Embed Size (px)
Citation preview

DRUG METABOLISM REVIEWS, 30(2), 421-434 (1998)
SEX STEROID HORMONES AND G EN ETI C S USC E PTI B I LlTY TO BREAST AND PROSTATE CANCER HEATHER SPENCER FEIGELSON,' RONALD K. ROSS,' MIMI C. YU,' GERHARD A. COETZEE,2 JUERGEN K. V. REICHARDT,3 and BRIAN E. HENDERSON'#* 'Department of Preventive Medicine 2Department of Urology U. S. C. /Norris Comprehensive Cancer Center 1441 Eastlake Avenue Los Angeles, California 90033-0800 9epartment of Biochemistry and Molecular Biology Institute for Genetic Medicine University of Southern California School of Medicine 201 1 Zonal Avenue Los Angeles, California 90033
I. INTRODUCTION ........................................................ 422
11. BREAST CANCER MODEL .......................................... 423 The Cytochrome P450c17a Gene (CYP17) .................... 424
The Estrogen Receptor Gene (ER) ............................... 426
A. B. The 17P-Hydroxysteroid Dehydrogenase 1 Gene
(HSD17Bl) ........................................................... 426 C.
*To whom correspondence should be addressed.
42 1
Copyright 0 1998 by Marcel Dekker, Inc.
Dru
g M
etab
olis
m R
evie
ws
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Mic
higa
n U
nive
rsity
on
10/3
1/14
For
pers
onal
use
onl
y.

422 FEIGELSON ET AL.
111. PROSTATE CANCER MODEL ...................................... 426 A. The Steroid Sa-Reductase Type I1 Gene (SRD5A2) ......... 427 B. The Androgen Receptor Gene (AR) ............................. 428 C. The Cytochrome P45Oc17a Gene (CYP17) .................... 430
IV. CONCLUSION ........................................................... 430
Acknowledgments ......................................................... 430
References. ................................................................. 430
I. INTRODUCTION
Second only to lung cancer, prostate and breast cancers are the leading causes of cancer death for men and women in the United States, respectively [I]. Whereas some cancers, such as lung and stomach cancers, have shown a decline in incidence from changing environmental exposures, hormone- related cancers, such as breast and prostate cancers, are becoming more prevalent. The development of this latter group of cancers appears to be closely related to endogenous exposure to circulating steroid hormones: estrogen, and perhaps progesterone, in the case of breast cancer, and tes- tosterone and its metabolites in the case of prostate cancer [2]. Although primary and secondary prevention trials using antihormonal agents are un- derway for both of these cancers, their long-term control requires a clearer understanding of the biosynthesis and metabolism of the relevant hormones in vivo [3].
Using the paradigms that have been developed from studies of bladder cancer [4] and using our existing knowledge about the hormone-dependent nature of breast and prostate cancer, we have proposed models for individual susceptibility to these two cancers. In developing these models, we have assumed that within and between ethnic groups there exist genetic differences that affect steroid hormone biosynthesis. Markers (i.e., genetic polymor- phisms) of these differences are likely to provide a more precise measure of risk than circulating levels of steroid hormones. The latter measurements are subject to variation by many factors, including time of day of specimen collection, age, menopausal status, disease status, and laboratory impreci- sion. Furthermore, the intracellular concentrations of the active hormones are not necessarily always affected by serum steroid levels. The multigenic models that we have developed for the hormone-related cancers assume that there are functionally important polymorphisms in genes that encode enzymes involved in steroid hormone biosynthesis and metabolism that lead to differ-
Dru
g M
etab
olis
m R
evie
ws
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Mic
higa
n U
nive
rsity
on
10/3
1/14
For
pers
onal
use
onl
y.

GENETIC SUSCEPTIBILITY TO CANCER 423
ences in individual susceptibility to breast and prostate cancers. We have selected candidate genes based on two criteria: (1) the gene has known polymorphisms that may be functionally relevant and (2) the product of the gene acts in a pathway that is physiologically relevant based on our under- standing the etiology of the cancer in question.
There is an important distinction between the genes proposed in these models and susceptibility genes such as BRCAl and BRCA2. Genes like BRCAl and BRCA2 that are necessary and sufficient to cause cancer are very rare and have low population attributable risks. Polymorphisms in “metabolic” genes, such as those described in the following models, are relatively common. So, whereas the at-risk genotypes may each cany a small risk relative to those associated with BRCAl and BRCA2, they likely con- vey much larger population-attributable risks.
In the following sections, we describe our proposed models of breast and prostate carcinogenesis. We also show recent data that are supportive of these models.
11. BREAST CANCER MODEL
Attempts to understand and quantify the role of estrogens in breast can- cer have been limited to some extent by our inability to accurately and re- producibly measure steroid hormones in circulating blood. Despite the dif- ficulty of conducting such studies, there is evidence from our group [5-71 and others [8] of significant elevations in circulating estradiol in breast cancer patients compared to controls in populations at both high or low risk of breast cancer. In addition, there are reproducible differences in plasma and urine estrogen levels between healthy high-risk U.S. and low-risk Asian women [6.7]. In one such study in which cycle length was controlled, estradiol levels were 20% higher in Los Angeles compared to Shanghai premenopausal women [6]; in a companion study of postmenopausal women, estradiol levels were 36% higher in Los Angeles than in their matched Japanese counter- parts [7]. Reasons for these differences remain poorly understood, but part of the explanation may be that there are genetic differences that affect me- tabolism or intracellular binding and transport of estradiol.
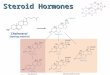
Based on the schematic presentation of estrogen metabolism in the ova- ries and breast epithelium shown in Fig. l , we describe (below) the func- tion of each of three candidate genes and their possible roles in breast can- cer etiology. The genes of interest are the cytochrome P450c17a gene (CYP17), the 17P-hydroxysteroid dehydrogenase 1 gene (HSD17Bl). and the estrogen receptor gene (ER).
Dru
g M
etab
olis
m R
evie
ws
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Mic
higa
n U
nive
rsity
on
10/3
1/14
For
pers
onal
use
onl
y.

424 FEIGELSON ET AL.
Choleaerol
1 plS(kl'l(CYPI7) Andmetandone
E s m r (El)
17HSD1 (HBDI7Bl)
Estradiol (E2)
17HSDl
E2
FIG. 1. Schematic presentation of 17P-hydroxysteroid dehydrogenase 1 (HSD17B1), cytochrome P450c17a (CYP17), and estrogen receptor (ER) genes in estrogen metabolism in the ovaries and breast epithelium.
A. The Cytochrome P450c17a Gene (CYP17)
CYP17 codes for the cytochrome P450c17a enzyme that mediates both steroid 17a-hydroxylase and 17-20-lyase -activities and functions at key branch points in human steroidogenesis [9]. 17a-Hydroxylase activity con- verts steroids to precursors of the glucocorticoid cortisol, and 17-20-lyase activity yields precursors to estradiol and testosterone.
The 5' untranslated region of CYP17 contains a single base-pair (bp) poly- morphism that creates an Spl-type (CCACC box) promoter site 34 bp up- stream from the initiation of translation, but downstream from the transcrip- tion start site [lo]. This base-pair change also creates a recognition site for the MspAI restriction enzyme. MspAI digestion of a PCR fragment has been used to arbitrarily designate two alleles, A1 (the published sequence) and A2 [lo]. Because this base-pair change creates a CCACC box and it is thought that the number of 5' promoter elements correlates with promoter activity [ 1 I], the A2 allele may result in an increased rate of transcription.
We have reported preliminary findings from a case-control study of breast cancer and this CYP17 polymorphism in which we found an increased risk of advanced breast cancer associated with the A2 allele [odds ratio (OR) = 2.5; 95% confidence interval (CI) = 1.07-5.941 1121. Further, we showed that the A2 allele is associated with an earlier age at menarche among con- trol subjects and that there was an interaction effect between CYP17 geno- type and age at menarche on the risk of breast cancer. As shown in Table 1, the protective effect of later age at menarche for breast cancer was more pronounced in women with the Al/Al genotype.
Dru
g M
etab
olis
m R
evie
ws
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Mic
higa
n U
nive
rsity
on
10/3
1/14
For
pers
onal
use
onl
y.

TABLE
1
Odd
s R
atio
s an
d 95
% C
onfid
ence
Intervals fo
r th
e Risk o
f B
reas
t C
ance
r A
ccor
ding
to
Age
at
Men
arch
e fo
r AU
Subjects a
nd S
ubgr
oups
Str
atifi
ed b
y C
YP1
7 Genotype
AlI
Al
AlI
A2
+ M
IA2
A
ll s
ubje
cts
geno
type
ge
noty
pe
Men
arch
e O
R"
95%
CI
OR"
95%
CI
OR"
95
% C
I
<13
yr
1 .o
-
1 .o
-
1 .o
-
2 13
yr
0.69
0.
47-1
.02
0.47
0.
22-0
.98
0.80
0.
5 1-
1.27
"Adj
uste
d fo
r ag
e, e
thni
city
, and
age
at
first
full-
term
pre
gnan
cy.
Sour
ce:
Ada
pted
fro
m R
ef.
12.
Dru
g M
etab
olis
m R
evie
ws
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Mic
higa
n U
nive
rsity
on
10/3
1/14
For
pers
onal
use
onl
y.

426 FEIGELSON ET AL.
These results need to be confirmed in a larger study, and further labo- ratory investigation is required to determine whether the A 1 /A2 polymor- phic site in CYP17 is, in fact, the causative mutation or is simply a marker for another mutation that affects enzymatic activity.
B. The 17B-Hydroxysteroid Dehydrogenase 1 Gene (HSD17Bl)
The HSD17B1 gene codes for the enzyme 17fLhydroxysteroid dehydro- genase type 1 (17HSD1) which catalyzes the final step of estradiol biosyn- thesis, namely the interconversion of estrone (El) into the more biologically active estrogen, estradiol (E2). 17HSDl acts in the theca cells of the ovary and is expressed in both normal and malignant breast epithelium [ 131.
Several polymorphisms have been identified in the HSD 17B 1 gene [14,15]. Normand et al. [14] described 11 polymorphisms, 2 of which are located in the coding region and result in amino acid changes. Neither poly- morphism appears to substantially affect the catalytic or the immunological properties of the enzyme [16], but one (a serine to glycine change at posi- tion 312) has been shown to be associated with a marginally statistically significant increased risk for breast cancer [ 151. We are currently evaluat- ing this polymorphism in a case-control study of breast cancer.
C. The Estrogen Receptor Gene (ER)
Several polymorphisms in the ER gene have been reported [17-201. The most promising evidence for an association between ER gene polymorphisms and breast cancer risk was reported by Andersen et al. [19], who examined three Restriction Fragment-Length Polymorphisms (RFLPs) and found that alleles having an XbaI restriction site were significantly more frequent in breast cancer patients than controls (OR = 2.0). This association was lim- ited to postmenopausal patients. Del Senno et al. [20] reported a highly poly- morphic (TA), repeat upstream from the region of the ER gene originally described as exon 1 in the transactivation domain. This is a potentially in- teresting region, as the ER has recently been shown to have at least three different promoters that exhibit tissue-specific expression [2 1-24]. Further characterization of this region could provide important clues for understand- ing the role of the ER gene in breast cancer. We have initiated studies of the XbaI polymorphism and the (TA), repeat in breast cancer.
111. PROSTATE CANCER MODEL
Increased cell division is the hallmark of the response of prostate epithelial cells to androgens. Specifically, cell division in the prostate is controlled by
Dru
g M
etab
olis
m R
evie
ws
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Mic
higa
n U
nive
rsity
on
10/3
1/14
For
pers
onal
use
onl
y.

GENETIC SUSCEPTIBILITY TO CANCER 427
4 4 P45Oc17 (CWI7)
Pregnenolone
Androstenedine
Testosterone (T) 4
Testicular Biosynthesis Rostate Qithelial Cell I Cholesterol 1 I
/ (k) _.) DNA
Dihydrotestosterone @HT)
1 sRD5A2
- -b Testosterone(T) - FIG. 2. Schematic presentation of steroid Sa-reductase (SRDSA2),
cytochrome P450c17a (CYP17). and androgen receptor (AR) activity in testosterone metabolism in the testicles and prostate epithelium.
testosterone (T), following its intracellular conversion to dihydrotestosterone (DHT), as shown in Fig. 2. This conversion is catalyzed by the enzyme steroid Sa-reductase type 11. DHT and, much less efficiently, T bind to the androgen receptor (AR) in the cytoplasm, and the ARAigand complexes are translocated to the nucleus for DNA binding and transactivation of target genes, including those that control cell division and death.
Ethnicity is an important determinant of prostate cancer risk. African- Americans have higher rates than whites who, in turn, have higher rates than U.S.-born Asians [25]. Differences among ethnic groups with respect to serum testosterone levels [26] and steroid Sa-reductase activities as measured by the DHT metabolite, Sa-androstane-3a, 17P-diol- 17P-glucuronide (AAG) have been suggested to explain, at least in part, the large ethnic variability in the incidence of prostate cancer [27]. The genes of interest to us currently are the steroid Sa-reductase gene type I1 (SRDSA2), the androgen receptor gene (AR), and the cytochrome P450c17a gene (CYP17).
A. The Steroid 5a-Reductase Type I1 Gene (SRD5A2)
There are two distinct steroid Sa-reductase enzymes encoded for by dif- ferent genes; the type I1 enzyme is active primarily in the prostate and in genital skin. Germline mutations of the type I1 gene (SRDSA2) have been identified, most notably in a sizable kindred in the Dominican Republic [28]. Boys with this mutated gene are phenotypically female with a persistent vaginal pouch until puberty, at which time there is some phallic enlargement and development of some secondary sex characteristics, but the prostate remains underdeveloped.
Davis and Russell [29] have described a polymorphism (TA repeat num- ber) in the 3' untranslated region of the SRDSA2 gene. We have confirmed
Dru
g M
etab
olis
m R
evie
ws
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Mic
higa
n U
nive
rsity
on
10/3
1/14
For
pers
onal
use
onl
y.

428 FEIGELSON ET AL.
this polymorphism [30], but find the prevalence of variant alleles using this polymorphic marker to be substantially higher than that reported by Davis and Russell.
We have also identified additional polymorphisms in the gene [31], in- cluding a functional substitution in exon 1 that changes the amino acid at position 89 from valine to leucine (V89L). As shown in Table 2, the leu- cine (L) allele is more common in Asians and is associated with lower lev- els of circulating AAG (a metabolite of DHT). These data may provide an explanation for the low risk of prostate cancer in Asian populations.
B. The Androgen Receptor Gene (AR)
It is likely that the AR also is involved in the racial-ethnic variation in prostate cancer incidence. The AR gene, on the long arm of the X chromo- some, belongs to the superfamily of ligand-dependent transactivation factors and, like others in this family, contains a C-terminal hormone-binding do- main, a central DNA-binding domain, and an N-terminal transcriptional modulatory domain which, for the AR gene, is encoded entirely by exon 1 [32].
Within exon 1 , there are two highly polymorphic microsatellites; one of these is a trinucleotide (CAG) repeat sequence of unknown function. Abnor- mal expansion of this satellite is causally associated with X-linked spinal and bulbar muscular atrophy (Kennedy’s disease) [33]. The normal size varies from 9 to 31 repeats, whereas the mutant range associated with Kennedy’s disease varies from 40 to 62. It was demonstrated that these mutant recep- tors have normal androgen binding, but in transfection assays, they trans- activate an androgen responsive reporter gene subnormally [34]. We and others have noted that African-Americans, U. S. whites, and Asian-Ameri- cans show population differences in the CAG repeat polymorphism of the AR locus, with the allele frequency distribution shifted toward shorter alle- les in the case of African-Americans and longer alleles in the case of Asians relative to whites [35,36]. We have hypothesized that this size polymorphism might correlate with, or cause changes in, AR function (smaller size, greater transactivity) and, therefore, possibly also be associated with prostate can- cer risk [36]. We and others have demonstrated this association in case- control comparisons in Caucasian populations [37-391. However, it is unclear whether this polymorphism is the functionally relevant change as described above, or a marker for another nearby functional polymorphism.
Dru
g M
etab
olis
m R
evie
ws
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Mic
higa
n U
nive
rsity
on
10/3
1/14
For
pers
onal
use
onl
y.

GENETIC SUSCEPTIBILITY TO CANCER 429
Dru
g M
etab
olis
m R
evie
ws
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Mic
higa
n U
nive
rsity
on
10/3
1/14
For
pers
onal
use
onl
y.

430 FEIGELSON ET AL.
C. The Cytochrome P450c17a Gene (CYP17)
As described in the breast cancer model, the cytochrome P450c17a en- zyme yields precursors to estradiol and testosterone. We are evaluating the CYP17 polymorphism to determine if it also plays a role in prostate cancer.
IV. CONCLUSION
Both breast and prostate cancers exhibit substantial unexplained differ- ences in risk across ethnic groups. The role of metabolic genes in carcino- genesis and in explaining racial-ethnic variation in risk is a relatively new area of research with scant information at present. Investigating functionally relevant polymorphisms in these genes should help us understand the under- lying etiology of these two cancers and lead to more refined definitions of “high-risk” individuals. Ultimately, this should enable us to better define who may benefit from specific interventions based on underlying genetic susceptibility and provide important targets for prevention and control.
ACKNOWLEDGMENTS
This work was support by grants R01 CA54281, CA63464, CA68581, and R35 CA53890 from the National Cancer Institute and by grant 2FB-0212 from the University of California Breast Cancer Research Program.
REFERENCES
1. S. L. Parker, T. Tong, S. Bolden, and P. A. Wingo, Cancer statis- tics, 1997, Ca Cancer J . Clin., 47, 5-27 (1997).
2. B. E. Henderson, R. K. Ross, M. C. Pike, and J. T. Casagrande, Endogenous hormones as a major factor in human cancer, Cancer Res., 42, 3232-3239 (1982).
3. B. E. Henderson, R. K. Ross, and M. C. Pike, Hormonal chemo- prevention of cancer in women, Science, 259, 633-638 (1993).
4. P. Vineis, Epidemiological models of carcinogenesis: the example of bladder cancer, Cancer Epidemiol. Biomarkers Prev., I , 149- 153 ( 1992).
Dru
g M
etab
olis
m R
evie
ws
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Mic
higa
n U
nive
rsity
on
10/3
1/14
For
pers
onal
use
onl
y.

GENETIC SUSCEPTIBILITY TO CANCER 43 1
5 .
6.
7.
8.
9.
10.
11.
12.
13.
14.
15.
B. E. Henderson, V. Gerkins, I. Rosario, J. T. Casagrande, and M. C. Pike, Elevated serum levels of estrogen and prolactin in daughters of patients with breast cancer, N. Engl. J. Med.. 293, 790-795 (1975). L. Bernstein, J. M. Yuan, R. K. Ross, M. C. Pike, R. Hanisch, R. Lobo. F. Stanczyk, Y. T. Gao, and B. E. Henderson, Serum hormone levels in premenopausal Chinese women in Shanghai and white women in Los Angeles: Results from two breast cancer case-control studies, Cancer Causes Cont., 1, 5 1-58 (1990). H. Shimizu, R. K. Ross, L. Berstein, M. C. Pike, and B. E. Henderson, Serum oestrogen levels in postmenopausal women: Com- parison of American whites and Japanese in Japan, Br. J. Cancer, 62,
P. England, L. Skinner, K. Cottrell, and R. A. Sellwood, Serum oestradiol-17P in women with benign and malignant breast disease, Br. J. Cancer, 30, 571-576 (1974). S. T. Brentano, J. Picado-Leonard, S. H. Mellon, C. C. D. Moore, and W. L. Miller, Tissue-specific, cyclic adenosine 3',5'-monophos- phate-induced, and phorbol ester-repressed transcription from the hu- man P45Oc17 promoter in mouse cells, Molec. Endocrinol., 4 , 1972- 1979 (1990). A. H. Carey, D. Waterworth, K. Patel, D. White, J. Little, P. Novelli, S . Franks, and R. Willamson, Polycystic ovaries and prema- ture male pattern baldness are associated with one allele of the steroid metabolism gene CYP17, Human Molec. Genet., 3 , 1873-1876 (1994). J. T. Kadonaga, K. A. Jones, and R. Tjian. Promoter-specific activa- tion of RNA polymerase I1 transcription by SpI, Trends Biochem. Sci.,
H. S. Feigelson, G. A. Coetzee, L. N. Kolonel, R. K. Ross, and B. E. Henderson, A polymorphism in the CYP17 gene increases the risk of breast cancer, Cancer Res., 57, 1063-1065 (1997). M. Poutanen, V. Isomaa, V.-P. Lehto, and R. Vihko, Immunologi- cal analysis of 17P-hydroxysteroid dehydrogenase in benign and ma- lignant human breast tissue, Int. J . Cancer, 50. 386-390 (1992). T. Normand, S. Narod, F. Labrie, and J. Simard, Detection of poly- morphisms in the estradiol 17P-hydroxysteroid dehydrogenase I1 gene at the EDH17B2 locus on 17qll-q21, Human Molec. Genet., 2. 479- 483 (1993). A. Mannermaa, H. Peltoketo, R. Winqvist, B. A. J. Ponder, H. Kiviniemi, D. F. Easton, M. Poutanen, V. Isomaa, and R. Vihko, Human familial and sporadic breast cancer: analysis of the coding regions of the 17P-hydroxysteroid dehydrogenase 2 gene (EDH 17B2)
451-453 (1990).
11, 20-23 (1986).
Dru
g M
etab
olis
m R
evie
ws
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Mic
higa
n U
nive
rsity
on
10/3
1/14
For
pers
onal
use
onl
y.

432 FEIGELSON ET AL.
16.
17.
18.
19.
20.
21.
22.
23.
24.
25.
26.
27.
using a single-strand conformation polymorphism assay, Human Gener., 93, 319-324 (1994). T. J. Puranen, M. H. Poutanen, H. E. Peltoketo, P. T. Vihko, and R. K. Vihko, Site-directed mutagenesis of the putative active site of human 17~-hydroxysteroid dehydrogenase type 1, Biochem. J., 304,
A. Castagnoli, I. Maestri, F. Bemardi, and L. Del Senno, PvuII RFLP inside the human estrogen receptor gene, Nucl. Acids Rex, 15, 866 (1987). T. Garcia, M. Sanchez, J. L. Cox, P. A. Shaw, J. B. A. Ross, S. Lehrer, and B. Schachter, Identification of a variant form of the hu- man estrogen receptor with an amino acid replacement, Nucl. Acids Res.. 17, 8364 (1989). T. I. Andersen, K. R. Heimdal, M. Skrede, K. Tveit, K. Berg, and A. L. Borresen, Oestrogen receptor (ESR) polymorphisms and breast cancer susceptibility, Human Gener., 94, 665-670 (1994). L. Del Senno, G. L. Aguiari, and R. Piva, Dinucleotide repeat poly- morphism in the human estrogen receptor (ESR) gene, Human Molec. Gener., 1, 354 (1992). M. Keaveney, J. Klug, M. T. Dawson, P. V. Nestor, J. G. Neilan, R. C. Forde, and F. Gannon, Evidence for a previously unidentified upstream exon in the human oestrogen receptor gene, J. Molec. Endocrinol., 6, 111-115 (1991). K. Grandien, Determination of transcription start sites in the human estrogen receptor gene and identification of a novel, tissue-specific, estrogen receptor-mRNA isoform, Molec. Cell Endocrinol., 116, 207- 212 (1996). K. F. H. Grandien, A. Berkenstam, S. Nilsson, and J. A. Gustafsson, Localization of DNase I hypersensitive sites in the human oestrogen receptor gene correlates with the transcriptional activity of two differ- entially used promoters, J . Molec. Endocrinol., 10, 269-277 (1993). K. Grandien, M. Backdahl, 0. Ljunggren, J. A. Gustafsson, and A. Berkenstam, Estrogen target tissue determines alternative promoter utilization of the human estrogen receptor gene in osteoblasts and tu- mor cell lines, Endocrinology, 136, 2223-2229 (1995). R. K. Ross and D. Schottenfeld, Prostate cancer, in Cancer Epidemology and Prevention, 2nd ed. (F. Fraumeni, ed.), Oxford University Press, Oxford, 1996, pp. 1180-1206. R. K. Ross, L. Bernstein, H. Judd, R. Hanisch, M. C. Pike, and B. E. Henderson, Serum testosterone levels in young black and white men, J. Narl. Cancer Insr., 76. 45-48 (1986). R. K. Ross, L. Bernstein, R. A. Lobo, H. Shimizu, F. Z. Stanczyk,
289-293 ( 1994).
Dru
g M
etab
olis
m R
evie
ws
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Mic
higa
n U
nive
rsity
on
10/3
1/14
For
pers
onal
use
onl
y.

GENETIC SUSCEPTIBILITY TO CANCER 433
28.
29.
30.
31.
32.
33.
34.
35.
36.
37 *
38.
M. C. Pike, and B. E. Henderson, 5-Alpha-reductase activity and risk of prostate cancer among Japanese and U.S. white and black males, Lancet, 339, 887-889 (1992). A. E. Thigpen, D. L. Davis, T. Gautier, J. Imperato-McGinley, and W. Russell, The molecular basis of steroid 5-alpha-reductase deficiency in a large Dominican kindren, N . Engl. J. Med., 327, 1216-1219 (1992). D. L. Davis and D. W. Russell, Unusual length polymorphisms in human steroid 5-a-reductase type 2 gene (SRD5A2), Human Mulec. Genet., 2, 820 (1993). J. K. V. Reichardt, N. Makridakis, B. E. Henderson, M. C. Yu, M. C. Pike, and R. K. Ross, Genetic variablity of the human SRD5A2 gene: Implications for prostate cancer risk, Cancer Res., 55, 3973- 3975 (1995). N . Makridakis, R. K. Ross, M. C. Pike, L. Chang, F. Z. Stanczyk, L. N. Kolonel, C. Y. Shi, M. C. Yu, B. E. Henderson, and J. K. V. Reichardt, A prevalent missense substitution that modulates activity of prostatic steroid 5a-reductase, Cancer Res., 57, 1020-1022 (1997). G. Jenster, H. A. G. M. Van der Korput, C. Van Vroonhoven, T. H. Van der Kwast, J. Trapman, and A. 0. Brinkmann, Domains of hu- man androgen receptor involved in steroid binding, transcriptional activation, and subcellular localization, Mulec. Enducrinul., 5 , 1396- 1404 (1991). A. R. La Spada, E. M. Wilson, D. B. Lubahn, A. E. Harding, and K. H. Fischback, Androgen receptor gene mutations in X-linked spi- nal and bulbar muscular atrophy, Nature, 352, 77-79 (1991). A. N. Mhatre, M. A. Trifiro, M. Kaufman, P. Kazemi-Esfarjani, D. Figlewicz, G. Rouleau, and L. Pinsky, Reduced transcriptional regu- latory competence of the androgen receptor in X-linked spinal and bulbar muscular atrophy, Nature Genet., 5, 184- 188 (1993). A. Edwards, H . A. Hammond, L. Jin, C. T. Caskey, and R. Chakraborty, Genetic variation at five trimeric and tandem repeat loci in four human population groups, Genumics, 12, 241-253 (1992). R. A. Irvine, M. C. Yu, R. K. Ross, and G. A. Coetzee, The CAG and GGC microsatellites of the androgen receptor gene are in linkage disequilibrium in men with prostate cancer, Cancer Res., 55, 1937- 1940 (1995). S. A. Ingles, R. K. Ross, M. C. Yu, R. A. Irvine, G. LaPera, R. W. Haile, and G. A. Coetzee, Association of prostate cancer risk with genetic polymorphisms in vitamin D receptor and androgen receptor, J, Nafl. Cancer Znst., 89, 166-170 (1997). J. L. Stanford, J. J. Just, M. Gibbs, K. G. Wicklund, C. L. Neal, B.
Dru
g M
etab
olis
m R
evie
ws
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Mic
higa
n U
nive
rsity
on
10/3
1/14
For
pers
onal
use
onl
y.

434 FEIGELSON ET AL.
A. Blumenstein, and E. A. Ostrander, Polymorphic repeats in the androgen receptor gene: molecular markers of prostate cancer risk, Cancer Res., 57, 1194-1 198 (1997).
39. E. Giovannucci, M. J. Stampfer, K. Krithivas, M. Brown, A. Brufsky, J. Talcott, C. H. Hennekens, and P. W. Kantoff, The CAG repeat within the androgen receptor gene and its relationship to prostate can- cer, Proc. Natl. Acad. Sci. USA., 94, 3320-3323 (1997).
Dru
g M
etab
olis
m R
evie
ws
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
Mic
higa
n U
nive
rsity
on
10/3
1/14
For
pers
onal
use
onl
y.