onthe actual cells involved in particular cancers. This has
handicappedour ability to translate our identification of mutations
into newtherapies.
A tumour can be viewed as an aberrant organ initiated by
atumorigenic cancer cell that acquired the capacity for
indefiniteproliferation through accumulated mutations. If one views
a tumouras an abnormal organ, then the principles of normal stem
cellbiology12,44 can be applied to understand better how tumours
develop(reviewed in ref. 45). In fact, many observations suggest
thatanalogies between normal stem cells and tumorigenic cells may
beappropriate. Both normal stem cells and tumorigenic cells
haveextensive proliferative potential and the ability to give rise
to new(normal or abnormal) t issues. Both tumours and normal
tissues arecomposed of heterogeneous combinations of cells, with
differentphenotypic characteri stics and di fferent proliferative
potentials 4649 .Because most tumours have a clonal origin 5052 ,
tumorigenic cancercells must give rise to phenotypically diverse
progeny, includingcancer cells with indefinite proliferative
potential, as well as cancercells with limited or no proliferative
potential. This suggests thattumorigenic cancer cells undergo
processes that are analogous to theself-renewal and differentiation
of normal stem cells.
Although some of the heterogeneity in tumours arises as a
result
of continuing mutagenesis, it is likely that heterogeneity also
arisesthrough the aberrant differentiation of cancer cells. It is
welldocumented that many types of tumours contain cancer cells
withheterogeneous phenotypes reflecting aspects of the
differentiationthat normally occurs in the tissues from which the
tumours arise.The variable expression of normal differentiation
markers by cancercells in a tumour suggests that some of the
heterogeneity in tumoursari ses as a result of the anomalous
differentiation of tumour cells.Examples of this include the
variable expression of myeloid markersin chronic myeloid leukaemia,
the variable expression of neuronalmarkers within peripheral
neurectodermal tumours, and thevariable expression of milk proteins
or the oestrogen receptorwithin breast cancer.
In other words, both normal stem cells and tumorigenic cells
give
rise to phenotypically heterogeneous cells that exhibit
variousdegrees of differentiation. Thus, tumorigenic cells can be
thought of as cancer stem cells that undergo an aberrant and poorly
regulatedprocess of organogenesis analogous to what normal stem
cells do. It isperhaps not surprising that tumorigenic cells behave
in ways that areanalogous to normal stem cells given that cancer
cells tend to displayfunctional and phenotypic attributes of the
normal cells from whichthey are derived 28.
Evidence for cancer stem cellsIt was first extensively
documented for leukaemia and multiplemyeloma that only a small
subset of cancer cells is capable of extensiveproliferation. For
example, when mouse myeloma cells wereobtained from mouse ascites,
separated from normal haematopoiet-
ic cells and put in clonal in vitro colony-forming assays, only
1 in10,000 to 1 in 100 cancer cells were able to form colonies 53.
Even whenleukaemic cells were transplanted in vivo , only 14% of
cells couldform spleen colonies 5456 . Because the differences in
clonogenicityamong the leukaemia cells mirrored the differences in
clonogenicityamong normal haematopoietic cells, the clonogenic
leukaemic cellswere described as leukaemic stem cells (for example,
see ref. 53). Buttwo formal possibil ities remained: either all
leukaemia cells had a lowprobability of proliferating extensively
in these assays such that allleukaemia cells had the potential to
behave as leukaemic stem cells, ormost leukaemia cells were unable
to proliferate extensively and only asmall , definable subset of
cells was consistently clonogenic.
To prove the second possibility, it would be necessary to
separatedifferent classes of leukaemia cell and show that one
subset is highlyenriched for clonogenic capacity and all other
cells are greatlydepleted for clonogenicity. This has been
accomplished by Dick and
colleagues 57, who showed that human AML stem cells could
beidentified prospectively and purified as CD34 +CD38 cells
frompatient samples. Despite the fact that these cells represented
a smallbut variable proportion of AML cells (0.2% in one patient) ,
they werethe only cells capable of transferring AML from human
patients toNOD/SCID mice in the vast majority of cases. This
excluded the firstpossibility that all AML cells had a similar
clonogenic capacity, andshowed that a small, predictable subset was
consistently enriched forthe ability to proli ferate and transfer
disease.
It has also been shown for solid cancers that the cells are
pheno-typically heterogeneous and that only a small proportion of
cells areclonogenic in culture and in vivo 4649,58 . For example,
only 1 in 1,000to 1 in 5,000 lung cancer, ovarian cancer or
neuroblastoma cells werefound to form colonies in soft agar 59.
Just as in the context of leukaemic stem cells, these observations
led to the hypothesis thatonly a few cancer cells are actually
tumorigenic and that thesetumorigenic cells could be considered as
cancer stem cells 59. But, asexplained above, two possibil ities
remain: either all solid cancer cellshave a low probability of
proliferating extensively and behaving inclonogenic assays as
cancer stem cells, or most cancer cells have only alimited proli
ferative potential and cannot behave as cancer stem cells,but a
small, definable subset of cells is enriched for the ability
toproliferate extensively and form tumours.
In both cases, some of the cancer cell heterogeneity would arise
asa result of environmental differences within the tumour
andcontinuing mutagenesis. The essential difference between
thesepossibil ities is the prediction, according to the second
possibili ty, thatwhatever the environment or mutational status of
the cells, only asmall, phenotypically distinct subset of cancer
cells has the ability toproliferate extensively or form a new
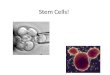
tumour (Fig. 4). It has not beenpossible to distinguish between
these models of solid cancer hetero-geneity, because as yet no one
has published the identity of purifiedsubsets of uncultured solid
cancer cells that are enriched for theabili ty to form new
tumours.
The implications of solid cancer stem cellsIf the growth of
solid cancers were driven by cancer stem cells, itwould have
profound implications for cancer therapy. At present, allof the
phenotypically diverse cancer cells are treated as though they
insight review articles
NATURE | VOL 414 |1 NOVEMBER 2001 | www.nature.com 10 9
Tumou r ce lls a reheterogeneous, but mostcells can
proliferateextensively and formnew tumours
Tumour ce lls a re heteroge neousand only the cancer stem
cellsubs et (CSC; yellow) has the a bilityto proliferate
extensively andform new tumours
a bCS C
CS C
CS C
CS C
Figure 4Two general m odels of heterogeneity in solid cancer c
ells.a, Cancer cells ofmany different phenotypes have the pot
ential to proliferate extensively, but acell would have a low
probability of exhibiting this potential in an assay
ofclonogenicity or tumorigenicity.b, Most c ancer cells have only
limited proliferativpotential, but a subset of cancer c ells
consistently proliferate extensively inclonogenic assays and can
form new tumours on transplantation. The modelinb predicts that a
distinct subset of cells is enriched for the ability to form ntumou
rs, whereas most c ells are depleted of this ability. Existing
therapeutic
approaches have been based largely on the model show n ina, but
the failure ofthese therapies to cure most solid cancers suggests
that the model shown inb maybe more accurate.
2001 Macmillan Magazines Ltd
8/10/2019 Stem cells article.pdf
7/7
self-renewal of normal stem cells, studies of stem cell biology
arelending insight into the origins of cancer and will ultimately
yieldnew approaches to fight this disease.
1. Spangrude, G. J., Heimfeld, S. & Weissman, I. L. Puri fi
cation and characteri zation of mousehematopoieti c stem cell s.
Science 241, 5862 (1988).
2. Morr ison, S. J. & Weissman, I. L. The long-term r
epopulating subset of hematopoietic stem cells isdeterministic and
isolatable by phenotype. Immunity 1, 661673 (1994).
3. Baum, C. M., Weissman, I. L., Tsukamoto, A. S., Buckle, A. M.
& Peault, B. Isolati on of a candidatehuman hematopoietic
stem-cell population. Proc. Natl Acad. Sci. USA 89, 28042808
(1992).
4. Osawa, M., Hanada, K., Hamada, H. & Nakauchi, H.
Long-term lymphohematopoietic
reconstitution by a single CD34-low/negative hematopoietic stem
cell. Science 273, 242245 (1996).5. Akashi, K. & Weissman, I.
L. in Developmental Biol ogy of Hematopoiesis (ed. Zon, L. I.) 1534
(Oxford
Univ. Press, New York, 2001).6. Petersen, B. E. et al. Bone
marrow as a potential source of hepatic oval cells. Science 284,
11681170 (1999).7. Brazelton, T. R., Rossi, F. M. V., Keshet, G. I.
& Blau, H. M . From marrow to brain: expression of
neuronal phenotypes in adult mice. Science 290, 17751779
(2000).8. Mezey, E., Chandross, K. J., Harta, G., Maki, R. A. &
McKercher, S. R. Turni ng blood into br ain: cells
beari ng neuronal anti gens generated in vivo from bone marrow.
Science 290, 17791782 (2000).9. Lagasse, E. et al. Purifi ed
hematopoietic stem cells can differentiate to hepatocytes in vivo .
Nature
Med. 6, 12291234 (2000).10.Krause, D. S. et al. Multi-organ,
multi-lineage engraftment by a single bone marrow derived stem
cell.
Cell 105, 369377 (2001).11.Morrison, S. J., Wandycz, A. M.,
Hemmati, H. D., Wright, D. E. & Weissman, I. L. Identification
of a
lineage of mult ipotent hematopoietic progenitor s. Development
124, 19291939 (1997).12.Weissman, I. L. Translating stem and
progenitor cell biology to the clini c: barr iers and oppor tuni
ties.
Science 287, 14421446 (2000).13.Mi ll er, C. L. & Eaves, C.
J. Expansion in vitro of adult muri ne hematopoietic stem cells
with tr ansplantable
lympho-myeloid reconstituting ability. Proc. Natl Acad. Sci. USA
94, 1364813653 (1997).
14.Domen, J., Gandy, K. L. & Weissman, I. L. Systemic
overexpression of BCL-2 in t he hematopoieti c systemprotects
transgenic mice from the consequences of lethal ir radiation. Blood
91, 22722282 (1998).
15.Domen, J. & Weissman, I . L. Hematopoi etic stem cell s
need two signals to pr event apoptosis; BCL-2can provide one of
these, Kit l/c-Ki t signaling the other. J. Exp. M ed. 192,
17071718 (2000).
16.Taipale, J. & Beachy, P. A. The Hedgehog and Wnt
signaling pathways in cancer. Nature 411,349354 (2001).
17.Varnum-Finney, B. et al. Pluripotent, cytokine-dependent,
hematopoietic stem cells are immortalizedby constitutive Notch1
signaling. Nature Med. 6, 12781281 ( 2000).
18.Karanu, F. N. et al. The Notch l igand Jagged-1 r epresents a
novel growth factor of humanhematopoieti c stem cell s. J. Exp. M
ed. 192, 13651372 (2000).
19.Bhardwaj, G. et al. Sonic hedgehog induces the proliferation
of pr imi tive human hematopoietic cellsvia BMP regulation. Nature
Immunol. 2, 172180 (2001).
20.Nusse, R. & Varmus, H. E. Many tumor s induced by the
mouse mammary tumor vir us contain aprovi rus integrated in the
same region of t he host genome. Cell 31, 99109 (1982).
21.Cadigan, K. M. & Nusse, R. Wnt signali ng: a common t
heme in animal development. Genes Dev. 11,32863305 (1997).
22.Reya, T. et al. Wnt signaling regulates B lymphocyte
proliferation through a LEF-1 dependentmechanism. Immunity 13, 1524
(2000).
23.Austin, T. W., Solar, G. P., Ziegler, F. C., Liem, L. &
Matthews, W. A rol e for the Wnt gene family inhematopoiesis:
expansion of multilineage progenitor cells. Blood 89, 36243635
(1997).
24.Van Den Berg, D. J., Sharma, A. K., Bruno, E. & H offman,
R. Role of members of the Wnt gene familyin human hematopoiesis.
Blood 92, 31893202 (1998).
25.Zhu , A. J. & Watt , F. M. -catenin signalli ng modulates
proliferative potential of human epidermalkeratinocytes
independently of intercellular adhesion. Development 126, 22852298
(1999).
26.Gat, U., DasGupta, R., Degenstein, L. & Fuchs, E. De novo
hair foll icle morphogenesis and hairtumors in mi ce expressing a
tr uncated -catenin in skin. Cell 95, 605614 (1998).
27.Korinek, V. et al. Depletion of epithelia stem-cell
compartments in the small i ntestine of mice lackingTcf-4. Natu re
Genet. 19, 15 (1998).
28.Sell, S. & Pierce, G. B. Maturation arrest of stem cell
differentiation is a common pathway for thecellular origin of
teratocarcinomas and epithelial cancers. Lab. I nvest. 70, 622
(1994).
29.Sawyers, C., Denny, C. & Wit te, O. Leukemia and the
disrupt ion of nor mal hematopoiesis. Cell 64,337350 (1991).
30.Kondo, M., Weissman, I. L. & Akashi, K . Identifi cation
of clonogenic common lymphoid progenitorsin mouse bone marrow. Cell
91, 661672 (1997).
31.Bonnet, D. & Dick, J. E. Human acute myeloid l eukemia is
organized as a hierarchy that originatesfrom a pri mit ive
hematopoietic cell. Nature M ed. 3, 730737 (1997).
32.Miyamoto, T., Weissman, I. L. & Akashi, K. AM
L1/ETO-expressing nonleukemic stem cells in acutemyelogenous
leukemia with 8;21 chromosomal translocation. Proc. Natl Acad. Sci.
U SA 97,75217526 (2000).
33.George, A. A. et al. Detection of leukemic cells in the CD34
+CD38 bone marrow progenitorpopulation in children wi th acute
lymphoblastic l eukemia. Blood 97, 39253930 (2001).
34.Mauro, M. J. & Druker, B. J. Chronic myelogenous
leukemia. Curr. Opin. On col. 13, 37 (2001).35.Lagasse, E. &
Weissman, I. L. bcl-2 inhibits apoptosis of neutrophils but not
their engulfment by
macrophages. J. Exp. M ed. 179, 10471052 ( 1994).36.Traver, D.,
Akashi, K., Weissman, I . L. & Lagasse, E. Mice defective in
two apoptosis pathways in the
myeloid li neage develop acute myeloblasti c leukemia. Immunity
9, 4757 (1998).37.Polakis, P. Wnt signali ng and cancer. Genes Dev.
14, 18371851 ( 2000).38.Tsukamoto, A., Grosschedl, R., Guzman, R.,
Parslow, T. & Varmus, H. E. Expression of the int- 1 gene
in tr ansgenic mice is associated with mammary gland hyperplasia
and adenocarcinomas in male andfemale mice. Cell 55, 619625
(1988).
39.Weissman, I. L. & Baird, S. in Neoplasti c Transformat
ion: Mechanisms and Consequences (ed.Korprowski, H.) 135152 (Dahlem
Conferenzen, Berlin, 1977).
40.McGrath, M. S. & Weissman, I. L. AKR leukemogenesis:
identification and biological significance of thymic lymphoma
receptors for AKR retroviruses. Cell 17, 6575 (1979).
41.McGrath, M. S., Pill emer, E. & Weissman, I. L. Muri ne
leukaemogenesis: monoclonal antibodies to T-cell determinants
arrest T-lymphoma cell proli feration. Nature 285, 259261
(1980).
42.Quinn, E. R. et al. The B cell receptor of a hepatit is C
virus associated non-Hodgki ns lymphomabinds the viral E-2 envelope
protein, i mpli cati ng immunoglobul in activation in
lymphomagenesis.Blood (in the press).
43.ONeill, H. C., McGrath, M . S., Alli son, J. P. &
Weissman, I. L. A subset of T cell receptors associated withL3T4
molecules mediates C6VL leukemia cell bindi ng of i ts cognate
retrovir us. Cell 49, 143151 (1987).
44.Morr ison, S. J., Shah, N. M. & Anderson, D. J. Regulator
y mechanisms in stem cell biology. Cell 88,287298 (1997).
45.Kummermehr, J. & Trott, K.-R. i n Stem Cell s (ed.
Potten, C. S.) 363399 (Academic, New York, 1997).46.Fidl er, I. J.
& Kri pke, M. L. Metastasis result s from preexisti ng vari ant
cell s wit hin a malignant t umor.
Science 197, 893895 (1977).47.Fidl er, I. J. & Hart , I. R.
Biological diversity in metastati c neoplasms: origins and impli
cations. Science
217, 9981003 (1982).48.Heppner, G. H. Tumor heterogeneity. Can
cer Res. 44, 22592265 (1984).49.Nowell , P. C. Mechanisms of tumor
progression. Cancer Res. 46, 22032207 (1986).50.Nowell , P. C. A
minute chromosome in human granulocytic leukemia. Science 132, 1497
(1960).51.Fialkow, P. J. Clonal origin of human tumors. Biochim. Bi
ophys. Acta 458, 283321 (1976).52.Fearon, E. R., Hamilton, S. R.
& Vogelstein , B. Clonal analysis of human color ectal t umor
s. Science
238, 193197 (1987).53.Park, C. H., Bergsagel, D. E. & McCull
och, E. A. Mouse myeloma tumor stem cells: a primary cell
culture assay. J. Natl Cancer I nst. 46, 411422 (1971).54.Bruce,
W. R. & Gaag, H. v. d. A quant it ative assay for the number of
muri ne lymphoma cell s capable
of proli feration in vivo . Nature 199, 7980 (1963).55.Wodinsky,
I ., Swiniarski, J. & Kensler, C. J. Spleen colony studies of
leukemia L1210. I . Growth
kinetics of lymphocytic L1210 cells in vivo as determined by
spleen colony assay. Cancer Chemother.Rep. 51, 415421 (1967).
56.Bergsagel, D. E. & Valeriote, F. A. Growth characteri
stics of a mouse plasma cell tumor. Can cer Res. 28,21872196
(1968).
57.Bonnet, D. & Dick, J. E. Human acute myeloi d leukemia is
organized as a hierarchy that or iginatesfrom a pr imi tive
hematopoietic cell. Nature M ed. 3, 730737 (1997).
58.Southam, C. M. & Brunschwig, A. Quantitative studies of
autotransplantation of human cancer.Cancer 14, 971978 (1961).
59.Hamburger, A. W. & Salmon, S. E. Primary bioassay of
human tumor stem cell s. Science 197,461463 (1977).
60.Salsbury, A. J. The signi fi cance of the circulati ng cancer
cell . Cancer Treatment Rev. 2, 5572 (1975).61.Williams, S. D.
Treatment of disseminated germ cell tumors with cisplatin,
bleomycin, and either
vinblastine or etoposide. N. Engl. J. Med. 316, 14351439
(1987).62.Stockler, M., Wi lcken, N. R. C., Ghersi, D. & Simes,
R. J. Systematic reviews of chemotherapy and
endocrine therapy in metastatic breast cancer. Cancer Treatment
Rev. 26, 151168 (2000).63.Lippman, M. E. High-dose chemotherapy
plus autologous bone marrow transplantation for
metastatic breast cancer. N. Engl. J. Med. 342, 11191120
(2000).64.Harrison, D. E. & Lerner, C. P. Most primitive
hematopoietic stem cells are stimulated to cycle rapidly
after treatment with 5-fl uorouracil. Blood 78, 12371240 (
1991).65.Bouwens, L. & DeBlay, E. Islet morphogenesis and stem
cell markers in rat pancreas. J. Histochem.
Cytochem. 44, 947951 (1996).66.Peters, R., Leyvraz, S. &
Perey, L. Apoptoti c regulati on in primit ive hematopoi etic
precursors. Blood
92, 20412052 (1998).67.Domen, J., Gandy, K. L. & Weissman,
I. L. Systemic overexpression of BCL-2 in the hematopoi etic
system
protects transgenic mice from the consequences of lethal ir
radiation. Blood 91, 22722282 (1998).68.Feuerhake, F., Sigg, W.,
Hoft er, E. A., Di mpfl , T. & Welsch, U. Immunohi stochemical
analysis of Bcl-2
and Bax expression in relation to cell turnover and epithelial
differenti ation markers in the non-lactati ng human mammary gland
epithelium. Cell Ti ssue Res. 299, 4758 (2000).
69.Zhou , S. et al. The ABC tr ansporter Bcrp1/ ABCG2 is
expressed in a wide variety of stem cells and is amolecular determi
nant of the side-populati on phenotype. Nature M ed. 7, 10281034
(2001).
70.Terskikh, A. V. et al. From hematopoiesis to neuropoiesis:
evidence of overl apping genetic programs.Proc. Natl Acad. Sci. U
SA 98, 79347939. (2001).
71.Bittner, M. et al. Mol ecular classification of cutaneous
mali gnant melanoma by gene expressionprofiling. Nature 406, 536540
(2000).
72.Perou, C. M. et al. Molecular port raits of human breast
tumours. Nature 406, 747752 (2000).73.Alizadeh, A. A. et al.
Distinct types of diffuse large B-cell lymphoma identified by gene
expression
profiling. Nature 403, 503511 (2000).74.Golub, T. R. Genome-wide
vi ews of cancer. N. Engl. J. Med. 344, 601602 (2001).75.Sgroi , D.
C. et al. In vivo gene expression profil e analysis of human breast
cancer progression. Cancer
Res. 59, 56565661 (1999).76.Leethanakul, C. et al. Distinct
pattern of expression of di fferentiation and growth-related genes
in
squamous cell carcinomas of the head and neck revealed by the
use of l aser capture microdissectionand cDNA arrays. Oncogene 19,
32203224 (2000).
77.Kogan, S. C. et al. BCL-2 cooperates with promyelocytic
leukemia retinoic acid receptor chimericprotein (PMLRAR ) to block
neutrophil differenti ation and ini tiate acute leukemia. J. Exp. M
ed. 193,531543 (2001).
78.Wechsler-Reya, R. J. & Scott , M. P. Control of neuronal
precursor proli ferati on in the cerebellum bySonic Hedgehog.
Neuron 22, 103114 (1999).
79.Zhang, Y. & Kalderon, D. Hedgehog acts as a somatic stem
cell factor i n the Drosophila ovary. Nature 410, 599604
(2001).
80.Henrique, D. et al. Maintenance of neuroepithelial progenitor
cells by Delta-Notch signalli ng in theembryonic chick retina.
Curr. Biol. 7, 661670 (1997).
81.Austin , J. & Kimble, J. glp-1 is required in the germ li
ne for regulation of t he decision between mitosisand meiosis in C.
elegans . Cell 51, 589599 (1987).
82.Chan, E. F., Gat, U., McNi ff, J. M. & Fuchs, E. A common
human skin tumour is caused by activati ngmutations in -catenin.
Natu re Genet. 21, 410413 (1999).
83.Wechsler-Reya, R. & Scott, M. P. The developmental
biology of brain tumors. Annu. Rev. Neurosci. 24,385428 (2001).
84.Gailani, M. R. & Bale, A. E. Acquired and inheri ted
basal cell carcinomas and the patched gene. Adv.Dermatol. 14,
261283 (1999).
85.Ell isen, L. W. et al. TAN-1, the human homolog of t he
Drosophila notch gene, is broken bychromosomal translocations in T
lymphoblastic neoplasms. Cell 66, 649661 (1991).
insight review articles
NATURE | VOL 414 |1 NOVEMBER 2001 | www.nature.com 11 1 2001 M
ill M g i Ltd

















![STEM CELLS EMBRYONIC STEM CELLS/INDUCED PLURIPOTENT STEM CELLS Stem Cells.pdf · germ cell production [2]. Human embryonic stem cells (hESCs) offer the means to further understand](https://img.dokumen.tips/doc/110x75/6014b11f8ab8967916363675/stem-cells-embryonic-stem-cellsinduced-pluripotent-stem-cells-stem-cellspdf.jpg)