Embed Size (px)
Citation preview

Neuroscience Letters, 20 (1980) 205-210 205 © Elsevier/North-Holland Scientific Publishers Ltd.
SOLUBILIZATION OF 20S ACETYLCHOLINESTERASE FROM THE CHICK CENTRAL NERVOUS SYSTEM
ANA BARAT, ELENA ESCUDERO, JAVIER GOMEZ-BARRIOCANAL and GALO RAMIREZ*
Centro de Biologia Molecular, CSIC-UAM, Canto Blanco, Madrid-34 (Spain)
(Received July 2nd, 1980; Revised version received August 6th, 1980; Accepted August 6th, 1980)
The ionic detergent sodium cholate, in the presence of 1 M NaCI, solubilizes a 20S acetylcholinesterase from chick retina and other brain tissues previously extracted with a buffered solution containing 1% Triton X-100 and 1 M NaCI. This 20S acetylcholinesterase appears to be a tailed form of the enzyme which, upon collagenase digestion, is converted to a 22S (mainly) form. This finding suggests that the vertebrate central nervous system does contain asymmetric, collagen-tailed forms of acetylcholinesterase, as is the case in skeletal muscle and cholinergic ganglia.
Acetylcholinesterase (acetylcholine hydrolase, EC 3.1.1.7) (ACHE), in its simplest monomeric form, has been characterized, in a number of biological systems, as a polypeptide chain with a molecular weight in the vicinity of 80,000 daltons [6]. Work in electrogenic tissue from electric fish has revealed, however, the existence of complex quaternary structural forms of the enzyme incorporating up to 12 catalytic subunits, arranged in discrete tetramers, and including at times (elongated or asymmetric membrane-bound forms) a second type of polypeptide chain, structurally related to collagen, integrated into a tail-like structure presumably involved in the attachment of the enzyme to the cell membrane (or extracellular basement membrane) matrix [1, 4, 7]. Although this general model has been confirmed so far in most cholinergic systems, it has not yet been possible to show conclusively the existence of tailed AChE forms in the vertebrate central nervous system [1, 3, 5, 8, 9]. Recently, we have shown [10] that collagenase, an enzyme known to cleave the tail protein in the asymmetric forms of AChE [1, 7, 8], can release two fast-sedimenting AChE species from chick retina and optic tectum (i.e. central nervous system) similar to those obtainable by collagenase digestion of the AChE tailed forms identified in chick peripheral nervous tissue and skeletal muscle [1, 2, 8, 9]. Having thus obtained indirect evidence as to the existence of tailed AChE species in chick retina and optic tectum, we have set out to solubilize them by means of non-proteolytic agents in order to confirm their existence and study their
* To whom correspondence should be addressed.

206
properties. We report here the solubilization of a 20S form of AChE from chick retina and other chick brain regions, by means of the ionic detergent sodium cholate, using a two-step extraction procedure.
The experiments have been carried out in 10-day-old chicks (10 _+ 1 days after hatching), hatched and raised as previously described [10, 11]. Retinas and other brain tissues were first homogenized in 10 vol. of 10 mM Tris. HC1 buffer, pH 7, containing 1% Triton X-100, 1 M NaCI and 50 mM MgCI2 (Tr i s -Tr i ton-sa l t buffer or homogenization buffer), at 4°C, to extract most of the AChE as l lS and 6S molecular forms [10, 11]. This homogenate was centrifuged at 110,000 x g, for 30 min, at 2°C. The supernatant was set aside and the pellet rehomogenized as before, and centrifuged again. This second pellet, representing the material insoluble in the initial homogenization buffer, was resuspended in a solution including 10 mM Tris • HCI, pH 7, 1% sodium cholate, 1 M NaCI and 50 mM MgCI2, and extracted for 30 rain. This cholate extract was freed of particulate material by centrifugation at 110,000 x g, as indicated above, and further analyzed by sedimentation in a linear sucrose gradient (5 -20%) prepared in the initial homogenization buffer (i.e. without cholate) [10, 11].
All the procedures used in sedimentation analysis, collagenase treatment and enzymatic assays, both AChE and sedimentation marker enzymes, have been described in detail in previous publications [10, 11]. Some additional experimental details are given in the legends to figures and tables.
Fig. 1, upper panel, shows the sedimentation pattern of the chick retina AChE quaternary structural forms in the first supernatant (material extracted by the Tr i s -Tr i ton - sa l t buffer); below, we can see the sedimentation behavior of the AChE molecular forms released by 1% sodium cholate, in 10 mM Tris • HCI buffer, pH 7, containing 1 M NaCI and 50 mM MgCI2, from the pellet remaining after the extraction and further washing of the retinal tissue with the initial homogenization buffer. As we see, a new 20S (19.9 _+ 0.2, in 9 different runs) form of ACHE, accounting for almost 50% of the residual enzyme activity (see Table I), is released by sodium cholate in this second extraction step (sodium cholate does not solubilize the 20S AChE when used in the initial homogenization step). This AChE form has therefore a sedimentation coefficient identical to that of the other fast-sedimenting molecular species of the enzyme previously characterized in chick ganglia, peripheral nerve and skeletal muscle [1, 2, 8, 9], believed to be the chick equivalent of the A12 tailed AChE [1] identified first in the electric fish and composed of 3 tetrameric clusters of catalytic subunits and a collagenase-sensitive 3-stranded tail [4, 7].
To confirm the tailed nature of the retinal 20S AChE we subjected it to collagenase digestion [10, 11]. The results are illustrated in Fig. 2, which shows the changes induced by collagenase in the structure and sedimentation behavior of the cholate-extracted 20S ACHE: the 20S peak disappears as such and most of the activity is now distributed in two new bands with sedimentation coefficients of
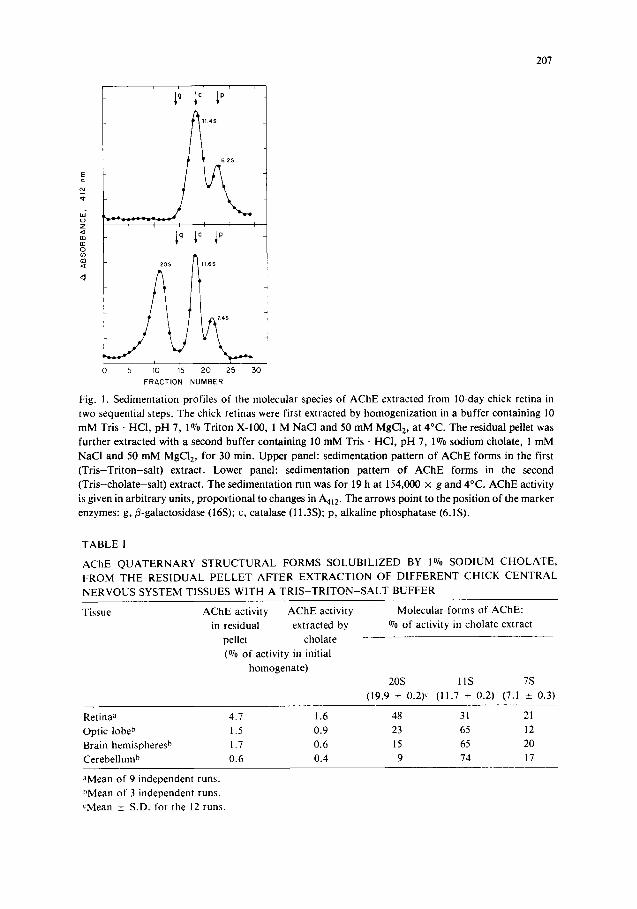
207
i r i T I c
11.45
c
w"
g 205 116$
5 10 15 2 0 2 5 5 0
F R A C T I O N N U M B E R
Fig. 1. Sedimentation profiles of the molecular species of AChE extracted from 10-day chick retina in two sequential steps. The chick retinas were firs[ extracted by homogenization in a buffer containing 10 mM Tris • HCI, pH 7, 1% Triton X-100, 1 M NaCl and 50 mM MgCI2, at 4°C. The residual pellet was further extracted with a second buffer containing 10 mM Tr i s . HC1, pH 7, 1070 sodium cholate, 1 mM
NaCI and 50 m M MgC12, for 30 min. Upper panel: sedimentation pattern of A C h E forms in the first
(Tr i s -Tr i ton-sa l t ) extract. Lower panel: sedimentation pattern o f AChE forms in the second (Tr is -chola te -sa l t ) extract. The sedimentation run was for 19 h at 154,000 x g and 4°C. AChE activity
is given in arbitrary units, proportional to changes in A412. The arrows point to the position o f the marker enzymes: g,/3-galactosidase (16S); c, catalase (11.3S); p, alkaline phosphatase (6.1S).
TABLE 1
AChE Q U A T E R N A R Y S T R U C T U R A L FORMS SOLUBILIZED BY 1°70 SODIUM C H O L A T E , FROM THE RESIDUAL PELLET AFTER EXTRACTION OF DIFFERENT CHICK CENTRAL
NERVOUS SYSTEM TISSUES WITH A T R I S - T R I T O N - S A L T BUFFER
Tissue AChE activity AChE activity
in residual extracted by pellet cholate (°70 of activity in initial
homogenate)
Molecular forms of ACHE:
% of activity in cholate extract
20S 11S 7S (19.9 _+ 0.2) c (11.7 _+ 0.2) (7.1 _+ 0.3)
Retina a 4.7 1.6 48 31 21 Optic lobe b 1.5 0.9 23 65 12 Brain hemispheres h 1.7 0.6 15 65 20 Cerebellum h 0.6 0.4 9 74 17
aMean of 9 independent runs. bMean of 3 independent runs.
cMean _+ S.D. for the 12 runs.

208
E c
z
o oo a3
<1
. . . . +~ +c +p
I ~ control
I i H 8S
i 21 as ~g ,c ,p
V co,+o° .....
O 5 ]O ]5 20 25 30 FRACTION NUMBER
Fig. 2+ Effects of collagenase on 20S retinal ACHE. The 20S AChE extracted by 1% sodium cholate (Fig. 1, lower panel) was dialyzed against homogenization buffer (without cholate), supplemented with CaCI2 up to 5 mM, and split into two equal portions: one was digested with 100 units/ml of protease-free cotlagenase (Sigma type VI, 840 units/mg) at 37°C for 30 rain [10], and the other one (control) simply incubated as such under the same conditions. Finally both samples were analyzed by sucrose gradient centrifugation. Total activities recovered in both gradients were practically identical. Upper panel: control sample. Lower panel: collagenase-digested sample.
21.8S and 16.9S. These values agree with the results o f o ther exper iments on
col lagenase t r ea tment o f chick tai led A C h E s [1, 8] and are to ta l ly consis tent with
our own previous f indings when digest ing direct ly with col lagenase the pellet
insoluble in the T r i s - T r i t o n - s a l t h o m o g e n i z a t i o n buf fe r [10]. Then , the two new
A C h E peaks co r r e spond most l ikely to the par t ia l ly tailless At2 and A8 A C h E s ,
respectively [1]. The small amoun t s o f l IS and 7S forms appea r ing both in the
cont ro l and in the col lagenase-d iges ted mater ia l poin t to the ac t iva t ion or
s imul taneous release o f some pro tease by the chola te ex t rac t ion step since the 21.5S
and 16.5S forms released direct ly by col lagenase f rom the T r i s - T r i t o n - s a l t -
insoluble res idue were s table to bo th 37°C incuba t ion and fur ther col lagenase
t r ea tment [10]. Besides sod ium cholate , we have t r ied a number o f detergents , chao t rop ic agents ,
enzymes and membrane -d i spe r s ing agents (e.g. sod ium deoxycho la te , po ta s s ium
b romide , perchlor ic acid salts, phospho l ipases , chondro i t inase , l i th ium
d i iodosa l icy la te , d igi tonin) . None o f these agents was effect ive in solubi l iz ing active
ta i led forms o f ACHE.

209
We have not yet been able to solubilize directly, f rom the chick central nervous system, the A8 form of AChE which has a sedimentation coefficient of about 15S in other chick cholinergic tissues [1]. However, the collagenase digestion of the retinal 20S AChE (A12) also yields a small proport ion of the 16.5-17S derivative of the A8 form (Fig. 2), presumably through the loss of a tetramer from the 21.5-22S digested form.
The fact that sodium cholate is not effective in solubilizing 20S AChE in the initial homogenization step, under the present experimental conditions, is somewhat surprising. It could be possible, however, that the initial extraction in the T r i s -Tr i t on - sa l t solution alters the membrane structure in such a way as to permit the observed action of sodium cholate in the second extraction step. This selective action of sodium cholate seems, nevertheless, genuine and specific. It is not just the effect of repetitive homogenizations and extractions since repeated homogenizat ion-centr i fugat ion cycles of the tissues in T r i s -Tr i t on - sa l t buffer (up to 3 times, and including incubation of the homogenate at 37°C [10]) does not result in the release of 20S ACHE. On the other hand, the addition of 1 °7o sodium cholate to the Triton-extracted l lS and 6S AChE forms does not either induce the artifactual format ion of any faster-sedimenting molecular species.
Finally, sodium cholate extracts equally the 20S form of AChE from different chick brain regions, under the same conditions, although the relative proport ions are different in each case (Table I). Thus, the occurrence of the Al2 tailed form of AChE is probably common to all vertebrate cholinergic structures, including the central nervous system. The apparently small proport ion of tailed AChE molecules in the chick central nervous system (less than 1°70 of the homogenate activity), in comparison with other chick cholinergic tissues [1, 2, 8, 9], may seem at first sight striking. Whether it reflects a more specific and selective localization of the 20S AChE in the chick central nervous system, or is simply due to non-optimal solubilization conditions, is being currently investigated in our laboratory.
This work was supported by grants from the Comisi6n Asesora de Investigaci6n Cientifica y T6cnica and Comisi6n Administradora del Descuento Complementar io.
1 Bon, S., Vigny, M. and Massouli~, J., Asymmetric and globular forms of acetylcholinesterase in mammals and birds, Proc. nat. Acad. Sci. (Wash.), 76 (1979) 2546-2550.
2 Couraud, J.Y,, Koenig, H. and DiGiamberardino, L., Formes mol~culaires de l'ac6tylcholinesterase dans le ganglion ciliaire du poulet; +volution apr6s d6nervation, axotomie et double section, C.R. Acad. Sci. (Paris), 288 (1979) 1199-1202.
3 Fern~mdez, H.L., Duell, M.J. and Festoff, B.W., Cellular distribution of 16S acetylcholinesterase, J. Neurochem., 32 (1979) 581-585.
4 Rieger, F., Bon, S., Massouli~, J., Cartaud, J., Picard, B. and Benda, P., Torpedo marmorata acetylcholinesterase; a comparison with the Electrophorus electricus enzyme, Europ. J. Biochem., 68 (1976) 513-521.

210
5 Rieger, F. and Vigny, M., Solubilization and physicochemical characterization of rat brain acetylcholinesterase: development and maturat ion of its molecular forms, J. Neurochem., 27 (1976)
121-129.
6 Rosenberry, T.L. , Acetylcholinesterase, Advanc. Enzymol. , 43 (1975) 103 218.
7 Rosenberry, T.L. and Richardson, J .M., Structure of 18S and 14S acetylcholinesterase. Identification of collagen-like subtmits that are linked by disulfide bonds to catalytic subunits,
Biochemistry, 16 (1977) 3550-3558.
8 Rottmdo, R.L. and Fambrough, D.M., Molecular forms of chicken embryo acetylcholinesterase in vitro and in vivo. Isolation and characterization, .1. biol. Chem., 254 (1979) 4790 4799.
9 Vigny, M., DiGiamberardino, k., Couraud, J.Y., Rieger, F. and Koenig, .1., Molecular forms of
chicken acetylcholinesterase: effect of denervation, FEBS kett. , 69 (1976) 277 280. 10 Villafruela, M.J . , Barat, A., Villa, S. and Ramirez, G., Molecular forms of acetylcholinesterase in
the chick visual system. Collagenasc-released 21.5S and 16.5S species, FEBS Lett., 110 (1980) 91-95.
11 Villafruela, M.J. , Barat, A., Manrique, E., Villa, S. and Ramirez, G., Molecular forms of acetylcholinesterse in the developing chick visual system, Develop. Neurosci., in press.





























