Embed Size (px)
Citation preview

JOURNAL OF VIROLOGY, Sept. 2003, p. 10015–10027 Vol. 77, No. 180022-538X/03/$08.00�0 DOI: 10.1128/JVI.77.18.10015–10027.2003Copyright © 2003, American Society for Microbiology. All Rights Reserved.
SR-Related Protein TAXREB803/SRL300 Is an Important CellularFactor for the Transactivational Function of Human
T-Cell Lymphotropic Virus Type 1 TaxHwang-Geum Youn,1 Jun Matsumoto,1 Yuetsu Tanaka,2 and Kunitada Shimotohno1*
Laboratory of Human Tumor Viruses, Department of Viral Oncology, Institute for Virus Research,Kyoto University, Kyoto 606-8507,1 and Department of Infectious Diseases and Immunology,
Okinawa-Asia Research Center of Medical Science, Faculty of Medicine,University of the Ryukyus, Okinawa 903-0215,2 Japan
Received 22 January 2003/Accepted 24 June 2003
Expression of the human T-cell lymphotropic virus type 1 (HTLV-1) genes is transcriptionally activated bythe cognate oncoprotein Tax which enhances the binding of the cyclin AMP-responsive element binding protein(CREB) to the Tax responsive element (TxRE) located in its long terminal repeat (LTR). TxRE is highlyhomologous to the cyclic AMP-responsive element (CRE) except for the GC-rich sequence flanking the CRE.We cloned the cDNA for a cellular factor, TAXREB803, of which the DNA-binding domain bound to TxRE andthe binding was dependent on the 3� GC-rich sequence in TxRE. TAXREB803 is an SR-related proteincomposed of 2,752 amino acids including numerous arginine/serine (RS) motifs. TAXREB803 enhanced boththe Tax dependent transcription and the CREB binding to TxRE in cooperation with Tax. The interaction ofTAXREB803 and Tax was detected by coimmunoprecipitation assays as well as by indirect immunofluorescenceassays. Significantly, Tax transactivation for the HTLV-1 LTR decreased dramatically when the expressionlevel of the endogenous TAXREB803 was suppressed by the small interfering RNA. These results suggest thatTAXREB803 functions as a transcriptional coactivator for Tax and plays a critical role in the expression ofHTLV-1 genes.
Human T-cell lymphotropic virus type 1 (HTLV-1) is a hu-man retrovirus which is the causative agent for adult T-cellleukemia/lymphoma. HTLV-1 is also etiologically related to aneurological disorder called tropical spastic paraparesis/HTLV-1-associated myelopathy as well as other diseases (re-viewed in reference 54). The HTLV-1 genome encodes the40-kDa viral transactivator protein Tax, which acts as a potentactivator for gene expression of HTLV-1 (10, 45).
Tax interacts with various cellular proteins, which are tran-scription-related factors and others, and modulates their tran-scriptional activities. The cellular genes, including those forinterleukin-2 (IL-2) and the IL-2 receptor � (29, 33) and c-fos(13, 14, 48), which possess nuclear factor �B (NF-�B) or se-rum-responsive factor (SRF) binding sites in their promoters,are transactivated by the interaction of Tax with NF-�B, I�B,IKK� (22, 50), or SRF (13, 48). Tax also interacts with factorsrelated to cell cycle progression and plays a role in cell growthcontrol. It transsuppresses transcription of genes such as cyclinD3 and CDK inhibitors (2, 21) and enhances the kinase activityof CDK4 resulting from the direct binding of Tax to CDK4 andp16INK-4A (19, 51). In the others, it is suggested that Taxbinding to the human homolog of the Drosophila Disc largetumor suppressor (hDLG) inhibited the tumor-suppressivefunction of hDLG (52) and network forming by the neuron-specific filament protein �-internexin is inhibited by its binding
to Tax (42). Although several cellular proteins have been re-ported to interact with Tax, the physiological effects of theirinteractions remain to be clarified.
The expression of the HTLV-1 genes by Tax depends onthree imperfectly repeated sequences of the 21-bp elementknown as the Tax-responsive element (TxRE) localized in theU3 region of the proviral long terminal repeats (LTRs) (5, 15,41, 47). The transactivational function of Tax with respect tothe TxREs is mediated by interaction with the cellular factorssuch as cyclic AMP-responsive element (CRE) binding proteinCREB/ATF family (49, 58), CREB-binding protein (CBP) (17,25), and p300/CBP-associated factor (PCAF) (23). The TxREconsists of the cellular CRE-like sequence flanked by GC-richsequences at its 3� terminus. The cellular CRE-containinggenes do not carry the GC-rich flanking sequences in theirpromoter regions and are not transactivated by Tax (56, 57).Tax interacts directly with CREB through its N terminus andstabilizes the binding of CREB to the TxRE (6, 18). Thebinding affinity of CREB to CBP is also increased in the pres-ence of Tax. Interestingly, the increase in CREB binding toTxRE or to CBP does not occur in the cellular CRE (26). It issuggested, therefore, that the functional interaction of Taxwith TxRE for transactivation depends on the GC-rich flankingsequences in TxRE. However, the mechanism of the specificinteractions mediated by the GC-rich sequence remains to beclarified.
We have previously isolated several TxRE-binding proteinsby Southwestern blotting, including TAXREB67/CREB-2/ATF-4, TAXREB302, TAXREB107, TAXREB703, andTAXREB803 (38, 39, 53). A piece of cDNA for TAXREB803
* Corresponding author. Mailing address: Laboratory of HumanTumor Viruses, Department of Viral Oncology, Institute for VirusResearch, Kyoto University, Kyoto 606-8507, Japan. Phone: 81 75 7514000. Fax: 81 75 751 3998. E-mail: [email protected].
10015
on February 14, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

was obtained from a peptide that specifically bound to TxRE(53). By searching GenBank for homologous genes, we founda partial cDNA clone with accession number AB002322. Weconstructed the full cDNA for TAXREB803 with the cDNA ofAB002322 and found that TXAREB803 has the capacity toencode a protein with 2,752 amino acid residues. The DNAbinding domain (DBD), the peptide initially isolated by us(53), resides between positions 1616 and 1718 of the aminoacid sequence of this putative protein.
While investigating the functions of TAXREB803, two pro-teins, SRL300 and SRm300, were identified by other groups. Inamino acid sequence, SRL300 is identical to TAXREB803, butSRm300 has a complicated structure, with many regions iden-tical to the TAXREB803. SRL300 was cloned as an RNA-binding protein bound to the 5� noncoding region of the AT-rich element binding factor 1 (ATBF1) mRNA (44). SRm300was isolated as a component of the SRm160/300 complex (SR-related matrix protein complex of 160 kDa and 300 kDa).Although the SRm160/300 complex was found to be capable ofassociating with the splicing complexes, only SRm160 couldactivate the splicing reaction, so the function of SRm300 hasremained unclear (4). In our study for the functions ofTAXREB803, we found that it enhanced Tax-mediated tran-scription, which was a significantly different feature from thepreviously reported characteristics of SRL300 and SRm300.Here we report the function of TAXREB803 as a coactivatorfor Tax-induced transcriptions and discuss the physiologicalroles of this protein.
Although TAXREB803 is identical to SRL300, we refer tothis cellular protein as TAXREB803 because we initially iso-lated its partial cDNA fragment and named the encoded pep-tide TAXREB803.
MATERIALS AND METHODS
Cell culture. SaOS-2, COS-7, HEK 293T, and HeLa cells were grown inDulbecco’s modified Eagle’s medium (DMEM; Nissui, Tokyo, Japan) supple-mented with 10% fetal calf serum (Invitrogen, Carlsbad, Calif.), 4 mM L-glu-tamine (Nacalai Tesque, Tokyo, Japan), and 0.1 mg/ml of kanamycin sulfate(Meiji, Tokyo, Japan). MT-2 cells were grown in RPMI 1640 medium (Nissui)supplemented as the same as DMEM except for 2 mM L-glutamine.
cDNA cloning and sequencing. The 5� and 3� coding regions of the mRNA forTAXREB803 were obtained from SaOS-2 by 5� rapid amplification of cDNAends (RACE) with the 5� Full RACE Core Set (TaKaRa, Otsu, Japan) andreverse transcription (RT)-PCR. The cDNAs were subcloned into pGEM-TEasy vector (Promega, Madison, Wis.) and their nucleotide sequences weredetermined with a DYEnamic ET terminator cycle sequencing premix kit (Am-ersham, Piscataway, N.J.).
Plasmids. The cDNA containing the 5�- and the 3�-end regions of TAXREB803was inserted into AB002322 cDNA (kindly provided by Kazusa DNA ResearchInst.) to make the full open reading frame of TAXREB803, and the resultingcDNA was cloned into the vector pBluescript SK� (Stratagene, La Jolla, Calif.).The eukaryotic expression plasmid for TAXREB803 was constructed by insertingthe full-length cDNA into vector pcDNA3 (Invitrogen) with KpnI and NotI sites.The Kozak consensus sequence was inserted between the KpnI site and theputative start codon of TAXREB803 to make pcDNA3-TAXREB803. pcDNA3-Flag-TAXREB803 was constructed by inserting the Flag tag between the Kozaksequence and the start codon of TAXREB803. pcDNA3-Tax was used for theexpression of Tax (1).
Several luciferase-expressing reporter plasmids were used. pTxRE-luc con-tains five copies of TxRE in its promoter region (1). pGL3-LTR-luc plasmid wasconstructed by inserting the HindIII fragment containing the U3 region of theLTR from pHTLV-1 LTR-CAT (47) into the pGL3-basic vector (Promega).pCRE-luc (a tetramer of CRE enhancer), pNF-�B-luc (a pentamer of NF-�Benhancer), pSRE-luc (a pentamer of SRE enhancer), and pFC-MEKK plasmids
were purchased from Stratagene, and phRL-TK-luc plasmid was purchased fromPromega.
Antibodies. The rabbit polyclonal antiserum to TAXREB803 was raised byinjecting the fusion protein of glutathione S-transferase (GST) and the peptideencoding amino acids 4 to 138 of TAXREB803, GST-TAXREB803(4-138), as animmunogenic antigen. Anti-Tax antibody is the mouse monoclonal antibodyagainst Tax. Rabbit polyclonal antiserum to Flag tag peptide was purchased fromSigma (Saint Louis, Mo.). Goat anti-mouse immunoglobulin lissamine rhoda-mine B conjugate and goat anti-rabbit immunoglobulin fluorescein isothiocya-nate were purchased from Molecular Probes (Eugene, Oreg.). Mouse monoclo-nal antisera to CREB and SC-35 were purchased from Santa Cruz Biotechnology(Santa Cruz, Calif.). Rabbit normal immunoglobulin G was purchased fromZymed Laboratories (San Francisco, Calif.).
Production of GST fusion proteins. The cDNA fragments encoding aminoacids 4 to 138 and 1616 to 1718 of the TAXREB803 open reading frame wereinserted into pGEX-6p vector (Amersham) to produce fusion proteins GST-TAXREB803(4-138) and GST-TAXREB803(1616-1718), respectively. The ex-pression plasmid for GST-CREB was provided by T. Ohshima (Institute forVirus Research, Kyoto University). These GST fusion proteins were produced inEscherichia coli BL21. GST-TAXREB803(4-138) was purified with glutathione-Sepharose resin (Amersham).
Detection of DNA-protein interaction by Southwestern blotting. The bindingof GST fusion proteins to 32P-labeled TxRE or TxRE mutant probe was analyzedas previously described (55). Crude extracts of GST, GST-TAXREB803(1616-1718), and GST-CREB were separated by SDS–10% PAGE and blotted onto apolyvinylidene difluoride membrane (Millipore, Bedford, Mass.). The membranewas submerged in 50 ml of binding buffer containing 25 mM NaCl, 5 mM MgCl2,25 mM HEPES (pH 7.9), and 0.5 mM dithiothreitol supplemented with 6 Mguanidine hydrochloride. After gentle shaking for 10 min, the solutions weredecanted and replaced with 50 ml of the same buffer. A second buffer containing6 M guanidine hydrochloride was diluted with a 100% dilution method aspreviously described (55); after this, the membrane was blocked by incubatingwith binding buffer containing 5% nonfat dry milk and then washed three timeswith binding buffer containing 0.25% dry milk. Next, the membrane was exposedto DNA probes by incubating overnight with binding buffer containing 0.25% drymilk, 2 �g of poly(dI-dC) (Amersham) per ml, and 2 � 105 cpm of 32P-labeledprobes per ml. The membrane was washed with binding buffer, air-dried, andsubjected to autoradiography.
The probes used for Southwestern blotting were prepared by labeling theannealed DNA of 5�-GACTAAGGCTCTGACGTCTCCCCCC-3� for the posi-tive strand and 5�-GACTGGGGGGAGACGTCAGAGCCTT-3� for the nega-tive strand for TxRE and of 5�-GACTAAGGCTCTGACGTCTTAATCG-3� forthe positive strand and 5�-AGCTCGATTAAGACGTCAGAGCCTT-3� for thenegative strand for mutant TxRE. The radiolabeling was performed by thefilling-in reaction of Klenow fragment (Toyobo, Osaka, Japan) with [�-32P]dCTP(Amersham).
Transient transfection and luciferase assays. Transfection of plasmids wasperformed with FuGene 6 transfection reagent (Roche Molecular Biochemicals,Indianapolis, Ind.). The reporter assays were performed with the luciferase assaysystem according to the manufacturer’s protocol (Promega). The firefly lucif-erase activity from pTxRE-luc, pCRE-luc, and pGL3-LTR-luc was measuredwith the luciferase assay system (Promega). Dual luciferase assay for firefly andRenilla luciferase was performed with the dual luciferase reporter assay system(Promega). pcDNA3 vector plasmid was used as a carrier DNA to equalize theDNA concentration for each transfection. All experiments were performed intriplicate, and results were obtained from at least three separate experiments.
RNA isolation and RT-PCR. Total RNA was isolated with Sepasol-RNA I(Nacalai Tesque) according to the manufacturer’s protocol. The RNA was fur-ther treated with RNase-free DNase I (Stratagene) for 30 min at 37°C, and theDNase I was inactivated by heating for 10 min at 80°C. RT for luciferase mRNAwas performed with Superscript II reverse transcriptase (Invitrogen) with theprimer 5�-GCTGATGTAGTCTCAGTGAG-3�, which is complementary to thecDNA sequence for the firefly luciferase, and followed by PCR with Taq poly-merase (Sigma) with a set of primers, 5�-CAGCCTACCGTGGTGTTC-3� and5�-CCTGAAGGCTCCTCAG-3�. Then, the PCR was conducted for 27 cycles,each cycle consisting of denaturation (94°C for 30 s), annealing (57°C for 30 s),and extension (72°C for 1 min). Reverse transcription was carried out in areaction volume of 20 �l in which 1 �g of total RNA was present. mRNA forglyceraldehyde 3-phosphate dehydrogenase (GAPDH) was used as the internalcontrol for the RT-PCR. RT-PCR for TAXREB803 mRNA in HeLa cells wasperformed with a One Step RNA PCR kit (TaKaRa) with the primers 5�-AAACCTGCAAGCCCCAAGAAG-3� and 5�-GCTGCTCTGCTGGTC-3�.
10016 YOUN ET AL. J. VIROL.
on February 14, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from
RNA transfection. RNA was transcribed in vitro with a MEGAscript SP6 kitor T7 kit (Ambion, Austin, Tex.) from a linearized plasmid template of pGL3-luc(basic) or pcDNA3-Tax. RNA transfection was performed with DMRIE-C re-agent (Invitrogen) according to the manufacturer’s protocol. The transfectionmedium was changed to normal culture medium 4 h after the transfection, andcells were harvested 24 h after the medium change.
DNA affinity precipitations. Biotin-labeled DNA probe was prepared by thefilling-in reaction of Klenow fragment with a template oligonucleotide, 5�-biotin-(TAAGGCTCTGACGTCTCCCCC)�3-3�, containing three repeats of TxREbiotinylated at the 5� end, and a primer oligonucleotide, 5�-GGGGGAGACGTCAGA-3�. The probe was composed of three repeats of CRE and was preparedwith a template oligonucleotide, 5�-biotin-(CCTTGGCTGACGTCAGAGAGA)�3-3�, which is biotinylated at the 5� end, and a primer oligonucleotide, 5�-TCTCTCTGACGTCAG-3�.
Nuclear extracts were prepared as previously described (38) and diluted withbinding buffer consisting of 10 mM HEPES (pH 7.9), 150 mM NaCl, 1 mMEGTA, 5% glycerol, and 1 mM dithiothreitol. The diluted nuclear extract wasmixed with 2 �g of biotinylated DNA probe and 15 �g of poly(dI-dC) and thenincubated for 30 min on ice. Streptavidin-coated MagneSphere paramagneticparticles (Promega) were added to the mixture. which was rotated for 30 min at4°C. Streptavidin beads were collected with the magnetic separation stands andwashed four times with cold binding buffer. The trapped proteins were dissolvedwith loading buffer and analyzed by SDS-PAGE followed by immunoblotting.
Immunoprecipitations and immunoblots. Cell pellets were lysed in 1 ml ofimmunoprecipitation buffer [50 mM Tris-HCl buffer (pH 8.0) containing 0.5%Nonidet P-40, 150 mM NaCl, 1 mM EDTA, 1 mM dithiothreitol, and 0.25 mMPMSF]. Cell lysates were precleared with protein G-Sepharose beads (Amer-sham) for 1 h and subjected to immunoprecipitation by incubation with thedesignated antibodies at 4°C overnight. The immunocomplexes were recoveredby adding protein G-Sepharose beads, and the trapped proteins were analyzed byWestern blotting as previously described (43).
DNA-protein coimmunoprecipitation assay. 293T cells transfected with theindicated plasmids were harvested and subjected to immunoprecipitation asdescribed above with anti-CREB antibody, anti-TAXREB803 antibody, anti-Taxantibody, anti-Flag antibody, or normal rabbit IgG. The protein G-Sepharosebeads containing immunocomplexes were suspended in 50 ml of TE buffer. DNAwas extracted from the immunocomplex with TE-saturated phenol and precipi-tated with ethanol. The DNA was then dissolved in distilled water, and 10% ofthe recovered DNA was used as a template for the PCR. The primers used toamplify the U3 region of HTLV-1 LTR in pGL3-LTR-luc were 5�-CCGAGAAACAGAAGTCTG-3� and 5�-GTGATGTCCACCTC-3�. The PCR was con-ducted for 30 cycles, and each cycle consisted of denaturation (94°C for 30 s),annealing (55°C for 30 s), and extension (72°C for 1 min).
Metabolic labeling of cells with [35S]methionine. Cells were cultured in me-thionine-free DMEM or RPMI 1640 (ICN Biomedicals, Aurora, Ohio) supple-mented with dialyzed fetal bovine serum (Invitrogen), glutamine, and 100 �Ci of
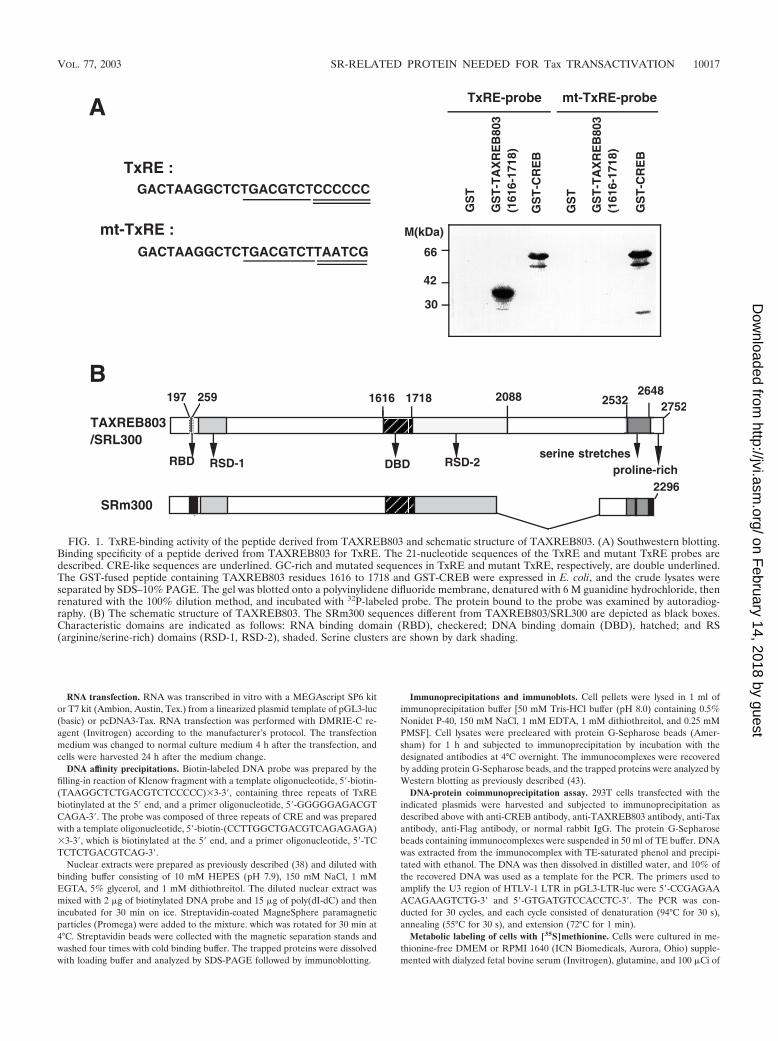
FIG. 1. TxRE-binding activity of the peptide derived from TAXREB803 and schematic structure of TAXREB803. (A) Southwestern blotting.Binding specificity of a peptide derived from TAXREB803 for TxRE. The 21-nucleotide sequences of the TxRE and mutant TxRE probes aredescribed. CRE-like sequences are underlined. GC-rich and mutated sequences in TxRE and mutant TxRE, respectively, are double underlined.The GST-fused peptide containing TAXREB803 residues 1616 to 1718 and GST-CREB were expressed in E. coli, and the crude lysates wereseparated by SDS–10% PAGE. The gel was blotted onto a polyvinylidene difluoride membrane, denatured with 6 M guanidine hydrochloride, thenrenatured with the 100% dilution method, and incubated with 32P-labeled probe. The protein bound to the probe was examined by autoradiog-raphy. (B) The schematic structure of TAXREB803. The SRm300 sequences different from TAXREB803/SRL300 are depicted as black boxes.Characteristic domains are indicated as follows: RNA binding domain (RBD), checkered; DNA binding domain (DBD), hatched; and RS(arginine/serine-rich) domains (RSD-1, RSD-2), shaded. Serine clusters are shown by dark shading.
VOL. 77, 2003 SR-RELATED PROTEIN NEEDED FOR Tax TRANSACTIVATION 10017
on February 14, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from
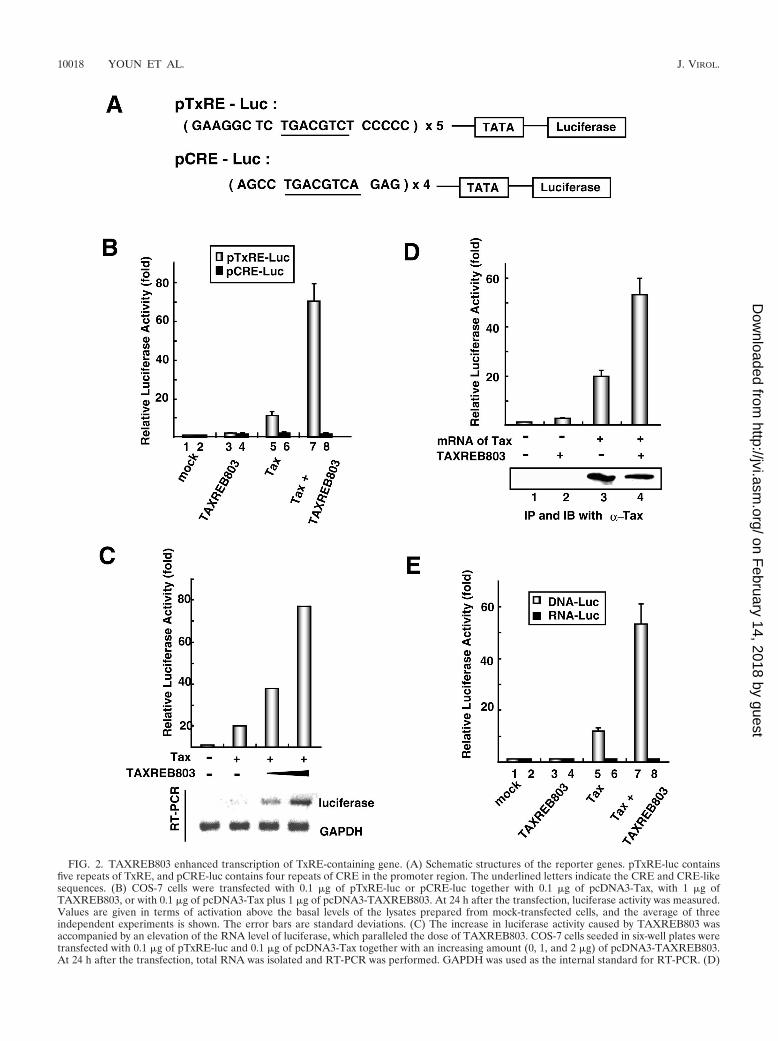
FIG. 2. TAXREB803 enhanced transcription of TxRE-containing gene. (A) Schematic structures of the reporter genes. pTxRE-luc containsfive repeats of TxRE, and pCRE-luc contains four repeats of CRE in the promoter region. The underlined letters indicate the CRE and CRE-likesequences. (B) COS-7 cells were transfected with 0.1 �g of pTxRE-luc or pCRE-luc together with 0.1 �g of pcDNA3-Tax, with 1 �g ofTAXREB803, or with 0.1 �g of pcDNA3-Tax plus 1 �g of pcDNA3-TAXREB803. At 24 h after the transfection, luciferase activity was measured.Values are given in terms of activation above the basal levels of the lysates prepared from mock-transfected cells, and the average of threeindependent experiments is shown. The error bars are standard deviations. (C) The increase in luciferase activity caused by TAXREB803 wasaccompanied by an elevation of the RNA level of luciferase, which paralleled the dose of TAXREB803. COS-7 cells seeded in six-well plates weretransfected with 0.1 �g of pTxRE-luc and 0.1 �g of pcDNA3-Tax together with an increasing amount (0, 1, and 2 �g) of pcDNA3-TAXREB803.At 24 h after the transfection, total RNA was isolated and RT-PCR was performed. GAPDH was used as the internal standard for RT-PCR. (D)
10018 YOUN ET AL. J. VIROL.
on February 14, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

[35S]methionine per ml (43.5 TBq/mmole, ICN Biomedicals). At 6 h after theculture, cells were harvested and used for immunoprecipitation as describedabove. The immunoprecipitants were analyzed by SDS–7% PAGE followed byautoradiography. Protein standards (Bio-Rad Laboratories, Hercules, Calif.)were used as a molecular weight marker.
Immunocytochemistry and confocal microscopic observation. COS-7 cellswere seeded on glass coverslips in six-well plates and transfected with 2 �g ofpcDNA3-Flag-TAXREB803 and 0.5 �g of pcDNA3-Tax. At 24 h after theculture, cells were washed twice in phosphate-buffered saline (PBS) and fixed bytreatment for 5 min at room temperature with 3.6% formaldehyde in PBS. Thenthey were permeabilized by treatment for 15 min with 0.1% Nonidet P-40 in PBS.Cells were washed in PBS and incubated with 3% bovine serum albumin in PBSfor 1 h at room temperature. Primary antibodies (rabbit anti-Flag antibody forFlag-tagged TAXREB803, mouse anti-Tax antibody, or mouse anti-SC35 anti-body) diluted in PBS were added and incubated for 1 h at room temperature.Then, secondary antibody (FITC 488 or rhodamine 568) was added and incu-bated for 40 min at room temperature in the dark. DNA was counterstained with4�,6�-diamidino-2-phenylindole (DAPI), and final preparations were mounted in90% glycerol in PBS containing 0.01% p-phenylenediamine. Observations wereperformed with a confocal microscope (Axio Vision; Carl Zeiss, Hallbergmoos,Germany).
RNA interference. A duplex of 21-nucleotide small interfering RNA (siRNA)corresponding to the sequence of mRNA for TAXREB803 (nucleotides 649 to669) with the addition of 2 nucleotides of 2-deoxythymidine attached to the 3�end was synthesized and annealed (B-Bridge International, San Jose, Calif.). Thesequence of the duplex is 5�-GCACAGGUCAGAAUCUGAGdTdT in the senseorientation and 5�-CUCAGAUGACCUGUGCdTdT in the antisense orienta-tion. HeLa cells were transfected with 25 �l of 20 �M siRNA duplex per 6-cmdish with Oligofectamine (Invitrogen) as previously described (9). Four hoursafter the siRNA transfection, the medium was replaced with the normal culturemedium. For the RNA analysis, 48 h after the siRNA transfection, total RNAwas isolated from the cells. For the protein analysis, cells were incubated for 66 hafter the siRNA transfection and subjected to metabolic labeling with [35S]me-thionine for 6 h. For the reporter assay, 24 h after the siRNA transfection, cellswere reseeded into 12-well dishes and incubated for another 24 h, and thenreporter plasmids were transfected with pcDNA3-Tax. At 24 h after the trans-fection of the reporter plasmids, the expressions of firefly luciferase from pGL3-LTR-luc and Renilla luciferase activity from phRL-TK-luc were evaluated withthe dual luciferase reporter assay system (Promega).
RESULTS
Cloning of full-length cDNA for TxRE-binding proteinTAXREB803. We previously cloned a partial cDNA for aTxRE-binding protein by Southwestern blotting with a probecontaining the 21-bp enhancer sequence (TxRE) residing inthe HTLV-1 LTR and named the product of the geneTAXREB803 (53). The cloned cDNA has an open readingframe coding a peptide with 103 amino acid residues. To an-alyze the region of TxRE which associates with this peptide, weperformed a Southwestern blotting analysis with 32P-labeledoligonucleotides for the wild-type TxRE and mutant TxRE inwhich the 3� GC-rich sequences were mutated. The GST-fusedprotein containing the 103 amino acids of TAXREB803 (ac-tually defined as amino acids 1616 to 1718 of the full sequence)bound to the probe of wild-type TxRE but failed to interactwith the mutant TxRE (Fig. 1A). GST-fused CREB protein
bound to both of the probes independently of the 3� GC-richsequences.
Previous analysis showed that TAXREB803 interacted withthe pyrimidine-rich region of the TxRE (data not shown), andthus, it was suggested that the sites for the interaction withTAXREB803 and CREB did not overlap at the TxRE. Inorder to clone the full-length cDNA for TAXREB803, wesearched GenBank and found one homologous sequence withaccession number AB002322. The cDNA of AB002322 is apartial clone lacking the 5� coding region of TAXREB803. Weconducted an analysis on the entire cDNA corresponding toAB002322 in cDNA from human SaOS-2 cells with RT-PCRand found a cDNA that was different from the 3� region ofAB002322. We obtained a cDNA for the 5� upstream regioncontaining the putative start codon by 5�-RACE. We con-structed the full-length cDNA for TAXREB803 with cDNAsof the 5� and 3� regions cloned from SaOS-2 cells and ofAB002322,
The protein deduced from the sequence of the cDNA con-sisted of 2,752 amino acid residues, with several unique fea-tures in its sequence. It contains many arginine/serine-richsequences (RS motifs) which are conserved in the SR familyproteins (12), two serine stretches (repeats of 25 and 42 resi-dues), and a proline-rich domain in the C terminus.TAXREB803 lacked the typical DNA binding motif as well asthe RNA recognition motif which is often seen in other SRfamily proteins. The DNA binding domain (DBD) is situatedbetween positions 1616 and 1718 of the amino acid sequence ofthis putative protein. The schematic structure of this protein,together with those of SRL300 and SRm300, is shown in Fig.1B.
TAXREB803 strongly enhanced Tax-induced transcriptionfrom HTLV-1 LTR. It is thought that Tax forms a ternarycomplex with CREB/ATF at TxRE sites and lead to transcrip-tional activation (56, 57). To investigate the effect ofTAXREB803 on Tax-mediated LTR transactivation, we ana-lyzed the transactivational function of Tax with the pTxRE-lucand pCRE-luc reporter genes. These contain five repeats ofTxRE and four repeats of cellular CRE in their promoterregions, respectively (Fig. 2A). No significant transcriptionalactivation of pTxRE-luc and pCRE-luc was observed whenTAXREB803 was overexpressed alone in COS-7 cells. How-ever, cotransfection of plasmids expressing TAXREB803 andTax resulted in the marked enhancement of Tax-induced ex-pression from pTxRE-luc (Fig. 2B, lanes 5 and 7) but not frompCRE-luc (Fig. 2B, lanes 6 and 8). This enhanced transactiva-tion of the luciferase gene with the TxRE promoter was de-pendent on the dose of TAXREB803 and accompanied anincrease in the mRNA for luciferase (Fig. 2C). These obser-
Enhanced transcription of HTLV-1 LTR by TAXREB803 is not due to an increase in Tax protein level. COS-7 cells were transfected with 0.1 �gof pTxRE-luc together with empty vector or 2 �g of pcDNA3-TAXREB803. Then 2 �g of in vitro-transcribed Tax mRNA was transfected 6 h afterthe DNA transfection, and luciferase activity was measured after another 24 h. The Tax protein levels were evaluated by immunoprecipitation andimmunoblotting with the anti-Tax antibody. (E) TAXREB803 did not affect luciferase expression at the posttranscriptional level. Two sets ofCOS-7 cells were transfected with 0.1 �g of pcDNA3-Tax with or without 1 �g of pcDNA3-TAXREB803 as indicated. Six hours after thetransfection, one set of cells was transfected with 0.1 �g of pTxRE-luc (DNA-luc) and the other with 2 �g of in vitro-transcribed luciferase mRNA(RNA-luc). Luciferase activity was measured 24 h after the second transfection. Error bars show standard deviations.
VOL. 77, 2003 SR-RELATED PROTEIN NEEDED FOR Tax TRANSACTIVATION 10019
on February 14, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from
vations clearly show that TAXREB803 enhances Tax transac-tivation but leave it unclear at which step it contributes.
We observed slightly enhanced expression of Tax whenpcDNA3-Tax was cotransfected with pcDNA3-TAXREB803,and it was also observed that the luciferase expression frompCMV-luc was slightly enhanced by TAXREB803 (data notshown), so there was a possibility that TAXREB803 enhancedthe expression of Tax through cytomegalovirus promoter ofpcDNA3 vector and that the increased luciferase activity byTax and TAXREB803 resulted from the increased productionof Tax. To analyze the context of this increase in luciferaseactivity more precisely, we prepared in vitro-transcribed RNAfor Tax and introduced it into COS-7 cells with the expressionplasmid of TAXREB803. At 24 h after the transfection, theexpression of Tax was examined by immunoprecipitation andimmunoblotting with an anti-Tax antibody. While the lucif-erase expression was enhanced by the addition of TAXREB803,the Tax expression levels were not affected (Fig. 2D, lanes 3
and 4). This result shows that TAXREB803 has a function ofenhancing the Tax-induced transcription independently of theexpression level of Tax.
Next, we examined the effect of TAXREB803 on the stabil-ity of luciferase RNA. In vitro-transcribed RNA for luciferase(RNA-luc) was transfected into the cells that had been trans-fected with the expression plasmids for TAXREB803 and/orTax at 6 h before the RNA transfection. The production ofluciferase from RNA-luc was not changed by the presence ofTax and/or TAXREB803 (Fig. 2E, lanes 4, 6, and 8). Theseresults indicate that the enhancement of transactivation of Taxby TAXREB803 occurs at the transcriptional level, not theposttranscriptional level.
TAXREB803 enhanced association of CREB with TxRE. Toinvestigate whether TAXREB803 plays a role in the binding ofCREB to TxRE, we used an in vitro DNA affinity precipitationassay. Nuclear extracts were prepared from 293T cells trans-fected with an empty vector, with TAXREB803 expression
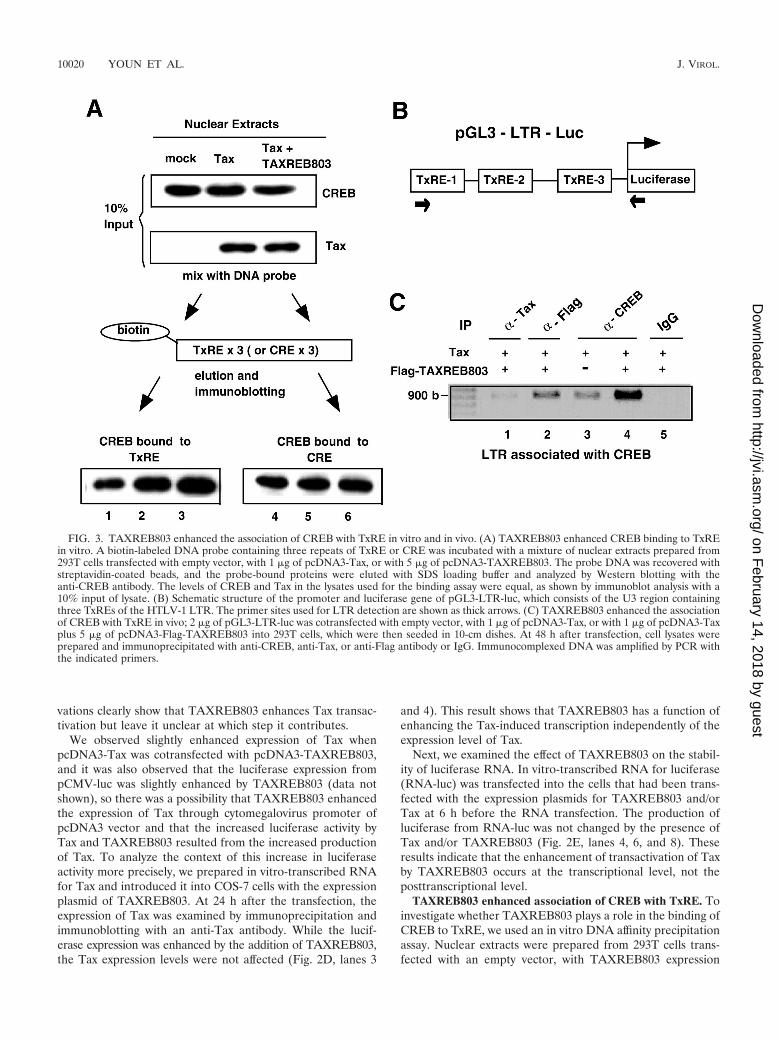
FIG. 3. TAXREB803 enhanced the association of CREB with TxRE in vitro and in vivo. (A) TAXREB803 enhanced CREB binding to TxREin vitro. A biotin-labeled DNA probe containing three repeats of TxRE or CRE was incubated with a mixture of nuclear extracts prepared from293T cells transfected with empty vector, with 1 �g of pcDNA3-Tax, or with 5 �g of pcDNA3-TAXREB803. The probe DNA was recovered withstreptavidin-coated beads, and the probe-bound proteins were eluted with SDS loading buffer and analyzed by Western blotting with theanti-CREB antibody. The levels of CREB and Tax in the lysates used for the binding assay were equal, as shown by immunoblot analysis with a10% input of lysate. (B) Schematic structure of the promoter and luciferase gene of pGL3-LTR-luc, which consists of the U3 region containingthree TxREs of the HTLV-1 LTR. The primer sites used for LTR detection are shown as thick arrows. (C) TAXREB803 enhanced the associationof CREB with TxRE in vivo; 2 �g of pGL3-LTR-luc was cotransfected with empty vector, with 1 �g of pcDNA3-Tax, or with 1 �g of pcDNA3-Taxplus 5 �g of pcDNA3-Flag-TAXREB803 into 293T cells, which were then seeded in 10-cm dishes. At 48 h after transfection, cell lysates wereprepared and immunoprecipitated with anti-CREB, anti-Tax, or anti-Flag antibody or IgG. Immunocomplexed DNA was amplified by PCR withthe indicated primers.
10020 YOUN ET AL. J. VIROL.
on February 14, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from
plasmid, or with Tax expression plasmid. Each nuclear extractwas mixed as designated in Fig. 3A (mock, Tax, or Tax plusTAXREB803) and then added with the 5�-biotinylated probecomposed of three repeats of the TxRE or CRE. In the inputof nuclear extracts, the amounts of CREB or Tax were ad-justed to the same level. The amount of CREB bound to TxREincreased in the presence of Tax (Fig. 3A, lanes 1 and 2), whichis consistent with previous reports (6, 18, 56, 57). This en-hanced binding of CREB to TxRE was further enhanced byTAXREB803 (Fig. 3A, lanes 2 and 3). But CREB binding toCRE was not changed (Fig. 3A, lanes 5 and 6).
Next, we analyzed the in vivo association of CREB andTxRE with a DNA-protein coprecipitation assay. We con-structed pGL3-LTR-luc, where the promoter included theunique 3� sequence (U3) containing three TxRE repeats of theHTLV-1 provirus (Fig. 3B). 293T cells were cotransfected withpGL3-LTR-luc together with the expression plasmids, asshown in Fig. 3C. After preparing the cell lysate, immunopre-cipitation was performed with the anti-CREB, anti-Flag, andanti-Tax antibodies and IgG. Then, we analyzed the immuno-complex-associated LTR. DNA was recovered from immuno-complexws and amplified by PCR with the primers indicated bya thick arrow in the diagram of pGL3-LTR-luc (Fig. 3B). Theexpected size of the PCR product was about 900 bases. LTRwas detected in the immunoprecipitant by anti-Tax antibody aswell as by anti-Flag antibody (Fig. 3B, lanes 1 and 2). The levelof CREB-associated LTR was increased to the higher level inthe cells overproducing both Tax and TAXREB803 than Taxalone (lanes 3 and 4). These results suggest that TAXREB803enhances the association of CREB with TxRE in cooperationwith Tax.
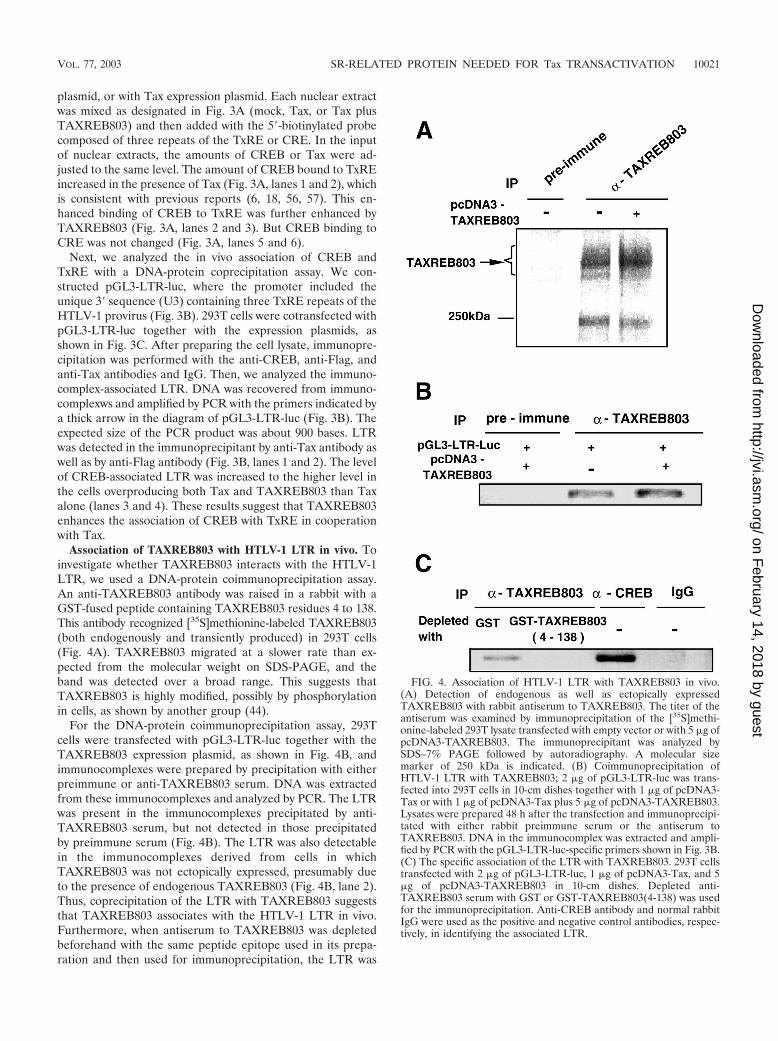
Association of TAXREB803 with HTLV-1 LTR in vivo. Toinvestigate whether TAXREB803 interacts with the HTLV-1LTR, we used a DNA-protein coimmunoprecipitation assay.An anti-TAXREB803 antibody was raised in a rabbit with aGST-fused peptide containing TAXREB803 residues 4 to 138.This antibody recognized [35S]methionine-labeled TAXREB803(both endogenously and transiently produced) in 293T cells(Fig. 4A). TAXREB803 migrated at a slower rate than ex-pected from the molecular weight on SDS-PAGE, and theband was detected over a broad range. This suggests thatTAXREB803 is highly modified, possibly by phosphorylationin cells, as shown by another group (44).
For the DNA-protein coimmunoprecipitation assay, 293Tcells were transfected with pGL3-LTR-luc together with theTAXREB803 expression plasmid, as shown in Fig. 4B, andimmunocomplexes were prepared by precipitation with eitherpreimmune or anti-TAXREB803 serum. DNA was extractedfrom these immunocomplexes and analyzed by PCR. The LTRwas present in the immunocomplexes precipitated by anti-TAXREB803 serum, but not detected in those precipitatedby preimmune serum (Fig. 4B). The LTR was also detectablein the immunocomplexes derived from cells in whichTAXREB803 was not ectopically expressed, presumably dueto the presence of endogenous TAXREB803 (Fig. 4B, lane 2).Thus, coprecipitation of the LTR with TAXREB803 suggeststhat TAXREB803 associates with the HTLV-1 LTR in vivo.Furthermore, when antiserum to TAXREB803 was depletedbeforehand with the same peptide epitope used in its prepa-ration and then used for immunoprecipitation, the LTR was
FIG. 4. Association of HTLV-1 LTR with TAXREB803 in vivo.(A) Detection of endogenous as well as ectopically expressedTAXREB803 with rabbit antiserum to TAXREB803. The titer of theantiserum was examined by immunoprecipitation of the [35S]methi-onine-labeled 293T lysate transfected with empty vector or with 5 �g ofpcDNA3-TAXREB803. The immunoprecipitant was analyzed bySDS–7% PAGE followed by autoradiography. A molecular sizemarker of 250 kDa is indicated. (B) Coimmunoprecipitation ofHTLV-1 LTR with TAXREB803; 2 �g of pGL3-LTR-luc was trans-fected into 293T cells in 10-cm dishes together with 1 �g of pcDNA3-Tax or with 1 �g of pcDNA3-Tax plus 5 �g of pcDNA3-TAXREB803.Lysates were prepared 48 h after the transfection and immunoprecipi-tated with either rabbit preimmune serum or the antiserum toTAXREB803. DNA in the immunocomplex was extracted and ampli-fied by PCR with the pGL3-LTR-luc-specific primers shown in Fig. 3B.(C) The specific association of the LTR with TAXREB803. 293T cellstransfected with 2 �g of pGL3-LTR-luc, 1 �g of pcDNA3-Tax, and 5�g of pcDNA3-TAXREB803 in 10-cm dishes. Depleted anti-TAXREB803 serum with GST or GST-TAXREB803(4-138) was usedfor the immunoprecipitation. Anti-CREB antibody and normal rabbitIgG were used as the positive and negative control antibodies, respec-tively, in identifying the associated LTR.
VOL. 77, 2003 SR-RELATED PROTEIN NEEDED FOR Tax TRANSACTIVATION 10021
on February 14, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from
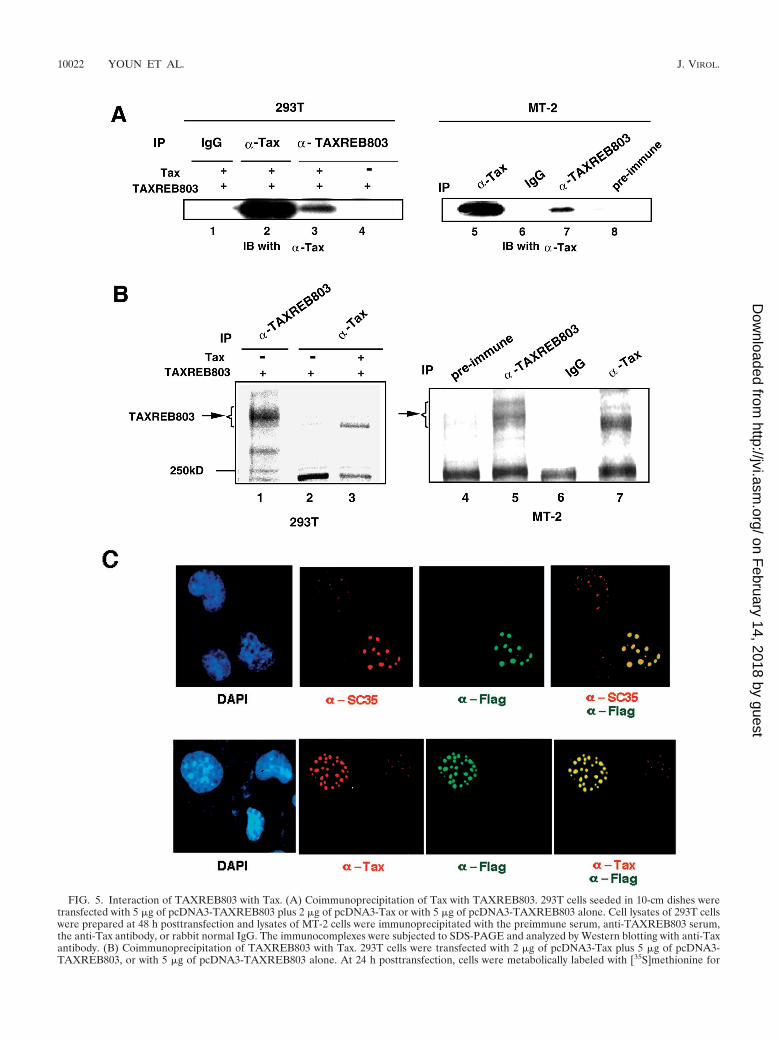
FIG. 5. Interaction of TAXREB803 with Tax. (A) Coimmunoprecipitation of Tax with TAXREB803. 293T cells seeded in 10-cm dishes weretransfected with 5 �g of pcDNA3-TAXREB803 plus 2 �g of pcDNA3-Tax or with 5 �g of pcDNA3-TAXREB803 alone. Cell lysates of 293T cellswere prepared at 48 h posttransfection and lysates of MT-2 cells were immunoprecipitated with the preimmune serum, anti-TAXREB803 serum,the anti-Tax antibody, or rabbit normal IgG. The immunocomplexes were subjected to SDS-PAGE and analyzed by Western blotting with anti-Taxantibody. (B) Coimmunoprecipitation of TAXREB803 with Tax. 293T cells were transfected with 2 �g of pcDNA3-Tax plus 5 �g of pcDNA3-TAXREB803, or with 5 �g of pcDNA3-TAXREB803 alone. At 24 h posttransfection, cells were metabolically labeled with [35S]methionine for
10022 YOUN ET AL. J. VIROL.
on February 14, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

not detected in the immunocomplex, but antiserum treatedwith GST could still precipitate the LTR (Fig. 4C, lanes 1 and2).
Association of TAXREB803 with Tax. In the promoter of theHTLV-1 gene, many factors, including Tax, CREB, p300, CBP,and AP-1 family members (17, 23, 25, 45, 49, 58), cooperateand promote transcription. We postulated that TAXREB803could be involved in their transcriptional complex, throughinteraction with certain factors. To verify our hypothesis, weexamined the association of TAXREB803 with Tax in cells. Inan immunoprecipitation assay, Tax was coimmunoprecipitatedwith TAXREB803 in MT-2 cells, HTLV-1-infected T cells, aswell as in 293T cells in which Tax and TAXREB803 weretransiently overproduced by expression plasmids (Fig. 5A,lanes 3 and 7), and in a reciprocal experiment, 35S-labeledTAXREB803 was detected in an immunocomplex precipitatedwith the anti-Tax antibody (Fig. 5B, lanes 3 and 7). Theseresults show that Tax and TAXREB803 interact in the cells,either directly or indirectly.
We also examined the cellular localization of these two pro-teins by expressing Tax and Flag tagged TAXREB803 inCOS-7 cells. TAXREB803 localized to the nuclear speckles ofinterchromatin granule clusters (IGCs)/RNA splicing bodies.We could identify these speckles as IGCs because SC-35, asplicing body component, colocalized with TAXREB803 (Fig.5C, upper panels). It has been reported that Tax also localizesto IGCs (46), and indeed we observed a tight colocalization ofTax with TAXREB803 in IGCs (Fig. 5C, lower panels). This isconsistent with our earlier observation of the in vivo associa-tion of Tax and TAXREB803 (Fig. 5A and B). Interestingly,IGC/RNA splicing bodies were enlarged in those cells in whichTAXREB803 was exogenously overexpressed (Fig. 5C).
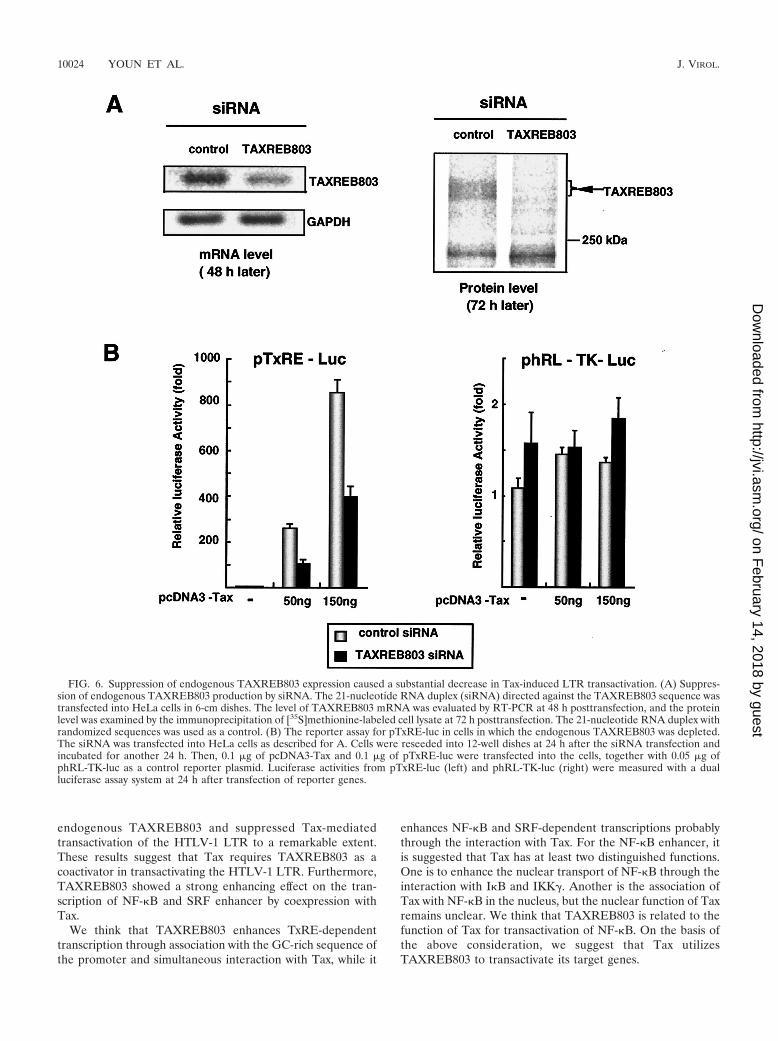
TAXREB803 is an important factor in Tax transactivation.The TAXREB803 gene is ubiquitously expressed in many dif-ferent cell lines and tissues (data not shown). The fact thatTAXREB803 is endogenously expressed in 293T cells (Fig.4B) allowed us to examine the capacity for endogenousTAXREB803-mediated enhancement of Tax-induced transac-tivation. To make cells in which the expression of endogenousTAXREB803 was repressed, we treated HeLa cells with smallinterfering RNA (siRNA), the duplex RNA encoding nucleo-tides 651 to 669 of TAXREB803. TAXREB803 expression wassignificantly suppressed at both the RNA and protein levels,while the level of other cellular protein such as GAPDH wasnot changed (Fig. 6A). TAXREB803 expression was not af-fected by the control siRNA with randomized sequences. In-terestingly, when expression of endogenous TAXREB803 wassuppressed, Tax-induced luciferase expression of pTxRE-lucdecreased by almost 60% (Fig. 6B, left panel, black bars).However, Renilla luciferase expression from phRL-TK-luc,
which contains the herpes simplex virus thymidine kinase pro-moter, showed no significant change (Fig. 6B, right panel).Taken together, these results show that TAXREB803 plays akey role in Tax-induced HTLV-1 LTR transactivation.
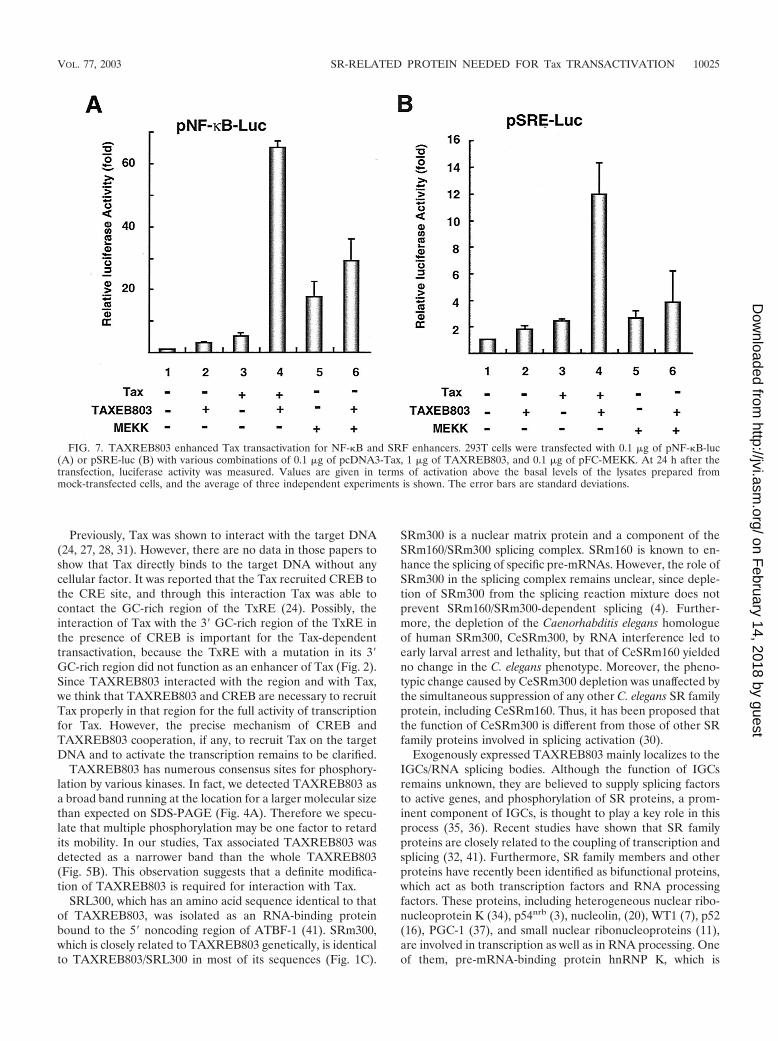
TAXREB803 enhanced Tax-mediated activation of NF-�Band SRE enhancer. In the above results, it was suggestedthat TAXREB803 was an important cellular factor for thetranscriptional function of Tax, so we examined whetherTAXREB803 affected Tax-dependent activation of genes con-taining an NF-�B or SRF enhancer. In the reporter gene assay,Tax-induced transactivation of pNF-�B-luc and pSRE-luc wasstrongly enhanced by TAXREB803 by approximately 10-fold(Fig. 7A and B, lanes 3 and 4). However, MEKK-derivedactivation of these genes was only slightly enhanced byTAXREB803 (about 1.5-fold; Fig. 7A and B, lanes 5 and 6).From this result, we suggest that TAXREB803 is an importantfactor for Tax transactivation for NF-�B and the SRF en-hancer as well as the HTLV-1 LTR.
DISCUSSION
In this paper, we report a new coactivator, TAXREB803, forTax-mediated transactivation of HTLV-1 gene expression.Transactivation of the HTLV-1 LTR by Tax requires the par-ticipations of various cellular transcription factors and coacti-vators (17, 23, 25, 49, 58). We had previously cloned a partialcDNA of TAXREB803 as a TxRE-binding protein by South-western blotting. This partial cDNA encoded 103 amino acidresidues, and the binding of the peptide to the TxRE requiredthe 3� GC-rich sequence in TxRE (Fig. 1A). In spite of itsTxRE-binding ability, a characteristic DNA-binding motif wasnot found in the peptide. On the other hand, TAXREB803 hasdistinctive amino acid sequences which include arginine/serine(RS)-rich domains scattered throughout. In addition to theseRS motifs, there are two clusters with an extraordinarily highnumber of repeated serine residues in the 3�-terminal region.
Exogenously expressed TAXREB803 markedly enhancedTax-induced transcription of the gene containing the HTLV-1LTR. We observed several characteristic features ofTAXREB803 which might be closely related to its ability toenhance transcription: TAXREB803 enhanced CREB bindingto TxRE both in vitro and in vivo, TAXREB803 associatedwith the TxRE-containing region of the LTR in vivo,TAXREB803 associated with Tax in vivo, and Tax andTAXREB803 very tightly colocalized to ICG/RNA splicingbodies. In order to analyze the contribution of endogenousTAXREB803 to enhancing Tax-mediated LTR transactiva-tion, we suppressed endogenous production of TAXREB803by RNA interference. The transfection of siRNA directedagainst TAXREB803 substantially reduced the expression of
6 h. MT-2 cells were also labeled for 6 h. The lysates prepared from the cells were immunoprecipitated with the anti-Tax antibody. The resultingimmunocomplex was analyzed by SDS–7% PAGE followed by autoradiography. The position of a protein of 250 kDa is shown as a molecular sizemarker. (C) The subcellular colocalization of Tax and TAXREB803. COS-7 cells were seeded on glass coverslips. After incubation for 24 h, 0.5�g of pcDNA3-Tax and 2 �g of pcDNA3-Flag-TAXREB803 were transfected. The cells were fixed at 24 h posttransfection, permeabilized, andstained by dual immunofluorescence with primary antibodies (rabbit polyclonal antibody to Flag and mouse monoclonal antibody to Tax andSC-35) and fluoroconjugated secondary antibodies. DNA was counterstained with DAPI, while Tax and SC-35 were stained with rhodamineconjugate (red), and TAXREB803 was stained with FITC conjugate (green). The overlay of TXAREB803 with SC-35 (upper panel) or with Tax(lower panel) is shown as yellow.
VOL. 77, 2003 SR-RELATED PROTEIN NEEDED FOR Tax TRANSACTIVATION 10023
on February 14, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from
endogenous TAXREB803 and suppressed Tax-mediatedtransactivation of the HTLV-1 LTR to a remarkable extent.These results suggest that Tax requires TAXREB803 as acoactivator in transactivating the HTLV-1 LTR. Furthermore,TAXREB803 showed a strong enhancing effect on the tran-scription of NF-�B and SRF enhancer by coexpression withTax.
We think that TAXREB803 enhances TxRE-dependenttranscription through association with the GC-rich sequence ofthe promoter and simultaneous interaction with Tax, while it
enhances NF-�B and SRF-dependent transcriptions probablythrough the interaction with Tax. For the NF-�B enhancer, itis suggested that Tax has at least two distinguished functions.One is to enhance the nuclear transport of NF-�B through theinteraction with I�B and IKK�. Another is the association ofTax with NF-�B in the nucleus, but the nuclear function of Taxremains unclear. We think that TAXREB803 is related to thefunction of Tax for transactivation of NF-�B. On the basis ofthe above consideration, we suggest that Tax utilizesTAXREB803 to transactivate its target genes.
FIG. 6. Suppression of endogenous TAXREB803 expression caused a substantial decrease in Tax-induced LTR transactivation. (A) Suppres-sion of endogenous TAXREB803 production by siRNA. The 21-nucleotide RNA duplex (siRNA) directed against the TAXREB803 sequence wastransfected into HeLa cells in 6-cm dishes. The level of TAXREB803 mRNA was evaluated by RT-PCR at 48 h posttransfection, and the proteinlevel was examined by the immunoprecipitation of [35S]methionine-labeled cell lysate at 72 h posttransfection. The 21-nucleotide RNA duplex withrandomized sequences was used as a control. (B) The reporter assay for pTxRE-luc in cells in which the endogenous TAXREB803 was depleted.The siRNA was transfected into HeLa cells as described for A. Cells were reseeded into 12-well dishes at 24 h after the siRNA transfection andincubated for another 24 h. Then, 0.1 �g of pcDNA3-Tax and 0.1 �g of pTxRE-luc were transfected into the cells, together with 0.05 �g ofphRL-TK-luc as a control reporter plasmid. Luciferase activities from pTxRE-luc (left) and phRL-TK-luc (right) were measured with a dualluciferase assay system at 24 h after transfection of reporter genes.
10024 YOUN ET AL. J. VIROL.
on February 14, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from
Previously, Tax was shown to interact with the target DNA(24, 27, 28, 31). However, there are no data in those papers toshow that Tax directly binds to the target DNA without anycellular factor. It was reported that the Tax recruited CREB tothe CRE site, and through this interaction Tax was able tocontact the GC-rich region of the TxRE (24). Possibly, theinteraction of Tax with the 3� GC-rich region of the TxRE inthe presence of CREB is important for the Tax-dependenttransactivation, because the TxRE with a mutation in its 3�GC-rich region did not function as an enhancer of Tax (Fig. 2).Since TAXREB803 interacted with the region and with Tax,we think that TAXREB803 and CREB are necessary to recruitTax properly in that region for the full activity of transcriptionfor Tax. However, the precise mechanism of CREB andTAXREB803 cooperation, if any, to recruit Tax on the targetDNA and to activate the transcription remains to be clarified.
TAXREB803 has numerous consensus sites for phosphory-lation by various kinases. In fact, we detected TAXREB803 asa broad band running at the location for a larger molecular sizethan expected on SDS-PAGE (Fig. 4A). Therefore we specu-late that multiple phosphorylation may be one factor to retardits mobility. In our studies, Tax associated TAXREB803 wasdetected as a narrower band than the whole TAXREB803(Fig. 5B). This observation suggests that a definite modifica-tion of TAXREB803 is required for interaction with Tax.
SRL300, which has an amino acid sequence identical to thatof TAXREB803, was isolated as an RNA-binding proteinbound to the 5� noncoding region of ATBF-1 (41). SRm300,which is closely related to TAXREB803 genetically, is identicalto TAXREB803/SRL300 in most of its sequences (Fig. 1C).
SRm300 is a nuclear matrix protein and a component of theSRm160/SRm300 splicing complex. SRm160 is known to en-hance the splicing of specific pre-mRNAs. However, the role ofSRm300 in the splicing complex remains unclear, since deple-tion of SRm300 from the splicing reaction mixture does notprevent SRm160/SRm300-dependent splicing (4). Further-more, the depletion of the Caenorhabditis elegans homologueof human SRm300, CeSRm300, by RNA interference led toearly larval arrest and lethality, but that of CeSRm160 yieldedno change in the C. elegans phenotype. Moreover, the pheno-typic change caused by CeSRm300 depletion was unaffected bythe simultaneous suppression of any other C. elegans SR familyprotein, including CeSRm160. Thus, it has been proposed thatthe function of CeSRm300 is different from those of other SRfamily proteins involved in splicing activation (30).
Exogenously expressed TAXREB803 mainly localizes to theIGCs/RNA splicing bodies. Although the function of IGCsremains unknown, they are believed to supply splicing factorsto active genes, and phosphorylation of SR proteins, a prom-inent component of IGCs, is thought to play a key role in thisprocess (35, 36). Recent studies have shown that SR familyproteins are closely related to the coupling of transcription andsplicing (32, 41). Furthermore, SR family members and otherproteins have recently been identified as bifunctional proteins,which act as both transcription factors and RNA processingfactors. These proteins, including heterogeneous nuclear ribo-nucleoprotein K (34), p54nrb (3), nucleolin, (20), WT1 (7), p52(16), PGC-1 (37), and small nuclear ribonucleoproteins (11),are involved in transcription as well as in RNA processing. Oneof them, pre-mRNA-binding protein hnRNP K, which is
FIG. 7. TAXREB803 enhanced Tax transactivation for NF-�B and SRF enhancers. 293T cells were transfected with 0.1 �g of pNF-�B-luc(A) or pSRE-luc (B) with various combinations of 0.1 �g of pcDNA3-Tax, 1 �g of TAXREB803, and 0.1 �g of pFC-MEKK. At 24 h after thetransfection, luciferase activity was measured. Values are given in terms of activation above the basal levels of the lysates prepared frommock-transfected cells, and the average of three independent experiments is shown. The error bars are standard deviations.
VOL. 77, 2003 SR-RELATED PROTEIN NEEDED FOR Tax TRANSACTIVATION 10025
on February 14, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

known to facilitate mRNA biogenesis, was found to act as atranscription factor (34). WT1 possesses both DNA bindingand transcriptional activity; moreover, it is incorporated intospliceosomes and interacts with the essential splicing factor(7). PGC-1, an SR-like protein containing both RNA bindingand RS domains, functions as a transcriptional coactivatorthrough association with promoters (37).
These bifunctional proteins are expected to participate inboth transcription and RNA processing and thereby to pro-mote efficient gene expression. TAXREB803 reported in thisreport may have the features of such bifunctional protein.Further characterization of TAXREB803 will elucidate thephysiological roles of this protein in gene expression.
ACKNOWLEDGMENTS
We are grateful to T. Morita for the initial study of TAXREB803, toKazusa DNA Research Inst. for the plasmid carrying the cDNA ofAB002322, and to M. Hijikata, Y. Ariumi, T. Ohshima, and othermembers of the Laboratory of Human Tumor Viruses for helpfulcomments and discussion.
This work was supported by Grants-in-Aid for Scientific Researchfrom the Ministry of Education, Culture, Sports and Technology ofJapan.
REFERENCES
1. Akagi, T., H. Ono, H. Nyunoya, and K. Shimotohno. 1997. Characterizationof peripheral blood T-lymphocytes transduced with HTLV-1 Tax mutantswith different trans-activating phenotypes. Oncogene 14:2071–2078.
2. Akagi, T., H. Ono, and K. Shimotohno. 1996. Expression of cell-cycle regu-latory genes in HTLV-1 infected T-cell lines: possible involvement of Tax1 inthe altered expression of cyclin D2, p18Ink4 and p21Waf1/Cip1/Sdi1. Oncogene12:1645–1652.
3. Basu, A., B. Dong, A. R. Krainer, and C. C. Howe. 1997. The intracisternalA-particle proximal enhancer-binding protein activates transcription and isidentical to the RNA- and DNA-binding protein p54nrb/NonO. Mol. Cell.Biol. 17:677–686.
4. Blencowe, B. J., G. Bauren, A. G. Eldridge, R. Issner, J. A. Nickerson, E.Rosonina, and P. A. Sharp. 2000. The SRm160/300 splicing coactivatorsubunits. RNA 6:111–120.
5. Brady, J., K.-T. Jeang, J. Duvall, and G. Khoury. 1987. Identification ofp40x-responsive regulatory sequences within the human T-cell leukemiavirus type I long terminal repeat. J. Virol. 61:2175–2181.
6. Brauweiler, A., P. Garl, A. A. Franklin, H. A. Giebler, and J. K. Nyborg.1995. A molecular mechanism for human T-cell leukemia virus latency andtax transactivation. J. Biol. Chem. 270:12814–12822.
7. Davies, R. C., C. Calvio, E. Bratt, S. H. Larsson, A. I. Lamond, and N. D.Hastie. 1998. WT1 interacts with the splicing factor U2AF65 in an isoform-dependent manner and can be incorporated into spliceosomes. Genes Dev.12:3217–3225.
8. Dignam, J. D., R. M. Lebovitz, and R. G. Roeder. 1983. Accurate transcrip-tion initiation by RNA polymerase II in a soluble extract from isolatedmammalian nuclei. Nucleic Acids Res. 11:1475–1489.
9. Elbashir, S. M., J. Harborth, W. Lendeckel, A. Yalcin, K. Weber, and T.Tuschi. 2001. Duplexes of 21-nucleotide RNAs mediate RNA interference incultured mammalian cells. Nature 411:494–498.
10. Felber, B. K., H. Paskalis, C. Kleinman-Ewing, F. Wong-Staal, and G. N.Pavlakis. 1985. The pX protein of HTLV-1 is a transcriptional activator of itslong terminal repeats. Science 229:675–679.
11. Fong, Y. W., and Q. Zhou. 2001. Stimulatory effect of splicing factors ontranscriptional elongation. Nature 414:929–933.
12. Fu, X.-D. 1995. The superfamily of arginine/serine-rich splicing factors. RNA1:663–680.
13. Fujii, M., J. Tsuchiya, T. Chuhjo, T. Akizawa, and M. Seiki. 1992. Interactionof HTLV-1 Tax1 with p67SRF causes the aberrant induction of cellularimmediate early genes through CArG boxes. Genes Dev. 6:2066–2076.
14. Fujii, M., P. Sassone-Corsi, and I. M. Verma. 1988. c-fos promoter transac-tivation by the tax1 protein of human T-cell leukemia virus type I. Proc. Natl.Acad. Sci. USA 85:8526–8530.
15. Fujisawa, J., M. Seiki, M. Sato, and M. Yoshida. 1986. A transcriptionalenhancer sequence of HTLV-1 is responsible for transactivation mediated byp40 chi HTLV-1. EMBO J. 5:713–718.
16. Ge, H., Y. Si, and A. P. Wolffe. 1998. A novel transcriptional coactivator, p52,functionally interacts with the essential splicing factor ASF/SF2. Mol. Cell2:751–759.
17. Giebler, H. A., J. E. Loring, K. van Orden, M. A. Colgin, J. E. Garrus, K. W.Escudero, A. Brauweiler, and J. K. Nyborg. 1997. Anchoring of CREBbinding protein to the human T-cell leukemia virus type 1 promoter: amolecular mechanism of Tax transactivation. Mol. Cell. Biol. 17:5156–5164.
18. Goren, I., O. J. Semmes, K.-T. Jeang, and K. Moelling. 1995. The aminoterminus of Tax is required for interaction with the cyclic AMP responseelement binding protein. J. Virol. 69:5806–5811.
19. Haller, K., Y. Wu, E. Derow, I. Schmitt, K.-T. Jeang, and R. Grassmann.2002 Physical interaction of human T-cell leukemia virus type 1 Tax withcyclin-dependant kinase 4 stimulates the phosphorylation of retinoblastomaprotein. Mol. Cell. Biol. 22:3327–3338.
20. Hanakahi, L. A., L. A. Dempsey, M.-J. Li, and N. Maizels. 1997. Nucleolinis one component of the B cell-specific transcription factor and switch regionbinding protein, LR1. Proc. Natl. Acad. Sci. USA 94:3605–3610.
21. Iwanaga, R., K. Ohtani, T. Hayashi, and M. Nakamura. 2001. Molecularmechanism of cell cycle progression induced by the oncogene product Tax ofhuman T-cell leukemia virus type I. Oncogene 20:2055–2067.
22. Jeang, K.-T. 2001. Functional activities of the human T-cell leukemia virustype I Tax oncoprotein: cellular signaling through NF-�B. Cytokine GrowthFactor Rev. 12:207–217.
23. Jiang, H., H. Lu, R. L. Schiltz, C. A. Pise-Masison, V. V. Ogryzko, Y.Nakatani, and J. N. Brady. 1999. PCAF interacts with Tax and stimulatesTax transactivation in a histone acetyltransferase-independent manner. Mol.Cell. Biol. 19:8136–8145.
24. Kimzey, A. L., and W. S. Dynan. 1998. Specific regions of contact betweenhuman T-cell leukemia virus type I tax protein and DNA identified byphotocross-linking. J. Biol. Chem. 273:13768–13775.
25. Kwok, R. P. S., J. R. Lundblad, J. C. Chrivia, J. P. Richards, H. P. Bach-inger, R. G. Brennan, S. G. E. Roberts, M. R. Green, and R. H. Goodman.1994. Nuclear protein CBP is a coactivator for the transcription factorCREB. Nature 370:223–226.
26. Kwok, R. P. S., M. E. Laurance, J. R. Lundblad, P. S. Goldman, H. Shih,L. M. Connor, S. J. Marriott, and R. H. Goodman. 1996. Control of cAMP-regulated enhancers by the viral transactivator Tax through CREB and thecoactivator CBP. Nature 380:642–646.
27. Lenzmeire, B. A., H. A. Giebler, and J. K. Nyborg. 1998. Human T-cellleukemia virus type 1 Tax requires direct access to DNA for recruitment ofCREB binding protein to the viral promoter. Mol. Cell. Biol. 18:721–731.
28. Lenzmeire, B. A., E. E. Baird, P. B. Dervan, and J. K. Nyborg. 1999. The Taxprotein-DNA interaction is essential for HTLV-1 transactivation in vitro. J.Mol. Biol. 291:731–744.
29. Leung, K., and G. J. Nabel. 1988. HTLV-1 transactivator induces interleu-kin-2 receptor expression through an NF-�B-like factor. Nature 333:776–778.
30. Longman, D., T. McGarvey, S. McCracken, I. L. Johnstone, B. J. Blencowe,and J. F. Caceres. 2001. Multiple interactions between SRm160 and SRfamily proteins in enhancer-dependent splicing and development of C. el-egans. Curr. Biol. 11:1923–1933.
31. Lundblad, J. R., P. S. Kwork, M. E. Laurance, M. S. Huang, J. P. Richards,R. G. Brennan, and R. H. Goodman. 1998. The human T-cell leukemiavirus-1 transcriptional activator Tax enhances cAMP-responsive element-binding protein (CREB) binding activity through interactions with the DNAminor groove. J. Biol. Chem. 273:19251–19259.
32. Maniatis, T., and R. Reed. 2002. An extensive network of coupling amonggene expression machines. Nature 416:499–506.
33. Maruyama, M., H. Shibuya, H. Harada, M. Hatakeyama, M. Seiki, T. Fujita,J. Inoue, M. Yoshida, and T. Taniguchi. 1987. Evidence for aberrant acti-vation of the interleukin-2 autocrine loop by HTLV-1-encoded p40X andT3/Ti complex triggering. Cell 48:343–350.
34. Michelotti, E. F., G. A. Michelotti, A. I. Aronsohn, and D. Levens. 1996.Heterogeneous nuclear ribonucleoprotein K is a transcription factor. Mol.Cell. Biol. 16:2350–2360.
35. Misteli, T., J. F. Caceres, and D. L. Spector. 1997. The dynamics of apre-mRNA splicing factor in living cells. Nature 387:523–527.
36. Misteli, T., J. F. Caceres, J. Q. Clement, A. R. Krainer, M. F. Wilkinson, andD. L. Spector. 1998. Serine phosphorylation of SR proteins is required fortheir recruitment to site of transcription in vivo. J. Cell Biol. 143:297–307.
37. Monsalve, M., Z. Wu, G. Adelmant, P. Puigserver, M. Fan, and B. M.Spiegelman. 2000. Direct coupling of transcription and mRNA processingthrough the thermogenic coactivator PGC-1. Mol. Cell 6:307–316.
38. Nyunoya, H., T. Morita, T. Sato, S. Honma, A. Tsujimoto, and K. Shimo-tohno. 1993. Cloning of a cDNA encoding a DNA-binding proteinTAXREB302 that is specific for the tax-responsive enhancer of HTLV-1.Gene 126:251–255.
39. Nyunoya, H., T. Morita, T. Sato, S. Honma, A. Tsujimoto, and K. Shimo-tohno. 1994. Corrigendum. Cloning of a cDNA encoding a DNA-bindingprotein TAXREB302 that is specific for the tax-responsive enhancer ofHTLV-1. Gene 148:371–373.
40. Paskalis, H., B. K. Felber, and G. N. Pavlakis. 1986. Cis-acting sequencesresponsible for the transcriptional activation of human T-cell leukemia virustype I constitute a conditional enhancer. Proc. Natl. Acad. Sci. USA 83:6558–6562.
10026 YOUN ET AL. J. VIROL.
on February 14, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

41. Patturajan, M., X. Wei, R. Berezney, and J. L. Corden. 1998. A nuclearmatrix protein interacts with the phosphorylated C-terminal domain of RNApolymerase II. Mol. Cell. Biol. 18:2406–2415.
42. Reddy, T. R., X. Li, Y. Jones, M. H. Ellisman, G. Y. Ching, R. K. H. Liem,and F. Wong-Staal. 1998. Specific interaction of HTLV tax protein and ahuman type IV neuronal intermediate filament protein. Proc. Natl. Acad.Sci. USA 95:702–707.
43. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecular cloning: alaboratory manual, 2nd ed. Cold Spring Harbor Laboratory, Cold SpringHarbor, N.Y.
44. Sawada, Y., Y. Miura, K. Umeki, T. Tamaoki, K. Fujinaga, and S. Ohtaki.2000. Cloning and characterization of a novel RNA-binding protein SRL300with RS domains. Biochim. Biophys. Acta 1492:191–195.
45. Seiki, M., J. Inoue, T. Takeda, and M. Yoshida. 1986. Direct evidence thatp40X of human T-cell leukemia virus type I is a trans-acting transcriptionalactivator. EMBO J. 5:561–565.
46. Semmes, O. J., and K.-T. Jeang. 1996. Localization of human T-cell leuke-mia virus type I Tax to subnuclear compartments that overlap with inter-chromatin speckles. J. Virol. 70:6347–6357.
47. Shimotohno, K., M. Takano, T. Teruuchi, and M. Miwa. 1986. Requirementof multiple copies of a 21-nucleotide sequence in the U3 regions of humanT-cell leukemia virus type I and type II long terminal repeats for transactingactivation of transcription. Proc. Natl. Acad. Sci. USA 83:8112–8116.
48. Shuh, M., and D. Derse. 2000. Ternary complex factors and cofactors areessential for human T-cell leukemia virus type 1 Tax transactivation of theserum response element. J. Virol. 74:11394–11397.
49. Suzuki, T., J. Fujisawa, M. Toita, and M. Yoshida. 1993. The transactivatorTax of human T-cell leukemia virus type 1 (HTLV-1) interacts with cAMP-responsive element (CRE) binding and CRE modulator proteins that bind tothe 21-base-pair enhancer of HTLV-1. Proc. Natl. Acad. Sci. USA 90:610–614.
50. Suzuki, T., H. Hirai, and M. Yoshida. 1994. Tax protein of HTLV-1 interactswith the Rel homology domain of NF-�B p65 and c-Rel proteins bound tothe NF-�B binding site and activates transcription. Oncogene 9:3099–3105.
51. Suzuki, T., S. Kitao, H. Matsushima, and M. Yoshida. 1996. HTLV-1 Taxprotein interacts with cyclin-dependant kinase inhibitor p16INK4A andcounteracts its inhibitory activity towards CDK4. EMBO J. 15:1607–1614.
52. Suzuki, T., Y. Ohsugi, M. Uchida-Toita, T. Akiyama, and M. Yoshida. 1999.Tax oncoprotein of HTLV-1 binds to the human homologue of Drosophiladiscs large tumor suppressor protein, hDLG, and perturbs its function in cellgrowth control. Oncogene 18:5967–5972.
53. Tsujimoto, A., H. Nyunoya, T. Morita, T. Sato, and K. Shimotohno. 1991.Isolation of cDNAs for DNA-binding proteins which specifically bind to atax-responsive enhancer element in the long terminal repeat of human T-cellleukemia virus type I. J. Virol. 65:1420–1426.
54. Uchiyama, T. 1997. Human T cell leukemia virus type I (HTLV-I) andhuman diseases. Annu. Rev. Immunol. 15:15–37.
55. Vinson, C. R., K. L. LaMarco, P. F. Johnson, W. H. Landschulz, and S. L.McKnight. 1988. In situ detection of sequence-specific DNA binding activityspecified by a recombinant bacteriophage. Genes Dev. 2:801–806.
56. Yin, M. J., and R. B. Gaynor. 1996. Complex formation between CREB andTax enhances the binding affinity of CREB for the human T-cell leukemiavirus type 1 21-base-pair repeats. Mol. Cell. Biol. 16:3156–3168.
57. Yin, M. J., and R. B. Gaynor. 1996. HTLV-1 21 bp repeat sequences facil-itate stable association between Tax and CREB to increase CREB bindingaffinity. J. Mol. Biol. 264:20–31.
58. Zhao, L. J., and C. Z. Giam. 1992. Human T-cell lymphotropic virus type I(HTLV-1) transcriptional activator, Tax, enhances CREB binding toHTLV-1 21-base-pair repeats by protein-protein interaction. Proc. Natl.Acad. Sci. USA 89:7070–7074.
VOL. 77, 2003 SR-RELATED PROTEIN NEEDED FOR Tax TRANSACTIVATION 10027
on February 14, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from





























