Embed Size (px)
Citation preview

Spiral-Wave Instability in a Medium with a Gradient in the Fibroblast Density:a Computational Study
Soling Zimik1, Rahul Pandit1
1 Centre for Condensed Matter Theory, Department of Physics, Indian Institute of Science,Bangalore, 560012, India
Abstract
Fibrosis, a process of fibroblast proliferation in cardiactissue, is a major concern for patients with diseases like is-chemia, heart failure, and cardiomyopathy, because of itsarrhythmogenic effects. Fibroblasts, in appreciable den-sities, are known to affect the electrical-wave dynamics incardiac tissue, because the coupling of fibroblasts with my-ocytes modulates the electrophysiological properties of themyocytes. Furthermore, in fibrotic hearts, the distributionof fibroblasts can be heterogeneous, so the density of fi-broblasts can vary from wounded regions (like infarcted orischemic zones) to normal regions of the heart. Such gra-dients in the fibroblast density (GFD) induce spatial varia-tion in the electrophysiological properties of the tissue,andthe latter can initiate and affect the dynamics of patholog-ical waves like spiral waves. We study the effects of GFDson the dynamics of spiral waves by using a state-of-the-art mathematical model for human-ventricular tissue. Wefind that, in the presence of GFDs, spiral waves are un-stable, i.e., a stable spiral wave breaks into multiple spiralwaves. We find that GFDs induce spatial variations in thelocal spiral-wave frequencyω in the medium. Such a vari-ation in ω leads to anisotropic thinning of the spiral armthat gives way to spiral-wave breaks. We also study thefactors that enhance the instability of the spiral waves inthe medium with GFD. Finally, we show that the presenceof GFD can spontaneously lead to spiral waves, via high-frequency pacing, in the medium.
1. Introduction
The occurrence of abnormal spatiotemporal patterns ofelectrical waves in cardiac tissue, in the form of spiralwaves, has been associated with cardiac arrhythmias [1].The existence of a single spiral wave is associated withtachycardia [2], and a multiple-spiral-wave state is asso-ciated with a life-threatening arrhythmia known as fibril-lation [3]. A single-spiral-wave state can transition intoa multiple-spiral-wave state if the spiral becomes unsta-
ble and breaks up, giving rise to many daughter spirals [1].Given that fibrillation is a more lethal condition than tachy-cardia, it is important to understand the mechanisms of thetransition process from a single- to a multiple-spiral-wavestate. Here, we present a mechanism of spiral-wave in-stability in a medium with a gradient in fibroblast density(GFD).
Fibroblasts are passive cells that are needed for theproper functioning of a heart, because they, along withother cells, form the extracellular matrix and ensure thestructural integrity of the heart. However, the abnormalproliferation of fibroblasts because of diseases like is-chemia, heart failure, and cardiomyopathy is consideredto be arrhythmogenic [4]. Fibroblasts, if they form gap-junctional coupling, can change the electrophysiology ofmyocytes in cardiac tissue [5]. This, in turn, can affect thedynamics of electrical waves in cardiac tissue. Many stud-ies have been performed to investigate the effects of fibrob-lasts on wave dynamics in cardiac tissue [6, 7]; however,most of the studies, which consider fibroblast-myocyte(FM) coupling, deal with a homogeneous distribution offibroblasts in the domain. But, the density of fibroblasts ina diseased cardiac tissue may not necessarily be homoge-neous [8]. Therefore, it is important to study the effects ofFM coupling on wave dynamics in a domain with hetero-geneous distributions of fibroblast density.
We investigate the effects of GFD on spiral-wave dy-namics in a mathematical model for human-ventricular tis-sue. We find that GFD induces a spatial variation of the lo-cal spiral-wave frequencyω in the domain. This variationof ω in the domain leads to spiral-wave instability, and thedegree of instability depends on the magnitude of the vari-ation ofω. We also investigate the factors that modulatethe variation ofω in the domain. We find that, for a givenGFD, the resting membrane potentialEf of the fibroblasts,and the number of fibroblastsNf that are coupled to a my-ocyte can change the spatial variation ofω in the domain.Finally, we also show that GFD can spontaneously initiatespiral waves via high-frequency pacing.
Computing in Cardiology 2017; VOL 44 Page 1 ISSN: 2325-887X DOI:10.22489/CinC.2017.034-043
2. Methods And Materials
We use the O’Hara-Rudy dynamic model [9] for ourhuman-ventricular myocyte cell. The fibroblasts are mod-elled as passive cells; for these fibroblasts, we use themodel given by MacCannell,et al. [10].
In our two-dimensional (2D) simulations the fibroblastsare attached atop the myocytes; thus, our 2D simulationdomain is a bilayer. The spatiotemporal evolution of themembrane potential (Vm) of the myocytes in tissue is gov-erned by a reaction-diffusion equation, which is the fol-lowing partial-differential equation (PDE):
∂Vm
∂t+
Iion + IgapCm
= D∇2Vm, (1)
whereD is the diffusion constant between the myocytes.Iion is the sum of all the ionic currents of the myocyte [9].Igap is the gap-junctional current between a myocyte anda fibroblast in an FM composite.Igap=0 if no fibroblast isattached to a myocyte.
We use the forward-Euler method for time marchingwith a five-point stencil for the Laplacian. We set D=0.0012cm2/ms. The temporal and spatial resolutions areset to beδx= 0.02 cm andδt=0.02 ms, respectively. Weuse a domain size of960× 960 grid points. We initiatethe spiral wave by using the conventional S1-S2 cross-fieldprotocol.
3. Results
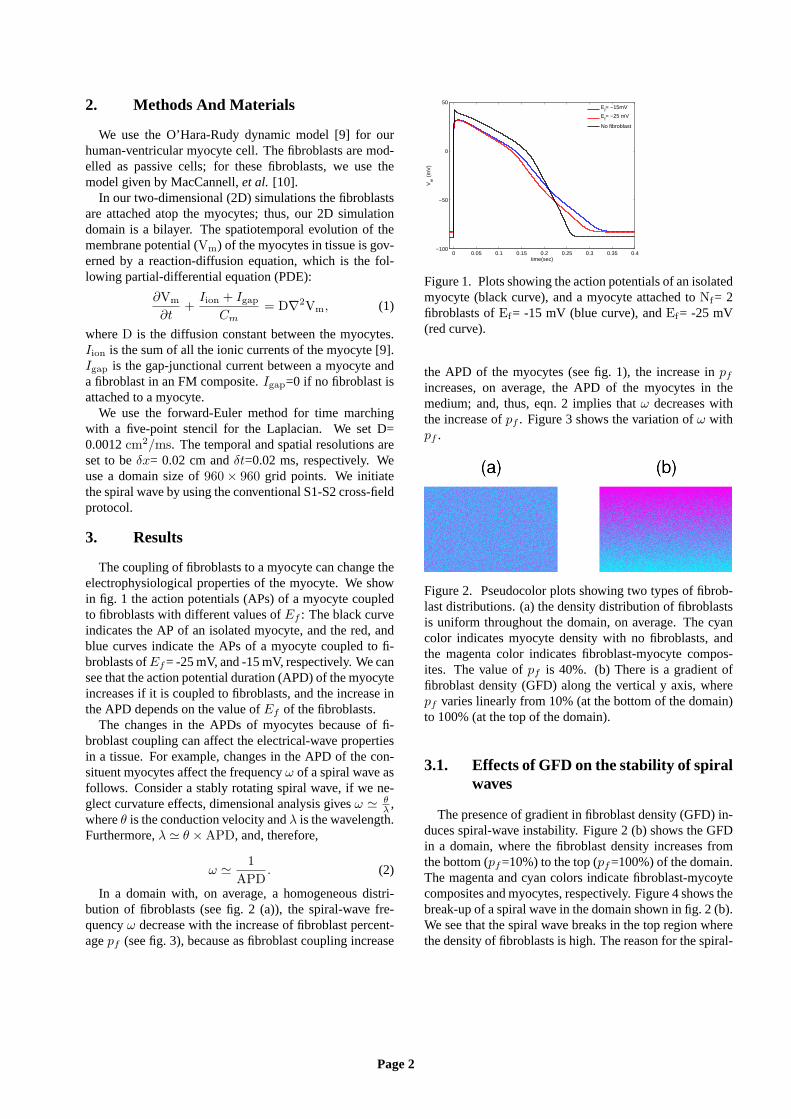
The coupling of fibroblasts to a myocyte can change theelectrophysiological properties of the myocyte. We showin fig. 1 the action potentials (APs) of a myocyte coupledto fibroblasts with different values ofEf : The black curveindicates the AP of an isolated myocyte, and the red, andblue curves indicate the APs of a myocyte coupled to fi-broblasts ofEf= -25 mV, and -15 mV, respectively. We cansee that the action potential duration (APD) of the myocyteincreases if it is coupled to fibroblasts, and the increase inthe APD depends on the value ofEf of the fibroblasts.
The changes in the APDs of myocytes because of fi-broblast coupling can affect the electrical-wave propertiesin a tissue. For example, changes in the APD of the con-situent myocytes affect the frequencyω of a spiral wave asfollows. Consider a stably rotating spiral wave, if we ne-glect curvature effects, dimensional analysis givesω ≃
θλ
,whereθ is the conduction velocity andλ is the wavelength.Furthermore,λ ≃ θ ×APD, and, therefore,
ω ≃1
APD. (2)
In a domain with, on average, a homogeneous distri-bution of fibroblasts (see fig. 2 (a)), the spiral-wave fre-quencyω decrease with the increase of fibroblast percent-agepf (see fig. 3), because as fibroblast coupling increase
0 0.05 0.1 0.15 0.2 0.25 0.3 0.35 0.4−100
−50
0
50
Vm
(m
V)
time(sec)
E
f= −15mV
Ef= −25 mV
No fibroblast
Figure 1. Plots showing the action potentials of an isolatedmyocyte (black curve), and a myocyte attached toNf= 2fibroblasts ofEf= -15 mV (blue curve), andEf= -25 mV(red curve).
the APD of the myocytes (see fig. 1), the increase inpfincreases, on average, the APD of the myocytes in themedium; and, thus, eqn. 2 implies thatω decreases withthe increase ofpf . Figure 3 shows the variation ofω withpf .
Figure 2. Pseudocolor plots showing two types of fibrob-last distributions. (a) the density distribution of fibroblastsis uniform throughout the domain, on average. The cyancolor indicates myocyte density with no fibroblasts, andthe magenta color indicates fibroblast-myocyte compos-ites. The value ofpf is 40%. (b) There is a gradient offibroblast density (GFD) along the vertical y axis, wherepf varies linearly from 10% (at the bottom of the domain)to 100% (at the top of the domain).
3.1. Effects of GFD on the stability of spiralwaves
The presence of gradient in fibroblast density (GFD) in-duces spiral-wave instability. Figure 2 (b) shows the GFDin a domain, where the fibroblast density increases fromthe bottom (pf=10%) to the top (pf=100%) of the domain.The magenta and cyan colors indicate fibroblast-mycoytecomposites and myocytes, respectively. Figure 4 shows thebreak-up of a spiral wave in the domain shown in fig. 2 (b).We see that the spiral wave breaks in the top region wherethe density of fibroblasts is high. The reason for the spiral-
Page 2

0 50 100
3.5
4
4.5
5
5.5
pf(%)
ω (
Hz)
(a)
E
f= −15mV
Ef= −25mV
0 50 1001
2
3
4
5
pf(%)
ω (
Hz)
(b)
Nf=1
Nf= 2
Nf= 3
Figure 3. Variation of the spiral-wave frequencyω withthe percentage of fibroblastspf , in a domain with a homo-geneous fibroblast distribution. (a) Plots showing the vari-ation ofω with pf for Ef= -15 mV (black curve), andEf=-25 mV (blue curve). (b) Plots showing the variation ofωwith pf for three different values ofNf=1 (black curve),Nf=2 (blue curve), andNf=3 (red curve).
wave instability is as follows. The presence of GFD in themedium induces a spatial variation of the local value ofω(refer fig. 3). In fig. 4 the local value ofω decreases fromthe bottom to the top of the domain. This anisotropic varia-tion ofω leads to an anisotropic thinning of the wavelengthof the spiral arms (see fig. 4 2.14 s), which gives way forspiral-wave break-up. The wave-thinning occurs in the up-per region, because it has large APD. As the upper regionhas a large APD, the conduction velocity is lower in that re-gion (conduction-velocty restitution property [11]); there-fore, in order to support the high-frequency wave trainscoming from the lower region, the wavelength of the wavesthins as it propagates towards the top region.
Figure 4. Pseudocolor plot ofVm showing the spatiotem-poral evolution of the break-up of a spiral in the presenceof GFD withpf going from 10%-100%, andEf = -25 mV.The spiral arm breaks up in the upper region, which is thelow-ω region.
As the instability of the spiral waves arises because ofthe spatial variation ofω in the medium, the degree of in-stability of the spiral waves depends on the magnitude ofthe gradient inω that is induced by GFD. We, therefore,study the factors that modulate the variation ofω with pf .We find that high values ofEf induce high variation ofωwith pf . Figure 3 (a) shows that the variation ofω with pfis more drastic forEf= -15 mV (black), as compared toEf= -25 mV (blue). Moreover, we also find that the varia-tion ofω with pf is enhanced if we couple more fibroblasts
numberNf to a myocyte. Figure 3 (b) shows the variationof ω with pf for three different values ofNf : Nf=1(black),Nf=2(blue), andNf=3(red). We see that variation ofω in-creases as we go fromNf=1 toNf=3. These results tellsus that, for a given GFD, a spiral wave is more vulnerableto break-up for high values ofEf andNf as compared tothe low values ofEf andNf .
3.2. Spontaneous initiation of spiral waves
The presence of GFD in the medium can also initiatespiral wave spontaneously if we pace the medium at a highfrequency. Figure 5 shows the initiation of spiral waves ina medium with GFD withpf going from 10%-100%. Weapply periodic line stimuli at the bottom edge of the do-main at a pacing cycle length PCL=250 ms. The reasonfor the spiral-wave initiation is as follows. The top regionof the medium, because it has a large APD, repolarizesslowly. Now, if we pace the medium, at high-frequency,there is a prominent waveback-wavefront interactions ofthe waves in the top region. This waveback-wavfront inter-actions induce corrugations in the wavefronts of the waves(because of the random distribution of fibroblasts), whicheventually leads to the initiation of spiral waves.
Figure 5. Pseudocolor plot ofVm showing the the sponta-neous initiation of a spiral wave, via high-frequency pac-ing, in a medium with GFD as shown in fig. 2 (b). Thepacing stimuli are applied at the bottom of the domain atPCL= 250 ms.
Such pacing-induced spiral waves occur at high-frequency pacing and not at low-frequency pacing, be-cause the wavefront-waveback interaction between theconsecutive waves is more prominent at high-frequencypacing. Figure 6 shows a stability diagram in theEF -PCLplane indicating the regions where spiral waves (magentacolor) occur and no spiral waves (black color) occur.
In conclusion, we have shown how fibroblast-myocyte(FM) coupling can affect the the electrical-wave propertiesin a tissue. We see that FM coupling changed the APD ofthe myocytes, and this, in turn, modulated the propertiesof the spiral waves in a tissue, like its frequencyω. Weshow thatω decreases with the increase of the percentageof fibroblastspf , because the FM coupling increases theAPD of the constituent myocytes. We then study how thegradient in the fibroblast density (GFD) affects the dynam-
Page 3

−20 −15 −10 −5200
220
240
260
280
300
320
No spiral waves
Spiral waves
Ef (mV)
PC
L (m
s)
Figure 6. Stability diagram in theEf -PCL plane. Figureshowing the regions where we observe spiral waves (blackcolor) and no spiral waves (magenta color) in theEf -PCLplane.
ics of the spiral waves. We find that GFD induces spiral-wave instability in the medium. We show that this insta-bility arises because of the spatial variation of the localvalue ofω in the medium induced by the GFD, and thespiral wave breaks in the low-ω region. Our finding ofspiral-wave break in the low-ω is consistent with the ex-perimental results of Campbell,et al., in monolayers ofneonatal-rat ventricular myocytes, where the gradeint inωis induced by varying theIKr ion-channel density. We alsoinvestigate the factors that affect the variation ofω withpf . We find that the variation ofω with pf is more for highvalues ofEf andNf as compared to when their values arelow. And thus, for a given GFD, the degree of spiral-waveinstability is higher at high values ofEf andNf as com-pared to their low values. Finally, we show how GFD in themedium can initiate spiral waves spontaneously via high-frequency pacing. We hope our results will lead to detailedstudies of GFD-induced spiral-wave instability at least inin vitro experiments on cell-cultures. At the simplest level,we suggest fibroblast analogs of the experiments of Camp-bell, etal. [12].
Acknowledgements
We thank the Department of Science and Technology(DST), India, and the Council for Scientific and IndustrialResearch (CSIR), India, for financial support, and the Su-percomputer Education and Research Centre (SERC, IISc)for computational resources.
References
[1] Fenton FH, Cherry EM, Hastings HM, Evans SJ. Multiplemechanisms of spiral wave breakup in a model of cardiacelectrical activity. Chaos An Interdisciplinary Journal ofNonlinear Science 2002;12(3):852–892.
[2] Efimov IR, Sidorov V, Cheng Y, Wollenzier B. Evidence of
three-dimensional scroll waves with ribbon-shaped filamentas a mechanism of ventricular tachycardia in the isolatedrabbit heart. Journal of cardiovascular electrophysiology1999;10(11):1452–1462.
[3] Bayly P, KenKnight B, Rogers J, Johnson E, Ideker R,Smith W. Spatial organization, predictability, and determin-ism in ventricular fibrillation. Chaos An InterdisciplinaryJournal of Nonlinear Science 1998;8(1):103–115.
[4] Manabe I, Shindo T, Nagai R. Gene expression in fibrob-lasts and fibrosis involvement in cardiac hypertrophy. Cir-culation research 2002;91(12):1103–1113.
[5] Nguyen TP, Xie Y, Garfinkel A, Qu Z, Weiss JN. Arrhyth-mogenic consequences of myofibroblast–myocyte cou-pling. Cardiovascular research 2012;93(2):242–251.
[6] Majumder R, Nayak AR, Pandit R. Nonequilibrium ar-rhythmic states and transitions in a mathematical model fordiffuse fibrosis in human cardiac tissue. PLoS one 2012;7(10):e45040.
[7] Greisas A, Zlochiver S. Modulation of spiral-wave dynam-ics and spontaneous activity in a fibroblast/myocyte hetero-cellular tissue—a computational study. IEEE Transactionson Biomedical Engineering 2012;59(5):1398–1407.
[8] Ashikaga H, Arevalo H, Vadakkumpadan F, Blake RC,Bayer JD, Nazarian S, Zviman MM, Tandri H, Berger RD,Calkins H, et al. Feasibility of image-based simulation toestimate ablation target in human ventricular arrhythmia.Heart Rhythm 2013;10(8):1109–1116.
[9] O’Hara T, Virag L, Varro A, Rudy Y. Simulation ofthe undiseased human cardiac ventricular action poten-tial: model formulation and experimental validation. PLoSComput Biol 2011;7(5):e1002061.
[10] MacCannell KA, Bazzazi H, Chilton L, Shibukawa Y, ClarkRB, Giles WR. A mathematical model of electrotonic inter-actions between ventricular myocytes and fibroblasts. Bio-physical journal 2007;92(11):4121–4132.
[11] Cherry EM, Fenton FH. Suppression of alternans and con-duction blocks despite steep apd restitution: electrotonic,memory, and conduction velocity restitution effects. Amer-ican Journal of Physiology Heart and Circulatory Physiol-ogy 2004;286(6):H2332–H2341.
[12] Campbell K, Calvo CJ, Mironov S, Herron T, Beren-feld O, Jalife J. Spatial gradients in action potential du-ration created by regional magnetofection of herg are asubstrate for wavebreak and turbulent propagation in car-diomyocyte monolayers. The Journal of physiology 2012;590(24):6363–6379.
Address for correspondence:
Soling ZimikCentre for Condensed Matter Theory, Department of Physics, In-dian Institute of Science, Bangalore, 560012, [email protected]
Page 4





























