Embed Size (px)
Citation preview

JOURNAL OF VIROLOGY, OCt. 1970, p. 406-413Copyright @ 1970 American Society for Microbiology
Vol. 6, No. 4Printed in U.S.A
Soluble Components of Adenovirus Type 81ANNA LENGYEL AND ISTVAN NASZ
Institute of Microbiology, Semmelweis University of Medicine, Budapest, Hungary
Received for publication 16 June 1970
Soluble components of type 8 adenovirus have been studied. Four different com-ponents were isolated by anion-exchange chromatography and purified by furtherchromatographic procedures, by zonal centrifugation, and by erythrocyte absorp-tion and elution. The four components exhibited the following characteristics. (i)Fiber antigen was trypsin-resistant and functioned as incomplete hemagglutinin(agglutinated rat and human erythrocytes only in the presence of certain types ofadenovirus antisera). (ii) The penton was trypsin-sensitive, exerted a cytotoxiceffect, and also showed incomplete hemagglutination, being active in the presenceof a majority of heterotypic adenovirus antisera studied. (iii) The group-specifichexon antigen reacted in complement fixation reaction and gel precipitation withsera prepared against other types of adenoviruses, besides showing characteristicsindicating the presence of a type-specific antigen component. (iv) The soluble com-plete hemagglutinin was trypsin-sensitive, displayed cytotoxic effect, adsorbed easilyto human and rat erythrocytes, and could be eluted from them by means of receptor-destroying enzyme. The three hemagglutinins of adenovirus type 8 proved to behighly unstable, and their demonstration was only successful by using a large quan-tity of freshly prepared concentrated virus material. Considering these conditions, amethod was developed for their concentration and purification.
The antigenic structure of several adenovirustypes has recently been elucidated by a numberof investigators (4, 5, 20, 22, 24, 25, 34), and yetinformation concerning the soluble componentsof adenovirus type 8 is very scarce (1).Adenovirus type 8 appears to diverge from
other adenovirus types in some biological aspects(16); for example, it is markedly reduced in in-fectivity. On the other hand, it is the causativeagent of a characteristic infectious disease, epi-demic keratoconjunctivitis (8, 17). On the basisof its deoxyribonucleic acid (DNA) compositionit was suggested that adenovirus type 8 may beoncogenic (29), although tumor induction withthis type was until now successful only in a singlecase (33).
Considering these features, it appeared to beof some interest to carry out a detailed study ofadenovirus type 8, and, despite the difficulties inits culturing, attempts were made to separate itssoluble antigens to compare their characteristicswith the known data of other adenovirus types.
MATERIALS AND METHODSTissue cultures and virus strains. HEp-2, HeLa, and
MAS-A permanent human cell lines, cultured as de-
1These experiments were carried out partly in the Microbio-logical Research Group of the Hungarian Academy of Sciences,Budapest.
scribed in previous reports (10, 17, 30), were used forvirus propagation and for studying the cytopathiceffect (CPE).The prototype strain of adenovirus type 8, strain
"Ijima" (kindly provided by R. Wigand and E.Jawetz), and two strains of adenovirus type 8 isolatedfrom epidemic keratoconjunctivitis cases in Budapest(17) and in Szeged (2) were included in our experi-ments. The four strains did not show any essentialdifferences.
Infected cells showing complete degeneration weresedimented by low-speed centrifugation, collected in0.01 M tris(hydroxymethyl)aminomethane (Tris)-hy-drochloride buffer (pH 7.6), frozen and thawed five tosix times, and disintegrated in a homogenizer (Meas-uring & Scientific Equipment, London, England). Thecell debris was removed by centrifugation, and thevirus pool obtained was stored at 4 C. The concen-trated virus material was treated in certain cases withfluorocarbon (Arcton 113), and the nonsoluble com-ponents were sometimes removed by a three-times-repeated, 1-hr-long run in the ultracentrifuge at Pi 5.4(23,000 X g). These treatments, however, damaged aconsiderable part of the soluble components.Immune sera. Rabbits were injected intravenously
with complete virus and intramuscularly with purifiedcomponents, adding Freund's complete adjuvant tothe material, as described earlier (12, 17, 19).
Hemagglutination, incomplete hemagglutination, andhemagglutination-inhibition (H) techniques. These ex-aminations were performed in the Takdtsy micro-titrator (32) by using 0.75% mixed rat or human 0
406
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 21
Dec
embe
r 20
21 b
y 5.
154.
57.1
92.

COMPONENTS OF ADENOVIRUS TYPE 8
erythrocytes. The hemagglutination enhancement(HE) titer of the heterotypic adenovirus antisera wasdetermined in chess-board titration, and 2 HE unitswas used for incomplete hemagglutination.To study the complete hemagglutinin (HA) in the
HI reaction, 8 units of antigen was allowed to incubatewith the immune serum for 30 min, and the erythro-cytes were added subsequently. To examine the in-complete HA, the antigen was subjected to prior in-cubation with the appropriate heterologous antiserumfor 1 hr before the HI reaction was performed in thesame way.
Complement fixation (CF) test. The reaction wasperformed in the Takatsy microtitrator by using 2units of complement and incubating overnight at 4 C.
Gel-diffusion precipitation. These tests were carriedout with the two-dimensional method of Ouchterlony;the procedure was described previously (19).
Ultraviolet (UV) absorption spectrum and proteincontent determination. The UV absorption spectrumwas determined in a Pye Unicam SP 800 spectropho-tometer at between 190 and 450 nm. The protein con-centration was determined by the procedure of Lowryet al. (15) by using bovine serum albumin as astandard.
Separation by ultracentrifugation. Experiments werecarried out in a Superspeed 50 ultracentrifuge (Meas-uring & Scientific Equipment). To separate the solubleand nonsoluble components, the virus material wascentrifuged three times for 1 hr at Pi 5.4 (23,000 X g)on a CsCl cushion (density, 1.4 g/ml) or was centri-fuged for 90 min on a discontinuous CsCl gradient(density, 1.4 and 1.2 g/ml) at Pi 7 (42,000 X g).Equilibrium centrifugation in CsCl was performed for24 hr at Pi 13.7 (83,000 X g). Some of the prepara-tions were subjected to zonal centrifugation for 5.5hr in a linear gradient of 5 to 20% (w/v) sucrose atPi 7.5 (45,000 X g).
Anion-exchange chromatography. Diethylamino-ethyl (DEAE)-Sephadex A-50 and A-25 (PharmaciaFine Chemicals) columns were used to separate thevirus components in anion-exchange chromatography.The results were more favorable with the A-25 col-umns; thus the majority ofour experiments were carriedout with this exchanger, in 0.04 M Tris-hydrochloridebuffer, pH 8.0. The components retained by the col-umn were eluted either stepwise with the same buffer,containing increasing concentrations of NaCl, or byintroducing a linear NaCl gradient. Fractions werecollected with an LKB fraction collector combinedwith a Uvicord I, and, in the case of gradient elution,the NaCl content of the fractions was checked in arefractometer.Enzyme treatments. For trypsin treatment, five
times crystallized trypsin (Serva) was added to thematerial to a final concentration of 0.1%. After incu-bating the mixture for 1 hr at 37 C, an equivalentquantity of soybean inhibitor (Serva) was added.
For receptor-destroying enzyme (RDE) treatment,the required dilution of standard cholera filtrate(Philips Roxane Inc.), determined by titration, wasadded to human erythrocytes, which had been pre-viously incubated with HA and subsequently centri-fuged. The HA-treated erythrocytes were incubated
for 2 hr with RDE and then removed from the eluateby low-speed centrifugation.
RESULTSTo separate the different soluble components of
adenovirus type 8, different techniques were com-bined. The first step was anion-exchange chro-matography.The fractions eluted from the columns were
subjected to the following procedures. The group-specific hexon antigen was determined by gelprecipitation or by CF reaction with antiseraprepared against the hexon antigens of adenovirustypes 5 and 8; the complete HA was demon-strated by hemagglutination with rat erythrocytes;and the two incomplete HA were revealed in thepresence of heterologous antisera. As adenovirustype 8 belongs to Rosen's subgroup II (31), itappeared to be necessary to apply a serum of theidentical subgroup (adenovirus type 13) and oneagainst a member of subgroup I (type 11).To study the early CPE, HeLa and MAS-A
cells were inoculated with 0.1 ml of the fractions.The UV absorption spectra of the fractions wererecorded as well, and their protein content wasdetermined occasionally.A chromatogram of adenovirus type 8 is pre-
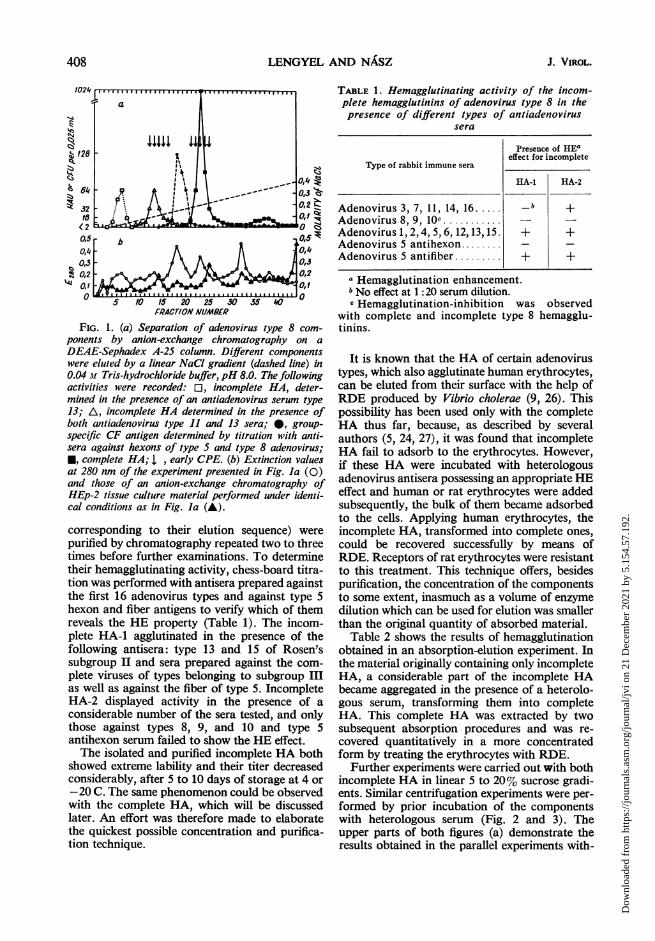
sented in Fig. 1. As noted, an incomplete HAwas the first to elute, being active with rat andhuman erythrocytes in the presence of antiadeno-virus type 13 serum. The elution of this compo-nent started to a slight degree, even withoutintroducing the NaCl, and was finished at a lowmolarity (0.03 to 0.04 M) of NaCl. The secondcomponent eluting was another incomplete HA,which agglutinated the erythrocytes in the pres-ence of both heterologous sera used in the experi-ments. This component showed a cytotoxic effectand was eluted at a concentration of NaCl of0.007 M.The hexon antigen was the third to elute, at a
concentration of NaCl of about 0.11 to 0.12 M,and was closely followed by the soluble completeHA, its elution being induced by a concentrationof NaCl of approximately 0.17 M. Fractions con-taining this component exhibited an early CPE.At the end of the chromatographic procedure, thehemagglutination activity increased slightly and asmall quantity of CF antigen appeared again,indicating the start of the elution of the non-soluble components.Four well-separated soluble components were
obtained in this manner, and, except for the firstone, there were certain overlaps. A detailed studyof these components required further purificationprocedures.
Incomplete HA. The two incomplete HA (desig-nated incomplete HA-1 and incomplete HA-2
407VOL. 6, 1960
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 21
Dec
embe
r 20
21 b
y 5.
154.
57.1
92.
LENGYEL AND NASZ
zaC
k 128
Ca.
64
3216<20,50,4
t 0,32
0,10'O
0,
0,.
00,O'
b ~~~~~~~~~~~~~0,
A
0,.
0l,
5 1O 15 20 25 30 55 410FRACrION NUMNEP
4
,2'
,5
,3
,2
FIG. 1. (a) Separation of adenovirus type 8 com-ponents by anion-exchange chromatography on aDEAE-Sephadex A-25 column. Different componentswere eluted by a linear NaCI gradient (dashed line) in0.04 M Tris-hydrochloride buffier, pH 8.0. The followingactivities were recorded: El, incomplete HA, deter-mined in the presence of an antiadenovirus serum type13; A, incomplete HA determined in the presence ofboth antiadenovirus type 11 and 13 sera; *, group-specific CF antigen determined by titrationi with anti-sera against hexons of type S and type 8 adenovirus;*, complete HA; I , early CPE. (b) Extinction valuesat 280 nm of the experiment presented in Fig. la (0)and those of an anion-exchange chromatography ofHEp-2 tissue culture material performed under identi-cal conditions as in Fig. la (A).
corresponding to their elution sequence) werepurified by chromatography repeated two to threetimes before further examinations. To determinetheir hemagglutinating activity, chess-board titra-tion was performed with antisera prepared againstthe first 16 adenovirus types and against type 5hexon and fiber antigens to verify which of themreveals the HE property (Table 1). The incom-plete HA-1 agglutinated in the presence of thefollowing antisera: type 13 and 15 of Rosen'ssubgroup II and sera prepared against the com-plete viruses of types belonging to subgroup IIIas well as against the fiber of type 5. IncompleteHA-2 displayed activity in the presence of aconsiderable number of the sera tested, and onlythose against types 8, 9, and 10 and type 5antihexon serum failed to show the HE effect.The isolated and purified incomplete HA both
showed extreme lability and their titer decreasedconsiderably, after 5 to 10 days of storage at 4 or-20 C. The same phenomenon could be observedwith the complete HA, which will be discussedlater. An effort was therefore made to elaboratethe quickest possible concentration and purifica-tion technique.
TABLE 1. Hemagglutinating activity of the incom-plete hemagglutinins of adenovirus type 8 in thepresence of different types of antiadenovirus
sera
Presence of HEaeffect for incomplete
Type of rabbit immune sera effec_forincopletHA-1 HA-2
Adenovirus 3, 7,11, 14, 16 b +Adenovirus 8, 9, lOc...........Adenovirus 1, 2, 4, 5, 6, 12,13,15.+ +Adenovirus 5 antihexon........Adenovirus 5 antifiber......... + +
a Hemagglutination enhancement.bNo effect at 1: 20 serum dilution.- Hemagglutination-inhibition was observed
with complete and incomplete type 8 hemagglu-tinins.
It is known that the HA of certain adenovirustypes, which also agglutinate human erythrocytes,can be eluted from their surface with the help ofRDE produced by Vibrio cholerae (9, 26). Thispossibility has been used only with the completeHA thus far, because, as described by severalauthors (5, 24, 27), it was found that incompleteHA fail to adsorb to the erythrocytes. However,if these HA were incubated with heterologousadenovirus antisera possessing an appropriate HEeffect and human or rat erythrocytes were addedsubsequently, the bulk of them became adsorbedto the cells. Applying human erythrocytes, theincomplete HA, transformed into complete ones,could be recovered successfully by means ofRDE. Receptors of rat erythrocytes were resistantto this treatment. This technique offers, besidespurification, the concentration of the componentsto some extent, inasmuch as a volume of enzymedilution which can be used for elution was smallerthan the original quantity of absorbed material.
Table 2 shows the results of hemagglutinationobtained in an absorption-elution experiment. Inthe material originally containing only incompleteHA, a considerable part of the incomplete HAbecame aggregated in the presence of a heterolo-gous serum, transforming them into completeHA. This complete HA was extracted by twosubsequent absorption procedures and was re-covered quantitatively in a more concentratedform by treating the erythrocytes with RDE.
Further experiments were carried out with bothincomplete HA in linear 5 to 20% sucrose gradi-ents. Similar centrifugation experiments were per-formed by prior incubation of the componentswith heterologous serum (Fig. 2 and 3). Theupper parts of both figures (a) demonstrate theresults obtained in the parallel experiments with-
408 J. VIROL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 21
Dec
embe
r 20
21 b
y 5.
154.
57.1
92.

COMPONENTS OF ADENOVIRUS TYPE 8
TABLE 2. Absorption-elution experiment with puri-fied incomplete hemagglutinin (HA)-2 of type 8adenovirus by using human erythrocytes and
receptor-destroying enzyme (RDE)
Treatment
None ..................
Antiadenovirus type 13serum.................
Absorption lb...........Absorption 2............Absorption 3............RDEC to cells of absorp-
tion 1.................RDE to cells of absorp-
tion 2.................
HA units per 0.025 ml of
CompleteI HA
4824
96
32
Incomplete HAtested in thepresence of
serum against
Adeno- Adeno-virus 13 virus 11
64 64
64 6424 244
128
48
16
8
114
2
(Za <2C.
16
8
4128 2
48 <2
a Less than 4.
h In each individual absorption, packed eryth-rocytes were added to a final concentration of10%. After incubation for 40 min, cells were re-moved by centrifugation and the supernatant wastested.
, A 1:4 dilution of cholera filtrate was added tothe packed erythrocytes in a volume correspond-ing to one-fourth of the original volume of theabsorbed material.
out adding heterologous serum, and the lowerparts (b) show the results obtained after incuba-tion with heterologous serum. The incompleteHA-2 also contained a small quantity of hexonantigen, its position being indicated in the figuresas well.As shown by Fig. 2 and 3, both incomplete HA,
after treatment with heterologous serum, reachedeven a higher concentration in the gradient be-cause of their aggregation. The connection pro-duced by the heterologous serum seems to berather unstable, as indicated by the HE effect ofthe heterologous sera with fractions 3 to 12 inFig. 3b.These findings suggest that the incomplete
HA-1 corresponds to the fiber of type 8 adeno-virus and HA-2 corresponds to the penton, aswith other types (21, 27). This is also supportedby the response of these components to trypsintreatment. Results of experiments with trypsintreatment are summarized in Table 3. Accordingto these data, the hemagglutinating activity of theincomplete HA-1, corresponding to the fiber, wasnot diminished by trypsin treatment; in fact, incertain experiments, similarly to other types (20),
5 1O 15- 720FTACTION NUMBLY
25
FIG. 2. Zonal centrifugation of the incomplete HA-Iofadenovirus type 8 in a linear 5 to 20% sucrose gradi-ent for 5.5 hr at Pi 7.5 (45.000 X g). Fractions were
collected dropwise through the bottom of the tube (left).Upper part (a) without heterologous serwn; lower part(b) after incubation with antiadenovirus type 13 serum.
Symbols correspond to those presented in Fig. 1.
an increase could even be observed. The activityof the penton, however, disappeared in the pres-
ence of type 11 serum together with its cytotoxiceffect. No changes were found in its hemaggluti-nating activity and cytotoxic effect after UVirradiation.Hexon antigen. The group-specific hexon anti-
gen was the third in the sequence of elution. Asshown by Fig. 1, this component was contami-nated by a certain amount of complete HA. Toeliminate the latter, erythrocyte absorption was
applied successfully. Thereafter, contaminationwith incomplete HA-2, corresponding to penton,was also found, with an overlap by the completeHA. The elimination of this factor was accom-plished by repeated absorptions, and occasionallyheterologous serum was added to the materialprior to adding the erythrocytes. The last absorp-tion procedure was followed by three or moresubsequent chromatographic treatments, and, ifno traces of either HA could be revealed in thefractions, they were considered to contain purehexon antigen.
Rabbits were immunized with hexon antigenprepared by several erythrocyte absorptions andby five-times-repeated anion-exchange chroma-tography. The serum reacted satisfactorily with
I 1 XI I I I I rII r
a0 0
_ ,z,~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~,,,,,,,,.,,,,,,,,, . ~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~b
a4_ ~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~.X
b
_=",a
VOL. 6, 1970 409
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 21
Dec
embe
r 20
21 b
y 5.
154.
57.1
92.

LENGYEL AND NASZ
:~~~~~~. IiIII'j
I'i
' 5 10 15 20FRACTION NVUM4EQ
25
FIG. 3. Zonal centrifugation of the incomplete HA-2of adenovirus type 8 under similar conditions as in Fig.2. Symbols correspond to those presented in Fig. 1, theonly difference being that group-specific hexon antigenwas tested in gel-diffusion precipitation test (0).
TABLE 3. Effect of trypsin treatment on the incom-plete hemagglutinins (HA) of adenovirus type 8
Designa-tion of in-completeHA
1
2
Trypsinatreat-ment
0+
+
HA units per 0.025 ml of
Com-pleteHA
Incomplete HA test-ed in the presenceof serum against
Adeno-virus 13
324864128
Adeno-virus 11
648
J. VIROL.
Earlycytopathic
effect
a Incubation with trypsin in a final concentra-tion of 0.1% for 1 hr at 37 C in a water bath.bLess than 4.
TABLE 4. Hemagglutination-inhibition (HI) testswith antiadenovirus sera and adenovirus type
8 antihexon immune serum
Reciprocals of HI titers ofAdeno- immune sera againstvirus Antigen prepntype Homologous
crude virus Type 8 hexon
8 Fiber 80 a8 Penton 1208 Complete HAb 3208 Virion 640 80 partial9 Crude virus 1,28010 Crude virus 64015 Crude virus 160
a Less than 1:10.bHemagglutinin.
different types of adenovirus strains in gel diffu-sion and CF tests up to the dilutions 1:128 and1:1,024, respectively. In neutralization tests, theserum failed to neutralize the CPE of adenovirustypes 5, 7, 9, and 10 but neutralized that of thehomologous type 8, up to the dilution 1:80.HI experiments were performed with the serum
prepared against the hexon antigen of type 8(Table 4). The serum failed to inhibit the hemag-glutination of three heterologous types, the sol-uble complete HA and both incomplete HA ofthe homologous type 8; however, it inhibitedpartially at a titer of 1:80 the type 8 virion, ob-tained by CsCl equilibrium centrifugation. Thispartial reactivity was revealed by the agglutina-tion of two-third of the erythrocytes only at thetiter mentioned. Table 4 indicates parallel titersof the sera against the complete adenovirus ofhomologous types.
Soluble complete HA. The complete HA was
the last in the sequence of elution, its elutionusually starting above 0.16 M NaCl and the maxi-mum being about 0.2 M NaCl (Fig. 1). Placed ontissue culture, this component also showed anearly CPE and was trypsin-sensitive both in re-spect to hemagglutinating activity and cytotoxiceffect. Results of trypsin treatment are presentedin Table 5. Thus, the HA lost its ability to causecomplete hemagglutination to a considerable ex-tent, and the majority of it transformed into in-complete HA corresponding to the fiber.The complete HA was easy to absorb with rat
and human erythrocytes and was eluted from thelatter with RDE (Table 6). It is noticeable that,while extracting the complete HA, an incompleteHA, covered up by the complete one, can betraced in these fractions besides the hexon con-tamination. The complete HA of type 8 adeno-virus was very unstable, and a four- to eightfoldtiter decrease could be observed within a few days
128
32
16
t 8
5 128
:Zt
32
16
8
' 2
410
I
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 21
Dec
embe
r 20
21 b
y 5.
154.
57.1
92.

COMPONENTS OF ADENOVIRUS TYPE 8
TABLE 5. Effect of trypsin treatment otn the purifiedsoluble complete hemagglutinint (HA) of
adenovirus type 8
HA units per 0.025 ml of
Trypsin Incomplete HA tested in Earlytreatmenta C l presence of serum against cytopathicComplete_________ effect
HAAdenovirus Adenovirus
13 11
No 768 768 768 +Yes 32 1,024 128
a Conditions similar to those presented inTable 3.
TABLE 6. Absorptionz-elution experimentts withfractions of anion-exchange chromatography
containinig soluble complete hemagglutinin(HA) of adenovirus type 8
HA units per 0.025 ml of
Incomplete HATreatment tested in presence Hexona
Complete of serum againstHA
Adeno- Adeno-vlrus 13 virus 11
No ............. 256 256 256 +Absorption lb ... -c 96 64 +Absorption 2...... 96 64 +RDE to cells of ab-
sorption ld. 384 384 384 -
a Determined in gel-diffusion precipitationtests with homologous and type 5 antihexon sera.bAbsorptions carried out as presented in
Table 2.c Less than 4.d A 1:4 dilution of cholera filtrate added to the
packed erythrocytes in a volume correspondingto half of the original volume of the absorbedmaterial.
both in the original fractions and in the RDEeluates. This finding supported the suggestionthat this incomplete HA resulted from the de-composition of the complete HA. However, bycollecting, concentrating, and submitting the frac-tions to repeated chromatography after the ab-sorption procedure, the position of this incom-plete HA remained unchanged and started toelute at 0.16 M NaCl (Fig. 4).
Results of the zonal centrifugation of an RDEeluate of complete HA (Fig. 5) show likewise theinstability of this component. The presence ofcomplete HA equally in both the upper and lowerregions of the gradient indicates the possibilitythat it consists of rapidly and slowly sedimenting
16 AA--027
5 01 1
FRACTION NUMACP
FIG. 4. Recovery of different biological aclivities offractions containing complete HA of adenovirus type 8,from which the complete HA had been removed byerythrocyte absorption. The conditions of the anion-exchange chromatography and symbols correspond tothose presented in Fig. 1.
32
'2 5
FPACT/ON NUMB/IA'FIG. 5. Zonal centrifugation of the complete HA of
adenovirus type 8 under similar conditions as in Fig. 2.Symbols as in Fig. 1.
components. The localization of incomplete HA-1and HA-2 indicates that the complete HA dis-integrated as early as during the few hours ofcentrifugation; this explains the phenomenon thatboth kinds of incomplete HA are present near thebottom of the tube, localized otherwise after a5.5-hr run in the upper regions of the tube, asshown in Fig. 2 and 3.
DISCUSSIONOur findings concerning the elution sequence of
adenovirus type 8 components in anion-exchangechromatography agree with those of other au-thors, who studied other members of Rosen'ssubgroup 11 (5, 6, 20, 24). A single reference is tobe found in the literature dealing with the anion-exchange chromatography of adenovirus type 8(7). The first successful separation of four differ-ent soluble components of this type was carriedout in our experiments. The common character-istic of these components, except the hexon, istheir extreme instability.The characteristics of the fiber antigen ob-
served in our experiments, i.e., the elution at alow molarity of NaCl, the trypsin resistance, andthe hemagglutinating activity displayed in the
411VOL. 6, 1970
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 21
Dec
embe
r 20
21 b
y 5.
154.
57.1
92.

LENGYEL AND NASZ
presence of heterologous adenovirus antisera be-longing to Rosen's subgroup II and III tally withthe characteristics observed with the types 9, 15,9-15, and 19 of the identical subgroup (5, 20, 24).
It was shown by several investigators (22, 27)that the penton and its aggregate the dodecon areresponsible for the early CPE exhibited by adeno-virus types of subgroups I and III. Early CPEwas also observed in our experiments with type 8of subgroup II, occurring likewise in the presenceof the two soluble components mentioned. Thiseffect of the penton was very unstable and disap-peared even before the decay of incomplete he-magglutinating activity (within approximately 7days). The trypsin sensitivity and the resistance toUV irradiation of the early CPE prove its toxicnature and contradict the possibility that it mightbe a phenomenon related to virus multiplication.Our observations on the penton component
indicate that every antiserum against the first 16adenovirus types, except types 8, 9, and 10, en-hances its hemagglutination. This supports thetheory (21) that the penton of the adenovirusesalso contains a group-specific antigen component,localized supposedly in the vertex capsomer.
Besides the lack of HE effect, the HI propertycould be demonstrated with antisera to types 8, 9,and 10, the titer of the latter two being consider-ably lower (1:40, and 1:20, respectively). Thisconfirms our previous observations on the cross-reactions among these three types (12, 13, 18).Results obtained in neutralization and HI tests(Table 4) with type 8 antihexon serum indicatethe possibility that it is not the hexon componentwhich is responsible for this cross-reaction butthe vertex projections. Various data are availablewith other adenovirus types on the type speci-ficity of hexon and the neutralization or virionHI effect of sera produced against purified hexon(3, 5, 11, 19, 23, 26, 35), whereas other authorsreport different results (28). Our observationsseem to support the previous findings.The trypsin sensitivity of the complete soluble
HA corresponds to that found with type 9 (24),which supports the reasonable suggestion thatthis component of type 8 is also a polymer ofpentons. The results obtained with purified com-plete HA in zonal centrifugation (Fig. 5) indicatethat, although polymers consisting of severalpentons, even dodecons, may be present, slowlysedimenting small units, occasionally dimers, mayassemble to build this component as well. Thelocalization of the slowly sedimenting incompleteHA is related to the rapid decomposition of thecomplete HA. Had they been present in the start-ing material in monomeric form, they would havebeen localized in fractions 10 to 13, taking theresults in Fig. 2 and 3 into consideration. An
eight-times-longer run was required for the iden-tical components of type 9 to move this far fromthe surface during gradient centrifugation (24).It may be presumed, on the basis of the findingspresented in Fig. 5, that the rapidly sedimentingpolymers are more unstable, than the slow ones.This is indicated by the fact that the fractionslocalized in the upper part of the tube, comprisingonly slowly sedimenting complete HA, whichconsists supposedly of dimers or trimers, containonly a small quantity, if any, of incomplete HAcovered up by the complete one.The problem of whether the soluble complete
HA of type 8 is a dodecon like those of the typesstudied of subgroup 11 (4, 20, 24) or a mixture ofpolymers consisting of different numbers of pen-tons, is questionable, but it seems worth indicatingthat slowly sedimenting complete HA (penton orfiber dimers) were found with other adenovirustypes as well; these, however, eluted in anion-exchange chromatography already separated fromthe dodecon (20, 25). Such separately elutingcomplete HA were not observed in our experi-ments; thus it might be supposed that the com-plete HA of type 8 is also a dodecon, which splitsin a very short time into smaller polymers ofdifferent sizes because of its instability.The soluble complete HA could be absorbed
with human erythrocytes from the solution andsubsequently eluted from them by RDE, similarlyto certain other adenovirus types (9, 20, 24). Thismethod can be used to purify the complete HA,as described with type 9 (24), and to concentrateit to a certain degree. It was also successfullyapplied for the purification and concentration ofthe incomplete HA, by incubating them before theabsorption with corresponding heterologous ade-novirus antisera. The technique provides the op-portunity not only for rapid isolation of theincomplete HA but also for obtaining favorablepurification results with the hexon antigen.The mechanism of the phenomenon which en-
abled us to develop this method of purification isprobably a linkage between the subgroup- orgroup-specific determinants (4 or /B) of two ormore incomplete HA (23, 27), accomplished bythe heterologous antibody, resulting in polymers.The aggregates thus acquiring two or more type-specific antigen determinants ('y), which are sup-posedly the knobs of the fibers, will be capable ofadsorbing onto the surface of erythrocytes, where-from they can be eluted by destroying the re-ceptors with RDE.No explanation was found for the late elution
of a part of the pentons (Fig. 4); it is possible thatsome kind of aggregation causes their elution withthe same NaCl concentration as the complete HAor their adsorption to some other virus (or tissue)
412 J. VIROLS.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 21
Dec
embe
r 20
21 b
y 5.
154.
57.1
92.

COMPONENTS OF ADENOVIRUS TYPE 8
component. It is known that the fiber is capable ofadsorbing to cell or virus DNA (14), and the fiberis actually one of the constituents of the penton.As no information is available at this time on thecomponent supposedly present inside the dodecon(21), a possible role can not be excluded.Although our experiments were not designed as
a detailed examination of the infectivity of adeno-virus type 8, the marked instability of the solublecomponents, some of them playing a part in theadsorption of virus to the cell (21), may perhapsaccount for the strikingly low infectivity of thistype.
ACKNOWLEDGMENTS
We are indebted to I. Be1adi for supplying virus material andfor the kind help of I. Cserba. The skilled technical assistance ofZ. Bakonyi and M. Lugasi is greatly appreciated.
LIRATURE CITED
1. Bauer, W., and R. Wigand. 1967. Vergleichende Unter-suchungen an Hamagglutininen von Adenovirusstiimmender Gruppen I und II. Arch. Gesamte Virusforsch. 21:11-24.
2. BeIldi, I., A. Kahan, E. Kukan, I. Mucsi, and I. Papai. 1963.Antigenic relationship of adenovirus type 8 to the epidemicof keratoconjunctivitis in Szeged. Acta Microbiol. Acad.Sci. Hung. 10:59-63.
3. Dohner, L. 1968. Untersuchungen zum Nachweis von sero-
lcgischen Unterschieden zwischen den Gruppenantigenenaus verschiedenen Adenovirus-typen. Z. Immunitaets-forsch. Exp. Ther. 136:404-414.
4. Gelderblom, H., H. Bauer, H. Frank, and R. Wigand. 1967.The structure of group lI adenoviruses. J. Gen. Virol. 1:553-560.
5. Gelderblom, H., W. Meiser, A. Lengyel, and R. Wigand.1968. The serological relationship of the soluble antigens ofadenovirus type 19. J. Gen. Virol. 2:331-340.
6. Gelderblom, H., R. Wigand, and H. Bauer. 1965. DEAE-chromatography of adenoviruses of Rosen's group II.
Nature (London) 205:625-626.7. Haruna, I., H. Yaoi, R. Kono, and I. Watanabe. 1961. Sepa-
ration of adenovirus by chromatography on DEAEPcellulose. Virology 13:264-267.
8. lawetz, E., S. J. Kimura, L. Hanna, V. R. Coleman, P.Thygeson, and A. Nicholas. 1955. Studies on the etiologyof epidemic keratoconjunctivitis. Amer. J. Ophthalmol.40:200-209.
9. Kasel, J. A., W. P. Rowe, and J. L. Nemes. 1960. Modifica-tion of erythrocyte receptors by a factor in adenovirussuspensions. Virology 10:388-391.
10. Kjellen, L. 1961. A study of adenovirus-host cell system bythe plaque technique. Virology 14:234-239.
11. Kjellen, L., and H. G. Pereira. 1968. Role of adenovirus anti-gens in the induction of virus neutralizing antibody. JGen. Virol. 2:177-185.
12. Lengyel, A., P. Dan, I. Cserba, and 1. Nasz. 1965. Die wech-selseitige heterotypische Hamagglutinations-Hemmungs-reaktion mit Adenoviren Typ 8, 9 und 10. Z. Immuni-taetsforsch. Exp. Ther. 129:34-41.
13. Lengyel, A., and I. Nasz. 1964. Adenovirus haemagglutina-tion-inhibiting antibodies in human sera. Acta Microbiol.Acad. Sci. Hung. 11:105-113.
14. Levine, A. J., and H. S. Ginsberg. 1968. Role of adenovirus
structural proteins in the cessation of host-cell biosyntheticfunctions. J. Virol. 2:430-439.
15. Lowry, 0. H., N. J. Rosebrough, A. L. Farr, and R. J.
Randall. 1951. Protein measurement with the Folin phenolreagent. J. Biol. Chem. 193:265-275.
16. Nasz, I. 1963. Some biological characteristics of adenovirustype 8 strains. Acta Microbiol. Acad. Sci. Hung. 10:355-363.
17. Nasz, I., G. Kulcsar, P. Dan., A. Lengyel, and 1. Cserba. 1963.Studies on the etiology of epidemic keratoconjunctivitisobserved in Budapest during winter 1961-1962. ActaMicrobiol. Acad. Sci. Hung. 10:35-40.
18. Nisz, I., A. Lengyel, P. Dan, and G. Kulcsir. 1963. Hetero-typic haemagglutination-inhibition in the adenovirus group.
Acta Microbiol. Acad. Sci. Hung. 10:379-385.19. Nasz, I., and H. G. Pereira. 1965. Studies on the antigen-
fractions of an oncogenic adenovirus type 7 strain (inHungarian). Proc. Med. Dep. Hung. Acad. Sci. 15:327-338.
20. Norrby, E. 1968. Comparative studies on the solusble com-
ponents of adenovirus types 9 and 15 and the intermediatestrain 9-15. J. Virol. 2:1200-1210.
21. Norrby, E. 1968. Biological significance of strtuctural adeno-
virus components. Curr. Top. Microbiol. Immuncl. 43:1-43.22. Norrby, E. 1968. Identification of soluble components of
adenovirus type 11. J. Gen. Virol. 2:123-133.23. Norrby, E. 1969. The structural and functional diversity of
adenovirus capsid compornents. J. Gen. Virol. 5:221- 236.24. Norrby, E., B. Nyberg, P. Skaaret, and A. Ler.gyel. 1967.
Separation and characterization of soluble ader-ovirus tvpe9 components. J. Virol. 1:1 101- 10.
25. Norrby, E., and P. Skaaret. 1968. Comparison betNieen solulblecomponents of adenovirus types 3 and 16 and of the i,iter-mediate strain 3-16 (the San Carlos agent). VirdAogy36:201-211.
26. Pereira, H. G. 1956. Typing of adenoidal-pharyngeal-con-junctival (APC) viruses by complement fixation. J. Pathol.Bacteriol. 72:105-109.
27. Pereira, H. G., and M. V. T. De Figueiredo. 1962. Mechanismof hemagglutination by adenovirus types 1, 2, 4, 5 and 6.Virology 18:1-8.
28. Pettersson, U., L. Philipson, and S. Hoglund. 1967. Struc-tural proteins of adenoviruses. I. Purification and charac-terization of the adenovirus type 2 hexon antigen. Virology33:575-590.
29. Pifia, M., and M. Green. 1965. Biochemical studies on adeno-virus multiplication. IX. Chemical and base compositionanalysis of 28 human adenoviruses. Proc. Nat. Acad. Sci.U.S.A. 54:547-571.
30. Pusztai, R., I. Blaidi, M. Bakay, and I. Mucsi. 1969. Effectof ultraviolet irradiation and heating on the interferon-inducing capacity of human adenoviruses. J. Gen. Virol.4:169-176.
31. Rosen, L. 1960. A hemagglutination-inhibition technique fortyping adenoviruses. Amer. J. Hyg. 71:120-128.
32. Takstsy, G. 1955. The use of spiral loops in serological andvirological micro-methods. Acta Microbiol. Acad. Sci.Hung. 3:191-202.
33. Trentin, J. J., G. L. Van Hoosier, Jr., and L. Samper. 1968.The oncogenicity of human adenoviruses in hamsters.Proc. Soc. Exp. Biol. Med. 127:683-689.
34. Valentine, R. C., and H. G. Pereira. 1965. Antigens andstructure of the adenovirus. J. Mol. Biol. 13:13-20.
35. Wilcox, W. C., and H. S. Ginsberg. 1963. Production ofspecific neutralizing antibody with soluble antigens of typeS adenovirus. Proc. Soc. Exp. Biol. Med. 114:37-42.
VOL. 6, 1970 413
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 21
Dec
embe
r 20
21 b
y 5.
154.
57.1
92.





























