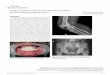
Skeletal and Dental Development Preserve Evidence of
Uploadothers
View
Download
Embed Size (px)
344 x 292
429 x 357
514 x 422
599 x 487
Citation preview
Skeletal and Dental Development Preserve Evidence of Energetic
Stress in the Moose of Isle RoyaleORIGINAL RESEARCH published: 29
September 2020 doi: 10.3389/fevo.2020.00272
LOAD MORE