Embed Size (px)
Citation preview

Journal of Archaeological Science (1999) 26, 651–659Article No. jasc.1998.0382, available online at http://www.idealibrary.com on
Stable Isotope Ecology and Palaeodiet in the Lake BaikalRegion of Siberia
M. Anne Katzenberg*
Department of Archaeology, University of Calgary, 2500 University Dr N. W., Calgary, Alberta T2N IN4,Canada
Andrzej Weber
Department of Anthropology, University of Alberta, Edmonton, Alberta T6G 2H4, Canada
Human and non-human faunal bone from the Neolithic and Early Bronze Age periods of Cis-Baikal were analysed forstable isotopes of carbon and nitrogen in order to study regional and temporal variation in diet and subsistence. Bothprehistoric and modern faunal species were analysed with all fish from the modern context. Results indicate thatregional variation is greater than temporal variation and can be attributed to proximity to Lake Baikal and its richaquatic resources. Stable isotope analyses of fish and seals from Lake Baikal indicate a wide range of variation inisotope values. ä15N values for the freshwater seals of Lake Baikal average 14&1·1‰. Freshwater fish are highlyvariable in the ä13C values ranging from "24·6‰ for the pelagic omul (Coregonus autumnalis migratorius) to "12·9‰for littoral species such as ide (Leuciscus idus). Terrestrial herbivores have much lower ä15N values, ranging from 4 to5‰ for deer and elk. These data demonstrate that even in temperate, inland regions, there is sufficient variation in stableisotope values of various food resources to provide useful information about variation in human palaeodiet.
? 1999 Academic Press
Keywords: LAKE BAIKAL, CARBON STABLE ISOTOPES, NITROGEN STABLE ISOTOPES,FRESHWATER ECOLOGY, NEOLITHIC, EARLY BRONZE AGE.
* For correspondence.
Introduction
R ecent archaeological research in the Cis-Baikalregion (Lake Baikal and surrounding area) ofSiberia, focusing on the Neolithic period, has
been carried out in order to address questions ofhuman subsistence and health. These issues havebecome particularly important since the identificationof a major culture change separating Early Neolithic(EN) adaptations from those of the Middle Neolithic(MN) and Early Bronze Age (EBA) cultures (Weber,1995). The Neolithic period in Siberia is marked not byagriculture, but by the appearance of pottery, the bowand arrow and stone polishing techniques (Khlobystin,1978). In this paper we present results of stable isotopeanalysis carried out on human burials from a numberof Neolithic and Early Bronze Age sites in the LakeBaikal, Angara and Lena River areas. Results of stableisotope analyses of numerous animal species used asfood in Neolithic times are also presented so thathuman diets can be reconstructed. What emerges is anew understanding of the range of variation in carbonand nitrogen stable isotopes in freshwater ecosystems.This variation, previously unrecognized in palaeodietstudies, is very important in explaining stable
6510305–4403/99/060651+09 $30.00/0
isotope values derived from human tissues and hencepalaeodietary interpretations.
Archaeological evidence indicates that people inthe Cis-Baikal subsisted on terrestrial herbivores, suchas red deer (Cervus elaphus), roe deer (Capreoluscapreolus) and elk (Alces alces) as well as numerous fishspecies from Lake Baikal and surrounding rivers.There is considerable archaeological evidence for theimportance of fishing in the form of various fish hooks,net sinkers and fish effigy figures (Okladnikov, 1950,1955). Some groups also hunted Lake Baikal seals(Phoca sibirica) (Okladnikov & Konopatskii, 1974–75;Weber, Konopatstii & Goriunova, 1993; Weber et al.,1998). Lake Baikal is one of the few places in the worldto have freshwater seals. Vegetation in the region usesthe C3 photosynthetic pathway so little variation incarbon isotopes is expected from terrestrial foods.Previous research on stable isotopes of nitrogen andcarbon from prehistoric freshwater fish bones in theGreat Lakes of North America indicates variationin nitrogen isotope ratios due to trophic level andrelatively less trophic level variation in carbon isotopes(Katzenberg, 1989). Recent ecological studies of otherfreshwater systems, however, suggest much greatercarbon isotope variation than has been acknowl-edged in palaeodiet studies (Kiyashko, Mamontov &
? 1999 Academic Press

652 M. A. Katzenberg and A. Weber
Chernyaev, 1991; Zohary et al., 1994; France, 1995a;Hecky & Hesslein, 1995).
Lam (1994) carried out a pilot study of stableisotope variation between Neolithic and Early BronzeAge peoples of the Cis-Baikal. He found that whilenitrogen isotopes were useful for distinguishingterrestrial animal foods from some aquatic animalfoods the observed carbon isotope variation in humanremains could not be explained by these data. Thepresent study is larger in scale in terms of sample sizes,species, time span and methods of analysis. These newdata support the findings of Lam but they also providean explanation for the carbon isotope variation, whichis due to variation in carbon isotope ratios of fish.
Lake Baikal is the deepest lake in the world at1634 m (Peeters et al., 1997), although estimates varyup to 1740 m. It contains 1/5 of the world’s freshwater,and lies between 51) and 56) N latitude and 103) and110)E longitude. The lake is 636 km long and up to80 km wide and occupies a part of the great Baikal Rift(Kozhov, 1963). The fauna of Lake Baikal are unusualand diverse. There are over 700 species including 52fish species and one species of freshwater seal. Manyspecies are unique to this lake while other organismsare characteristic of the region.
The climate of the Cis-Baikal can be characterized asincluding long cold winters with minimum tempera-tures of "37 to "40)C and short summers withmaximum temperatures around 38)C, but more fre-quently 19–20)C. The lake provides a moderating effecton temperatures and areas away from the lake, such asthe Upper Lena drainage, are more extreme. The lakeis in a mountainous region so that weather conditionscan be highly variable within a day and from one dayto the next. Precipitation is 300–400 mm/year withmost falling in the summer months (Kozhov, 1963).
Water comes into the lake from more than 300rivers, mainly from the north and east. Larger sourcesare the Selenga, Upper Angara, Barguzin and Turka.The only outflow of Lake Baikal is the Angara River,which flows from the southwestern part of the lake.The mountains to the west send short streams into thelake but larger rivers, such as the Lena, have theirsource in the steep slopes of the Baikal range and flownorth to the Arctic Ocean, not connecting with LakeBaikal.
A large-scale archaeological investigation of theCis-Baikal that focuses on the Neolithic period iscurrently being carried out under the direction ofAndrzej Weber, with the involvement of a numberof specialists. The project began with an evaluation ofexisting archaeological interpretations of chronologyand a revision of these based on a new series ofradiocarbon dates from human remains. The firstdetailed chronology for the region was developed inthe 1950s (Okladnikov, 1950, 1955). Weber’s revisedculture history model, based on a large series ofradiocarbon dates, places the Early Neolithic Kitoiculture between 5800 and 4900 , the Middle
Neolithic Servo culture between 4200 and 3000 , andthe Early Bronze Age Glazkovo culture between 3400and 1700 . Of particular interest is an apparentdecrease in population size and density around 4900 (Weber, 1995) and a discontinuity between Kitoiand later groups in terms of both culture and biology.This new chronology is quite different from that ofOkladnikov, which had been challenged by otherspreviously (reviewed in Weber, 1995), and evenreverses the order of some cultures.
There have not been many palaeodiet studies con-ducted in temperate regions where there is no evidencefor the use of either marine foods or C4 plants. VanKlinken, Richards & Hedges (in press) have carriedout such studies on European peoples and have foundthat nitrogen isotope variation is sufficient to makestable isotope studies feasible and useful in temperateregions with predominantly C3 vegetation. Other OldWorld studies in which coastal and C4 resources arenot a factor include those by Bocherens et al. (1991a)on Mediaeval peoples, and Bocherens et al. (1991b)on Neanderthals. Human groups utilizing freshwaterresources have not been studied extensively.Katzenberg (1989, unpubl. work) gathered data fromGreat Lakes fish in an attempt to better understandvariation observed in the prehistoric Ontario peoples.Results suggest that there is sufficient variation innitrogen isotope ratios in freshwater ecosystemsbetween primary and secondary consumers to have animpact on human nitrogen isotope ratios.
The plants and animals of Lake Baikal have beenstudied for several hundred years with a particularemphasis during the 1920s to 1950s. Much of this workis synthesized and presented in English by Kozhov(1963). In recent years there has been a great deal ofinterest in the ecology of Lake Baikal due to the urgentproblem of industrial pollution in the region. Since thelate 1980s there have been several international teamsinvestigating such aspects as species diversity, sealmigration, physical limnology and heavy metal accu-mulation (e.g., Weiss, Carmack & Koropalov, 1991;Gurulev, 1994; Peeters et al., 1997). Of particularinterest to the present study is work carried out byKiyashko et al. (1991) on food web analysis of LakeBaikal fish using stable carbon isotopes.
MaterialsSites for which there are human skeletal samples areshown in Figure 1 and listed with approximate dates inTable 1. Sample sizes vary among the different sitesand our largest samples are from Lokomotiv, a site onthe Angara River near Lake Baikal, and Ust’-Ida,located further north on the Angara River. Faunalbone comes from both archaeological and moderncontexts. All fish bones are modern.
The human skeletal samples are from curated collec-tions in museums in Siberia. Excavated as mortuary

Stable Isotope Ecology in the Lake Baikal Region 653
sites, these collections do not include associatedfaunal samples. Prehistoric mammalian bonesamples are from a number of other sites in the regiondating within the Neolithic. It was not possible tosample archaeological fish bones. Excavations beingcarried out currently will seek to include faunalremains in association with mortuary sites wheneverpossible.
Methods of Analysis
Belaia
Kitoi
Irkut
Obkhoi
Ust'-Ida
Lokomotiv
Ust'-BelaiaKhotoruk
Borki
Khuzhir
Nikol'skaia SkalaMakrushina
Selenga
Lake
B
aika
l
Irkutsk
Kachug
BratskL
ena
Turuka
Ang
ara
Ilim
Ust'-Ilimsk
Ust'-KutKirensk
Kirenga
Angara
0 250 km
Lena
Figure 1. Map of the region showing the location of sites. Lake Baikal lies between 51) and 56) N latitude and 103) and 110) E longitude.
Collagen extraction and analysis by mass spectrometry
Bone protein was isolated following the methoddescribed by Sealy (1986) in which chunks of bone aresoaked in a weak hydrochloric acid until all mineral isdissolved. Collagen ‘‘ghosts’’ are then soaked in weakNaOH to remove any humic contaminants. The

654 M. A. Katzenberg and A. Weber
remaining material is then combusted in a Carlo Erbagas analyser and CO2 and N2 gases are analysed bymass spectrometry. Most materials were analysed on aMicromass Prism mass spectrometer but some wereanalysed on a more recently acquired Finnegan MatTracer Mat mass spectrometer. Fish bones wereanalysed on the Tracer Mat. C:N ratios were deter-mined from sample combustion in a Carlo Erba gasanalyser. All modern samples were also soaked indiethyl ether for 4–6 h prior to treatment with HCl andNaOH in order to remove lipids. Precision is &0·1‰for ä13C and &0·2‰ for ä15N on the Prism and&0·3‰ for ä13C and &0·4‰ for ä15N on the TracerMat.
Biological apatite preparation and analysisBiological apatite was isolated following the proceduredescribed by Lee-Thorp (1989) in which ground bone ispretreated with sodium hypochlorite to remove anyorganic matter, and acetic acid to remove adsorbedand diagenetic carbonates. The sample is then reactedwith phosphoric acid in order to convert the carbonatefrom apatite to CO2. The resulting CO2 was analysedin a Sira mass spectrometer.
Predicted Variation in Stable Isotope RatiosThere are no known terrestrial C4 plants in the region.There was no access to marine foods with the possibleexception of anadromous fish swimming up the LenaRiver from the Arctic Ocean, but there is little evidenceof fish exploitation among the inhabitants of theNeolithic Upper Lena sites. Therefore we expectedto see little variation in carbon isotope ratios whichwere expected to be around "20‰ for humans. Theunknown factor was carbon isotope variation inthe lake fauna. It has generally been assumed thatorganisms living in freshwater lakes had similar carbonisotope ratios to organisms living in terrestrial, C3plant-based environments.
Over the last 10 years, ecologists have becomeinterested in using stable isotopes to characterize
aquatic food webs and to trace carbon sources (Fry &Sherr, 1989; Boutton, 1991; Zohary et al., 1994;France, 1995a; Hecky & Hesslein, 1995). We nowknow that aquatic systems are considerably more com-plex than terrestrial systems and this is discussedbelow.
Earlier work has shown nitrogen isotope vari-ation among freshwater fish species in prehistoric(Katzenberg, 1989) and modern fish (Hesslein et al.,1991; France, 1995b; Vander Zanden & Rasmussen,1996; Vander Zanden, Cabana & Rasmussen, 1997)and this was expected to be the major source ofvariation in Lake Baikal given the presence of fresh-water seals as well as numerous fish species. Nitrogenisotope ratios in humans were expected to vary basedon reliance on terrestrial mammals, plants and aquaticresources including fish and seals.
Table 1. Sites sampled for human bone, listed by time period andregion. Site locations are shown on Figure 1
Region Site
Early Neolithic sites (5800–4900 )Lake Baikal KhotorukAngara River Lokomotiv, Ust’-BelaiUpper Lena River Makrushina, TurukaMiddle Neolithic sites (4200–3000 )Angara River Ust’-IdaUpper Lena River Nikol’skaia SkalaEarly Bronze Age sites (3400–1700 )Lake Baikal KhuzhirAngara River Ust’-IdaUpper Lena River Obkhoi, Borki
Ecology of freshwater systemsAn excellent review of the stable isotope ecology offreshwater systems is provided by Hecky & Hesslein(1995) and this is the source of some of the informationpresented in this section. Terrestrial plants take incarbon from CO2 (ä13C= "7‰) in the atmosphere. C3plants fractionate CO2 to a greater degree than do C4plants due to the greater need in C4 plants to conservewater. The distributions of ä13C values of C3 and C4plants are discrete (Deines, 1980).
Unlike terrestrial plants, aquatic plants have anumber of sources of carbon. These include atmos-pheric CO2, CO2 in the water, bicarbonate and carbon-ate from rocks and soils as dissolved inorganic carbon,and organic carbon from plants and animals that havedecayed in the water and faecal pellets from variousaquatic fauna (Zohary et al., 1994). The amount ofcarbon in water in these different forms depends, inpart on the pH of the water and on the temperature,and may vary seasonally. With each transition (e.g.,from atmospheric CO2 to dissolved inorganic carbon),where carbon crosses a boundary between air andwater, there is fractionation. Carbonate and bicarbo-nate are enriched in 13C by 6–8‰ relative to theatmosphere (around 0‰ versus "7‰). Plants take upCO2 from water and most use the C3 pathway(Boutton, 1991). If there was full atmospheric equi-librium, plant tissues could be "37‰. However,there are a number of factors that interfere with fullequilibrium including photosynthesis, respiration, theboundary layers at interfaces of air, water and solidsand the thermal discontinuities in the water column(Hecky & Hesslein, 1995).
Fractionation of available carbon can vary due toa number of reasons (Deines, 1980: 339). ä13C ofplankton and temperature of water are related.Plankton preferentially utilizes CO2 rather than HCO3unless the former is not readily available. CO2 fraction-ation when taken up by plankton is 18–19‰ such thatplankton is depleted in the heavy isotope. It is not the
Stable Isotope Ecology in the Lake Baikal Region 655
temperature which determines fractionation but therate of CO2 supply. CO2 supply increases with decreas-ing water temperature. When available CO2 decreases,fractionation decreases (down to as low as 6‰)(Deines, 1980: 339).
Fish are a few steps up the trophic web fromphytoplankton and macrophytes and vary in terms oftheir diets and habitats. Variation in fish isotope valuesthen must relate to these differences in the diet andhabitat as well as variation within species depending onwhat part of the lake they are taken from and at whattime of year. There will also be trophic level differencesamong fish species and diets of individual fish may varyover their life span. Within Lake Baikal there areseveral levels to the food web with seals on top. Seals(Phoca sibirica) feed on fish such as sculpins (genusCottocomephorus) and bullhead (genus Comephorus),who feed on zooplankton, who feed on phytoplankton(Kiyashko et al., 1991; Kozhov, 1963).
To summarize, for aquatic plants in general, Hecky& Hesslein (1995: 634) describe the situation asfollows:
‘‘In contrast to the relatively simple situation for mostterrestrial plants which have a relatively narrow range ofä13C signatures dependent primarily on their photosyn-thetic fractionation, i.e., C3 versus C4, aquatic plantsproduce a broad and potentially continuous range ofsignatures because of the interplay of physical as well asbiological factors’’. (Hecky & Hesslein, 1995: 634.)
In a study of carbon isotopes and trophic level inLake Malawi, Hecky & Hesslein (1995) found vari-ation in fish similar to our findings in Lake Baikal.Both lakes are very deep and occupy rifts. In LakeMalawi, littoral fish species are isotopically heavier(ä13C ranges from "6 to "21‰) than are the pelagicspecies (ä13C ranges from "26 to "22·5‰), much aswe and others have found in Lake Baikal (Kiyashko etal., 1991). France (1995a) found differences in the ä13C
values of littoral and pelagic consumers in Canadianshield lakes. Stable isotope ratios of fish from temper-ate and Arctic lakes in Canada are variable but are lessenriched in 13CO2 than are fish from Lakes Baikaland Malawi. For example, in temperate lakes on theCanadian shield (NW Ontario) fish ä13C ranges from"32 to "21‰ (Hecky & Hesslein, 1995).
Results and Discussion
Table 2. Results of stable isotope analyses (ä13C and ä15N) by region and time period. Faunal samples are from various sites and time periodswithin the Neolithic and Early Bronze Age Cis-Baikal
Carbon from collagenä13C(‰)
Nitrogen from collagenä15N(‰)
Carbon from bioapatiteä13C(‰)
Collagen-apatitespacingSpecies Region Period n Mean .. n Mean .. n Mean ..
Humans Angara EN 20 "15·8 1·1 21 14·4 1·0 4 "10·8 1·0 5·0Humans Angara MN 4 "19·2 0·3 4 11·5 0·2 6 "11·8 1·1 7·3Humans Angara EBA 5 "19·9 1·0 5 10·2 1·2 2 "12·1 7·7Humans Baikal EN 3 "17·6 0·9 2 12·6 3 "12·3 1·7 5·3Humans Baikal EBA 6 "18·4 0·4 6 14·3 0·5 2 "13·8 4·5Humans U. Lena EN 11 "20·2 0·6 11 12·0 0·8 12 "12·3 0·9 7·9Humans U. Lena MN 3 "19·5 0·4 3 10·9 0·7 3 "12·6 0·9 6·9Humans U. Lena EBA 13 "19·5 0·4 13 10·1 0·8 14 "13·9 0·6 5·6Alces Prehistoric 3 "19·3 1·2 3 4·1 1·4 3 "10·1 0·7 9·2Cervus Prehistoric 8 "19·0 0·6 7 5·0 1·1 7 "10·3 0·8 8·7Phoca Prehistoric 8 "22·0 0·9 8 14·0 1·1 8 "15·6 1·5 6·4Capreolus Prehistoric 8 "20·1 0·5 8 5·4 1·0 4 "12·3 1·6 7·9
Carbon isotopes—collagen
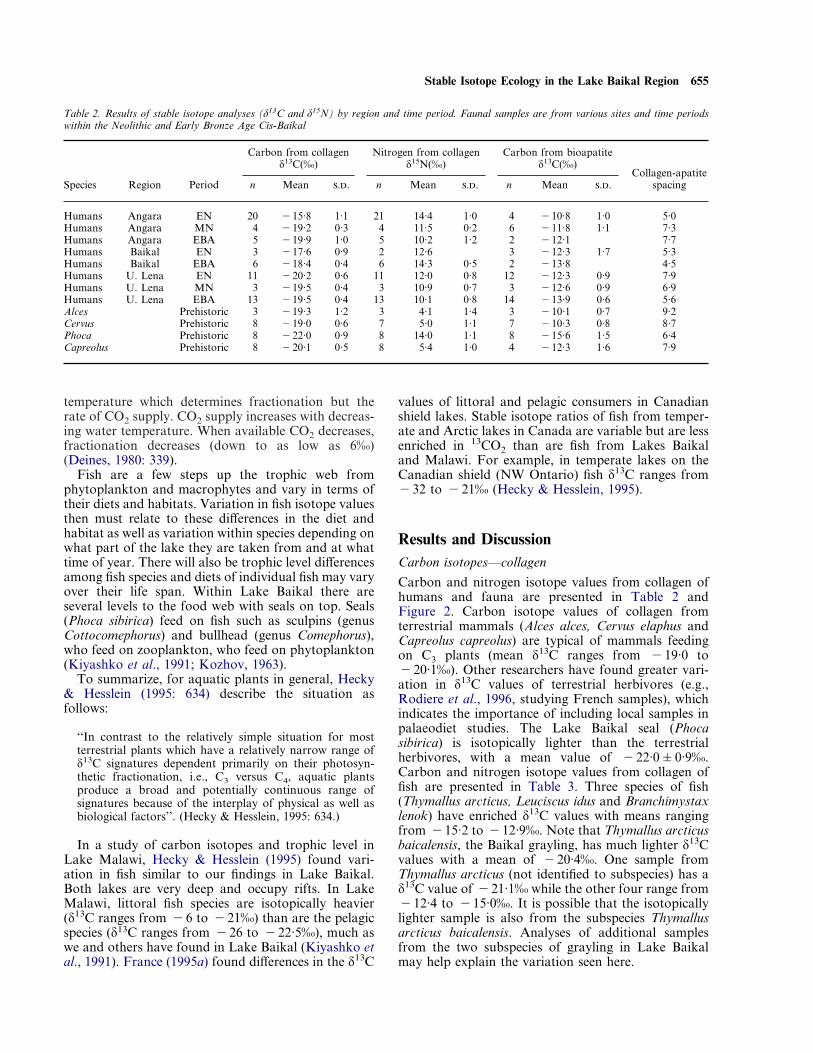
Carbon and nitrogen isotope values from collagen ofhumans and fauna are presented in Table 2 andFigure 2. Carbon isotope values of collagen fromterrestrial mammals (Alces alces, Cervus elaphus andCapreolus capreolus) are typical of mammals feedingon C3 plants (mean ä13C ranges from "19·0 to"20·1‰). Other researchers have found greater vari-ation in ä13C values of terrestrial herbivores (e.g.,Rodiere et al., 1996, studying French samples), whichindicates the importance of including local samples inpalaeodiet studies. The Lake Baikal seal (Phocasibirica) is isotopically lighter than the terrestrialherbivores, with a mean value of "22·0&0·9‰.Carbon and nitrogen isotope values from collagen offish are presented in Table 3. Three species of fish(Thymallus arcticus, Leuciscus idus and Branchimystaxlenok) have enriched ä13C values with means rangingfrom "15·2 to "12·9‰. Note that Thymallus arcticusbaicalensis, the Baikal grayling, has much lighter ä13Cvalues with a mean of "20·4‰. One sample fromThymallus arcticus (not identified to subspecies) has aä13C value of "21·1‰ while the other four range from"12·4 to "15·0‰. It is possible that the isotopicallylighter sample is also from the subspecies Thymallusarcticus baicalensis. Analyses of additional samplesfrom the two subspecies of grayling in Lake Baikalmay help explain the variation seen here.
656 M. A. Katzenberg and A. Weber
–12
16
2–26
δ13C (‰)
δ15N
(‰
)
–16
6
14
12
10
8
4
–24 –22 –20 –18 –14
Cor.a.m.
Caras au
Capr. cap.
Angara LLena L
Lena MAngara M
Esox luc.Baikal E
Baikal LPhoca sib.
Thymus a.b.Perca flu.
Cervus el.Alces alc.
Lena E
Angara E
Bran. len
Thym. arcLeuc. idu
Figure 2. Carbon and nitrogen stable isotope ratios for protein from human and non-human faunal bone. Human samples are plotted byregion and time period (from data in Table 2). The three regions are Baikal, Angara and Lena following the groups presented in Table 1. Timeperiods are Early Neolithic (E), Middle Neolithic (M) and Early Bronze Age (L).
Table 3. Carbon (ä13C) and nitrogen (ä15N) stable isotope results for modern fish from Lake Baikal. Habitat and feeding habits are those that aremost predominant (Kozhov, 1963; Scott & Crossman, 1973; Kiyashko et al., 1991)
Latin name Common name n ä13C(‰) .. ä15N(‰) .. Habitat and feeding habit
Thymallus arcticus Grayling 5 "15·2 3·4 10·1 0·6 Littoral and pelagic omnivoreLeuciscus idus Ide 2 "12·9 9·4 Littoral detritivore and planktivoreBranchimystax lenok Lenok 4 "14·2 0·8 11·1 0·7 Littoral omnivoreEsox lucius Pike 2 "22·0 12·0 Littoral and pelagic picivoreThymallus arcticus baicalensis Baikal grayling 1 "20·4 13·7 Pelagic omnivoreCarassius auratus Carp 1 "21·4 7·3 Benthic detritivorePerca fluviatalis Perch 2 "21·0 11·9 Pealgic and littoral omnivoresCoregonus autumnalis migratorius Omul 4 "24·6 0·4 11·5 0·7 Pelagic planktivore
Kiyashko et al. (1991) also found variation in ä13Cvalues that separates pelagic species, such as omul(Coregomus), from benthic species such as bullhead inLake Baikal. In their study, the omul are isotopicallylightest, and lenok (Branchimystax lenok), whitegrayling (Thymallus arcticus brevipinnis) and dace(Leuciscus leuciscus) are isotopically more enriched,as in the present study (if we assume that the en-riched Thymallus arcticus are white grayling). Stablecarbon isotope variation in pelagic versus littoral andbenthic species has been attributed to the difference indiscrimination of 13C at the level of primary producersin both marine and freshwater systems (France,1995a).
The range of mean ä13C values for humans is from"15·8‰ at the Early Neolithic Lokomotiv site,which is located on the Angara River near its source inLake Baikal, to "20·2‰ in the Upper Lena RiverEarly Neolithic sites of Makrushina and Turuka. Asexpected, the Upper Lena River peoples were relyingon terrestrial herbivores as their main source of pro-tein. The enriched ä13C values seen in the people livingclose to Lake Baikal must result from consumption offish species with enriched ä13C values. These fish are allinhabitants of the littoral zone (shallow waters) ofLake Baikal. The pelagic (open water) and benthic(deep-water) species have lighter carbon isotope values("20·4 to "24·6‰).

Stable Isotope Ecology in the Lake Baikal Region 657
Nitrogen isotopes—collagenNitrogen isotope ratios are lowest in the terrestrialherbivores (mean ä15N ranges from 4·1 to 5·4‰) andhighest in the Lake Baikal seals (ä15N=14·0&1·0‰).Fish show a wide range of variation, which may belargely attributed to differences in trophic level. Thedetritivore, carp, is lowest at 7·3‰ while the lenok,which is at the top of the littoral trophic chain, are11·1&0·7‰ and the Baikal grayling, which is highon the trophic chain of pelagic fish, has a ä15N valueof 13·7‰.
Nitrogen isotope values for humans are highest forthe three groups living in closest proximity to LakeBaikal. The Early Neolithic Angara River samplecomes from the Lokomotiv site, near the source of theAngara River at Lake Baikal. The Early NeolithicLake Baikal sample is from Khotoruk, a site on thewestern shore of the lake. The Early Bronze Age LakeBaikal sample is from Khuzhir, located on Ol’khonIsland in Lake Baikal. The region of the lake betweenthe western shore and Ol’khon Island is called theMaloye More (minor, or little sea) (Kozhov, 1963). Itis one of the shallower parts of the lake and is rich inaquatic resources.
The lightest nitrogen isotope values among thehuman groups are found in the Upper Lena River sitesand the Early Bronze Age Angara River sample. Thesesites are located the farthest from Lake Baikal. Faunalresources must have been largely terrestrial herbivoresand possibly some fish. Based on the carbon isotopevalues from these sites, these would not include any ofthe fish species with high ä13C values.
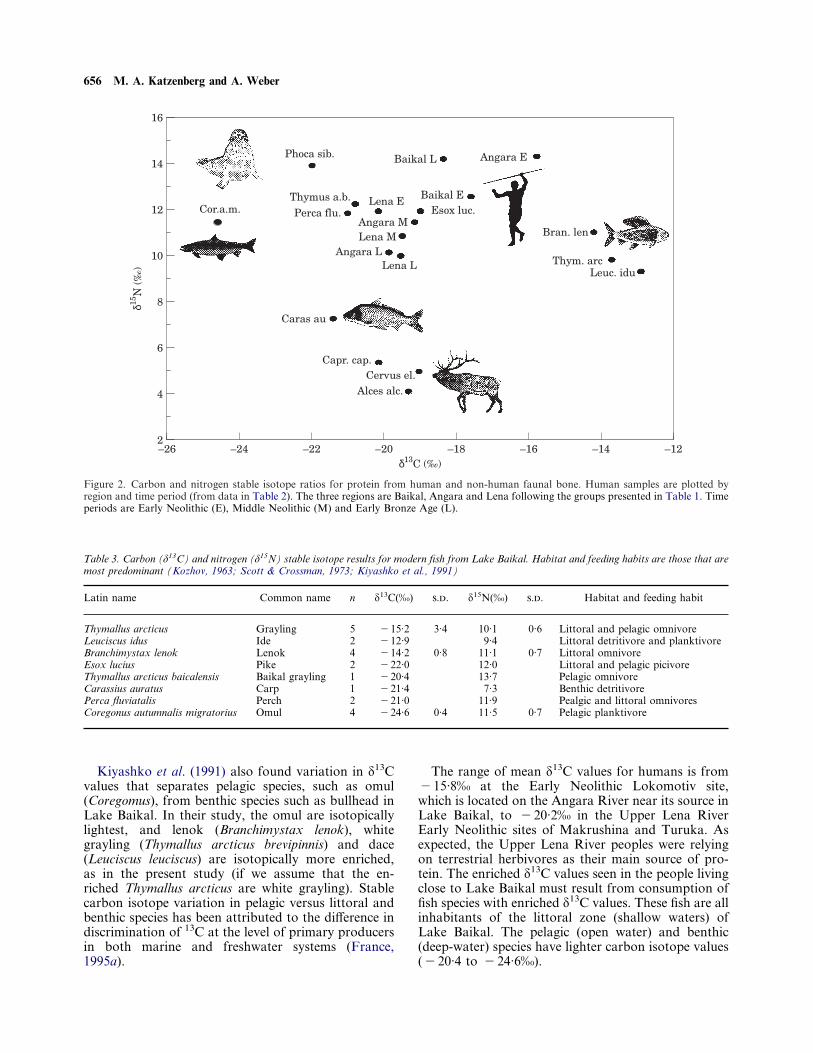
Carbon isotopes—biological apatiteThe ä13C values from biological apatite provideanother source of information about reliance on differ-ent food resources. The correlation coefficient betweenmean values (from Table 2) for ä13C of collagen andbiological apatite is 0·52. This suggests that the twocarbon sources are providing different information, assuggested by Krueger & Sullivan (1984) and demon-strated in controlled feeding experiments carried out byAmbrose & Norr (1993) and Tieszen & Fagre (1993).Specifically, the carbon in collagen comes largely fromprotein in the diet, while the carbon in biologicalapatite reflects the total diet. The difference in the ä13Cvalues of these two carbon sources is related to trophiclevel with herbivores showing a greater difference andcarnivores a smaller difference (Krueger & Sullivan,1984; Lee-Thorp, Sealy & van der Merwe, 1989).Therefore, the spacing between collagen and biologicalapatite correlates roughly with the ä15N values sinceboth are related to reliance on animal protein(Figure 3). This is a negative correlation (r= "0·8)since ä15N increases as the spacing between collagenand apatite ä13C decreases. The smallest differencebetween these two carbon sources is in the EarlyBronze Age Lake Baikal site, followed by the Early
Neolithic Lokomotiv site on the Angara, near LakeBaikal. The Early Neolithic Lake Baikal site is next,followed by the seals. The largest spacing is found inAlces alces, followed by Cervus elaphus and Capreoluscapreolus. The highest ä15N values are found amongthe groups with the smallest difference between ä13C ofcollagen and biological apatite, and the lowest ä15Nvalues are found among the groups with the largestdifference between ä13C of collagen and biologicalapatite. This trend does not occur for the Upper Lenasites. This is probably because their source of animalprotein is the large terrestrial herbivores (deer and elk)that have low ä15N values in comparison with animalprotein from lake and riverine sources. Because of thediversity of protein sources within this study regionä15N values may differ while reflecting similar relianceon animal foods. This is because the large terrestrialfauna have lower ä15N values than do the fish and sealsof Lake Baikal and surrounding rivers.
16
10
44
δ15N (‰)
Col
lage
n–a
pati
te s
paci
ng
(‰)
6
5
9
8
7
6
8 10 12 14
Terrestrialherbivores
Angara
U. Lena
U. Lena
U. Lena
Baikal
Seals
Angara
Baikal
Angara
Figure 3. Plot of ä15N values in comparison with the differencebetween ä13C of collagen and ä13C of biological apatite. Data arefrom Table 2.
ConclusionsThe three lines of stable isotope evidence forprehistoric diet in the Cis-Baikal suggest both regionaland temporal variation in diet and subsistence. Thosepeople living closest to Lake Baikal differ from peopleliving along the Upper Lena and the Angara River sitesmore distant from the lake. The sites closest to thelake (Early Neolithic Lokomotiv site on the AngaraRiver and the Early Neolithic and Early Bronze Agesites on the shore and on Ol’khon Island) have thehighest ä13C and ä15N values for collagen, and thesmallest difference between ä13C of collagen and bio-logical apatite of all human samples. All indicatorssuggest reliance on the fish and seals of Lake Baikal.People further down the Angara River and along theUpper Lena River appear to have relied mainly onterrestrial herbivores for their protein source, with lessreliance on riverine fish species.
From the perspective of subsistence behaviour, thisargues for reliance on food resources close to home.

658 M. A. Katzenberg and A. Weber
Regularly scheduled migration or hunting trips to LakeBaikal for exploitation of its food resources does notappear to have been practiced by peoples of theAngara or Upper Lena valleys. Presumably they hadadequate food resources in closer proximity to theirhabitation sites. While there is no archaeological evi-dence for fishing among the Upper Lena sites thenitrogen isotope evidence indicates some input of fishinto the diet since ä15N values of humans are 2–3‰higher than expected if protein were mainly fromterrestrial herbivores.
Reliance on terrestrial mammals and locally avail-able plants would result in human isotope valuesaround 7·5‰ for ä15N and around "20‰ for ä13C incollagen. We do not see these values in any of theburials analysed. The low values for ä15N (10–11‰)occur in the riverine sites far from Lake Baikal. Thecarbon values are consistent with predictions but theelevated ä15N values indicate at least a minimal intakeof fish. The higher ä15N values from the sites in andaround Lake Baikal are consistent with much greaterreliance on fish and exploitation of seals. This isconsistent with the faunal evidence from those samesites (see Weber et al. (1998), on prehistoric sealhunting at these sites). The higher ä13C values of thesesites indicate some reliance on the isotopically enrichedfish such as white grayling (Thymallus arcticus brev-ipinnis), ide (Leuciscus idus) and lenok (Branchimystaxlenok). These higher ä13C values were not expected andit was only through sampling a range of fish speciesfrom the lake that their source was determined. Thefact that the people of the Early Neolithic Lokomotivsite on the Angara River are isotopically similar to thepeople of the Lake Baikal sites must relate to the factthat the upper reaches of the Angara, where it leavesLake Baikal, are very cold and have a very similarecology to that of the lake. Foods consumed by thepeople of the Lokomotiv site would then be expectedto have similar isotope ratios to those consumed bypeople living along the shores of the lake.
Temporal variation among the Upper Lena Riversites can be seen in slightly higher ä15N values in theEarly Neolithic sites in comparison with the MiddleNeolithic and Early Bronze Age sites. This suggestsgreater reliance on fish in the Early Neolithic. The veryhigh ä15N values in the Early Bronze Age samples fromthe Khuzhir site on Ol’khon Island reflect consumptionof seal, which is supported by faunal evidence for sealhunting (Weber et al., 1998). The small amount oftemporal variation among the Upper Lena sites is dueto the fact that there is little variation in the isotopicsignatures of the food resources in that area.
The larger than expected variation in the stablecarbon isotope values for the fauna in and aroundLake Baikal explains the variation observed in humanremains. Stable isotope variation in terrestrial andfreshwater systems has the potential to provide infor-mation on palaeodiet, even in temperate regions. Itseems, from the available literature and from our
results, that deep lakes will provide more variation incarbon isotope ratios due to the various sources ofcarbon available at the lower levels of the trophic web.For the Neolithic and Early Bronze Age peoples of theLake Baikal region, varying emphasis on aquatic andterrestrial resources may be observed, and may becorrelated with faunal remains and artefacts used inprocuring these resources. Unknown factors in palaeo-diet reconstruction include exploitation of plants. Withfurther sampling it may be possible to determinechanges in subsistence behaviour among the people ofthis region. Our current data suggest some subsistencedifferences in the Early Neolithic in comparison withlater times, but these differences are very subtle.
A knowledge of the stable isotope ecology of fresh-water systems can aid in understanding sources ofvariation in carbon isotope ratios at a number oflevels. The applications to aquatic ecology includeunderstanding trophic levels and food webs, andunderstanding the dynamics of carbon sources in lakesand rivers. For archaeology, this adds another clue tounderstanding past human subsistence behaviour.
AcknowledgementsThis research was funded by grants from the SocialSciences and Humanities Research Council of Canada(nos 410–93–0201 and 410–96–0353) to AndrzejWeber. Additional funding is from the Associationof Universities and Colleges of Canada and theUniversity of Alberta Central Research fund (toAndrzej Weber). Stable isotope work was carried outat the University of Calgary. Sharon Whitley andRoman Harrison of the Department of Archaeology,University of Calgary, prepared bone samples. Stableisotope analyses were performed in the Stable IsotopeLaboratory, Department of Physics, University ofCalgary, under the direction of Dr H. R. Krouse withassistance from Nenita Lozano and Maria Mihailescu.Bone samples were kindly provided by V. I. Bazaliiskiiand N. Berdnikova of Irkutsk State Universityand A. K. Konopatskii of the Russian Academy ofScience, Novosibirsk. Thanks also to AcademiciansA. P. Derevianko of the Russian Academy ofScience, Novosibirsk and G. I. Medvedev of IrkutskState University for their continuous support. A.Katzenberg is very grateful to colleagues, J. R. Postand F. E. R. McCauley, Department of BiologicalSciences, University of Calgary for helpful discussionson aquatic ecology.
ReferencesAmbrose, S. H. & Norr, L. (1993). Experimental evidence for the
relationship of the carbon isotope ratios of whole diet and dietaryprotein to those of bone collagen and carbonate. In (J. B. Lambert& G. Grupe, Eds) Prehistoric Human Bone: Archaeology at theMolecular Level. Berlin: Springer Verlag, pp. 1–37.

Stable Isotope Ecology in the Lake Baikal Region 659
Bocherens, H., Fizet, M., Mariotti, A., Olive, C., Bellon, G. &Billiou, D. (1991a). Application de la biogeochimie isotopique(13C, 15N) a la determination du regime alimentaire despopulations humaines et animales durant les periodes antique etmedievale. Archives des Sciences 44, 329–340.
Bocherens, H., Fizet, M., Mariotti, A., Lange-Badre, B.,Vandermeersch, B., Borel, J. P. & Bellon, G. (1991b). Isotopicbiogeochemistry (13C, 15N) of fossil vertebrate collagen: applica-tion to the study of a past food web including Neandertal man.Journal of Human Evolution 20, 481–492.
Boutton, T. W. (1991). Stable carbon isotope ratios of naturalmaterials: II. Atmospheric, terrestrial, marine and freshwaterenvironments. In (D. C. Coleman & B. Fry, Eds) Carbon IsotopeTechniques. San Diego: Academic Press, Inc., pp. 173–185.
Deines, P. (1980). The isotopic composition of reduced organiccarbon. In (P. Fritz & J. C. Fontes, Eds) Handbook of Environ-mental Isotope Geochemistry. Amsterdam: Elsevier, pp. 329–406.
France, R. L. (1995a). Differentiation between littoral and pelagicfood webs in lakes using stable carbon isotopes. Limnology andOceanography 40, 1310–1313.
France, R. L. (1995b). Stable nitrogen isotopes in fish: literaturesynthesis on the influence of ecotonal coupling. Estuarine, Coastaland Shelf Science 41, 737–742.
Fry, B. & Sherr, E. B. (1989). ä13C measurements as indicators ofcarbon flow in marine and freshwater ecosystems. In (P. W.Rundel, J. R. Ehleringer & K. A. Nagy, Eds) Stable Isotopes inEcological Research. Ecological Studies No. 68. New York:Springer Verlag, pp. 196–229.
Gurulev, S. A. (1994). The Face of Baikal: Water. Irkutsk: Name.Hecky, R. E. & Hesslein, R. H. (1995). Contributions of benthic
algae to lake food webs as revealed by stable isotope analysis.Journal of the North American Benthological Society 14, 631–653.
Hesslein, R. H., Capel, M. J., Fox, D. E. & Hallard, K. A. (1991).Stable isotopes of sulfur, carbon and nitrogen as indicators oftrophic level and fish migration in the Lower Mackenzie RiverBasin, Canada. Canadian Journal of Fisheries and Aquatic Sciences48, 2258–2265.
Katzenberg, M. A. (1989). Stable isotope analysis of archaeologicalfaunal remains from southern Ontario. Journal of ArchaeologicalScience 16, 319–329.
Khlobystin, L. P. (1978). Vozrast i sootnoshenie neoliticheskikhkul’tur Vostochnoi Sibiri (The age and relationship of Neolithiccultures of eastern Siberia). Kratkie Soobshcheniia InstitutaArkheologii 153, 93–99.
Kiyashko, S. I., Mamontov, A. M. & Chernyaev, M. Zh. (1991).Food web analysis of Lake Baikal fish by ratios of stable carbonisotopes. Doklady, Biological Sciences 318, 274–276.
Kozhov, M. (1963). Lake Baikal and its Life. MonographiaeBiologicae XI. The Hague: Dr. W. Junk.
Krueger, H. W. & Sullivan, C. H. (1984). Models for carbon isotopefractionation between diet and bone. In (J. E. Turnlund & P. E.Johnson, Eds) Stable Isotopes in Nutrition. American ChemicalSociety Symposium Series No. 258. Washington, D.C: ACS,pp. 205–222.
Lam, Y. M. (1994). Isotopic evidence for change in dietary patternsduring the Baikal Neolithic. Current Anthropology 35, 185–190.
Lee-Thorp, J. A. (1989). Stable carbon isotopes in deep time: thediets of fossil fauna and hominids. Ph.D. Thesis. University ofCape Town.
Lee-Thorp, J. A., Sealy, J. C. & van der Merwe, M. J. (1989). Stablecarbon isotope ratio differences between bone collagenand bone apatite, and their relationship to diet. Journal ofArchaeological Science 16, 585–599.
Okladnikov, A. P. (1950). Neolit i bronzovyi vek Pribaikal’ia (chast’I i II) (The Neolithic and Bronze age of the Cis-Baikal (part I and
II). Materialy i Issledovaniia po Arkheologii SSSR 18, Vol 43.Moscow Izdatel’stvo Akademii nauk SSSR.
Okladnikov, A. P. (1955). Neolit i bronzovyi vek Pribaikal’ia (chast’III) (The Neolithic and Bronze Age of the Lake Baikal region(part III). Materialy i Issledovaniia po Arkheologii SSSR 18,Vol. 43. Moscow: Izdatel’stvo Akademii nauk SSSR.
Okladnikov, A. P. & Konopatskii, A. K. (1974–1975). Hunters forseal on the Baikal Lake in the Stone and Bronze Ages. Folk 16–17,299–308.
Peeters, F., Kipfer, R., Hohmann, R., Hofer, M., Imboden, D. M.Kodenev, G. G. & Khozder, T. (1997). Modeling transportrates in Lake Baikal: gas exchange and deep water renewal.Environmental Science and Technology 31, 2973–2982.
Rodiere, E., Bocherens, H., Angibault, J. M. & Mariotti, A. (1996).Particularites isotopiques de I’azote chez le chevreuil (Caprioluscapriolus L.): implications pour les reconstitutions paleoenviron-nementales. Comptes Rendus de l’Academie des Sciences 323,179–185.
Scott, W. B. & Crossman, E. J. (1973). Freshwater Fishes of CanadaFisheries Research Board of Canada Bulletin No. 184. Ottawa:Fisheries Research Board of Canada.
Sealy, J. C. (1986). Stable carbon isotopes and prehistoric diets in thesouthwestern Cape Province, South Africa. BAR InternationalSeries No. 293, Oxford: BAR.
Tieszen, L. L. & Fagre, T. (1993). Effect of diet quality andcomposition on the isotopic composition of respiratory CO2 bonecollagen, bioapatite and soft tissues. In (J. B. Lambert & G. Grupe,Eds) Prehistoric Human Bone: Archaeology at the Molecular Level.Berlin: Springer Verlag, pp. 121–155.
Vander Zanden, M. J. & Rasmussen, J. B. (1996). A trophicposition model of pelagic food webs: impact on contaminantbioaccumulation in lake trout. Ecological Monographs 66,451–477.
Vander Zanden, M. J., Cabana, G. & Rasmussen, J. B. (1997).Comparing trophic position of freshwater fish calculated usingstable nitrogen isotope ratios (ä15N) and literature dietarydata. Canadian Journal of Fisheries and Aquatic Sciences 54,1142–1158.
Van Klinken, G. J., Richards, M. P. & Hedges, R. E. M. (in press).An overview of causes for stable isotopic variations in pastEuropean human populations: environmental, ecophysiological,and cultural effects. In (S. H. Ambrose & M. A. Katzenberg, Eds)Close to the Bone: Biogeochemical Approaches to PaleodietaryAnalysis in Archaeology. Advances in Archaeological and MuseumScience. Plenum Press.
Weber, A. (1995). The Neolithic and Early Bronze Age of the LakeBaikal region: a review of recent research. Journal of WorldPrehistory 9, 99–165.
Weber, A., Konopatskii, A. K. & Goriunova, O. I. (1993).Prehistoric seal hunting on Lake Baikal: methodology andpreliminary results of the analysis of canine sections. Journal ofArchaeological Science 20, 629–644.
Weber, A., Link, D. W., Goriunova, O. I. & Konopatskii, A. K.(1998). Patterns of prehistoric procurement of seal at Lake Baikal:a zooarchaeological contribution to the study of past foragingeconomies in Siberia. Journal of Archaeological Science 25,215–227.
Weiss, R. F., Carmack, E. C. & Koropalov, V. M. (1991).Deep-water renewal and biological production in Lake Baikal.Nature 349, 665–669.
Zohary, T., Erez, J., Gophen, M., Berman-Frank, I. & Stiller, M.(1994). Seasonality of stable carbon isotopes within the pelagicfood web of Lake Kinneret. Limnology and Oceanography 39,1030–1043.





























