Embed Size (px)
Citation preview

Specialized piRNA PathwaysAct in Germline and Somatic Tissuesof the Drosophila OvaryColin D. Malone,1,2,5 Julius Brennecke,1,2,5,6 Monica Dus,1,2,5,7 Alexander Stark,3,4,8 W. Richard McCombie,1
Ravi Sachidanandam,1,9 and Gregory J. Hannon1,2,*1Watson School of Biological Sciences2Howard Hughes Medical Institute
Cold Spring Harbor Laboratory, 1 Bungtown Road, Cold Spring Harbor, NY 11724, USA3Broad Institute of Massachusetts Institute of Technology and Harvard University, Cambridge, MA 02141, USA4Computer Science and Artificial Intelligence Laboratory, Massachusetts Institute of Technology, Cambridge, MA 02139, USA5These authors contributed equally to this work6Present address: Institute of Molecular Biotechnology of the Austrian Academy of Sciences (IMBA), Dr. Bohr-Gasse 3,
A-1030 Vienna, Austria7Present address: Skirball Institute for Biomolecular Medicine, New York University School of Medicine, 540 First Avenue, New York,
NY 10016, USA8Present address: Institute of Molecular Pathology, Dr. Bohr-Gasse 7, 1030 Vienna, Austria9Present address: Department of Genetics and Genomic Sciences, Mount Sinai School of Medicine, 1425 Madison Avenue, New York,
NY 10029, USA
*Correspondence: [email protected] 10.1016/j.cell.2009.03.040
SUMMARY
In Drosophila gonads, Piwi proteins and associatedpiRNAs collaborate with additional factors to forma small RNA-based immune system that silencesmobile elements. Here, we analyzed nine DrosophilapiRNA pathway mutants for their impacts on bothsmall RNA populations and the subcellular localiza-tion patterns of Piwi proteins. We find that distinctpiRNA pathways with differing components functionin ovarian germ and somatic cells. In the soma, Piwiacts singularly with the conserved flamenco piRNAcluster to enforce silencing of retroviral elementsthat may propagate by infecting neighboring germcells. In the germline, silencing programs encodedwithin piRNA clusters are optimized via a slicer-dependent amplification loop to suppress a broadspectrum of elements. The classes of transposonstargeted by germline and somatic piRNA clusters,though not the precise elements, are conservedamong Drosophilids, demonstrating that the archi-tecture of piRNA clusters has coevolved with thetransposons that they are tasked to control.
INTRODUCTION
Eukaryotic genomes harbor a wide variety of transposons,
whose maintenance and spread throughout the population
requires the colonization of new genomic locations in germ cells.
For the host, the deleterious consequences of transposon pro-
522 Cell 137, 522–535, May 1, 2009 ª2009 Elsevier Inc.
pagation range from insertional mutagenesis and reductions in
the long-term fitness of their progeny to an acute loss of germ
cell integrity and sterility.
Transposable elements can be broadly categorized as retro-
transposons (class I), which move via an RNA intermediate, or
DNA transposons (class II), which mobilize through a ‘‘cut-and-
paste’’ mechanism (Slotkin and Martienssen, 2007). Transpo-
sons within these classes differ in their structure, evolutionary
origins, and both their tissue and developmental expression
patterns. Many transposons are expressed in germ cells, where
movement can lead to heritable expansions in their number.
Examples in Drosophila include TAHRE, TART, HetA, copia,
and the I element (Brennecke et al., 2008; Chambeyron et al.,
2008; Shpiz et al., 2009; Shpiz et al., 2007; Vagin et al., 2004).
Some transposons are exclusively or additionally expressed in
somatic cells of the ovary, with gypsy, ZAM, and idefix occupying
this category in the Drosophila ovary (Desset et al., 2008; Desset
et al., 2003; Mevel-Ninio et al., 2007; Pelisson et al., 2007;
Prud’homme et al., 1995; Sarot et al., 2004). The diversity of
transposition strategies and the overall similarities to host
protein-coding genes pose a substantial challenge to their selec-
tive silencing (Malone and Hannon, 2009).
In animals, suppression of mobile elements is accomplished by
an elegant, small RNA-based immune system, which displays
both genetically encoded and adaptive aspects. Its core compo-
nents are Piwi family proteins, and their associated Piwi-interact-
ing RNAs (piRNAs). Like other members of the Argonaute family,
Piwi proteins use bound small RNAs as guides for substrate
recognition and target cleavage (Carmell et al., 2002). In
Drosophila, the major sources of piRNAs are discrete heterochro-
matic loci termed piRNA clusters (Supplemental Glossary avail-
able online) (Brennecke et al., 2007). These are characterized

by an exceptional density of nested, fragmented, and immobi-
lized transposon remnants. Thus, the generation of piRNAs
from these loci inherently targets the three Drosophila Piwi
proteins, Piwi, Aub, and AGO3, toward mobile elements.
Most piRNA clusters contain transposon fragments in sense
and antisense orientations and produce piRNAs from both
genomic strands. Nevertheless, piRNAs overall tend to be anti-
sense to transposons (Brennecke et al., 2007). Piwi- and Aub-
associated piRNAs reflect the antisense bias of the system,
whereas AGO3-bound piRNAs are typically sense to transpo-
sons. Sense and antisense piRNAs bound by AGO3 and Aub,
respectively, show a prevalent relationship with their 50 ends,
overlapping by precisely 10 nt.
These observations coalesced into a model in which Piwi
proteins engage in a Slicer-dependent amplification loop (the
ping-pong cycle—Supplemental Glossary) between piRNA clus-
ters and active elements (Brennecke et al., 2007; Gunawardane
et al., 2007). Cleavage of a transposon transcript by Aub, loaded
with an antisense piRNA, triggers production of an AGO3-bound
sense piRNA, whose 50 end is offset by 10 nt. The AGO3-bound
piRNA can then catalyze the production of more silencing-
competent piRNAs, which associate with Aub, via cleavage of
antisense transposon sequences within cluster transcripts. Over-
all, the ping-pong cycle optimizes the piRNA response against
transposons active in a given cell and at a given developmental
time point. Signatures of the ping-pong cycle are conserved
throughout animals, suggesting that it is a fundamental property
of the piRNA pathway (Aravin et al., 2007b; Houwing et al., 2007;
Murchison et al., 2008).
Though the majority of Aub- and AGO3-bound piRNAs appear
to be generated via the ping-pong cycle, only a small proportion
of Piwi-bound piRNAs display ping-pong signatures. Yet, Piwi-
bound piRNAs still exhibit a strong antisense bias. This has led
to the concept of primary piRNA biogenesis, wherein Piwi acts
as a possible recipient of cluster-derived piRNAs that are gener-
ated via yet unknown mechanisms.
Primary piRNAs have been proposed as one initiator of the
ping-pong cycle. However, a recent study has also highlighted
the importance of maternally inherited piRNA populations (Bren-
necke et al., 2008). For two transposons that were examined, the
I and P elements, the lack of a maternal piRNA program pre-
vented silencing in progeny, and this was associated with the
lack of a robust ping-pong response. Thus, primary and mater-
nally deposited piRNAs serve as inputs into the pathway, which
initiate a cycle of interactions between piRNA clusters and trans-
poson mRNAs.
We sought to determine whether this model applied univer-
sally, not only in germ cells but also in somatic support cells
wherein a subset of transposons are regulated by Piwi, the sole
family member expressed in this compartment. By comparing
germline-specific piRNA populations to those derived from whole
ovaries, we show that a distinct, ping-pong-independent piRNA
pathway operates in somatic cells. Analysis of piRNA profiles
from mutant ovaries strongly supports this model and indicates
that the somatic pathway depends exclusively upon Piwi and
the flamenco piRNA cluster. We also probed the roles of addi-
tional factors within the piRNA pathway, examining the impacts
of nine such mutants on piRNA populations, on the operation of
the pathway, and on the localization of pathway components in
germ and somatic cells. We find that Piwi function in the germline
depends on the RNA helicase Armitage. The ping-pong cycle
acts independently of Piwi and Armitage but requires the function
of Aubergine, the RNA helicases Spindle-E and Vasa, and the
Tudor-domain protein Krimper. Through these studies, we begin
to assemble a scaffold model of the piRNA pathway, which differs
substantially in the germline and somatic compartments of the
ovary.
RESULTS
Transposons Display Tissue-SpecificControl MechanismsIn the Drosophila ovary, all three Piwi-family members (Piwi, Aub,
and AGO3) are expressed ingermline cells (Brennecke etal., 2007;
Coxetal., 2000; Gunawardaneet al., 2007;Harris andMacdonald,
2001; Saito et al., 2006), while Piwi alone is expressed in gonadal
somatic cells. This implied possible differences in the architecture
of the piRNA pathway,and perhaps the elements that it controls, in
germline and somatic tissues. We therefore sought to separately
analyze piRNAs present in these two compartments.
Germline cells within the Drosophila egg chamber are syncy-
tial, and nearly all of the cytoplasmic contents of nurse cells
are incorporated into late-stage oocytes (Spradling, 1993). In
contrast, the follicular epithelium is shed from the laid egg.
Thus, we could infer somatic and germline piRNA pools by
comparing small RNA libraries derived from wild-type ovaries
to those from 0–2 hr old embryos, prior to the activation of the
zygotic genome (Brennecke et al., 2008).
piRNAs (see Figure S1 available online) were mapped to the
Repbase collection of known Drosophila melanogaster elements
(allowing up to three mismatches) (Supplemental Experimental
Procedures). We chose to focus on the 86 elements most heavily
targeted by the piRNA pathway. This corresponds to 75% of all
elements and includes �99% of all transposon-derived piRNAs.
piRNAs were also assigned to their generative clusters, including
only those small RNAs mapping unambiguously to a single site
within the Drosophila genome. Our key question was the extent
to which piRNAs present in the mixed ovarian sample were
maternally deposited.
For the majority of transposons, the piRNA content of early
embryos mirrored that of total ovary (Figure 1A). As exemplified
by roo and the F element, not only the overall abundance but
also the distribution of ovarian piRNAs targeting each transposon
was faithfully retained inearlyembryos (Figure 1A, right).However,
piRNAs targeting a number of transposons (e.g., ZAM and gypsy;
Figure 1A, right; Figure S2A) were substantially underrepresented
in the embryonic piRNA pool. The transposons targeted by these
small RNAs are likely subject to selective control by the piRNA
system in somatic cells.
Elements with piRNA patterns characteristic of somatic
control populate the gypsy family of long terminal repeat (LTR)
retrotransposons. Among these are ZAM, gypsy, and idefix, all
of which are regulated by the flamenco piRNA cluster. Across
the entire spectrum of transposons, a lack of maternally depos-
ited piRNAs correlated strongly with the presence of corre-
sponding transposon fragments in flamenco (Figure 1A, left). In
Cell 137, 522–535, May 1, 2009 ª2009 Elsevier Inc. 523
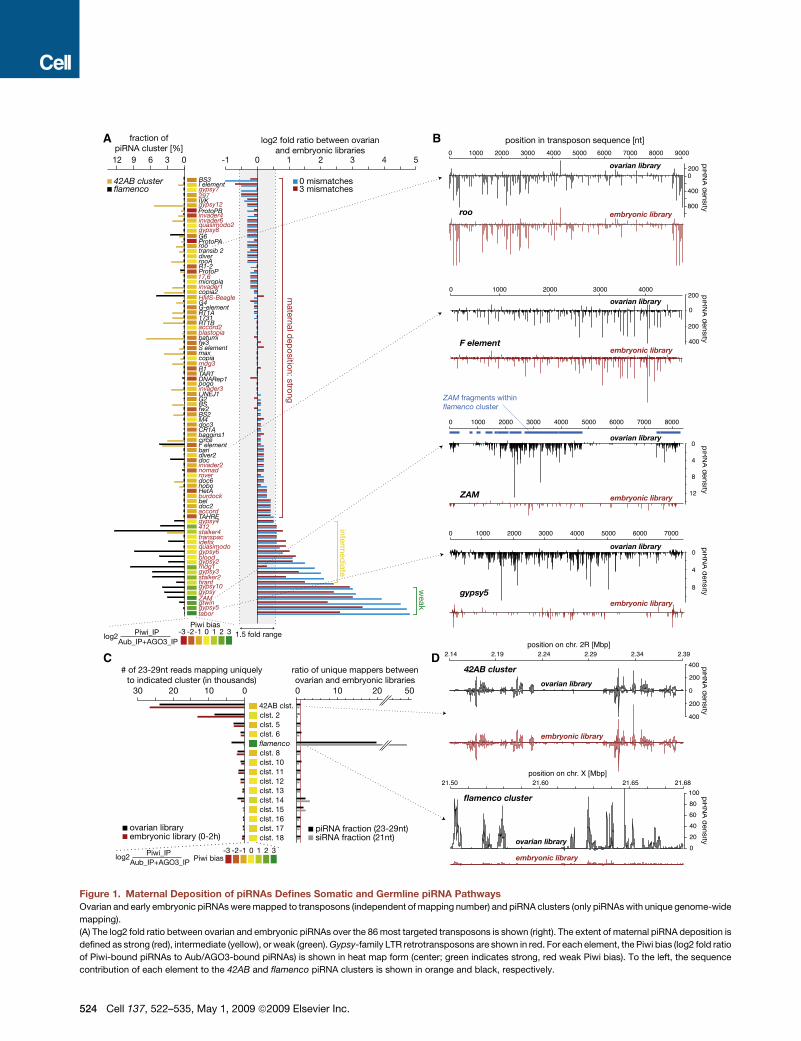
Figure 1. Maternal Deposition of piRNAs Defines Somatic and Germline piRNA Pathways
Ovarian and early embryonic piRNAs were mapped to transposons (independent of mapping number) and piRNA clusters (only piRNAs with unique genome-wide
mapping).
(A) The log2 fold ratio between ovarian and embryonic piRNAs over the 86 most targeted transposons is shown (right). The extent of maternal piRNA deposition is
defined as strong (red), intermediate (yellow), or weak (green). Gypsy-family LTR retrotransposons are shown in red. For each element, the Piwi bias (log2 fold ratio
of Piwi-bound piRNAs to Aub/AGO3-bound piRNAs) is shown in heat map form (center; green indicates strong, red weak Piwi bias). To the left, the sequence
contribution of each element to the 42AB and flamenco piRNA clusters is shown in orange and black, respectively.
524 Cell 137, 522–535, May 1, 2009 ª2009 Elsevier Inc.

contrast, elements with fragments lying within other piRNA clus-
ters (e.g., the cluster at cytological position 42AB; Figure 1A, left)
contribute piRNAs to both ovarian and early embryonic libraries,
indicating active control in germline cells.
These observations suggested tissue-specific regulation of
certain element classes. This correlated with tissue-specific
expression of piRNA clusters. Small RNAs derived from flamenco
were highly depleted from early embryonic populations, irrespec-
tive of whether piRNA or siRNA pools were analyzed (Figure 1C).
All remaining major clusters showed relatively equivalent contri-
bution to ovary and embryo libraries. As with individual elements,
the relative pattern of piRNAs mapping to clusters apparently
expressed in germ cells was mirrored in embryo libraries
(Figure 1D, right).
The overall degree to which transposons display Piwi-biased
association (Piwi bias—Supplemental Glossary) strongly corre-
lated with lower representation in maternally deposited small
RNA populations (Figures 1A, 1B, and S2A). Also, while the
majority of piRNA clusters load small RNAs into all three Piwi
family members, flamenco-derived piRNAs almost exclusively
occupy Piwi complexes (Figures 1C, 1D, and S2B).
These data suggest the existence of two separate piRNA path-
ways in germline and somatic cells of the gonad. In the soma,
Piwi appears to be programmed exclusively or predominantly
by the flamenco cluster to target elements from the gypsy family.
In the germline, a variety of clusters collaborate with all three
Piwi-clade proteins to control a broad range of elements and
to contribute a heritable collection of piRNAs that maintains
resistance across generations.
The Ping-Pong Cycle Is Germ Cell SpecificPing-pong constitutes a feed-forward loop that optimizes the
piRNA response against elements active in a given strain and
simultaneously creates characteristic relationships between
small RNAs that reveal their participation in the cycle (Brennecke
et al., 2007; Gunawardane et al., 2007). The strongest ping-pong
interactions are found between sense-oriented piRNAs in AGO3
and antisense piRNAs in Aub (Brennecke et al., 2007). In contrast,
the participation of Piwi in the ping-pong cycle is less obvious. We
therefore probed the degree to which individual transposons
participate in the ping-pong cycle and correlated this with their
bias toward control by individual Piwi proteins.
For each transposon, we quantified its ping-pong signature
(Supplemental Experimental Procedures). In short, this was
defined as the likelihood, in percent, for the average piRNA
mapping to an element to have a complementary ping-pong
partner. If plotted against the degree to which an element has
corresponding piRNAs in Piwi versus Aub/AGO3 complexes,
we find that Piwi-biased elements show no significant evidence
of ping-pong (Figure 2A).
To probe the correlation between ping-pong signatures and
maternal inheritance (Supplemental Glossary), we divided trans-
posons into three groups (Figures 1A and 2A). Those with strong
maternal deposition (red) are considered to have a dominant
germline silencing component. Those with intermediate levels
of maternal deposition (yellow) are considered to be expressed
and targeted in both germline and follicle cells, while those with
weak maternal deposition (green) are defined as having a domi-
nant somatic silencing component (Supplemental Glossary).
Strong ping-pong signatures correlate with germline silencing
(Figure 2A). Somatically silenced elements, such as gypsy5, show
no enrichment for ping-pong pairs (Figure 2B), while elements
such as idefix with mixed germline and somatic silencing show
a weak but evident ping-pong signature. Predominantly germline
elements such as ProtoP-B and F element show strong ping-
pong signals (Figure 2B). For F element, idefix, and gypsy5, we
plotted the distribution of piRNAs along each element consensus
and then split the total population into piRNAs with an identified
ping-pong partner and those without (Figure 2C). While piRNAs
that have a ping-pong partner do show an overall antisense
bias, this is much more pronounced for piRNAs that appear to
arise via primary biogenesis. While an understanding is emerging
for how the antisense bias is created for gypsy5 (see below), we
still cannot explain how strand information for germline elements
is incorporated into the pathway as a whole.
Although somatic Piwi lacks detectable ping-pong activity, we
could not rule out roles for Piwi in the ping-pong cycle of germline
elements. We therefore compared the impact of mutations in aub
and piwi on ping-pong signatures. In a panel of 21 representative
transposons with dominant germline expression, loss of Piwi
showed no significant impact on ping-pong signals, while loss
of Aub essentially ablated the cycle (Figure 2D). Elements tar-
geted in somatic cells fail to enter the cycle (Figure 2D, right).
Elements with apparently mixed expression patterns lose their
ping-pong signatures in aub mutants but often show elevated
signals in piwi mutants. This suggests that piwi and aub muta-
tions impact different piRNA populations and that the germ-
line-specific ping-pong cycle operates independently of Piwi.
Piwi resides in the nuclei of both germ and follicle cells (Cox
et al., 2000). Aub and AGO3 concentrate in nuage, perinuclear
RNP granules characteristic of germ cells (Figures 2E and S3)
(Brennecke et al., 2007; Gunawardane et al., 2007; Harris and
Macdonald, 2001). This has led to the speculation that that the
ping-pong cycle might operate in nuage (Klattenhoff and Theur-
kauf, 2008). Loss of Aub leads to delocalization of AGO3 from
nuage and to its accumulation in discrete cytoplasmic foci,
a pattern not seen in piwi mutant germ cells (Figure 2E). These
results underscore the link between Aub and AGO3 in germ cells
and support a role for nuage in the ping-pong cycle.
Flamenco Programs the Somatic piRNA PathwayMany features distinguish flamenco from other generative loci.
The flamenco locus shows an extreme orientation bias of the
elements it harbors (Figure 1B), and unlike clusters expressed
(B) Ovarian (black) and embryonic (red) piRNAs were plotted over elements with strong (roo and F element) or weak (ZAM and gyspy 5) maternal piRNA deposition
(identical y axes). For ZAM, a strong correlation between piRNA density and sequence fragments present in the flamenco cluster (blue) was found.
(C) Total number (left) and ratio (right) of ovarian and embryonic piRNAs mapping to the 15 major piRNA clusters is shown. The corresponding Piwi-bias heat map
is shown for each cluster as in (A).
(D) piRNA densities over the 42AB and flamenco piRNA clusters (identical y axes) are shown.
Cell 137, 522–535, May 1, 2009 ª2009 Elsevier Inc. 525

Figure 2. Transposons Segregate into Distinct Regulatory Classes
Transposons segregated by maternal deposition and Piwi bias display differential levels of ping-pong amplification. (A)–(C) display piRNAs from the wK strain.
(A) Ping-pong signal and Piwi bias of transposons with strong (red), intermediate (yellow), or weak (green) maternal piRNA deposition are displayed as a scatter
plot (0 MM, zero mismatches).
(B) Depiction of the ping-pong signature for indicated transposons. Graphs indicate the likelihood (in percent) that a complementary piRNA exists with a 50 end at
the indicated distance (x axis) for the average piRNA mapping to a particular transposon. The ping-pong signal was defined as the value at position 10 nt.
(C) piRNA densities over the indicated transposons are shown in black. Those are split into piRNAs with (red) and piRNAs without (green) a sequenced ping-pong
partner.
(D) Ping-pong signals (value at 10 nt in [B]) are displayed as heat maps for aub and piwi heterozygote (+/�) and mutant (�/�) libraries.
(E) Aub, AGO3, and Piwi protein localization in wild-type, aubaubQC42/HN2, and piwi1/2 mutant ovaries. We note some variability in piwi mutants due to their
aberrant morphology.
526 Cell 137, 522–535, May 1, 2009 ª2009 Elsevier Inc.

Figure 3. Evolutionary Conservation of the flamenco piRNA Cluster
Transposon composition and chromosomal organization of the somatic flamenco piRNA cluster.
(A) Schematic of the Drosophila melanogaster X chromosome with the flamenco cluster enlarged. Below, the transposon annotation at the flamenco loci in two
other Drosophilid species is shown. Uniquely mapping D. erecta piRNAs are plotted over the putative flamenco cluster (bottom).
(B) The transposon makeup of the 42AB, flamenco, and putative flamenco clusters are displayed. Pie charts display transposon orientation percentages. All
graphs display the percentage of total annotated Repbase transposons in the cluster. Known and putative errantiviruses are indicated by black and gray dots.
in germ cells, this bias seems to have been evolutionarily
hardwired. flamenco comprises �180 KB of pericentromeric
heterochromatin on the X chromosome, in which the majority of
transposon fragments (�85%) are similarly oriented (Figure 3A).
Moreover, flamenco-derived piRNAs are produced exclusively
from the plus strand of the genome, indicating transcription
from the DIP1 gene toward the centromere. Analysis of P element
insertions suggests that flamenco generates a long, continuously
transcribed precursor, which is converted into a preponderance
of antisense piRNAs by primary processing (Figure S4).
Cell 137, 522–535, May 1, 2009 ª2009 Elsevier Inc. 527
In D. yakuba and D. erecta (Drosophila 12 Genomes Consor-
tium et al., 2007), putative flamenco loci could be identified via
their proximity to DIP1, and genomic assemblies were suffi-
ciently complete to allow informative analysis (Figure 3A). In
both cases, the DIP1-proximal region was enriched in trans-
poson fragments with a consistent genomic orientation, such
that transcription from DIP1 across these loci would produce
antisense transposon information (Figure 3A). To confirm that
these syntenic regions actually represent functional piRNA clus-
ters, we sequenced a small RNA library from D. erecta ovaries.
Abundant, uniquely mapping species could be assigned to the
putative D. erecta flamenco locus, and as in D. melanogaster,
they were derived from only one genomic strand (Figure 3A).
Moreover, these RNAs showed no substantial evidence of an
active amplification cycle (1U/10A partners with a 10 nt, 50 over-
lap) (data not shown). Thus, natural selection seems to have
shaped the flamenco clusters of Drosophilids to encode anti-
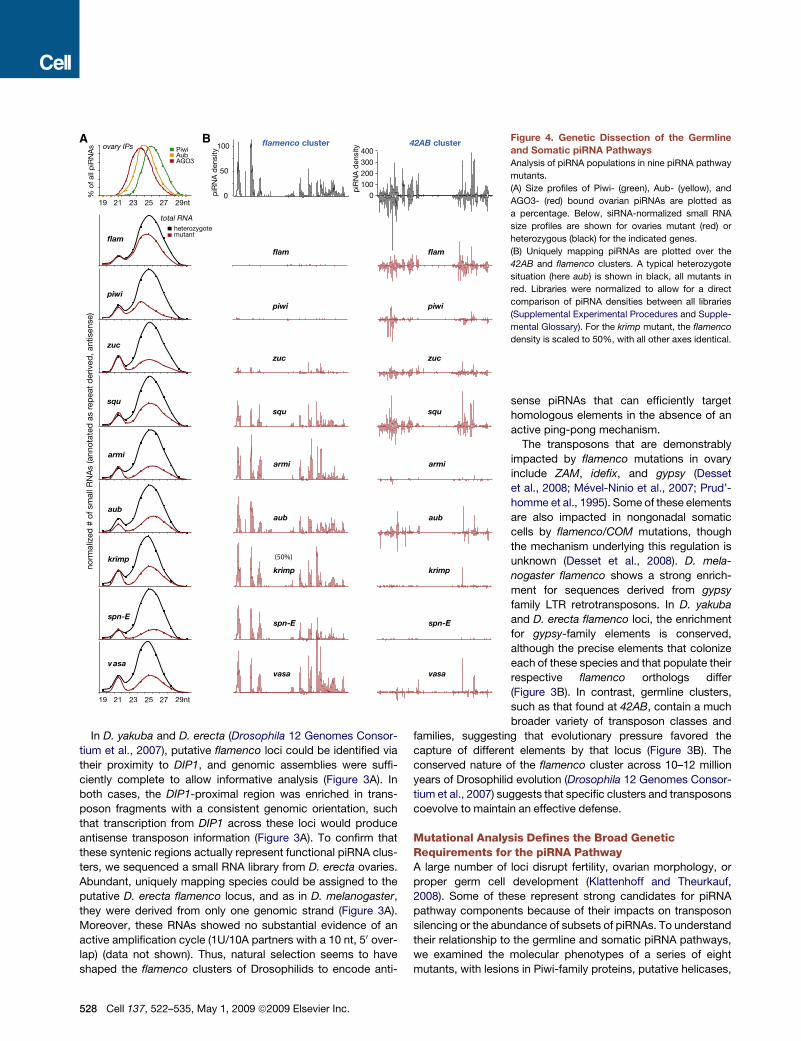
Figure 4. Genetic Dissection of the Germline
and Somatic piRNA Pathways
Analysis of piRNA populations in nine piRNA pathway
mutants.
(A) Size profiles of Piwi- (green), Aub- (yellow), and
AGO3- (red) bound ovarian piRNAs are plotted as
a percentage. Below, siRNA-normalized small RNA
size profiles are shown for ovaries mutant (red) or
heterozygous (black) for the indicated genes.
(B) Uniquely mapping piRNAs are plotted over the
42AB and flamenco clusters. A typical heterozygote
situation (here aub) is shown in black, all mutants in
red. Libraries were normalized to allow for a direct
comparison of piRNA densities between all libraries
(Supplemental Experimental Procedures and Supple-
mental Glossary). For the krimp mutant, the flamenco
density is scaled to 50%, with all other axes identical.
sense piRNAs that can efficiently target
homologous elements in the absence of an
active ping-pong mechanism.
The transposons that are demonstrably
impacted by flamenco mutations in ovary
include ZAM, idefix, and gypsy (Desset
et al., 2008; Mevel-Ninio et al., 2007; Prud’-
homme et al., 1995). Some of these elements
are also impacted in nongonadal somatic
cells by flamenco/COM mutations, though
the mechanism underlying this regulation is
unknown (Desset et al., 2008). D. mela-
nogaster flamenco shows a strong enrich-
ment for sequences derived from gypsy
family LTR retrotransposons. In D. yakuba
and D. erecta flamenco loci, the enrichment
for gypsy-family elements is conserved,
although the precise elements that colonize
each of these species and that populate their
respective flamenco orthologs differ
(Figure 3B). In contrast, germline clusters,
such as that found at 42AB, contain a much
broader variety of transposon classes and
families, suggesting that evolutionary pressure favored the
capture of different elements by that locus (Figure 3B). The
conserved nature of the flamenco cluster across 10–12 million
years of Drosophilid evolution (Drosophila 12 Genomes Consor-
tium et al., 2007) suggests that specific clusters and transposons
coevolve to maintain an effective defense.
Mutational Analysis Defines the Broad GeneticRequirements for the piRNA PathwayA large number of loci disrupt fertility, ovarian morphology, or
proper germ cell development (Klattenhoff and Theurkauf,
2008). Some of these represent strong candidates for piRNA
pathway components because of their impacts on transposon
silencing or the abundance of subsets of piRNAs. To understand
their relationship to the germline and somatic piRNA pathways,
we examined the molecular phenotypes of a series of eight
mutants, with lesions in Piwi-family proteins, putative helicases,
528 Cell 137, 522–535, May 1, 2009 ª2009 Elsevier Inc.

Table 1. Overview of the Effect of Mutations on the piRNA Pathway
piRNA Loss Piwi Protein Localization:
Mutant Function Expression S GL Aubergine AGO3 Piwi Ping-Pong piRNA Size
flamenco piRNA cluster S +++ +/� nuage nuage nuclear functional YY
piwi Piwi family protein S, GL +++ ++ nuage nuage n.d. functional YYY
zucchini putative nuclease unknown ++ ++ nuage nuage nuclear functional Y
squash putative nuclease unknown +/� + nuage nuage nuclear functional /
armitage RNA helicase GL +/� ++ nuage nuage lost in GL functional [
aubergine Piwi family protein GL +/� ++ n.d. dispersed nuclear very weak [[
krimper Tudor-domain
containing
GL +/� +++ dispersed dispersed nuclear weak [[
spindle-E RNA helicase;
Tudor domain
GL +/� +++ dispersed dispersed low in GL very weak [[
vasa RNA helicase GL +/� ++ dispersed dispersed low in GL very weak [[
S, somatic cells; GL, germline; +++, strong; ++, intermediate; +, weak; +/�, none; n.d., not detectable; [, increase (in size); Y, decrease (in size); /, no
change (in size).
nucleases, and proteins of unknown biochemical function (Klat-
tenhoff and Theurkauf, 2008). For reference, we compared these
to a line mutant for the somatic piRNA cluster flamenco.
We isolatedand sequenced small RNAs (18–29nt in length) from
ovaries of age-matched flies, mutant for the genes shown in
Figure 4 and Table 1. To avoid confounding our analysis because
of interstrain variability in transposon content, we compared each
mutant to its heterozygous siblings. Small RNA libraries were
normalized using a subset of endogenous, AGO2-bound siRNAs
as a reference (Tables S1 and S2, Supplemental Experimental
Procedures, and Supplemental Glossary).
An examination of total piRNA levels and of piRNAs mapping to
the major germline and somatic piRNA clusters revealed strong
impacts on various aspects of the piRNA pathway for every
mutant examined (Figures 4 and S4–S10, Tables 1 and S3–S6,
Supplemental Experimental Procedures). Loss of piwi caused
a substantial reduction in overall piRNA populations and also
resulted in a shift in the overall size of the population to that
more characteristic of Aub/AGO3-bound populations (Figure 4A).
piwi loss virtually eliminated piRNAs mapping uniquely to the
flamenco locus, consistent with the hypothesis that it is the sole
family member that acts with this cluster in the somatic pathway.
piwi also impacted piRNA production from the germline 42AB
cluster, suggesting that it plays an important role in this compart-
ment as well (Figure 4B). Mutation of the flamenco locus itself
slightly reduced overall piRNA levels and shifted their average
size, consistent with this somatic cluster contributing a substan-
tial fraction of ovarian, Piwi-bound piRNAs (Figure 4A, Table 1).
As expected, it had no impact on the production of piRNAs
from other piRNA clusters such as 42AB (Figures 4B and S5).
Loss of aubergine strongly impacted overall piRNA popula-
tions, this time shifting their average size more toward that
characteristic of Piwi-bound species (Figure 4A). flamenco was
virtually untouched by the aub mutation. While piRNA production
from 42AB was affected to a degree similar to that seen in piwi
mutants, the subset of piRNAs that remain were quite different
(Figure 4B). 42AB piRNAs remaining in the piwi mutant are Aub-
sized and robustly participate in ping-pong, while in the aub
mutant, 42AB piRNAs are Piwi-sized and display no ping-pong
signatures (data not shown).
A number of mutations shared aubergine’s strong impact on
the germline 42AB cluster with minimal impact on the somatic
piRNA pathway. Loss of spn-E (Gillespie and Berg, 1995), vasa
(Liang et al., 1994; Styhler et al., 1998) (Figure S11), krimp (Barbosa
et al., 2007; Lim and Kai, 2007), and armi (Cook et al., 2004; Tomari
et al., 2004) left the output of piRNAs from flamenco intact while
suppressing, to varying degrees, piRNAs uniquely assignable to
42AB (Figure4B,Table1).With theexceptionof armi�/�, all of these
mutants strongly impacted the ping-pong cycle (FigureS8, Table 1)
with a concomitant delocalization of Aub and AGO3 from nuage
(Figures S9 and S10). Moreover, the average size of piRNAs in
each mutant shifted toward that characteristic of Piwi, consistent
with an impact on Aub/AGO3-bound populations (Figure 4A).
zucchini and squash were identified in a screen for female-
sterile mutants (Schupbach and Wieschaus, 1991). Both were
previously shown to impact the levels of a few abundant piRNAs
by northern blotting (Pane et al., 2007). Mutations in zuc reduced
somatic and germline piRNA pathways, though both systems
still produce some piRNAs in mutant animals. squash shows
the least severe impact of all the mutants examined. Substantial
production of piRNAs from flamenco and 42AB persist in squash
mutants but overall piRNA levels do fall detectably.
The impacts of mutations in each of the genes we examined
correlated largely with the expression patterns of the proteins,
which they encoded (Table 1). Piwi appears in the nuclei of both
germ and somatic cells, while Aubergine, like AGO3, is restricted
to germ cells. Vasa is only expressed in germ cells, and similarly
restricted expression is seen for Krimper, Spindle-E (M.D. and
G.J.H., unpublished data), and Armitage (Cook et al., 2004; Gil-
lespie and Berg, 1995; Lim and Kai, 2007). Expression patterns
of Zucchini and Squash are unknown.
spindle-E and flamenco Mutations Define Propertiesof the Germline and Somatic piRNA PathwaysSpindle-E (spn-E) is a putative DExH-box RNA helicase that is
critical for silencing of transposons in the Drosophila germline
Cell 137, 522–535, May 1, 2009 ª2009 Elsevier Inc. 529
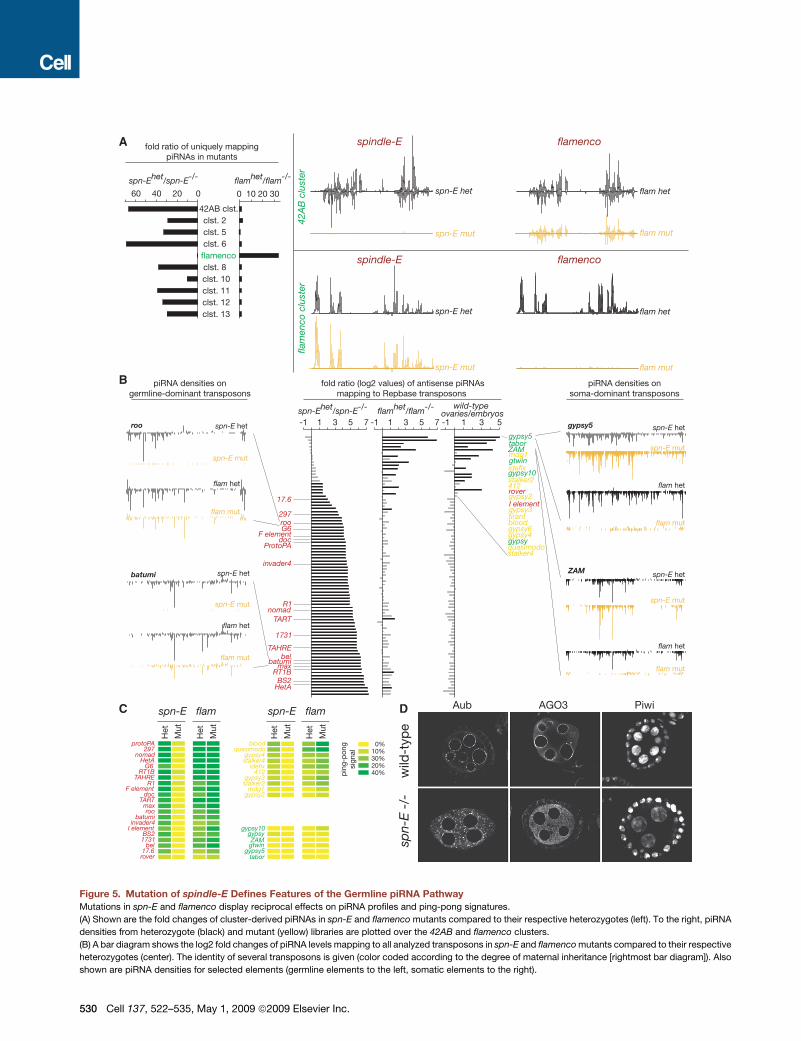
Figure 5. Mutation of spindle-E Defines Features of the Germline piRNA Pathway
Mutations in spn-E and flamenco display reciprocal effects on piRNA profiles and ping-pong signatures.
(A) Shown are the fold changes of cluster-derived piRNAs in spn-E and flamenco mutants compared to their respective heterozygotes (left). To the right, piRNA
densities from heterozygote (black) and mutant (yellow) libraries are plotted over the 42AB and flamenco clusters.
(B) A bar diagram shows the log2 fold changes of piRNA levels mapping to all analyzed transposons in spn-E and flamenco mutants compared to their respective
heterozygotes (center). The identity of several transposons is given (color coded according to the degree of maternal inheritance [rightmost bar diagram]). Also
shown are piRNA densities for selected elements (germline elements to the left, somatic elements to the right).
530 Cell 137, 522–535, May 1, 2009 ª2009 Elsevier Inc.

(Aravin et al., 2004; Aravin et al., 2001; Gillespie and Berg, 1995;
Kennerdell et al., 2002; Klenov et al., 2007; Vagin et al., 2006). Its
strong impact on the 42AB cluster extended to every other
prominent piRNA cluster, save flamenco (Figure 5A). Impacts
on the germline piRNA pathway were considerably stronger
than those observed for aub mutants, suggesting that spn-E
also affects piRNA populations bound by AGO3 and Piwi
(Figure 4). In fact, in spn-E mutant ovaries (and those lacking
other germline components), we consistently detected lower
levels of Piwi in germline nuclei as compared to their heterozy-
gote siblings (Figure S9). Thus, spn-E and flamenco mutants
can be considered as archetypes for loss of germline and
somatic pathways, respectively, allowing us to compare the
participation of individual transposons in each system.
Elements were sorted according to the fold change in corre-
sponding piRNAs in spn-E mutants, as compared to their hetero-
zygous siblings. Only a few elements responded to loss of
flamenco but not spn-E, and these almost perfectly overlapped
with those that showed predominantly somatic control and
a lack of maternal deposition of corresponding piRNAs (Fig-
ure 5B, right). In contrast, the vast majority of elements
responded strongly to the spn-E mutation (Figure 5B). These
included a wide variety of class I and class II elements,
comprising the majority of Drosophila transposons and families
(Figure 5B). Examples are roo and batumi. Both are unaffected
in flamenco mutants but lose essentially all corresponding piR-
NAs in spn-E mutants (Figure 5B). Virtually all of the elements,
which respond to spn-E, showed a strong maternal deposition
of homologous piRNAs, indicative of germline control. A few
elements were impacted by both flamenco and spn-E mutations,
consistent with the proposal that some elements are regulated in
both somatic and germ cells.
Since the ping-pong cycle operates only in germ cells, it was
not surprising that flamenco mutations show little impact on
ping-pong signatures for elements that depend heavily on the
germline pathway or that are strongly restricted to the soma
(Figure 5C, elements designated in red and green, respectively).
For elements that appear to be regulated by both germ cell and
somatic pathways, flamenco mutations actually increased the
proportion of piRNAs with a ping-pong signature. This is expected
since the relative contribution from the somatic pathway would
havebeen lost inflamencomutants.Forall elementswithadetect-
able ping-pong signal, spn-E mutations greatly reduced or elimi-
nated detectable partners (Figure 5C).
Mutations in other components of the germ cell pathway
impacted both piRNA production and ping-pong signatures simi-
larly to spn-E. For example, loss of the Tudor-domain protein
Krimper had very pronounced impacts on piRNA levels and
ping-pong signatures. Ovaries mutant for the RNA helicase
Vasa resembled aubergine mutants in that both strongly affected
the ping-pong cycle, though the impacts on piRNA levels on the
germline specific elements was less dramatic than in spn-E and
krimp mutants (Figure S6 and S7).
Mutations, which disrupted the ping-pong cycle, shared the
impact of aub lesions on the localization of piRNA pathway
components to nuage. spn-E mutant cells have altered Aub and
AGO3 staining, while maintaining the characteristic localization
of Piwi in the nuclei of both somatic and germ cells (Figure 5D).
Similar impacts are seen for mutations in the germline pathway
components krimp and vasa (Figure S9). Mutations in zuc or
squ had no discernable impact on the localization of any Piwi-
family protein, despite the impact of the former on the production
of piRNAs in both the soma and germline (Figure S10). Notably,
these mutations also had no (zuc) or a relatively mild (squ) impact
on ping-pong signatures (Figure S8).
Armitage Is Important for Piwi Function in the GermlineDespite the lack of a role for Piwi in the ping-pong cycle, several
germline transposons exhibit a strong loss of piRNAs in piwi
mutant animals. To probe the underlying basis of these effects,
we wished to examine a situation in which the function of Piwi
was specifically impaired in germ cells. We noted that mutations
in armitage caused a loss of Piwi from germ cell nuclei (Fig-
ure 6A). Armitage encodes a homolog of SDE3, an RNA helicase
that is involved in RNAi in Arabidopsis (Tomari et al., 2004) and
armi mutations disrupt translational repression and localization
of oskar mRNA in Drosophila oocytes (Cook et al., 2004).
In armi mutant ovaries, loading of flamenco-derived piRNAs
and piRNAs targeting gypsy-family transposons into Piwi is
unaffected, consistent with the maintenance of Piwi expression
and localization in somatic cells (Figures 6B and 6C, Table S7).
However, we could no longer detect Piwi-associated piRNAs
derived from 42AB or piRNAs corresponding to germline-regu-
lated elements in Piwi complexes from mutant animals (Figures
6B and 6C).
Because of its selective effect in the germline, those piRNAs
remaining Piwi-associated in armi mutants must represent
somatic species. This allowed us to re-evaluate requirements
for the somatic pathway among the remaining eight mutants
that we characterized (Figure S12). In accord with our prior
conclusions, only piwi, flamenco, and zucchini impacted this
selected set of RNAs.
An examination of total RNA from armi mutants indicated that
piRNAs contributed from all germline clusters were generally
depleted (Figure 6D). Of the many germline elements that rely
heavily upon the integrity of spn-E for piRNA production, many
were also impacted by armi (e.g, RT1B, Figures 6E and 6F).
However, a number of elements, including protoP-A, F element,
and doc, were relatively insensitive to armi mutations (Figures 6E
and 6F). Nearly all germline elements required spn-E for robust
ping-pong. In contrast, a minority of elements depended upon
armi for their participation in the amplification cycle, indicating
that it is not required for the cycle, per se (Figures 6E and 6F).
In fact, a detailed examination of piRNA densities across the
F element indicated that the complex mixture of small RNAs
seen in wild-type ovaries could be split genetically into two
pools. armi mutants retain almost exclusively ping-pong pairs,
while aubergine mutants retain the antisense-biased pool of
small RNAs that lack ping-pong signatures and that likely repre-
sent primary piRNAs (Figures 6F and 6G).
(C) Heat maps indicating ping-pong signals for typical germline (red), intermediate (yellow), and somatic transposons (green) in flamenco and spn-E mutants.
(D) Immunocytochemical analysis of Aub, AGO3, and Piwi protein localization in wild-type and spn-E1/100.37 mutant ovaries.
Cell 137, 522–535, May 1, 2009 ª2009 Elsevier Inc. 531

Figure 6. Piwi Localization and Loading in Germline Cells Requires Armitage
(A) Piwi protein localization in wild-type and armi1/72.1 mutant ovaries.
(B) Densities of uniquely mapping, Piwi-bound piRNAs over the 42AB and flamenco clusters from the wild-type (Oregon R, black) and armi mutants (yellow).
(C) Annotation of repeat derived, Piwi-bound piRNAs from wild-type and armi mutant ovaries.
(D) Log2 fold changes of cluster-derived piRNAs in an armi mutant total RNA library compared to heterozygote.
(E) Log2 fold changes of piRNAs mapping antisense to indicated transposons in armi and spn-E mutants compared to respective heterozygotes are shown in heat
map form. Corresponding ping-pong signal heat maps are shown (right).
(F) piRNA densities over indicated transposons in heterozygote (black) and armi mutant (yellow) libraries.
(G) F element ping-pong profiles in armi heterozygote (black), armi mutant (yellow), and aub mutant (blue) libraries.
532 Cell 137, 522–535, May 1, 2009 ª2009 Elsevier Inc.

Overall, these data were consistent with a model in which Piwi
functions analogously in the germline and in the soma, accepting
primary piRNAs produced as a result of processing of cluster
transcripts. For some elements, these primary piRNAs appear
to be essential for sustaining a robust response. However, for
others, Aub and AGO3 can sustain a ping-pong response and
support substantial piRNA populations in the absence of Piwi
complexes loaded with cluster-derived piRNAs (Figures S5–
S8). This may be due, in part, to the priming of the pathway
against those elements by maternally contributed piRNAs.
DISCUSSION
The piRNA pathway forms an evolutionarily conserved mecha-
nism for recognizing and selectively silencing mobile genetic
elements. We have worked to deepen our understanding of the
piRNA pathway by comparing piRNA populations from ovaries
to early embryos and by examining small RNA populations in
nine Drosophila mutants. These studies have revealed the exis-
tence of two related but distinct piRNA pathways that operate in
the somatic and germline compartments of the Drosophila ovary.
Similar conclusions were reached by Zamore and colleagues
(Li et al., 2009) though their detailed analysis of piRNA populations
AGO3 mutants. Each pathway showed unique features and
distinct genetic dependencies.
In germ cells, the integrity of the piRNA pathway was strongly
affected by seven different mutations, including piwi, auberinge,
spindle-E, vasa, krimper, armitage, and zucchini. For mostof these
genes, loss of function strongly reduced overall piRNA levels. All
but piwi, zucchini, and armitage had a substantial impact on the
operation of the ping-pong amplification cycle that forms the
adaptive arm of the pathway.
Nuage are a signature feature of germ cells in animals
(al-Mukhtar and Webb, 1971; Eddy, 1974; Mahowald, 1968;
Snee and Macdonald, 2004; Wilsch-Brauninger et al., 1997),
which all share a need to guard their genomes from mobile
elements. Previous studies have indicated that Aubergine and
AGO3 occupy these structures. We found that this localization
was disrupted specifically by mutations that reduce the operation
of the ping-pong cycle. In Drosophila germ cells, nuage may
concentrate both piRNAs and their targets to facilitate selective
amplification of piRNAs targeting active elements (Klattenhoff
and Theurkauf, 2008). However, since these structures also
contain proteins that have not yet been linked to the piRNA
pathway, nuage might also play additional roles in germline RNA
metabolism.
Although the piRNA clusters that operate in the germline
generally produce small RNAs from both genomic strands and
contain element fragments in random orientations, the overall
system is strongly biased toward antisense species (Brennecke
et al., 2007). This bias is even more evident in small RNAs formed
independently of the ping-pong cycle, indicating that primary
biogenesis from piRNA clusters somehow perceives strand
information. The mechanism by which this occurs remains
mysterious, since none of the mutations that we evaluated selec-
tively impacted this feature of the germline pathway.
In somatic follicle cells, a simplified version of the piRNA
pathway is driven by the flamenco piRNA cluster. Unlike loci
that operate in germ cells, the flamenco cluster generates
piRNAs in the absence of ping-pong from only one strand and
from a precursor RNA that contains element fragments in
a uniform orientation (Brennecke et al., 2007). This arrangement
is superficially similar to pachytene piRNA clusters in mammals.
However, these do not play a role in transposon control, and their
relevant targets remain unknown (Aravin et al., 2007a). Thus,
these generative loci share only a propensity to generate piRNAs
via a primary biogenesis mechanism.
Unlike the germline system, the antisense bias of the somatic
system appears evolutionarily determined by selection for inser-
tion of transposons in a preferred orientation within flamenco.
Support for this hypothesis came from our analysis of two related
species, D. yakuba and D. erecta. In both of these, flamenco
shares a uniformity of transposon orientation and the production
of predominantly antisense piRNAs from onlyone genomic strand.
In addition to its structure, the content of flamenco loci is
conserved in all three species examined. This locus specifically
targets LTR retrotransposons of the gypsy family. Many elements
within the gypsy family are classified as errantiviral transposons
(Song et al., 1994), which retain the ability to express envelope
proteins. Thus, it has been proposed that gypsy-family elements
have colonized the somatic gonadal niche and propagate in the
population by infecting underlying germ cells with viral particles
produced in follicular epithelial cells (Kim et al., 1994; Lecher
et al., 1997; Song et al., 1997). In accord with this notion, many
of the elements that show a Piwi/flamenco pattern of control
canencode envelope proteins (Chalvetetal., 1999). Thissuggests
that retroelements that can potentially form viral particles have
long occupied a follicle cell niche and that this strategy has
coevolved with a flamenco-directed silencing system in the soma.
Overall, the studies presented here have revealed unexpected
complexities in the piRNA pathway. Two distinct pathways
with different strategies and different genetic compositions are
responsible for the silencing of different transposon classes in
Drosophila ovaries. This suggests that the pathway adapts
specifically to the structure and habits of each element to effec-
tively protect the germline from transposon activity.
EXPERIMENTAL PROCEDURES
Antibodies and Immunocytochemistry
Rabbit polyclonal antisera directed against the N termini of Piwi, Aub, and
AGO3 were previously described (Brennecke et al., 2007). Primary antibodies
were diluted 1:500 for immunohistochemistry (Findley et al., 2003). In brief,
ovaries were fixed in formaldehyde, rinsed, permeabilized in 0.1% Triton and
washed. Ovaries were blocked in bovine serum albumin (BSA) and incubated
overnight with primary antibodies. Ovaries were then washed and incubated
in secondary antibody and washed. Finally, DNA was stained with TO-PRO
(Molecular Probes), washed, and mounted in glycerol. Images were acquired
with a Carl Zeiss Confocal LSM 510 miscroscope. See the Supplemental
Experimental Procedures for further information.
Fly Stocks
The wild-type Drosophila melanogaster strains used in this study are Oregon R
and the I element reactive, wK strain (a kind gift of Silke Jensen), (Hazelrigg et al.,
1984; Luning, 1981). The following allelic combinations were used for immuno-
localization, western, and RNA analyses: armitage, armi1/72.1 (Cook et al., 2004;
Tomari et al., 2004), aubergine, aubQC42/HN (Schupbach and Wieschaus, 1991;
Wilson et al., 1996), flamenco, flamKG00476 (Mevel-Ninio et al., 2007), krimper,
Cell 137, 522–535, May 1, 2009 ª2009 Elsevier Inc. 533

krimpf06583 (Barbosa et al., 2007; Lim and Kai, 2007), piwi, piwi1/2 (Cox et al.,
1998; Lin and Spradling, 1997), spindle-E, spn-E1/100.37 (Gillespie and Berg,
1995), squash, squHE47/PP33 (Pane et al., 2007), vasa, vasD5/PH165 (Liang et al.,
1994; Styhler et al., 1998), zucchini, zucHM27/Df(2I)PRL (Pane et al., 2007). See
the Supplemental Experimental Procedures for further information.
Small RNA Cloning and Analysis
Small RNA libraries were generated as previously described (Brennecke et al.,
2007). In brief, ovaries of the respective genotype were dissected in 10–50 ml
batches into ice cold phosphate-buffered saline (PBS). Total RNA was
extracted with Trizol (Invitrogen) and two phenol:chloroform:isoamyl alcohol
(ROCHE) extraction steps. For each genotype, 50 mg of total RNA was sepa-
rated on a 12% denaturing polyacrylamide gel and 18–29nt small RNAs
were isolated for cloning. A detailed protocol for the generation of small
RNA libraries is available upon request. Corresponding heterozygote libraries
were prepared from ovarian RNA of heterozygous siblings (with mutant alleles
balanced), which were collected from the same crosses. Only sequences
matching the Drosophila release 5 genome (excluding ArmUextra) 100%
were retained. Libraries were normalized to a subset of endo-siRNAs (Supple-
mental Experimental Procedures and Supplemental Glossary) to allow for
cross-analysis. We mapped all 23–29 nt small RNAs to known piRNA clusters
(Brennecke et al., 2007) and to the complete collection of D. melanogaster
transposable elements (Repbase) (Jurka et al., 2005). See the Supplemental
Experimental Procedures for further information. Previously published Piwi/
AGO3/Aub-ovarian-IP libraries from Oregon R flies (GSE6734) (Brennecke
et al., 2007) and total RNA libraries from the wK wild-type strain (GSE13081)
(Brennecke et al., 2008) were also analyzed.
ACCESSION NUMBERS
Small RNA libraries are deposited at Gene Expression Omnibus (accession
number GSE15186, data sets GSM379050–GSM379067 and GSM379301).
SUPPLEMENTAL DATA
Supplemental Data include Supplemental Experimental Procedures, Supple-
mental Glossary, 12 figures, and seven tables and can be found with this article
online at http://www.cell.com/supplemental/S0092-8674(09)00377-8.
ACKNOWLEDGMENTS
We thank members of the Hannon laboratory for helpful discussions and Oliver
Tam for computational assistance. We are grateful to Michelle Rooks, Emily
Hodges, Danea Rebolini, Laura Cardone, and Mike Regulski (Cold Spring
Harbor Laboratory) for help with deep sequencing. We also thank Attilio
Pane and Trudi Schupbach for providing the zuc and squ fly stocks. C.D.M.
is a Beckman fellow of the Watson School of Biological Sciences and is sup-
ported by a National Science Foundation Graduate Research Fellowship. J.B.
is supported by a fellowship from the Ernst Schering foundation. M.D. is an
Engelhorn fellow of the Watson School of Biological Sciences. A.S. is sup-
ported by a Human Frontier Science Program postdoctoral fellowship. This
work was supported in part from grants from the National Institutes of Health
to G.J.H. and W.R.M. and a kind gift from Kathryn W. Davis (G.J.H.).
Received: December 19, 2008
Revised: March 3, 2009
Accepted: March 24, 2009
Published online: April 23, 2009
REFERENCES
al-Mukhtar, K.A., and Webb, A.C. (1971). An ultrastructural study of primordial
germ cells, oogonia and early oocytes in Xenopus laevis. J. Embryol. Exp.
Morphol. 26, 195–217.
Aravin, A.A., Naumova, N.M., Tulin, A.V., Vagin, V.V., Rozovsky, Y.M., and
Gvozdev, V.A. (2001). Double-stranded RNA-mediated silencing of genomic
534 Cell 137, 522–535, May 1, 2009 ª2009 Elsevier Inc.
tandem repeats and transposable elements in the D. melanogaster germline.
Curr. Biol. 11, 1017–1027.
Aravin, A.A., Klenov, M.S., Vagin, V.V., Bantignies, F., Cavalli, G., and
Gvozdev, V.A. (2004). Dissection of a natural RNA silencing process in the
Drosophila melanogaster germ line. Mol. Cell. Biol. 24, 6742–6750.
Aravin, A.A., Hannon, G.J., and Brennecke, J. (2007a). The Piwi-piRNA
pathway provides an adaptive defense in the transposon arms race. Science
318, 761–764.
Aravin, A.A., Sachidanandam, R., Girard, A., Fejes-Toth, K., and Hannon, G.J.
(2007b). Developmentally regulated piRNA clusters implicate MILI in trans-
poson control. Science 316, 744–747.
Barbosa, V., Kimm, N., and Lehmann, R. (2007). A maternal screen for genes
regulating Drosophila oocyte polarity uncovers new steps in meiotic progres-
sion. Genetics 176, 1967–1977.
Brennecke, J., Aravin, A.A., Stark, A., Dus, M., Kellis, M., Sachidanandam, R.,
and Hannon, G.J. (2007). Discrete small RNA-generating loci as master
regulators of transposon activity in Drosophila. Cell 128, 1089–1103.
Brennecke, J., Malone, C.D., Aravin, A.A., Sachidanandam, R., Stark, A., and
Hannon, G.J. (2008). An epigenetic role for maternally inherited piRNAs in
transposon silencing. Science 322, 1387–1392.
Carmell, M.A., Xuan, Z., Zhang, M.Q., and Hannon, G.J. (2002). The Argonaute
family: tentacles that reach into RNAi, developmental control, stem cell
maintenance, and tumorigenesis. Genes Dev. 16, 2733–2742.
Chalvet, F., Teysset, L., Terzian, C., Prud’homme, N., Santamaria, P.,
Bucheton, A., and Pelisson, A. (1999). Proviral amplification of the Gypsy
endogenous retrovirus of Drosophila melanogaster involves env-independent
invasion of the female germline. EMBO J. 18, 2659–2669.
Chambeyron, S., Popkova, A., Payen-Groschene, G., Brun, C., Laouini, D.,
Pelisson, A., and Bucheton, A. (2008). piRNA-mediated nuclear accumulation
of retrotransposon transcripts in the Drosophila female germline. Proc. Natl.
Acad. Sci. USA 105, 14964–14969.
Cook, H.A., Koppetsch, B.S., Wu, J., and Theurkauf, W.E. (2004). The
Drosophila SDE3 homolog armitage is required for oskar mRNA silencing
and embryonic axis specification. Cell 116, 817–829.
Cox, D.N., Chao, A., Baker, J., Chang, L., Qiao, D., and Lin, H. (1998). A novel
class of evolutionarily conserved genes defined by piwi are essential for stem
cell self-renewal. Genes Dev. 12, 3715–3727.
Cox, D.N., Chao, A., and Lin, H. (2000). piwi encodes a nucleoplasmic factor
whose activity modulates the number and division rate of germline stem cells.
Development 127, 503–514.
Desset, S., Meignin, C., Dastugue, B., and Vaury, C. (2003). COM, a hetero-
chromatic locus governing the control of independent endogenous retrovi-
ruses from Drosophila melanogaster. Genetics 164, 501–509.
Desset, S., Buchon, N., Meignin, C., Coiffet, M., and Vaury, C. (2008). In
Drosophila melanogaster the COM locus directs the somatic silencing of
two retrotransposons through both Piwi-dependent and -independent
pathways. PLoS ONE 3, e1526.
Drosophila 12 Genomes Consortium, Clark, A.G., Eisen, M.B., Smith, D.R.,
Bergman, C.M., Oliver, B., Markow, T.A., Kaufman, T.C., Kellis, M., Gelbart, W.,
et al. (2007). Evolution of genes and genomes on the Drosophila phylogeny.
Nature 450, 203–218.
Eddy, E.M. (1974). Fine structural observations on the form and distribution of
nuage in germ cells of the rat. Anat. Rec. 178, 731–757.
Findley, S.D., Tamanaha, M., Clegg, N.J., and Ruohola-Baker, H. (2003). Mael-
strom, a Drosophila spindle-class gene, encodes a protein that colocalizes
with Vasa and RDE1/AGO1 homolog, Aubergine, in nuage. Development
130, 859–871.
Gillespie, D.E., and Berg, C.A. (1995). Homeless is required for RNA localiza-
tion in Drosophila oogenesis and encodes a new member of the DE-H family
of RNA-dependent ATPases. Genes Dev. 9, 2495–2508.
Gunawardane, L.S., Saito, K., Nishida, K.M., Miyoshi, K., Kawamura, Y.,
Nagami, T., Siomi, H., and Siomi, M.C. (2007). A slicer-mediated mechanism

for repeat-associated siRNA 50 end formation in Drosophila. Science 315,
1587–1590.
Harris, A.N., and Macdonald, P.M. (2001). Aubergine encodes a Drosophila
polar granule component required for pole cell formation and related to
eIF2C. Development 128, 2823–2832.
Hazelrigg, T., Levis, R., and Rubin, G.M. (1984). Transformation of white locus
DNA in drosophila: dosage compensation, zeste interaction, and position
effects. Cell 36, 469–481.
Houwing, S., Kamminga, L.M., Berezikov, E., Cronembold, D., Girard, A., van
den Elst, H., Filippov, D.V., Blaser, H., Raz, E., Moens, C.B., et al. (2007). A role
for Piwi and piRNAs in germ cell maintenance and transposon silencing in
Zebrafish. Cell 129, 69–82.
Jurka, J., Kapitonov, V.V., Pavlicek, A., Klonowski, P., Kohany, O., and
Walichiewicz, J. (2005). Repbase Update, a database of eukaryotic repetitive
elements. Cytogenet. Genome Res. 110, 462–467.
Kennerdell, J.R., Yamaguchi, S., and Carthew, R.W. (2002). RNAi is activated
during Drosophila oocyte maturation in a manner dependent on aubergine and
spindle-E. Genes Dev. 16, 1884–1889.
Kim, A., Terzian, C., Santamaria, P., Pelisson, A., Purd’homme, N., and
Bucheton, A. (1994). Retroviruses in invertebrates: the gypsy retrotransposon
is apparently an infectious retrovirus of Drosophila melanogaster. Proc. Natl.
Acad. Sci. USA 91, 1285–1289.
Klattenhoff, C., and Theurkauf, W. (2008). Biogenesis and germline functions
of piRNAs. Development 135, 3–9.
Klenov, M.S., Lavrov, S.A., Stolyarenko, A.D., Ryazansky, S.S., Aravin, A.A.,
Tuschl, T., and Gvozdev, V.A. (2007). Repeat-associated siRNAs cause chro-
matin silencing of retrotransposons in the Drosophila melanogaster germline.
Nucleic Acids Res. 35, 5430–5438.
Lecher, P., Bucheton, A., and Pelisson, A. (1997). Expression of the Drosophila
retrovirus gypsy as ultrastructurally detectable particles in the ovaries of flies
carrying a permissive flamenco allele. J. Gen. Virol. 78, 2379–2388.
Li, C., Vagin, V.V., Lee, S., Xu, J., Ma, S., Xi, H., Seitz, H., Horwich, M.D.,
Syrzycka, M., Honda, B.M., et al. (2009). Collapse of germline piRNAs in the
absence of Argonaute3 reveals somatic piRNAs in flies. Cell 137, in press.
10.1016/j.cell.2009.04.027.
Liang, L., Diehl-Jones, W., and Lasko, P. (1994). Localization of vasa protein to
the Drosophila pole plasm is independent of its RNA-binding and helicase
activities. Development 120, 1201–1211.
Lim, A.K., and Kai, T. (2007). Unique germ-line organelle, nuage, functions to
repress selfish genetic elements in Drosophila melanogaster. Proc. Natl.
Acad. Sci. USA 104, 6714–6719.
Lin, H., and Spradling, A.C. (1997). A novel group of pumilio mutations affects
the asymmetric division of germline stem cells in the Drosophila ovary. Devel-
opment 124, 2463–2476.
Luning, K. (1981). Genetics of inbred Drosophila melanogaster. Hereditas 95,
181–188.
Mahowald, A.P. (1968). Polar granules of Drosophila. II. Ultrastructural
changes during early embryogenesis. J. Exp. Zool. 167, 237–261.
Malone, C.D., and Hannon, G.J. (2009). Small RNAs as guardians of the
genome. Cell 136, 656–668.
Mevel-Ninio, M., Pelisson, A., Kinder, J., Campos, A.R., and Bucheton, A.
(2007). The flamenco locus controls the gypsy and ZAM retroviruses and is
required for Drosophila oogenesis. Genetics 175, 1615–1624.
Murchison, E.P., Kheradpour, P., Sachidanandam, R., Smith, C., Hodges, E.,
Xuan, Z., Kellis, M., Grutzner, F., Stark, A., and Hannon, G.J. (2008). Conser-
vation of small RNA pathways in platypus. Genome Res. 18, 995–1004.
Pane, A., Wehr, K., and Schupbach, T. (2007). zucchini and squash encode
two putative nucleases required for rasiRNA production in the Drosophila
germline. Dev. Cell 12, 851–862.
Pelisson, A., Payen-Groschene, G., Terzian, C., and Bucheton, A. (2007).
Restrictive flamenco alleles are maintained in Drosophila melanogaster popu-
lation cages, despite the absence of their endogenous gypsy retroviral targets.
Mol. Biol. Evol. 24, 498–504.
Prud’homme, N., Gans, M., Masson, M., and Terzian, C. (1995). Flamenco,
a gene controlling the gypsy retrovirus of Drosophila melanagaster. Genetics
139, 697–711.
Saito, K., Nishida, K.M., Mori, T., Kawamura, Y., Miyoshi, K., Nagami, T.,
Siomi, H., and Siomi, M.C. (2006). Specific association of Piwi with rasiRNAs
derived from retrotransposon and heterochromatic regions in the Drosophila
genome. Genes Dev. 20, 2214–2222.
Sarot, E., Payen-Groschene, G., Bucheton, A., and Pelisson, A. (2004).
Evidence for a piwi-dependent RNA silencing of the gypsy endogenous
retrovirus by the Drosophila melanogaster flamenco gene. Genetics 166,
1313–1321.
Schupbach, T., and Wieschaus, E. (1991). Female sterile mutations on the
second chromosome of Drosophila melanogaster. II. Mutations blocking
oogenesis or altering egg morphology. Genetics 129, 1119–1136.
Shpiz, S., Kwon, D., Rozovsky, Y., and Kalmykova, A. (2009). rasiRNA pathway
controls antisense expression of Drosophila telomeric retrotransposons in the
nucleus. Nucleic Acids Res. 37, 268–278.
Shpiz, S., Kwon, D., Uneva, A., Kim, M., Klenov, M., Rozovsky, Y., Georgiev, P.,
Savitsky, M., and Kalmykova, A. (2007). Characterization of Drosophila
telomeric retroelement TAHRE: transcription, transpositions and RNAi-based
regulation of expression. Mol. Biol. Evol. 24, 2535–2545.
Slotkin, R.K., and Martienssen, R. (2007). Transposable elements and the
epigenetic regulation of the genome. Nat. Rev. Genet. 8, 272–285.
Snee, M.J., and Macdonald, P.M. (2004). Live imaging of nuage and polar
granules: evidence against a precursor-product relationship and a novel role
for Oskar in stabilization of polar granule components. J. Cell Sci. 117,
2109–2120.
Song, S.U., Gerasimova, T., Kurkulos, M., Boeke, J.D., and Corces, V.G.
(1994). An env-like protein encoded by a Drosophila retroelement: evidence
that gypsy is an infectious retrovirus. Genes Dev. 8, 2046–2057.
Song, S.U., Kurkulos, M., Boeke, J.D., and Corces, V.G. (1997). Infection of
the germ line by retroviral particles produced in the follicle cells: a possible
mechanism for the mobilization of the gypsy retroelement of Drosophila.
Development 124, 2789–2798.
Spradling, A.C. (1993). Germline cysts: communes that work. Cell 72,
649–651.
Styhler, S., Nakamura, A., Swan, A., Suter, B., and Lasko, P. (1998). vasa is
required for GURKEN accumulation in the oocyte, and is involved in oocyte
differentiation and germline cyst development. Development 125, 1569–1578.
Tomari, Y., Du, T., Haley, B., Schwarz, D.S., Bennett, R., Cook, H.A.,
Koppetsch, B.S., Theurkauf, W.E., and Zamore, P.D. (2004). RISC assembly
defects in the Drosophila RNAi mutant armitage. Cell 116, 831–841.
Vagin, V.V., Klenov, M.S., Kalmykova, A.I., Stolyarenko, A.D., Kotelnikov, R.N.,
and Gvozdev, V.A. (2004). The RNA interference proteins and vasa locus are
involved in the silencing of retrotransposons in the female germline of
Drosophila melanogaster. RNA Biol. 1, 54–58.
Vagin, V.V., Sigova, A., Li, C., Seitz, H., Gvozdev, V., and Zamore, P.D. (2006).
A distinct small RNA pathway silences selfish genetic elements in the germline.
Science 313, 320–324.
Wilsch-Brauninger, M., Schwarz, H., and Nusslein-Volhard, C. (1997). A
sponge-like structure involved in the association and transport of maternal
products during Drosophila oogenesis. J. Cell Biol. 139, 817–829.
Wilson, J.E., Connell, J.E., and Macdonald, P.M. (1996). aubergine enhances
oskar translation in the Drosophila ovary. Development 122, 1631–1639.
Cell 137, 522–535, May 1, 2009 ª2009 Elsevier Inc. 535


















![local piRNA biogenesis · presumably through phased piRNA biogenesis [37–39] , since transcripts derived from unique genomic regions flanking the TE will not be the direct target](https://img.dokumen.tips/doc/110x75/5f51f802441d0649ae60009e/local-pirna-biogenesis-presumably-through-phased-pirna-biogenesis-37a39-since.jpg)










