Embed Size (px)
Citation preview

SPECIAL ARTICLE
The Electrical Potential Difference Acrossthe Cell Membrane of Heart Muscle
Biophysical Considerations
By ERNEST PAGE, M.D.
T HE CELL MEMBRANE of mammalianheart muscle cells may be characterized
by three interrelated observations. In the firstplace, the membrane separates an intracellularsolution having a high potassium (K) and alow sodium (Na) concentration from an ex-tracellular solution with a high Na and lowK concentration. Secondly, K and Na ionscan be shown with radioactive tracer technicsto be continuously passing into and out of thecell; to do so, these ions. must cross the cellmembrane, which is therefore said to be per-mneable to them. In spite of the apparent per-meability of the membrane to both Na and K,the intracellular concentrations of these ionsare observed to remain remarkably constant.Finally, an electrical potential difference of80 to 90 millivolts can be measured across thecell membrane of quiescent (noncontracting)heart muscle cells between an electrode in theintra.cellular solution and a sinmilar electrodein the solution bathinlg the exterior of thecell. This electrical potential difference can-not be detected by measurenments from theoutside of intact cells as in the electrocardio-gram. It may, however, be observed either asthe "injury potential" of cells with damagedcell membranes or as the somewhat larger"resting potential" in cells whose cell mem-brane has been pierced with a microelectrode.The sign of the electrical potential difference
From the Biophysical Laboratory, Harvard MedicalSchool, Boston, Massachusetts.
582
across the cell mnembrane undergoes periodicreversals called "action potentials." Thetime course of the vectorial summation ofsuch action potentials fromn many cells is re-flected in the QRST complexes of the surfaceelectrogram and conventional electrocardio-gram.
TheoryThe study of the electrical potential differ-
ence across ion-permeable membranes sep-arating two salt solutions of different ioniccomposition is a province of physics and ofphysical chemistry, which attempt to formu-late in mathematical terms an exact and rig-orous description of the processes involved.1' 2The present article explains the same physicaland physicochemical processes in nontechni-cal language in order to promote a betterunderstanding of cell-membrane phenomenaamong clinical cardiac physiologists.
Since many aspects of the behavior of cellmembranes cani be reproduced experimentallywith artificial membranes,3-6 it is useful tobegin bv inquiring how electrical potentialdifferences may arise across such artificialmembranes. To take an apparently simplemodel, consider two well-stirred reservoirs ofequal and fixed volume separated by a porouspartition or membrane (fig. la). Let the leftreservoir be filled with a solution of potassiunichloride (KCI) at a conceentration of 200 mil-limoles per liter and the right reservoir witha KCI solution at a concentration of 5 milli-moles per liter. Assume that movements of
Circulation, Volume XXVI, October 1962
by guest on June 4, 2018http://circ.ahajournals.org/
Dow
nloaded from

ELECTRICAL POTENTIAL DITFFERENCE
la.
KCI 200 mM
KCI 5mM
MEMBRANE MEMBRANE LESS MEMBRANEEQUALLY PERMEABLE PERMEABLE IMPERMEABLE
TO Ci- AND K+ TO Cl- THAN TO K+ TO Cl-
rigure 1The electrical potential difference produced during diffusion of a KCl solution across
a model membrane.
wa.ter through the porous partit,ion are pre-
vented. Only KCl can then move through themembrane. When diss,olved in water, this saltseparates into two particles of equal and op-
posit.e charge, the K and Cl ions. Each ionsurrounds itself with a number of moleculesof the solvent, water (the so-called hydrationshell), and henceforth behaves as an inde-pendent molecule, an important restrictionbeing that for the solution as a whole or any
microscopic portion of it the sum of the Kions be, always, equal to the sum of the Cl ions.The unequal partition of dissolved KCI on
the two sides of the membrane will give riseto a, redistribution of the dissolved salt by a
random process called diffusion. Since theconcentration of KCl in the left reservoir isinitially 40 times as great as that in the rightreservoir, there will initially be 40 times as
great a tendency for KCI to diffuse from leftto right as from right to left. As a conse-
quence, a net movement of KCl from left toright is set up. This net movement will con-
tinue until the concentrations in the tworeservoirs have become equal, i.e., until thegradient of concentration has been abolishedby the trans.fer of KCI from left to right. If
Circulation, Volume XXVI, October 1962
the positively charged K ions and the nega-
tively charged Cl ions move through the mem-brane with equal ea-se, the membrane is saidto be, equally permeable to K and Cl. Thesepositively and negatively charged particleswill then pass t.ogether through the pores inthe membrane, electrically neutralizing each
other. The net movement of KCI will occur
without a separation of positive from negativecharge, and a device for measuring the elec-trical potential difference across the mem-
brane will accordingly register zero volts whilethe diffusion is in progress (fig. lb).
Suppose, now, the membrane is altered so
that the passage of the Cl ion across the mem-
brane is made more difficult than that of theK ion. Then, as K and Cl move from left toright under the force set up by the gradientof concentration, a thin cross-section of themembrane will show the advancing front ofthe net diffusion process to consist of thereadily penetrating, positively charged K ions.These K ions will drag behind them by theforce of electrostatic attraction the negativelycharged Cl ions whose movement is impededby the membrane (fig. ie). KCI now diffusesas a dipole, oriented so that the positive pole
lb.
I
0
0
NO POTENTIALDIFFERENCE
Ic.
I
DIFFUSIONPOTENTIAL
id.
IMAXI MALPOTENTIALDIFFERENCE
583
by guest on June 4, 2018http://circ.ahajournals.org/
Dow
nloaded from

PAGE
(the K ion) is leading and the negative pole(the Cl ion) is following. A sufficiently sensi-tive voltmeter across the thin section of thememnbrane will indicate that the right face ofthe membrane is electrieally positive withrespect to the left face. The reading oni thevoltmeter reflects a separation of positivefromn ilegative charges such that the positivelycharrged K ions are at a relatively higher con-
centration toward the right face of the mem-
brane and the negatively charged Cl ions aremore concentrated toward the left face. Anelectrical potential differenee arising in thisway is called a diffusion potential. It stemsfrom the difference iii the mobility withinthe membrane pores of the positive anidnegative ions of a salt diffusing throughthe memnbrane. The spatial separation pro-
duced by the membrane between the K amidCl ions need be very slight, a separationof a small fraction of an Angstromn unitinivolving only a small number of KCInolecules sufficing to gemierate a very appre-
ciable potential difference. [One Angstronmunit (abbreviated A - 10-1i nmeters = one
ten billionth of a mueter.] A diffusion potentialexists while an actual net moveunelit of EC1through the miieinbrane is unider way. Theexistence of a diffusion potential and the factthat its magnitude cail be shown to dependon the coneentration of a given ion are there-fore evidenee that the mnembrane is permneableto that particular ion.
In the interaction betweeni menmbranes andsalts like KCI there is a continuous gradationin the degree to which the membra.ne restrictsthe nlovements of either positively chargedions (cations) or negatively charged ions(anions). At the onie extreme, the membranebrings about no separa.tion of positive froniimegative charge and no diffusion potential(fig. lb); the mobilities of cation and anioliin the membrane are equal. At the other ex-
treme, the memnbrane is permeable only to ionsof one charge (e.g., to the cation, K) andtotally impermeable to ions of the oppositecharge (e.g., to the anion, Cl). The separatiomof charge and hemmee the electrical potential
difference produced are maximal (fig. ld).Intermediate between these two extremes isthe type of diffusion potential (fig. lc) thatcorresponds to the situation at many eell mem-branes (See Appendix 1).
The three cases are summarized mathematicallyby the Nernst Equation. For the diffusion of apotassiuml salt at 370 C this equation takes theform
vm =- U61.5log ([K] [K]0)u-I- u-
in which Vm is the electrical potential differenceacross the mnembrane, u+ and u- are the mobilities(velocities per unit force) of K and Cl withinthe membrane, and [K] i and [K]. are the con-centrations of K in the solutions on either sideof the membrane (the "inside" and "outside" solu-tions, respectively). When the electroheemical mlo-bilities of cation and anion are equal (u+ - u-), thefraction cointaining the mobility termns vanishesand Vm = 0. When the anion mnobility in themnembrane, u-, is zero (the imlemibrane is imperme-able to Cl), the u's cancel and Vm becomes equalto -61.5 log (L[K]I/[K0). Under these condi-tions Vm depends only on the ratio of the Kconcentrations in the solutions bathing the twosides of the membrane. Finally, if us and u- areunequal and neither is zero, Vm is the type ofdiffusion potential believed to exist across heartnusele cell membrane (See Appendix 2).
It is pertinent to consider next by whatmechanismts a porous mnembrane niay impedethe diffusion of one ion of salt like KCl so asto bring about a separation of positive fromnegative charges and, hence, to set up a dif-fusion potential. Diffusion of ions throughthe membrane probably occurs via the water-filled pores in the mnembrane. A membranemay therefore be expected to impede thetransit of ions of a size large compared tothe diameter of its pores, while permitting themovement of ions whose diameter is relativelysmall. Figure 2 compares the hydrated iondiameters (the diameter of the ion plus theshell of water molecules that moves with it)of K, Na, and Cl with the diameter of thepores in the cell membrane of eat ventricularmuscle. These pores have recently been shownto have a diameter not in excess o;f 8A.7 Itwill be seen from figure 2 that the membraneinight be expected on the basis of relative ionsize to be more permeable to K and Cl ions
Circulation, Volume XXVI, October 1962
584
by guest on June 4, 2018http://circ.ahajournals.org/
Dow
nloaded from

ELECTRICAL POTENTIAL DIFFERENCE
than to the larger Na ion. The diameters ofthe K and Cl ions, however, are sufficientlysimilar that the pores of the membrane cannotdiscriminate between them by virtue of theirsize. Since such discrimination is required inorder to generate a diffusion potential, it isnecessary to look for an additional mechanismfor separatnig the positively charged K ionfrom itis negatively charged ion partner.In figure 3 a simplified model of a mem-
brane pore is depicted as lined with fixednegative electrical charges, that is, with nega-tively charged molecules built into the sub-stance of the walls of the pore. These fixedcharges attract, by the force of electrostaticat-traction, ions having an electrical chargeopposite to their own charge, and repel, bythe force of electrostatic repulsioni, ions hav-ing the same charge as their own. The sitesat which the fixed charge groups abut oni thewater-filled lumen of the pore become coatedwith oppositely charged ions from the solu-tioln, the counter-ions. Unlike the fixedcharges themselves, counter-ions are free toexchange with other K ions in the solution.If the pore is wide enough (fig. 3a), the elec-trostatic effects of the fixed charges do notappreciably influence the diffusion of ionsthrough the membrane. As the pore becomesprogressively narrower, however, the fixedcharges become increasingly effective in ex-eluding ions of like sign (fig. 3b). At a cer-tain critical pore diameter, only ions of op-posite sign, in this case, K ions, can pass (fig.3e). The existence of fixed charges in cellmembranes is strongly suggested by theirselective permeability to ions and by the find-ing that, isolated membrane fragments containlipid-carbohydrate-protein complexes withelectrically charged lipid components.8The nature of a diffusLion potential has been
discussed with reference to the two-compart-ment. model of figure la in which a porousmembrane separates a 200 mM KCl solutionfrom a 5 mM KCl solution. It is apparentthat with the passage of time the model willtend to run down, i.e., when both K and Clcan -permeate, the diffusion will come to an
Circulation, Volume XXVI, October 1962
DIAMETER OF PORESOF CAT HEART
MUSCLE CELL MEMBRANE:8.0 A OR LESS.
ION
Na
K
CI
H20( NOT( IONIZED J
DIAMETER A
0
A
5.12
3.96
3.86
1.5 0
Figure 2Comparison of size of ions and water with poresize of cat heart muscle cell membrane.7' 34, 35 *Iondiameters include the hydration shell of the ion.
end as the concentrations of KCl on the twosides of the nmembrane approa.ch equality.What is required for a maintained diffusionalnd therefore for a. maintainted diffusion po-tential is a mechanism that continually re-plenishes the KCl coneentration in the 200nM reservoir and continually depletes theKCI cloneentration in the 5 mM reservoir. Therates of repletion and depletion must be suffi-cient to keep up with the rate at which KCldiffuses from left to right, down the concen-tration gradient, through the pores in themembrane. Sueh a mechanism would set upa steady state in which, by some energy-con-suming process, KCl is transported "uphill"against the concentration gradient from the5 mM compartment into the 200 mM compart-ment as fast as it is lost from the 200 mMcompartment by diffusion "downhill."The situation is analogous to that of two
lakes with different water levels separated bya leaky dam. For the purposes of this analogy,the difference in water levels is maintainedby pumping water from the lower lake intothe upper at a rate equal to that at which
585
by guest on June 4, 2018http://circ.ahajournals.org/
Dow
nloaded from

PAGE
@30303(3 (3 (3 (D (3
(D (3 (D se
Wide pore.K+ and Cl-pass readily.No potential difference.
0.
(39(D( (3(3(D E (±- (D (
Intermediate pore.
Cl- partially excluded.Diffusion potential.
b.
0000F KCI5 mM
Narrow pore.Cl- totally excluded.
Maximum potential difference.
C.
Figure 3Effect of fixed charges onz permea-bility of membrane to ions. The schematically indi-cated pores are shown as lined with fixed negatite electrical chalrges. KCO is diffusingfrom the 200 mM solution on the left into the 5 mM solution on the right. Modifiedafter Sollner.3
water leaks through the damti froimi the upper
into the lower lake. The same pumping meeh-anism that miiailitains the difference of waterlevels also perpetuates the leak.
The cell membranie of heart mnuscle, in com-
mon with that of virtually all living cells,9is the site of energy-consuming mechanismisfor mnoving ions and other molecules againstconcentration and electrical gradients. Thesetuphill" processes, ofteni referred to as "ac-
tive transport," derive their energy supplyfrom cellular metabolic reactions, most prob-ably fromn the terminal high-enlergy phosphatebonid of adenosine triphosphate. By contrast,the "downhill" diffusion, which the "uphill"processes serve to maintain, oceurs sponta-neously without the necessity for a specialmetabolie energy input, and such "downhill"diffusion is consequenitly termed "passivetransport " (See Appendix 3). Unlike " down-hill" diffusion, active transport does not occur
through water-filled pores in the membrane.Instead, for the purpose of transport againstconcentration and electrical gradients, K andNa appear to combine at the inembrane sur-
face with certain molecules called carriers or,
colloquially, "ion pumps," which are part of
the substance (matrix) of the membrane it-self. In heart muscle this conibination, usuallyreferred to as anl ion-carrier complex, willhave to remain hvpothetical umntil it is chem-
ically i(lenL-tified. In tissues in which such ion-carrier complexes have been more closelvstudied, 11 they displav mnany of the char-acteristies of enzyvmes.
Experimental Approaches'The theoretical considerations presented so
far suggest that the ion permeability of heartmuscle-cell menubrane should be accessible tomeasurement bv three experimeental ap-
proaches. These include the measurement ofintracellular ion colncentrations and cell vol-umes, of the electrical poten-tial differeneeacross the cell membrane, and of the rate atwhich radioactively labeled ionis pass throughthe inembrane into and out of the cell.
Figure 4a shows the intracellular K anid Naconcentrations in papillary mLuscles from theright ventricles of cat hearts, 12 iimmersed ina solution having the approximnate K and Naconcentration of cat plasma. It is, apparetitfrom the figure that the cell membrane is thesite of a steep cooncentrationi gradient of 200mM to 5.3 mM favoring the diffusion of K outof the cell, and an almost equally steep con-
centration gradient of 178 muM to 5 mnM favor-inig the diffusion of Na into the cell. Nowassume that the ainion, Cl, will passively ae-
company any outward diffusioni of K or in-ward diffusion of Na. Then-, in terms of themlodel previously developed, the concentrationgradients for K anid Na produce a curious
Circulation, Volume XXVI, October 1962
KCI200 mM
586
by guest on June 4, 2018http://circ.ahajournals.org/
Dow
nloaded from

EliECTRICAL POTENTIAL DIFFERENCE
b.
INTRACELLULAR EXTRACELLULAR INTRACELLULAR EXTRACELLULAR
K200
Na 5
No 178
K 5.3
Figure 4(a) Intracellular and extracellular K and Na concentrations in resting cat papillarymuscle.7 12 (b) Steady state model corresponding to (a). K and Na diffuse "downhill"through the pores in the membrane in the direction indicated by the solid arrows;
they are returned "uphill" by arn active transport mechanism located within thesubstance of the membrane.
situation (fig. 4b), with the possibility of twooppositely directed diffusion processes settingup diffusion potentials of opposite algebraicsign. The electrical potential difference actu-ally measured a,cros,s the cell membrane willrepresent a summation of these two opp,ositediffusion potentials. Since only ions that can
diffuse through the membrane can be effectivein producing a diffusion potential, the rela-tive contributions of K and Na to the elec-trical potential difference in heart muscledepend on the relative permeability of thecell membrane to these two cations (See Ap-pendix 4).
Figure 5 illustrates an experimental methodfor comparing the K and Na permeabilitiesof the cell membrane of quiescent cat papil-lary muscle.7 12 The experimental procedure
was, to raise the K concentration of the solu-tion ba,thing the external surface of the cellmembrane a,pproximately 10-fold by a mole-for-nmole substitution of K for Na, keepingthe total concentration (K + Na) constant.After a period of equilibration sufficientlylong for completion of all net movements ofions and water, the ecell volumes and intra-cellular ion concentra,tions, of the muscle were
determine,d. When K was substituted for Nain this way, the cells behaved as, if they hadbeen placed in a hypotonic solution, i.e., theytook up water and swelled. Chemical analysisof the muscle revealed the intracellular K
Circulation, Volume XXVI, October 1962
concentra,tion to be unchanged, while thecellular water and K content ha,d increased.The cellular uptake of K, Cl, and water showsthat the cells, are permeable to these threemolecules. Looked at in another way,, theexperiment demonstrates that resting heartmuscle is normally sufficiently impermeableto the Na ion for the extracellular Na to be-have osmotically like an impermeant sub-stance. Thus, when the external Na coneen-tration is lowered to 133 mM, the deficit intonicity cannot be made up by an equivalentconcentration of K, since K, unlike Na, is an
ion to, which the cell membrane is readilypermeable.As previously stated, those ions to which the
cell membrane is, most permeable will contrib-ute most significantly to the observed electricalpotential difference a-cross the membrane.Figure 6 illustrates the dependence of theelectrical potential difference across the cellmembrane of quiescent ca,t papillary muscleon the concent-ration of external K13, shownby the experiments just described to be a rela-tively permenant cation. In figure 6, the fullline is the value the potential difference shouldhave if it, depended only on the ratio of theintracellular and extracellular K concentra-tions according to the relation, Vm = -59.6log( [K] i/ [K]o). The broken line connectingthe experimental points indicates. the ele.c-trical potential difference actually measured
FK 200 mM| Na 178 mM|
\ 5.3mN c 5mM| K5. M
LL
F
i
51-87
by guest on June 4, 2018http://circ.ahajournals.org/
Dow
nloaded from

588
IOX NORMAL EXTERNALK CONCENTRATION
(SUBSTITUTE K FOR NO)
[K]1 = 200 mM
[K]o = 50 mM
[Na]o = 133 mM
[K]0+ [No]o = 183 mM
Figure 5Comparison of permeability of cat p,apillarymunscle cell membrane to K and Na.7 Left: Cellvolume acnd intracellular K concentration in solu-tion with normal external K concentration; rigbt:in solution with 1OX normal external K concen-
tration, made by a mole for mole substitution ofK for external Na. Increase in cell volume exag-
gerated for illustrative purposes.
with the intracellular microelectrode technic.In this techn-iie a glass electrode with a tip ofless than 0.5 muieron is introduced into thecell; the hole so nmade in the cell memnbraneis, well sealed and small enough so that thereis no significant admixture of intracellularand extracellular solutions. The observed po-
tential differenee is large at physiological (5.3mM) and lower external K colncentrations; itdinminishes progressively as the external K
concentration is raised, i.e., as the differencein K coneentrations on the two sides of themembrane beeomnes smaller. At the higherexternal K concentrationis the slope of thebroken line approaches that of the theoreticalline; at physiological anid lower external Kconceltratiomis the experimental slope is muchless steep tha.n the theoretical, and the ob-served electrical potential differen-ce is pro-
gressively less than that predicted from theratio of the intracellular and extracellular Kconcentrations. These phenonmena are con-
veniently iliterpreted in terms of the two-compartment mnodel of figure 4b, which repre-
150_
E
z _*
u _
fL_
CI50 _
1 2 3 5 10 20 30 50 iGSEXTERNAL K CONC (mM)
Figure 6D)ependen cc of electrical potential difference acrossthe cell membrane of quiescent cat papillarymuscle on external K concentration.3 The intra-cellular K concentration is constant at 200 mM.Experimental points connected by brok-en lineare the ralues of the electrical potential differenceobtained with the intracellular microelectrodetechnic. The solid line is the value the electricalpotential difference would have, if it dependedexclusively on the ra'tio o f intracellular to extra-cellular K concentrations according to the re-lation Vr,n = -59.6 log ([K]i1/[K]j (tempera-ture 27.5' C).
sents a sumiationi of two oppositely directeddiff-usioni processes setting up a potential dif-ference whose magnitude depends on the rela-tive permneabilities to K and Na. The experi-mnental observation is that at low external Kcon-centrations the electrical potential differ-eicce across the nmembrane does not dependsolely on the outward diffusion of K. Themembrane permneability to Na in the restingsteady state, though small relative to that forK, is sufficienitly important that the inwarddiffusion of NaCI contributes significantly tothe potential difference.A third method for characterizing the cell
mnemubrane is the nmeasureient of the rates ofmovenment of ions aeross the membrane byuse of radioactive tracer technies.14 The rateof movement of an ionI across a membrane isreferred to as an ion flux; such a flux mnightbe expressed, for example, as picomoles of Kcrossing unit area of cell inembrane per unittinme (pmnol/em2 mini). (One picomole - 10-12mole -one millionth nmillionth mole.) A sub-
Circulation, Volume XXVI, October 1962
NORMAL EXTERNALK CONCENTRATION
[K]0 = 5 mM
[No] = 178 mM
[K]0+ [Na]o = 183 mm
PAGE
o
I
by guest on June 4, 2018http://circ.ahajournals.org/
Dow
nloaded from

ELECTRICAL POTENTIAL DIFFERENCE
stance to which the membralne is permeableis continuously pass,ing through it in both di-rections. Accordingly, the ion movement maybe considered as made up of two components,one directed into the cell, called the influx,and one directed outward, called the outflux.The net flux, repres,enting the difference be-tween influx and outflux, determines whetherthe cell is gaining or los,ing the substance be-ing traced. The net flux is zero in the steadystate, an important special case in which theinflux and outflux are equal, i.e., in which therate of cellular accumulation equals the rateof loss. This condition, which prevails in thequiescent heart muscle cell, is necessary if theintracellular ion concentrations and the re-lated electrical potential difference are to re-main c.onstant. In terms of the model of figure4b, the Na, influx occurs "downhill" with re-spect to electrical aand concentration gradi-ents; it must therefore in the steady state beequal to the Na outflux a,gainst the electro-chemical gradient, and, similarly, the "down-hill" K outflux must be balanced by an equaland opposite "uphill" K influx.As indicated in figure 4b, the "downhill"
fluxes of Na and K probably occur throughwater-filled pores in the membrane. Accord-ingly, these "downhill" movements may beexpected to obey in a gene.ral way the physi-coehemical laws applicable to electrolytes, inaqueous solutions.15 16 The "uphill" fluxes,on the other hand, appear to involve surfa.cereactions with the substance of the membraneand obey a different set of physicochemicallaws applicable to such surfa,ces. Consequent-ly, the criterion of whether a particular ionflux obeys the phys,icochemical laws of aque-ous solutions or of reactions a,t surfaces isuseful in identifying the flux as passive orae.tive.17' 18 A detailed consideration of thisand other criteria of a.ctive and passive trans-port, as well as of the carrier hypothesis ofactive transport, is, beyond the scope of thisarticle. It is, however, instructive to point outcertain other characteristics, of the energy-consuming "uphill" fluxes. An inadequateoxygen supply, inhibition of respiration with
Circulation, Volume XXVI, October 1962
carbon monoxide or cyanide, failure to supplya substrate to act as a "metabolic fuel," orthe uncoupling of oxidation from phosphory-lation with dinitrophenol, all inhibit activetransport by depriving it of its energy supply.Similarly, the cardiac glycosides appear tointerfere with active transport, possibly bycompeting with the ion transported for thesite of combination with the carrier mole-cule.'9 20 Regardless of the mechanism, inter-ference with active transport reduces or abol-ishes the ea-pa,city of the cell to take up K andextrude Na in an "uphill" direction at a ratesufficient to compensate for the "downhill"diffus;ion of the-se ions. The consequent, netcellular K loss and Na uptake result in a de-creased intracellular K concentration and anincreased intracellular Na concentration.
The Action PotentialThe discussion to this point has. dealt with
the electrical potential difference during thesteady state prevailing in quiescent heartmuscle. It is expedient to consider next thedisturbances, of the steady state called actionpotentials, the clinical counterparts of whichare the QRST complexes of the electrocardio-gramn. The experimental observations, ob-tained with the intracellular microelectrodetechnic, are comparatively simple (fig. 7).The electrical potential difference across thecell membrane of the resting mammalian ven-tricular muscle cell bathed in solutions havingthe same K concentration as that of plasmais between 80 and 90 millivolts, the interiorof the resting cell being negative with respectto the extracellular solution. The action po-tential is propagated along the length of theheart muscle-cell membrane. The electricalpotential difference across. the membrane isobserved to revers,e its electrical sign, chang-ing from its resting value of -80 to -90 mVto a value of +20 to +40 mV, the cell interiorbecoming positive with respect to the extra-cellular solution. After the initial reversal,the potential difference falls to its, restingvalue with a delayed time course that resultsin the characteristic plateau configuration ofthe repolarization phase. It is instructive to
589
by guest on June 4, 2018http://circ.ahajournals.org/
Dow
nloaded from
PAGE
+20
0
POTENTIAL -20DIFFERENCE O
3
(millivolts) -40 -
-60
-80 4
-100 - 200 MILLISECONDS -1Figure 7
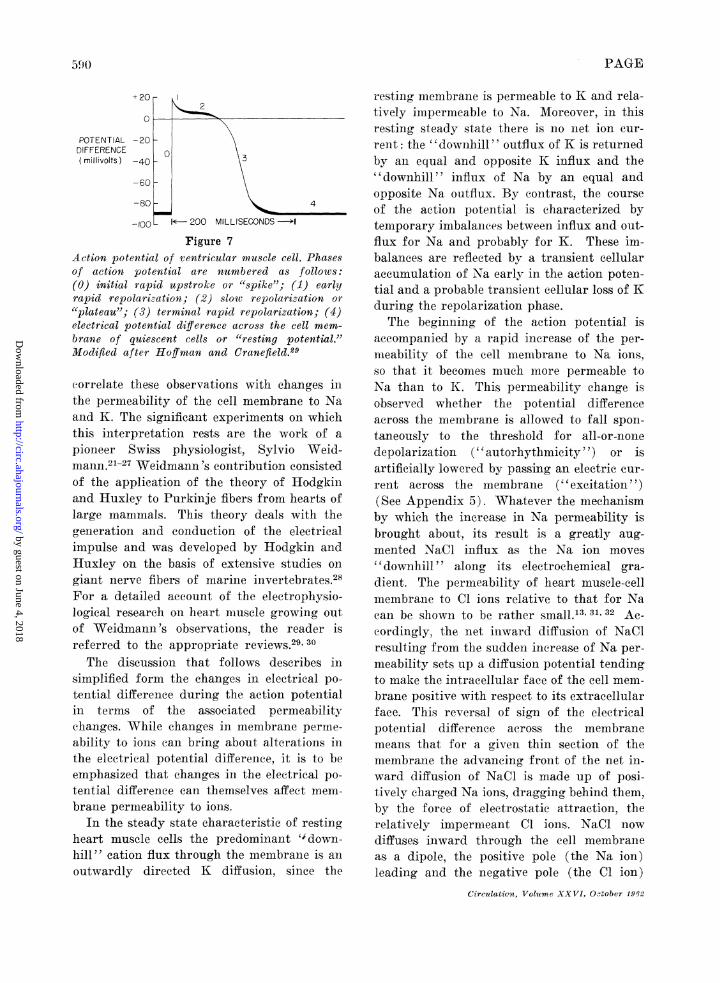
Action potential of ventricular muscle cell. Phasesof action potential are numbered as follows:(0) initial rapid upstroke or "spike"; (1) earlyrapid repolarization; (2) slow repolarization or"{plateau"; (3) terminal rapid repolarization; (4)electrical potential difference across the cell mem-brane of quiescent cells or "resting potential."Modified after Hoffman and Cranefield.29
eorrelate these observations with changes inthe permeability of the cell membrane to Naand K. The significant experiments on whichthis interpretation rests are the work of apioneer Swiss physiologist, Sylvio Weid-mann.21-27 Weidmann 's contribution consistedof the application of the theory of Hodgkiniand Huxley to Purkinje fibers from hearts oflarge imanmmals. This theory deals with thegeneration and conduction of the electricalimpulse and was developed by Hodgkin andHuxley on the basis of extensive studies ongiant nerve fibers of marine invertebrates.28For a detailed account of the electrophysio-logical research on heart muscle growing outof Weidmaun 's observations, the reader isreferred to the appropriate reviews.29 30The discussion that follows describes in
simplified formn, the changes, in electrical po-tential difference during the action potentialin terms of the ass,ociated permeabilitychanges. While changes in membrane pernme-ability to ions can bring a-bout alterations inthe electrical potential difference, it is to beemphasized that changes. in the electrical po-tential difference can themselves affect mem-brane permeability to ions.
In the steady state characteristic of restingheart muscle cells the predominant " down-hill" cation flux through the membrane is anoutwardly directed K diffusion, since the
resting nmembrane is permeable to K and rela-tively impermeable to Na. Moreover, in thisresting steady state there is no net ion cur-rent: the "downhill" outflux of K is returnedby an equal and opposite K influx and the"downhill" influx of Na by an equal andopposite Na outflux. By contrast, the courseof the action potential is characterized bytemporary imbalances between influx and out-flux for Na and probably for K. These im-balances are reflected by a transient cellularaccumulation of Na early in the action poten-tial and a probable transient cellular loss of Kduring the repolarization phase.The beginning of the action potential is
accompanied by a rapid increase of the per-nieability of the cell membrane to Na, ions,so that it beconmes much miore permeable toNa thain to K. This permeability change isobserved whether the potential differenceacross the membrane is allowed to fall spon-taneously to the threshold for all-or-nonedepolarizat,ion (" autorhythmicity ") or isartificially lowered by passing an electric cur-rent across the membrane ("excitation "(See Appendix 5). Whatever the mechanismby which the increase in Na permeability isbrought about, its result is a greatly aug-mented Na,Cl influx as the Na ion moves"downhill ' along its, electrochemical gra-dient. The permeability of heart muscle-cellmemtbrane to Cl ions relative to that for Nacan be shown to be rather small.'3 3, 32 Ac-cordingly, the net inward diffusion of NaClresulting from the sudden increase of Na per-meability sets up a diffusion potential tendingto make the intracellular face of the cell mem-brane positive with respect to its extracellularface. This reversal of sign of the electricalpotential difference across the membranemneans that for a given thin section of themembrane the advancing front of the net in-ward diffusion of Na.Cl is made up of posi-tively charged Na ions, dragging behind them,by the force of electrostatic at-traction, therelatively impermeant Cl ions.. NaCl nowdiffuses inward through the cell membraneas, a dipole, the positive pole (the Na ion)leading and the negative pole (the Cl iona)
Circulation. Volume XXVI, 0O-ober 1962
9O7(
by guest on June 4, 2018http://circ.ahajournals.org/
Dow
nloaded from
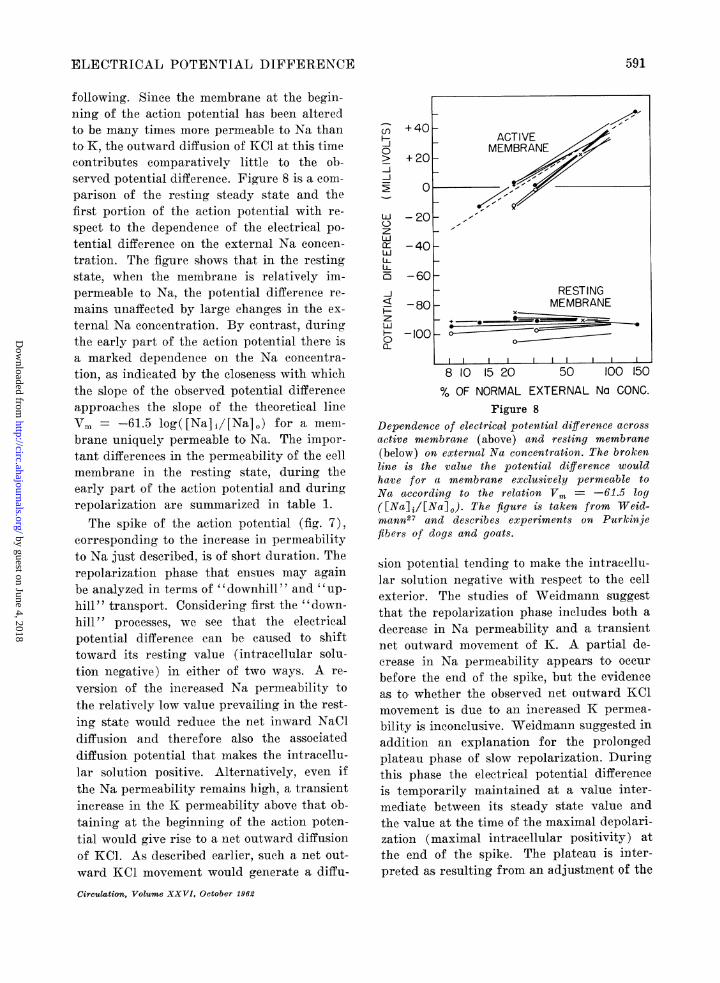
ELECTRICAL POTENTIAL DIFFERENCE
f'ollowing. Since the membrane at the begin-ning of the action potential has been alteredto be many times more permeable to Na thanto K, the outward diffusion of KCl at this timecontributes eomparatively little to the ob-served potential difference. Figure 8 is a com-parison of the res.ting steady state and thefirst portion of the acetion potential with re-spect. to the dependence of the electrical po-tential difference on the external Na concen-t.ration. The figure shows that in the restingstate, when the membrane is relatively im-permeable to Na, the potential difference re-mains unaffected by large changes in the ex-ternal Na concentration. By contrast, duringthe early part of the action potential there isa marked dependence on the Na concentra-tion, as indicated by the closeness with whichthe slope of the observed potential differenceapproaches the slope of the theoretical lineVm = -61.5 log( [Na] i/[Na] o) for a. men-brane uniquely permeable to, Na. The impor-t.ant differences in the permeability of the cellmembrane in the resting state, during theearly part of the action potential and duringrepolarization are summarized in table 1.The spike of the action potential (fig. 7),
corresponding to the increase in permeabilityto Na just described, is of short duration. Therepolarization pha,se that ensues may againbe analyzed in terms of "downhill" and "up-hill" transport. Considering first the "down-hill" processes., we s.ee that the electricalpotential difference can be caused to shifttoward its resting value (intracellular solu-tion negative) in either of two ways. A re-version of the increased Na pernmeability tot,he relatively low value preva,iling in the rest-ing state would reduce the net inward NaCldiffusion and therefore also the associateddiffusion. potential that, makes the intracellu-lar solution positive. Alternatively, even ifthe Na permeability remains high, a transientinerea-se in the K permeability above that ob-taining at the beginning of the action poten-tial would give rise to a net outward diffusionof KCl. As, described earlier, such a net out-ward KCl movement would generate a diffu-Circulation, Volume XXVI, October 1962
co+40-H ~~~~ACTIVE+20 ~MEMBRANE> +20 ,
uj - 20_ , ,z _LU -4011o -60 -
2 - RESTING- -80 - MEMBRANE
LUi
C -1005 6
8 10 15 20 50 100 150
% OF NORMAL EXTERNAL Na CONC.Figure 8
Dependence of electrical potential difference acrossactive membrane (above) and resting membrane(below) on external Na concentration. The brokenline is the value the potential difference wouldhave for a membrane exclusively permeable toNa according to the relation Vm, = -61.5 log([Na]j/[Na] j. The figure is taken from TVeid-mann27 and describes experiments on Purkinjefibers of dogs and goats.
sion potentia,l tending to make the intracellu-la,r solution negative with respect t,o the cellexterior. The studies of Weidmann suggestthat the repolarization phase includes both adecreas.e in Na, permeability and a, transientnet. outward movement of K. A partial de-crease in Na permeability appears, to occurbefore the end of the spike, but the evidenceas, to whether the observed net outward KClmovement is due to an increased K permea-bility is inconclusive. Weidmann suggested inaddition an explanation for the prolongedplateau phase of slow repolarization. Duringthis phase the electrical potential differenceis temporarily maintained at a value inter-mediate between its steady state value andthe value at the time of the maximal depolari-zation (maximal intracellular positivity) atthe end of the spike. The plateau is inter-preted as resulting from an adjustment of the
591
by guest on June 4, 2018http://circ.ahajournals.org/
Dow
nloaded from

PAGE
-~~~~~44-1 rc4
-~~~ra~~~~~~~-~~C
ct~~~
bcc
~~~~4-
S~ ~ ~ ~ ~~4
ci)
z+-aD.0
44-a C.
0 S: .e e~
9 N) c" P:D:-o a:)O c)
C) C) -4-- CZA= :S ':0 ;-
C)° .=~~~~~~~~~~~ r-4'4° h- U -4.0
4- )~
co -4¢ tmoQ
Ca
4
P4
m
,o
p- QAx AX P
Na and K permneabilities such that the diffu-sion- potential set up by the iniward net NaCldiffusion is temporarily approximnately equalto the diffusion potential of opposite electricalsign generated by the niet outward diffusionof KCI. Experimental evidence to supportthis hypothesis has so far proved difficult toobtain. The definitive determnination of themagnitude and time sequence of the changesin cation permeability durinig repolarization,anid their exact correlation with particul-arsegments of the action potential remain aschallenging unsolved problems."Uphill" transport during the repolariza-
tion phase includes the extrusion from the cellof Na that has elntered during the period ofincreased Na permeability, and the uptake bythe cell of K lost in any net outward K move-inents that may have occurred. The compara-tively short duration of these transient per-mileability changes, as well as the relativelvlow inembrane permeability to Cl, probablykeeps cellular Na accumulationi small and lim-its cellular K loss. Nonietheless, the Na gainanid K loss must be reversed by transportagainst the electrocheinical gradient. For thisreasomi the portions of the action potentialassociated with restoration of the steady stateshould be sensitive to interference with theactive transport mechanisul, whether by oxy-gen lack, metabolic inhibitors, or digitalisglycosides. It is not known at what point inthe repolarization sequence "uphill" trans-port processes become active, or whether res-toration of the steady state is complete at thetime the electrical potential difference hasreturned to its resting value.The transient net ion movements associated
with the action potential should be accom-pamnied by net water movements, and thereforeby changes in cell volume. Moreover, com-plexities in the structure of the cell mem-brane33 and in the biophysical properties ofthe extracellular regions of heart muscle7raise the possibility of non-uniform ion dis-tributions localized near the cell surface.When methods for the experimental measure-ment of such distributions are developed, it is
Circulation, Volume XXVI, October 1.962
0
0
ct0
r.d0
02Cc)
cd
"ci
0
a
0
ccci
I Sci:
0O
0*t
5(9
cQCc
ciz
,_d
P.
-.
_ac
a.24
a
a
as
ai
*
N
Cd
C) )_.. Q~
0op
"-c
as W co"-
bc0 by guest on June 4, 2018
http://circ.ahajournals.org/D
ownloaded from

ELECTRICAL POTENTIAL DIFFERENCE
to be expected that the elemenitary scheme pre-sented here may require modification.
SummaryThe electrica.l potential difference across
the cell membrane of heart muscle cells is adiffusion potential aris,ing from the interactionof fixed charges. within the membrane poreswith NaCl and KCI diffusing into and out ofthe cell. The magnitude and sign of this po-tential difference depend on the relative per-meabilities, of the membrane to K, Na, and Cl.The resting membrane is predominantly per-mea.ble to K, while the action potential ischaracterized by a transient increase in Napermeability. An intracellular solution ofhigh K concentration and low Na concentra-tion is separated by the cell membrane froman extracellular solution having a low K con-centration and a high Na concentration. Theseconcentration differences are maintained byactive transport processes. that utilize theenergy derived from cellular metabolic reac-tions to transport Na and K "uphill" againstelectrical and concentration gradients.
References1. TEORELL, T.: Transport processes and electrical
phenomena in ionic membranes. Progress inBiophysics 3: 305, 1953.
2. KEDEM, O., AND KATCHALSKY, A.: A physicalinterpretation of the phenomenologic coeffici-ents of membrane permeability. J. Gen.Physiol. 45: 143, 1961.
3. SOLLNER, K.: The electrochemistry of porousmembranes. In Electrochemistry in Biologyand Medicine, (T. Shedlovsky, editor), NewvYork, John Wiley and Sons, 1955, p. 33.
4. HELFFERICH, F.: Tonenaustauseher, Volume I,chapter 8: Ionenaustauschermembranen. Wein-heim/Bergstr. (Germany), 1959, p. 305.
5. MEARES, P., AND USSING, H. H.: The fluxes ofsodium and chloride ions across a cationexchange resin membrane. Part 1-Effect ofa concentration gradient. Transactions of theFaraday Society 55: 142, 1959.
6. MEARES, P., AND USSING, H. H.: The fluxes ofsodium and chloride ions across a cationexchange resin membrane. Part 2-Diffusionwith electric current. Transactions of theFaraday Society 55: 244, 1959.
7. PAGE, E.: Cat heart muscle in vitro. III. Theextracellular space. J. Gen Physiol. 1962. Inpress.
Circulation, Volume XXVI, October 1962
8. FOLCH, J., LEES, M., AND SLOANE-STANLEY,G. H.: The role of acidic lipids in the electro-lyte balance of the nervous system of mam-mals. In Metabolism of the Nervous System(D. Richter, editor). New York, PergamonPress, 1957, p. 174.
9. USSING, H. H.: The Alkali Metal Ions inBiology. I. The Alkali Metal Ions in IsolatedSystems and Tissues. Handbuch der Experi-mentellen Pharmakologie, Ergainzungswerk,Berlin, Springer Verlag, 1960.
10. DUNHAM, E. T., AND GLYNN, I. M.: Adenosine-triphosphatase activity and the active move-ments of alkali metal ions. J. Physiol. 156:274, 1961.
11. SKOU, J. C.: The relationship of a (Mg2+ andNa+) -activated, KR-stimulated enzyme or en-zyme system to the active, linked transportof Na4 and K4 across the cell membrane.In Membrane Transport and Metabolism (A.Kleinzeller and A. Kotyk, editors). New York,Academic Press, 1961, p. 228.
12. PAGE, E., AND SOLOMION, A. K.: Cat heart musclein vitro. I. Cell volumes and intracellularconcentrations in papillary muscle. J. Gen.Physiol. 44: 327, 1960.
13. PAGE, E.: Cat heart muscle in vitro. II. Thesteady state resting potential difference inquiescent papillary muscle. J. Gen. Physiol.1962. In press.
14. SOLOMON, A. K.: Compartmenital methods ofkinetic analysis. In Mineral Metabolism, Vol-ume I, Part A. New York, Academic Press,1960, p. 119.
15. USSING, H. H.: The distinction by means oftracers between active travsport and diffusion.Acta physiol. Scandinav. 19: 43, 1949.
16. HARRIS, E. J.: Transport and Accumulation inBiological Systems. Ed. 2. New York, AcademicPress, 1960.
17. KEDEM, O.: Criteria of active transport. InMembrane Transport and Metabolism (A.Kleinzeller and A. Kotyk, Editors). NewYork, Academic Press, 1961, p. 87.
18. GLYNN, I. M.: The ionic permeability of th)ered cell membrane. Progress in Biophysics8: 241, 1957.
19. SOLOMnON, A. K., GILL, T. J., AND GOLD, G. L.:The kinetics of cardiac glycoside inhibitionof potassium transport in human erythrocytes.J. Gen. Physiol. 40: 327, 1957.
20. GLYNN, I. M.: The action of cardiac glyco-sides on sodium and potassium movements inhuman red cells. J. Physiol. 136: 148, 1957.
21. DRAPER, M. H., AND WEIDMANN, S.: Cardiacresting and action potentials recorded withan intracellular electrode. J. Physiol. 115:74, 1951.
593
by guest on June 4, 2018http://circ.ahajournals.org/
Dow
nloaded from

PAGE
22. WEIDMANN, S.: Effect of current flow on themembrane potential of cardiac muscle. J.Physiol. 115: 227, 1951.
23. WEIDMANN, S.: The electrical constants ofPurkinje fibers. J. Physiol. 118: 348, 1952.
24. COREABOEUF, E., AND WEIDMANN, S.: Tenmper a-ture effects on the electrical activity ofPurkinje fibers. Helvet. physiol. acta 12: 32,1954.
25. WEIDMANN, S.: The effect of the cardiac mem-brane potential on the rapid availability ofthe sodium-carrying system. J. Physiol. 127:213, 1955.
26. WEIDMANN, S.: Effects of calcium ions andlocal anesthetics on electrical properties ofPurkinje fibers. J. Physiol. 129: 568, 1955.
27. 1NEIDMNN, S.: Elektrophysiologie der Herz-nmuskelfaser. Berin, Mediziniseher Verlag HansHuber, 1956.
28. HODGKIN, A. L.: The Croonian Lecture: Ionicatovements anid electrical activity in gianterive fibers. Proc. Roy. Soc., Ser. B. 148:
1, 1958.29. HOFFMAN, B. F., AND CRXNEFIELD, P. F.: Elec-
trophysiology of the Heart. New York, McGrawHill Book Co., 1960.
30. CORABOET Fi, E.: Aspects cellulaires de 1'6lectro-genese cardiaque clez les vrte6br6s. J. Physiol.,Paris 52: 323, 1960.
31. HUTTER, 0. F., AND NOBLE, D.: Anion con-ductance of cardiac nmusele. J. Physiol. 157:335, 1961.
32. CARMEI.ET, E. E.: Chloride ions arid the mem-brane potential of Purkinje fibers. J. Physiol.156: 375, 1961.
33. SiMpsoN, F. 0., AND OERTELIS, S. J.: The finestructure of sheep inyocardial cells; sarcolem-mal invaginiations and the transverse tubularsystem . J. Cell. Biol. 12: 91, 1962.
34. STERN, K. H., XND Amis, E. S.: Iontic size.ChemB. Reviews 59: 1, 1959.
35. ROBINSON, R. A., A-ND STOKES, R. H.: ElectrolyteSolutions. Ed. 2. New York, Academic Press,Inc., 1959, p. 11.
36. EISENATAN, G., RUDIN, D. O., AND CASBY, J. U.:Glass electrode for measuring sodium ion.Science 126: 831, 1957.
37. IHINKE, J. A. M.: The measurement of sodiumanld potassium activities in squid axon bymeanls of cation-selective glass micro-electrodes.J. Physiol. 156: 314, 1961.
38. GOLDMAN, D. E.: Potential, impedance andrectification in membranes. J. Gen. Physiol.27: 37, 1943.
39. HODGKIN, A. L., AND KATZ, B.: The effect ofsodium on the electrical activity of the giantaxon of the squid. J. Physiol. 108: 37, 1949.
Appendices1. The electrical potential difference across a
membrane may include, in addition to the diffu-sion potential described above, a contribution fromso-called Donnan potentials due to fixed ehargesat the boundary of the nmnembrane with the solu-tions on either side.1 In order to simplify thepresent discussion, such Donnan potentials areconsidered negligible.
2. The use of the K concentrations in thisequation is an approximation. To be exact, theconcentration terms [K] should be multipliedby a factor, y, called an activity coefficient, theproduct y [K] being defined as the activity ofK in the solution. The activity may be thoughtof as an effective conicentration, i.e., as that frac-tion of the total K concentration which is effec-tive in producing an electrical potential difference.At the concentrations encountered in tissue fluids,the activity of ions is always less than theirconcentration. The usual flamle photometric de-termiination of K and Na nmeasures concentrations.The K and Na activities of solutions may, how-ever, be measured directly with special K and Naglass electrodes,36' 3 just as the activity of hydro-gen ion is conventionally measured with a pHglass electrode.
3. In deciding for a given biological membranewhether the transport of a charged particle likethe K ion is an "aetive," "uphill" or energy-consuming process rather than a "passive," "down-hill" or spontaneous process it is necessary totake into consideration not only the gradient ofconcentration but also the gradient of electricalpotential. Thus, movement of K from an extra-eellular coneentration of 5 mM into a cellularsolution at a concentration of 200 mM is "uphill"with respect to the concentration gradient; sincethe cellular solution is at a negative electricalpotential coiipared with the extracellular solution,the transport into the cell of positively chargedparticles like the K ion is, however, "downhill"with respect to the gradient of potential. Whenboth concentration and potential gradients areconsidered together for the siniultaneous move-rnents of K and Na, it appears likely that boththese ions are actively transported in mammialianheart muscle. Throughout this paper the con-ventional assunmption that the Cl ion is passivelytransported is followed. A consequence of thisassumption is that in the steady state at 370C the distribution of Cl ions between the intra-cellular and extracellular solutions can be predictedfrom the electrical potential difference across thecell membrane by the relation, Vm = -61.5 log([Cl] 0/[Cl] i). For a discussion of the validityof this assunmiption, see Page.7
Circulation, Volume XXVI, October 1962
594
by guest on June 4, 2018http://circ.ahajournals.org/
Dow
nloaded from

ELECTRICAL POTENTIAL DIFFERENCE
4. An approximationi com-monly used to expressthese relative contributions is the equation de-veloped by Goldman38 and by Hodgkin and Katz.39If movements of Cl may be assumed to be pas-sive, this equation takes the form,
Vm = -61.5 log pK [K]i + PNa [Na]
in which Vm is the electrical potential differenceacross the cell membrane at 370 C, [K] i and[Na]i are the intracellular K and Na concentra-tions, [K]0 and [Na]0 the corresponding extra-cellular concentrations, and PK and PNa the per-meabilities of the membrane to K and Na, respect-tively.
5. Changes in the a.pparently small electricalpotential differences across cell mnembranes ofexcitable tissues may alter the permeability of themembrane to ions because the short distancesacross which such potential differences exist setup relatively large forces. The force aeting on acharged particle in the membrane is called an
electric field and may be approximated by theelectrical potential difference divided by the thick-ness of the membrane. For an electrical potentialdifference of 100 mV across a cell membrane100 A in thickness this calculation gives a forceof 100,000 volts per centimeter. In an electric fieldof this magnitude positively charged moleculeswithin the substance of the membrane will tendto move toward the negatively charged face ofthe membrane and negatively charged moleculeswill attempt to migrate to the positively chargedface. Since the charged molecules making up thesubstance of the memnbrane are fixed, they areprevented from moving in these directions. Suchmolecules, however, may have a certain amountof freedom to rotate about an axis. This freedompermits them to orient themselves so that theirfixed charge groups tend to point toward theface of the memubrane toward which they wouldmigrate if they were free to move. Variations inthe electrical potential difference may alter thisorientation of fixed charges, suggesting a pos-sible mechanism for permeability clhanges.
If our feeling constantly puts the question why, our reason shows us that only thequestion how is within our range; for the moment, then, only the question how concernsmen of science and experimenters. If we cannot know why opium and its alkaloids putus to sleep, we can learn the mechanism of sleep and know how opium or its ingredientsputs us to sleep; for sleep takes place only because an active substance enters intocontact with certain organic substances which it changes. Learning these changes willgive us the means of producing or preventing sleep, and we shall be able to act on thephenomenon and regulate it at pleasure.-CLAUDE BERNARD. An Introduction to the Studyof Experimental Medicine. New York, The Macmillan Company, 1927, p. 82.
Circulation, Volume XXVI. October 1962
595
by guest on June 4, 2018http://circ.ahajournals.org/
Dow
nloaded from

ERNEST PAGEBiophysical Considerations
The Electrical Potential Difference Across the Cell Membrane of Heart Muscle:
Print ISSN: 0009-7322. Online ISSN: 1524-4539 Copyright © 1962 American Heart Association, Inc. All rights reserved.
is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Circulation doi: 10.1161/01.CIR.26.4.582
1962;26:582-595Circulation.
http://circ.ahajournals.org/content/26/4/582located on the World Wide Web at:
The online version of this article, along with updated information and services, is
http://circ.ahajournals.org//subscriptions/
is online at: Circulation Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. and Rights Question and Answer
Permissionsthe Web page under Services. Further information about this process is available in thewhich permission is being requested is located, click Request Permissions in the middle column ofClearance Center, not the Editorial Office. Once the online version of the published article for
can be obtained via RightsLink, a service of the CopyrightCirculationoriginally published in Requests for permissions to reproduce figures, tables, or portions of articlesPermissions:
by guest on June 4, 2018http://circ.ahajournals.org/
Dow
nloaded from









