Embed Size (px)
Citation preview

Publications of the University of Eastern Finland
Dissertations in Health Sciences
isbn 978-952-61-1320-3
Publications of the University of Eastern FinlandDissertations in Health Sciences
Polyamines are ubiquitious
molecules essential for cell
proliferation. However, the
role of polyamine metabolism
in the immune response,
hematopoiesis, and bone
remodeling is largely unknown
and was thus addressed in this
study. The study showed that
enhanced activity of spermidine/
spermine N1-acetyltransferase
(SSAT), a catabolic regulator of
polyamines, was associated with
disturbed hematopoiesis and bone
remodeling in mice and myeloid
leukemias in humans.
dissertatio
ns | 207 | S
ini P
irn
es-Ka
rh
u | Sperm
idine/spermine N
1-acetyltransferase in Mouse H
ematopoiesis and B
one Rem
odeling...
Sini Pirnes-KarhuSpermidine/spermine
N1-acetyltransferase in Mouse Hematopoiesis
and Bone Remodeling and in Human Leukemias
Sini Pirnes-Karhu
Spermidine/spermine N1-acetyltransferase in Mouse Hematopoiesis and Bone Remodeling and in Human Leukemias

SINI PIRNES-KARHU
Spermidine/spermine N1-acetyltransferase
in Mouse Hematopoiesis and Bone
Remodeling and in Human Leukemias
To be presented by permission of the Faculty of Health Sciences, University of Eastern Finland for
public examination in Tietoteknia Auditorium (TTA), Kuopio,
on Saturday, December 14th 2013, at 1 pm
Publications of the University of Eastern Finland
Dissertations in Health Sciences
Number 207
Department of Biotechnology and Molecular Medicine
A.I.Virtanen Institute for Molecular Sciences
Faculty of Health Sciences
University of Eastern Finland
Tampere
2013

Juvenes Print – Suomen Yliopistopaino Oy
Tampere, 2013
Series Editors:
Professor Veli-Matti Kosma, M.D., Ph.D.
Institute of Clinical Medicine, Pathology
Faculty of Health Sciences
Professor Hannele Turunen, Ph.D.
Department of Nursing Science
Faculty of Health Sciences
Professor Olli Gröhn, Ph.D.
A.I. Virtanen Institute for Molecular Sciences
Faculty of Health Sciences
Professor Kai Kaarniranta, M.D., Ph.D.
Institute of Clinical Medicine, Ophthalmology
Faculty of Health Sciences
Lecturer Veli-Pekka Ranta, Ph.D. (pharmacy)
School of Pharmacy
Faculty of Health Sciences
Distributor:
University of Eastern Finland
Kuopio Campus Library
P.O.Box 1627
FI-70211 Kuopio, Finland
http://www.uef.fi/kirjasto
ISBN 978-952-61-1320-3 (print)
ISBN 978-952-61-1321-0 (pdf)
ISSN 1798-5706 (print)
ISSN 1798-5714 (pdf)
ISSN-L 1798-5706

III
Author’s address: Biotechnology and Molecular Medicine
A.I.Virtanen Institute
University of Eastern Finland
KUOPIO
FINLAND
e-mail: [email protected]
Supervisors: Professor Leena Alhonen, Ph.D.
A.I.Virtanen Istitute
University of Eastern Finland
KUOPIO
FINLAND
Anne Uimari, Ph.D.
A.I.Virtanen Institute
University of Eastern Finland
KUOPIO
FINLAND
Reviewers: Docent Jarmo Wahlfors, Ph.D.
Research Council for Health
Academy of Finland
HELSINKI
FINLAND
Docent Juhani Sand, Ph.D.
Division 2
Tampere University Hospital
TAMPERE
FINLAND
Opponent: Professor Leif C. Andersson, MD, Ph.D.
Haartman institute
University of Helsinki
HELSINKI
FINLAND

IV

V
Pirnes-Karhu, Sini. Spermidine/spermine N1-acetyltransferase in Mouse Hematopoiesis and Bone Remodeling and in
Human Leukemias. University of Eastern Finland, Faculty of Health Sciences
Publications of the University of Eastern Finland. Dissertations in Health Sciences 207. 2013. 56 p.
ISBN 978-952-61-1320-3 (print)
ISBN 978-952-61-1321-0 (pdf)
ISSN 1798-5706 (print)
ISSN 1798-5714 (pdf)
ISSN-L 1798-5706
ABSTRACT
Polyamines are ubiquitous molecules essential for cell proliferation. Induction of the
catabolic enzyme of polyamines, spermidine/spermine N1-acetyltransferase (SSAT), is
successfully used to limit cancer cell growth in cell culture. In animal models, however,
SSAT overexpression has been shown to increase susceptibility to cancer. In the present
study, the role of enhanced polyamine catabolism in homeostasis of hematopoietic and
bone systems were investigated in mice overexpressing SSAT (SSAT mice) and its
association to hematopoietic malignancies was studied with patient samples.
The basal levels of serum and hepatic inflammatory markers of SSAT mice were
comparable to those encountered in wild-type littermates. When exposed to endotoxin
SSAT mice displayed a slightly enhanced anti-inflammatory response but the survival and
symptoms of the SSAT mice were similar to those of wild-type mice. Further studies of cells
and tissues of the immune system revealed that the hematopoietic phenotype of SSAT mice
fulfilled the criteria of mouse myeloproliferative disease.
The myeloproliferation of SSAT mice resulted from both the intrinsic SSAT
overexpression of hematopoietic cells and microenvironmental factors. Characterization of
the bone phenotype of SSAT mice showed altered structure of bones of SSAT mice to
provide larger niche for hematopoietic cells. SSAT mice also showed cell-autonomously
impaired osteoblastogenesis and increased number of premature osteoblasts.
The association between SSAT activity and myeloproliferation was supported by
findings from human leukemia patients. The SSAT activity in peripheral blood
mononuclear cells of these patients was associated with the leukocyte number in myeloid
leukemias but not in lymphoid leukemia. Further investigation of SSAT mice provided
evidence that the association between SSAT and myeloproliferation might be mediated
through epigenetic factors.
Our results show that enhanced SSAT activity leads to aberrant bone formation and
subsequent changes in bone marrow microenvironment. In conjunction with enhanced
intrinsic SSAT activity of hematopoietic cells, this results in a myeloproliferative disease in
SSAT mice. The role of SSAT activity in myelopoiesis is further supported by the
association between SSAT activity and the white blood cell count in human myeloid
leukemias. National Library of Medicine Classification: QU 141, QU 475, WE 200, WH 140, WH 250, WH 380
Medical Subject Headings: Acetyltransferases/metabolism; Bone Marrow; Epigenesis, Genetic; Hematopoiesis;
Human; Leukemia, Myeloid; Mice; Myeloproliferative Disorders; Osteogenesis; Oxidative stress; Polyamines

VI

VII
Pirnes-Karhu Sini. Spermidiini/spermiini-N1-asetyylitransferaasi hiirten veren- ja luunmuodostuksessa sekä
ihmisten leukemioissa. Itä-Suomen yliopisto, Terveystieteiden tiedekunta
Publications of the University of Eastern Finland. Dissertations in Health Sciences 207. 2013. 56 s.
ISBN 978-952-61-1320-3 (print)
ISBN 978-952-61-1321-0 (pdf)
ISSN 1798-5706 (print)
ISSN 1798-5714 (pdf)
ISSN-L 1798-5706
TIIVISTELMÄ
Polyamiinit ovat solujen kasvulle välttämättömiä molekyylejä. Niitä hajottavan entsyymin,
spermidiini/spermiini-N1-asetyylitransferaasin (SSAT), indusointia on käytetty rajoittamaan
viljeltyjen syöpäsolujen kasvua. Eläinmalleissa puolestaan SSAT-geenin yli-ilmentäminen
on myös lisännyt alttiutta syövän synnylle. Tässä väitöskirjatutkimuksessa selvitettiin
kohonneen polyamiinikatabolian roolia veren- ja luunmuodostuksessa SSAT-geeniä yli-
ilmentävillä hiirillä (SSAT-hiiret) ja sen yhteyttä verisyöpiin potilasnäytteillä.
SSAT-hiirten seerumista ja maksasta mitattujen tulehdusmarkkereiden perustasot olivat
verrattavia villityypin hiirten markkereiden tasoon. SSAT-hiirten vaste endotoksiinille oli
kuitenkin lievästi tulehdusta hillitsevä. Tästä huolimatta eloonjääminen ja oireet olivat
verrattavia villityypin hiiren vastaaviin. Immunologisten solujen ja kudosten
lisätutkimukset osoittivat että SSAT-hiirten ilmiasu täytti hiirten myeloproliferatiivisen
sairauden kriteerit.
SSAT-hiirten myeloproliferatio syntyi yhdessä valkosolujen sisäisen SSAT:n yli-
ilmentämisen ja luuytimen mikroympäristön tekijöiden seurauksena. SSAT-hiirten luiden
lähempi tarkastelu osoitti niiden muuttuneen rakenteen muodostavan suuremman tilan
veren soluille. SSAT-hiirillä havaittiin lisäksi luuta muodostavien solujen (osteoblastien)
erilaistumisen olevan ympäristöstä riippumattomasti häiriintynyt ja epäkypsien
osteoblastien määrän kasvaneen.
SSAT-aktiivisuuden ja myeloproliferaation välillä havaittua yhteyttä vahvisti ihmisen
leukemianäytteiden tutkimus. Leukemiapotilaiden perifeerisen veren mononukleaaristen
valkosolujen SSAT-aktiivisuus oli yhteydessä valkosolumäärään myeloista mutta ei
lymfoista leukemiaa sairastavilla potilailla. Lisätutkimukset SSAT-hiirillä viittasivat
epigeneettisten tekijöiden olevan yhteydessä SSAT-geenin ja myeloproliferaation välillä
havaittuun suhteeseen.
Saamamme tulokset osoittavat kohonneen SSAT-aktiivisuuden johtavan epänormaaliin
luunmuodostukseen ja siitä johtuen poikkeavaan luuytimen mikroympäristöön. Yhdessä
veren solujen kohonneen sisäisen SSAT-aktiivisuuden kanssa tämä johtaa
myeloproliferatiivisen taudin kehittymiseen SSAT-hiirillä. SSAT-aktiivisuuden osuutta
myelopoieesissa tukee lisäksi havainto SSAT-aktiivisuuden yhteydestä valkosolumäärään
myeloista leukemiaa sairastavilla potilailla.
Yleinen Suomalainen asiasanasto: epigenetiikka; hiiret; ihmiset; immuunijärjestelmä; leukemia; luuydin;
luusto; polyamiinit; veri

VIII

IX
Acknowledgements
This work was carried out in the A.I.Virtanen Institute for Molecular Sciences, University of
Eastern Finland, during the years 2009‒2013.
I wish to express my sincere gratitude to my principal supervisor Professor Leena
Alhonen, PhD, who offered me an interesting project to work with and has shown belief in
my work ever since. My warm thanks belong also to my supervisor Anne Uimari, PhD, for
her inexhaustible guidance and all the stimulating discussions both in science and in
personal life. It has been great deal of fun to work with you!
I wish to thank Docent Jarmo Wahlfors, PhD, and Docent Juhani Sand, PhD, the official
reviewers of my thesis for their valuable comments to improve the manuscript. My thanks
belong also to Ewen Macdonald, PhD, and Sara Wojciechowski, B.Sc., who revised the
language of this manuscript.
I wish to express my gratitude to all our collaborators whose expertise in their own fields
has provided a strong foundation for my studies. I acknowledge Reijo Sironen, MD, PhD,
for his rigorous work with my first manuscript. I am very thankful for Docent Esa
Jantunen, MD, and Pentti Mäntymaa, MD, for their invaluable expertise in hematopoiesis
and hematopoietic malignancies as well as for Jorma Määttä, PhD, and Mikko Finnilä, MSc,
for their enthusiasm, work and advice with bone-related studies. I am grateful for Petri
Mäkinen, PhD, and Sara Wojciechowski for their kind and knowledgeable assistance with
flow cytometry. I wish to thank also Sohvi Hörkkö, MD, PhD, and Satu Mustjoki, MD, PhD,
for their scientific effort on my second and third manuscripts.
My warm thanks belong to all the present and former members of Leena’s laboratory
with whom I have had the pleasure to work. Thank you Tuomo Keinänen, PhD, Mervi
Hyvönen, PhD, Marko Pietilä, PhD, Taina Koponen, MSc, Marc Cerrada-Gimenez, PhD,
Eija Pirinen, PhD, Anita Lampinen, MSc, Tekele Fashe, MSc, Susanna Vuohelainen, MSc,
Maija Tusa, Lic.Phil., for helping me with my projects and making the working
environment so pleasant. I wish to direct my special thanks to the technical personnel who
participated in this study. Thank you, Anne Karppinen, Arja Korhonen, Marita Heikkinen,
Sisko Juutinen and Tuula Reponen, for your indefatigable and skillful help in the
laboratory and keeping the lab functional with all the practical essentials. In addition, I
would like to acknowledge Eeva Hakala, Jouko Mäkäräinen, Riitta Sinervirta, Helena Pernu
and all the personnel of National Laboratory Animal Center, especially Arja Konttinen,
Teija Oinonen and Virve Immonen, for efficiently managing the practical details.
My loving thanks belong to my family, especially to my husband Tero for making
everyday life so much fun. I also admire your calm response to stressful situations and I am
grateful for how you are able to make me calmer too. I am thankful for my parents, Maija
and Veijo, for encouraging me to pursue a quality education and follow my own path. I am
grateful for my beloved sisters Suvi, Säde and Sara and “brother” Teemu and my friends
both inside and outside of the scientific world, Ansku, Hanna-Maija and Eini, for sharing
the ups and downs of my work and personal life. I feel so privileged to have you all in my
life!
Lahti, November 2013

X

XI
List of the original publications
This dissertation is based on the following original publications:
I Pirnes-Karhu S, Sironen R, Alhonen L and Uimari A. Lipopolysaccharide-
induced anti-inflammatory acute phase response is enhanced in
spermidine/spermine N1-acetyltransferase (SSAT) overexpressing mice. Amino
Acids 42: 473-484, 2012.
II Pirnes-Karhu S, Mäntymaa P, Sironen R, Mäkinen PI, Wojciechowski S, Juutinen
S, Koistinaho J, Hörkkö S, Jantunen E, Alhonen L and Uimari A. Enhanced
polyamine catabolism disturbs hematopoietic lineage commitment and leads to a
myeloproliferative disease in mice overexpressing spermidine/spermine N1-
acetyltransferase. Amino Acids Epub Jul 9, 2013.
III Pirnes-Karhu S, Jantunen E, Mäntymaa P, Mustjoki S, Alhonen L and Uimari A.
Spermidine/spermine N1-acetyltransferase activity associates with white blood
cell count in myeloid leukemias. Manuscript.
IV Pirnes-Karhu S, Määttä J, Finnilä M, Alhonen L and Uimari A. Overexpression of
Spermidine/spermine N1-acetyltransferase Impairs Osteoblastogenesis and Alters
Mouse Bone Phenotype. Manuscript.
The following papers are referred to in the text by their Roman numeral.
The publications were adapted with the permission of the copyright owners.

XII

XIII
Contents
1 INTRODUCTION ...................................................................................................................... 1
2 REVIEW OF THE LITERATURE ............................................................................................. 3
2.1 POLYAMINE METABOLISM ............................................................................................................................ 3
2.1.1 Biosynthesis of polyamines............................................................................................................................. 4
2.1.1.1 Ornithine decarboxylase ......................................................................................................................... 4
2.1.1.2 S-adenosylmethionine decarboxylase................................................................................................... 5
2.1.1.3 Spermidine and spermine synthases .................................................................................................... 5
2.1.2 Polyamine catabolism ...................................................................................................................................... 5
2.1.2.1 Spermidine/spermine N1-acetyltransferase .......................................................................................... 6
2.1.2.2 Acetylpolyamine oxidase and spermine oxidase ................................................................................ 6
2.1.3 Polyamine transport ........................................................................................................................................ 7
2.1.4 Cellular functions of polyamines and SSAT ................................................................................................. 9
2.1.5 Polyamine catabolism in disease .................................................................................................................. 10
2.1.6 Polyamine metabolism in hematopoiesis ................................................................................................... 11
2.1.7 Polyamine metabolism in bone remodeling ............................................................................................... 11
2.2 HEMATOPOIESIS .............................................................................................................................................. 11
2.2.1 Hematopoietic stem cells .............................................................................................................................. 13
2.2.2 Maintenance of self-renewal capacity of hematopoietic stem cells ......................................................... 13
2.2.3 Lineage commitment of hematopoietic cells .............................................................................................. 14
2.2.4 Regulation of myeloid and megakaryocyte-erythroid lineage commitment ......................................... 16
2.2.5 Aging of the hematopoietic system ............................................................................................................. 16
2.2.6 Bone marrow microenvironment ................................................................................................................. 17
2.3 BONE STRUCTURE, FUNCTION AND DEVELOPMENT ......................................................................... 18
2.3.1 Bone remodeling ............................................................................................................................................ 18
2.3.2 Osteoblasts and bone formation................................................................................................................... 19
2.3.3 Osteoclasts and bone resorption .................................................................................................................. 20
3 AIMS OF THE STUDY ............................................................................................................ 21
4 MATERIALS AND METHODS ............................................................................................ 23
4.1 PATIENT SAMPLES .......................................................................................................................................... 23
4.2 MICE .................................................................................................................................................................... 23
4.3 ISOLATION OF BONE MARROW CELLS FROM MICE............................................................................. 23
4.4 EXPANSION OF FACS-SORTED LSK CELLS ............................................................................................... 24
4.5 DIFFERENTIATION OF MESENCHYMAL STROMAL CELLS TO OSTEOBLASTS .............................. 24
4.6 DIFFERENTIATION OF HEMATOPOIETIC CELLS TO OSTEOCLASTS ................................................ 24
4.7 STATISTICAL ANALYSES ............................................................................................................................... 24
4.8 ANALYTICAL METHODS ............................................................................................................................... 24
5 RESULTS .................................................................................................................................... 27
5.1 EFFECTS OF ACTIVATED POLYAMINE CATABOLISM IN IMMUNE RESPONSE (I) ........................ 27
5.2 EFFECTS OF ACTIVATED POLYAMINE CATABOLISM ON HEMATOPOISIS IN MICE (II) ............ 28
5.2.1 Polyamine cycle is enhanced in hematopoietic cells of SSAT mice ......................................................... 28
5.2.2 SSAT mice exhibit myeloproliferative phenotype ..................................................................................... 28
5.2.3 The myeloproliferative phenotype of SSAT mice results from both the intrinsic factors of bone
marrow cells and the bone marrow microenvironment ........................................................................................ 29

XIV
5.3 SSAT ACTIVITY IN MYELOID AND LYMPHOID LEUKEMIAS AND EPIGENETICS IN
HEMATOPOIETIC CELLS OF SSAT OVEREXPRESSING MICE (III) .................................................................... 29
5.3.1 Polyamine metabolism of human leukemic blood cells is disturbed ...................................................... 29
5.3.2 SSAT mice show epigenetic changes in hematopoietic cells and differential response to drugs
affecting epigenetic factors......................................................................................................................................... 29
5.4 EFFECTS OF ACTIVATED POLYAMINE CATABOLISM ON BONE REMODELING (IV) .................. 30
5.4.1 Polyamine metabolism of osteoblasts and osteoclasts is disturbed ........................................................ 30
5.4.2 SSAT overexpression affects osteoblastogenesis but not osteoclastogenesis cell-autonomously ....... 30
5.4.3 Induction of SSAT with α-methylspermidine disturbs wild-type osteoblastogenesis ......................... 31
5.4.4 Bone phenotype of SSAT mice reveals striking changes .......................................................................... 31
6 DISCUSSION ............................................................................................................................ 33
7 SUMMARY AND CONCLUSIONS ..................................................................................... 39
8 REFERENCES ............................................................................................................................ 41
ORIGINAL PUBLICATIONS I─IV .............................................................................................. 57

XV
Abbreviations
AdoMet S-adenosylmethionine
AGP alpha-1-acid glycoprotein
ALAT alanine aminotransferase
ALL acute lymphoid leukemia
ALP alkaline phosphatase
AML acute myeloid leukemia
APAO acetylpolyamine oxidase
AZ antizyme
AZI antizyme inhibitor
BMD bone mass density
B.Pm bone perimeter
BSP bone sialoprotein
BV bone volume
C/EBP CCAAT-enhancer-binding
protein
CLP common lymphoid
progenitor
CML chronic myeloid leukemia
CMP common myeloid progenitors
CRP C-reactive protein
Cs.Th cross-sectional thickness
CTSK cathepsin K
CTX carboxy-terminal cross-
linking telopeptide
dcAdoMet decarboxylated S-
adenosylmethionine
DFMO difluoromethylornithine
DNMT DNA methyltransferases
GM-CSF granulocyte-macrophage
colony-stimulating factor
GMP granulocyte-macrophage
progenitors
HAT histone acetyltransferase
HDAC histone deacetylase
HMT histone methyltransferase
HSC hematopoietic stem cell
IL-1β interleukin-1beta
IL-6 interleukin-6
IL-10 interleukin-10
INF-γ interferon gamma
LMPP lymphoid-primed
multipotent progenitor
LPS lipopolysaccharide
LSK lineage-negative Sca-1+ c-Kit+
cells
LT-HSC long-term hematopoietic stem
cell
MEP megakaryocyte-erythrocyte
progenitors
MPP multipotent progenitor
MSC mesenchymal stromal cell
MTA 5’-deoxy-5’-
methylthioadenosine
MT-SSAT spermidine/spermine N1-
acetyltransferase under
metallothionein promoter
XVI
NFATc1 nuclear factor of activated T-
cells cytoplasmic 1
NF-κB nuclear factor kappa-B
OC osteocalcin
ODC ornithine decarboxylase
OPN osteopontin
OSX osterix
PBMC peripheral blood
mononuclear cell
PINP procollagen type I propeptide
Po.V total volume of porosity
RUNX2 runt-related transcription
factor 2
ROS reactive oxygen species
SAA serum amyloid A
SAMDC S-adenosylmethionine
decarboxylase
SCF stem cell factor
SMI structural model index
SMO spermine oxidase
SRS Snyder-Robinson syndrome
SSAT spermidine/spermine N1-
acetyltransferase
SSATKO spermidine/spermine N1-
acetyltransferase knockout
ST-HSC short-term hematopoietic
stem cell
Tb.Th trabecular thickness
Tb.N trabecular number
TNF-α tumor necrosis factor alpha
TPO thrombopoietin
TRAcP tartrate-resistant acid
phosphatase
TV tissue volume
WBC white blood cell

1 Introduction
Cancers in general result from disturbances of several normal cellular functions at the same
time. Tissue homeostasis is disrupted also during the normal aging process due to
alterations occurring at the molecular level and these changes can predispose to cancer
development. In addition to cell-autonomous factors, the surrounding stroma may
contribute to the malignancy of neoplasias. Thus, elucidation of the function of factors
predisposing to cancer represents one way to prevent cancer development.
All blood cells, including platelets, red blood cells and white blood cells, are derived
from multi-potent hematopoietic stem cells. Platelets, which originate from
megakaryocytes, are involved in blood coagulation process whereas the function of red
blood cells is to bind oxygen and deliver it to tissues. White blood cells, further subdivided
into myeloid and lymphoid lineages, are responsible for the immune defense of body. In
the hematopoietic system the disruption of tissue homeostasis with aging leads not only to
an increased propensity to anemia but also to the attenuation of lymphoid lineage and
further to dominance by myeloid lineages (Woolthuis et al. 2011). The domination of
myeloid lineages is connected to the increased incidence of myeloid leukemias observed in
the elderly. In addition to cell-intrinsic factors, recently much attention has been focused on
the role of the micro-environment in the development of hematological malignancies
(Askmyr et al. 2011; Carlesso and Cardoso 2010).
Bone is a connective tissue consisting of cellular components as well as of an organic and
inorganic matrix. Osteoblasts, osteoclasts and osteocytes are the cellular components of
bone. Bone serves a variety of functions e.g. providing structural support and protection for
internal organs as well as a suitable environment where hematopoiesis can take place. Bone
is constantly being renewed by a process termed remodeling (Eriksen 2010; Kular et al.
2012). This is carried out by bone-resorbing osteoclasts and bone-forming osteoblasts. The
osteoclasts break down the old or damaged bone and osteoblasts re-fill the gap in the bone
matrix. In healthy young bone the process of bone resorption and bone formation are
tightly coupled to maintain homeostasis and overall bone mass. Thus, a defect in either of
the events can evoke an imbalance in bone structure, leading to a disease state.
Polyamines are small cationic molecules which have been conserved during the course
of evolution. Due to their cationic nature, polyamines interact with negatively charged
macromolecules (Igarashi and Kashiwagi 2010; Watanabe et al. 1991). Through these
interactions, polyamines are involved in many cellular functions, with the best known
being their function in cell proliferation. Polyamines have been linked also to neoplastic
growth as biosynthesis of polyamines is enhanced in cancer cells as compared to healthy
tissue (Wallace et al. 2003). In cell cultures, the induction of the activity of the catabolic
enzyme of polyamines, spermidine/spermine N1-acetyltransferase (SSAT), has been
successfully used to limit cancer cell growth (Kee et al. 2004b; Vujcic et al. 2000). In animal
models, however, also opposite results are found; SSAT overexpression has been reported
to enhance the animal’s susceptibility to skin and intestinal cancer (Coleman et al. 2002;

2
Tucker et al. 2005). Additionally, human breast cancer patients have been shown to exhibit
increased SSAT activity in their tumor cells compared to healthy tissue (Wallace et al. 2000).
This study aimed to reveal the role of disturbed polyamine metabolism in tissue
homeostasis of the hematopoietic system. Thereby, polyamine levels and activity of their
metabolic enzymes were measured from leukemia patient samples. Furthermore, the
immunological and hematopoietic phenotypes of SSAT overexpressing mice were
examined. These mice were shown to develop a myeloproliferative disease with the bone
marrow microenvironment partly contributing to its development. Thus, in order to clarify
how the microenvironmental factors could affect hematopoiesis the bone remodeling status
of SSAT mice were also characterized.

3
2 Review of the literature
2.1 POLYAMINE METABOLISM
Polyamines are amino acid-derived aliphatic molecules found in all eukaryotes and nearly
all prokaryotes (Cohen 1998). The three polyamines present in mammalian cells, putrescine,
spermidine and spermine, contain two to four amino groups and at physiological pH they
are fully protonated and possess cationic characteristics (Figure 1). Unlike the divalent
inorganic cations, like Ca2+, the positive charge in polyamines is distributed along the
carbon backbone. Thus, polyamines bind readily to negatively charged macromolecules
such as DNA, RNA and proteins and therefore the proportion of free cellular polyamines is
small (Igarashi and Kashiwagi 2010; Watanabe et al. 1991). Spermidine and spermine
interact more strongly with macromolecules than putrescine. The ratio of spermidine and
spermine is claimed to be important for cellular functions (Pegg and Michael 2010). The
cellular content of spermine equals or exceeds that of spermidine only in animal tissues
whereas not all prokaryotic species contain spermine at all. Putrescine is often considered
as the precursor molecule and spermine as a reservoir molecule for spermidine. However,
both of these molecules have been shown to possess their own individual functions.
In view of their ability to form ionic bonds with other molecules and, additionally, to
serve as precursors for other molecules, polyamines are involved in many cellular
functions, such as transcription, translation and cell signaling. As a result of their important
role in these functions, it is recognized that polyamines are essential for normal cell growth
and development of organisms (Nishimura et al. 2002; Pendeville et al. 2001). The cellular
polyamine content and the activity of their biosynthetic enzymes increase rapidly when
proliferation is induced (Wallace 2009). The depletion of the cellular polyamine content
triggers growth arrest by reducing synthesis of nucleic acids and proteins whereas high
concentrations of polyamines may be toxic to cells (Davis et al. 1992; Persson 2009). For this
reason, cellular concentrations of polyamines are very tightly regulated. In addition to their
requirement in cell proliferation, the conservation of polyamines across evolution is also
evidence emphasizing biological significance of these molecules (Wallace et al. 2003).

4
Figure 1 Mammalian polyamines, putrescine, spermidine and spermine. Polyamines are small
aliphatic molecules with two to four cationic amino groups. Positively charged amino groups
enable interaction with negatively charged macromolecules such as DNA and RNA.
2.1.1 Biosynthesis of polyamines
The major source of polyamines in mammals is via de novo biosynthesis, whereas dietary
polyamines and polyamine production by gut microflora make a smaller contribution
(Wallace 2009). The biosynthesis of polyamines uses two amino acids, methionine and
arginine, as precursors (Cohen 1998). Methionine is converted to S-adenosylmethionine
(AdoMet) and L-arginine to L-ornithine and both are then further decarboxylated by S-
adenosylmethionine decarboxylase (SAMDC) and ornithine decarboxylase (ODC),
respectively. The decarboxylation of L-ornithine results in the formation of the diamine
putrescine. Spermidine is synthesized from putrescine, and spermine further from
spermidine, by transfer of aminopropyl group which is provided by decarboxylated S-
adenosylmethionine. The enzymes which catalyze the transfer are spermidine synthase and
spermine synthase. The metabolic pathway of polyamines is presented in Figure 2.
2.1.1.1 Ornithine decarboxylase
Ornithine decarboxylase (ODC) is a pyridoxal phosphate -dependent enzyme which
catalyzes the decarboxylation of L-ornithine to form the diamine putrescine. ODC is one of
the two rate-limiting enzymes of polyamine biosynthesis. The knockout of the gene and
subsequent loss of intracellular polyamine pools is lethal in mice (Pendeville et al. 2001).
ODC functions as a homodimer with two active sites (Coleman et al. 1994). The association
between the two ODC subunits is relatively weak and the dimers exist in rapid equilibrium
with the inactive monomers. ODC has a very rapid turnover rate for a mammalian enzyme.
The depletion of cellular polyamine pools stabilizes the enzyme whereas in excess of
polyamines ODC is directed to degradation. The degradation is brought about by 26S
proteasome but instead of ubiquitination ODC associates with protein termed antizyme
(AZ) that directs it to proteosomal degradation. AZ is synthesized and its degradation is

5
inhibited in response to an increase in cellular polyamine content. It binds the ODC
monomer preventing formation of dimer and, thus, it inhibits enzymatic activity. On the
other hand, AZ activity is regulated by antizyme inhibitor (AZI) which binds AZ more
tightly than ODC and thereby releases ODC from ODC-AZ complex (Murakami et al.
1996). ODC protein levels are controlled also by high cellular polyamine content through
translational regulation.
2.1.1.2 S-adenosylmethionine decarboxylase
The second rate-limiting enzyme of biosynthesis of polyamines is S-adenosylmethionine
decarboxylase (SAMDC) (Nishimura et al. 2002). Inactivation of mouse SAMDC gene and
the subsequent loss of intracellular polyamine pools is lethal at early stage of development.
SAMDC is synthesized as proenzyme which after intramolecular cleavage reactions forms
α and β subunits (Pegg 2009). In mammals, SAMDC functions as a tetramer of two pairs of
α and β subunits and, additionally, it contains a covalently bound prosthetic group,
pyruvate. SAMDC catalyzes the decarboxylation of AdoMet producing an aminopropyl
donor for synthesis of higher polyamines (Pegg et al. 1998). Decarboxylated AdoMet
(dcAdoMet) is required in polyamine biosynthesis whereas the non-decarboxylated form of
AdoMet is needed as a methyl group donor in methyl transfer reactions. The
decarboxylated form is not suitable for this purpose. For this reason, the steady-state level
of dcAdoMet is kept very low and SAMDC activity is highly regulated by polyamines at
the transcriptional, translational and protein levels. Putrescine enhances the proenzyme
processing and catalytic activity of SAMDC whereas high concentrations of spermidine and
spermine suppress transcription and translation and accelerate turn-over rate of SAMDC
(Kameji and Pegg 1987; Pegg 2009). SAMDC is degraded by 26S proteasome after
polyubiquitination.
2.1.1.3 Spermidine and spermine synthases
Spermidine and spermine synthases are aminopropyl transferases that function as
homodimers and use dcAdoMet as a donor of aminopropyl group (Ikeguchi et al. 2006;
Pegg and Michael 2010; Wu et al. 2007). Mammalian aminopropyltransferases are highly
specific for their acceptor molecules, putrescine and spermidine, from which spermidine
and spermine are formed, respectively. The by-product of the process, 5’-deoxy-5’-
methylthioadenosine (MTA), strongly inhibits both spermidine and spermine synthase
(Ikeguchi et al. 2006; Wu et al. 2008). However, MTA is normally rapidly degraded to
adenosine and methionine and, thus, does not have a great inhibitory effect in vivo.
Spermidine and spermine synthases are expressed constitutively and, unlike ODC and
SAMDC, not readily induced (Ikeguchi et al. 2006; Pegg and Michael 2010). Rather their
enzymatic activity is limited by the availability of their common substrate, dcAdoMet.
2.1.2 Polyamine catabolism
Spermine is converted to spermidine and spermidine to putrescine by the concerted action
of two enzymes, spermidine/spermine N1-acetyltransferase (SSAT) and acetylpolyamine
oxidase (APAO) (Casero and Pegg 1993). SSAT adds acetyl groups to aminopropyl end(s)
of spermidine and spermine. The acetylated forms are then either exported or oxidized by
APAO which cleaves the molecules to generate N-acetylaminopropanal and the smaller
polyamine. Additionally, spermine can be converted directly to spermidine by spermine

6
oxidase (SMO) (Vujcic et al. 2002). Due to its ability to directly convert spermine to
spermidine, SMO can be considered as a way to recycle the higher polyamines (Wallace
2009). The spermidine/spermine N1-acetyltransferase (SSAT) pathway, on the other hand,
can be considered to be mainly utilized for depletion of the polyamines as
acetylpolyamines produced by SSAT are readily excreted. For this reason, acetylpolyamines
are only rarely found in normal cells, however, these molecules can be found in high
concentrations in cancer cells (Kingsnorth and Wallace 1985). Furthermore, acetylation of
polyamines reduces their charge and thus decreases their functional ability and potency to
bind anionic molecules.
High levels of SSAT have been shown to increase polyamine flux (Janne et al. 2006; Kee
et al. 2004b). The activity of SSAT decreases polyamine levels and to compensate the
polyamine depletion ODC and SAMDC are induced. The increased flux increases
consumption of ATP and acetyl-CoA in addition to increased production of toxic
compounds such as H2O2.
2.1.2.1 Spermidine/spermine N1-acetyltransferase
Spermidine/spermine N1-acetyltransferase is considered as a key catabolic enzyme of
polyamines (Pegg 2008). SSAT is a part of the GCN5-related N-acetyltransferase (GNAT)
family. It is primarily a cytosolic enzyme, although there is evidence of its localization in
mitochondria and nucleus as well (Holst et al. 2008; Uimari et al. 2009). SSAT is active as a
homodimer. Using acetyl-CoA as a donor for acetyl group SSAT is able to acetylate N1-
position of spermidine and either end of spermine as spermine is a symmetrical molecule
(Matsui and Pegg 1981). In addition, other molecules with the aminopropyl structure are
excellent substrates for SSAT, whereas for example, putrescine with its aminobutyl group
does not (Della Ragione and Pegg 1983).
SSAT is stringently regulated by polyamines and other factors at many levels, e.g.
transcription, mRNA processing, translation and protein stability (Casero and Pegg 2009;
Pegg 2008). Polyamine analogues, several molecules (growth factors, toxic compounds,
drugs) and various pathophysiological stimuli affect SSAT either directly or indirectly
through the regulation of polyamine levels. In the presence of high levels of polyamines,
transcription and translation of SSAT are increased whereas degradation of the protein and
incorrect splicing of the mRNA are reduced (Pegg 2008). Cellular SSAT activity is very low
under resting conditions but it can be readily induced by several factors and the stability of
the otherwise short-lived protein can be enhanced (Matsui and Pegg 1981; Persson and
Pegg 1984). When polyamines and polyamine analogues bind the SSAT protein, its half-life
is greatly prolonged since this prevents the ubiquitination and subsequent degradation by
the 26S proteasome (Coleman and Pegg 1997). Many polyamine analogs have strong effects
on the regulation of SSAT and as they are not substrates for SSAT, their content is not
reduced in response to its induction (Pegg 2008).
2.1.2.2 Acetylpolyamine oxidase and spermine oxidase
APAO is a peroxisomal FAD-dependent enzyme in the polyamine catabolic pathway
catalyzing the turn-over of N1-acetylated polyamines back to spermidine or putrescine
(Casero and Pegg 2009). Spermine can be converted back to spermidine also directly,
without the acetylation step, through the action of SMO (Vujcic et al. 2002). Both reactions,
oxidation by APAO or SMO, produce highly toxic by-products, hydrogen peroxide (H2O2)

7
and 3-acetoamidopropanal or 3-aminopropanal, respectively. APAO is located in
peroxisomes and this means that any oxidizing compounds produced by APAO activity
may be immediately detoxified by peroxisomal catalase. SMO, however, is located in the
cytosol and nucleus. Thus, the reactive oxygen species (ROS) produced by its activity can
be potentially more harmful to the cell (Murray-Stewart et al. 2008). APAO is constitutively
expressed and its activity is limited by the availability of its substrates, the acetylated forms
of polyamines, which are produced by action of SSAT (Holtta 1977; Vujcic et al. 2003).
APAO has very poor ability to oxidize non-acetylated polyamines. SMO, on the other hand,
is highly inducible and all of the analogues inducing SSAT also induce SMO expression
(Casero and Pegg 2009). APAO activity has been shown to be low in breast cancer patients
in comparison to healthy controls, thus partly contributing to the accumulation of cellular
acetylated polyamines (Kingsnorth and Wallace 1985; Wallace et al. 2000).
2.1.3 Polyamine transport
In addition to biosynthesis and catabolism, polyamine uptake and export are important
pathways in the regulation of intracellular polyamine content. If polyamine synthesis is
prevented, then the transport system becomes critical for normal cellular growth. The
polyamine transporters have been well documented in prokaryotes but the transport
mechanisms in mammalian cells are not well characterized and no mammalian transporter
genes have been cloned yet. Food contains large quantities of polyamines which are
absorbed from the intestine and can be transported for use by cells throughout the body
(Bardocz 1993). Additionally intestinal bacteria produce and excrete polyamines which can
be taken up and utilized (Wallace et al. 2003). Due to the positive charge of polyamines,
they require a transport system to permit exogenous uptake. Thus, cells take up polyamines
from the environment through an active, energy-requiring, transport system (Seiler et al.
1996). Two transport systems for uptake of polyamines have been recognized: a sodium-
dependent form that has a preference for putrescine (although it can transport also
spermidine and spermine) and a sodium-independent form that is capable of transporting
spermidine and spermine. Additionally, there has been postulated to be an endocytosis-
mediated mechanism for polyamine transport (Soulet et al. 2002).
The uptake and export of polyamines seem to be mediated by different transporters
since polyamine uptake-deficient mutant cells are able to excrete polyamines (Hyvonen et
al. 1994). However, both systems are regulated by the same factor, AZ, which also functions
as an ODC inhibitor (Sakata et al. 2000). The main polyamines to be exported are putrescine
and acetylated polyamines (Seiler et al. 1996). Acetylated polyamines do not function as
substrates for the polyamine uptake system and thus once exported, they are not
reaccumulated into cells (Byers and Pegg 1989). Exported acetylpolyamines are transported
to the kidneys and excreted in urine. In many cancers, it is known that there are elevated
urinary acetylpolyamine levels (Seiler et al. 1981).

8
Figure 2 Mammalian polyamine metabolism. Polyamine synthesis uses amino acids as
precursors. The polyamine metabolism can be considered as a cycle (red arrows). The higher
polyamines are produced from the smaller forms (spermidine from putrescine and spermine
from spermidine) by the action of biosynthetic enzymes, ODC, SAMDC, SPDSy, SPMSy, and can
again be converted back to the smaller forms by catabolic enzymes SMO, SSAT, APAO. The
polyamine cycle consumes metabolites such as Ac-Co and ATP and produces toxic compounds
(ROS and reactive aldehydes). Ac-coA, acetyl-coenzyme A; APAO, acetylpolyamineoxidase;
ARG, arginine; ATP, adenosine triphosphate; AZ, antizyme; dcAdoMet, decarboxylated S-
adenosylmethionine; MAT, methionine adenosyltransferase; MTA, methylthioadenosine; ODC,
ornithine decarboxylase; ROS, reactive oxygen species; S-AdoMet, S-adenosylmethionine;
SAMDC, S-adenosylmethionine decarboxylase; SMO, spermine oxidase; SPDSy, spermidine
synthase; SPMSy, spermine synthase; SSAT, spermidine/spermine N1-acetyltransferase

9
2.1.4 Cellular functions of polyamines and SSAT
Polyamines can stabilize DNA and protect it from damage that is caused by for example
oxidation and radiation (Ha et al. 1998b; Newton et al. 1996; TABOR 1962). Additionally,
among the polyamines, spermine has been reputed to exert protective roles also during
inflammation as it inhibits post-transcriptionally pro-inflammatory cytokine production in
human mononuclear cells (Ha et al. 1998a; Zhang et al. 1997b). Moreover, spermine has
been shown to reduce carrageenan-induced edema and partially protect from cecal ligation
and puncture -induced sepsis in mice (Zhang et al. 1997b; Zhu et al. 2009). Polyamines
modify gene activation and gene silencing by modulating the transition of DNA from the B-
to the Z-conformation, inducing chromosome condensation by neutralizing the charges on
histone and chromatin phosphate and by regulating activity of the histone
acetyltransferases (Fredericq et al. 1991; Hougaard et al. 1987; Thomas and Messner 1986).
Polyamines are known to be able to influence protein synthesis at various stages by
changing the structure of mRNA, tRNA and rRNA [reviewed in (Igarashi and Kashiwagi
2010)]. Furthermore, polyamines can change protein conformation by undergoing
interactions with acidic protein structures thereby disturbing their functions. Polyamines
regulate also membrane potential of cells by interacting with glutamate receptors and ion
channels (Donevan and Rogawski 1995; Williams 1997).
The best recognized function of polyamines is their role in cell proliferation. Similar to
the content of known cell cycle regulators, cyclins and cyclin-dependent kinases, also
polyamine concentration and activities of polyamine metabolic enzymes fluctuate during
the cell cycle (Oredsson 2003). The activity of ODC, and subsequently the polyamine levels,
peak at G1 and G2 phases whereas AZ and SSAT expressions are up-regulated during the
G2/M phase. The dramatic changes in putrescine concentration during cell cycle suggest
that this divalent polyamine directly drives cells through G1 restriction point to S phase
(Bettuzzi et al. 1999). Additionally, a specific role of spermidine in eukaryotic cell
proliferation is its posttranscriptional protein modification function in the formation of
eukaryotic elongation factor 5A, eIF5A. Spermidine serves as the sole precursor of
hypusine, a rare amino acid, which is required in the maturation of eIF5A (Park et al. 1981).
The maturation of eIF5A is prevented not only by depleted spermidine pools but also by
accumulated putrescine (Tome et al. 1997). Polyamines have a bivalent role in the
regulation of cellular functions as, in addition to cell proliferation, they can regulate also
apoptosis. The studies covering their role in inducing apoptosis are, however,
contradictory. The depletion of polyamines has been shown to both induce and prevent
apoptosis (Schipper et al. 2000). One can conclude from these studies that it is essential for
cell survival to maintain cellular polyamine content within a narrow range.
In addition to its central role in polyamine catabolism, the SSAT enzyme also possess
other functions. SSAT is known to autoacetylate its own lysine residue (Bewley et al. 2006).
Furthermore, SSAT can acetylate the deoxyhypusine/hypusine residue of eIF5A and,
thereby, regulate its activity (Lee et al. 2011). It is possible that also other molecules function
as substrates for SSAT. A recent study revealed SSAT to have an additional role in
repression of expression of other proteins, however, the repression was restricted to
exogenous proteins and only transiently transfected SSAT was able to exert any effects (Lee
et al. 2010). The mechanism, however, is not yet known but the effect of SSAT action does
not seem to depend on acetylpolyamines or on the depletion of polyamines. In addition,
SSAT activity and the concomitant depletion of local polyamines is believed to induce

10
inward rectifier potassium channel Kir4.2 (Chen et al. 2004; deHart et al. 2008).
Furthermore, SSAT binds α9β1 integrin and increased SSAT activity enhances cell
migration through colocalization of α9β1 integrin and Kir4.2 (Chen et al. 2004).
Furthermore, SSAT is linked to inflammatory responses through tumor-necrosis factor-α
(TNF-α) and non-steroidal anti-inflammatory drugs (NSAIDs) which are able to induce
SSAT activity (Babbar et al. 2007). Activation of SSAT will lead to polyamine depletion and
hence to a reduction in cell growth and triggering of apoptosis. This may be beneficial in
conditions of inflammatory stress.
2.1.5 Polyamine catabolism in disease
Studies of transgenic animals overexpressing or deficient for SSAT have pointed to a role
for SSAT in many disease states where the effects of disturbed polyamine metabolism are
mediated differently depending on the diseased tissue. A rapid depletion of spermidine
and spermine increases the susceptibility to pancreatitis (Alhonen et al. 2000; Hyvonen et
al. 2007). The accumulation of putrescine in hair follicles has evoked hair loss and skin
problems in SSAT overexpressing mice (Pietila et al. 1997; Pietila et al. 2001). A similar
polyamine profile and skin changes has been reported also in a patient with keratosis
follicularis spinulosa decalvans (KSFD) (Gimelli et al. 2002). A lower consumption of acetyl-
CoA as a result of reduced SSAT enzyme activity has caused insulin resistance in SSAT
knockout mice (Jell et al. 2007; Niiranen et al. 2006) whereas SSAT overexpressing mice
exhibited an increased consumption of acetyl-CoA and ATP and, thus, they represented
the opposite phenotype (Pirinen et al. 2007). Increased production of ROS as a consequence
of increased polyamine catabolism has been implicated in pathogenesis of
ischemia/reperfusion injury in stroke and acute renal tubular necrosis (Tomitori et al. 2005;
Zahedi et al. 2010).
Since polyamines display such a strong positive correlation with cell growth, it is not
surprising that the growth rate of cancer cells is also closely linked to the polyamine content
of the cell. An increase in ODC activity and the subsequent increase in cellular polyamine
pools have been considered to be an early event in several cancer cell types (Kingsnorth et
al. 1984a; Kingsnorth et al. 1984b). It might be anticipated that inhibition of biosynthesis or
enhancing the catabolism of polyamines would impair neoplastic growth. Decreased tumor
incidence was indeed found in a tumorigenesis initiation study with mice overexpressing
SSAT (Pietila et al. 2001). Furthermore, crossbreeding SSAT overexpressing mice with
TRAMP mice, which develop prostate tumors reduced the incidence of tumors (Kee et al.
2004a). However, SSAT overexpression has been linked also to increased susceptibility to
tumor growth. The tumor cells of breast cancer patients have increased SSAT activity and
decreased APAO activity in comparison to normal tissue (Wallace et al. 2000). The mouse
model of K6/SSAT mice overexpressing SSAT only in skin exhibited increased tumor
incidence in response to a two-stage tumorigenesis initiation protocol (Coleman et al. 2002).
In a study where APCmin mice were crossed with SSAT overexpressing mice, the SSAT
overexpression was associated with a susceptibility towards intestinal tumor growth
(Tucker et al. 2005). However, in such a situation the effect of SSAT seemed to be indirect
since prolonged exposure of intestine to increased levels of bile acid is known to predispose
to carcinogenesis (Reddy et al. 1977) and the SSAT mice have been shown to suffer from an
increased excretion of bile acids (Pirinen et al. 2010).

11
2.1.6 Polyamine metabolism in hematopoiesis
In addition to other neoplasias , polyamine metabolism has also been linked to leukemias.
The erythrocyte spermine content has prognostic value in childhood acute lymphoid
leukemia (ALL) (Bergeron et al. 1997). In addition, the ODC activity in peripheral blood
mononuclear cells from common myeloid leukemia (CML) patients has been shown to
reflect the neoplastic proliferation activity of leukemic cells (Tripathi et al. 2002).
Furthermore, the catabolic side of polyamine metabolism has been implicated in leukemias
since SSAT is differentially expressed during different stages of CML pointing to an
involvement of SSAT in disease transformation (Janssen et al. 2005). Additionally, SSAT
expression is increased in the transition phase of acute megakaryoblastic leukemia (AMKL),
a form of acute myeloid leukemia, common in individuals with Down syndrome (Lightfoot
et al. 2004). Furthermore, SSAT is super-induced in response to treatment with 12-0-
tetradecanoylphorbol-13-acetate (TPA), a compound which triggers the differentiation of
human myeloblastic leukemia cell line ML-1. Thus, SSAT was speculated to be an
important regulator of myeloid differentiation (Wang et al. 1998).
2.1.7 Polyamine metabolism in bone remodeling
There are a few experiments describing the role of polyamines in bone formation (Tjabringa
et al. 2008) and bone resorption (Yamamoto et al. 2012). Tjabringa et al (2008) showed that
addition of spermine could enhance osteoblastogenesis in cultured mesenchymal stromal
cells (MSC). Yamamoto et al. (2012) found spermidine or spermine treatment to prevent
bone loss in ovariectomized mice. The effect of administered polyamines was mediated
through disruption of osteoclast maturation. Polyamines did not have any substantial effect
on osteoblasts in that study. Polyamine metabolism has been lined to bone homeostasis also
in a human disorder called Snyder-Robinson syndrome (SRS) which is associated with
mutations in the spermine synthase gene (Becerra-Solano et al. 2009; Cason et al. 2003; de
Alencastro et al. 2008). SRS patients were reported to suffer from osteoporosis and the
authors claimed that it resembled osteogenesis imperfecta, although, the diagnosis was not
confirmed (Arena et al. 1996; Becerra-Solano et al. 2009; de Alencastro et al. 2008). SRS
patients exhibit only a modest reduction in spermine but their spermidine content is greatly
elevated (Cason et al. 2003). Thus, the spermine:spermidine ratio is reduced and the total
polyamine content increased with the putrescine level being decreased. Additionally, mice
lacking spermine synthase activity, termed gyro (Gy) mice, have been described to have a
similar cellular spermine:spermidine ratio as SRS patients and also to suffer a defect in bone
development in addition to other symptoms (Lyon et al. 1986; Mackintosh and Pegg 2000;
Meyer et al. 1998). However, Gy mice have a deletion of a part of the X chromosome that
inactivates not only the spermine synthase gene but also Phex, a gene that regulates
phosphate metabolism. Therefore, caution is necessary in the use of Gy mice to study the
role of spermine in bone formation (Wang et al. 2004).
2.2 HEMATOPOIESIS
Hematopoietic stem cells (HSC), like all other multi-potent stem cells in the body, are able
to both self-renew and differentiate into more mature cells. Self-renewal is essential for
maintaining the HSC compartment and hence is a prerequisite for lifelong hematopoiesis.
HSC divisions can be either symmetrical or asymmetrical (Blank et al. 2008). During

12
symmetrical cell division, both daughter cells retain stem cell properties whereas in
asymmetrical cell division one of the resulting daughter cells maintains stem cell properties
but the other commits to a more differentiated outcome. As a result of symmetrical cell
division, the stem cell pool is expanded. On the other hand, asymmetrical cell division
maintains the HSC compartment. The more differentiated progeny progressively lose their
self-renewal potential and become more restricted in their differentiation capacity. All
circulating blood cells, including platelets, red blood cells and white blood cells, are
descendants of multi-potent hematopoietic stem cells. Platelets (also called thrombocytes)
are important participants in the blood coagulation process. They are derived by budding
from large precursors called megakaryocytes. Red blood cells, or erythrocytes, bind oxygen
and deliver it to tissues. During differentiation erythrocytes undergo denucleation in order
to gain space for hemoglobin, the molecule able used for binding and transporting oxygen.
White blood cells, also called leukocytes, are responsible for the immune defense in the
body. They are further subdivided into myeloid and lymphoid lineages which both have
several subpopulations with different specialized tasks in immune defense. The
commitment of hematopoietic lineages is presented in Figure 3.
Figure 3 Origin of hematopoietic and bone cells. Hematopoietic and bone cells both originate
from the bone marrow. All the mature hematopoietic cells (right side of figure) develop from
myeloid and lymphoid progenitors that differentiated from the hematopoietic stem cells. The
bone destructing osteoclasts also have a hematopoietic origin whereas the bone forming
osteoblasts derive from the mesenchymal stromal cells. CLP, common lymphoid progenitor;
CMP, common myeloid progenitor; GMP, granulocyte-macrophage progenitor; HSC,
hematopoietic stem cell; LMPP, lymphoid-primed multipotent progenitor; MEP, megakaryocyte-
erythrocyte progenitor; MPP, multipotent progenitor; MSC, mesenchymal stromal cell; OB,
osteoblast; OC, osteoclast

13
2.2.1 Hematopoietic stem cells
The prevailing definition for murine hematopoietic stem cells is Lineage- Sca-1+ c-Kit+,
designated as LSK cells (Ikuta and Weissman 1992; Li and Johnson 1995) (Figure 4.). These
cells lack the lineage-associated surface markers but the expression of stem cell antigen 1
(Sca-1) and c-Kit (also termed receptor for stem cell factor or CD117) is high. HSCs can be
further classified into most primitive self-renewing stem cells, termed long-term
hematopoietic stem cells (LT-HSC), and into more differentiated short-term hematopoietic
stem cells (ST-HSC) and multipotent progenitors (MPP). LT-HSCs are able to reconstitute
the bone marrow of a transplanted animal. The ST-HSCs and MPPs, however, are capable
to only short-term reconstitution. The LT-HSCs reside in Thy1.1lo, CD34-, CD38+, Flt3- or
Hoechstlo fraction of the LSK population (Adolfsson et al. 2001; Goodell et al. 1996;
Morrison and Weissman 1994; Osawa et al. 1996; Randall et al. 1996). There seems to be no
clear-cut phenotypic definition, however, to distinguish ST-HSCs and MPPs from each
other (Iwasaki and Akashi 2007a). HSC compartment is heterogenous in respect of the
lineage and self-renewal potential of the cells and HSCs comprise distinct lineage biased
clonal subtypes (Beerman et al. 2010b). These HSCs with differential lineage potential can
be purified based on the expression of CD150 (Slamf1) (Beerman et al. 2010a). The human
HSCs are defined by the expression of surface markers CD34 and CD38, the most primitive
HSCs (LT-HSC) being lineage- CD34+ CD38- rhodaminelow (McKenzie et al. 2007).
2.2.2 Maintenance of self-renewal capacity of hematopoietic stem cells
Self-renewal capacity, i.e. a daughter cell maintaining its stem cell properties, is sustained
by both the intrinsic factors present in the stem cells and the extrinsic cues of the
microenvironment surrounding the cells. Many transcription factors, such as Gfi-1, Pbx1,
interferon regulatory factor-2, thioredoxin-interacting protein and Nurr-1 are known to be
important regulators of HSC quiescence (Li 2011). For example, Gfi-1 knockout in mice
leads to an increase in the numbers of cycling cells within the HSC pool and the function of
HSCs in bone marrow transplantation experiments is decreased (Hock et al. 2004). The
homeobox (Hox) transcription factors, such as Hoxb4 and Hoxa9, are widely studied
transcription factors indispensable for hematopoiesis. They are expressed preferentially in
HSC and immature progenitor cell populations and are down-regulated during the
differentiation of hematopoietic cells (Argiropoulos and Humphries 2007). The Hox family
transcription factors have overlapping functions and the loss of one gene may be
compensated for by the other members of this family. Quiescent HSCs have also a higher
expression of GATA-2, although its exact role in quiescence regulation in vivo is not clear
(Li 2011; Venezia et al. 2004). Additionally, p53 is important in ensuring that HSCs remain
quiescent and thus the increased expression of p53 is related to aging of hematopoietic cells
(Dumble et al. 2007). Several cytokines, such as IL-3, IL-6, IL-11, Flt-3 ligand,
thrombopoietin (TPO) and stem-cell factor (SCF, c-kit ligand), have been proposed as
regulators of HSC self-renewal and used in the in vitro expansion of HSCs (Blank et al.
2008). However, only studies with TPO and SCF have provided sufficient evidence to
support their important role as positive regulators of stem cell pool. The receptors for TPO
and SCF, c-mpl and c-kit respectively, are expressed on HSCs. Many signaling pathways
have been reported to regulate HSC self-renewal or promote expansion of these cells in
vitro, but nonetheless, their function in vivo has proved to be less pronounced. This
indicates that there are overlapping pathways to secure the control of HSC self-renewal.

14
The maintenance of the self-renewal capacity of HSCs and the promotion of
differentiation of committed cells are regulated also by epigenetic factors, such as DNA
methylation, histone modifications and non-coding RNA (Cedar and Bergman 2011; Scholz
and Marschalek 2012). DNA methyltransferases (DNMTs) catalyze the addition of a methyl
group to the 5’ position of the cytosine ring of DNA, especially to CpG dinucleotide
sequences. The N-terminal histone tails are subject to a vast array of covalent modifications,
including acetylation, methylation, ubiquitination, sumolation and phosphorylation. The
acetylation of histones H3 and H4 by histone acetyltransferases (HAT) promotes the
activation of transcription which is opposed by histone deacetylases (HDACs) that remove
acetyl groups, resulting in the formation of heterochromatin. Histone methylation by
histone methyltransferases (HMTs) can be a signal for either eu- or heterochromatin (active
or inactive chromatin). Methylation of H3K4 is known to signal for euchromatin whereas
methylation of H3K9 and/or H3K27 promotes the association of repressive proteins, such as
polycomb-group (PcG) proteins. The PcG proteins repress the genes that are involved in
cell-cycle regulation and differentiation (Radulovic et al. 2013). Bmi1 is one of the best
studied members of PcG family. It is specifically expressed in non-differentiated
hematopoietic cells and is required for HSC self-renewal. Although, there is no evidence
that Bmi1 overexpression could itself induce leukemia, it is believed to be an important
collaborating factor in leukemic transformation. Furthermore, other participants in
epigenetics have been shown to take part in leukemogenesis (Plass et al. 2008). Thus,
therapeutic agents targeting the enzymes of DNA methylation (DNMT inhibitors such as
decitabine (5-aza-2'-deoxycytidine)) and histone modifications (HDAC inhibitors such as
trichostatin A) are being actively developed, with some already in clinical use.
Since HSCs reside in an environment where the oxygen tension is very low, the
maintenance of low intrinsic oxidant levels has been proposed to be an important way of
keeping HSCs quiescent. The ROS level of HSCs is lower than the ROS level of committed
progenitor cells (Tothova et al. 2007). Forkhead transcription factors, FoxOs, have been
reported to protect HSCs from oxidative stress and triple knockout of FoxO1, FoxO3 and
FoxO4 resulted in an accelerated cycling of HSCs in mice (Tothova et al. 2007).
Additionally, mice deficient for ATM, a cell-cycle checkpoint regulator directly
downstream of FoxO3A, were shown to display increased ROS levels in HSCs which led to
a defect in maintenance of HSC quiescence (Ito et al. 2004).
2.2.3 Lineage commitment of hematopoietic cells
The expression of Sca-1 decreases in the more differentiated cell populations being, low in
common lymphoid progenitors (CLP) and negative in common myeloid progenitors (CMP,
Figure 4). The receptor for interleukin 7 (IL-7Rα) can be used to distinguish these two
populations from each other. IL-7Rα, an essential cytokine for T and B cell development, is
up-regulated in CLPs and absent in CMPs (Akashi et al. 2000; Bhatia et al. 1995; Kondo et
al. 1997). The mature blood cells of lymphoid lineage consist of small, round cells: T, B and
natural killer (NK) cells. Early lymphoid progenitors reside in bone marrow but the true
maturation of T cell lineage takes place in the thymus. Myeloid progenitors can be further
divided into three subsets on the basis of the expression of CD34 and FcγRII/III:
CD34+FcγRII/IIIlo CMPs, CD34-FcγRII/IIIlo megakaryocyte-erythrocyte progenitors (MEPs)
and CD34+FcγRII/IIIhi granulocyte-macrophage progenitors (GMPs). CMPs can produce all
types of myeloid colonies whereas MEPs and GMPs, which differentiate from the CMPs,

15
can generate only granulocytes (i.e. neutrophils, basophils and eosinophils), mast and
macrophage-dendritic cells or megakaryocytes and erythrocytes, respectively (Arinobu et
al. 2005; Iwasaki et al. 2005a). Dendritic cells can be derived also from CLPs (Manz et al.
2001). The monopotent megakaryocyte lineage-committed progenitors, giving rise to
platelets, have been identified in the CD9+ MEP fraction whereas the surface markers of
monopotent erythrocyte progenitors, giving rise to red blood cells, have not been
characterized (Iwasaki and Akashi 2007b; Nakorn et al. 2003).
Although the above definition for the distinct common lymphoid and myeloid
progenitors derived from MPPs is widely used, there is some evidence that the
differentiation routes of hematopoietic cells may be more complex [reviewed in (Iwasaki
and Akashi 2007a)]. The postulated model implies that myeloid differentiation can occur
not only through MPP-CMP pathway but also through lymphoid-primed multipotent
progenitors (LMPP) (Adolfsson, Mansson et al. 2005, Lai, Kondo 2006). LMPPs, defined by
the expression of Fms-like tyrosine kinase 3 (Flt3) and vascular cell adhesion protein 1
(VCAM-1), are lymphoid-biased but possess a granulocyte-macrophage potential. The
myeloid-potential is gradually down-regulated and eventually silenced during the course
of lymphocyte differentiation. The commitment to the lymphoid lineage is finally
established when the cell loses all of its abilities to produce cells with a myeloid lineage, i.e.
at the CLP stage. Megakaryocyte-erythrocyte differentiation is traditionally thought to
occur through the MPP-CMP pathway but there is also evidence that MEPs can be
generated directly from HSC-MPP compartment (Adolfsson et al. 2005; Friedman 2007;
Iwasaki et al. 2005b). LMPPs lack megakaryocyte-erythrocyte potential.
Figure 4 Differentiation path of murine hematopoietic stem and progenitor cells. The
hematopoietic stem cells and the myeloid and lymphoid progenitors can be identified and
divided into subtypes based on the expression levels of the surface markers c-Kit, Sca-1, CD34,
IL-7R and FcγRII/III. CLP, common lymphoid progenitor; CMP, common myeloid progenitor;
GMP, granulocyte-macrophage progenitor; LT-HSC, long-term hematopoietic stem cell; MEP,
megakaryocyte-erythrocyte progenitor; MPP, multipotent progenitor; ST-HSC, short-term
hematopoietic stem cell

16
2.2.4 Regulation of myeloid and megakaryocyte-erythroid lineage commitment
The lineage commitment is controlled by transcription factors that selectively activate or
silence a set of genes. The order in which certain transcription factors become expressed
and the expression level of transcription factor can decide the lineage outcome (Graf and
Enver 2009). The expression level and timing of transcription factors depend on both
intrinsic and extrinsic signals. Additionally, transcription factors may undergo cross-
antagonistic interaction with each other, inactivating transcription factors that direct the
differentiation of other cell types (Dahl et al. 2003; Nerlov et al. 2000; Rekhtman et al. 2003;
Zhang et al. 1999).
PU.1 is one of the most important regulators of hematopoietic lineage commitment. It is
highly expressed in myelomonocytic cells, B cells and at a lower level in their precursors,
such as HSCs, CMPs and CLPs (Akashi et al. 2000). PU.1 regulates myelolymphoid
differentiation and thereby its deficiency impairs the development of granulocytes,
macrophages and lymphocytes but does not have any impact on the development of
megakaryocytes or erythrocytes (McKercher et al. 1996; Scott et al. 1994). Conditional
knock-out models have demonstrated PU.1 to be necessary for the maintenance of HSCs
and indispensable in allowing MPPs to proceed to CMPs, GMPs and CLPs (Dakic et al.
2005; Iwasaki et al. 2005b). Moreover, PU.1 expression favors myeloid over lymphoid
development and monopoiesis over granulopoiesis (Dahl et al. 2003; DeKoter and Singh
2000). C/EBPs are expressed predominantly in granulocytes, monocytes and eosinophils
and C/EBPα also in HSCs, CMPs and GMPs but not in CLPs or MEPs (Friedman 2007;
Traver et al. 2001). Mice deficient in C/EBPα lack neutrophils and eosinophils whereas this
deficiency does not impact on lymphoid or megakaryocyte-erythrocyte development
(Zhang et al. 1997a; Zhang et al. 2004). Moreover, C/EBPα is required to allow CMPs to
proceed to GMPs as mice with a conditional deficiency of C/EBPα lack GMPs but the
development of CMPs, MEPs and CLPs is normal. However, C/EBPα is not absolutely
required for GMPs to differentiate into mature granulocyte-macrophages as demonstrated
by disruption of C/EBPα at GMP stage (Zhang et al. 2004). Disruption of PU.1 at GMP
stage, on the other hand, results in differentiation arrest (Iwasaki et al. 2005b).
GATA factors are essential transcription factors for the development of megakaryocytes
and erythrocytes (Iwasaki and Akashi 2007a). Enforced expression of GATA-1 in HSCs has
been shown to lead to an increase in megakaryocyte-erythrocyte differentiation (Zhang et
al. 2004). For further lineage specification, bipotent megakaryocyte-erythrocyte progenitors
require the presence of additional signals. Erythroid Krüppel-like factor (Kfl-1) and friend
leukemia integration 1 (Fli-1) have been identified as transcription factors directing
erythrocyte and megakaryocyte commitment, respectively (Mancini et al. 2012). GATA-2 is
crucial for the maintenance and proliferation of HSCs and multipotent progenitors (Vicente
et al. 2012). It is normally down-regulated as hematopoiesis proceeds and it is likely
repressed by GATA-1 (Crispino 2005).
2.2.5 Aging of the hematopoietic system
Similar to the homeostasis of other cellular systems, the cellular homeostasis of
hematopoietic system becomes disrupted with age. The most striking feature of an aging
hematopoietic system is the skewing towards a myeloid-biased output. The lymphoid
output, on the contrary, is decreased and adaptive immunity concomitantly diminished.
The incidence of myeloproliferative diseases increases with age whereas lymphoid

17
leukemias are more common in the young (Beerman et al. 2010b; Woolthuis et al. 2011).
Elderly humans and mice have also a greater propensity to anemia. The age-related
changes of hematopoietic system begins from HSCs whose population expands in
advancing age and the repopulation capacity diminishes (Beerman et al. 2010a; Rossi et al.
2005). During aging, the myeloid-biased (CD150high) HSC population expands whereas the
proportion of HSCs with balanced lineage output (CD150low) diminishes. A comparison of
old mice with their young counterparts has additionally revealed that the GMP population
expands with age whereas the frequencies of CMPs and MEPs do not differ from those of
young mice (Rossi et al. 2005). The gene expression profile of hematopoietic stem cells
changes with age and epigenetic factors have been postulated as being important in this
phenomenon (Woolthuis et al. 2011).
2.2.6 Bone marrow microenvironment
In recent years, it has become evident that the stromal cells surrounding solid tumors are in
close contact with the tumor cells and they can promote the transformation of these cells
into a malignant form. Hence, it is not surprising that also the microenvironment around
hematopoietic cells has been claimed to play a role in development of hematological
malignancies (Askmyr et al. 2011; Carlesso and Cardoso 2010). There is also evidence that
leukemic cells are able to influence bone cells and thereby may further contribute to the
severity of the hematopoietic disease (Edwards et al. 2008; Frisch et al. 2012).
Early in life, hematopoiesis occurs in the yolk sac, the aorta-gonadal-mesonephros region
and the liver. After bone forming cells (osteoblasts) have initiated the mineralization of the
bone extracellular matrix, the hematopoietic stem cells move to the bone environment and
in adults, hematopoiesis is localized in bone marrow. It is directed there by the calcium-
sensing receptor expressed on hematopoietic stem cells that enables cells to respond to
extra-cellular ionic calcium concentrations (Adams et al. 2006). The bone marrow
microenvironment is formed not only of the bone mineral matrix but also of cellular
components, such as mesenchymal stromal cells, bone forming and destroying cells,
perivascular cells and sympathetic neurons. Cells of the nervous system have been shown
to support HSC quiescence by influencing the molecular and cellular components of the
microenvironment (Katayama et al. 2006; Yamazaki et al. 2011). The vasculature of bone
marrow is also recognized as being an important factor for hematopoietic homeostasis
(Kopp et al. 2005). The most primitive HSCs are thought to line the endosteal walls of bones
whereas the more differentiated progenitors reside in close proximity to the vasculature
(Kopp et al. 2005; Zhang et al. 2003). HSCs from elderly individuals have been found to
localize more distantly from endosteum due to their reduced adhesion to stromal cells
(Kohler et al. 2009). Osteoblasts, lining the endosteal walls of bones, have been reported to
be one of the key cellular components maintaining hematopoiesis (Calvi et al. 2003). The
ablation of these cells leads to the concomitant loss of bone marrow cells (Visnjic et al.
2004). Osteoblasts maintain HSCs by secreting thrombopoietin (TPO) and by expressing
angiopoietin-1 (Ang-1), stem cell factor (SCF, c-kit ligand) and Jagged-1 on their surface
(Blank et al. 2008; Li 2011). Additionally, the adhesion molecules present in osteoblasts such
as N-cadherin, integrins and osteopontin anchor HSCs to the osteoblastic niche and
regulate the cell-cycle quiescence of HSCs. Adipocytes, on contrary, have been considered
to be negative regulators of hematopoiesis (Naveiras et al. 2009). In aging organisms,
adipocytes seem to displace the hematopoietic cells in the bone marrow.

18
2.3 BONE STRUCTURE, FUNCTION AND DEVELOPMENT
Bone serves a variety of functions providing, e.g. structural support and protection of
internal organs. In addition, it maintains the body’s mineral homeostasis and serves as
environment for hematopoiesis. Based on its structure, bone can be divided into cortical
and trabecular compartments (Bilezikian et al. 2008). Cortical bone is called also compact
bone due to its dense structure. It forms the hard outer layer of bones. Trabecular bone, on
the other hand, is a porous network inside the area that cortical bone has bordered. As a
result of its appearance, trabecular bone is often called spongy bone. In the long bones of
the limbs, trabeculae can be found at the ends of the bones (epiphyses).
Bone is a connective tissue consisting of cellular components, organic matrix and
inorganic elements (Bilezikian et al. 2008). Osteoblasts, osteoclasts and osteocytes are the
cellular components of bone, the latter accounting for 90 percent of bone cellularity. The
most abundant protein in the organic bone matrix, representing over 90% of all the bone
proteins, is type I collagen which is involved in the aligning of the mineral matrix. In order
to assemble collagen molecules into fibrils, procollagen needs to be cleaved by specific
proteinases. This process releases two propeptides, the amino-terminal PINP and carboxy-
terminal PICP whose serum concentrations reflect the rate of synthesis of type I collagen
and therefore have been used as markers of bone formation (Pollmann et al. 2007; Rissanen
et al. 2008). In addition, bone has also several non-collagenous proteins, such as osteopontin
(OPN), bone sialoprotein (BSP) and osteocalcin (OC). In fact, circulating levels of OC can be
used as a specific marker for osteoblastic activity and bone formation (Bilezikian et al.
2008). Bone proteins are bound to hydroxyapatite crystals, the inorganic calcium phosphate
constituent of the bone (accounting for approximately 60% of bone), and they serve
important functions in the regulation of mineralization.
During growth, bone formation occurs via two distinct mechanisms: intramembranous
and endochondral ossification (Clarke 2008). The flat bones are formed by
intramembranous ossification, a mechanism of direct bone formation. In endochondral
ossification, on the other hand, the pre-existing cartilage template is replaced by the
mineralized bone matrix. The cartilage growth plate has an integral role in the postnatal
linear growth of the long bones, nevertheless, the long bones are formed by combination of
these two mechanisms. The peak bone mass is achieved in adulthood, but thereafter bone is
lost as a result of the normal aging process. This means that the bone formation exceeds
bone loss in early life but the situation is reversed in the elderly.
2.3.1 Bone remodeling
After growth, bone appears to be a static organ. However, it is being constantly moulded
and renewed by a process termed remodeling (Eriksen 2010; Kular et al. 2012). Bone
remodeling allows repair of microdamage and replacement of old bone with new. It is
generally thought that the purpose of bone remodeling is to maintain the mechanical
properties of the skeleton as well as regulating mineral homeostasis. Remodeling is carried
out by the bone-resorbing osteoclasts, breaking down the old or damaged bone, and the
bone-forming osteoblasts, re-filling the gaps in the bone matrix. Signals from osteoclasts
and the products of the resorbed matrix partly regulate osteoblastogenesis and bone
formation (Martin and Sims 2005). However, human patients and mice with osteopetrosis
due to impaired osteoclast function have normal or increased numbers of functional

19
osteoblasts, thus indicating that it is the number of osteoclasts, rather than their activity
which is more crucial for osteoblastic bone formation (Karsenty 2000). Mesenchyme
derived cells, such as osteoblasts and osteocytes, are important regulators of
osteoclastogenesis and there are many regulatory factors influencing osteoclast formation.
Some of these influence the cells indirectly by stimulating expression of
osteoblastic/osteocytic receptor activator of nuclear factor kappa B ligand (RANKL).
Osteoblasts regulate osteoclastogenesis additionally by secreting macrophage colony
stimulating factor (M-CSF) (Udagawa et al. 1990). Previously, osteoblasts were thought to
be the major regulatory cell type involved in osteoclastogenesis but in recent years, the
important role of osteocytes has also been recognized (Xiong and O'Brien 2012).
Additionally, osteocytes secrete sclerostin, a protein which inhibits bone formation (van
Bezooijen et al. 2004).
In healthy young bone, the process of resorption and formation are tightly coupled to
maintain homeostasis and overall bone mass. Thus, a defect in either will cause an
imbalance in bone structure leading to a disease state. An imbalance in cellular processes of
bone remodeling is involved in serious bone diseases such as osteoporosis, osteosclerosis,
osteopetrosis and Paget’s disease of bone (Kular et al. 2012). Osteosclerosis and
osteopetrosis are diseases characterized by a marked increase in bone mass due to
stimulated osteoblast activity and osteoclast dysfunction, respectively. Osteoporosis, the
most common bone disease, on the other hand, is characterized by decreased bone mineral
density and by deranged levels and forms of bone proteins. Osteoporosis is classified into
primary and secondary diseases, with the primary form being further classified into
postmenopausal (in women) and age-related (both women and men) osteoporosis. The
pathophysiology of senile bone loss is characterized by bone formation exceeding bone
resorption on the periosteal surface and bone resorption exceeding bone formation on the
endosteal surface (Bilezikian et al. 2008; Clarke 2008). Consequently, bones increase in
diameter and the marrow space expands in the elderly. Osteoporosis is preceded by a state
called osteopenia in which there is a reduced bone mass, but not so extensive to fulfill the
criteria of osteoporosis.
2.3.2 Osteoblasts and bone formation
Osteoblasts are cells of mesenchymal origin (Dennis et al. 1999; Pittenger et al. 1999) (Figure
3). They are responsible for initial bone formation during development and later in bone
remodeling. The maturation of osteoblasts from mesenchymal progenitors consists of three
distinct phases: proliferation, extra-cellular matrix maturation and matrix mineralization.
After mineralization, the majority of osteoblasts undergo apoptosis whereas the remainder
of the population differentiates into osteocytes or quiescent bone lining cells. Osteocytes are
cells that become trapped inside the bone-matrix. They are able to sense a mechanical load
and subsequently send signals which cause bone to be either resorbed or formed.
The runt-related transcription factor 2 (Runx2) is viewed as the master regulator in
embryonic bone development as well as in postnatal differentiation and bone formation of
osteoblasts (see (Ducy et al. 1999; Komori 2003; Stein et al. 2004) for a review). Mice lacking
Runx2 do not produce any osteoblasts. Despite the essential role of Runx2 in osteoblast
commitment, it is not needed for the maintenance of the major bone matrix genes in mature
osteoblasts. Instead, it seems to function as a negative regulator of the terminal
differentiation of osteoblasts (Liu et al. 2001; Marie 2008; Maruyama et al. 2007). Among

20
many other signaling pathways, Wnt/β-catenin pathway has been shown to be an
important promoter of osteoblastogenesis through the stimulation of Runx2 (Gaur et al.
2005). The bone cell phenotype is further reinforced by transcription factor Osterix (OSX)
(Nakashima et al. 2002). OSX is believed to act downstream of Runx2 during osteoblast
differentiation since it is not expressed in Runx2-deficient embryos but Runx2 is expressed
in Osx-deficient mice. After proliferation, the expression of bone-matrix genes, such as type
I collagen and fibronectin, increase in pre-osteoblasts and these cells turn into bone-matrix-
secreting osteoblasts. The expression of bone-matrix proteins osteocalcin, octeopontin and
bone sialoprotein is increased in the mineralization phase in response to Runx2 promotion
(Ganss et al. 1999; Neve et al. 2013; Sodek et al. 2000). These bone-matrix proteins have high
affinity for the mineral constituent of bone, the hydroxyapatite crystal. The mineralization
of the secreted matrix requires the presence of tissue non-specific alkaline phosphatase
(TNAP/ALP), whose expression is increased along with the genes encoding bone-matrix
proteins (Orimo 2010). ALP hydrolyzes pyrophosphate and provides the inorganic
phosphate needed for hydroxyapatite crystallization. The hydroxyapatite crystals are
formed within the matrix vesicles of osteoblasts (and chondrocytes) and deposited between
collagen fibrils. Deactivating mutations in the TNAP gene are responsible for poorly
mineralized cartilage and bones and also the accumulation of pyrophosphate.
2.3.3 Osteoclasts and bone resorption
Osteoclasts are multinucleated gigantic cells derived from the monocyte-macrophage
lineage of hematopoietic cells (Walker 1975) (Figure 3). Despite sharing some cell-surface
markers with macrophages, osteoclasts differ clearly from these cells, most distinctively by
their ability to excavate bone (Chambers 2000). Upon reaching the bone matrix,
mononuclear precursors fuse to form multinucleated osteoclasts which then become
polarized. These polarized osteoclasts generate an isolated environment (sealing zone)
between themselves and the bone and begin the degradation with the resorbing membrane
(ruffled border) (Vaananen and Horton 1995). Osteoclasts secrete protons to lower the pH
and, thus, are able to dissolve minerals from the bone-matrix (Bilezikian et al. 2008).
Additionally, the matrix osteoclasts secrete enzymes, such as matrix metalloproteinases
(MPPs) and cysteine proteinases, especially cathepsin K (CTSK), which degrade the organic
components. When CTSK degrades the bone-matrix, collagen fragments, such as carboxy-
terminal cross-linking telopeptide (CTX), are released and thus the serum concentration of
CTX reflects the resorbing activity of osteoclasts. The bone degradation products are
transcytosed into vesicles via the resorbing osteoclasts from the ruffled border to the basal
membrane. Tartrate-resistant acid phosphatase (TRAcP) is believed to function in the
vesicles to undertake the final steps in the destruction of matrix components. The serum
concentration of TRAcP5b has been shown to reflect the number of osteoclasts (Alatalo et
al. 2004). After resorption, osteoclasts are subjected to apoptosis. The master transcriptional
regulator of osteoclastogenesis is nuclear factor of activated T cells cytoplasmic 1 (NFATc1)
(Zhao et al. 2010). NFATc1 regulates the expression of genes encoding adhesion molecules,
proton pumps and bone-matrix degradation proteins, thus, modulating osteoclast
migration, adhesion, differentiation and activation. NFATc1 is activated by RANKL
signaling which leads to the activation of several downstream molecules, for example NF-
κB, c-jun and c-fos.

21
3 Aims of the study
The main purpose of this study was to investigate the effects of activated polyamine
catabolism on hematopoiesis and bone remodeling, and to evaluate the interplay between
these two processes.
The specific aims were:
To investigate the response of SSAT overexpressing mice to an immunological
challenge
To characterize the hematopoietic phenotype of SSAT overexpressing mice and to
assess the factors affecting the phenotype
To assess the role of SSAT activity in human lymphoid and myeloid leukemias
To characterize the bone phenotype of SSAT overexpressing mice and to elucidate
the molecular mechanisms underlying the changes

22

23
4 Materials and methods
4.1 PATIENT SAMPLES
Peripheral blood mononuclear cells (PBMCs) were obtained from 21 CML, 28 AML and 33
ALL patients at time of diagnosis. Blood samples from AML and ALL patients were
collected in the Department of Medicine at Kuopio University hospital in the years
2001─2004. PBMCs were isolated (CPTTM tubes, BD Biosciences, Franklin Lakes, NJ) and
stored at the Laboratory of Eastern Finland. Blood samples from CML patients were
collected at the Helsinki University Central Hospital (Helsinki, Finland) during years
2004─2011. PBMCs from CML patients were separated with Ficoll centrifugation (GE
Healthcare Bio-Sciences, San Jose, CA, USA). Control PBMCs were obtained from healthy
donors (n=5─12). The use of mononuclear cells taken at diagnosis in AML and ALL patients
was accepted by the Ethical Committee of the North Savo Hospital District and by the
National Supervisory Authority for Welfare and Health. The collection of samples from
CML patients was approved by the Helsinki University Central Hospital Ethics. Written
informed consents were obtained from all patients prior to sample collection.
4.2 MICE
Mice overexpressing the SSAT gene under its endogenous promoter (SSAT mice), their
wild-type littermates (wt) and mice deficient in SSAT (SSATKO mice) were used in the
experiments (Niiranen et al. 2006; Pietila et al. 1997). SSAT and wild-type mice were in
C57BL/6JOlaHsd and SSATKO in C57BL/6J background. All animal experiments were
approved by the National Animal Experiment Board of Finland.
In the LPS experiments, mice were given a single intraperitoneal injection of
lipopolysaccharide (LPS, E. coli strain 0111:B4, Sigma-Aldrich, 2 mg/kg as the non-lethal
dose, 15 mg/kg or 30 mg/kg as the lethal dose) or saline (controls), n=3─10/group. Mice were
monitored once every 1 to 2 hours and sacrificed at different time-points (and if in a
moribund state). In bone marrow transplantation, the recipient mice (n=8─10/group) were
lethally irradiated twice with a dose of 5.5 Gy (fractions 3 hours apart) one day before the
transplantation. Donor bone marrow cells (5x106 cells/ 200 µl of PBS-2% FBS) were injected
into the tail vein of the recipient mice under anesthesia. In cell proliferation studies, 1 mg of
BrdU (BrdU Flow Kit, 557891, BD Biosciences, San Jose, CA) in PBS was administered
intraperitoneally ten to eleven hours before sacrifice (n=6/group). In studies treating mice
with drugs affecting epigenetic factors, the animals were subjected to daily intraperitoneal
injections of decitabine (0.2 mg/kg, 5-Aza-2’-deoxycytidine, Sigma-Aldrich, St. Louis, MO,
n=5─11) or trichostatin A (1 mg/kg, Selleck Chemicals, Houston, TX, n=7─9) for seven and
ten days, respectively. Control animals received intraperitoneal PBS at a dose of 10 ml/kg.
4.3 ISOLATION OF BONE MARROW CELLS FROM MICE
Bone marrow cells were isolated from the bone marrow cavities of the hind legs by flushing
with PBS-10% FBS. The cells were filtered through 40 µm strainers before being used in

24
flow cytometry (n=6─8 mice/group) or bone cell (osteoblasts and osteoclasts) differentiation
experiments.
4.4 EXPANSION OF FACS-SORTED LSK CELLS
Sorted LSK cells pooled from six to eight mice were grown for 16 hours at 37ºC in 5% CO2
in IMDM (Iscove’s modified Dulbeccos medium) supplemented with 1% BSA, 0.1 mM 2-
mercaptoethanol, 2 mM glutamine, human transferrin (200 µg/ml), bovine pancreatic
insulin (10 µg/ml), low density lipoproteins (40 µg/ml), human Flt-3 ligand (100 ng/ml),
human IL-11(100 ng/ml) and murine SCF (50 ng/ml).
4.5 DIFFERENTIATION OF MESENCHYMAL STROMAL CELLS TO OSTEOBLASTS
Osteoblast differentiation from mesenchymal stromal cells was conducted as described
previously (Heino and Hentunen 2008). Shortly, bone marrow cells isolated from 4‒5 weeks old
mice (n=4─6) were cultured in αMEM supplemented with 15% FBS, 10 mM HEPES, 50
µg/ml gentamycin and 10 nM dexamethasone for one week at 37°C with 5 % CO2. Free-
floating cells were removed at day two of culture. Adherent cells, enriched for
mesenchymal stromal cells, were detached by trypsin treatment and plated in six-well
plates with 150 000 cells /well or in 24-well plates with 30 000 cells /well. Differentiation of
osteoblasts was initiated by adding 10 mM Na-β-glycerophosphate (Sigma-Aldrich, St.
Louis, MO) and 70 µg/ml ascorbate-2-phosphate (Sigma-Aldrich, St. Louis, MO).
Dexamethasone was omitted on the third day of differentiation. The medium was changed
on every third day and the cells were differentiated for up to 21 days.
4.6 DIFFERENTIATION OF HEMATOPOIETIC CELLS TO OSTEOCLASTS
Non-adherent bone marrow cells isolated from 4‒5 weeks old mice (n=4─6) were cultured
in αMEM supplemented with 10% FBS, 50 µg/ml gentamycin and 5 ng/ml M-CSF (Merck
Millipore, Billerica, MA) at 37°C with 5 % CO2. After 24 hours, the medium was changed to
contain 30 ng/ml M-CSF. After an additional 48 hours, the cells were plated in equal
numbers and osteoclast formation was induced by addition of 35 ng/ml RANKL (Merck
Millipore, Billerica, MA).
4.7 STATISTICAL ANALYSES
Data are presented as mean ± SD when applicable. GraphPad Prism 5.03 software package
(GraphPad Software Inc., LaJolla, CA) was used to perform the unpaired two-tailed
Student’s t test and Pearson’s correlation analyses. P value < 0.05 was considered to be
statistically significant.
4.8 ANALYTICAL METHODS
Peripheral blood samples were collected from hind leg saphenous vein or directly from the
hearts of sacrificed mice into EDTA (ethylenediaminetetraacetic acid) -coated or serum
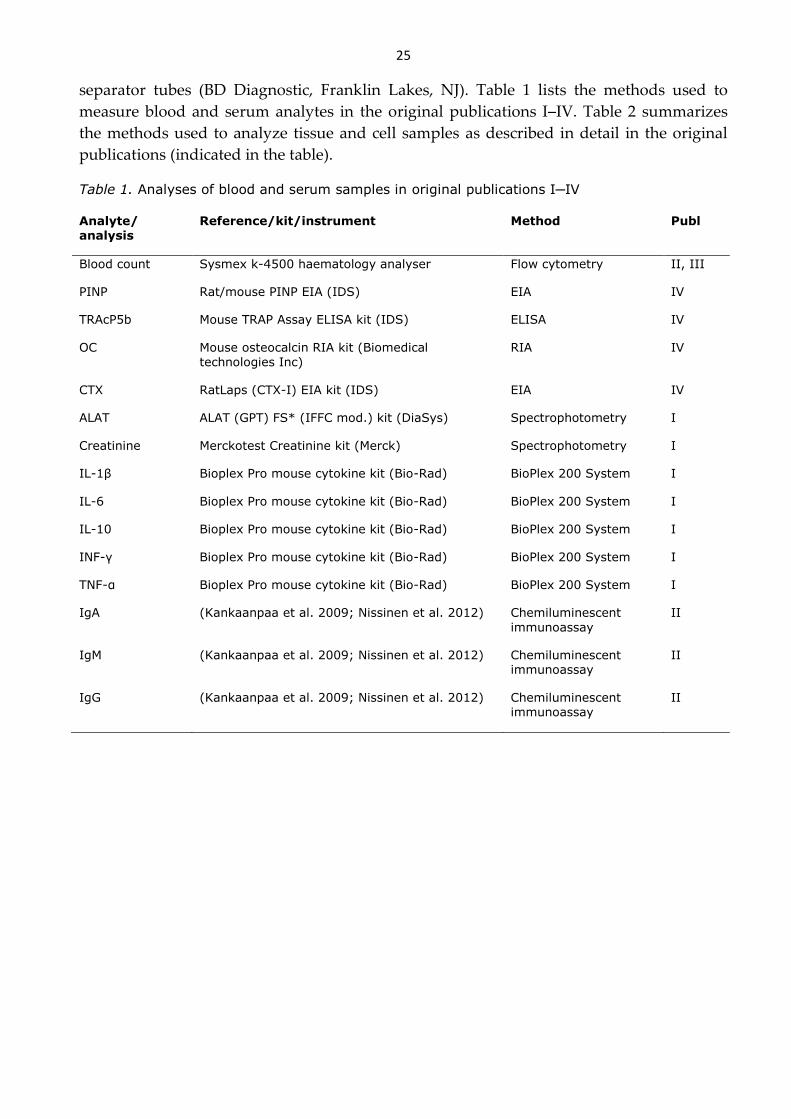
25
separator tubes (BD Diagnostic, Franklin Lakes, NJ). Table 1 lists the methods used to
measure blood and serum analytes in the original publications I─IV. Table 2 summarizes
the methods used to analyze tissue and cell samples as described in detail in the original
publications (indicated in the table).
Table 1. Analyses of blood and serum samples in original publications I─IV
Analyte/ analysis
Reference/kit/instrument Method Publ
Blood count Sysmex k-4500 haematology analyser Flow cytometry II, III
PINP Rat/mouse PINP EIA (IDS) EIA IV
TRAcP5b Mouse TRAP Assay ELISA kit (IDS) ELISA IV
OC Mouse osteocalcin RIA kit (Biomedical
technologies Inc)
RIA IV
CTX RatLaps (CTX-I) EIA kit (IDS) EIA IV
ALAT ALAT (GPT) FS* (IFFC mod.) kit (DiaSys) Spectrophotometry I
Creatinine Merckotest Creatinine kit (Merck) Spectrophotometry I
IL-1β Bioplex Pro mouse cytokine kit (Bio-Rad) BioPlex 200 System I
IL-6 Bioplex Pro mouse cytokine kit (Bio-Rad) BioPlex 200 System I
IL-10 Bioplex Pro mouse cytokine kit (Bio-Rad) BioPlex 200 System I
INF-γ Bioplex Pro mouse cytokine kit (Bio-Rad) BioPlex 200 System I
TNF-α Bioplex Pro mouse cytokine kit (Bio-Rad) BioPlex 200 System I
IgA (Kankaanpaa et al. 2009; Nissinen et al. 2012) Chemiluminescent
immunoassay
II
IgM (Kankaanpaa et al. 2009; Nissinen et al. 2012) Chemiluminescent immunoassay
II
IgG (Kankaanpaa et al. 2009; Nissinen et al. 2012) Chemiluminescent immunoassay
II

26
Table 2. Methods used to analyze samples in the original publications
Method/analysis Tissue/cells Reference/kit Publ
Cell smear PB, BM REASTAIN Quick-Diff kit (Reagena)
II
Histology L, K, S, Th, LN, BM I─II
von Kossa staining OB (Leskela et al. 2003) IV
Bone length and diameter F, T IV
Ash weight T IV
Micro-computed tomography
T (Bouxsein et al. 2010; Maatta et al. 2013)
IV
Biomechanical analysis F, T (Jamsa et al. 2001) IV
ALP activity OB (Leskela et al. 2003) IV
Polyamine concentration
(HPLC)
L, K, S, LN, BM, PBMC, OB, OC (Hyvonen et al. 1992) I─IV
SSAT activity L, K, S, LN, BM, PBMC, OB, OC (Bernacki et al. 1995) I─IV
ODC activity L, K, S, LN, BM, PBMC, OB, OC (Janne and Williams-Ashman 1971)
I─IV
RNA analysis L TRIzol reagent (Invitrogen), High Capacity cDNA Reverse
Transcription Kit (AB)
I
Quantitative real-time PCR LSK RNeasy Mini Kit (Qiagen), High Capacity cDNA Reverse
Transcription Kit (AB)
II
Quantitative real-time PCR OB, OC TRIzol reagent (Invitrogen), High Capacity cDNA Reverse Transcription Kit (AB)
IV
Western blot PBMC III
Protein concentration L, K, S, LN, BM, LSK, PBMC, OB, OC
Protein assay kit (Bio-Rad) I─IV
Magnetic separation BM Mouse hematopoietic progenitor cell enrichment kit (Stemcell Technologies)
II
Flow cytometry PB, S, BM, LSK II
PB, peripheral blood; BM, bone marrow; L, liver; K, kidney; S, spleen; Th, thymus; LN, lymph node; F, femur; T, tibia; OB, osteoblast; OC, osteoclast; PBMC, peripheral blood mononuclear cell; LSK, Lineage-
Sca-1+cKit+ hematopoietic stem cell

27
5 Results
5.1 EFFECTS OF ACTIVATED POLYAMINE CATABOLISM IN IMMUNE
RESPONSE (I)
To monitor whether the immune response of SSAT mice was altered compared to wild-type
mice, the immune system of these mice was challenged with lipopolysaccharide (LPS). LPS
is a structural constituent of the outer membrane of gram-negative bacteria. It activates
immune and liver cells to produce pro- and anti-inflammatory mediators and ultimately
causes endotoxemia leading to septic shock (Pinsky 2004). SSAT mice and wild-type mice
had similar survival rates after administration of a lethal dose of LPS (I, Fig 1). However,
SSAT mice begun to show early signs of inflammation (e.g. diarrhea and lethargy) slightly
before their wild-type littermates. The histopathological changes observed in liver and
kidneys after a non-lethal LPS challenge were similar in SSAT and wild-type mice (I, Fig 3
and 4). The creatinine levels were also similar in both genotypes whereas serum ALAT
activities were significantly higher in SSAT mice in comparison with the wild-type
littermates (I, Fig 2). Thus, the liver of SSAT mice was affected to a greater extent than the
liver of the wild-type mice to the LPS challenge. Serum concentrations of pro-inflammatory
cytokines TNF-α and IL-6 were similar in SSAT and wild-type mice after a non-lethal LPS
challenge whereas the increase in IL-1β and INFγ concentrations was greater in the wild-
type than in the SSAT mice (I, Fig 5). On the other hand, the concentration of the anti-
inflammatory cytokine IL-10 was higher in SSAT mice than in wild-type mice. The
expression levels of hepatic acute-phase proteins C-reactive protein (CRP), haptoglobin and
alpha-1-acid glycoprotein (AGP1) were increased in SSAT mice after a non-lethal LPS
challenge whereas the level of expression of serum amyloid A (SAA1) was comparable to
that of wild-type mice (I, Fig 6). In the basal state, levels of serum cytokines and hepatic
acute-phase proteins showed no differences between genotypes. The lethal LPS challenge
increased ODC activity in kidneys and liver in both genotypes (I, Tables 1 and 3). SSAT
activity, on the other hand, was enhanced only in SSAT mice. Polyamine levels changed in
accordance with the enzyme activities, i.e. putrescine was increased in both tissues in both
genotypes, spermidine was decreased in liver and spermine in kidneys of SSAT mice. A
non-lethal LPS challenge increased hepatic ODC activity and renal SSAT activity in SSAT
mice (I, Table 2 and 4). The wild-type mice displayed no major changes in enzyme levels in
either tissue. (Renal ODC activity was not analysed.) However, putrescine accumulated in
both tissues in both genotypes. In addition, wild-type mice showed an increased level of
spermidine in both tissues after the LPS challenge. N1-acetylspermidine accumulated
mainly only in SSAT mice.

28
5.2 EFFECTS OF ACTIVATED POLYAMINE CATABOLISM ON HEMATOPOISIS IN MICE (II)
5.2.1 Polyamine cycle is enhanced in hematopoietic cells of SSAT mice
Polyamine metabolism was disturbed in SSAT overexpressing mice in all of the
hematopoietic tissues investigated, i.e. bone marrow, spleen, peripheral blood mononuclear
cells, lymph nodes and macrophages (II, Fig 4, bone marrow). SSAT activity was
significantly elevated as also was the activity of the biosynthetic enzyme ODC. Putrescine
became highly accumulated and, uncharacteristically for mammalian cells, its concentration
exceeded that of spermidine and spermine. In fact, the spermidine and spermine
concentrations were decreased. Furthermore, the activity of SSAT increased N1-
acetylspermidine levels. The level of bone marrow SSAT activity was observed to be
associated with the number of cells in bone marrow and with the weight of spleen.
5.2.2 SSAT mice exhibit myeloproliferative phenotype
The blood count of SSAT mice started to exhibit changes at sexual maturation and the
changes became more drastic at an older age (II, Table 1). The number of mature
neutrophils increased with age at the expense of lymphocytes which resulted in an elevated
proportion of neutrophils. The number of platelets was significantly elevated after two
months of age and in old SSAT mice there was evidence of anemia, due to a decreased
number of erythrocytes. The splenic proportion of T cells (CD3+) was decreased and the
proportion of B cells (B220+) was increased in SSAT mice (II, Table 2). Additionally, serum
IgG levels were increased in SSAT mice compared to the wild-type. The ratio of CD4-
positive helper T cells to CD8-positive cytotoxic T cells (CD4+/CD8+) was significantly
reduced and the proportion of regulatory T cells (CD4+CD25+FoxP3+) from all T cells was
significantly higher in the peripheral blood and spleen of SSAT mice. Additionally, spleens
were enlarged in SSAT mice (II, Fig 3). However, the histological sections of spleen, thymus
and liver revealed no morphological differences between SSAT and wild-type mice and no
infiltration of leukocytes or extramedullary hematopoiesis was observed in the liver of
these mice. Bone marrow cellularity was significantly higher in SSAT mice and their bone
marrow exhibited enhanced myelopoiesis and thrombocytopoiesis (II, Fig 2 and 3). The
bone marrow architecture, however, was normal and no fibrosis was detected. The
proportion of blast cells in the bone marrow remained unchanged. The numbers and
proportion of common myeloid progenitors (CMP) and megakaryocyte-erythrocyte
progenitors (MEP) from all bone marrow cells were significantly decreased whereas the
numbers and proportion of granulocyte-macrophage progenitors (GMP) were elevated in
SSAT mice (II, Fig 6). The number and proportion of LSK cells (including hematopoietic
stem cells and multipotent progenitors) and more specifically, long-term hematopoietic
stem cells (LT-HSC) also increased in SSAT mice. Proliferation of LSK cells was found to be
increased in SSAT mice. However, there were no changes in the phases of cell cycle or in
apoptosis of sorted LSK cells expanded in cell culture for 4 days. The expression of HSC
self-renewal regulators Gfi1, Bmi1, GATA-2 and HoxB4 was comparable between wild-type
and SSAT LSK cells as was the expression of a myeloid lineage commitment regulator
C/EBPα. The expression of the megakaryocyte-erythrocyte commitment factor GATA-1 was
increased whereas that of the myelo-lymphoid commitment regulator PU.1 declined in LSK
cells.

29
5.2.3 The myeloproliferative phenotype of SSAT mice results from both the intrinsic
factors of bone marrow cells and the bone marrow microenvironment
Blood count analysis revealed that after short-term transplantation (up to ten weeks) wild-
type and SSAT recipients injected with SSAT bone marrow as well as the SSAT recipients
receiving wild-type cells displayed an increased proportion of neutrophils compared with
the wild-type animals transplanted with wild-type bone marrow (II, Fig 5). Neutrophilia
remained present in all SSAT recipients after long-term reconstitution whereas the wild-
type environment attenuated blood count in long-term reconstitution in wild-type mice
that received SSAT bone marrow. The most prominent phenotype was observed in those
SSAT mice transplanted with SSAT bone marrow. The number of bone marrow cells was
increased only in the SSAT recipients at the time of sacrifice (26 weeks). However, the size
of the spleen was significantly increased in all SSAT recipients as well as in wild-type
recipients transplanted with SSAT bone marrow in comparison to wild-type mice
transplanted with wild-type bone marrow. The bone marrow cells of SSAT recipients and
the wild-type mice transplanted with SSAT bone marrow had increased SSAT activity, an
increased amount of putrescine as well as reduced levels of spermidine compared to wild-
type mice transplanted with wild-type bone marrow.
5.3 SSAT ACTIVITY IN MYELOID AND LYMPHOID LEUKEMIAS AND EPIGENETICS IN HEMATOPOIETIC CELLS OF SSAT OVEREXPRESSING MICE (III)
5.3.1 Polyamine metabolism of human leukemic blood cells is disturbed
Levels of spermidine and spermine were significantly increased in peripheral blood
mononuclear cells (PBMCs) of acute myeloid leukemia (AML), chronic myeloid leukemia
(CML) and acute lymphoid leukemia (ALL) (III, Fig 1) patients compared with the healthy
controls and the levels correlated positively with peripheral blood white blood cell (WBC)
count and blast percentage. Additionally, the activity of SSAT was increased in all leukemia
types. Interestingly, in myeloid leukemia patients (AML and CML), but not ALL patients,
with the peripheral WBC count above the reference range SSAT activity was increased
compared with the patients with normal or low peripheral blood WBC count. The SSAT
activity correlated positively with the peripheral blood WBC count. After treatment, both
the PBMC SSAT activity and WBC count of the follow-up samples of CML patients
declined to the level of healthy controls. The activity of ODC and the level of putrescine
were elevated only in ALL patients and the ODC activity correlated negatively with WBC
count and blasts percentage.
5.3.2 SSAT mice show epigenetic changes in hematopoietic cells and differential
response to drugs affecting epigenetic factors
The amount of methylation was significantly increased in tri-methylated lysine 36, 9 and 4
residues and also in di-methylated lysine 79 residue of histone 3 in the bone marrow cells of
SSAT mice compared to wild-type mice (III, Fig 4). The acetylation of histone 3 in lysine 9
and lysine 14 residues and the level of histone deacetylase 1 (HDAC1) were also slightly
increased in the bone marrow cells of SSAT mice compared to wild-type mice. Seven day
treatment with decitabine, a DNA methyltransferase inhibitor used as an antileukemic drug
in the therapy of human myeloid malignancies, evened up the changes in the peripheral

30
blood granulocytic and lymphoid cells between SSAT and wild-type littermates (III, Fig 2).
In addition, the bone marrow cell number of SSAT mice was equal to that of the wild-type
mice after treatment with decitabine. The SSAT activity of the bone marrow cells was not
affected by the treatment. Ten day treatment with trichostatin A, a histone deacetylase
inhibitor, did not affect WBCs of SSAT mice but significantly decreased the proportion and
number of lymphocytes and increased the proportion of neutrophils in wild-type mice (III,
Fig 3). The treatment also reduced the total WBC count in wild-type animals whereas in
SSAT mice, the WBC count was unaltered.
5.4 EFFECTS OF ACTIVATED POLYAMINE CATABOLISM ON BONE REMODELING (IV)
5.4.1 Polyamine metabolism of osteoblasts and osteoclasts is disturbed
Polyamine metabolism of in vitro differentiated osteoblasts of SSAT overexpressing mice
showed significantly increased SSAT activity, accumulation of putrescine and a decreased
level of spermidine (IV, Fig 5). The spermine level was unaltered and there was no
accumulation of the acetylated forms of polyamines. Interestingly, ODC activity was
reduced in SSAT osteoblasts differentiated for 14 days but it increased as differentiation
proceeded further. In SSATKO osteoblasts, ODC activity was decreased, the putrescine
level was significantly reduced whereas spermidine and spermine levels remained
unaltered when compared to wild-type osteoblasts. Polyamine metabolism of in vitro
differentiated SSAT osteoclasts showed increased SSAT and ODC activities and
accumulation of putrescine (unpublished data). Spermidine and spermine levels were
unchanged and no acetylated forms of polyamines were detected.
5.4.2 SSAT overexpression affects osteoblastogenesis but not osteoclastogenesis cell-
autonomously
Mesenchymal stromal cells of SSAT mice differentiated into osteoblasts in cell culture
showed decreased ALP enzyme activity and a decreased number of mineralized calcium
nodules as compared to osteoblasts of wild-type littermates indicating impairment in
osteoblastogesis (IV, Fig 4). In contrast to SSAT osteoblasts, SSATKO osteoblasts showed
increased osteoblastogenesis. The expression of the osteoblastogenic regulator OSX as well
as of the osteoblastogenic marker genes ALP, OC, OPN and BSP were decreased in SSAT
osteoblasts. On the other hand, expression of RUNX2, the master regulator of
osteoblastogenesis, was not changed. However, as differentiation proceeded, the expression
of RUNX2 and ALP increased, whereas that of the other genes was still significantly lower
in SSAT osteoblasts as compared to the level of expression in wild-type cells. In vivo, the
serum concentration of PINP, a marker of collagen deposition and thus bone formation,
was slightly elevated in SSAT mice compared to wild-type mice (IV, Fig 3). The serum
concentration of OC, a protein binding bone mineral matrix, on the other hand, was
reduced in SSAT mice. The numbers of osteoclasts, differentiated in vitro from bone
marrow cells, were comparable in wild-type and SSAT mice. The expression of the
osteoclastogenic regulator NFATC1 was significantly increased in the osteoclasts of SSAT
mice (IV, Fig 4). However, that did not reflect to the expression levels of the
osteoclastogenic markers, TRACP and CTSK, as their levels were unaltered. In vivo, the
concentration of the bone resorption marker reflecting the number of osteoclasts, TRACP5b,

31
was decreased in SSAT mice whereas the concentration of a marker of osteoclast activity,
CTX, was comparable with that found in wild-type mice (IV, Fig 3).
5.4.3 Induction of SSAT with α-methylspermidine disturbs wild-type
osteoblastogenesis
Treatment of SSAT osteoblasts with α-methylspermidine did not have any effect on their
defective differentiation potential (IV, Fig 6). The ALP activity of SSATKO osteoblasts was
slightly, but statistically significantly, decreased after treatment with the analogue.
Nevertheless, it remained markedly increased as compared to ALP activity of wild-type
and SSAT osteoblasts. ALP activity and the number of mineralized nodules of wild-type
osteoblasts, however, decreased to the level of those in SSAT osteoblasts after treatment
with the analogue. α-Methylspermidine induced SSAT activity in wild-type and SSAT cells
but not in SSATKO cells lacking a functional SSAT gene. The activity of ODC was
decreased similarly in all three genotypes compared to their untreated counterparts. The
levels of natural polyamines decreased drastically in all three genotypes and they were
replaced by the analogue.
5.4.4 Bone phenotype of SSAT mice reveals striking changes
The skeleton of SSAT mice started to show changes at sexual maturation revealing
kyphosis (hunched back). The diameter of femurs and tibiae were larger in SSAT mice (IV,
Table 1). The length of the bones was increased only in males at the age of 5 months. The
diameter and length of long bones of SSATKO mice, however, were unaltered. The bone
mineral content of tibiae was increased in young SSAT mice but comparable between the
three genotypes at older age. The bones of SSAT mice broke more easily than the bones of
wild-type mice when subjected to compression whereas biomechanical studies showed that
in response to a bending force, the bones of female SSAT mice had greater stiffness and
strength (IV, Fig 2). The bones of male SSAT mice showed increased tibial but not femoral
stiffness and strength. In contrast to SSAT mice, the bone strength of SSATKO mice in
response to bending was slightly decreased in the tibiae in females and in the femur necks
of the males. An investigation of tibiae of SSAT and SSATKO mice by computational micro-
tomography revealed drastic differences in their structure compared to wild-type mice (IV,
Fig 1). The endosteal tissue volume (TV) of cortical bone was significantly larger in SSAT
mice and the BV/TV ratio was significantly decreased. Additionally, the bone perimeter
(B.Pm) showed a significant increase in SSAT mice whereas the cross-sectional thickness
(Cs.Th) was reduced. The trabecular thickness (Tb.Th) was reduced and total volume of
porosity (Po.V) was larger in SSAT mice. Additionally, in the 7 month old SSAT mice, the
trabecular number (Tb. N) was increased. In SSATKO mice, both the cortical BV and TV
were reduced resulting in a BV/TV ratio which was comparable to that in the wild-type
mice. B.Pm and Cs.Th were unaltered in SSATKO mice. The Tb. N was reduced in SSATKO
mice compared to wild-type. The bone mass density (BMD) was similar in SSAT and wild-
type mice whereas it was decreased in SSATKO mice when compared to wild-type mice.

32

33
6 Discussion
SSAT is the key catabolic enzyme of polyamines. Activation of SSAT in mice concurrently
enhances the whole polyamine cycle in many tissues which in addition to affecting
polyamine levels, influences the availability of metabolites shared with other metabolic
pathways and increases the production of toxic metabolites. The present work has
characterized the role of enhanced polyamine catabolism in immune defense,
hematopoiesis and bone remodeling.
I Inflammatory challenge of SSAT mice
Neutrophils are the first cells recruited to acute inflammatory sites. They bind surface
structures of pathogens, such as lipopolysaccharide (LPS) of gram-negative bacteria. In
consequence, neutrophils begin to produce cytokines to recruit other immune cells as well
as enzymes and reactive oxygen species (ROS) to kill ingested bacteria. During the early
stages of endotoxic shock neutrophils are known to be essential for suppression of
bacteremia and the preservation of tissue function (Hoesel et al. 2005). However, in the later
stages of endotoxic shock, neutrophils are themselves significant contributors to tissue
damage and organ failure. These present results show that after exposing animals to a
lipopolysaccharide (LPS) -induced endotoxic shock, the anti-inflammatory actions were
enhanced and pro-inflammatory actions inhibited earlier in SSAT mice than in wild-type
mice. Although, SSAT mice revealed an enhanced anti-inflammatory response at the
molecular level during the early stage of LPS challenge, this had no effect on the mortality
of these mice as it was comparable to their wild-type counterparts. The increased
neutrophil numbers of SSAT mice (II) and the harmful effects of neutrophils at later stages
of endotoxic shock could explain the end result of the inflammatory challenge. In further
support of this proposal, SSAT mice had increased alanine aminotransferase (ALAT) values
at 24 hours after the LPS-challenge, evidence of greater liver damage in the late stage of
endotoxic shock.
II-IV SSAT in myeloproliferation and association of hematopoiesis and bone phenotype
in SSAT mice
These present studies demonstrated that overexpression of SSAT in mice evoked a
myeloproliferative phenotype that emerged at sexual maturity and progressed with age.
The hematological phenotype of SSAT mice fulfilled the criteria of a myeloproliferative
disease, according to the Bethesda proposal for classification of murine nonlymphoid
hematopoietic neoplasms (Kogan et al. 2002). This was supported by the immunological
studies which revealed that the basal levels of serum cytokines and acute phase protein
expressions were comparable between SSAT and wild-type mice (I), thus suggesting that
SSAT mice do not suffer from a systemic chronic inflammation. Furthermore, the
hematological phenotype of SSAT mice was shown to originate partly from the intrinsic
SSAT overexpression of hematopoietic cells as demonstrated by transplantation
experiments. However, the bone marrow microenvironment was also shown to contribute
to the development of myeloproliferation in SSAT mice with the most prominent
hematopoietic phenotype being encountered when both the SSAT bone marrow cells and

34
the SSAT microenvironment were present. Nevertheless, the changes seen in bone structure
and in bone forming cells of SSAT mice were in line with the increased hematopoiesis (IV).
The myeloproliferative phenotype of SSAT mice implied that there had been changes in
lineage commitment. This was indeed apparent already at the hematopoietic stem cell
(HSC) level as the expression level of the lineage marker PU.1 was decreased and that of
GATA-1 increased when compared to the expression levels in wild-type HSCs. PU.1 is an
important regulator of the myelolymphoid lineage commitment and GATA-1 an essential
transcription factor for the lineage commitment of megakaryocytes and erythrocytes at the
HSC level (Iwasaki et al. 2005b; Iwasaki and Akashi 2007b). Mature granulocytic cells were
extensively accumulated in the hematopoietic tissues of SSAT mice at the expense of
lymphoid cells. This myeloid domination was apparent already at the progenitor level as
the numbers of granulocyte-macrophage progenitors (GMPs) were elevated in SSAT mice.
In addition to granulocytic accumulation, SSAT overexpression led to an increase in the
number of platelets in SSAT mice as compared to wild-type mice whereas the numbers of
MEPs, the common progenitor for megakaryocytes and red blood cells, were reduced as
were those of mature erythrocytes. The findings from the mouse model were
complemented by the studies of polyamine metabolism in human leukemia samples which
revealed that the SSAT activity was associated with the white blood cell count in patients
with myeloid leukemias (III). In contrast in lymphoid leukemia patients, the increased
peripheral blood mononuclear cell SSAT activity did not correlate with the white blood cell
count. Furthermore, the follow-up samples from treated chronic myeloid leukemia (CML)
patients exhibited a normalized peripheral blood white blood cell count and additionally
normalized peripheral blood mononuclear cell SSAT activity. An earlier study by Janssen et
al (Janssen et al. 2005) has also pointed to a role for SSAT in myeloid leukemias as its
expression was shown to differ during the different stages of CML (being highest in the
chronic phase in comparison to the diagnosis or that in a blast crisis). Moreover, the
expression of SSAT has been shown to be super-induced in response of 12-0-
tetradecanoylphorpol-13-acetate (TPA) induced differentiation of human myeloblastic
leukemia cell line ML-1 (Wang et al. 1998). Interestingly, increased SSAT expression has
also been related to megakaryocyte malignancy in a study which characterized genes in the
transition phase of acute megakaryoblastic leukemia (AMKL), a form of acute myeloid
leukemia in children with Down syndrome (Lightfoot et al. 2004). Taken together, the
earlier studies and these present findings from mouse and human studies indicate that
increased SSAT activity is linked to myeloid and megakaryocyte differentiation.
Polyamines have been shown to play essential roles in the differentiation of many cell
types (Facchini et al. 2012; Pietila et al. 2005; Vuohelainen et al. 2010). In SSAT mice, the
accumulation of putrescine has been reported to disturb the differentiation of keratinocytes
(Pietila et al. 2005). Also the hematopoietic cells of SSAT mice accumulated high levels of
putrescine whereas the levels of spermidine and spermine were reduced. Despite the
marked changes in the polyamine levels of hematopoietic cells, the polyamines do not seem
to substantially affect hematopoietic cell differentiation or the hematopoietic phenotype of
SSAT mice. Firstly, in a preliminary study, where wild-type and SSAT mice were treated
for three months with 2% difluoromethylornithine (DFMO, an ODC inhibitor) to decrease
the biosynthesis of polyamines, there were no changes detected in blood counts or bone
marrow cell counts, i.e. SSAT mice still exhibited neutrophilia as compared to wild-type
mice (unpublished data). However, DFMO treatment did reduce the levels of putrescine

35
and spermidine as well as the activity of SSAT in bone marrow cells of both genotypes.
Nonetheless, the SSAT activity of the cells of SSAT mice remained higher than that of wild-
type mice. Secondly, the blood counts and spleen weights of SSATKO mice were evaluated
but these mice did not exhibit any notable hematopoietic phenotype despite the alterations
in their hematopoietic cell polyamine levels (decreased putrescine and increased
spermidine concentrations, unpublished data). Thus, these results suggest that increased
SSAT activity influences the myeloproliferation in SSAT mice which is in line with the
present findings from studies of human leukemia samples described in earlier chapter.
SSAT has been reported to possess also other functions in addition to regulation of
cellular polyamine levels. For example, SSAT is able to bind integrin α9β1 and binding of
SSAT with integrin α9β1 could potentially influence the differentiation of hematopoietic
cells of SSAT mice as increased SSAT activity has been shown to enhance cell migration
through binding of this integrin (Chen et al. 2004). Integrin α9β1 is widely expressed in
many cell types, including neutrophils and hematopoietic stem cells (Grassinger et al. 2009;
Hoye et al. 2012). Mice lacking integrin α9β1 have been shown to exhibit a dramatic
impairment in neutrophil differentiation due to a defect in granulocyte colony-stimulating
factor (G-CSF) mediated signaling (Chen et al. 2006). The numbers of granulocyte precursor
cells and mature neutrophils are reduced in these mice. The results of Chen et al (2006),
however, indicate that SSAT binding does not affect the interaction of integrin and G-CSF.
Preliminary data from granulocytic cells of SSAT mice indicated that apoptosis of these
cells seemed to be decreased as compared to wild-type granulocytes (unpublished data).
Interestingly, in addition to playing a role in migration and differentiation, α9β1 is known
to regulate cell survival of neutrophils by inhibiting apoptosis through vascular cell
adhesion protein 1 (VCAM-1) binding (Ross et al. 2006). Thus, it is plausible that
hematopoietic phenotype of SSAT mice is partly influenced by the interplay between
integrin α9β1 and enhanced SSAT activity either through increased migration potential or
decreased apoptosis.
The enhanced catabolism in hematopoietic cells of SSAT mice enhanced the whole
polyamine metabolic cycle as, in addition to increased SSAT activity, also the ODC activity
was increased in the cells. The enhanced polyamine cycle could influence the availability of
metabolites, such as acetyl-CoA, which are shared with other metabolic pathways and in
this way it could affect hematopoietic cell differentiation (Jell et al. 2007; Kee et al. 2004a;
Pegg 2008). The consumption of acetyl-coA by SSAT could potentially interfere with the
acetylation processes conducted by histone acetyltransferases. However, it was found that
the increased activity of SSAT did not decrease the acetylation status of H3 lysine 9/14 in
bone marrow cells of SSAT overexpressing mice but rather increased it slightly (III). The
protein level of histone deacetylase 1 (HDAC1), was also increased in bone marrow cells of
SSAT mice compared to wild-type mice and the cells did show slight changes in their
histone methylation profile. Additionally, the myeloproliferative phenotype of SSAT mice
was attenuated when they were exposed to decitabine, a DNA methyltransferase drug that
is in clinical use. These data indicate that the myeloproliferation found in SSAT
overexpressing mice is related to epigenetic changes in the hematopoietic cells.
The enhanced polyamine cycle and concomitantly the increased acetylpolyamine oxidase
activity lead to the production of ROS which could potentially increase the level of
oxidative stress in SSAT mice. As a matter of fact, recently our group reported mice over-
expressing SSAT under the control of a metallothionein promoter (MT-SSAT mice) had an

36
increased cellular protein carbonyl content which was interpreted as an indication of
enhanced oxidative stress (Cerrada-Gimenez et al. 2011). Additionally, overall MT-SSAT
mice have a very similar phenotype to the SSAT mice used in the present experiments,
including similar changes in their blood cell count (unpublished data). Interestingly, ROS
have been implicated in hematopoietic malignancies as there is evidence for the presence of
chronic oxidative stress in chronic myeloid leukemia (CML) and acute myeloid leukemia
(AML) (Ahmad et al. 2008; Sallmyr et al. 2008). Furthermore, in normal hematopoiesis,
ROS are indicated to have signaling functions being linked especially to differentiation of
myeloid lineage cells, e.g. ROS are involved in promoting differentiation of Drosophila
hematopoietic cells akin to the mammalian myeloid progenitors (Owusu-Ansah and
Banerjee 2009). In addition, in murine myeloid progenitors, the ROS level has been shown
to be higher than that of hematopoietic stem cells and increased levels of intracellular ROS
in hematopoietic cells led to a myeloproliferative phenotype in mice which had a
conditional deletion of all FoxO isoforms (Tothova et al. 2007). In addition to differentiation
of myeloid cells, ROS have been shown to contribute to the lineage commitment of
megakaryocytes (Eliades et al. 2012; Sardina et al. 2010). Thus, increased oxidative stress, if
present in SSAT mice, could represent a possible mechanism to account the increased
myeloproliferation seen in these mice.
The bone marrow microenvironment is implicated to be associated with myeloid lineage
commitment of hematopoietic cells. In mouse models it has been shown that the bone
marrow microenvironment contributes to the development of myeloproliferation (Askmyr
et al. 2011; Walkley et al. 2007). Moreover, humans with myeloid diseases suffer from
related bone defects. However, the exact cell type within the microenvironment influencing
the myeloproliferation is not known in either case. The key component supporting HSC,
however, is known to be the osteoblastic cell (Calvi et al. 2003; Zhang et al. 2003). The first
experiments highlighting the importance of osteoblasts in HSC maintenance was conducted
with mice having a conditional inactivation of bone morphogenetic protein receptor type
IA (BMPRIA) (Calvi et al. 2003). These mice show, similarly to the situation in SSAT mice,
an increase in their LT-HSC numbers (II) which was evoked by an increase in the numbers
of early osteoblastic cells. Furthermore, a subsequent study by Raaijmakers et al
(Raaijmakers et al. 2010; Zhang et al. 2003) emphasized the importance of osteoprogenitors
in sustaining hematopoiesis. Interestingly, although osteoblasts of SSAT mice had an
impaired osteogenic potential, the serum concentration of a bone formation marker PINP
was increased, thus, indicating that the number of osteoblastic cells was increased in vivo in
SSAT mice. In support of this proposal, SSAT mice had also an increased number of
trabeculae which are the main sites for osteoblastic cells. SSAT HSCs showed increase in
proliferation rate compared to wild-type HSCs and decrease in expression of PU.1 which is
not only a differentiation marker but also a regulator of HSC maintenance (Iwasaki et al.
2005b). However, no changes were observed between wild-type and SSAT mice in other
self-renewal markers, GFI, BMI, GATA-2 or HOXB4, or in the proportion of apoptotic
HSCs. Thus, the expansion of the HSC pool in SSAT mice was not due to promotion of self-
renewal or to a block of apoptosis. All of these findings indicate that it is the osteoblastic
cells, rather than some intrinsic mechanisms which are likely to be responsible for the
increase in HSC number in SSAT mice. Furthermore, osteoblasts may take part also in the
regulation of lineage commitment in these mice.

37
IV Bone remodeling and structure in SSAT mice
SSAT overexpression elicited striking changes in the structure of bones in SSAT mice.
Outwardly the most apparent skeletal change was the hunched back. The long bones of
SSAT mice were larger in perimeter and the cortex became strikingly thin with age. The
bones of old SSAT mice snapped more easily than those of wild-type or SSATKO mice
when subjected to compression. The striking diminution of cortex explains the fragility to
compression. However, SSAT bones tolerated bending slightly better than wild-type or
SSATKO mice, which is understandable as the cortical bone of SSAT mice was larger in the
perimeter. SSAT osteoblasts were indicated to have a defect in the mineralization steps of
bone formation. However, there were no differences in the total bone mineral content of old
wild-type, SSAT and SSATKO mice but since the bones of SSAT mice were of a different
size than those of wild-type or SSATKO mice and the bone volume (BV) was increased in
these mice, there may be differences in the relative mineral density between the genotypes.
Thereby, since the bone mineral density of SSAT mice could not be confirmed, the bone
phenotype of these mice could not be designated as osteopenic/osteoporotic.
The osteoblastogenesis of SSAT mice was cell-autonomously impaired as the activity of
alkaline phosphatase (ALP), an enzyme required for generation of inorganic phosphate for
hydroxyapatite crystallization, was decreased in in vitro differentiated SSAT osteoblasts.
Additionally, calcium nodules were nearly absent and there was reduced gene expression
of late osteogenic markers. The osteoblasts of mice deficient for SSAT showed opposite
features, thus, supporting the view that the level of SSAT activity affected
osteoblastogenesis. Serum markers supported the in vitro results of impaired mineralization
steps of bone formation in SSAT mice as there was a reduction in the concentration of
osteocalcin, a protein which binds the hydroxyapatite mineral of bone matrix. However,
even though the maturation of osteoblasts was impaired, the circulating PINP was slightly
increased. PINP is a byproduct of collagen cleavage and thus a marker of bone formation
rate. The increased activity of PINP, thereby, indicated the numbers of osteoblastic cells
were increased in SSAT mice. SSAT osteoclasts displayed no notable changes in their
number in cell culture, but the reduced serum tartrate-resistant acid phosphatase
(TRAcP5b) concentration indicated that their number in vivo was decreased (Alatalo et al.
2004). Despite the decreased number of osteoclasts, their activity, as measured by a marker
of collagen degradation (CTX), remained comparable between wild-type and SSAT mice.
This indicated that the relative activity of osteoclasts (activity per osteoclast) was increased
in SSAT mice. The expression of the master transcriptional regulator of osteoclastogenesis,
NFATc1, was increased in SSAT osteoclasts. However, this was not reflected in the
expressions of either TRAcP or cathepsin K (CTSK). The lack of effect on osteoclastogenic
markers may have been due to the early time-point used in the experiment. Taken together,
the bone phenotype of SSAT mice resulted, for the most part, from an impaired function of
osteoblasts.
A recent study has shown that polyamines enhance osteoblastogenesis (Lee et al. 2013).
However in SSAT mice, although the osteoblastic concentration of spermidine was
reduced, the mechanism leading to defective osteoblastogenesis appeared to be related to
the induction of SSAT enzyme activity. First of all, the treatment of SSAT osteoblasts with a
spermidine analogue did not improve osteoblastogenesis in the SSAT mice. Secondly,
SSATKO osteoblasts showed a significantly higher osteoblastogenic potential than that of
SSAT mice, even though their polyamine levels did not dramatically differ from each other.

38
Finally, the osteoblastogenesis in the wild-type osteoblasts was decreased to the level of
SSAT osteoblasts in response to addition of α-methylspermidine to the culture media and
the common feature with the analogue-treated wild-type and SSAT osteoblasts was the
high enzymatic activity of SSAT. In contrast to the situation in hematopoietic cells, the
increased SSAT activity did not affect the acetylation status of histone protein 3 in the
osteoblasts of SSAT overexpressing mice (data not shown). Thus, the effect of polyamine
catabolism on osteoblastogenesis could be mediated through increased oxidative stress. The
MT-SSAT mice, which have been reported to have increased oxidative stress, express also
increased levels of p53 and age prematurely (Cerrada-Gimenez et al. 2011). Similarly to
SSAT mice, MT-SSAT mice display skeletal changes such as kyphosis (unpublished data).
Age-related changes in the bone phenotype, such as expansion of marrow space and
thinning of cortex, are known to result from increased oxidative stress in mice (Almeida et
al. 2007a; Levasseur et al. 2003). Aging mice suffer defects in osteoblastogenesis and,
additionally, the number of osteoclasts declines with age (Almeida et al. 2007b).
Additionally in humans, osteoblastogenic potential of mesenchymal stromal cells (MSCs) is
reduced with age (D'Ippolito et al. 1999). Moreover, oxidative stress has been demonstrated
to impair osteoblastogenesis also in rabbit MSCs and gamma-glutamyl transpeptidase
(GGT) deficient mice (Bai et al. 2004; Levasseur et al. 2003). Therefore, premature aging and
oxidative stress would seem to be plausible explanations for the changes seen in
osteoblastogenesis and in bone phenotype of SSAT mice.

39
7 Summary and conclusions
Not much is known about the role of polyamines, let alone about SSAT, in the immune
response, hematopoiesis and bone formation. These present studies have shed some light
on the role of enhanced polyamine catabolism in these processes. The results demonstrated
that it was the presence of enhanced polyamine catabolism, rather than the polyamines
themselves, which disturbed bone formation in SSAT overexpressing mice. Aberrant bone
formation evoked changes in the bone marrow microenvironment which, together with
intrinsic SSAT overexpression of hematopoietic cells, resulted in a myeloproliferative
disease in these mice. The observation that SSAT activity was associated with the white
blood cell count in human myeloid leukemias is further support for the view that SSAT
activity is connected to myeloid cell development.
I SSAT mice displayed no difference in their response to endotoxic shock with respect to
survival or symptoms. However, at the molecular level, SSAT mice showed a slightly
enhanced anti-inflammatory response. The basal levels of serum cytokines and acute-phase
protein expressions of SSAT mice were comparable to those in their wild-type littermates
which indicated SSAT mice did not suffer from a systemic chronic inflammation in the
basal state.
II The investigation of hematopoietic phenotype of SSAT mice revealed an enhanced
blood leukocyte count dominated by mature neutrophils at the expense of lymphocytes.
The blood platelet number was also increased whereas the red blood cell count decreased
with age, causing anemia. In addition, the spleens of SSAT mice were larger than those of
their wild-type counterparts and the bone marrow cellularity was increased revealing the
presence of enhanced myelopoiesis and thrombocytopoiesis. The numbers of long-term
hematopoietic stem cells and granulocyte-macrophage progenitors were increased in the
bone marrows of SSAT mice whereas the number of megakaryocyte-erythrocyte
progenitors was reduced. Altogether, the hematopoietic phenotype of SSAT mice fulfilled
the criteria of mouse myeloproliferative disease. The myeloproliferative phenotype of SSAT
mice resulted from both the intrinsic SSAT overexpression and from microenvironmental
factors. The myeloproliferative phenotype of SSAT mice indicated that the enhanced SSAT
activity was participating in myeloid differentiation.
III Patients with chronic myeloid leukemia, acute myeloid leukemia and acute lymphoid
leukemia showed increased levels of spermidine and spermine and enhanced SSAT activity
in their peripheral blood mononuclear cells. The increased SSAT activity was associated
with the increased number of white blood cells in myeloid leukemias but not in lymphoid
leukemia. This supported the findings from SSAT mice that SSAT activity was associated
with myeloproliferation. The myeloproliferation of these mice was considered to be
connected to epigenetic factors as the SSAT mice showed a differential response to human
leukemia drugs affecting epigenetics and the acetylation and methylation patterns of bone
marrow cells differed from those found in their wild-type controls.
IV The bones of SSAT mice broke more easily than those of wild-type mice when
compressed. The long bones of SSAT mice were enlarged at the perimeter yet had a
strikingly thinned cortical wall. The thickness of the trabeculae was also decreased in SSAT

40
mice but their number was increased. Osteoblastogenesis was cell-autonomously impaired
in SSAT mice. Nevertheless, there appeared to be a greater number of premature
osteoblasts. Altogether, the cellular and structural changes in the bones meant that there
was a larger niche for hematopoietic cells. Thus, the bone phenotype studies supported the
view that the myeloproliferation of SSAT mice was partly due to bone marrow
microenvironmental factors. The study also indicated that SSAT activity impaired
osteoblastogenesis through some mechanism other than depletion of polyamine levels. This
mechanism could potentially be through excess production of ROS.
These findings increase our understanding of the role of polyamine metabolism in
hematopoiesis and bone remodeling. Together with earlier studies, these present findings
emphasize the important role of SSAT in myeloid cell differentiation. In addition, these
studies contribute to the body of research investigating the interplay between
hematopoietic cells and bone microenvironment. Importantly, they also highlight the
contribution of the catabolic part of the polyamine cycle in pathophysiological processes. In
particular, highlighting the possibility that functions other than polyamine depletion may
be responsible for the disturbed cellular homeostasis in bone forming cells and perhaps also
in hematopoietic cells.

41
8 References
Adams GB, Chabner KT, Alley IR, Olson DP, Szczepiorkowski ZM, Poznansky MC, Kos CH,
Pollak MR, Brown EM, Scadden DT (2006) Stem cell engraftment at the endosteal niche is specified
by the calcium-sensing receptor. Nature 439:599-603.
Adolfsson J, Mansson R, Buza-Vidas N, Hultquist A, Liuba K, Jensen CT, Bryder D, Yang L, Borge
OJ, Thoren LA, Anderson K, Sitnicka E, Sasaki Y, Sigvardsson M, Jacobsen SE (2005)
Identification of Flt3+ lympho-myeloid stem cells lacking erythro-megakaryocytic potential a
revised road map for adult blood lineage commitment. Cell 121:295-306.
Adolfsson J, Borge OJ, Bryder D, Theilgaard-Monch K, Astrand-Grundstrom I, Sitnicka E, Sasaki
Y, Jacobsen SE (2001) Upregulation of Flt3 expression within the bone marrow Lin(-)Sca1(+)c-
kit(+) stem cell compartment is accompanied by loss of self-renewal capacity. Immunity 15:659-669.
Ahmad R, Tripathi AK, Tripathi P, Singh S, Singh R, Singh RK (2008) Malondialdehyde and protein
carbonyl as biomarkers for oxidative stress and disease progression in patients with chronic myeloid
leukemia. In Vivo 22:525-528.
Akashi K, Traver D, Miyamoto T, Weissman IL (2000) A clonogenic common myeloid progenitor
that gives rise to all myeloid lineages. Nature 404:193-197.
Alatalo SL, Ivaska KK, Waguespack SG, Econs MJ, Vaananen HK, Halleen JM (2004) Osteoclast-
derived serum tartrate-resistant acid phosphatase 5b in Albers-Schonberg disease (type II autosomal
dominant osteopetrosis). Clin Chem 50:883-890.
Alhonen L, Parkkinen JJ, Keinanen T, Sinervirta R, Herzig KH, Janne J (2000) Activation of
polyamine catabolism in transgenic rats induces acute pancreatitis. Proc Natl Acad Sci U S A
97:8290-8295.
Almeida M, Han L, Martin-Millan M, O'Brien CA, Manolagas SC (2007a) Oxidative stress
antagonizes Wnt signaling in osteoblast precursors by diverting beta-catenin from T cell factor- to
forkhead box O-mediated transcription. J Biol Chem 282:27298-27305.
Almeida M, Han L, Martin-Millan M, Plotkin LI, Stewart SA, Roberson PK, Kousteni S, O'Brien
CA, Bellido T, Parfitt AM, Weinstein RS, Jilka RL, Manolagas SC (2007b) Skeletal involution by
age-associated oxidative stress and its acceleration by loss of sex steroids. J Biol Chem 282:27285-
27297.
Arena JF, Schwartz C, Ouzts L, Stevenson R, Miller M, Garza J, Nance M, Lubs H (1996) X-linked
mental retardation with thin habitus, osteoporosis, and kyphoscoliosis: linkage to Xp21.3-p22.12.
Am J Med Genet 64:50-58.
Argiropoulos B, Humphries RK (2007) Hox genes in hematopoiesis and leukemogenesis. Oncogene
26:6766-6776.

42
Arinobu Y, Iwasaki H, Gurish MF, Mizuno S, Shigematsu H, Ozawa H, Tenen DG, Austen KF,
Akashi K (2005) Developmental checkpoints of the basophil/mast cell lineages in adult murine
hematopoiesis. Proc Natl Acad Sci U S A 102:18105-18110.
Askmyr M, Quach J, Purton LE (2011) Effects of the bone marrow microenvironment on
hematopoietic malignancy. Bone 48:115-120.
Babbar N, Murray-Stewart T, Casero RA,Jr (2007) Inflammation and polyamine catabolism: the
good, the bad and the ugly. Biochem Soc Trans 35:300-304.
Bai XC, Lu D, Bai J, Zheng H, Ke ZY, Li XM, Luo SQ (2004) Oxidative stress inhibits osteoblastic
differentiation of bone cells by ERK and NF-kappaB. Biochem Biophys Res Commun 314:197-207.
Bardocz S (1993) The role of dietary polyamines. Eur J Clin Nutr 47:683-690.
Becerra-Solano LE, Butler J, Castaneda-Cisneros G, McCloskey DE, Wang X, Pegg AE, Schwartz
CE, Sanchez-Corona J, Garcia-Ortiz JE (2009) A missense mutation, p.V132G, in the X-linked
spermine synthase gene (SMS) causes Snyder-Robinson syndrome. Am J Med Genet A 149A:328-
335.
Beerman I, Bhattacharya D, Zandi S, Sigvardsson M, Weissman IL, Bryder D, Rossi DJ (2010a)
Functionally distinct hematopoietic stem cells modulate hematopoietic lineage potential during aging
by a mechanism of clonal expansion. Proc Natl Acad Sci U S A 107:5465-5470.
Beerman I, Maloney WJ, Weissmann IL, Rossi DJ (2010b) Stem cells and the aging hematopoietic
system. Curr Opin Immunol 22:500-506.
Bergeron C, Bansard JY, Le Moine P, Bouet F, Goasguen JE, Moulinoux JP, Le Gall E, Catros-
Quemener V (1997) Erythrocyte spermine levels: a prognostic parameter in childhood common acute
lymphoblastic leukemia. Leukemia 11:31-36.
Bernacki RJ, Oberman EJ, Seweryniak KE, Atwood A, Bergeron RJ, Porter CW (1995) Preclinical
antitumor efficacy of the polyamine analogue N1, N11-diethylnorspermine administered by multiple
injection or continuous infusion. Clin Cancer Res 1:847-857.
Bettuzzi S, Davalli P, Astancolle S, Pinna C, Roncaglia R, Boraldi F, Tiozzo R, Sharrard M, Corti A
(1999) Coordinate changes of polyamine metabolism regulatory proteins during the cell cycle of
normal human dermal fibroblasts. FEBS Lett 446:18-22.
Bewley MC, Graziano V, Jiang J, Matz E, Studier FW, Pegg AE, Coleman CS, Flanagan JM (2006)
Structures of wild-type and mutant human spermidine/spermine N1-acetyltransferase, a potential
therapeutic drug target. Proc Natl Acad Sci U S A 103:2063-2068.
Bhatia SK, Tygrett LT, Grabstein KH, Waldschmidt TJ (1995) The effect of in vivo IL-7 deprivation
on T cell maturation. J Exp Med 181:1399-1409.
Bilezikian JP, Raisz LG, Martin TJ (2008) Principles of bone biology Vol 1.
Blank U, Karlsson G, Karlsson S (2008) Signaling pathways governing stem-cell fate. Blood
111:492-503.
Bouxsein ML, Boyd SK, Christiansen BA, Guldberg RE, Jepsen KJ, Muller R (2010) Guidelines for
assessment of bone microstructure in rodents using micro-computed tomography. J Bone Miner Res
25:1468-1486.

43
Byers TL, Pegg AE (1989) Properties and physiological function of the polyamine transport system.
Am J Physiol 257:C545-53.
Calvi LM, Adams GB, Weibrecht KW, Weber JM, Olson DP, Knight MC, Martin RP, Schipani E,
Divieti P, Bringhurst FR, Milner LA, Kronenberg HM, Scadden DT (2003) Osteoblastic cells
regulate the haematopoietic stem cell niche. Nature 425:841-846.
Carlesso N, Cardoso AA (2010) Stem cell regulatory niches and their role in normal and malignant
hematopoiesis. Curr Opin Hematol 17:281-286.
Casero RA, Pegg AE (2009) Polyamine catabolism and disease. Biochem J 421:323-338.
Casero RA,Jr, Pegg AE (1993) Spermidine/spermine N1-acetyltransferase--the turning point in
polyamine metabolism. FASEB J 7:653-661.
Cason AL, Ikeguchi Y, Skinner C, Wood TC, Holden KR, Lubs HA, Martinez F, Simensen RJ,
Stevenson RE, Pegg AE, Schwartz CE (2003) X-linked spermine synthase gene (SMS) defect: the
first polyamine deficiency syndrome. Eur J Hum Genet 11:937-944.
Cedar H, Bergman Y (2011) Epigenetics of haematopoietic cell development. Nat Rev Immunol
11:478-488.
Cerrada-Gimenez M, Pietila M, Loimas S, Pirinen E, Hyvonen MT, Keinanen TA, Janne J, Alhonen
L (2011) Continuous oxidative stress due to activation of polyamine catabolism accelerates aging
and protects against hepatotoxic insults. Transgenic Res 20:387-396.
Chambers TJ (2000) Regulation of the differentiation and function of osteoclasts. J Pathol 192:4-13.
Chen C, Huang X, Atakilit A, Zhu QS, Corey SJ, Sheppard D (2006) The Integrin alpha9beta1
contributes to granulopoiesis by enhancing granulocyte colony-stimulating factor receptor signaling.
Immunity 25:895-906.
Chen C, Young BA, Coleman CS, Pegg AE, Sheppard D (2004) Spermidine/spermine N1-
acetyltransferase specifically binds to the integrin alpha9 subunit cytoplasmic domain and enhances
cell migration. J Cell Biol 167:161-170.
Clarke B (2008) Normal bone anatomy and physiology. Clin J Am Soc Nephrol 3 Suppl 3:S131-9.
Cohen SS (1998) A Guide to the Polyamines. Oxford University Press, New York.
Coleman CS, Pegg AE, Megosh LC, Guo Y, Sawicki JA, O'Brien TG (2002) Targeted expression of
spermidine/spermine N1-acetyltransferase increases susceptibility to chemically induced skin
carcinogenesis. Carcinogenesis 23:359-364.
Coleman CS, Pegg AE (1997) Proteasomal degradation of spermidine/spermine N1-acetyltransferase
requires the carboxyl-terminal glutamic acid residues. J Biol Chem 272:12164-12169.
Coleman CS, Stanley BA, Viswanath R, Pegg AE (1994) Rapid exchange of subunits of mammalian
ornithine decarboxylase. J Biol Chem 269:3155-3158.
Crispino JD (2005) GATA1 in normal and malignant hematopoiesis. Semin Cell Dev Biol 16:137-
147.

44
Dahl R, Walsh JC, Lancki D, Laslo P, Iyer SR, Singh H, Simon MC (2003) Regulation of
macrophage and neutrophil cell fates by the PU.1:C/EBPalpha ratio and granulocyte colony-
stimulating factor. Nat Immunol 4:1029-1036.
Dakic A, Metcalf D, Di Rago L, Mifsud S, Wu L, Nutt SL (2005) PU.1 regulates the commitment of
adult hematopoietic progenitors and restricts granulopoiesis. J Exp Med 201:1487-1502.
Davis RH, Morris DR, Coffino P (1992) Sequestered end products and enzyme regulation: the case
of ornithine decarboxylase. Microbiol Rev 56:280-290.
de Alencastro G, McCloskey DE, Kliemann SE, Maranduba CM, Pegg AE, Wang X, Bertola DR,
Schwartz CE, Passos-Bueno MR, Sertie AL (2008) New SMS mutation leads to a striking reduction
in spermine synthase protein function and a severe form of Snyder-Robinson X-linked recessive
mental retardation syndrome. J Med Genet 45:539-543.
deHart GW, Jin T, McCloskey DE, Pegg AE, Sheppard D (2008) The alpha9beta1 integrin enhances
cell migration by polyamine-mediated modulation of an inward-rectifier potassium channel. Proc
Natl Acad Sci U S A 105:7188-7193.
DeKoter RP, Singh H (2000) Regulation of B lymphocyte and macrophage development by graded
expression of PU.1. Science 288:1439-1441.
Della Ragione F, Pegg AE (1983) Studies of the specificity and kinetics of rat liver
spermidine/spermine N1-acetyltransferase. Biochem J 213:701-706.
Dennis JE, Merriam A, Awadallah A, Yoo JU, Johnstone B, Caplan AI (1999) A quadripotential
mesenchymal progenitor cell isolated from the marrow of an adult mouse. J Bone Miner Res 14:700-
709.
D'Ippolito G, Schiller PC, Ricordi C, Roos BA, Howard GA (1999) Age-related osteogenic potential
of mesenchymal stromal stem cells from human vertebral bone marrow. J Bone Miner Res 14:1115-
1122.
Donevan SD, Rogawski MA (1995) Intracellular polyamines mediate inward rectification of Ca(2+)-
permeable alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptors. Proc Natl Acad
Sci U S A 92:9298-9302.
Ducy P, Starbuck M, Priemel M, Shen J, Pinero G, Geoffroy V, Amling M, Karsenty G (1999) A
Cbfa1-dependent genetic pathway controls bone formation beyond embryonic development. Genes
Dev 13:1025-1036.
Dumble M, Moore L, Chambers SM, Geiger H, Van Zant G, Goodell MA, Donehower LA (2007)
The impact of altered p53 dosage on hematopoietic stem cell dynamics during aging. Blood
109:1736-1742.
Edwards CM, Zhuang J, Mundy GR (2008) The pathogenesis of the bone disease of multiple
myeloma. Bone 42:1007-1013.
Eliades A, Matsuura S, Ravid K (2012) Oxidases and reactive oxygen species during hematopoiesis:
a focus on megakaryocytes. J Cell Physiol 227:3355-3362.
Eriksen EF (2010) Cellular mechanisms of bone remodeling. Rev Endocr Metab Disord 11:219-227.

45
Facchini A, Borzi RM, Olivotto E, Platano D, Pagani S, Cetrullo S, Flamigni F (2012) Role of
polyamines in hypertrophy and terminal differentiation of osteoarthritic chondrocytes. Amino Acids
42:667-678.
Fredericq E, Hacha R, Colson P, Houssier C (1991) Condensation and precipitation of chromatin by
multivalent cations. J Biomol Struct Dyn 8:847-865.
Friedman AD (2007) Transcriptional control of granulocyte and monocyte development. Oncogene
26:6816-6828.
Frisch BJ, Ashton JM, Xing L, Becker MW, Jordan CT, Calvi LM (2012) Functional inhibition of
osteoblastic cells in an in vivo mouse model of myeloid leukemia. Blood 119:540-550.
Ganss B, Kim RH, Sodek J (1999) Bone sialoprotein. Crit Rev Oral Biol Med 10:79-98.
Gaur T, Lengner CJ, Hovhannisyan H, Bhat RA, Bodine PV, Komm BS, Javed A, van Wijnen AJ,
Stein JL, Stein GS, Lian JB (2005) Canonical WNT signaling promotes osteogenesis by directly
stimulating Runx2 gene expression. J Biol Chem 280:33132-33140.
Gimelli G, Giglio S, Zuffardi O, Alhonen L, Suppola S, Cusano R, Lo Nigro C, Gatti R, Ravazzolo
R, Seri M (2002) Gene dosage of the spermidine/spermine N(1)-acetyltransferase ( SSAT) gene with
putrescine accumulation in a patient with a Xp21.1p22.12 duplication and keratosis follicularis
spinulosa decalvans (KFSD). Hum Genet 111:235-241.
Goodell MA, Brose K, Paradis G, Conner AS, Mulligan RC (1996) Isolation and functional
properties of murine hematopoietic stem cells that are replicating in vivo. J Exp Med 183:1797-1806.
Graf T, Enver T (2009) Forcing cells to change lineages. Nature 462:587-594.
Grassinger J, Haylock DN, Storan MJ, Haines GO, Williams B, Whitty GA, Vinson AR, Be CL, Li
S, Sorensen ES, Tam PP, Denhardt DT, Sheppard D, Choong PF, Nilsson SK (2009) Thrombin-
cleaved osteopontin regulates hemopoietic stem and progenitor cell functions through interactions
with alpha9beta1 and alpha4beta1 integrins. Blood 114:49-59.
Ha HC, Sirisoma NS, Kuppusamy P, Zweier JL, Woster PM, Casero RA,Jr (1998a) The natural
polyamine spermine functions directly as a free radical scavenger. Proc Natl Acad Sci U S A
95:11140-11145.
Ha HC, Sirisoma NS, Kuppusamy P, Zweier JL, Woster PM, Casero RA,Jr (1998b) The natural
polyamine spermine functions directly as a free radical scavenger. Proc Natl Acad Sci U S A
95:11140-11145.
Heino TJ, Hentunen TA (2008) Differentiation of osteoblasts and osteocytes from mesenchymal
stem cells. Curr Stem Cell Res Ther 3:131-145.
Hock H, Hamblen MJ, Rooke HM, Schindler JW, Saleque S, Fujiwara Y, Orkin SH (2004) Gfi-1
restricts proliferation and preserves functional integrity of haematopoietic stem cells. Nature
431:1002-1007.
Hoesel LM, Neff TA, Neff SB, Younger JG, Olle EW, Gao H, Pianko MJ, Bernacki KD, Sarma JV,
Ward PA (2005) Harmful and protective roles of neutrophils in sepsis. Shock 24:40-47.
Holst CM, Nevsten P, Johansson F, Carlemalm E, Oredsson SM (2008) Subcellular distribution of
spermidine/spermine N1-acetyltransferase. Cell Biol Int 32:39-47.

46
Holtta E (1977) Oxidation of spermidine and spermine in rat liver: purification and properties of
polyamine oxidase. Biochemistry 16:91-100.
Hougaard DM, Bolund L, Fujiwara K, Larsson LI (1987) Endogenous polyamines are intimately
associated with highly condensed chromatin in vivo. A fluorescence cytochemical and
immunocytochemical study of spermine and spermidine during the cell cycle and in reactivated
nuclei. Eur J Cell Biol 44:151-155.
Hoye AM, Couchman JR, Wewer UM, Fukami K, Yoneda A (2012) The newcomer in the integrin
family: integrin alpha9 in biology and cancer. Adv Biol Regul 52:326-339.
Hyvonen MT, Merentie M, Uimari A, Keinanen TA, Janne J, Alhonen L (2007) Mechanisms of
polyamine catabolism-induced acute pancreatitis. Biochem Soc Trans 35:326-330.
Hyvonen T, Seiler N, Persson L (1994) Characterization of a COS cell line deficient in polyamine
transport. Biochim Biophys Acta 1221:279-285.
Hyvonen T, Keinanen TA, Khomutov AR, Khomutov RM, Eloranta TO (1992) Monitoring of the
uptake and metabolism of aminooxy analogues of polyamines in cultured cells by high-performance
liquid chromatography. J Chromatogr 574:17-21.
Igarashi K, Kashiwagi K (2010) Modulation of cellular function by polyamines. Int J Biochem Cell
Biol 42:39-51.
Ikeguchi Y, Bewley MC, Pegg AE (2006) Aminopropyltransferases: function, structure and genetics.
J Biochem 139:1-9.
Ikuta K, Weissman IL (1992) Evidence that hematopoietic stem cells express mouse c-kit but do not
depend on steel factor for their generation. Proc Natl Acad Sci U S A 89:1502-1506.
Ito K, Hirao A, Arai F, Matsuoka S, Takubo K, Hamaguchi I, Nomiyama K, Hosokawa K, Sakurada
K, Nakagata N, Ikeda Y, Mak TW, Suda T (2004) Regulation of oxidative stress by ATM is required
for self-renewal of haematopoietic stem cells. Nature 431:997-1002.
Iwasaki H, Akashi K (2007a) Myeloid lineage commitment from the hematopoietic stem cell.
Immunity 26:726-740.
Iwasaki H, Akashi K (2007b) Hematopoietic developmental pathways: on cellular basis. Oncogene
26:6687-6696.
Iwasaki H, Mizuno S, Mayfield R, Shigematsu H, Arinobu Y, Seed B, Gurish MF, Takatsu K,
Akashi K (2005a) Identification of eosinophil lineage-committed progenitors in the murine bone
marrow. J Exp Med 201:1891-1897.
Iwasaki H, Somoza C, Shigematsu H, Duprez EA, Iwasaki-Arai J, Mizuno S, Arinobu Y, Geary K,
Zhang P, Dayaram T, Fenyus ML, Elf S, Chan S, Kastner P, Huettner CS, Murray R, Tenen DG,
Akashi K (2005b) Distinctive and indispensable roles of PU.1 in maintenance of hematopoietic stem
cells and their differentiation. Blood 106:1590-1600.
Jamsa T, Viluksela M, Tuomisto JT, Tuomisto J, Tuukkanen J (2001) Effects of 2,3,7,8-
tetrachlorodibenzo-p-dioxin on bone in two rat strains with different aryl hydrocarbon receptor
structures. J Bone Miner Res 16:1812-1820.

47
Janne J, Alhonen L, Pietila M, Keinanen TA, Uimari A, Hyvonen MT, Pirinen E, Jarvinen A (2006)
Genetic manipulation of polyamine catabolism in rodents. J Biochem 139:155-160.
Janne J, Williams-Ashman HG (1971) On the purification of L-ornithine decarboxylase from rat
prostate and effects of thiol compounds on the enzyme. J Biol Chem 246:1725-1732.
Janssen JJ, Klaver SM, Waisfisz Q, Pasterkamp G, de Kleijn DP, Schuurhuis GJ, Ossenkoppele GJ
(2005) Identification of genes potentially involved in disease transformation of CML. Leukemia
19:998-1004.
Jell J, Merali S, Hensen ML, Mazurchuk R, Spernyak JA, Diegelman P, Kisiel ND, Barrero C, Deeb
KK, Alhonen L, Patel MS, Porter CW (2007) Genetically altered expression of spermidine/spermine
N1-acetyltransferase affects fat metabolism in mice via acetyl-CoA. J Biol Chem 282:8404-8413.
Kameji T, Pegg AE (1987) Effect of putrescine on the synthesis of S-adenosylmethionine
decarboxylase. Biochem J 243:285-288.
Kankaanpaa J, Turunen SP, Moilanen V, Horkko S, Remes AM (2009) Cerebrospinal fluid
antibodies to oxidized LDL are increased in Alzheimer's disease. Neurobiol Dis 33:467-472.
Karsenty G (2000) Bone formation and factors affecting this process. Matrix Biol 19:85-89.
Katayama Y, Battista M, Kao WM, Hidalgo A, Peired AJ, Thomas SA, Frenette PS (2006) Signals
from the sympathetic nervous system regulate hematopoietic stem cell egress from bone marrow.
Cell 124:407-421.
Kee K, Foster BA, Merali S, Kramer DL, Hensen ML, Diegelman P, Kisiel N, Vujcic S, Mazurchuk
RV, Porter CW (2004a) Activated polyamine catabolism depletes acetyl-CoA pools and suppresses
prostate tumor growth in TRAMP mice. J Biol Chem 279:40076-40083.
Kee K, Vujcic S, Merali S, Diegelman P, Kisiel N, Powell CT, Kramer DL, Porter CW (2004b)
Metabolic and antiproliferative consequences of activated polyamine catabolism in LNCaP prostate
carcinoma cells. J Biol Chem 279:27050-27058.
Kingsnorth AN, Wallace HM (1985) Elevation of monoacetylated polyamines in human breast
cancers. Eur J Cancer Clin Oncol 21:1057-1062.
Kingsnorth AN, Lumsden AB, Wallace HM (1984a) Polyamines in colorectal cancer. Br J Surg
71:791-794.
Kingsnorth AN, Wallace HM, Bundred NJ, Dixon JM (1984b) Polyamines in breast cancer. Br J
Surg 71:352-356.
Kogan SC, Ward JM, Anver MR, Berman JJ, Brayton C, Cardiff RD, Carter JS, de Coronado S,
Downing JR, Fredrickson TN, Haines DC, Harris AW, Harris NL, Hiai H, Jaffe ES, MacLennan IC,
Pandolfi PP, Pattengale PK, Perkins AS, Simpson RM, Tuttle MS, Wong JF, Morse HC,3rd,
Hematopathology subcommittee of the Mouse Models of Human Cancers Consortium (2002)
Bethesda proposals for classification of nonlymphoid hematopoietic neoplasms in mice. Blood
100:238-245.
Kohler A, Schmithorst V, Filippi MD, Ryan MA, Daria D, Gunzer M, Geiger H (2009) Altered
cellular dynamics and endosteal location of aged early hematopoietic progenitor cells revealed by
time-lapse intravital imaging in long bones. Blood 114:290-298.

48
Komori T (2003) Requisite roles of Runx2 and Cbfb in skeletal development. J Bone Miner Metab
21:193-197.
Kondo M, Weissman IL, Akashi K (1997) Identification of clonogenic common lymphoid
progenitors in mouse bone marrow. Cell 91:661-672.
Kopp HG, Avecilla ST, Hooper AT, Rafii S (2005) The bone marrow vascular niche: home of HSC
differentiation and mobilization. Physiology (Bethesda) 20:349-356.
Kular J, Tickner J, Chim SM, Xu J (2012) An overview of the regulation of bone remodelling at the
cellular level. Clin Biochem 45:863-873.
Lee MJ, Chen Y, Huang YP, Hsu YC, Chiang LH, Chen TY, Wang GJ (2013) Exogenous
polyamines promote osteogenic differentiation by reciprocally regulating osteogenic and adipogenic
gene expression. J Cell Biochem 114:2718-2728.
Lee SB, Park JH, Folk JE, Deck JA, Pegg AE, Sokabe M, Fraser CS, Park MH (2011) Inactivation of
eukaryotic initiation factor 5A (eIF5A) by specific acetylation of its hypusine residue by
spermidine/spermine acetyltransferase 1 (SSAT1). Biochem J 433:205-213.
Lee SB, Park JH, Woster PM, Casero RA,Jr, Park MH (2010) Suppression of exogenous gene
expression by spermidine/spermine N1-acetyltransferase 1 (SSAT1) cotransfection. J Biol Chem
285:15548-15556.
Leskela HV, Risteli J, Niskanen S, Koivunen J, Ivaska KK, Lehenkari P (2003) Osteoblast
recruitment from stem cells does not decrease by age at late adulthood. Biochem Biophys Res
Commun 311:1008-1013.
Levasseur R, Barrios R, Elefteriou F, Glass DA,2nd, Lieberman MW, Karsenty G (2003) Reversible
skeletal abnormalities in gamma-glutamyl transpeptidase-deficient mice. Endocrinology 144:2761-
2764.
Li CL, Johnson GR (1995) Murine hematopoietic stem and progenitor cells: I. Enrichment and
biologic characterization. Blood 85:1472-1479.
Li J (2011) Quiescence regulators for hematopoietic stem cell. Exp Hematol 39:511-520.
Lightfoot J, Hitzler JK, Zipursky A, Albert M, Macgregor PF (2004) Distinct gene signatures of
transient and acute megakaryoblastic leukemia in Down syndrome. Leukemia 18:1617-1623.
Liu W, Toyosawa S, Furuichi T, Kanatani N, Yoshida C, Liu Y, Himeno M, Narai S, Yamaguchi A,
Komori T (2001) Overexpression of Cbfa1 in osteoblasts inhibits osteoblast maturation and causes
osteopenia with multiple fractures. J Cell Biol 155:157-166.
Lyon MF, Scriver CR, Baker LR, Tenenhouse HS, Kronick J, Mandla S (1986) The Gy mutation:
another cause of X-linked hypophosphatemia in mouse. Proc Natl Acad Sci U S A 83:4899-4903.
Maatta JA, Buki KG, Gu G, Alanne MH, Vaaraniemi J, Liljenback H, Poutanen M, Harkonen P,
Vaananen K (2013) Inactivation of estrogen receptor alpha in bone-forming cells induces bone loss
in female mice. FASEB J 27:478-488.
Mackintosh CA, Pegg AE (2000) Effect of spermine synthase deficiency on polyamine biosynthesis
and content in mice and embryonic fibroblasts, and the sensitivity of fibroblasts to 1,3-bis-(2-
chloroethyl)-N-nitrosourea. Biochem J 351 Pt 2:439-447.

49
Mancini E, Sanjuan-Pla A, Luciani L, Moore S, Grover A, Zay A, Rasmussen KD, Luc S, Bilbao D,
O'Carroll D, Jacobsen SE, Nerlov C (2012) FOG-1 and GATA-1 act sequentially to specify
definitive megakaryocytic and erythroid progenitors. EMBO J 31:351-365.
Manz MG, Traver D, Miyamoto T, Weissman IL, Akashi K (2001) Dendritic cell potentials of early
lymphoid and myeloid progenitors. Blood 97:3333-3341.
Marie PJ (2008) Transcription factors controlling osteoblastogenesis. Arch Biochem Biophys
473:98-105.
Martin TJ, Sims NA (2005) Osteoclast-derived activity in the coupling of bone formation to
resorption. Trends Mol Med 11:76-81.
Maruyama Z, Yoshida CA, Furuichi T, Amizuka N, Ito M, Fukuyama R, Miyazaki T, Kitaura H,
Nakamura K, Fujita T, Kanatani N, Moriishi T, Yamana K, Liu W, Kawaguchi H, Nakamura K,
Komori T (2007) Runx2 determines bone maturity and turnover rate in postnatal bone development
and is involved in bone loss in estrogen deficiency. Dev Dyn 236:1876-1890.
Matsui I, Pegg AE (1981) Effect of inhibitors of protein synthesis on rat liver spermidine N-
acetyltransferase. Biochim Biophys Acta 675:373-378.
McKenzie JL, Takenaka K, Gan OI, Doedens M, Dick JE (2007) Low rhodamine 123 retention
identifies long-term human hematopoietic stem cells within the Lin-CD34+CD38- population. Blood
109:543-545.
McKercher SR, Torbett BE, Anderson KL, Henkel GW, Vestal DJ, Baribault H, Klemsz M, Feeney
AJ, Wu GE, Paige CJ, Maki RA (1996) Targeted disruption of the PU.1 gene results in multiple
hematopoietic abnormalities. EMBO J 15:5647-5658.
Meyer RA,Jr, Henley CM, Meyer MH, Morgan PL, McDonald AG, Mills C, Price DK (1998) Partial
deletion of both the spermine synthase gene and the Pex gene in the X-linked hypophosphatemic,
gyro (Gy) mouse. Genomics 48:289-295.
Morrison SJ, Weissman IL (1994) The long-term repopulating subset of hematopoietic stem cells is
deterministic and isolatable by phenotype. Immunity 1:661-673.
Murakami Y, Ichiba T, Matsufuji S, Hayashi S (1996) Cloning of antizyme inhibitor, a highly
homologous protein to ornithine decarboxylase. J Biol Chem 271:3340-3342.
Murray-Stewart T, Wang Y, Goodwin A, Hacker A, Meeker A, Casero RA,Jr (2008) Nuclear
localization of human spermine oxidase isoforms - possible implications in drug response and
disease etiology. FEBS J 275:2795-2806.
Nakashima K, Zhou X, Kunkel G, Zhang Z, Deng JM, Behringer RR, de Crombrugghe B (2002) The
novel zinc finger-containing transcription factor osterix is required for osteoblast differentiation and
bone formation. Cell 108:17-29.
Nakorn TN, Miyamoto T, Weissman IL (2003) Characterization of mouse clonogenic
megakaryocyte progenitors. Proc Natl Acad Sci U S A 100:205-210.
Naveiras O, Nardi V, Wenzel PL, Hauschka PV, Fahey F, Daley GQ (2009) Bone-marrow
adipocytes as negative regulators of the haematopoietic microenvironment. Nature 460:259-263.

50
Nerlov C, Querfurth E, Kulessa H, Graf T (2000) GATA-1 interacts with the myeloid PU.1
transcription factor and represses PU.1-dependent transcription. Blood 95:2543-2551.
Neve A, Corrado A, Cantatore FP (2013) Osteocalcin: skeletal and extra-skeletal effects. J Cell
Physiol 228:1149-1153.
Newton GL, Aguilera JA, Ward JF, Fahey RC (1996) Polyamine-induced compaction and
aggregation of DNA--a major factor in radioprotection of chromatin under physiological conditions.
Radiat Res 145:776-780.
Niiranen K, Keinanen TA, Pirinen E, Heikkinen S, Tusa M, Fatrai S, Suppola S, Pietila M, Uimari
A, Laakso M, Alhonen L, Janne J (2006) Mice with targeted disruption of spermidine/spermine N1-
acetyltransferase gene maintain nearly normal tissue polyamine homeostasis but show signs of
insulin resistance upon aging. J Cell Mol Med 10:933-945.
Nishimura K, Nakatsu F, Kashiwagi K, Ohno H, Saito T, Igarashi K (2002) Essential role of S-
adenosylmethionine decarboxylase in mouse embryonic development. Genes Cells 7:41-47.
Nissinen AE, Laitinen LM, Kakko S, Helander A, Savolainen MJ, Horkko S (2012) Low plasma
antibodies specific for phosphatidylethanol in alcohol abusers and patients with alcoholic
pancreatitis. Addict Biol 17:1057-1067.
Oredsson SM (2003) Polyamine dependence of normal cell-cycle progression. Biochem Soc Trans
31:366-370.
Orimo H (2010) The mechanism of mineralization and the role of alkaline phosphatase in health and
disease. J Nippon Med Sch 77:4-12.
Osawa M, Hanada K, Hamada H, Nakauchi H (1996) Long-term lymphohematopoietic
reconstitution by a single CD34-low/negative hematopoietic stem cell. Science 273:242-245.
Owusu-Ansah E, Banerjee U (2009) Reactive oxygen species prime Drosophila haematopoietic
progenitors for differentiation. Nature 461:537-541.
Park MH, Cooper HL, Folk JE (1981) Identification of hypusine, an unusual amino acid, in a protein
from human lymphocytes and of spermidine as its biosynthetic precursor. Proc Natl Acad Sci U S A
78:2869-2873.
Pegg AE, Michael AJ (2010) Spermine synthase. Cell Mol Life Sci 67:113-121.
Pegg AE (2009) S-Adenosylmethionine decarboxylase. Essays Biochem 46:25-45.
Pegg AE (2008) Spermidine/spermine-N(1)-acetyltransferase: a key metabolic regulator. Am J
Physiol Endocrinol Metab 294:E995-1010.
Pegg AE, Xiong H, Feith DJ, Shantz LM (1998) S-adenosylmethionine decarboxylase: structure,
function and regulation by polyamines. Biochem Soc Trans 26:580-586.
Pendeville H, Carpino N, Marine JC, Takahashi Y, Muller M, Martial JA, Cleveland JL (2001) The
ornithine decarboxylase gene is essential for cell survival during early murine development. Mol Cell
Biol 21:6549-6558.
Persson L (2009) Polyamine homoeostasis. Essays Biochem 46:11-24.

51
Persson L, Pegg AE (1984) Studies of the induction of spermidine/spermine N1-acetyltransferase
using a specific antiserum. J Biol Chem 259:12364-12367.
Pietila M, Pirinen E, Keskitalo S, Juutinen S, Pasonen-Seppanen S, Keinanen T, Alhonen L, Janne J
(2005) Disturbed keratinocyte differentiation in transgenic mice and organotypic keratinocyte
cultures as a result of spermidine/spermine N-acetyltransferase overexpression. J Invest Dermatol
124:596-601.
Pietila M, Parkkinen JJ, Alhonen L, Janne J (2001) Relation of skin polyamines to the hairless
phenotype in transgenic mice overexpressing spermidine/spermine N-acetyltransferase. J Invest
Dermatol 116:801-805.
Pietila M, Alhonen L, Halmekyto M, Kanter P, Janne J, Porter CW (1997) Activation of polyamine
catabolism profoundly alters tissue polyamine pools and affects hair growth and female fertility in
transgenic mice overexpressing spermidine/spermine N1-acetyltransferase. J Biol Chem 272:18746-
18751.
Pinsky MR (2004) Dysregulation of the immune response in severe sepsis. Am J Med Sci 328:220-
229.
Pirinen E, Gylling H, Itkonen P, Yaluri N, Heikkinen S, Pietila M, Kuulasmaa T, Tusa M, Cerrada-
Gimenez M, Pihlajamaki J, Alhonen L, Janne J, Miettinen TA, Laakso M (2010) Activated
polyamine catabolism leads to low cholesterol levels by enhancing bile acid synthesis. Amino Acids
38:549-560.
Pirinen E, Kuulasmaa T, Pietila M, Heikkinen S, Tusa M, Itkonen P, Boman S, Skommer J,
Virkamaki A, Hohtola E, Kettunen M, Fatrai S, Kansanen E, Koota S, Niiranen K, Parkkinen J,
Levonen AL, Yla-Herttuala S, Hiltunen JK, Alhonen L, Smith U, Janne J, Laakso M (2007)
Enhanced polyamine catabolism alters homeostatic control of white adipose tissue mass, energy
expenditure, and glucose metabolism. Mol Cell Biol 27:4953-4967.
Pittenger MF, Mackay AM, Beck SC, Jaiswal RK, Douglas R, Mosca JD, Moorman MA, Simonetti
DW, Craig S, Marshak DR (1999) Multilineage potential of adult human mesenchymal stem cells.
Science 284:143-147.
Plass C, Oakes C, Blum W, Marcucci G (2008) Epigenetics in acute myeloid leukemia. Semin Oncol
35:378-387.
Pollmann D, Siepmann S, Geppert R, Wernecke KD, Possinger K, Luftner D (2007) The amino-
terminal propeptide (PINP) of type I collagen is a clinically valid indicator of bone turnover and
extent of metastatic spread in osseous metastatic breast cancer. Anticancer Res 27:1853-1862.
Raaijmakers MH, Mukherjee S, Guo S, Zhang S, Kobayashi T, Schoonmaker JA, Ebert BL, Al-
Shahrour F, Hasserjian RP, Scadden EO, Aung Z, Matza M, Merkenschlager M, Lin C, Rommens
JM, Scadden DT (2010) Bone progenitor dysfunction induces myelodysplasia and secondary
leukaemia. Nature 464:852-857.
Radulovic V, de Haan G, Klauke K (2013) Polycomb-group proteins in hematopoietic stem cell
regulation and hematopoietic neoplasms. Leukemia 27:523-533.
Randall TD, Lund FE, Howard MC, Weissman IL (1996) Expression of murine CD38 defines a
population of long-term reconstituting hematopoietic stem cells. Blood 87:4057-4067.

52
Rekhtman N, Choe KS, Matushansky I, Murray S, Stopka T, Skoultchi AI (2003) PU.1 and pRB
interact and cooperate to repress GATA-1 and block erythroid differentiation. Mol Cell Biol
23:7460-7474.
Reddy BS, Watanabe K, Weisburger JH, Wynder EL (1977) Promoting effect of bile acids in colon
carcinogenesis in germ-free and conventional F344 rats. Cancer Res 37:3238-3242.
Rissanen JP, Suominen MI, Peng Z, Morko J, Rasi S, Risteli J, Halleen JM (2008) Short-term
changes in serum PINP predict long-term changes in trabecular bone in the rat ovariectomy model.
Calcif Tissue Int 82:155-161.
Ross EA, Douglas MR, Wong SH, Ross EJ, Curnow SJ, Nash GB, Rainger E, Scheel-Toellner D,
Lord JM, Salmon M, Buckley CD (2006) Interaction between integrin alpha9beta1 and vascular cell
adhesion molecule-1 (VCAM-1) inhibits neutrophil apoptosis. Blood 107:1178-1183.
Rossi DJ, Bryder D, Zahn JM, Ahlenius H, Sonu R, Wagers AJ, Weissman IL (2005) Cell intrinsic
alterations underlie hematopoietic stem cell aging. Proc Natl Acad Sci U S A 102:9194-9199.
Sakata K, Kashiwagi K, Igarashi K (2000) Properties of a polyamine transporter regulated by
antizyme. Biochem J 347 Pt 1:297-303.
Sallmyr A, Fan J, Rassool FV (2008) Genomic instability in myeloid malignancies: increased
reactive oxygen species (ROS), DNA double strand breaks (DSBs) and error-prone repair. Cancer
Lett 270:1-9.
Sardina JL, Lopez-Ruano G, Sanchez-Abarca LI, Perez-Simon JA, Gaztelumendi A, Trigueros C,
Llanillo M, Sanchez-Yague J, Hernandez-Hernandez A (2010) p22phox-dependent NADPH oxidase
activity is required for megakaryocytic differentiation. Cell Death Differ 17:1842-1854.
Schipper RG, Penning LC, Verhofstad AA (2000) Involvement of polyamines in apoptosis. Facts
and controversies: effectors or protectors?. Semin Cancer Biol 10:55-68.
Scholz B, Marschalek R (2012) Epigenetics and blood disorders. Br J Haematol 158:307-322.
Scott EW, Simon MC, Anastasi J, Singh H (1994) Requirement of transcription factor PU.1 in the
development of multiple hematopoietic lineages. Science 265:1573-1577.
Seiler N, Delcros JG, Moulinoux JP (1996) Polyamine transport in mammalian cells. An update. Int J
Biochem Cell Biol 28:843-861.
Seiler N, Graham A, Bartholeyns J (1981) Enhanced urinary excretion of N1-acetylspermidine and
the presence of tumors. Cancer Res 41:1572-1573.
Sodek J, Ganss B, McKee MD (2000) Osteopontin. Crit Rev Oral Biol Med 11:279-303.
Soulet D, Covassin L, Kaouass M, Charest-Gaudreault R, Audette M, Poulin R (2002) Role of
endocytosis in the internalization of spermidine-C(2)-BODIPY, a highly fluorescent probe of
polyamine transport. Biochem J 367:347-357.
Stein GS, Lian JB, van Wijnen AJ, Stein JL, Montecino M, Javed A, Zaidi SK, Young DW, Choi JY,
Pockwinse SM (2004) Runx2 control of organization, assembly and activity of the regulatory
machinery for skeletal gene expression. Oncogene 23:4315-4329.

53
TABOR H (1962) The protective effect of spermine and other polyamines against heat denaturation
of deoxyribonucleic acid. Biochemistry 1:496-501.
Thomas TJ, Messner RP (1986) A left-handed (Z) conformation of poly(dA-dC).poly(dG-dT)
induced by polyamines. Nucleic Acids Res 14:6721-6733.
Tjabringa GS, Zandieh-Doulabi B, Helder MN, Knippenberg M, Wuisman PI, Klein-Nulend J (2008)
The polymine spermine regulates osteogenic differentiation in adipose stem cells. J Cell Mol Med
12:1710-1717.
Tome ME, Fiser SM, Payne CM, Gerner EW (1997) Excess putrescine accumulation inhibits the
formation of modified eukaryotic initiation factor 5A (eIF-5A) and induces apoptosis. Biochem J 328
( Pt 3):847-854.
Tomitori H, Usui T, Saeki N, Ueda S, Kase H, Nishimura K, Kashiwagi K, Igarashi K (2005)
Polyamine oxidase and acrolein as novel biochemical markers for diagnosis of cerebral stroke.
Stroke 36:2609-2613.
Tothova Z, Kollipara R, Huntly BJ, Lee BH, Castrillon DH, Cullen DE, McDowell EP, Lazo-
Kallanian S, Williams IR, Sears C, Armstrong SA, Passegue E, DePinho RA, Gilliland DG (2007)
FoxOs are critical mediators of hematopoietic stem cell resistance to physiologic oxidative stress.
Cell 128:325-339.
Traver D, Miyamoto T, Christensen J, Iwasaki-Arai J, Akashi K, Weissman IL (2001) Fetal liver
myelopoiesis occurs through distinct, prospectively isolatable progenitor subsets. Blood 98:627-635.
Tripathi AK, Chaturvedi R, Ahmad R, Asim M, Sawlani KK, Singh RL, Tekwani BL (2002)
Peripheral blood leucocytes ornithine decarboxylase activity in chronic myeloid leukemia patients:
prognostic and therapeutic implications. Leuk Res 26:349-354.
Tucker JM, Murphy JT, Kisiel N, Diegelman P, Barbour KW, Davis C, Medda M, Alhonen L, Janne
J, Kramer DL, Porter CW, Berger FG (2005) Potent modulation of intestinal tumorigenesis in
Apcmin/+ mice by the polyamine catabolic enzyme spermidine/spermine N1-acetyltransferase.
Cancer Res 65:5390-5398.
Udagawa N, Takahashi N, Akatsu T, Tanaka H, Sasaki T, Nishihara T, Koga T, Martin TJ, Suda T
(1990) Origin of osteoclasts: mature monocytes and macrophages are capable of differentiating into
osteoclasts under a suitable microenvironment prepared by bone marrow-derived stromal cells. Proc
Natl Acad Sci U S A 87:7260-7264.
Uimari A, Keinanen TA, Karppinen A, Woster P, Uimari P, Janne J, Alhonen L (2009) Spermine
analogue-regulated expression of spermidine/spermine N1-acetyltransferase and its effects on
depletion of intracellular polyamine pools in mouse fetal fibroblasts. Biochem J 422:101-109.
Vaananen HK, Horton M (1995) The osteoclast clear zone is a specialized cell-extracellular matrix
adhesion structure. J Cell Sci 108 ( Pt 8):2729-2732.
van Bezooijen RL, Roelen BA, Visser A, van der Wee-Pals L, de Wilt E, Karperien M, Hamersma
H, Papapoulos SE, ten Dijke P, Lowik CW (2004) Sclerostin is an osteocyte-expressed negative
regulator of bone formation, but not a classical BMP antagonist. J Exp Med 199:805-814.
Venezia TA, Merchant AA, Ramos CA, Whitehouse NL, Young AS, Shaw CA, Goodell MA (2004)
Molecular signatures of proliferation and quiescence in hematopoietic stem cells. PLoS Biol 2:e301.

54
Vicente C, Conchillo A, Garcia-Sanchez MA, Odero MD (2012) The role of the GATA2
transcription factor in normal and malignant hematopoiesis. Crit Rev Oncol Hematol 82:1-17.
Visnjic D, Kalajzic Z, Rowe DW, Katavic V, Lorenzo J, Aguila HL (2004) Hematopoiesis is
severely altered in mice with an induced osteoblast deficiency. Blood 103:3258-3264.
Vujcic S, Liang P, Diegelman P, Kramer DL, Porter CW (2003) Genomic identification and
biochemical characterization of the mammalian polyamine oxidase involved in polyamine back-
conversion. Biochem J 370:19-28.
Vujcic S, Diegelman P, Bacchi CJ, Kramer DL, Porter CW (2002) Identification and characterization
of a novel flavin-containing spermine oxidase of mammalian cell origin. Biochem J 367:665-675.
Vujcic S, Halmekyto M, Diegelman P, Gan G, Kramer DL, Janne J, Porter CW (2000) Effects of
conditional overexpression of spermidine/spermine N1-acetyltransferase on polyamine pool
dynamics, cell growth, and sensitivity to polyamine analogs. J Biol Chem 275:38319-38328.
Vuohelainen S, Pirinen E, Cerrada-Gimenez M, Keinanen TA, Uimari A, Pietila M, Khomutov AR,
Janne J, Alhonen L (2010) Spermidine is indispensable in differentiation of 3T3-L1 fibroblasts to
adipocytes. J Cell Mol Med 14:1683-1692.
Walker DG (1975) Control of bone resorption by hematopoietic tissue. The induction and reversal of
congenital osteopetrosis in mice through use of bone marrow and splenic transplants. J Exp Med
142:651-663.
Walkley CR, Olsen GH, Dworkin S, Fabb SA, Swann J, McArthur GA, Westmoreland SV,
Chambon P, Scadden DT, Purton LE (2007) A microenvironment-induced myeloproliferative
syndrome caused by retinoic acid receptor gamma deficiency. Cell 129:1097-1110.
Wallace HM (2009) The polyamines: past, present and future. Essays Biochem 46:1-9.
Wallace HM, Fraser AV, Hughes A (2003) A perspective of polyamine metabolism. Biochem J
376:1-14.
Wallace HM, Duthie J, Evans DM, Lamond S, Nicoll KM, Heys SD (2000) Alterations in polyamine
catabolic enzymes in human breast cancer tissue. Clin Cancer Res 6:3657-3661.
Wang X, Ikeguchi Y, McCloskey DE, Nelson P, Pegg AE (2004) Spermine synthesis is required for
normal viability, growth, and fertility in the mouse. J Biol Chem 279:51370-51375.
Wang Y, Gong B, Dai W, Lu L (1998) Identification of immediate early genes during TPA-induced
human myeloblastic leukemia ML-1 cell differentiation. Gene 216:293-302.
Watanabe S, Kusama-Eguchi K, Kobayashi H, Igarashi K (1991) Estimation of polyamine binding to
macromolecules and ATP in bovine lymphocytes and rat liver. J Biol Chem 266:20803-20809.
Williams K (1997) Interactions of polyamines with ion channels. Biochem J 325 ( Pt 2):289-297.
Woolthuis CM, de Haan G, Huls G (2011) Aging of hematopoietic stem cells: Intrinsic changes or
micro-environmental effects?. Curr Opin Immunol 23:512-517.
Wu H, Min J, Zeng H, McCloskey DE, Ikeguchi Y, Loppnau P, Michael AJ, Pegg AE, Plotnikov AN
(2008) Crystal structure of human spermine synthase: implications of substrate binding and catalytic
mechanism. J Biol Chem 283:16135-16146.

55
Wu H, Min J, Ikeguchi Y, Zeng H, Dong A, Loppnau P, Pegg AE, Plotnikov AN (2007) Structure
and mechanism of spermidine synthases. Biochemistry 46:8331-8339.
Xiao L, Celano P, Mank AR, Griffin C, Jabs EW, Hawkins AL, Casero RA,Jr (1992) Structure of the
human spermidine/spermine N1-acetyltransferase gene (exon/intron gene organization and
localization to Xp22.1). Biochem Biophys Res Commun 187:1493-1502.
Xiong J, O'Brien CA (2012) Osteocyte RANKL: new insights into the control of bone remodeling. J
Bone Miner Res 27:499-505.
Yamamoto T, Hinoi E, Fujita H, Iezaki T, Takahata Y, Takamori M, Yoneda Y (2012) The natural
polyamines spermidine and spermine prevent bone loss through preferential disruption of
osteoclastic activation in ovariectomized mice. Br J Pharmacol 166:1084-1096.
Yamazaki S, Ema H, Karlsson G, Yamaguchi T, Miyoshi H, Shioda S, Taketo MM, Karlsson S,
Iwama A, Nakauchi H (2011) Nonmyelinating Schwann cells maintain hematopoietic stem cell
hibernation in the bone marrow niche. Cell 147:1146-1158.
Zahedi K, Barone S, Kramer DL, Amlal H, Alhonen L, Janne J, Porter CW, Soleimani M (2010) The
role of spermidine/spermine N1-acetyltransferase in endotoxin-induced acute kidney injury. Am J
Physiol Cell Physiol 299:C164-74.
Zhang DE, Zhang P, Wang ND, Hetherington CJ, Darlington GJ, Tenen DG (1997a) Absence of
granulocyte colony-stimulating factor signaling and neutrophil development in CCAAT enhancer
binding protein alpha-deficient mice. Proc Natl Acad Sci U S A 94:569-574.
Zhang J, Niu C, Ye L, Huang H, He X, Tong WG, Ross J, Haug J, Johnson T, Feng JQ, Harris S,
Wiedemann LM, Mishina Y, Li L (2003) Identification of the haematopoietic stem cell niche and
control of the niche size. Nature 425:836-841.
Zhang M, Caragine T, Wang H, Cohen PS, Botchkina G, Soda K, Bianchi M, Ulrich P, Cerami A,
Sherry B, Tracey KJ (1997b) Spermine inhibits proinflammatory cytokine synthesis in human
mononuclear cells: a counterregulatory mechanism that restrains the immune response. J Exp Med
185:1759-1768.
Zhang P, Behre G, Pan J, Iwama A, Wara-Aswapati N, Radomska HS, Auron PE, Tenen DG, Sun Z
(1999) Negative cross-talk between hematopoietic regulators: GATA proteins repress PU.1. Proc
Natl Acad Sci USA 96:8705-8710.
Zhang P, Iwasaki-Arai J, Iwasaki H, Fenyus ML, Dayaram T, Owens BM, Shigematsu H, Levantini
E, Huettner CS, Lekstrom-Himes JA, Akashi K, Tenen DG (2004) Enhancement of hematopoietic
stem cell repopulating capacity and self-renewal in the absence of the transcription factor C/EBP
alpha. Immunity 21:853-863.
Zhao Q, Wang X, Liu Y, He A, Jia R (2010) NFATc1: functions in osteoclasts. Int J Biochem Cell
Biol 42:576-579.
Zhu S, Ashok M, Li J, Li W, Yang H, Wang P, Tracey KJ, Sama AE, Wang H (2009) Spermine
protects mice against lethal sepsis partly by attenuating surrogate inflammatory markers. Mol Med
15:275-282.

Publications of the University of Eastern Finland
Dissertations in Health Sciences
isbn 978-952-61-1321-0
Publications of the University of Eastern FinlandDissertations in Health Sciences
Polyamines are ubiquitious
molecules essential for cell
proliferation. However, the
role of polyamine metabolism
in the immune response,
hematopoiesis, and bone
remodeling is largely unknown
and was thus addressed in this
study. The study showed that
enhanced activity of spermidine/
spermine N1-acetyltransferase
(SSAT), a catabolic regulator of
polyamines, was associated with
disturbed hematopoiesis and bone
remodeling in mice and myeloid
leukemias in humans.
dissertatio
ns | 207 | S
ini P
irn
es-Ka
rh
u | Sperm
idine/spermine N
1-acetyltransferase in Mouse H
ematopoiesis and B
one Rem
odeling...
Sini Pirnes-KarhuSpermidine/spermine
N1-acetyltransferase in Mouse Hematopoiesis
and Bone Remodeling and in Human Leukemias
Sini Pirnes-Karhu
Spermidine/spermine N1-acetyltransferase in Mouse Hematopoiesis and Bone Remodeling and in Human Leukemias





























