Embed Size (px)
Citation preview

BIOTECHNOLOGY AND BIOENGINEERING VOL. VII, PAGES 279-253 (1965)
Starter Populations as Determined Under Steady State Conditions*
HOLGER W. .JAP\’KASCH, Woods Hole Oceanographic Institution, Woods Hole, Massachiaetts
Summary Continuous cultures of some heterotrophic Spirilli ( Sp . serpas , Sp. curvaturn,
Sp . s p . ) exhibit threshold concentrations of lactate, the growth limiting carbon and energy source, around 15 mg./l. This value corresponds to a threshold popiilation density of 0.92 mg. dry weight/l. a t a dilution rate of D = 0.185 hr.-I ( = 0.5 maximum growth rate). The minimum population density, which is regarded as equivalent to the starter population in batch culture, was dependent on the growth rate and the redox potential. The existence of minimum popu- lations and the pronounced effect of population density on growth at low nutrient levels seem to be widespread phenomena closely related to growth under sub- optimal conditions.
INTRODUCTION
In discussing the theoretical basis of continuous culture, usually i t is assumed that the overall culture conditions are not influenced by growth activities of the organisms. This ideal situation seeins to be described by the term “optimal” growth conditions. Most technical batch culture processes, however, start off under pronounced sub-optimal growth conditions which may bc obscured at high popula- tion densities. In such cases, growth is stiiiiulated, or inhibitory factors are overeoiiie, by metabolic activities of a population greater than a certain miriiiiiuiii inoculum. Such starter populations of several strains of hcterotrophic Spirilli were studied in a well-aerated mineral medium with the addition of lactate as the growth limiting carbon and energy source.
S61 of the National Science Foundation. * Contribution No. 1561 from the WHOI. Supported in part by Grant No.
279
280 H. W. JANNASCH
EXPERIMENT A L
Experiments in batch culture, employing different concentrations of washed cells, were impeded by cryptic growth, one part of the population feeding on autolytic products of the other. In chemostat experiments, Spirilli proved to be extremely advantageous in regard t.0 homogenous mixing, negligible wall growth, e t ~ . ~ When the
1 /
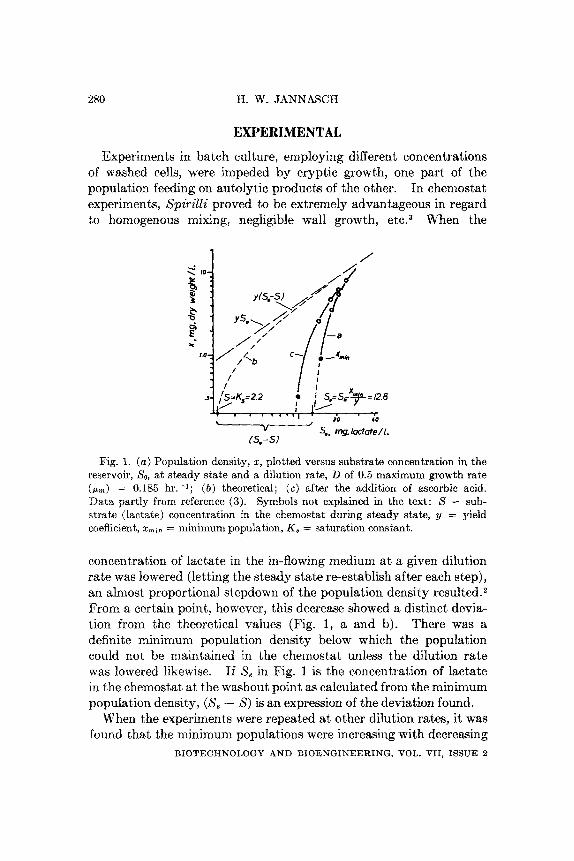
Fig. 1. ( a ) Population density, 2, plotted versus substrate concentration in the reservoir, So, a t steady state and a dilution rate, D of 0.5 maximum growth rate ( p m ) = 0.185 hr.-'; ( b ) theoretical; ( c ) after the addition of ascorbic acid. Data partly from reference (3). Symbols not explained in the text: S = sub- strate (lactate) concentration in the chemostat during steady state, y = yield coefficient, zmin = minimum population, K , = saturation constant.
concent.ration of lactate in the in-flowing medium at a given dilution rate was lowered (letting the steady state re-establish after each step), an almost proportional stepdown of the population density r e s ~ l t e d . ~ From a certain point, however, this decrease showed a distinct devia- tion from the theoretical values (Fig. 1, a and b). There was a definite minimum population density below which the population could not be maintained in the chemostat unless the dilution rate was lowered likewise. If S, in Fig. 1 is the concentration of lactate in the chemostat at the washout point as calculated from the minimum population densit,y, (S, - S ) is an expression of the deviation found.
When the experiments were repeated at other dilution rates, it was found that the minimum populations were increasing with decreasing
BIOTECHNOLOGY AND BIOENGINEERING, VOL. VII, ISSUE 2
STARTER POPULATIONS 28 1
r I 0.1
I I 7 /
(l/Se-S) Umg. Lacide
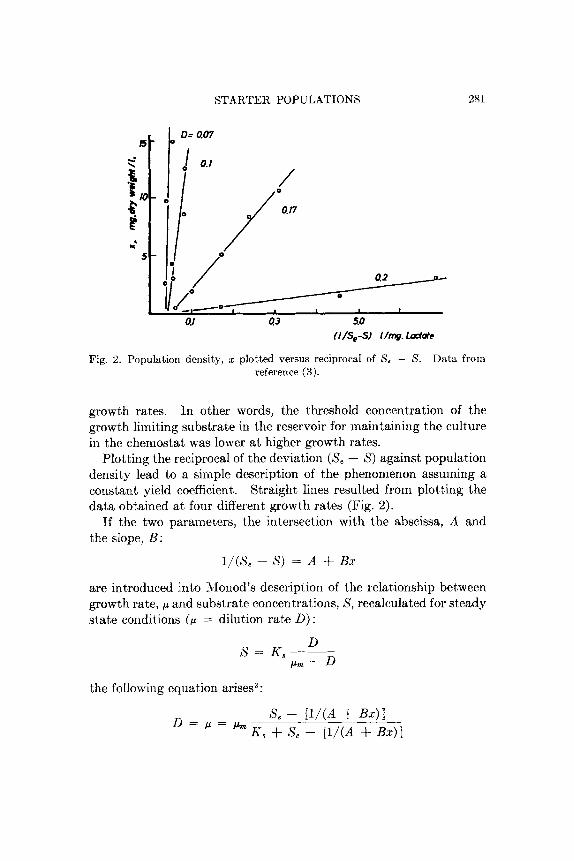
Fig. 2. Population density, x plotted versus reciprocal of S, - S. Data front reference ( 3 ) .
growth rates. In other words, the threshold concentration of the growth limiting substrate in the reservoir for maintaining the culture in the chemostat was lower at higher growth rates.
Plotting the reciprocal of the deviation (S, - S ) against population density lead to a siinple description of the phenomenon assuming a constant yield coefficient. Straight, lines resulted froin plotting the data obtained at four different growth raies (Fig. 2).
If the two parameters, the intersection with the abscissa, A and the slope, B :
l/(Se - S) = A + BX
are introduced into ;\lonod's description of the relationship between growth rate, p and substrate concentrations, S, recalculated for steady state conditions ( p = dilution rate 0) :
the following equation arises3:
282 H. W. JANNASCH
\ \ \ \ \ \
. h
I
0.07 0.1 Ql7 92 0.25 0-3 0.j D = p , hr.-‘
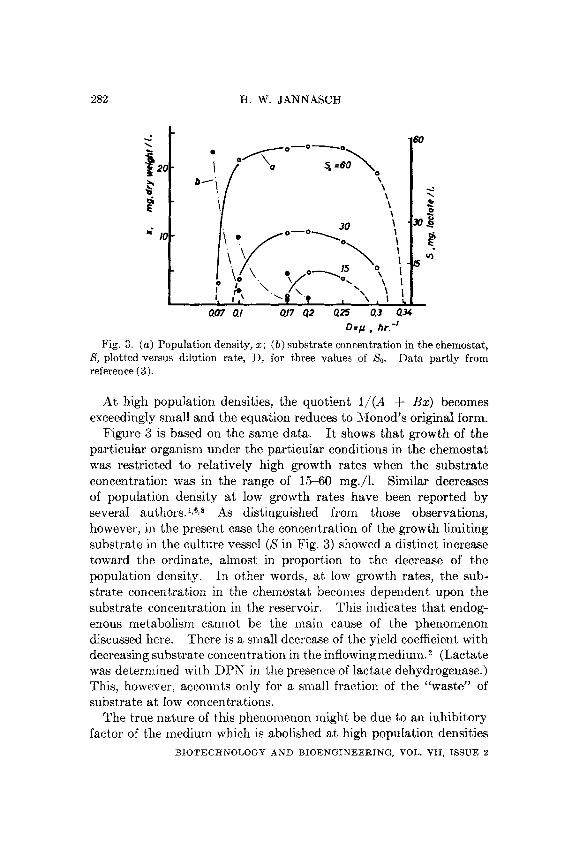
Fig. 3. ( a ) Population density, 2; ( b ) substrate concentration in the chemostat, I h t a partly from S, plotted versus dilution rate, I), for three values of So.
reference (3).
At high population densities, the quotient l / (A + Bz) becomes exceedingly small and the equation reduces to Xlonod’s original form.
Figure 3 is based on the same data. It shows that growth of the particular organism under the particular conditions in the chemostat was restricted to relatively high growth rates when the substrate concentration was in the range of 15-60 mg./l. Similar decreases of population density a t low growth rates have been reported by several authors. lr6F8 As distinguished from those observations, however, in the present case the concentration of the growth limiting substrate in the culture vessel (X in Fig. 3) showed a distinct increase toward the ordinate, almost in proportion to the decrease of the population density. In other words, a t low growth rates, the sub- strate concentration in the cheniostat becomes dependent upon the substrate concentration in the reservoir. This indicates that endog- enous metabolism cannot be the main cause of the phenomenon discussed here. There is a small decrease of the yield coefficient with decreasing substrate concentration in the inflowingmedium.2 (Lactate was determined with DPS in the presence of lactate dehydrogenase.) This, however, accounts only for a small fraction of the “waste” of substrate at low concentrations.
The true nature of this phenomenon might be due to an inhibitory factor of the medium which is abolished a t high population densities
BIOTECHNOLOGY AND BIOENGINEERING. VOL. VII, ISSUE 2

STARTER POPULATIONS 283
by metabolic activity of the organisms. This internal factor, or positive feedback, becomes perceptible only when the concentration of the growth limiting nutrient-and, as a consequence, the popula- tion density-drops below a certain value at which the production of a proposed metabolite does not meet the demand for maintenance of the set growth conditions. The culture, in an indirect sense, becomes “product limited.” This explanation is supported by the fact that the elimination of the inhibitory effect increases with increasing metabolic activity.
After Lodge and Hinshelwood5 had first mentioned stimulation of growth by excreted metabolites, several types of “activators” have been de~cribed.~,~,g With the present cultures, it was observed that a redox potential of -230 mV. was produced and maintained. When the population decreased to the critical level, the medium became less reducing. Restoring the reducing conditions by addition of 0.002% ascorbic acid eliminated part of the inhibit.ory effect (Fig. 1, c). The strains used clearly exhibited microaerophilic behavior. Thus, suboptimal growth conditions were represented by air satura- tion of the medium. At high population densities, the culture was not affected by external changes of the concentrat.ion of dissolved oxygen or the redox potential in a similar range.
In general, at suboptimal conditions, growth of a bacterial culture at low nutrient concentrations is likely to be affected by “threshold” population densities. The possibility of assessing these minimal populations in continuous culture under steady state conditions offers a means of determining starter populations of the corresponding batch cultures.
References
1. Herbert, D., Intern. Congr. Microbwl., 7ch, Stockholm, 381 (1958). 2. Jannasch, H. w., Nature, 196, 496 (1962); 197, 1322 (1963). 3. Jannasch, H. W., Arch. Mikrobiol., 45, 323 (1963). 4. Koburger, J. A., and I‘. J. Claydon, J . Dairy Sci., 44, 1 (1961). 5. Lodge, R. &I., and C. N. Hinshelwood, J . Chem. Soc., 1943,213. 6. Marr, A. G., E. H. Nilson, and D. J. Clark, Ann. N . Y . Acud. Sci., 102,
7. Meyrath, J., Experieniiu, 18, 41 (1962). 8. Schulze, K. L., and R. S. Lipe, Arch. Mikrobiol., 48, 1 (1964). 9. Walker, J. R., J. B. R.eeved, and C. D. Lankford, Racteriol. Proc., 1962, 37.
536 (1963).
Received December 16, 1964





























